 Yaru Hou
Yaru Hou Xiaoyu Zhang*
Xiaoyu Zhang* Zhenfeng Gao
Zhenfeng Gao Lixin Zhang
Lixin Zhang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 24 March 2023
Sec. Food Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1132271
This article is part of the Research Topic Postharvest Disease Management in Fruits and Vegetables: Recent Advances and Mechanisms View all 6 articles
Postharvest decay is an urgent problem that affects the storage of pears. Low temperature storage is one of the most important methods to reduce the prevalence of fruit diseases during storage. In this study, the microbial diversity of postharvest Yuluxiang pear (Pyrus × michauxii “Yu Lu Xiang”) fruits stored at low temperature for different lengths of times was analyzed. Illumina MiSeq high-throughput sequencing was used to analyze the composition and diversity of fungal communities. The results showed that the fungi within fruit were classified into 6 phyla, 18 classes, 40 orders, 72 families, and 92 genera based on the 97% sequence similarity level. They belonged to 6 phyla, 18 classes, 40 orders, 72 families, and 92 genera. The highest richness of fungi was obtained after 30 d of treatment. The β-diversity index showed that the fungal community composition of these fruit was significantly different at the beginning of storage compared with the different timepoints of samples at low temperature during storage. The comparison of fungal composition at the phylum level indicated that Ascomycota was dominant in the different timepoints of samples at low temperature, while Alternaria was the primary fungus at the genus level. A correlation analysis was used to further explore the correlation between fungi and fruit firmness, titratable acid, and solid soluble contents at low temperatures during storage. Aureobasidium and Didymella positively correlated with the soluble solids and hardness. Phoma positively correlated with the titratable acid, and Aspergillus positively correlated with titratable acid and hardness. This study can guide the industrial production of Yulu pear and also provide a theoretical basis to prevent and control diseases during the storage period of Yulu pear.
Yuluxiang pear (Pyrus × michauxii “Yu Lu Xiang”) is the primary fruit grown in orchards in Shanxi Province, China. It is also one of the few varieties that is grown independently of China. Experts in the national pear industry system recognize it as China’s No. 1 pear (Ma et al., 2019; Zhang et al., 2020). The Yuluxiang pear has an important place in reorganizing the planting industry structure in Shanxi Province and serving as a strategic pillar industry to decrease the poverty of growers. However, owing to its thin peel and natural richness in nutrients, some types of pathogenic fungi and opportunistic pathogens can attach to its surface, and diseases, such as mold, can occur owing to improper temperature and humidity settings during the harvesting and storage processes (Sun et al., 2017). During the process of transportation and low temperature storage, fungal infection can cause common diseases, such as pear pulp browning disease, brown rot, and soft rot among others (Yu et al., 2012; Li et al., 2016). These conditions seriously affect the commodity value of fruits. Thus, the study of fungal diversity of fruits under low temperature storage has become an important issue.
In recent years, with the rapid development of molecular biology techniques and the popularization of extensive data analyses, high-throughput sequencing (HTS) technology is becoming an indispensable tool to study microbial community structure and composition (Xu et al., 2016; Dai et al., 2019; Abdel-Wahab et al., 2021). It provides a convenient method to examine the diversity of organisms in nature and enables researchers to more comprehensively and accurately measure the relative abundance of various microorganisms in the biological environment (Chen C. K. et al., 2020). It is less labor-intensive and more efficient than previous culture-based techniques, and it provides deeper insights into the diversity of communities (Chen L. et al., 2020). A variety of fungi, such as Alternaria, Fusarium, and Penicillium among others, were mostly used in previous reports on Yuluxiang pear fungi based on culture technology (Hou et al., 2022). As far as we know, they only quantified the proportion of fungi that belonged to specific culturable taxa based on the media used. However, fungal diversity can be affected by differences in plant types, sample sizes, sampling time points, and extraction methods. Now, HTS has been utilized to study the community structure and microbial diversity on the surface of fruits and vegetables (Burgos et al., 2017), including apple (Malus domestica), grape (Vitis vinifera), lettuce (Lactuca sativa), corn meal, peels, peach (Prunus persica), pepper (Capsicum spp.), spinach (Spinacea oleracea), strawberry (Fragaria x ananassa), tomato (Solanum lycopersicum), and pear (Pyrus spp.) among others (Leff et al., 2013; Lopez-Velasco et al., 2013; Chen et al., 2019). In addition, it is widely used in agriculture, industry, medicine, and other fields (Salmaso et al., 2018; Zang et al., 2018).
Therefore, this study used Illumina MiSeq (Illumina, San Diego, CA, United States) next-generation sequencing technology to analyze the changes in fungal community structure and diversity at different stages after harvest of Yuluxiang pear, which laid a foundation for preventing postharvest disease and preserving Yulu pear, which is crucial to promoting the development of a circular economy in Yuluxiang pear.
Yuluxiang pears were harvested in Zhaizi Township, Xixian County, Linfen City, Shanxi Province, China, in September 2019. Fruits of uniform size that no obvious diseases, pests, or mechanical damage were selected. In addition, fruit with a soluble solids content of 12.5 to 16.1%, total sugar of 8.7 to 9.8%, and titratable acid of 0.08 to 0.17% were transported to the laboratory in a refrigerated truck within 24 h and stored in a cold storage facility at 4°C ± 0.5°C. Mixtures of the peels and pulp of pears that were stored in a cold storage at 4°C ± 0.5°C were cut into sections of approximately 1 cm × 1 cm every 30 d with a sterile knife and designated (A) for the initial 30 d sample (A), and then 60 d (B), 90 d (C), and 120 (D). Each treatment was conducted in triplicate.
The firmness of the pulp was measured with a GY-1 fruit firmness tester (Shandong, China). PAL-BX/ACIDF5 (Beijing, China) sugar and the integrated acid machine were used to determine the contents of titratable acid and soluble solids in the pulp.
The samples were ground in liquid nitrogen to ensure uniformity. DNA kits (MoBio Laboratories, Inc., Carlsbad, CA, United States) were used to extract total DNA from the samples in the four storage periods. Simultaneously, a Nanodrop spectrophotometer (ND-1000; Thermo Fisher Scientific, Waltham, MA, United States) was utilized. The DNA was quantified, and the quality of DNA extraction was examined by 1.2% agarose gel electrophoresis (Zhao et al., 2020). The ribosomal ribonucleic acid (rRNA) gene ITS region using ITS1F (5’-CTTGGTCATAGAGAGTAA-3′) and ITS2 (5’-GCTGCGTTCTTCATC GATGC-3′) were amplified by PCR (Tang et al., 2022; Wang et al., 2022).
The sequencing libraries were created using Illumina’s TruSeq Nano DNA LT Library Prep Kit. The quality of library was tested on an analyzer with a DNA kit before sequencing. The qualified sequencing library was diluted in a gradient, mixed, and denatured by NaOH to single-stranded DNA for sequencing. The optimal length of the target fragment for sequencing was 200–450 bp.
Diversity analyses of the community composition were conducted, including α-diversity and β-diversity. α-Diversity is primarily used to analyze the diversity and abundance of fungal communities in the samples (Xue et al., 2021). The Observed Species and Chao1 indices reflect the abundance of fungal flora between different timepoints of samples at low temperature, and the Shannon and Simpson indices reflect the diversity of fungi in each treatment. Studies of the α-diversity index also included dilution curves, a rarefaction curve, and coverage index to detect the degree of sequencing coverage of the operational taxonomic units (OTUs) (Zhang et al., 2019). The β-diversity analysis primarily included a principal coordinates analysis (PCoA) and a β-diversity index between-group difference analysis (Wang et al., 2019). The operational taxonomic unit (OTU) classification table primarily provided the distribution of OTU composition of each sample at the taxonomic level (phylum, class, order, family, genus, and species), which reflects the community structure of the models at different taxonomic levels (Wang et al., 2017).
