 María Teresa Llorente1,2
María Teresa Llorente1,2 Raquel Escudero2Raquel Ramiro1María Antonia Remacha3Rocío Martínez-Ruiz4
Raquel Escudero2Raquel Ramiro1María Antonia Remacha3Rocío Martínez-Ruiz4 Fátima Galán-Sánchez5
Fátima Galán-Sánchez5 Mónica de Frutos6Matilde Elía7Isabel Onrubia8
Mónica de Frutos6Matilde Elía7Isabel Onrubia8 Sergio Sánchez1*
Sergio Sánchez1*- 1Reference and Research Laboratory on Food and Waterborne Bacterial Infections, National Center for Microbiology, Institute of Health Carlos III, Madrid, Spain
- 2Reference and Research Laboratory on Special Pathogens, National Center for Microbiology, Institute of Health Carlos III, Madrid, Spain
- 3Servicio de Microbiología Clínica, Complejo Asistencial Universitario de León, León, Spain
- 4Servicio de Microbiología y Parasitología, Hospital Puerta de Hierro Majadahonda, Majadahonda, Spain
- 5Servicio de Microbiología, Hospital Universitario Puerta del Mar, Cádiz, Spain
- 6Servicio de Microbiología, Hospital Universitario del Río Hortega, Valladolid, Spain
- 7Servicio de Microbiología Clínica, Hospital Universitario de Navarra, Pamplona, Spain
- 8Pediatría, Centro de Salud Valle de la Oliva, Majadahonda, Spain
Background: Enteroaggregative Escherichia coli (EAEC) is increasingly associated with domestically acquired diarrheal episodes in high-income countries, particularly among children. However, its specific role in endemic diarrhea in this setting remains under-recognized and information on molecular characteristics of such EAEC strains is limited. We aimed to investigate the occurrence of EAEC in patients with non-travel related diarrhea in Spain and molecularly characterize EAEC strains associated with illness acquired in this high-income setting.
Methods: In a prospective multicenter study, stool samples from diarrheal patients with no history of recent travel abroad (n = 1,769) were collected and processed for detection of EAEC and other diarrheagenic E. coli (DEC) pathotypes by PCR. An additional case–control study was conducted among children ≤5 years old. Whole-genome sequences (WGS) of the resulting EAEC isolates were obtained.
Results: Detection of DEC in the study population. DEC was detected in 23.2% of patients aged from 0 to 102 years, with EAEC being one of the most prevalent pathotypes (7.8%) and found in significantly more patients ≤5 years old (9.8% vs. 3.4%, p < 0.001). Although not statistically significant, EAEC was more frequent in cases than in controls. WGS-derived characterization of EAEC isolates. Sequence type (ST) 34, ST200, ST40, and ST10 were the predominant STs. O126:H27, O111:H21, and O92:H33 were the predominant serogenotypes. Evidence of a known variant of aggregative adherence fimbriae (AAF) was found in 89.2% of isolates, with AAF/V being the most frequent. Ten percent of isolates were additionally classified as presumptive extraintestinal pathogenic E. coli (ExPEC), uropathogenic E. coli (UPEC), or both, and belonged to clonal lineages that could be specifically associated with extraintestinal infections.
Conclusion: EAEC was the only bacterial enteric pathogen detected in a significant proportion of cases of endemic diarrhea in Spain, especially in children ≤5 years old. In particular, O126:H27-ST200, O111:H21-ST40, and O92:H33-ST34 were the most important subtypes, with all of them infecting both patients and asymptomatic individuals. Apart from this role as an enteric pathogen, a subset of these domestically acquired EAEC strains revealed an additional urinary/systemic pathogenic potential.
1. Introduction
Diarrheal disease is a significant cause of hospitalization and economic losses due to sick leave in developed countries (Guarino et al., 2012; Ridderstedt et al., 2018). The etiological agents include a wide range of bacteria, viruses, and parasites. Among bacterial pathogens, strains of Escherichia coli that cause diarrhea in humans are known collectively as diarrheagenic E. coli (DEC) and traditionally classified into individual pathotypes, with Shiga toxin-producing E. coli (STEC), enteroaggregative E. coli (EAEC), enteropathogenic E. coli (EPEC), enterotoxigenic E. coli (ETEC), and enteroinvasive E. coli (EIEC) being the most important ones (Kaper et al., 2004).
Concretely, EAEC strains colonize the intestinal mucosa via the aggregative adherence fimbriae (AAF), which include five variants (designated I-V) (Nataro et al., 1993; Czeczulin et al., 1997; Bernier et al., 2002; Boisen et al., 2008; Jønsson et al., 2015) and are transcriptionally regulated by the AggR activator (Elias et al., 1999; Boisen et al., 2008). AggR promotes the expression of both chromosomal and plasmid-encoded EAEC virulence factors and is therefore considered as the central regulator of virulence functions in EAEC (Dudley et al., 2006; Harrington et al., 2006). Two examples of genes commonly found in EAEC that are regulated by AggR include aatA, encoding a component of the dispersin transport system, and aaiC, encoding a type VI secretion system (Dudley et al., 2006; Harrington et al., 2006; Petro et al., 2020). Strains harboring the AggR regulon or its components have been termed typical EAEC (Nataro, 2003), and many studies have strongly associated them with diarrhea (Wilson et al., 2001; Pabst et al., 2003; Cohen et al., 2005; Denno et al., 2012). These strains were the focus of our work and therefore, from now on, the term EAEC will specifically refer to “typical EAEC.” Strains showing an aggregative-adherence pattern but not carrying AggR-regulated genes are termed atypical EAEC (Nataro, 2003), and they are considered of uncertain pathogenicity (Tokuda et al., 2010; Boisen et al., 2020), despite having been isolated from food-borne outbreaks of gastrointestinal illness (Itoh et al., 1997). Additionally, EAEC strains often harbor a variable number of serine protease autotransporters of the Enterobacteriaceae (SPATEs) (Boisen et al., 2009).
Although often underdiagnosed, EAEC is frequently detected in both symptomatic and asymptomatic children in developing countries (Rogawski et al., 2017; Manhique-Coutinho et al., 2022) and considered one of the leading causes of travelers’ diarrhea (Estrada-Garcia and Navarro-Garcia, 2012). Additionally, there is increasing evidence that EAEC is also associated with domestically acquired diarrheal episodes in high-income countries, particularly among children (Pabst et al., 2003; Shazberg et al., 2003; Cohen et al., 2005; Nataro et al., 2006; Tobias et al., 2015). However, the specific role of EAEC in endemic diarrhea in industrialized countries remains under-recognized. In Spain, as EAEC infections are not notifiable and no surveillance has been conducted to date, the actual burden of disease is unknown, apart from several studies dealing with the etiology of travelers’ diarrhea (Vargas et al., 1998; Palmeiro et al., 2012). To better understand the importance of EAEC as etiological agent of endemic diarrhea in Spain we undertook a prospective study to investigate its occurrence in 1,769 patients with non-travel related diarrhea. Additionally, we investigated the clinical significance of EAEC infections especially among children ≤5 years old, by comparing EAEC prevalence in children with diarrhea (n = 256) and in healthy controls (n = 133). Furthermore, we performed whole-genome sequencing (WGS) on the resulting isolates (n = 120) to determine the molecular characteristics of EAEC strains associated with illness acquired in this setting.
2. Materials and methods
2.1. Study design and sample collection
A prospective multicenter study was performed from June 2015 to December 2016 in collaboration with five public tertiary hospitals located in the provinces of Madrid (central Spain), Navarra (northern Spain), Cádiz (southern Spain), Valladolid (central-western Spain), and León (north-western Spain). The collaborating laboratories were asked to submit unduplicated fresh stools from patients of any age with diarrhea and no history of recent travel abroad testing negative to other bacterial enteric pathogens after microbiologic examination. Our case definition included patients with diarrhea, either acute (≥3 liquid or semi-liquid stools in 24 h, or at least one with presence of mucus, blood, or pus for up to 2 weeks) or chronic (>4 weeks duration with decreased consistency and increased stool frequency). Cases were recruited from the emergency department, inpatient, and outpatient clinics. The samples were collected according to availability and submitted to the National Center for Microbiology (NCM, Majadahonda, Spain) once a week, up to a minimum of 350 samples per hospital. Before being shipped to NCM, all stools were routinely tested at their respective home laboratory for Salmonella spp., Shigella spp., Campylobacter spp., Yersinia enterocolitica, and Aeromonas spp. (but not for DEC) using conventional microbiological methods.
We conducted an additional case–control study among children ≤5 years old living in one of the provinces included in the prospective prevalence study (Madrid). The study population consisted of all the children ≤5 years old with diarrhea for whom occurrence of DEC had been previously investigated over the prospective prevalence study from June 2015 to February 2016 (cases) (n = 256), and a group of randomly selected children ≤5 years old with no history of diarrhea or use of antibiotics for at least 14 days and no history of recent travel abroad (controls) (n = 133). These unpaired control subjects were recruited from June 2016 to February 2017 during primary care pediatric consultations in a health center belonging to the same health care district than the hospital that had provided the cases.
2.2. Ethical statement
Since the prospective multicenter study was approved as a part of the routine diagnostic practice, neither specific approval of the respective hospital ethics committees nor informed consent from patients was needed. As for control subjects, ethical approval and permission for the study was obtained from the health care district management (Comisión Central de Investigación, Gerencia Asistencial de Atención Primaria, Servicio Madrileño de Salud; date: May 11, 2016/Reference: 03/2016) and written informed consent was obtained from parents/legal guardians.
2.3. Microbiological analysis
Upon receipt, a stool impregnated cotton swab was inoculated in 5 mL of buffered peptone water (BPW, Oxoid, Basingstoke, United Kingdom) and overnight incubated at 37°C. After this non-selective enrichment step, the BPW culture was subcultured on both MacConkey agar (MAC, Becton Dickinson, Franklin Lakes, NJ, United States) and tryptic soy agar (TSA, Becton Dickinson) and overnight incubated at 37°C. A loopful of bacterial growth taken from the first streaking area of the TSA plate was suspended in 0.5 mL of sterile distilled water, boiled for 5 min to release the DNA, and centrifuged at 10,000 rpm for 5 min.
The supernatant was used directly as a template in eight in-house conventional PCR assays for the specific amplification of genes defining each DEC pathotype (Table 1), using DreamTaq DNA Polymerase (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s instructions. An additional gapA-specific PCR was also run concurrently with the diagnostic PCR assays to ensure that all samples had sufficient bacterial DNA present and no PCR inhibition occurred (Table 1). Thermal cycler conditions consisted of 25 cycles of denaturation at 94°C for 30 s, annealing at 56°C for 40 s, and extension at 72°C for 1 min. Our diagnostic criterion for EAEC infection was the presence of aatA gene, considering its historical specificity (Pabst et al., 2003; Beczkiewicz et al., 2019). According to this criterion, the study focused only on typical EAEC. The diagnostic criteria for other DEC infections were as follows: for STEC, the presence of stx1, and/or stx2, and/or stx2f, and possible additional gene eae, with STEC primers targeting the specific subtypes stx1a, stx1c, stx1d, stx2a, stx2b, stx2c, stx2d, stx2e, stx2f, and stx2g; for EPEC, the presence of eae and possible additional gene bfpA, with the absence of bfpA indicating atypical EPEC (aEPEC); for ETEC, the presence of eltA and/or estA; for EIEC, the presence of ipaH.
