 Frédéric De Meyer1
Frédéric De Meyer1 Aurélien Carlier1,2*
Aurélien Carlier1,2*- 1Laboratory of Microbiology, Department of Biochemistry and Microbiology, Ghent University, Ghent, Belgium
- 2LIPME, Université de Toulouse, INRAE, CNRS, Castanet-Tolosan, France
Serine protease inhibitors are a large family of proteins involved in important pathways and processes, such as inflammatory responses and blood clotting. Most are characterized by a precise mode of action, thereby targeting a narrow range of protease substrates. However, the serine-protease inhibitor ecotin is able to inhibit a broad range of serine proteases that display a wide range of specificities. This specificity is driven by special structural features which allow unique flexibility upon binding to targets. Although frequently observed in many human/animal-associated bacteria, ecotin homologs may also be found in plant-associated taxa and environmental species. The purpose of this review is to provide an update on the biological importance, role in host–microbe interactions, and evolutionary relationship between ecotin orthologs isolated from Eukaryotic and Prokaryotic species across the Tree of Life.
Introduction
Serine-protease inhibitors (serpins) are a large family of protease inhibitors with members in bacteria, fungi, plants, and humans (Silverman et al., 2001; Spence et al., 2021). Serpins are primarily known for playing a role in controlling serine protease activity in biological processes (Gettins, 2002). Targets of serine proteases, in turn, participate in the regulation of a wide variety of complex physiological pathways, such as inflammation, fibrinolysis, and blood coagulation (Stein and Carrell, 1995). The structural mechanism by which serpins inhibit their protease substrates is well understood (Huntington et al., 2000). Target proteases interact with serpins, cleaving a reactive center loop (RCL) which protrudes from the serpin body. Following cleavage, but before hydrolysis of the acyl enzyme intermediate, the RCL inserts into the center of the serpin body, effectively trapping the protease. Through conformational changes, involving major reorganization of exposed protease recognition loops, serpins form a noncovalent complex upon binding (Gettins, 2000). Serpin protease inhibitors usually display high specificity (Gettins and Ofson, 2009), but some serpins are capable of inhibiting a broad range of serine proteases (Ksiazek et al., 2015).
Ecotin (Escherichia coli trypsin inhibitor) is a member of the serpin superfamily and a potent inhibitor of serine proteases, first isolated from E. coli (Chung et al., 1983). Its 16 kDa structure consists of a monomer that includes a 20 amino acid signal peptide which targets the protein to the periplasm (McGrath et al., 1991). High-resolution crystal structures revealed that two ecotin monomers assemble into a contralateral dimer which binds to two target protease molecules at opposite ends to form a heterotetramer (Pál et al., 1996; Yang et al., 1998). Each ecotin monomer inhibits its respective target via binding at two different surface contact sites: a primary and a secondary site (McGrath et al., 1994; Yang et al., 1998). This 1:1 stoichiometric configuration of two ecotin monomers for two protease units is unique among all the known structures and mechanisms of serine-protease inhibitor complexes (Figure 1A; Yang et al., 1998). Therefore, ecotin does not belong to one of the already established serpin families and has been classified in the MEROPS database as inhibitor family I11, clan IN (Rawlings et al., 2018). Interestingly, ecotin does not target endogenous E. coli proteases, and is therefore unlikely to be involved in the regulation of protease activity in this organism (Eggers et al., 2004). Rather, ecotin likely plays a role in protection against exogenous proteolytic attacks in environments such as the mammalian gastrointestinal tract (Chung et al., 1983; Seymour et al., 1994). In keeping with this hypothesis, ecotin is a reversible inhibitor of a wide range of mammalian and human derived serine proteases including trypsin, chymotrypsin, neutrophil elastase (NE), cathepsin G, granzyme B, and mannan-binding lectin-associated serine proteases (MASPs; Chung et al., 1983; Waugh et al., 2000; Nagy et al., 2019).
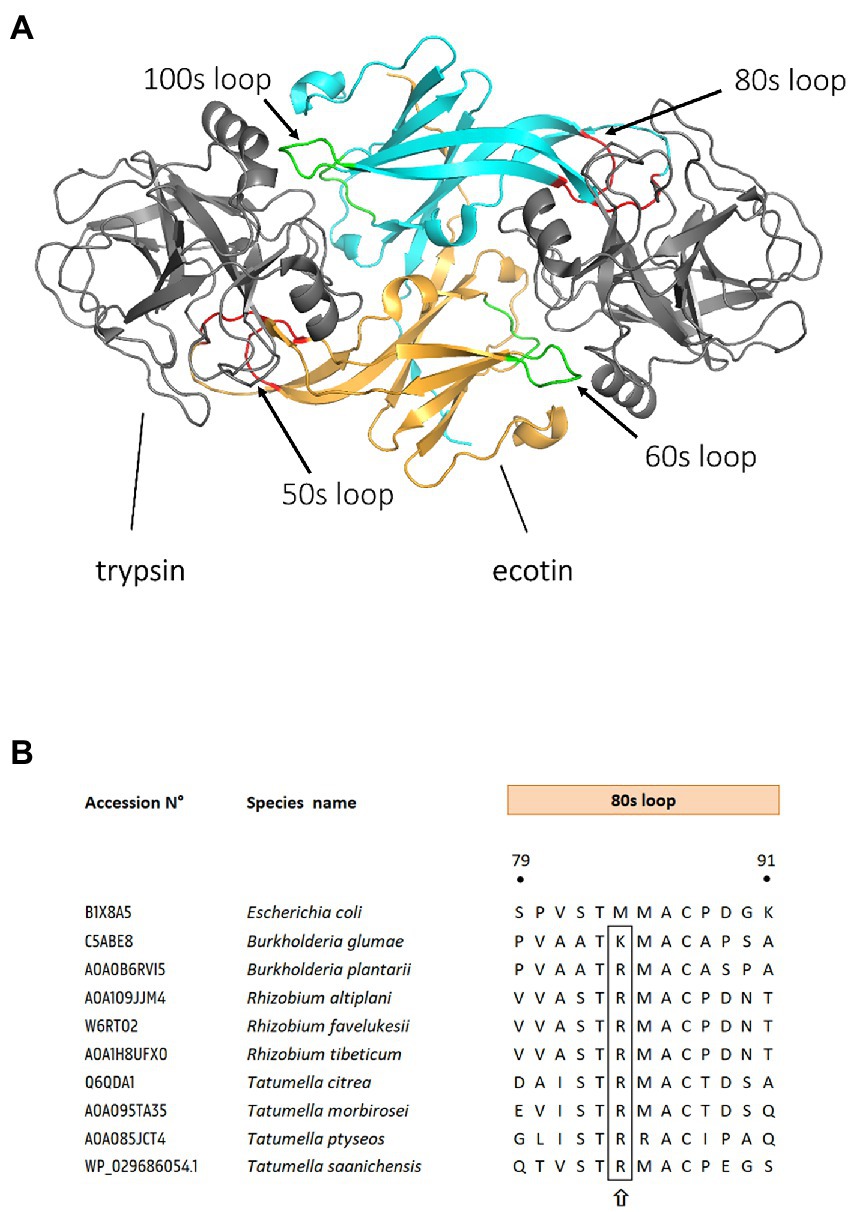