The sequences were optimized with Trimmomatic (v 0.4), Flash (v 1.2.11), USEARCH 10.0 and QIIME (v1.9.1) and a Perl program that was developed by Shanghai Yuanxin Biologics (Shanghai, China) that clustered the effective sequences according to 97% consistency that then became operational taxa. QIIME (v 1.9.1) and an RDP Classifier Bayesian algorithm performed classification analysis on 97% similar OTU representative sequences to calculate the index of fungal diversity. The R (v 4.0.3) language Vegdist and Hclust were used for community heatmap mapping, and Bray-Curtis distance calculations (Bray and Curtis, 1957) were used to conduct a cluster analysis. The R (v 4.0.3) language was used for the PCoA and PERMANOVA analyses. The relationship between fungal diversity and community structure and fruit hardness, titratable acid and soluble solids content was studied by a Spearman correlation analysis. The R language tool was used to create a correlation heatmap diagram.
A statistical analysis of the test data was conducted using Microsoft Excel 2007 (Redmond, WA, United States) and SPSS 23.0 (IBM, Inc., Armonk, NY, United States), and Origin2021 (OriginLab, Northampton, MA, United States) was used to draw the horizontal distribution map of the phyla and genera. Duncan’s new complex range method was used to test the significance of difference.
The hardness, soluble solids, and titratable acid content of Yuluxiang pear fruits during storage are shown in Table 1. The fruit firmness, soluble solids, and titratable acid content decreased as the storage period was extended. In early storage, the fruit hardness decreased rapidly, and it then remained stable. The decrease in fruit hardness could be related to the gradual degradation of cellulose and the decomposition of pectin and starch in the fruit. In addition, the content of soluble solids also decreased as the time of storage was prolonged, which could be related to the gradual consumption of organic matter, such as soluble sugars, in the fruit that was converted into CO2 and H2O during the metabolic process. The titratable acid content decreased rapidly, and there were significant differences in different storage periods, which could be the result of ethanol absorption by the fruits that interacted with the organic acids (Table 1).
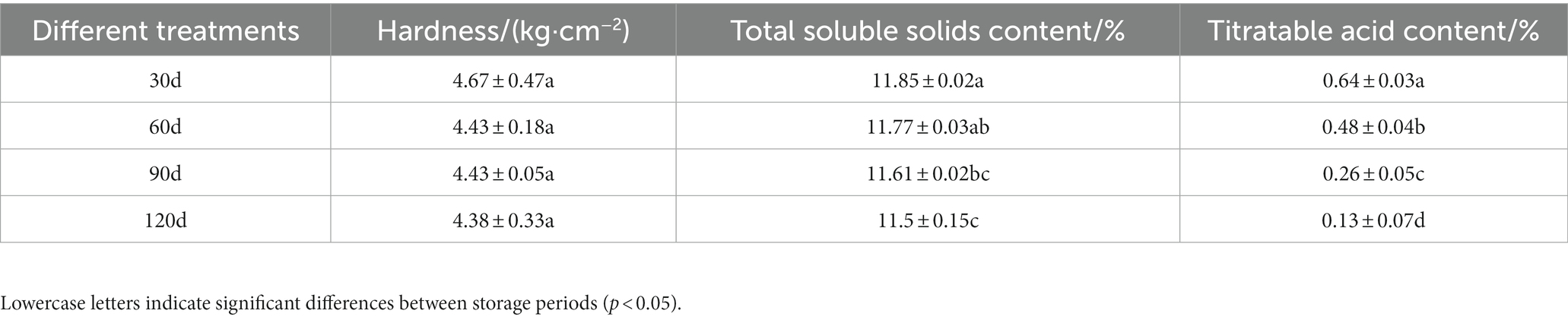
Table 1. Hardness, titratable acid, and solid soluble content of Yuluxiang pears in different storage periods.
A total of 1,792,803 valid sequences were obtained for total sequencing, with an average of 149,400 ± 16,282 sequences per sample. Based on the α-analysis of the sample, the rates of coverage of the 30 d(A), 60 d(B), 90 d(C) and 120 d(D) libraries were relatively high (>99.9%), indicating that sequencing increased the coverage of this species. The results were reliable, and the data met the sequencing requirements. Observed species and Chao1 were used to evaluate the abundance of fungal flora in different timepoints of samples at low temperature. Shannon and Simpson indices were used to assess the diversity of microorganisms in each treatment. The Observed species (78 ± 8.75) and Chao1 (78.33 ± 8.81) values at 30 d(A) were greater than those at 60 d(B), 90 d(C), and 120 d(D), indicating that the 30 d(A) treatment had the most fungi, followed by the 60 d(B), 90 d(C), and 120 d(D) groups. The Shannon and Simpson indices at 30 d(A) were 3.98 ± 0.04 and 0.089 ± 0.002, respectively, followed by lower values at 90 d(C), 60 d(B), and 120 d(D), indicating that the 30 d(A) treatment had the highest degree of fungal diversity. The Shannon and Simpson indices indicated there were specific differences in fungal diversity among different timepoints of samples at low temperature. Compared with other samples, the 30 d(A) samples exhibited a higher richness and diversity of fungi, indicating that there were more species in these samples, and the abundance and diversity of fungal flora in Yuluxiang pear changed with the storage time (Table 2).
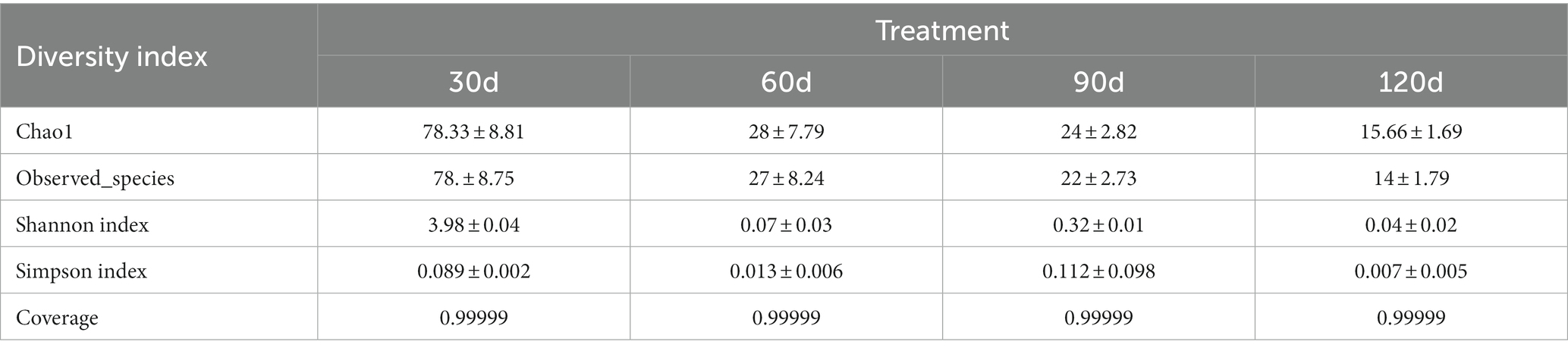
Table 2. Alpha diversity index of different treatments.