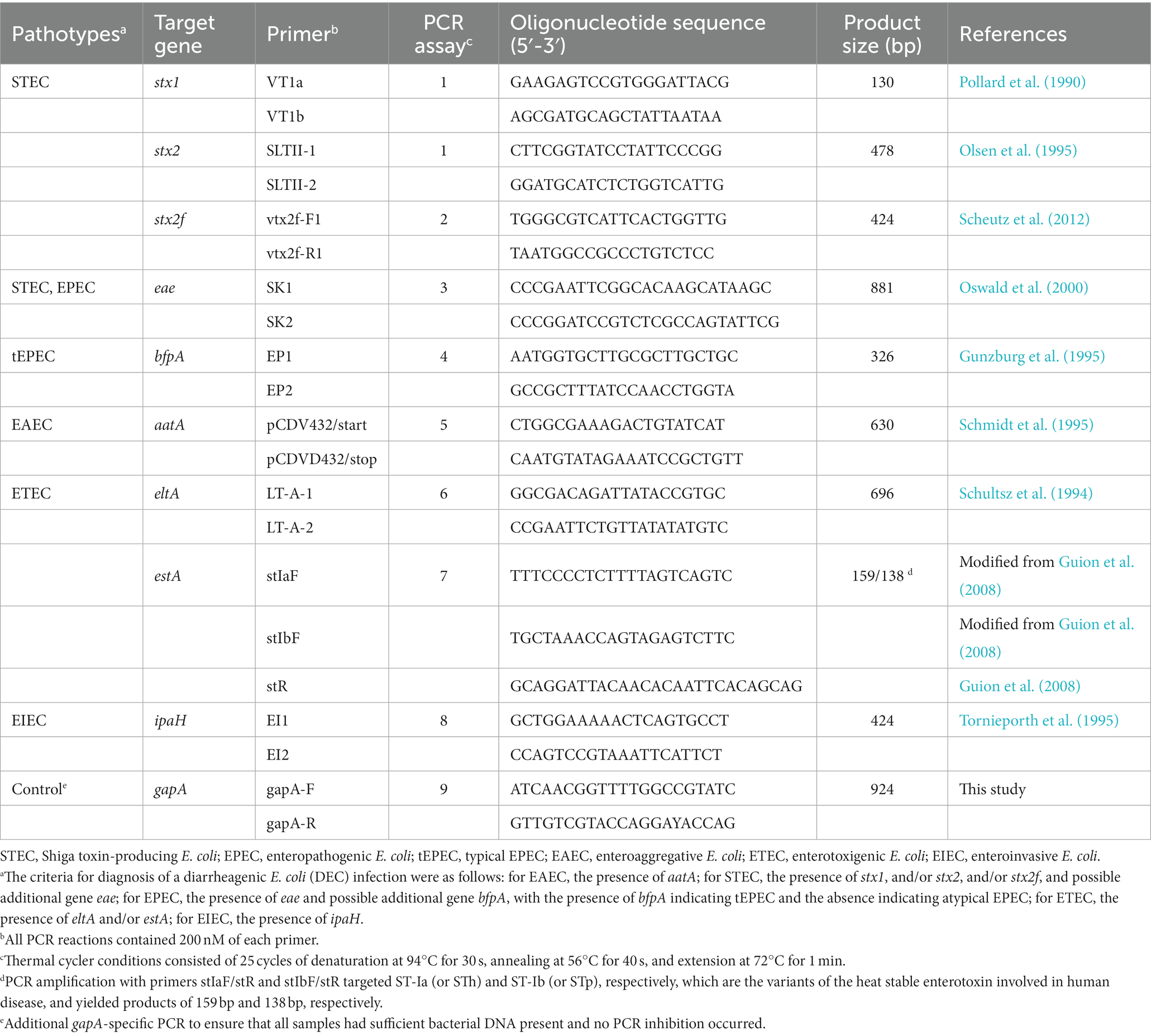
Table 1. Primer pairs and target genes used for detection and isolation of diarrheagenic Escherichia coli pathotypes in patients with endemic diarrhea and asymptomatic controls.
When culture tested EAEC-positive, up to 20 individual E. coli-like colonies obtained from MAC plates were tested by PCR to obtain the isolate, which was further confirmed biochemically as E. coli by the API 20E system (bioMérieux, Marcy l’Etoile, France).
2.4. Whole-genome sequencing
Genomic DNA was purified from the EAEC isolates using the QIAamp DNA Mini Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions. A DNA library was generated using the Nextera XT DNA Sample Preparation Kit (Illumina, San Diego, CA, United States) according to the manufacturer’s instructions and run on a NextSeq 500 (Illumina) for generating paired-end 150 bp reads, aiming at a coverage of at least 200-fold. The reads were trimmed (FastP, 0.23.2) and filtered according to quality criteria (FastQC, 0.11.9), and the quality-filtered reads were de novo assembled by using Unicycler (v0.4.8) (Wick et al., 2017).
2.5. Data analysis and molecular characterization
The O and H serogenotypes (in silico serotypes), virulence genes, sequence types (STs), and acquired antibiotic resistance genes, were identified by uploading the reads to SerotypeFinder 2.0, VirulenceFinder 2.0, MLST 2.0, and ResFinder 4.1, respectively, available on the Center for Genomic Epidemiology (CGE) website.1 The threshold of sequence identity was set to 85% and the percentage of minimum overlapping gene length to 60%. MLST tool used the seven loci (adk, gyrB, fumC, icd, mdh, purA, and recA) scheme. When SerotypeFinder did not predict O antigen it was considered not typeable (ONT). The E. coli phylogroups were determined using the ClermonTyping tool available on the Iame-research Center website.2 The presence of the colonization factor CS22 structural gene (cseA, accession no. AF145205.1) was determined by searching the assembled contigs with BLASTn. The presumptive extraintestinal pathogenic E. coli (ExPEC) status was assigned to those isolates positive for ≥2 of the following virulence genes: papA and/or papC, sfa-focDE, afa-draBC, iutA, and kpsMII (Johnson et al., 2003). For this purpose, isolates were considered positive for afa-draBC if a combination of afaB or nfaE and also afaC was identified by WGS and positive for sfa-focDE if a combination of focC or sfaE and also focI or sfaD was identified (Malberg Tetzschner et al., 2020). Likewise, the uropathogenic E. coli (UPEC) status was assigned to those isolates positive for ≥2 of the following genes: chuA, fyuA, vat, and yfcV (Spurbeck et al., 2012; Malberg Tetzschner et al., 2020).
2.6. Phylogenomic analysis
The 120 EAEC genomes analyzed in this study were compared with 195 previously sequenced EAEC genomes originating from the United Kingdom, Egypt, Kenya, or Peru (Do Nascimento et al., 2017; Ellis et al., 2020; Petro et al., 2020), as well as the EAEC reference genomes 17–2, 042, and 55,989, and six ExPEC reference genomes (Supplementary Table 1) using Snippy 4.6.0 as previously described3. Snippy identified 690,639 conserved SNPs, compared against the reference genome of the E. coli strain IAI39 (accession no. NC_011750.1), that were used to infer a maximum likelihood phylogeny using IQ-Tree 2.1.4 (Minh et al., 2020) with a TVM model and 1,000 bootstrap iterations. The phylogeny was midpoint rooted and annotated with iTOL 6.6 (Letunic and Bork, 2019).
Additionally, a more specific SNP analysis was performed for each of the most important serogenotype-ST combinations identified in the present study, including isolates from both the present and previous studies (Do Nascimento et al., 2017; Ellis et al., 2020; Petro et al., 2020). The analysis was carried out by uploading the reads to CSI Phylogeny 1.4, available on the CGE website, with the following settings: a minimum depth of 10 at SNP positions, a minimum relative depth of 10% at SNP positions, a minimum distance of 10 bp between SNPs (prune), a minimum SNP quality of 30, a minimum read quality of 25, and a minimum Z-score of 1.96. According to KmerFinder 3.2 results, the published genomes of E. coli strains A41 (accession no. NZ_CP028735.1), ESBL 15 (accession no. NZ_CP041678.1), BR1220 (accession no. NZ_CP093068.1), and H3 (accession no. NZ_CP028732.1) were used as a reference for EAEC strains belonging to O126:H27-ST200, O111:H21-ST40, O92:H33-ST34, and O3:H2-ST10, respectively. The percentage of the reference genome covered by all isolates of the same serogenotype-ST combination ranged between 82.3 and 90.3%. From the aligned sequences of concatenated SNPs, we calculated maximum likelihood phylogenetic trees with RAxML 8.2.12 (Stamatakis, 2014) with a GTR model and 1,000 bootstrap iterations. The respective consensus trees were midpoint rooted and annotated with iTOL.
2.7. Sequence availability
FASTQ sequences were deposited in the National Center for Biotechnology Information Short Read Archive under the BioProject PRJNA863489. Accession numbers for each sequence are listed in Supplementary Table 2.
2.8. Statistical analysis
The sample size for determining the DEC prevalence in the prospective multicenter study was calculated using the website tool www.openepi.com, with a confidence level of 95%, a precision value of 3%, and an anticipated frequency of 9% (Nataro et al., 2006). The case–control study was conducted with as many healthy controls fulfilling the inclusion criteria as possible. Proportions were compared by a two-tailed chi-square test or Fisher’s exact test and odds ratios with 95% confidence intervals were determined. A p value <0.05 was considered statistically significant.
3. Results
3.1. Detection of diarrheagenic Escherichia coli in the study population
The prospective study included 1,769 patients (mean age 14.5 years, range 0–102 years, standard deviation (SD) 24.4 years, male 54%), with 68.6% of participants being ≤5 years old (n = 1,213). The case–control study included 256 children with diarrhea (mean age 18.1 months, SD 16.2 months, male 55.1%) and 133 asymptomatic children (mean age 16.4 months, SD 14 months, male 59.4%).
At least one DEC pathotype was detected in 23.2% (410/1,769) of patients and more than one (up to three) was detected in 2.3% (40/1,769) (Table 2). EAEC was one of the pathotypes most commonly found (7.8%, 138/1,769), second only to aEPEC (13.3%, 235/1,769) and followed by STEC (3.6%, 63/1,769), with ETEC and EIEC being anecdotal (0.6%, 10/1,769, and 0.3%, 5/1,769, respectively). The most common DEC co-infection was the combination of EAEC and aEPEC, detected in 62.5% (25/40) of co-infection episodes. There were no statistically significant differences between male and female patients neither in relation to general DEC prevalence nor in relation to EAEC (data not shown). However, DEC infection was found in significantly more patients ≤5 years old (27.7%, 336/1,213, vs. 13.3%, 74/556, p < 0.001), with EAEC and aEPEC infections being particularly associated with this age group (9.8%, 119/1,213, vs. 3.4%, 19/556, p < 0.001 for EAEC; 16.2%, 197/1,213, vs. 6.8%, 38/556, p < 0.001 for aEPEC; 2.1%, 25/1,213, vs. 0%, 0/556, p < 0.001 for EAEC+aEPEC co-infection) (Table 2).