Figure 1. Binding sites of ecotin and ecotin-like proteins to their substrates. (A) Structure of a homodimeric ecotin in complex with trypsin protease. Ecotin (in orange and cyan) forms a stable homodimer, which binds two trypsin proteases simultaneously (in gray). The ecotin dimer contains two distinct protease-binding sites, each composed of two loops. The primary binding site (red) is composed of the 80s loop and the 50s loop, while the secondary binding site is composed of the 60s and 100 s loops (shown in green). Figure is adapted from Gillmor et al. (2000), using Protein Database (PDB) entry 1EZU. (B) Sequence alignment of ecotin orthologs primary binding site. Ecotin ortholog protein sequences were aligned, using MUSCLE (Edgar, 2004) in MEGA11 (Tamura et al., 2021). Residue position numbering is according to E. coli ecotin. The P1 position from the 80s loop (the primary specificity determinant) is designated by a upward arrow. UniProt or NCBI GenBank accession numbers are mentioned for each bacterial ecotin sequence.
The structure and substrate binding properties of ecotin have been analyzed by X-ray crystallography and protease-binding assays (McGrath et al., 1994; Shin et al., 1996; Perona et al., 1997; Wang et al., 2001, 2003; Clark et al., 2011; Gaboriaud et al., 2013). These studies have primarily focused on the E. coli ecotin protein, but homologs from other taxa (e.g., Yersinia pestis) display similar protein quaternary structure and binding partners despite considerable primary sequence divergence (Clark et al., 2011). However, it is increasingly recognized that the genomes of diverse bacterial and eukaryotic taxa encode homologs of ecotin, which play various biological and functional roles (Eggers et al., 2004; Ireland et al., 2014; Verma et al., 2018; Nagy et al., 2019). This review aims at giving an update on the biological importance, role in host–microbe interactions, and evolutionary relationship between ecotin orthologs isolated from prokaryotic and eukaryotic species across the Tree of Life, as well as its potential in medical biotechnology.
Taxonomic distribution of ecotin
Ecotin has been studied for nearly four decades (Chung et al., 1983), and more than 600 protein homologs have since been discovered across the Bacterial and Eukaryotic kingdoms (Figure 2; Eschenlauer et al., 2009; Ireland et al., 2014; Nagy et al., 2019; Garcia et al., 2020). Ecotin sequences diverge along the major taxonomic lineages, indicating that homologs were present in the ancestors of Proteobacteria (Figure 2C). Interestingly, sequences from other phyla (e.g., Bacteroidetes) are nested within Proteobacteria clusters, indicating that ecotin-like proteins were acquired via horizontal gene transfer (Figure 2C).

Figure 2. Taxonomic distribution of organisms containing ecotin orthologs. (A) More than six hundred species are present throughout the Tree of Life and encode at least one ecotin homolog. Protein sequences were extracted from InterPro (query: IPR036198) and validated as belonging to the same orthogroup with eggNOG-Mapper (E-value: 0.0001, minimum bit score: 60, % identity >40, query coverage >20%; Blum et al., 2021; Cantalapiedra et al., 2021). Orthology assignments were double checked in EggNOG database using EggNOG OGs. Remaining protein sequences were checked by the presence of ecotin domains as predicted by Pfam using eggNOG-Mapper (Mistry et al., 2021). One sequence for each species was retained, after which they were further aligned based on their amino acid sequence using MUSCLE (Edgar, 2004) in MEGA11 (Tamura et al., 2021). (A) Among Bacteria, the class of Proteobacteria (502 species) represents the majority of ecotin-expressing species. Note that this analysis is not comprehensive and homologs may have been missed due to biases in database representation and/or search parameters. (B) Among the Eukaryotes, both the Trypanosoma and Leishmania species, belonging to the Kinetoplastida class, contain over 10 species that are causative agents of several and widespread infectious diseases. The figure was made using KronaTools 2.7.1 (Ondov et al., 2011). (C) Unrooted phylogenetic tree of ecotin orthologs. Only one representative sequence per identified EggNOG OG was retained. Sequences were aligned based on their amino acid sequences using MUSCLE (Edgar, 2004) in MEGA11 (Tamura et al., 2021). A phylogenetic tree was constructed using the Maximum Likelihood (ML) method and LG + G4 substitution model in IQ-TREE (Minh et al., 2020). The SH-aLRT support (%) and the percentage of replicate trees in which the associated taxa cluster together in the bootstrap test (1,000 replicates) are shown next to the branches. The scale bar indicates the number of substitutions per site. Additional annotations are presented showing which taxa contain animal pathogenic (orange squares), plant-associated (green squares), and environmental (blue squares) strains.