The fungal community composition of four groups of pear fruit samples was compared and analyzed. The diversity of four groups of pear fruit fungi was analyzed by a PCoA using the Jaccard distance algorithm to evaluate the similarity and difference of community composition. As shown in Figure 1, the fungal communities in the samples of the same treatment group were clustered, while the fungal communities in the samples of different treatment groups could be significantly separated, indicating that there were obvious differences between the fungi in the samples of different treatment groups. A PERMANOVA experimental analysis showed that the overall p-value was greater than 0.05, indicating that there were significant differences in OTU composition and relative abundance (Figure 1).
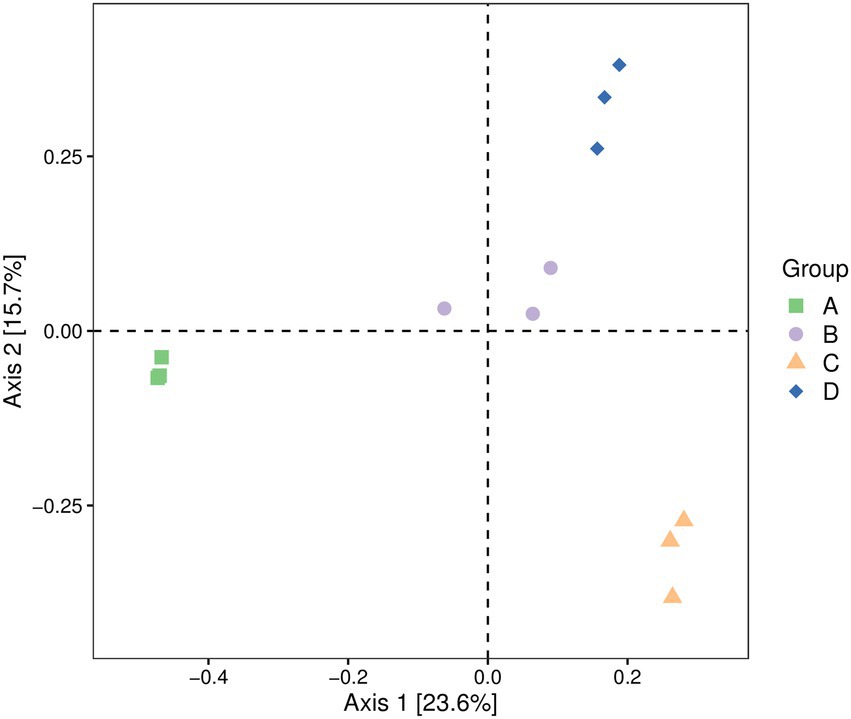
Figure 1. PCoA of fungal communities in Yuluxiang pears after storage at low temperatures for 30 d(A), 60 d(B), 90 d(C), and 120 d(D). Each point represents a sample, and the dots of different colors indicate different samples (groups). The percentages in the parentheses on the axis represent the proportion of sample difference data (distance matrix) that can be interpreted by the corresponding axis. PCoA, Principal coordinates analysis.
A Venn diagram can intuitively reflect the differences and similarities of the OTU composition of the fungal community in different Yuluxiang pear treatment groups. There were 46.57% (95) and 15.2% (31) of the specific fungal community OTU on 60 d(B), 6.37% (13) of the specific fungal community OTU on 90 d(C), and 10.29% of the 120 d(D) OTU (21). In addition, the 30 d(A), 60 d(B), 90 d(C), and 120 d(D) groups shared six OTUs (2.94%) (Figure 2). The unique OTUs of the 30 d(A) treatment were higher than those of the other groups, indicating that the fungal community after 30 d(A) treatment was more diverse than those in the 60 d(B), 90 d(C) and 120 d(D) groups.
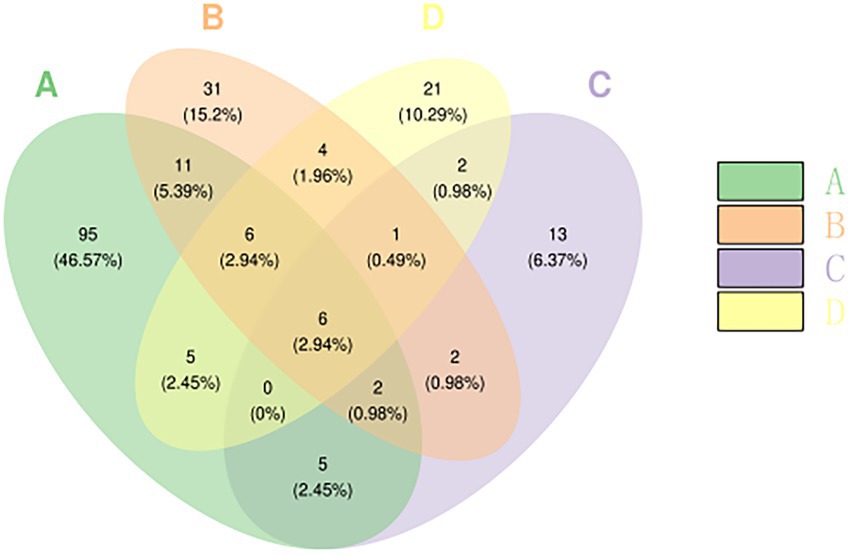
Figure 2. Microbial Venn plot of different samples based on the OTU level. Each ellipse represents a sample (group). The overlapping area between the ellipses indicates the common OTUs between the samples (group), and the number of each block indicates the number of OTUs contained in the block. OTU, operational taxonomic unit.
According to the OTU annotation results of the four groups of samples of Yuluxiang pear, the fungi in the samples were identified as 6 phyla, 18 classes, 40 orders, 72 families, and 92 genera. The species diversity information in the samples was analyzed, and the histogram of the taxonomic abundance of the flora is shown in Figure 3. According to all the sequences in the library, the distribution of the four treatment groups of the Yuluxiang pear at the gate level is shown in Figure 3. The community distribution of the four treatment groups of Yuluxiang pear was primarily Ascomycota, and the abundance of Ascomycota in the four treatment groups was 96, 97, 97 and 97% at 30 d(A), 60 d(B), 90 d(C), and 120 d(D), respectively. The abundance of Ascomycota at the phylum level increased in parallel with the storage time. The 30 d(A) treatment was dominated by Ascomycota and Basidiomycota, with Basidiomycota accounting for 0.85%, while the abundances of Basidiomycota for the 60 d(B), 90 d(C) and 120 d(D) groups were 0.05, 0.05 and 0.14%, respectively. Basidiomycota in the 30 d(A) treatment group was more abundant than those in the 60 d(B), 90 d(C), and 120 d(D) groups. The abundance of Basidiomycota slowly decreased over time. The 30 d(A), 90 d(C), 120 d(D) groups did not contain Mortierellomycota or Olpidiomycota, and these phyla were present at relatively low abundance in the 60 d(B) group.