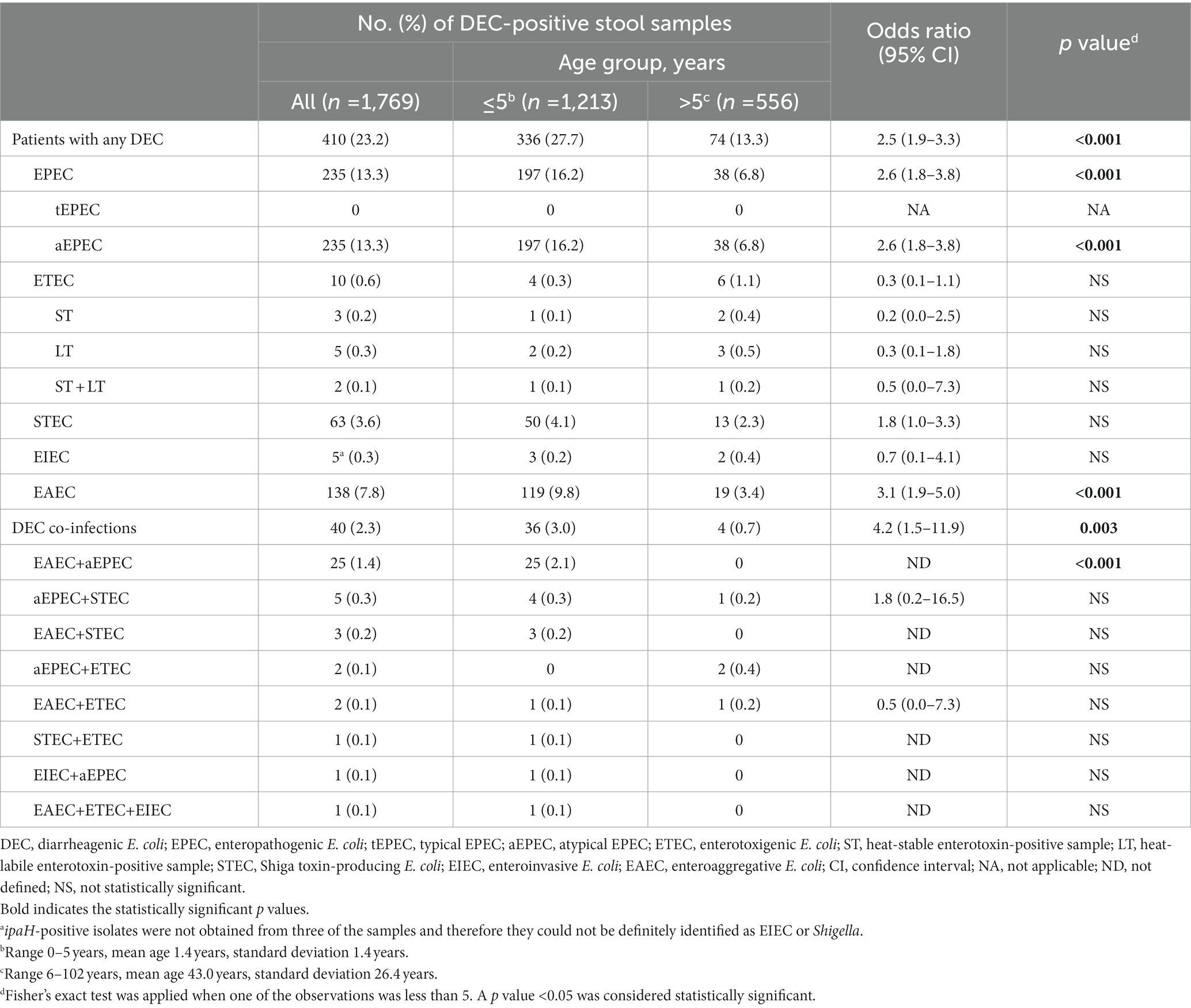
Table 2. Detection of diarrheagenic Escherichia coli pathotypes among patients with endemic diarrhea and differences in prevalence by age.
In the case–control study, conducted only with children ≤5 years old, EAEC was more frequently detected in cases than in controls (12.5%, 32/256, vs. 10.5%, 14/133), although this difference was not statistically significant (Table 3). Only aEPEC and DEC co-infections in general were found in significantly more cases than controls (16.8%, 43/256, vs. 8.3%, 11/133, p = 0.021 for aEPEC; 5.5%, 14/256, vs. 0%, 0/133, p = 0.005 for DEC co-infections).
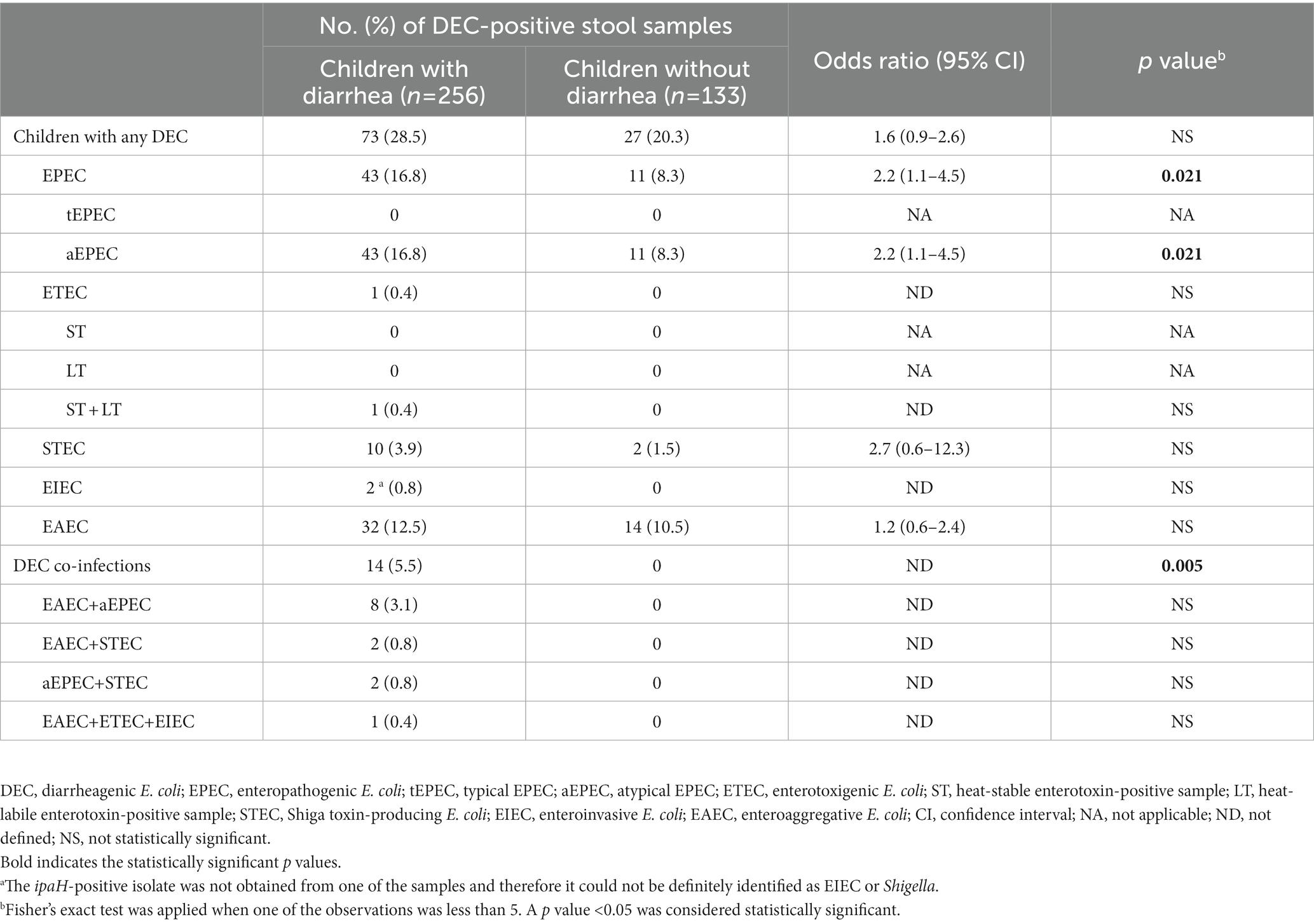
Table 3. Detection of diarrheagenic Escherichia coli pathotypes among patients with endemic diarrhea and asymptomatic controls ≤5 years old and association of specific pathotypes with diarrhea.
3.2. WGS-derived characterization of EAEC isolates
EAEC was isolated from 110 (79.7%) of the 138 EAEC-positive fecal samples from patients and 10 (71.4%) of the 14 EAEC-positive samples from asymptomatic individuals. Characteristics of the 120 resulting EAEC isolates are summarized in Supplementary Table 2. Only four of the eight E. coli phylogroups sensu stricto recognized so far (A, B1, B2, C, D, E, F, and G) (Clermont et al., 2019) were represented among the 120 EAEC isolates. The predominant phylogroups were B1 (47%) and A (44%), although D (8%) and B2 (1%) were also identified (Supplementary Table 2). The presumptive ExPEC and/or UPEC status was found only in isolates belonging to phylogroups A (58%), D (33%), and B2 (9%). All isolates negative for both AAF and CS22 belonged to phylogroup A. Thirteen different STs and 26 different serogenotypes were found. The most common ST was ST34 (27%), followed by ST200 (20%), ST40 (18%), and ST10 (17%) (Figure 1A). The predominant serogenotype was O126:H27 (18%), followed by O111:H21 (17%) and O92:H33 (14%) (Figure 1B). Notably, three of the six serotypes defining the EAEC prototype strains were found in our collection (JM221 serotype O92:H33, 042 serotype O44:H18, and 17–2 serotype O3:H2) (Boisen et al., 2020).