Ecotin homologs are widespread among Bacteria, particularly within the Gammaproteobacteria (Nagy et al., 2019). Among these, the genomes of several human pathogenic species such as Yersinia pestis, Klebsiella oxytoca, Salmonella enterica, Citrobacter rodentium, Pseudomonas aeruginosa, and Burkholderia pseudomallei and several species of Enterobacter encode ecotin homologs (Darby et al., 2014; Ireland et al., 2014; Tseng et al., 2018; Nagy et al., 2019; Salimiyan Rizi et al., 2019; Subramaniam et al., 2019; Vogt et al., 2019). Also in the Gammaproteobacteria, most genomes of Xenorhabdus species encode ecotin. Although this bacterial genus is known for symbiotic associations with nematodes, some members are also pathogenic to insects (Chaston et al., 2011). In addition, ecotin homologs have recently been characterized in pathogenic strains of Campylobacter rectus and Campylobacter showae (Epsilonproteobacteria), which reside in the oral cavity (Thomas et al., 2020).
Although mostly studied in the context of infection, ecotin proteins are not exclusive to animal pathogens. For instance, ecotin homologs are common in members of the genus Pseudomonas, which includes species without known pathogenic strains such as Pseudomonas fluorescens or Pseudomonas protegens (Figure 2A). Pseudomonas species are known for their metabolic diversity and ability to colonize a wide range of environmental niches (Spiers et al., 2000; Vartapetian et al., 2011), as well as the pathogenic Pseudomonas aeruginosa. Several other genera with few known pathogenic members encode ecotin homologs, including Chryseobacterium (more than 50 species) and Shewanella (more than 30 species). Although Chryseobacterium species have been isolated from diseased fish and human wounds, their main habitat is freshwater and soil (Gallego et al., 2006; Zhou et al., 2007; Cho et al., 2010). Similarly, Shewanella species are aquatic microorganisms with a worldwide distribution, with species adapted to extreme environments and are rarely host-associated (Abboud et al., 2005; Hau and Gralnick, 2007). Interestingly, ecotin homologs are also found in a few, taxonomically diverse plant-associated taxa. Tamlana fucoidanivorans, Elizabethkingia argenteiflava, and Sphingobacterium nematocida have been isolated from the endosphere or from plant surfaces (Liu et al., 2012; Li et al., 2020; Hwang et al., 2021). Furthermore, some of these species have a plant-pathogenic lifestyle. For example, Pseudomonas fuscovaginae, Burkholderia glumae, and Burkholderia plantarii are important pathogens of rice, while Ganoderma boninense and Gibbsiella quercinecans cause stem rot of oil palm and acute oak decline, respectively (Brady et al., 2016; Isha et al., 2020).
The genomes of several Eukaryotes also encode ecotin orthologs, especially within the phylum Euglenozoa, which contains the insect-borne parasitic genera Trypanosoma and Leishmania (Peña et al., 2017; Verma et al., 2017, 2018; Garcia et al., 2020; Levy et al., 2021). These genera contain species such as Trypanosoma cruzi, which causes Chagas disease (Brener, 1973), Leishmania major causing zoonotic cutaneous leishmaniasis (El-On et al., 1984), and Leishmania donovani the causative agent of visceral leishmaniasis, traditionally known as kala-azar (“black fever”; van Griensven and Diro, 2012). In contrast to prokaryotes which usually encode only 1 ortholog of ecotin, the eukaryotic unicellular Leishmania pathogens harbor three distinct paralogs, named ISP1, ISP2 and ISP3 (Eschenlauer et al., 2009; Morrison et al., 2012). In L. major, ISP1 is located on the same transcription unit upstream to ISP2 and ISP3 (Eschenlauer et al., 2009). ISP1 and ISP2 encode proteins of around 17 kDa, which is comparable to the 16 kDa mature form isolated from E. coli ecotin. ISP3, on the other hand, encodes a protein which is more than double the size (41.8 kDa), with an ecotin-like N-terminal domain, and a C-terminal domain of unknown function (Eschenlauer et al., 2009). Ecotin proteins of Euglenozoa form a distinct phylogenetic clade, nested within proteins of Gamma- and Betaproteobacteria (Figure 2C). Thus, ecotin proteins in these lineages seem to have evolved from a single ancestor, perhaps acquired from Proteobacteria via horizontal gene transfer. Interestingly, protease contact residues and substrate-like loops of Leishmania and Trypanosoma ecotin proteins display low identity to residues from bacterial homologs, indicating functional divergence (Peña et al., 2017).