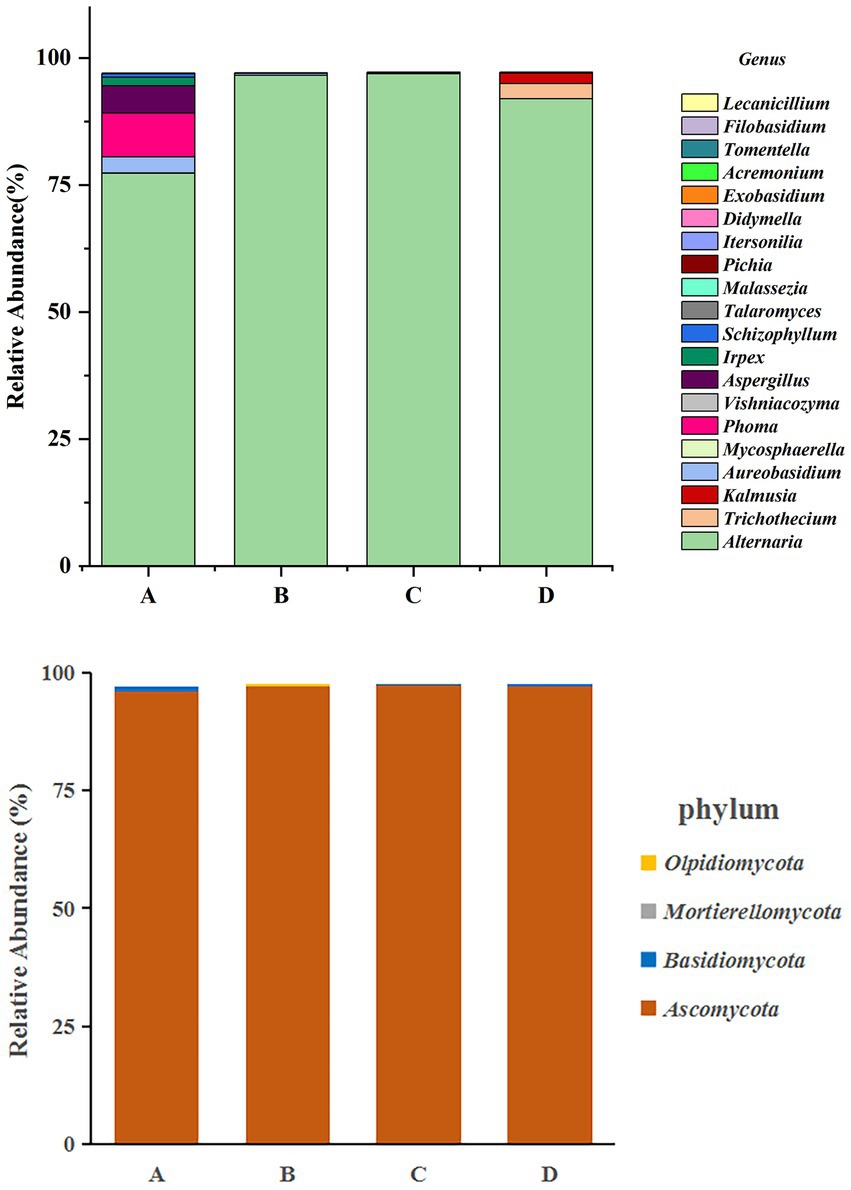
Figure 3. Analysis of fungal community diversity at the phylum level and genus level of Yuluxiang pears stored at low temperature for 30 d(A), 60 d(B), 90 d(C), 120 d(D), and different colored dots indicated different samples (groups).
In terms of the horizontal composition of the genera, the relative abundances of Alternaria at 30 d(A), 60 d(B), 90 d(C) and 120 d(D) were 77.4, 96.6, 96.9 and 92%, respectively, and Alternaria was the dominant genus. In addition, the abundances of Phoma, Aureobasidium, Aspergillus and Irpex were 8.64, 3.2, 5.33 and 1.73% in the 30 d(A) treatment, and 0.39% in the 60 d(B) treatment. The abundance of Mycosphaerella in the 90 d(C) treatment was 0.26%, while it was 3.1% in the 120 d(D) treatment. The abundance of Kalmusia was 2.1%. As shown in Figure 3, Alternaria was the overwhelmingly dominant community in the four groups of samples. It was distributed across all the samples and dominant in most of them at the genus level. There was a gradual decline in the viability of Phoma, Aureobasidium, and Aspergillus in the 30 d(A), 60 d(B), 90 d(C), and 120 d(D) groups, which could have been owing to the poor culture conditions that hampered the growth of fungi. Thus, the viability of the fungi had been greatly reduced, or they even died. However, different pathogens, such as Trichothecium and Kalmusia, appeared later. They were present at relatively low levels (relative abundance<0.1%), so they were not analyzed.
Figure 4 shows a heatmap analysis of the distribution of the 20 OTUs with the highest abundance between groups. A heatmap can visually reflect the similarity and difference of the fungal community composition of each group through color changes, which is more convenient to find genera that were more or less distributed in the sample. A redder color indicates that a higher proportion of the genus was contained in the sample. A bluer color indicates a lower proportion. A lateral homogenization treatment was conducted. As shown in Figure 4, A1 and A3 were clustered in A; B1 and B3 were clustered in B, and D1 and D3 were clustered in D. The similarity of their microbiota was relatively higher than those of the other samples in the same group. However, C1, C2 and C3 were separated in the C treatment, indicating that there were differences between the fungal communities in the same batch of Yuluxiang pear, and there were also differences between the fungal communities of different batches. In addition, LDA histograms (default threshold: LDA > 2) and evolutionary clade plots (Figure 5) were outputted to use LEfSe to identify the major groups that were specific between the groups. There was no differentiation of species in the 60 d(B) samples.
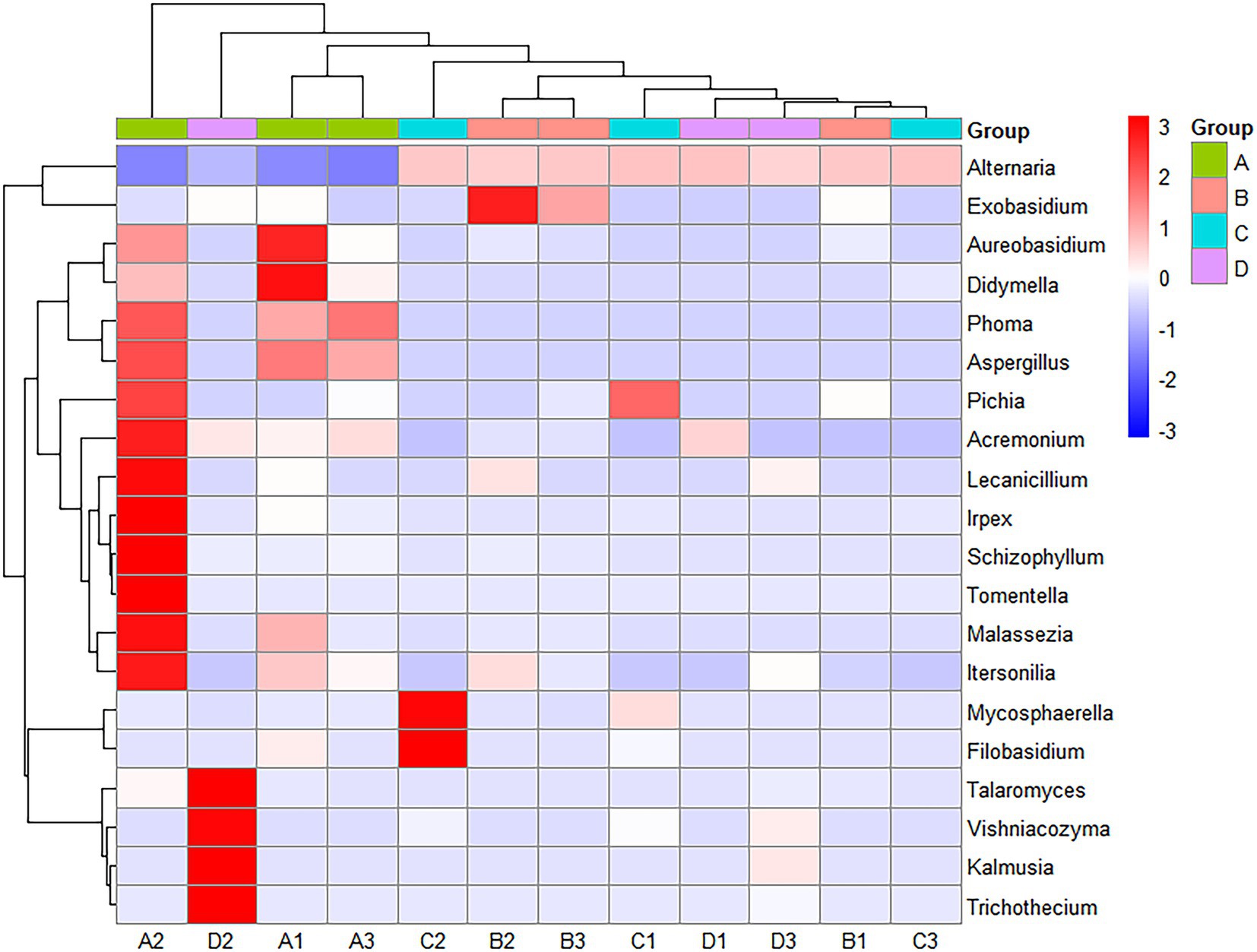
Figure 4. A cluster heatmap of the flora at the genus level. The red patch in the figure indicates that the genus is more abundant in this sample than in the other samples, and the blue patch indicates that the genus is less abundant in this sample than in the other samples.