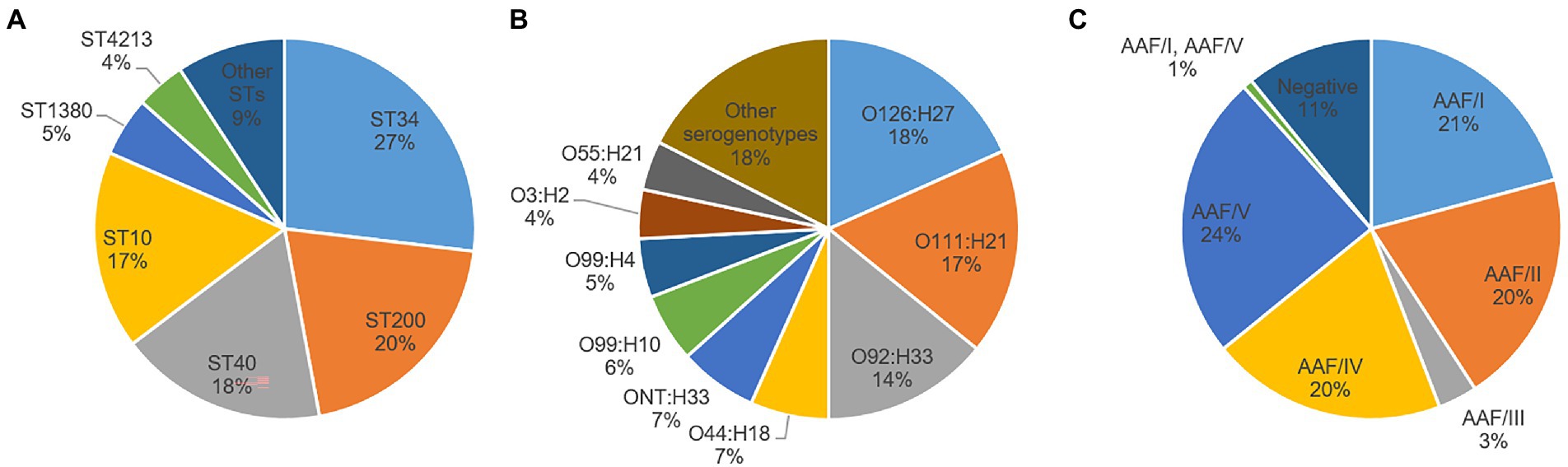
Figure 1. Distribution of different sequence types (ST) (A), serogenotypes (B), and aggregative adherence fimbriae (AAF) variants (C) among enteroaggregative Escherichia coli isolates. Other serogenotypes and STs (with less than five isolates each) are summarized in Supplementary Table 2. When O antigen was not predicted, it was considered not typeable (ONT).
Regarding the distribution of EAEC-associated putative virulence genes, the resulting EAEC isolates are typical EAEC in the formal definition, as they harbored at least aatA gene, per study protocol. Apart from aatA, the dispersin gene aap was the most commonly detected EAEC-associated gene (100%), followed by aggR (90.8%), the genes encoding the proteins ORF3 and ORF4 (89.2% each), aaiC (85%), the SPATE gene pic (83.3%), and the AggR-activated regulator aar (71.7%). Other SPATE genes detected were sat (41.7%), sepA (40.8%), and pet (20%) (Table 4). No statistically significant differences could be observed in the presence and frequency of specific putative virulence genes between isolates obtained from patients and those from controls (Table 4). Evidence of a known AAF variant was found in 89.2% of the isolates, with AAF/V being the most frequently observed (24%), followed by AAF/I (21%), AAF/II (20%), AAF/IV (20%), and AAFIII (3%) (Table 4, Figure 1C). Remarkably, the sequence of one isolate (4519–15) contained the genes for both AAF/I (aggA, aggB, aggC, and aggD) and AAF/V (agg3C, agg3D, and agg5A) variants (Supplementary Table 2). Thirteen isolates (10.8%) were negative for any genes attributed to the five known AAF variants, despite harboring aggR and/or other AggR-regulated genes. Notably, one of these AAF-negative isolates (2018–2015) had a gene identical (100% homologous) to cseA (Supplementary Table 2), indicative of the presence of the non-fimbrial ETEC colonization factor CS22, instead of AAF.
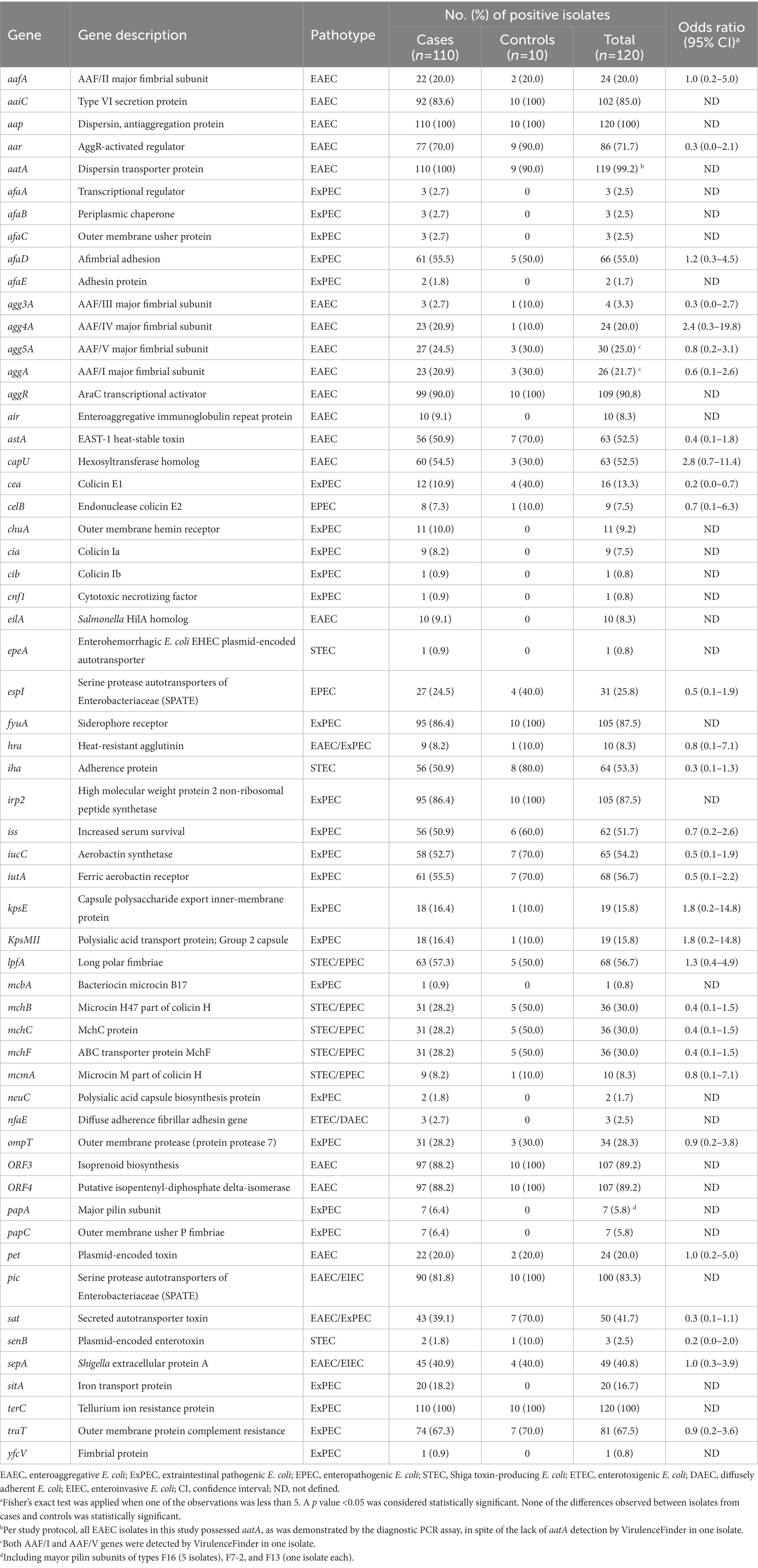
Table 4. Distribution of putative virulence genes among the 120 enteroaggregative Escherichia coli isolates obtained from patients with endemic diarrhea (cases) and asymptomatic individuals (controls).
Apart from EAEC-associated putative virulence genes, a substantial number of genes typically associated with other E. coli pathotypes were detected (Table 4). In particular, 27 of the 44 genes associated with ExPEC included in the CGE E. coli virulence gene database were detected among EAEC isolates. The most frequent ExPEC-associated genes were terC (100%), fyuA (87.5%), irp2 (87.5%), traT (67.5%), iutA (56.7%), afaD (55%), iucC (54.2%), iss (51.7%), and ompT (28.3%). Other relevant ExPEC-associated genes detected were kpsMII (15.8%), chuA (9.2%), papA (5.8%), papC (5.8%), afaB (2.5%), afaC (2.5%), and yfcV (0.8%). According to two main operational definitions (Johnson et al., 2003; Spurbeck et al., 2012), nine EAEC isolates were classified as presumptive ExPEC, one isolate was classified as UPEC, and two isolates were classified as both ExPEC and UPEC (Supplementary Table 2). Therefore, 10% of EAEC isolates in our collection revealed an additional urinary/systemic pathogenic potential.
Regarding the acquired antibiotic resistance genes profiles, 40 (33.3%) of 120 isolates harbored genes conferring resistance to at least one antibiotic category and 28 (23.3%) harbored genes conferring resistance to three or more categories and were therefore considered multidrug resistant (MDR) based on the WGS prediction (Supplementary Table 2). There were no common resistance determinant profiles and the highest number of isolates that shared the same genotypic resistance determinant profile (aph(3″)-Ib/aph(6)-Id/sul2/blaTEM-1C) was seven. As for the presence of extended-spectrum beta-lactamases (ESBLs), conferring resistance to third generation cephalosporins, the blaCTX-M-15 and blaCTX-M-14 genes were harbored by two and one isolate, respectively (Supplementary Table 2).
4. Discussion
We undertook a large prospective study of diarrheal disease at five hospitals located in different Spanish provinces widely distributed geographically, with the purpose of determining the role of EAEC among patients seeking medical care. The study demonstrated that EAEC is frequently detected among patients with diarrhea in Spain (7.8%), especially in children ≤5 years old, among which EAEC prevalence reached 9.8%. This finding corresponds well with previous studies demonstrating a remarkable predisposition to EAEC infection in children ≤5 years of age and suggesting that the prevalence and significance of EAEC infections depend on age (Pabst et al., 2003; Cohen et al., 2005). Although it is possible that some EAEC detected in this study are not pathogenic or represent colonization rather than infection, the presence of other more established bacterial enteric pathogens (e.g., Salmonella spp., Campylobacter spp.) was ruled out per study protocol, and other clinically relevant DEC pathotypes (STEC, ETEC, and EIEC) were co-detected in only 4.3% of cases of EAEC infection. Furthermore, 90% of our EAEC isolates met the new molecular definition of EAEC comprising E. coli strains harboring AggR and its adhesin dependent factors (AAF(I-V) or CS22), recently proposed by Boisen et al. (2020), and could thus be considered as true EAEC. Therefore, EAEC was the only bacterial enteric pathogen detected in a significant proportion of cases of diarrhea, none of which had a history of recent travel abroad, thus suggesting that EAEC is an important domestically acquired bacterium responsible for endemic diarrhea in Spain. This proportion of patients with non-travel related diarrhea who were demonstrated to be infected with EAEC was unexpectedly high, as EAEC prevalence is expected to be higher among travelers from industrialized countries visiting less-developed regions. However, our findings are supported by studies conducted in other high-income countries that also showed relatively frequent detection of EAEC among patients with diarrhea (Supplementary Table 3), with detection rates ranging from 1.9 to 5.9% in the general population (Wilson et al., 2001; Nataro et al., 2006; Hardegen et al., 2010; Tam et al., 2012; Cybulski et al., 2018; Hebbelstrup Jensen et al., 2018) and up to 11.9% in children ≤5 years old (Pabst et al., 2003; Nataro et al., 2006; Tobias et al., 2015). In particular, Pabst et al. (2003) and Cohen et al. (2005) detected EAEC from children ≤5 years with diarrhea significantly more frequently than from healthy children (11.9% vs. 2.2 and 9.2% vs. 3.3%, respectively), although the association of EAEC with diarrhea did not achieve statistical significance in our case–control study. On the contrary, as expected, our detection rates are substantially lower than those generally reported in developing countries, with EAEC prevalences up to 39% in this setting (Gonzalez et al., 1997; Okeke et al., 2000; Modgil et al., 2021; Manhique-Coutinho et al., 2022).