Function of ecotin in animal-associated microorganisms
Although the molecular structure and protease targets of ecotin are well characterized for a few organisms, its precise ecological function remains unclear. In E. coli, ecotin may play a role in protecting the cell against host proteases. Ecotin is translocated to the periplasmic space, where it can protect the cell against NE that may have permeated through the damaged outer cell membrane of Gram-negative bacteria (Eggers et al., 2004). Aside from a role in protection against the host immune system, ecotin from E. coli may play a role in microbe-microbe interactions, with E. coli ecotin knock-out strains more susceptible to T6SS-mediated killing by Vibrio cholerae (Myint et al., 2021). However, the precise mode of action remains to be elucidated. In P. aeruginosa, ecotin was shown to be released into the extracellular milieu via cell lysis during biofilm formation (Webb et al., 2003; Wang et al., 2013; Tseng et al., 2018). Ecotin homologs from P. aeruginosa and Y. pestis also inhibit NE (Eggers et al., 2001; Clark et al., 2011; Tseng et al., 2018). Similarly, ecotin of B. pseudomallei is essential for intracellular survival in murine macrophages, probably by inhibiting host proteases of the early endosome (Ireland et al., 2014). P. aeruginosa as well as B. cepacia are two major pathogens causing chronic infections in adult cystic fibrosis (CF) patients, both of which possess ecotin homologs (Govan and Deretic, 1996; Rajan and Saiman, 2002). Interestingly, P. aeruginosa and B. cepacia are opportunistic human pathogens that thrive in the lung environment as biofilms, and as such are not exposed to digestive or plasma proteases (Lavoie et al., 2011; Yaghi et al., 2020). However, pulmonary infections are mostly associated with increased numbers of degranulating neutrophils, and therefore by high concentrations of NE (Schaaf et al., 2000). A similar situation is also observed in CF, where there is an influx of neutrophils (Goldstein and Doring, 1986; Birrer et al., 1994; Witko-Sarsat et al., 1999). P. aeruginosa ecotin, released to the extracellular milieu via cell lysis during biofilm formation, directly binds to Psl, a component of the biofilm exopolysaccharide matrix (Tseng et al., 2018). Ecotin might therefore protect the biofilm from antimicrobial effectors and proteolytic degradation (Tseng et al., 2018; Nagy et al., 2019). This may represent a novel mechanism of protection for biofilms to increase their tolerance against the innate immune response. Interestingly, P. aeruginosa ecotin, together with the human protease inhibitor SERPINB1, has recently been proposed to act as a barrier to SARS-CoV-2 infection in CF lungs by inhibiting priming of the S protein by TMPRSS2 (Stanton et al., 2020). Although this role of ecotin as a protective layer may have intriguing consequences for the host, this remains to be proven experimentally.
The main function of the ecotin-like homologs ISP1, ISP2, and ISP3 of eukaryotic parasites T. cruzi and L. major is likely protection against intestinal proteases, such as neutrophil elastase in the gut of an insect vector (Lima and Mottram, 2010; Alam et al., 2016; Verma et al., 2017). However, a potential role for these proteins in survival inside the insect vector remains to be tested. Protozoan metacaspases (MCAs) of Leishmania species can be inhibited by the peptide ecotin-like ISP3 inhibitor from L. amazonensis and L. major. ISP3 interferes with the trypsin-like activity, resulting in significantly reduced parasite cell death (Peña et al., 2017; Shadab et al., 2017). In addition, serine proteases with MCA activity also play a role in the programmed cell death (PCD) in Leishmania donovani (Das et al., 2014), Trypanosoma brucei (T. brucei; Szallies et al., 2002), and other Leishmania species (Lee et al., 2007; Khademvatan et al., 2011; Castanys-Muñoz et al., 2012). Ecotin-like protein ISP2 of L. major inhibits mannan-binding lectin (MBL)-associated serine protease (MASP)-2 in addition to NE in the host cell (Verma et al., 2018). MASP-2 is involved in the cleavage of proteins in the complement system as well as in the coagulation cascade through cleavage of prothrombin to thrombin (Thiel et al., 1997; Krarup et al., 2007). In addition to L. major, E. coli, Y. pestis, and P. aeruginosa homologs also display inhibition of MASP2, but also of MASP1 and MASP3 with Ki values ranging from 10−5 to 10−9 (Cortesio and Jiang, 2006; Gaboriaud et al., 2013; Nagy et al., 2019). MASP1, MASP2, and MASP3 participate to the lectin pathway of the complement system, an essential part of the innate immune system that acts as the first line of defense against pathogens (Héja et al., 2012; Dobó et al., 2016). This function of ecotin seems surprisingly conserved, as endogenous ecotin also protects E. coli against attack from the lectin pathway (Nagy et al., 2019). Most recently, ISP2 from T. brucei has been identified as a virulence factor, contributing to the reduction of NO-producing myeloid cells and of IFN-γ-producing NK-cells: Mice infected with ΔISP2 mutants strains displayed lower blood parasitemia, delayed symptoms, and survived longer (Levy et al., 2021). Moreover, ISP2 of T. cruzi contributes to evasion and replication in macrophages (Garcia et al., 2020). These findings indicate that ISP2 is a virulence factor in mice and attenuates the inflammatory response during early infection. One of the major roles of ecotin in animal pathogens may be to inactivate key proteases of the immune system.