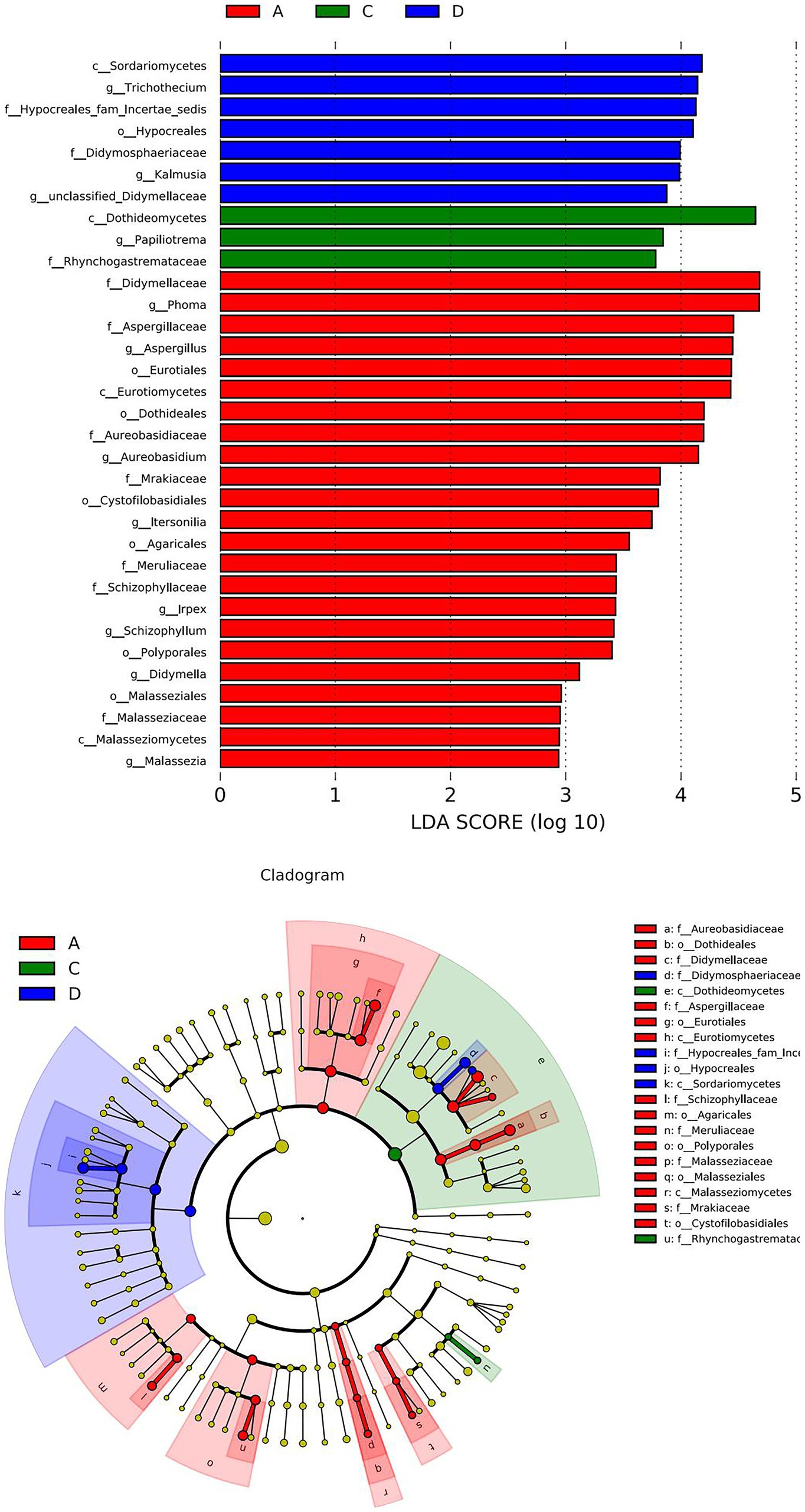
Figure 5. Histogram of the LDA effect values of marker species and differential taxa between groups. The LDA ordinate is a taxa with significant differences between the groups, and the abscissa visually displays the logarithmic score values of the LDA analysis for each taxa in a bar chart. The taxonomic clade shows the taxonomic hierarchical relationships of the main taxa in the sample community from the phylum to genus (from inner circle to outer circle). The node size corresponds to the average relative abundance of that taxa. Hollow nodes represent taxa with insignificant intergroup differences, while the nodes of other colors indicate that these taxa exhibit significant between-group differences and are more abundant in the grouped sample represented by that color. Letters identify the names of taxa that differ significantly between groups. LDA, linear discriminant analysis.
The LDA histogram shows the fungal microorganisms with significant differences in 30 d(A), 90 d(C), and 120 d(D) of the sample, and a longer bar for the taxa indicated that they were more significant. The color of the bar chart indicated that the group of samples with the highest abundance corresponded to the taxon. c_Sordariomycetes, g_Trichothecium, f_Hypocreales_fam_Incertae_sedis, o_Hypocrea les, f_Didymosphaeriaceae, g_Kalmusia and g_unclassified_Didymellaceae were more prevalent at 120 d(D) than in 30 d(A) and 60 d(B). There was a significant difference in 90 d(C). The c_Dothideomycetes, g_Papiliotrema and f_Rhyncho gastremataceae in 90 d(C) were significantly different from those in the 30 d(A), 60 d(B) and 120 d(D) groups. f_Didymellaceae, g_Phoma, f_Aspergillus, o_Eurotiales, c_Eurotiomycetes, o_Dothideales, g_Aureobasidium, f_Mrakiaceae, o_Cystofilobasidiales, g_Itersonilia, o_Agaricales, f_ in 30D Meruliaceae, g_Irpex, g_Schizophyllum, o_Polyporales, g_Didymella and g_Malassezia were significantly different from those in the 60 d(B), 90 d(C) and 120 d(D) groups.