Although there were 26 different serogenotypes among the 120 isolates, 82% were restricted to only nine serogenotypes, very homogeneous with respect to ST, with some particularly common serogenotype-ST combinations, such as O126:H27-ST200, O111:H21-ST40, and O92:H33-ST34, comprising 50% of isolates. This finding is in contrast to previous studies reporting a higher diversity in terms of serotyping and MLST among EAEC clinical isolates from the United States or the United Kingdom (Do Nascimento et al., 2017; Beczkiewicz et al., 2019), probably because such studies did not rule out travel-related infections. These same serotypes are among the most frequently reported in EAEC strains from other high-income countries (Shazberg et al., 2003; Tobias et al., 2015; Hebbelstrup Jensen et al., 2016; Imuta et al., 2016; Do Nascimento et al., 2017; Beczkiewicz et al., 2019), and even linked to outbreaks of gastroenteritis (Yatsuyanagi et al., 2002; Harada et al., 2007; Scavia et al., 2008; Dallman et al., 2012), although rarely identified among EAEC strains from developing countries (Boisen et al., 2020; Petro et al., 2020). Indeed, in the whole-genome phylogeny including 324 EAEC genomes from developing (Egypt, Kenya, and Peru) and high-income (Spain and the United Kingdom) countries (Figure 2), isolates of the most common serogenotype-ST combinations clustered together in independent groups consisting exclusively (O126:H27-ST200 and O111:H21-ST40) or almost exclusively (O92:H33-ST34) of isolates originating from high-income countries. Moreover, as revealed in the whole-genome phylogenies specific for each of these subtypes (Supplementary Figures 1–3), isolates from the United Kingdom were interleaved with those from Spain belonging to the same serogenotype-ST combination. As isolates in the present study were not travel-related, this suggests that O126:H27-ST200, O111:H21-ST40, and O92:H33-ST34 are the most important domestically acquired EAEC subtypes in Spain, and probably also in other high-income countries. In particular, O111:H21-ST40 strains have been recently proposed to have a higher intrinsic potential to cause diarrheal disease in the United Kingdom (Ellis et al., 2020). Nevertheless, we found O111:H21-ST40, and all the aforementioned common EAEC subtypes, both in isolates obtained from patients and those from asymptomatic controls. Indeed, most of the isolates originating from asymptomatic carriage in the present study showed combinations of phylogroup, serogenotype, ST, virulence genes, and antibiotic resistance genes already found among clinical isolates (Supplementary Table 2). This similarity between isolates from cases and controls was also revealed in the O126:H27-ST200, O111:H21-ST40, and O92:H33-ST34 specific phylogenies (Supplementary Figures 1–3), in which isolates from controls were interleaved with those from cases belonging to the same serogenotype-ST combination. Although these findings could be influenced by the scarce number of isolates obtained from asymptomatic controls in this study (n = 10), they suggest that the same EAEC strains infected both patients and asymptomatic individuals. This is in contrast to the hypothesis that EAEC strains isolated from patients with diarrhea would belong to different subtypes and/or harbor putative virulence factors distinct from, or more commonly than, those isolated from asymptomatic controls (Boisen et al., 2012, 2020).
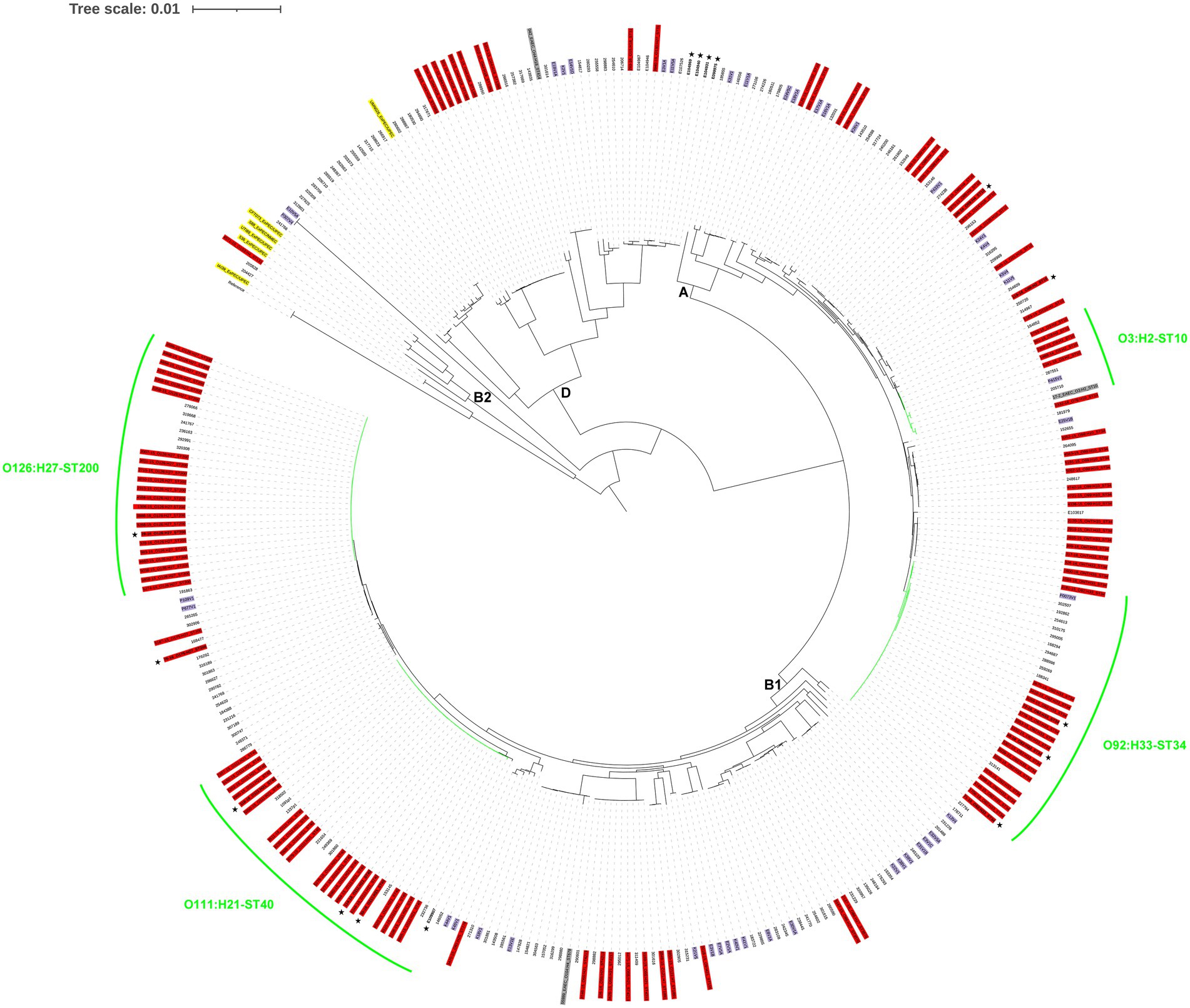
Figure 2. Phylogenomic analysis of the enteroaggregative Escherichia coli genomes. The whole-genome phylogeny was constructed from 690,639 conserved SNP sites per genome that were identified compared against the reference genome of the E. coli strain IAI39 (GenBank accession no. NC_011750.1). The isolates from Spain (this study) are colored in red, those from Egypt, Kenya, or Peru are colored in purple, and those from the United Kingdom are not colored. EAEC and ExPEC reference genomes are colored in gray and yellow, respectively. Isolates obtained from asymptomatic controls are in bold and indicated by a star in the outer ring of labels. The most important serogenotype-ST combinations identified in this study are highlighted in green. The E. coli phylogroups are designated by letters (A, B1, B2, and D) on the interior of the phylogeny based on the inclusion of both EAEC isolates sequenced in this study and reference strains. The tree scale indicates the distance of 0.01 nucleotide changes per site.
The molecular characterization of isolates together with their origin and collection date could suggest possible unnoticed episodes of transmission of the most important domestically acquired EAEC subtypes (Supplementary Table 2) and this could be assessed from the whole-genome phylogenies specific for each of these subtypes. According to criteria proposed by Pightling et al. (2018) for interpreting WGS analyses of foodborne bacteria for outbreak investigations, monophyletic groups of E. coli isolates with a median pairwise distance of 20 or fewer SNPs, a bootstrap support of 90 or higher, and some epidemiological evidence support transmission episodes. In this study, such analyses revealed four possible episodes of EAEC O126:H27-ST200 transmission involving 2–6 patients (Supplementary Figure 1), one possible episode of EAEC O111:H21-ST40 transmission involving two patients (Supplementary Figure 2), and two possible episodes of EAEC O92:H33-ST34 transmission involving 2–3 patients (Supplementary Figure 3).
AAF/V was the most prevalent variant in our collection, as previously reported in Denmark (Hebbelstrup Jensen et al., 2017), with 70% of our AAF/V-harboring isolates belonging to the predominant EAEC subtype O111:H21-ST40 and thus explaining its predominance in our setting. Likewise, 92% of our AAF/II-harboring isolates and 68% of our AAF/I-harboring isolates belonged to the predominant EAEC subtypes O126:H27-ST200 and O92:H33-ST34, respectively, also explaining their predominance in our setting. Of particular interest is the frequent detection of AAF/IV in our isolates, all of them lacking aar and harboring sepA, as such strains have been recently proposed to be more diarrheagenic than other EAEC (Boisen et al., 2020; Petro et al., 2020). However, certain AAF/IV-harboring isolates in our study showed what appears to be a novel AAF/IV fimbrial cluster where the minor pilin subunit gene agg4B has been replaced by afaD. In particular, this apparently new organization of AAF/IV was found in all O55:H21-ST4213 isolates (n = 5), all O44:H18-ST2959 isolates (n = 2), all O121:H27-ST1891 isolates (n = 2), and some O99:H4-ST10 isolates (n = 3). This finding was also confirmed in five O55:H21-ST4213 isolates and one O44:H18-ST2959 isolate originating from the United Kingdom (Do Nascimento et al., 2017; Supplementary Table 4), thus supporting the idea that the epidemiological scenario of endemic EAEC infections would be very similar in different industrialized countries. The identification and characterization of the genetic environment of this apparently novel AAF/IV fimbrial cluster warrants further investigation. Notably, one isolate was found to harbor the genes for both AAF/I and AAF/V, a phenomenon described previously only for AAF/III and AAF/V (Jønsson et al., 2017; Petro et al., 2020). Again, the identification and characterization of the genetic environment of both AAF variants in this particular EAEC isolate warrants further investigation. One of the isolates without a known AAF variant harbored the cseA gene, indicative of the presence of the non-fimbrial ETEC colonization factor CS22 (Pichel et al., 2000), recently identified in strains lacking an identifiable AAF but harboring different putative EAEC virulence factors and being considered typical EAEC by genomic criteria (Boisen et al., 2020; Petro et al., 2020). This cseA-positive isolate belonged to O9:H21-ST155, which has been recently identified among CS22-like harboring EAEC strains originating from Kenya (Petro et al., 2020) and Mozambique (Boisen et al., 2020).