The role of ecotin in plant-associated bacteria
Although the role of ecotin in protecting pathogens or parasites against the mammalian innate immunity is relatively clear, there is a dearth of data about the targets of ecotin outside mammal systems. Some notorious plant pathogens encode ecotin homologs, for example Burkholderia plantarii, Burkholderia glumae, Pseudomonas fuscovaginae, or Tatumella citrea (Ta. citrea). So far, a single study by Eggers et al. analyzed the contribution of ecotin to virulence of the plant pathogen Ta. citrea (Eggers et al., 2001). Interestingly, ecotin from Ta. citrea, the phytopathogen responsible for pink disease in pineapples (Cha et al., 1997), displayed 1,000-fold weaker inhibition against human NE compared to proteins of E. coli, Y. pestis, or P. aeruginosa (Eggers et al., 2004). Despite this lower anti-NE activity, ecotin from Ta. citrea maintains inhibitory activity against trypsin, and may thus protect the bacteria against digestive proteases in an animal vector (Eggers et al., 2004). The first line of defense of plants is drastically different from the mammalian immune system. For example, NE is known to be involved in the non-oxidative pathway of innate defense which represents the first line of defense against invading microorganisms in mammals (Burg and Pillinger, 2001; Hargaden and Singer, 2012; Juul-Madsen et al., 2013), but there is no equivalent in plants (Thomas et al., 1988). Plants have developed various other mechanisms to protect themselves from pathogens. After the first contact with a pathogen, plant cells react by releasing reactive oxygen intermediates, salicylic acid, nitric oxide, ethylene, and/or jasmonic acid (Conrath et al., 2002). These signals orchestrate different downstream responses like the activation of cell wall reinforcement proteins or synthesis of antimicrobial peptides and phenolics, depending on the nature of the injury or infection (Jones and Dangl, 2006). Perhaps because of this divergence of targets, ecotin proteins in animal pathogens commonly display aliphatic methionine (M) or leucine (L) residues in the ecotin contact binding loop, whereas plant pathogens like Ta. citrea and B. glumae display conserved cationic arginine (R) and lysine (K) residues, respectively. Accordingly, ecotin of Ta. citrea only weakly inhibited mammalian thrombin, indicating the importance of other contact residues in the selectivity and binding specificity of the target (Eggers et al., 2004). Interestingly, an R residue is also present in the ecotin contact binding loop of all Rhizobium and some Tatumella species such as Ta. morbirosei, Ta. ptyseos, and Ta. Saanichensis (Figure 1B).
Serine proteases are abundant in plants, participating in numerous crucial processes such as plant immunity (reviewed in Figueiredo et al., 2018). Plant subtilases, a large family of plant serine proteases (MEROPS subfamily S8A), have a broad range of biological functions in plant development, but also in response to biotic and abiotic stresses (Schaller et al., 2012). Interestingly, some plant subtilases are key to the response to pathogen attack and PCD (Ryan and Pearce, 2003; Huffaker et al., 2006; Huffaker and Ryan, 2007; Vartapetian et al., 2011). As a countermeasure, some pathogens secrete inhibitors that target extracellular subtilases to avoid recognition. For example, tomato apoplastic S8 subtilases, P69B and P69C, are PR proteins that play a role in response to Phytophthora infestans and Phytophthora syringae infection (Jordá et al., 1999; Jorda et al., 2000). Kazal-like inhibitors EPI1 and EPI10 from Phytophthora infestans inactivate P69B, suggesting that protease inhibition is an important strategy for plant pathogens (Tian et al., 2004, 2005). Moreover, there are several other serine protease involved in different steps during plant immunity (Figueiredo et al., 2014; Van Der Hoorn & Klemenčič, 2021), but also in other processes during in plant defense activated through abiotic stimuli (Figueiredo et al., 2018). Metacaspases, for example, are also possible candidate targets as they are orthologs of metazoan caspases, restricted to fungi, protozoa and plants (Uren, 2000). Caspases are a family of cysteine proteases (C14), with a catalytic cysteine and histidine dyad essential for enzyme activity, playing a crucial role in PCD in plants (Del Pozo and Lam, 1998). Cysteine proteases have a distinct catalytic mechanism from serine proteases, and are important hubs in plant immunity (Misas-Villamil et al., 2016). Although there are no documented ecotin targets in plant immunity-related processes, the ecotin-like protein ISP3 from L. amazonensis has recently been suggested to also inhibit cysteine proteases such as MCAs (Peña et al., 2017). The presence of ecotin in the proteomes of plant pathogens is therefore intriguing, and could indicate a role in circumventing or preventing plant defenses. However, prediction of potential ecotin targets is difficult, and in addition several of the plant pathogens mentioned above are related to clinical pathogens that cause severe infections in humans. For instance, there are reports of B. glumae clinical infections (Weinberg et al., 2007), Tatumella ptyseos is known as a foodborne opportunistic pathogen and Ta. saanichensis has been isolated from a CF patient (Mardaneh et al., 2014; Tracz et al., 2015; Bourlond et al., 2019). Whether ecotin plays an adaptive role in plant pathogenicity or is rather a vestige from ancestral animal-associated lifestyles remains unanswered.
Conclusion
Although ecotin homologs are widespread in the Bacterial and Eukaryotic kingdoms with diverse host-associated or environmental lifestyles, only a handful potential targets have been identified. Exploring and characterizing new ecotin targets could therefore be of importance in understanding several poorly studied pathways or protease reaction cascade mechanisms. Characterizing ecotin targets in new systems, especially plants, may provide novel insights into host immunity, and perhaps new ways to manage infections.
Author contributions
FD and AC drafted and edited the manuscript. All authors contributed to the article and approved the submitted version.