Figure 6 is a heatmap created by a correlation analysis of the 20 OTUs with the highest levels of genera and fruit hardness, titratable acid and soluble solids. Aureobasidium and Didymella positively correlated with the soluble solids and hardness. Phoma positively correlated with the titratable acid, and Aspergillus positively correlated with titratable acid and hardness.
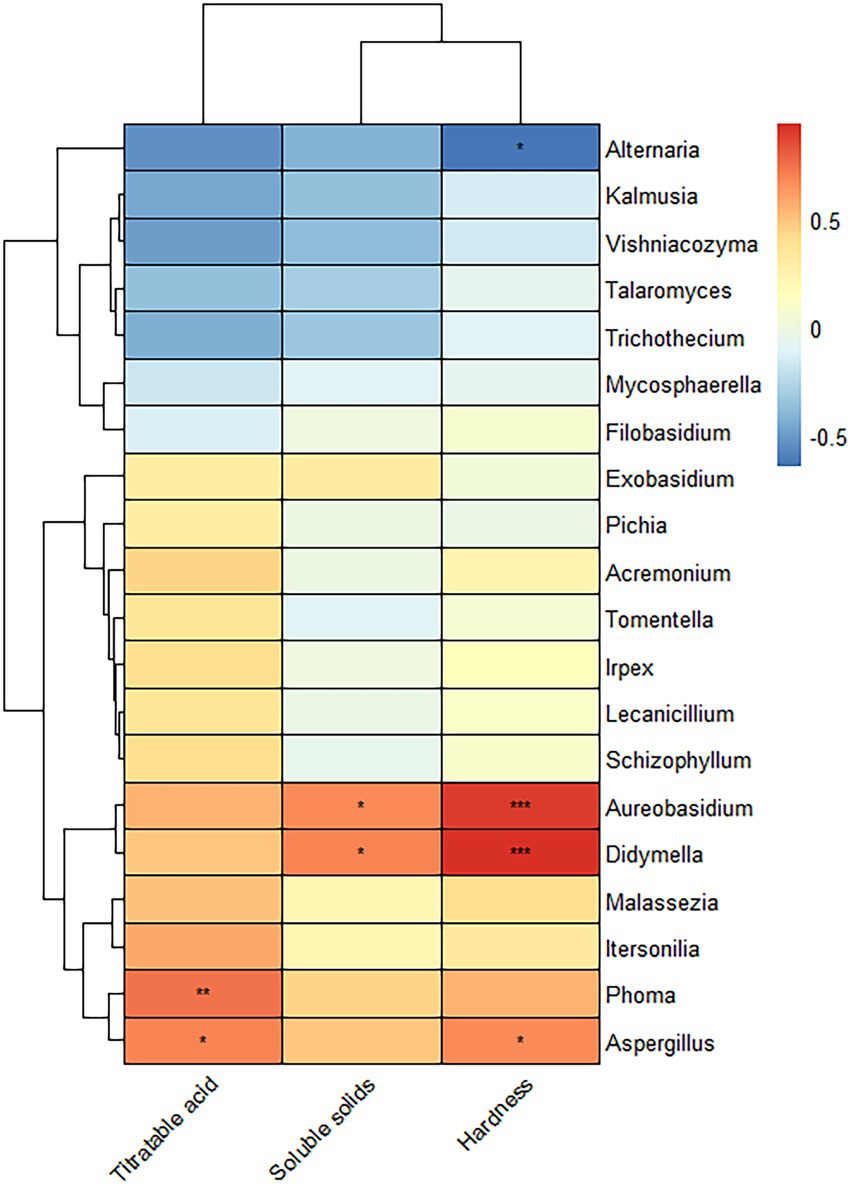
Figure 6. Correlation analysis of the fungi of Yuluxiang pear with fruit hardness, titratable acid, and soluble solids in different storage periods. Each column represents a bacterium. Blue is a negative correlation, and red is a positive correlation. * is a distinctive identifier.
In China, pears are harvested in season, and most of them are stored refrigerated for long-term sales. During the past 10 years, the implementation of refrigerated transportation has extended the postharvest storage of pears and their shelf life. However, with the extension of the storage period, pears will have some diseases owing to the invasion of fungi. From a general perspective, cold storage is often thought to decrease the fruit ripening process and delay the growth of spoilage fungi. However, some plant pathogenic fungi can cause postharvest deterioration, which seriously affects the safety and quality of fruits (Prasanna et al., 2007; Morales et al., 2010; Shen et al., 2018). Therefore, identifying the fungal diversity of pear fruits has become an important object of fruit research, which can provide a new perspective to predict and control postharvest diseases in pear fruits.
In this study, HST was used to analyze the structure and diversity of fungal colonization of Yuluxiang pear fruits in different storage periods. The sparsity curve suggests that the current sequencing depth is deep enough to detect most of the fungi on pears.
The comparison of the three biodiversity indices of Chao, Shannon, and Simpson revealed that the highest abundance of flora and fungal diversity was observed after 30 d(A) of treatment. A ß-diversity analysis showed that the fungal communities in the samples of the same treatment group were clustered together. The fungal communities in the samples of different treatment groups could be significantly separated, indicating that there were clear differences in fungi between the samples of different treatment groups. Furthermore, low temperature can delay the growth of spoilage pathogens, and the effect of storage time on the fungal community is complex (Dadzie et al., 2021; Tian et al., 2021). The results showed that cryogenic storage reduced the α-diversity of microorganisms over time. It is hypothesized that some fungi will become intolerant with prolonged storage, inhibiting their growth.
In the community structure analysis of the Yuluxiang pear fruit fungi at different storage stages, it emerged that the level of composition of fungi at the phylum level was relatively simple and was primarily composed of Ascomycota, Basidiomycota, and Mortierellomycota. Among them, Ascomycota dominated the number of community compositions. There were differences in the level of Basidiomycota among the groups at 30 d(A), 60 d(B), and 90 d(C), presumably owing to the colonization of pathogenic fungi. HST was used to comprehensively analyze the samples, and Ascomycota was the primary phylum that was detected. It is the largest phylum in the fungal kingdom and was present in all the samples (Abdelfattah et al., 2016; Shen et al., 2018). There are studies that have shown that the main fungi on the surface of pear are Ascomycota, which grow quickly and can survive harsh conditions with low nutrient levels (Challacombe et al., 2019). They can adapt to a wide range of substrates in challenging environments, such as ultraviolet light, water and high temperature stress (Jaber et al., 2012). At the level of genus, Alternaria was more abundant in all the different timepoints of samples at low temperature. In addition, Hou et al. (2022) obtained Alternaria during the isolation and identification of postharvest pathogenic fungi of Yuluxiang pear, indicating that Alternaria is the dominant genus that causes postharvest disease on Yuluxiang pear. Alternaria alternata is highly adaptable to Yuluxiang pears and is also the primary pathogenic fungus that causes pear black spot disease in China (Roberts, 2005; Wang and Zhang, 2010; Ttpabc et al., 2019). As shown in the figure, fungal diversity at the genus level slowly stabilized, and studies have shown that after harvesting, the fungal microbiome changes significantly during storage and becomes an aging plant microbiome. It is characterized by a decrease in the diversity of fungal microbial diversity and an increase in the degradation and decomposition of the special fungal microflora (Kusstatscher et al., 2020). A LEfSe analysis yielded species that varied between the different sample groups. As compared with the other three groups of different species, the 30 d(A) treatment had the highest abundance of fungi. The results indicate that the fungal community structure is the most complex during the early stages of preservation of Yuluxiang pear. With the extension of storage time, the fungal community structure was relatively stable.