Apart from its role as an enteric pathogen, EAEC has emerged as a causative agent of urinary tract infection (UTI) and bacteremia in the last years (Boll et al., 2020; Mandomando et al., 2020). In particular, phylogroup A and AAF/I have been associated with uropathogenicity and AAF/V with bacteremia (Nunes et al., 2017; Mandomando et al., 2020). In the present study, 11 EAEC isolates were classified as presumptive ExPEC and four of them specifically belonged to serotype O3:H2, phylogroup A, and ST10 and harbored AAF/I. Additionally, we analyzed four previously sequenced EAEC O3:H2-ST10 genomes, including the EAEC reference strain 17–2, and three of them were also classified as presumptive ExPEC (Supplementary Table 5). It should be noted that EAEC O3:H2-ST10 isolates classified as ExPEC consistently harbored AAF/I (with the only exception of one isolate harboring AAF/V), whereas isolates not classified as ExPEC harbored AAF/III and clustered together, far from the ExPEC isolates harboring AAF/I and AAF/V, in the specific phylogeny (Supplementary Figure 4), thus suggesting the importance of AAF/I in extraintestinal EAEC infections. Although the EAEC prototype strain 17–2 had been previously proposed to present some ExPEC/UPEC characteristics (Gomes et al., 1995; Schüroff et al., 2021), to the best of our knowledge, this is the first study to reveal that phylogroup A EAEC O3:H2-ST10 harboring AAF/I is a clonal lineage of EAEC that could be specifically associated with extraintestinal infections. Furthermore, one of the two EAEC isolates classified as ExPEC/UPEC belonged to serotype O153:H4, phylogroup B2, and ST131 and harbored AAF/V and fimH27 (data not shown). It was the only EAEC isolate that belonged to phylogroup B2 in our collection and clustered together with ExPEC/UPEC reference strains belonging to phylogroup B2 but far from the rest of diarrheagenic EAEC isolates and reference strains (Figure 2), thus supporting the idea that authentic enteric and urinary/systemic pathogens can be found among strains meeting the definition of EAEC (Boisen et al., 2020). Indeed, the ST131 H27 sublineage is a novel subclone of E. coli ST131 that has acquired the EAEC diarrheagenic phenotype, spread across multiple continents, and caused multiple outbreaks of community-acquired bacteremia and recurrent UTIs (Boll et al., 2020; Mandomando et al., 2020).
Multidrug-resistance defined as antibiotic resistance to at least three antibiotic categories (Magiorakos et al., 2012) is widespread among foodborne and waterborne enteric pathogens, including EAEC (Hebbelstrup Jensen et al., 2014, 2016; Do Nascimento et al., 2017; Beczkiewicz et al., 2019; Boisen et al., 2020). In our study, 23.3% of the EAEC isolates were considered MDR based on the WGS prediction, and the majority of them originated from children ≤5 years old. As expected, this level of MDR was much lower than that detected in previous studies conducted in developing countries, with MDR detection rates close to 80% (Boisen et al., 2020). However, this finding is in contrast to previous studies also based on the WGS prediction of antibiotic resistance and conducted in high-income countries like the United Kingdom and the United States (Do Nascimento et al., 2017; Beczkiewicz et al., 2019), with MDR detection rates of 56.8 and 51.6%, respectively, again probably because such studies did not rule out travel-associated infections. Of special concern are the abundant ESBL production and the increased resistance to quinolones in EAEC strains (Herrera-León et al., 2015; Imuta et al., 2016; Guiral et al., 2019). In particular, the presence of CTX-M ESBL variants (blaCTX-M-15 and blaCTX-M-14 genes) was detected only in 2.5% of EAEC isolates obtained from cases of endemic diarrhea in Spain. Again, this finding is in contrast to the 20% detected among EAEC isolates from patients with gastrointestinal symptoms in the United Kingdom, and this is probably due to the extremely high percentage of patients reporting travel abroad within 7 days of onset of symptoms in that study (Do Nascimento et al., 2017). While treatment of EAEC infection is not based on antibiotics in the majority of cases, as many EAEC infections are self-limited, evaluating antibiotic susceptibility is important in cases where antibiotic use is clinically indicated.
Our study had several strengths. It is the first Spanish study to explore the role of EAEC in endemic diarrhea and one of the largest studies conducted in an industrialized country to date. As samples were collected from five provinces widely distributed geographically, our results might be representative of the whole country. Unlike most previous studies, we ruled out travel-related diarrheal episodes and those in which other bacterial pathogens were present. We generated one of the most complete characterizations of EAEC strains associated with illness acquired in industrialized countries to date. However, it also had several limitations. Detection of EAEC was based on PCR amplification of a well-known EAEC target but functional testing using the Hep-2 adherence assay was not performed. Although the adherence test remains the “gold standard” for diagnosing EAEC infection, it is resource intensive and requires strict adherence to protocol and specialized facilities. This issue could have underestimated the EAEC prevalence in our study and could be especially significant for isolates negative for both AAF and CS22 not meeting the new molecular definition of EAEC. It was not possible to elucidate the exact etiology of the disease outcome, as samples were not tested for the presence of Clostridioides difficile toxins, parasites, or viruses. The scarce number of control subjects fulfilling the inclusion criteria may have compromised the accuracy of some of our results. We did not collect comprehensive data on symptoms, treatments, outcomes, or risk factors. Finally, phenotypic resistance profile information was not available as conventional antimicrobial susceptibility testing was not performed.
5. Concluding remarks
EAEC was the only bacterial enteric pathogen detected in a significant proportion of cases of endemic diarrhea in Spain, especially in children ≤5 years old. In particular, O126:H27-ST200, O111:H21-ST40, and O92:H33-ST34 were the most important subtypes, with all of them infecting both patients and asymptomatic individuals. A subset of these domestically acquired EAEC strains were additionally classified as ExPEC, UPEC, or both, and belonged to clonal lineages that could be specifically associated with extraintestinal infections, thus revealing an additional urinary/systemic pathogenic potential. These data highlight the convenience of routinely testing for EAEC especially for children ≤5 years old with diarrheal disease and those patients in which no other pathogen can be identified.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethics statement
The studies involving human participants were reviewed and approved by Comisión Central de Investigación, Gerencia Asistencial de Atención Primaria, Servicio Madrileño de Salud. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author contributions
SS and RE conceived and designed the study. Material preparation, data collection and analysis were performed by MR, RM-R, FG-S, MF, ME, IO, RR, and ML. The first draft of the manuscript was written by SS and all authors commented on previous versions of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Institute of Health Carlos III, Spanish Ministry of Economy and Competitiveness (MPY-1042/14, PI14CIII/00051, and PI18CIII/00043) and the SHARP Joint Action (2019 – March 2023) co-funded by the Health Programme of the European Union. SS performed this work while under a research contract from the Miguel Servet program from the Spanish Ministry of Economy and Competitiveness (CP13/00237).
Acknowledgments
The authors thank A. Zaballos and M. P. Jiménez (Genomic Unit, ISCIII) for technical support in the whole-genome sequencing, I. Cuesta and S. Monzón (Bioinformatics Unit, ISCIII) for quality assessment and pre-processing of raw sequencing reads, de novo assembly, and phylogenomic analysis, and A. J. Martín-Galiano (National Center for Microbiology, ISCIII) for BLAST screening. The authors also thank D. Carmena (National Center for Microbiology, ISCIII) for critical review of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1120285/full#supplementary-material
Footnotes
References
Beczkiewicz, A., Cebelinski, E., Decuir, M., Lappi, V., Wang, X., Smith, K., et al. (2019). High relative frequency of enteroaggregative Escherichia coli among patients with reportable enteric pathogens, Minnesota, 2016-2017. Clin. Infect. Dis. 69, 473–479. doi: 10.1093/cid/ciy890
Bernier, C., Gounon, P., and Le Bouguenec, C. (2002). Identification of an aggregative adhesion fimbria (AAF) type III-encoding operon in enteroaggregative Escherichia coli as a sensitive probe for detecting the AAF-encoding operon family. Infect. Immun. 70, 4302–4311. doi: 10.1128/IAI.70.8.4302-4311.2002
Boisen, N., Østerlund, M. T., Joensen, K. G., Santiago, A. E., Mandomando, I., Cravioto, A., et al. (2020). Redefining enteroaggregative Escherichia coli (EAEC): genomic characterization of epidemiological EAEC strains. PLoS Negl. Trop. Dis. 14:e0008613. doi: 10.1371/journal.pntd.0008613
Boisen, N., Ruiz-Perez, F., Scheutz, F., Krogfelt, K. A., and Nataro, J. P. (2009). Short report: high prevalence of serine protease autotransporter cytotoxins among strains of enteroaggregative Escherichia coli. Am. J. Trop. Med. Hyg. 80, 294–301. doi: 10.4269/ajtmh.2009.80.294
Boisen, N., Scheutz, F., Rasko, D. A., Redman, J. C., Persson, S., Simon, J., et al. (2012). Genomic characterization of enteroaggregative Escherichia coli from children in Mali. J. Infect. Dis. 205, 431–444. doi: 10.1093/infdis/jir757