Funding
AC acknowledges support from the Flemish Fonds Wetenschappelijk Onderzoek under grant G017717N. AC also acknowledges support from the French National Research Agency under grant agreement ANR-19-TERC-0004-01 and from the French Laboratory of Excellence project “TULIP” (ANR-10-LABX-41; ANR-11-IDEX-0002-02). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abboud, R., Popa, R., Souza-Egipsy, V., Giometti, C. S., Tollaksen, S., Mosher, J. J., et al. (2005). Low-temperature growth of Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 71, 811–816. doi: 10.1128/AEM.71.2.811-816.2005
Alam, M. N., Das, P., De, T., and Chakraborti, T. (2016). Identification and characterization of a Leishmania donovani serine protease inhibitor: possible role in regulation of host serine proteases. Life Sci. 144, 218–225. doi: 10.1016/j.lfs.2015.12.004
Birrer, P., Mcelvaney, N. G., Rüdeberg, A., Wirz Sommer, C., Liechti-Gallati, S., Kraemer, R., et al. (1994). Protease-antiprotease imbalance in the lungs of children with cystic fibrosis. Am. J. Respir. Crit. Care Med. 150, 207–213. doi: 10.1164/ajrccm.150.1.7912987
Blum, M., Chang, H. Y., Chuguransky, S., Grego, T., Kandasaamy, S., Mitchell, A., et al. (2021). The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 49, D344–D354. doi: 10.1093/nar/gkaa977
Bourlond, B., Croxatto, A., Donner, V., Prodhom, G., and Guery, B. (2019). Tatumella ptyseos causing human sepsis: report of the first case in Switzerland and review of the literature. SN Compr. Clin. Med. 1, 200–202. doi: 10.1007/s42399-018-0021-8
Brady, C., Allainguillaume, J., Denman, S., and Arnold, D. (2016). Rapid identification of bacteria associated with acute oak decline by high-resolution melt analysis. Lett. Appl. Microbiol. 63, 89–95. doi: 10.1111/lam.12593
Brener, Z. (1973). Biology of Trypanosoma cruzi. Annu. Rev. Microbiol. 27, 347–382. doi: 10.1146/annurev.mi.27.100173.002023
Burg, N. D., and Pillinger, M. H. (2001). The neutrophil: function and regulation in innate and humoral immunity. Clin. Immunol. 99, 7–17. doi: 10.1006/clim.2001.5007
Cantalapiedra, C. P., Hernandez Plaza, A., Letunic, I., Bork, P., and Huerta-Cepas, J. (2021). eggNOG-mapper v2: functional annotation, Orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 38, 5825–5829. doi: 10.1093/MOLBEV/MSAB293
Castanys-Muñoz, E., Brown, E., Coombs, G. H., and Mottram, J. C. (2012). Leishmania mexicana metacaspase is a negative regulator of amastigote proliferation in mammalian cells. Cell Death Dis. 3:e385. doi: 10.1038/cddis.2012.113
Cha, J. S., Pujol, C., and Kado, C. I. (1997). Identification and characterization of a Pantoea citrea gene encoding glucose dehydrogenase that is essential for causing pink disease of pineapple. Appl. Environ. Microbiol. 63, 71–76. doi: 10.1128/aem.63.1.71-76.1997
Chaston, J. M., Suen, G., Tucker, S. L., Andersen, A. W., Bhasin, A., Bode, E., et al. (2011). The Entomopathogenic bacterial endosymbionts Xenorhabdus and Photorhabdus: convergent lifestyles from divergent genomes. PLoS One 6:e27909. doi: 10.1371/journal.pone.0027909
Cho, S. H., Lee, K. S., Shin, D. S., Han, J. H., Park, K. S., Lee, C. H., et al. (2010). Four new species of Chryseobacterium from the rhizosphere of coastal sand dune plants, Chryseobacterium elymi sp. nov., Chryseobacterium hagamense sp. nov., Chryseobacterium lathyri sp. nov. and Chryseobacterium rhizosphaerae sp. nov. Syst. Appl. Microbiol. 33, 122–127. doi: 10.1016/j.syapm.2009.12.004
Chung, C. H., Ives, H. E., Almeda, S., and Goldberg, A. L. (1983). Purification from Escherichia coli of a periplasmic protein that is a potent inhibitor of pancreatic proteases. J. Biol. Chem. 258, 11032–11038. doi: 10.1016/s0021-9258(17)44382-1
Clark, E. A., Walker, N., Ford, D. C., Cooper, I. A., Oyston, P. C. F., and Acharya, K. R. (2011). Molecular recognition of chymotrypsin by the serine protease inhibitor ecotin from Yersinia pestis. J. Biol. Chem. 286, 24015–24022. doi: 10.1074/jbc.M111.225730
Conrath, U., Pieterse, C. M. J., and Mauch-Mani, B. (2002). Priming in plant-pathogen interactions. Trends Plant Sci. 7, 210–216. doi: 10.1016/S1360-1385(02)02244-6
Cortesio, C. L., and Jiang, W. (2006). Mannan-binding lectin-associated serine protease 3 cleaves synthetic peptides and insulin-like growth factor-binding protein 5. Arch. Biochem. Biophys. 449, 164–170. doi: 10.1016/j.abb.2006.02.006
Darby, A., Lertpiriyapong, K., Sarkar, U., Seneviratne, U., Park, D. S., Gamazon, E. R., et al. (2014). Cytotoxic and pathogenic properties of Klebsiella oxytoca isolated from laboratory animals. PLoS One 9:e100542. doi: 10.1371/journal.pone.0100542
Das, P., De, T., and Chakraborti, T. (2014). Leishmania donovani secretory serine protease alters macrophage inflammatory response via COX-2 mediated PGE-2 production. Indian J. Geo-Marine Sci. 51, 542–551.