Temperature, relative humidity, pH, soluble solids, and hardness are the main factors that influence fungal colonization (Li et al., 2022). The results of this study on the correlation between fungal communities and fruit quality illustrate the hardness, titratable acid, and solid soluble content of Yuluxiang pears at different postharvest storage periods. A downward trend was observed, which was similar to the findings of Zhang et al. (2021). The results of the study on the correlation between fungal communities and fruit quality indicated that Aureobasidium and Didymella increased when the content of soluble solids and hardness increased. Similarly, the abundances of Aureobasidium and Didymella also increased when the content of soluble solids and hardness increased. The increase in Phoma correlated with the fruit titratable acid. The fruit titratable acid and hardness increased in parallel with the abundance of Aspergillus, Aureobasidium, Phoma, and Aspergillus, which were all abundant at 30 d(A), indicating that the fungal diversity of the sample at 30 d(A) was closely related to fruit hardness, titratable acid and soluble solids.
HTS was used to analyze the community structure and fungal diversity of Yuluxiang Pear at different storage periods. The diversity of treated microflora was found to differ as the storage time was extended. Compared with the treatment at the storage stage, the composition of the fungal community was significantly different from that at the beginning of storage. Comparison of the fungal phyla showed that Ascomycota was the dominant phylum in different treatment groups, with Alternaria dominating at the genus level. In addition, the relationship between fungal community structure and hardness, titratable acid and soluble solids at different storage stages was also discussed. To improve the prevention and control of fruit diseases and quality control of Yuluxiang pear, this study provides assistance for the industrial production of Yulu pear.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/, PRJNA909451.
All authors listed have made a substantial, direct, and intellectual contribution to the work and approved it for publication.
This work was supported by the Natural Science Foundation of Shanxi Province (201901D111451) and the Research Innovation Team Training Project of Shanxi Academy of Agricultural Sciences (YGC2019TD05).
I would like to thank TC and LZ for their help in data collection and experimental design of my paper, ZG for his help in materials, instruments, and equipment in the experimental process, and XZ for her guidance in the experimental process and her control over the writing of the paper.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abdelfattah, A., Wisniewski, M., Droby, S., and Schena, L. (2016). Spatial and compositional Vari ation in the fungal communities of organic and conventionally grown apple fruit at the consumer point-of-purchase. Hort. Res. 3:16047. doi: 10.1038/HORTRES.2016.47
Abdel-Wahab, M. A., Bahkali, A. H., Elgorban, A. M., and Jones, E. B. (2021). High-throughput amplicon sequencing of fungi and microbial eukaryotes associated with the seagrass Halophila stipulacea (Forssk.) Asch. From Al-Leith mangroves, Saudi Arabia. Mycol. Prog. 20, 1365–1381. doi: 10.1007/S11557-021-01744-2
Bray, J. R., and Curtis, J. T. (1957). An ordination of the upland forest communities of southern Wisconsin. Ecological monographs 27, 326–349. doi: 10.2307/1942268
Burgos, M., Aguayo, M., Pulido, R. P., Gálvez, A., and Lucas, R. (2017). Analysis of the bacterial biodiversity of peaches under refrigerated storage after treatment by high hydrostatic pressure. Food Bioprod. Process. 102, 55–61. doi: 10.1016/j.fbp.2016.12.003
Challacombe, J. F., Hesse, C. N., Bramer, L. M., McCue, L. A., and Kuske, C. R. (2019). Genomes and secretomes of Ascomycota fungi reveal diverse functions in plant biomass decomposition and pathogenesis. BMC Genomics 20:976. doi: 10.1186/s12864-019-6358-x
Chen, L., Li, Y. Z., Jin, L., He, L., Ao, X. L., Liu, S. L., et al. (2020). Analyzing bacterial community in pit mud of Yibin baijiu in China using high throughput sequencing. Peer J. 8:e9122. doi: 10.7717/peerj.9122
Chen, J. R., Yan, R. X., Hu, Y. F., Zhang, N., and Hu, H. Y. (2019). Compositional shifts in the fungal diversity of garlic scapes during postharvest transportation and cold storage. LWT 115:108453. doi: 10.1016/j.lwt.2019.108453
Chen, C. K., Zhang, H. J., Zhang, X. J., Dong, C. H., Xue, W. T., and Xu, W. T. (2020). The effect of different doses of ozone treatments on the postharvest quality and biodiversity of cantaloupes. Postharvest Biol. Technol. 163:111124. doi: 10.1016/j.postharvbio.2020.111124
Dadzie, R. G., Amoah, R. S., Ampofo-Asiama, J., Quaye, B., Kizzie-Hayford, N., and Abano, E. E. (2021). Improving the storage quality of eggplants (solanum Aethiopicum L.) fruit using aloe Vera gel coating. J. Food Technol. Res. 8, 58–66. doi: 10.18488/journal.58.2021.82.58.66
Dai, C. W., Xu, Y., Wu, C. J., and Cai, B. (2019). Application progress in food-borne pathogens detection based on high-throughput sequencing technology. J. Food Safe. Qual. 10, 6006–6012. doi: 10.19812/j.cnki.jfsq11-5956/ts.2019.18.007
Hou, Y. R., Gao, Z. F., Yang, Z. G., Chen, T., Zhang, Y., Guan, J. F., et al. (2022). Isolation, identification and biological characteristics of postharvest pathogens of Yuluxiang. Pear. Sci. Technol. Food Ind. 43, 122–129. doi: 10.13386/j.issn1002-0306.2021110364
Jaber, B. M., Al-Silawi, R., and Al-Najjar, T. (2012). Isolation and molecular identification of ascomycetes in sediments and waters of the Gulf of Aqaba. Red Sea. Natural ence 04, 555–561. doi: 10.4236/ns.2012.48074
Kusstatscher, P., Cernava, T., Abdelfattah, A., Gokul, J., Korsten, L., and Berg, G. (2020). Microbio me approaches provide the key to biologically control postharvest pathogens and storability of fruits and vegetables. FEMS Microbiol. Ecol. 96:fiaa119. doi: 10.1093/femsec/fiaa119
Leff, J. W., Noah, F., and Gabriele, B. (2013). Bacterial communities associated with the surfaces of fresh fruits and vegetables. PLoS One 8:e59310. doi: 10.1371/journal.pone.0059310
Li, H., Zhang, Y., Gao, C., Gao, Q., Cheng, Y., Zhao, M., et al. (2022). Mycotoxin production and the relationship between microbial diversity and mycotoxins in Pyrus bretschneideri Rehd cv. Huangguan Pear. Toxins. 14:699. doi: 10.3390/toxins14100699