Boisen, N., Struve, C., Scheutz, F., Krogfelt, K. A., and Nataro, J. P. (2008). New adhesin of enteroaggregative Escherichia coli related to the Afa/Dr/AAF family. Infect. Immun. 76, 3281–3292. doi: 10.1128/IAI.01646-07
Boll, E. J., Overballe-Petersen, S., Hasman, H., Roer, L., Ng, K., Scheutz, F., et al. (2020). Emergence of enteroaggregative Escherichia coli within the ST131 lineage as a cause of extraintestinal infections. MBio 11. doi: 10.1128/mBio.00353-20
Clermont, O., Dixit, O. V. A., Vangchhia, B., Condamine, B., Dion, S., Bridier-Nahmias, A., et al. (2019). Characterization and rapid identification of phylogroup G in Escherichia coli, a lineage with high virulence and antibiotic resistance potential. Environ. Microbiol. 21, 3107–3117. doi: 10.1111/1462-2920.14713
Cohen, M. B., Nataro, J. P., Bernstein, D. I., Hawkins, J., Roberts, N., and Staat, M. A. (2005). Prevalence of diarrheagenic Escherichia coli in acute childhood enteritis: a prospective controlled study. J. Pediatr. 146, 54–61. doi: 10.1016/j.jpeds.2004.08.059
Cybulski, R. J., Bateman, A. C., Bourassa, L., Bryan, A., Beail, B., Matsumoto, J., et al. (2018). Clinical impact of a multiplex gastrointestinal polymerase chain reaction panel in patients with acute gastroenteritis. Clin. Infect. Dis. 67, 1688–1696. doi: 10.1093/cid/ciy357
Czeczulin, J. R., Balepur, S., Hicks, S., Phillips, A., Hall, R., Kothary, M. H., et al. (1997). Aggregative adherence fimbria II, a second fimbrial antigen mediating aggregative adherence in enteroaggregative Escherichia coli. Infect. Immun. 65, 4135–4145. doi: 10.1128/iai.65.10.4135-4145.1997
Dallman, T., Smith, G. P., O’Brien, B., Chattaway, M. A., Finlay, D., Grant, K. A., et al. (2012). Characterization of a verocytotoxin-producing enteroaggregative Escherichia coli serogroup O111:H21 strain associated with a household outbreak in Northern Ireland. J. Clin. Microbiol. 50, 4116–4119. doi: 10.1128/JCM.02047-12
Denno, D. M., Shaikh, N., Stapp, J. R., Qin, X., Hutter, C. M., Hoffman, V., et al. (2012). Diarrhea etiology in a pediatric emergency department: a case control study. Clin. Infect. Dis. 55, 897–904. doi: 10.1093/cid/cis553
Do Nascimento, V., Day, M. R., Doumith, M., Hopkins, K. L., Woodford, N., Godbole, G., et al. (2017). Comparison of phenotypic and WGS-derived antimicrobial resistance profiles of enteroaggregative Escherichia coli isolated from cases of diarrhoeal disease in England, 2015-16. J. Antimicrob. Chemother. 72, 3288–3297. doi: 10.1093/jac/dkx301
Dudley, E. G., Thomson, N. R., Parkhill, J., Morin, N. P., and Nataro, J. P. (2006). Proteomic and microarray characterization of the AggR regulon identifies a pheU pathogenicity island in enteroaggregative Escherichia coli. Mol. Microbiol. 61, 1267–1282. doi: 10.1111/j.1365-2958.2006.05281.x
Elias, W. P., Czeczulin, J. R., Henderson, I. R., Trabulsi, L. R., and Nataro, J. P. (1999). Organization of biogenesis genes for aggregative adherence fimbria II defines a virulence gene cluster in enteroaggregative Escherichia coli. J. Bacteriol. 181, 1779–1785. doi: 10.1128/JB.181.6.1779-1785.1999
Ellis, S. J., Crossman, L. C., McGrath, C. J., Chattaway, M. A., Holken, J. M., Brett, B., et al. (2020). Identification and characterisation of enteroaggregative Escherichia coli subtypes associated with human disease. Sci. Rep. 10:7475. doi: 10.1038/s41598-020-64424-3
Estrada-Garcia, T., and Navarro-Garcia, F. (2012). Enteroaggregative Escherichia coli pathotype: a genetically heterogeneous emerging foodborne enteropathogen. FEMS Immunol. Med. Microbiol. 66, 281–298. doi: 10.1111/j.1574-695X.2012.01008.x
Gomes, T. A., Abe, C. M., and Marques, L. R. (1995). Detection of HeLa cell-detaching activity and alpha-hemolysin production in enteroaggregative Escherichia coli strains isolated from feces of Brazilian children. J. Clin. Microbiol. 33:3364. doi: 10.1128/jcm.33.12.3364-3364.1995
Gonzalez, R., Diaz, C., Marino, M., Cloralt, R., Pequeneze, M., and Perez-Schael, I. (1997). Age-specific prevalence of Escherichia coli with localized and aggregative adherence in Venezuelan infants with acute diarrhea. J. Clin. Microbiol. 35, 1103–1107. doi: 10.1128/jcm.35.5.1103-1107.1997
Guarino, A., Dupont, C., Gorelov, A. V., Gottrand, F., Lee, J. K., Lin, Z., et al. (2012). The management of acute diarrhea in children in developed and developing areas: from evidence base to clinical practice. Expert. Opin. Pharmacother. 13, 17–26. doi: 10.1517/14656566.2011.634800
Guion, C. E., Ochoa, T. J., Walker, C. M., Barletta, F., and Cleary, T. G. (2008). Detection of diarrheagenic Escherichia coli by use of melting-curve analysis and real-time multiplex PCR. J. Clin. Microbiol. 46, 1752–1757. doi: 10.1128/JCM.02341-07
Guiral, E., Goncalves Quiles, M., Munoz, L., Moreno-Morales, J., Alejo-Cancho, I., Salvador, P., et al. (2019). Emergence of resistance to quinolones and beta-lactam antibiotics in enteroaggregative and enterotoxigenic Escherichia coli causing traveler’s diarrhea. Antimicrob. Agents Chemother. 63. doi: 10.1128/AAC.01745-18
Gunzburg, S. T., Tornieporth, N. G., and Riley, L. W. (1995). Identification of enteropathogenic Escherichia coli by PCR-based detection of the bundle-forming pilus gene. J. Clin. Microbiol. 33, 1375–1377. doi: 10.1128/jcm.33.5.1375-1377.1995
Harada, T., Hiroi, M., Kawamori, F., Furusawa, A., Ohata, K., Sugiyama, K., et al. (2007). A food poisoning diarrhea outbreak caused by enteroaggregative Escherichia coli serogroup O126:H27 in Shizuoka Japan. Jpn. J. Infect. Dis. 60, 154–155.
Hardegen, C., Messler, S., Henrich, B., Pfeffer, K., Wurthner, J., and MacKenzie, C. R. (2010). A set of novel multiplex Taqman real-time PCRs for the detection of diarrhoeagenic Escherichia coli and its use in determining the prevalence of EPEC and EAEC in a university hospital. Ann. Clin. Microbiol. Antimicrob. 9:5. doi: 10.1186/1476-0711-9-5
Harrington, S. M., Dudley, E. G., and Nataro, J. P. (2006). Pathogenesis of enteroaggregative Escherichia coli infection. FEMS Microbiol. Lett. 254, 12–18. doi: 10.1111/j.1574-6968.2005.00005.x
Hebbelstrup Jensen, B., Adler Sørensen, C., Hebbelstrup Rye Rasmussen, S., Rejkjær Holm, D., Friis-Møller, A., Engberg, J., et al. (2018). Characterization of diarrheagenic enteroaggregative Escherichia coli in Danish adults—antibiotic treatment does not reduce duration of diarrhea. Front. Cell. Infect. Microbiol. 8:306. doi: 10.3389/fcimb.2018.00306
Hebbelstrup Jensen, B., Poulsen, A., Hebbelstrup Rye Rasmussen, S., Struve, C., Engberg, J. H., Friis-Møller, A., et al. (2017). Genetic virulence profile of enteroaggregative Escherichia coli strains isolated from Danish children with either acute or persistent diarrhea. Front. Cell. Infect. Microbiol. 7:230. doi: 10.3389/fcimb.2017.00230
Hebbelstrup Jensen, B., Stensvold, C. R., Struve, C., Olsen, K. E., Scheutz, F., Boisen, N., et al. (2016). Enteroaggregative Escherichia coli in daycare—a 1-year dynamic cohort study. Front. Cell. Infect. Microbiol. 6:75. doi: 10.3389/fcimb.2016.00075
Herrera-León, S., Llorente, M. T., and Sánchez, S. (2015). Plasmid-mediated quinolone resistance in different diarrheagenic Escherichia coli pathotypes responsible for complicated, noncomplicated, and traveler’s diarrhea cases. Antimicrob. Agents Chemother. 60, 1950–1951. doi: 10.1128/AAC.02909-15
Hebbelstrup Jensen, B., Olsen, K. E., Struve, C., Krogfelt, K. A., and Petersen, A. M. (2014). Epidemiology and clinical manifestations of enteroaggregative. Escherichia coli. Clin Microbiol Rev, 27(3), 614–630. doi: 10.1128/CMR.00112-13
Imuta, N., Ooka, T., Seto, K., Kawahara, R., Koriyama, T., Kojyo, T., et al. (2016). Phylogenetic analysis of enteroaggregative Escherichia coli (EAEC) isolates from Japan reveals emergence of CTX-M-14-producing EAEC O25:H4 clones related to sequence type 131. J. Clin. Microbiol. 54, 2128–2134. doi: 10.1128/JCM.00711-16
Itoh, Y., Nagano, I., Kunishima, M., and Ezaki, T. (1997). Laboratory investigation of enteroaggregative Escherichia coli O untypeable:H10 associated with a massive outbreak of gastrointestinal illness. J. Clin. Microbiol. 35, 2546–2550. doi: 10.1128/jcm.35.10.2546-2550.1997
Johnson, J. R., Murray, A. C., Gajewski, A., Sullivan, M., Snippes, P., Kuskowski, M. A., et al. (2003). Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob. Agents Chemother. 47, 2161–2168. doi: 10.1128/AAC.47.7.2161-2168.2003
Jønsson, R., Struve, C., Boisen, N., Mateiu, R. V., Santiago, A. E., Jenssen, H., et al. (2015). Novel aggregative adherence fimbria variant of enteroaggregative Escherichia coli. Infect. Immun. 83, 1396–1405. doi: 10.1128/IAI.02820-14
Jønsson, R., Struve, C., Boll, E. J., Boisen, N., Joensen, K. G., Sørensen, C. A., et al. (2017). A novel pAA virulence plasmid encoding toxins and two distinct variants of the fimbriae of enteroaggregative Escherichia coli. Front. Microbiol. 8:263. doi: 10.3389/fmicb.2017.00263
Kaper, J. B., Nataro, J. P., and Mobley, H. L. (2004). Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123–140. doi: 10.1038/nrmicro818
Letunic, I., and Bork, P. (2019). Interactive tree of life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 47, W256–W259. doi: 10.1093/nar/gkz239
Magiorakos, A. P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., et al. (2012). Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. doi: 10.1111/j.1469-0691.2011.03570.x
Malberg Tetzschner, A. M., Johnson, J. R., Johnston, B. D., Lund, O., and Scheutz, F. (2020). In silico genotyping of Escherichia coli isolates for extraintestinal virulence genes by use of whole-genome sequencing data. J. Clin. Microbiol. 58. doi: 10.1128/JCM.01269-20