Del Pozo, O., and Lam, E. (1998). Caspases and programmed cell death in the hypersensitive response of plants to pathogens. Curr. Biol. 8, 1129–1132. doi: 10.1016/S0960-9822(98)70469-5
Dobó, J., Szakács, D., Oroszlán, G., Kortvely, E., Kiss, B., Boros, E., et al. (2016). MASP-3 is the exclusive pro-factor D activator in resting blood: the lectin and the alternative complement pathways are fundamentally linked. Sci. Rep. 6:31877. doi: 10.1038/srep31877
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Eggers, C. T., Murray, I. A., Delmar, V. A., Day, A. G., and Craik, C. S. (2004). The periplasmic serine protease inhibitor ecotin protects bacteria against neutrophil elastase. Biochem. J. 379, 107–118. doi: 10.1042/BJ20031790
Eggers, C. T., Wang, S. X., Fletterick, R. J., and Craik, C. S. (2001). The role of ecotin dimerization in protease inhibition. J. Mol. Biol. 308, 975–991. doi: 10.1006/jmbi.2001.4754
El-On, J., Jacobs, G. P., Witztum, E., and Greenblatt, C. L. (1984). Development of topical treatment for cutaneous leishmaniasis caused by Leishmania major in experimental animals. Antimicrob. Agents Chemother. 26, 745–751. doi: 10.1128/AAC.26.5.745
Eschenlauer, S. C. P., Faria, M. S., Morrison, L. S., Bland, N., Ribeiro-Gomes, F. L., Dosreis, G. A., et al. (2009). Influence of parasite encoded inhibitors of serine peptidases in early infection of macrophages with Leishmania major. Cell. Microbiol. 11, 106–120. doi: 10.1111/j.1462-5822.2008.01243.x
Figueiredo, A., Monteiro, F., and Sebastiana, M. (2014). Subtilisin-like proteases in plant–pathogen recognition and immune priming: A perspective. Front. Plant Sci. 5:739. doi: 10.3389/fpls.2014.00739
Figueiredo, J., Sousa Silva, M., and Figueiredo, A. (2018). Subtilisin-like proteases in plant defence: the past, the present and beyond. Mol. Plant Pathol. 19, 1017–1028. doi: 10.1111/mpp.12567
Gaboriaud, C., Gupta, R. K., Martin, L., Lacroix, M., Serre, L., Teillet, F., et al. (2013). The serine protease domain of MASP-3: enzymatic properties and crystal structure in complex with Ecotin. PLoS One 8:e67962. doi: 10.1371/journal.pone.0067962
Gallego, V., García, M. T., and Ventosa, A. (2006). Chryseobacterium hispanicum sp. nov., isolated from the drinking water distribution system of Sevilla. Spain. Int. J. Syst. Evol. Microbiol. 56, 1589–1592. doi: 10.1099/ijs.0.64264-0
Garcia, F. B., Cabral, A. D., Fuhlendorf, M. M., da Cruz, G. F., dos Santos, J. V., Ferreira, G. C., et al. (2020). Functional and structural characterization of an ecotin-like serine protease inhibitor from Trypanosoma cruzi. Int. J. Biol. Macromol. 151, 459–466. doi: 10.1016/j.ijbiomac.2020.02.186
Gettins, P. G. W. (2000). Keeping the serpin machine running smoothly. Genome Res. 10, 1833–1835. doi: 10.1101/gr.168900
Gettins, P. G. W. (2002). Serpin structure, mechanism, and function. Chem. Rev. 102, 4751–4804. doi: 10.1021/cr010170+
Gettins, P. G. W., and Ofson, S. T. (2009). Exosite determinants of serpin specificity. J. Biol. Chem. 284, 20441–20445. doi: 10.1074/jbc.R800064200
Gillmor, S. A., Takeuchi, T., Yang, S. Q., Craik, C. S., and Fletterick, R. J. (2000). Compromise and accommodation in ecotin, a dimeric macromolecular inhibitor of serine proteases. J. Mol. Biol. 299, 993–1003. doi: 10.1006/jmbi.2000.3812
Goldstein, W., and Doring, G. (1986). Lysosomal enzymes from polymorphonuclear leukocytes and proteinase inhibitors in patients with cystic fibrosis. Am. Rev. Respir. Dis. 134, 49–56. doi: 10.1164/arrd.1986.134.1.49
Govan, J. R. W., and Deretic, V. (1996). Microbial pathogenesis in cystic fibrosis: mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev. 60, 539–574. doi: 10.1128/MMBR.60.3.539-574.1996
Hargaden, M., and Singer, L. (2012). “Anatomy, physiology, and behavior” in The laboratory rabbit, Guinea pig, hamster, and other rodents, Eds. Mark A. Suckow, Karla A. Stevens and Ronald P. Wilson, 575–602.