Li, W. H., Zhang, H. Y., Li, P. X., Tibiru, A. M., Yang, Q. Y., Peng, Y. P., et al. (2016). Biocontrol of postharvest green mold of oranges by hanseniaspora uvarum Y3 in combination with phosphatidylc holine. Biol. Control 103, 30–38. doi: 10.1016/j.biocontrol.2016.07.014
Lopez-Velasco, G., Carder, P. A., Welbaum, G. E., and Ponder, M. A. (2013). Diversity of the spin ach (Spinacia oleracea) spermosphere and phyllosphere bacterial communities. FEMS Microbiol. Lett. 346, 146–154. doi: 10.1111/1574-6968.12216
Ma, F. L., Du, Y. M., Wang, Y., Tong, W., Liu, B. L., Wang, W. H., et al. (2019). Effect of 1-Methylcyclopropene(1-MCP) on quality and chlorophyll maintenance of postharvest 'Yuluxiang' pear. Acta Horticult. Sin. 46, 2299–2308. doi: 10.16420/j.issn.0513-353x.2018-0676
Morales, H. S., Marín,, Ramos, A. J., and Sanchis, V. (2010). Influence of post-harvest technologies applied during cold storage of apples in Penicillium expansum growth and patulin accumulation: a review. Food Control 21, 953–962. doi: 10.1016/j.foodcont.2009.12.016
Prasanna, V., Prabha, T. N., and Tharanathan, R. N. (2007). Fruit ripening phenomena–an overview. Crit. Rev. Food Sci. Nutr. 47, 1–19. doi: 10.1080/10408390600976841
Roberts, R. G. (2005). Alternaria yaliinficiens sp. nov. on Ya Li pear fruit: from interception to iden tification. Plant Dis. 89, 134–145. doi: 10.1094/PD-89-0134
Salmaso, N., Albanese, D., Capelli, C., Boscaini, A., Pindo, M., and Donati, C. (2018). Diversity and cyclical seasonal transitions in the bacterial community in a large and deep perialpine lake. Microb. Ecol. 76, 125–143. doi: 10.1007/s00248-017-1120-x
Shen, Y., Nie, J., Dong, Y., Kuang, L., Li, Y., and Zhang, J. (2018). Compositional shifts in the surface fungal communities of apple fruits during cold storage. Postharvest Biol. Technol. 144, 55–62. doi: 10.1016/j.postharvbio.2018.05.005
Shen, Y. M., Nie, J. Y., Li, Z. X., Li, H. F., Wu, Y. L., Dong, Y. F., et al. (2018). Differentiated surface fungal communities at point of harvest on apple fruits from rural and peri-urban orchards. Sci. Rep. 8, 2165–2112. doi: 10.1038/s41598-017-17436-5
Sun, P. P., Cui, J. C., Jia, X. H., and Wang, W. H. (2017). Complete genome sequence of bacillus velezensis l-1, which has antagonistic activity against pear diseases. Genome Announc. 5:e01271. doi: 10.1128/genomeA.01271-17
Tang, L., Wei, W. L., Zhao, Y. J., Yu, W. P., Wu, Z. Y., Zeng, L., et al. (2022). Analysis of physicochemical characteristics and fungal community diversity during fermentation of industrial radish kimchi. Food Ferment. Ind. 48, 25–31. doi: 10.13995/j.cnki.11-1802/ts.028275
Tian, F., Cai, E. W., Kou, X. H., Fan, G. J., and Li, T. (2021). Surface fungal community diversity change and potential pathogens of Ginkgo biloba seed during cold storage. Food Biosci. 41:100981. doi: 10.1016/j.fbio.2021.100981
Ttpabc, E., Ec, E., Dwsabc, D., Jp, E., and Hpab, C. (2019). Pathogenetic process monitoring and early detection of pear black spot disease caused by alternaria alternata using hyperspectral imaging. Postharvest Biol. Technol. 154, 96–104. doi: 10.1016/j.postharvbio.2019.04.005
Wang, X. D., Ban, S. D., Hu, B. D., Qiu, S. Y., and Zhou, H. X. (2017). Bacterial diversity of Moutai-flavour Daqu based on high-throughput sequencing method. J. Inst. Brew. 123, 138–143. doi: 10.1002/jib.391
Wang, B. T., Yang, W. L., Lei, G., Li, S. S., and Liu, D. H. (2022). Analysis of fungal diversity in rhizosphere soil of konjac based on high-throughput sequencing. Southwest China J. Agric. Sci. 35, 804–811. doi: 10.16213/j.cnki.scjas.2022.4.010
Wang, Y. B., and Zhang, Y. X. (2010). Salicylic acid induces the accumulation of defense-related enzym es in Whangkeumbae pear and protects from pear black spot. Front. Agric. China 4, 215–219. doi: 10.1007/s11703-010-0002-5
Wang, M. Y., Zhao, Q. S., Su, C., and Yang, J. G. (2019). Analysis of the microbial community structure during brewing of Sichuan Xiaoqu baijiu. J. Am. Soc. Brew. Chem. 77, 210–219. doi: 10.1080/03610470.2019.1605033
Xu, S., Lu, W. J., Liu, Y. T., Ming, Z. Y., Liu, Y. J., Meng, R. H., et al. (2016). Structure and div ersity of bacterial communities in two large sanitary landfills in China as revealed by high-throughput sequencing (miseq). Waste Manag. 63, 41–48. doi: 10.1016/j.wasman.2016.07.047
Xue, B., Yu, J. J., Zhang, J. C., Hao, F., Zhang, X. M., Dong, J. H., et al. (2021). Microbial diversity analysis of vineyard son the eastern foothills of the Helan Mountain region using high-th roughput sequencing. Food Sci. Technol. 42:66320. doi: 10.1590/fst.66320
Yu, T., Chen, Y., Lu, H., Zunun, M., Chen, F., Tao, Z., et al. (2012). Effect of cryptococcus laurentii and calcium chloride on control of penicillium expansum and botrytis cinerea infections in pear fruit. Biol. Control 61, 169–175. doi: 10.1016/j.biocontrol.2012.01.012
Zang, J. H., Xu, Y. S., Xia, W. S., Yu, D. W., Gao, P., Jiang, Q. X., et al. (2018). Dynamics and diversity of microbial community succession during fermentation of suan yu, a chinese traditional fermented fish, determined by high throughput sequencing. Food Res. Int. 111, 565–573. doi: 10.1016/j.foodres.2018.05.076
Zhang, S. W., Chen, X., Zhong, Q. D., Zhuang, X. L., and Bai, Z. H. (2019). Microbial community analyses associated with nine varieties of wine grape carposphere based on high-throughput sequencin g. Microorganisms. 7:668. doi: 10.3390/microorganisms7120668
Zhang, Q., Shi, W. C., Zhou, B., Du, H. Y., Xi, L. Q., Zou, M., et al. (2020). Variable characteristics of microbial communities on the surface of sweet cherries under different storage conditions. Postharvest Biol. Technol. 173:111408. doi: 10.1016/j.postharvbio.2020.111408
Zhang, W., Zhao, Y. L., Yang, Z. G., and Wang, L. (2021). Effect of different temperature storage on fruit Q uality and chlorophyll fluorescence parameters of Yuluxiang pears in Xixian County. J. Shanxi Agricul. Sci. 49, 236–242. doi: 10.3969/j.issn.1002-2481.2021.02.23
Keywords: pear, high-throughput sequencing, fungal diversity, low temperature, diversity analysis
Citation: Hou Y, Zhang X, Gao Z, Chen T and Zhang L (2023) Relationships between fungal diversity and fruit quality of Yuluxiang pear during low temperature storage. Front. Microbiol. 14:1132271. doi: 10.3389/fmicb.2023.1132271
Edited by:
Zhihui Chen, University of Dundee, United KingdomReviewed by:
Mohamed Mannaa, Cairo University, EgyptCopyright © 2023 Hou, Zhang, Gao, Chen and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoyu Zhang, eGlhb3l1emhhbmcyMDA1QDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.