Mandomando, I., Vubil, D., Boisen, N., Quinto, L., Ruiz, J., Sigauque, B., et al. (2020). Escherichia coli ST131 clones harbouring AggR and AAF/V fimbriae causing bacteremia in Mozambican children: emergence of new variant of fimH27 subclone. PLoS Negl. Trop. Dis. 14:e0008274. doi: 10.1371/journal.pntd.0008274
Manhique-Coutinho, L., Chiani, P., Michelacci, V., Taviani, E., Bauhofer, A. F. L., Chissaque, A., et al. (2022). Molecular characterization of diarrheagenic Escherichia coli isolates from children with diarrhea: a cross-sectional study in four provinces of Mozambique. Int. J. Infect. Dis. 121, 190–194. doi: 10.1016/j.ijid.2022.04.054
Minh, B. Q., Schmidt, H. A., Chernomor, O., Schrempf, D., Woodhams, M. D., von Haeseler, A., et al. (2020). IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37, 1530–1534. doi: 10.1093/molbev/msaa015
Modgil, V., Chaudhary, P., Bharti, B., Mahindroo, J., Yousuf, M., Koundal, M., et al. (2021). Prevalence, virulence gene profiling, and characterization of enteroaggregative Escherichia coli from children with acute diarrhea, asymptomatic nourished, and malnourished children younger than 5 years of age in India. J. Pediatr. 234:e105, 106–114.e5. doi: 10.1016/j.jpeds.2021.03.010
Nataro, J. P. (2003). “Enteroaggregative Escherichia coli” in Emerging infections. eds. W. M. Scheld, J. M. Hughes, and B. E. Murray, vol. 6 (Washington, DC: American Society for Microbiology)
Nataro, J. P., Mai, V., Johnson, J., Blackwelder, W. C., Heimer, R., Tirrell, S., et al. (2006). Diarrheagenic Escherichia coli infection in Baltimore, Maryland, and New Haven Connecticut. Clin. Infect. Dis. 43, 402–407. doi: 10.1086/505867
Nataro, J. P., Yikang, D., Giron, J. A., Savarino, S. J., Kothary, M. H., and Hall, R. (1993). Aggregative adherence fimbria I expression in enteroaggregative Escherichia coli requires two unlinked plasmid regions. Infect. Immun. 61, 1126–1131. doi: 10.1128/iai.61.3.1126-1131.1993
Nunes, K. O., Santos, A. C. P., Bando, S. Y., Silva, R. M., Gomes, T. A. T., and Elias, W. P. (2017). Enteroaggregative Escherichia coli with uropathogenic characteristics are present in feces of diarrheic and healthy children. Pathog. Dis. 75. doi: 10.1093/femspd/ftx106
Okeke, I. N., Lamikanra, A., Czeczulin, J., Dubovsky, F., Kaper, J. B., and Nataro, J. P. (2000). Heterogeneous virulence of enteroaggregative Escherichia coli strains isolated from children in Southwest Nigeria. J. Infect. Dis. 181, 252–260. doi: 10.1086/315204
Olsen, J. E., Aabo, S., Hill, W., Notermans, S., Wernars, K., Granum, P. E., et al. (1995). Probes and polymerase chain reaction for detection of food-borne bacterial pathogens. Int. J. Food Microbiol. 28, 1–78. doi: 10.1016/0168-1605(94)00159-4
Oswald, E., Schmidt, H., Morabito, S., Karch, H., Marches, O., and Caprioli, A. (2000). Typing of intimin genes in human and animal enterohemorrhagic and enteropathogenic Escherichia coli: characterization of a new intimin variant. Infect. Immun. 68, 64–71. doi: 10.1128/IAI.68.1.64-71.2000
Pabst, W. L., Altwegg, M., Kind, C., Mirjanic, S., Hardegger, D., and Nadal, D. (2003). Prevalence of enteroaggregative Escherichia coli among children with and without diarrhea in Switzerland. J. Clin. Microbiol. 41, 2289–2293. doi: 10.1128/JCM.41.6.2289-2293.2003
Palmeiro, A., Toro, C., Dhabi, G., Amor, A., Iglesias, N., Trevisi, P., et al. (2012). Diarrhoeagenic Escherichia coli pathotypes in travellers attending a tropical medicine unit in a Spanish hospital. J. Med. Microbiol. 61, 1485–1487. doi: 10.1099/jmm.0.044461-0
Petro, C. D., Duncan, J. K., Seldina, Y. I., Allue-Guardia, A., Eppinger, M., Riddle, M. S., et al. (2020). Genetic and virulence profiles of enteroaggregative Escherichia coli (EAEC) isolated from deployed military personnel (DMP) with travelers’ diarrhea. Front. Cell. Infect. Microbiol. 10:200. doi: 10.3389/fcimb.2020.00200
Pichel, M., Binsztein, N., and Viboud, G. (2000). CS22, a novel human enterotoxigenic Escherichia coli adhesin, is related to CS15. Infect. Immun. 68, 3280–3285. doi: 10.1128/IAI.68.6.3280-3285.2000
Pightling, A. W., Pettengill, J. B., Luo, Y., Baugher, J. D., Rand, H., and Strain, E. (2018). Interpreting whole-genome sequence analyses of foodborne bacteria for regulatory applications and outbreak investigations. Front. Microbiol. 9:1482. doi: 10.3389/fmicb.2018.01482
Pollard, D. R., Johnson, W. M., Lior, H., Tyler, S. D., and Rozee, K. R. (1990). Rapid and specific detection of verotoxin genes in Escherichia coli by the polymerase chain reaction. J. Clin. Microbiol. 28, 540–545. doi: 10.1128/jcm.28.3.540-545.1990
Ridderstedt, F., Widerstrom, M., Lindh, J., and Lilja, M. (2018). Sick leave due to diarrhea caused by contamination of drinking water supply with Cryptosporidium hominis in Sweden: a retrospective study. J. Water Health 16, 704–710. doi: 10.2166/wh.2017.311
Rogawski, E. T., Guerrant, R. L., Havt, A., Lima, I. F. N., Medeiros, P., Seidman, J. C., et al. (2017). Epidemiology of enteroaggregative Escherichia coli infections and associated outcomes in the MAL-ED birth cohort. PLoS Negl. Trop. Dis. 11:e0005798. doi: 10.1371/journal.pntd.0005798
Scavia, G., Staffolani, M., Fisichella, S., Striano, G., Colletta, S., Ferri, G., et al. (2008). Enteroaggregative Escherichia coli associated with a foodborne outbreak of gastroenteritis. J. Med. Microbiol. 57, 1141–1146. doi: 10.1099/jmm.0.2008/001362-0
Scheutz, F., Teel, L. D., Beutin, L., Piérard, D., Buvens, G., Karch, H., et al. (2012). Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 50, 2951–2963. doi: 10.1128/JCM.00860-12
Schmidt, H., Knop, C., Franke, S., Aleksic, S., Heesemann, J., and Karch, H. (1995). Development of PCR for screening of enteroaggregative Escherichia coli. J. Clin. Microbiol. 33, 701–705. doi: 10.1128/jcm.33.3.701-705.1995
Schultsz, C., Pool, G. J., van Ketel, R., de Wever, B., Speelman, P., and Dankert, J. (1994). Detection of enterotoxigenic Escherichia coli in stool samples by using nonradioactively labeled oligonucleotide DNA probes and PCR. J. Clin. Microbiol. 32, 2393–2397. doi: 10.1128/jcm.32.10.2393-2397.1994
Schüroff, P. A., Salvador, F. A., Abe, C. M., Wami, H. T., Carvalho, E., Hernandes, R. T., et al. (2021). The aggregate-forming pili (AFP) mediates the aggregative adherence of a hybrid-pathogenic Escherichia coli (UPEC/EAEC) isolated from a urinary tract infection. Virulence 12, 3073–3093. doi: 10.1080/21505594.2021.2007645
Shazberg, G., Wolk, M., Schmidt, H., Sechter, I., Gottesman, G., and Miron, D. (2003). EnteroaggregativeEscherichia coliSerotype O126:H27, Israel. Emerg. Infect. Dis. 9, 1170–1173. doi: 10.3201/eid0909.020695
Spurbeck, R. R., Dinh, P. C., Walk, S. T., Stapleton, A. E., Hooton, T. M., Nolan, L. K., et al. (2012). Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect. Immun. 80, 4115–4122. doi: 10.1128/IAI.00752-12
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Tam, C. C., O’Brien, S. J., Tompkins, D. S., Bolton, F. J., Berry, L., Dodds, J., et al. (2012). Changes in causes of acute gastroenteritis in the United Kingdom over 15 years: microbiologic findings from 2 prospective, population-based studies of infectious intestinal disease. Clin. Infect. Dis. 54, 1275–1286. doi: 10.1093/cid/cis028
Tobias, J., Kassem, E., Rubinstein, U., Bialik, A., Vutukuru, S. R., Navaro, A., et al. (2015). Involvement of main diarrheagenic Escherichia coli, with emphasis on enteroaggregative E. coli, in severe non-epidemic pediatric diarrhea in a high-income country. BMC Infect. Dis. 15:79. doi: 10.1186/s12879-015-0804-4
Tokuda, K., Nishi, J., Imuta, N., Fujiyama, R., Kamenosono, A., Manago, K., et al. (2010). Characterization of typical and atypical enteroaggregative Escherichia coli in Kagoshima, Japan: biofilm formation and acid resistance. Microbiol. Immunol. 54, 320–329. doi: 10.1111/j.1348-0421.2010.00210.x
Tornieporth, N. G., John, J., Salgado, K., de Jesus, P., Latham, E., Melo, M. C., et al. (1995). Differentiation of pathogenic Escherichia coli strains in Brazilian children by PCR. J. Clin. Microbiol. 33, 1371–1374. doi: 10.1128/jcm.33.5.1371-1374.1995
Vargas, M., Gascón, J., Gallardo, F., Jimenez De Anta, M. T., and Vila, J. (1998). Prevalence of diarrheagenic Escherichia coli strains detected by PCR in patients with travelers’ diarrhea. Clin. Microbiol. Infect. 4, 682–688. doi: 10.1111/j.1469-0691.1998.tb00652.x
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 13:e1005595. doi: 10.1371/journal.pcbi.1005595
Wilson, A., Evans, J., Chart, H., Cheasty, T., Wheeler, J. G., Tompkins, D., et al. (2001). Characterisation of strains of enteroaggregative Escherichia coli isolated during the infectious intestinal disease study in England. Eur. J. Epidemiol. 17, 1125–1130. doi: 10.1023/A:1021224915322
Keywords: enteroaggregative Escherichia coli, diarrheagenic Escherichia coli, non-travel related diarrhea, children, whole-genome sequencing, molecular characterization, extraintestinal infection
Citation: Llorente MT, Escudero R, Ramiro R, Remacha MA, Martínez-Ruiz R, Galán-Sánchez F, de Frutos M, Elía M, Onrubia I and Sánchez S (2023) Enteroaggregative Escherichia coli as etiological agent of endemic diarrhea in Spain: A prospective multicenter prevalence study with molecular characterization of isolates. Front. Microbiol. 14:1120285. doi: 10.3389/fmicb.2023.1120285
Edited by:
Axel Cloeckaert, Institut National de recherche pour l’agriculture, l’alimentation et l’environnement (INRAE), FranceReviewed by:
Elna Buys, University of Pretoria, South AfricaNadia Boisen, Statens Serum Institut (SSI), Denmark
Jorge Blanco, University of Santiago de Compostela, Spain
Enrique Joffré, Karolinska Institutet (KI), Sweden
Copyright © 2023 Llorente, Escudero, Ramiro, Remacha, Martínez-Ruiz, Galán-Sánchez, de Frutos, Elía, Onrubia and Sánchez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sergio Sánchez, c2VyZ2lvLnNhbmNoZXpAaXNjaWlpLmVz