Hau, H. H., and Gralnick, J. A. (2007). Ecology and biotechnology of the genus Shewanella. Annu. Rev. Microbiol. 61, 237–258. doi: 10.1146/annurev.micro.61.080706.093257
Héja, D., Kocsis, A., Dobó, J., Szilágyi, K., Szász, R., Závodszky, P., et al. (2012). Revised mechanism of complement lectin-pathway activation revealing the role of serine protease MASP-1 as the exclusive activator of MASP-2. Proc. Natl. Acad. Sci. U. S. A. 109, 10498–10503. doi: 10.1073/pnas.1202588109
Huffaker, A., Pearce, G., and Ryan, C. A. (2006). An endogenous peptide signal in Arabidopsis activates components of the innate immune response. Proc. Natl. Acad. Sci. U. S. A. 103, 10098–10103. doi: 10.1073/pnas.0603727103
Huffaker, A., and Ryan, C. A. (2007). Endogenous peptide defense signals in Arabidopsis differentially amplify signaling for the innate immune response. Proc. Natl. Acad. Sci. U. S. A. 104, 10732–10736. doi: 10.1073/pnas.0703343104
Huntington, J. A., Read, R. J., and Carrell, R. W. (2000). Structure of a serpin–protease complex shows inhibition by deformation. Nature 407, 923–926. doi: 10.1038/35038119
Hwang, J.-H., Kim, J., Kim, J.-H., and Mo, S. (2021). Elizabethkingia argenteiflava sp. nov., isolated from the pod of soybean, Glycine max. Int. J. Syst. Evol. Microbiol. 71:767. doi: 10.1099/ijsem.0.004767
Ireland, P. M., Marshall, L., Norville, I., and Sarkar-Tyson, M. (2014). The serine protease inhibitor Ecotin is required for full virulence of Burkholderia pseudomallei. Microb. Pathog. 67-68, 55–58. doi: 10.1016/j.micpath.2014.01.001
Isha, A., Yusof, N. A., Shaari, K., Osman, R., Abdullah, S. N. A., and Wong, M. Y. (2020). Metabolites identification of oil palm roots infected with Ganoderma boninense using GC–MS-based metabolomics. Arab. J. Chem. 13, 6191–6200. doi: 10.1016/j.arabjc.2020.05.026
Jones, J. D. G., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Jordá, L., Coego, A., Conejero, V., and Vera, P. (1999). A genomic cluster containing four differentially regulated subtilisin-like processing protease genes is in tomato plants. J. Biol. Chem. 274, 2360–2365. doi: 10.1074/JBC.274.4.2360
Jorda, L., Conejero, V., and Vera, P. (2000). Characterization of P69E and P69F, two differentially regulated genes encoding new members of the subtilisin-like proteinase family from tomato plants. Plant Physiol. 122, 67–74. doi: 10.1104/pp.122.1.67
Juul-Madsen, H. R., Viertlböeck, B., Härtle, S., Smith, A. L., and Göbel, T. W. (2013). Chapter 7-Innate Immune Responses. Avian Immunology: Second Edition, Eds. K. A. Schat, B. Kaspers and P. Kaiser, 121–147.
Khademvatan, S., Neisi, N., Maraghi, S., and Saki, J. (2011). Diagnosis and identification of Leishmania spp. from giemsa-stained slides, by real-time PCR and melting curve analysis in south-west of Iran. Ann. Trop. Med. Parasitol. 105, 559–565. doi: 10.1179/2047773211Y.0000000014
Krarup, A., Wallis, R., Presanis, J. S., Gál, P., and Sim, R. B. (2007). Simultaneous activation of complement and coagulation by MBL-associated serine protease 2. PLoS One 2:e623. doi: 10.1371/journal.pone.0000623
Ksiazek, M., Mizgalska, D., Enghild, J. J., Scavenius, C., Thogersen, I. B., and Potempa, J. (2015). Miropin, a novel bacterial serpin from the periodontopathogen Tannerella forsythia, inhibits a broad range of proteases by using different peptide bonds within the reactive center loop. J. Biol. Chem. 290, 658–670. doi: 10.1074/jbc.M114.601716
Lavoie, E. G., Wangdi, T., and Kazmierczak, B. I. (2011). Innate immune responses to Pseudomonas aeruginosa infection. Microbes Infect. 13, 1133–1145. doi: 10.1016/j.micinf.2011.07.011
Lee, N., Gannavaram, S., Selvapandiyan, A., and Debrabant, A. (2007). Characterization of metacaspases with trypsin-like activity and their putative role in programmed cell death in the protozoan parasite Leishmania. Eukaryot. Cell 6, 1745–1757. doi: 10.1128/EC.00123-07
Levy, D. J., Goundry, A., Laires, R. S. S., Costa, T. F. R., Novo, C. M., Grab, D. J., et al. (2021). Role of the inhibitor of serine peptidase 2 (ISP2) of Trypanosoma brucei rhodesiense in parasite virulence and modulation of the inflammatory responses of the host. PLoS Negl. Trop. Dis. 15:e0009526. doi: 10.1371/journal.pntd.0009526
Li, J., Xu, Y., Feng, J., Zhong, M., Xie, Q., Peng, T., et al. (2020). Tamlana fucoidanivorans sp. nov., isolated from algae collected in China. Int. J. Syst. Evol. Microbiol. 70, 1496–1502. doi: 10.1099/ijsem.0.003850
Lima, A. P. C. A., and Mottram, J. C. (2010). Trypanosomatid-encoded inhibitors of peptidases: unique structural features and possible roles as virulence factors. Open Parasitol. J. 4, 132–138. doi: 10.2174/1874421401004010132
Liu, J., Yang, L. L., Xu, C. K., Xi, J. Q., Yang, F. X., Zhou, F., et al. (2012). Sphingobacterium nematocida sp. nov., a nematicidal endophytic bacterium isolated from tobacco. Int. J. Syst. Evol. Microbiol. 62, 1809–1813. doi: 10.1099/ijs.0.033670-0
Mardaneh, J., Soltan Dallal, M. M., Taheripoor, M., and Rajabi, Z. (2014). Isolation, identification and antimicrobial susceptibility pattern of Tatumella ptyseos strains isolated from powdered infant formula Milk consumed in neonatal intensive care unit: first report from Iran. Jundishapur J. Microbiol. 7:e10608. doi: 10.5812/jjm.10608
McGrath, M. E., Erpel, T., Bystroff, C., and Fletterick, R. J. (1994). Macromolecular chelation as an improved mechanism of protease inhibition: structure of the ecotin-trypsin complex. EMBO J. 13, 1502–1507. doi: 10.1002/j.1460-2075.1994.tb06411.x
McGrath, M. E., Hines, W. M., Sakanari, J. A., Fletterick, R. J., and Craik, C. S. (1991). The sequence and reactive site of ecotin: A general inhibitor of pancreatic serine proteases from Escherichia coli. J. Biol. Chem. 266, 6620–6625. doi: 10.1016/s0021-9258(18)38162-6
Minh, B. Q., Schmidt, H. A., Chernomor, O., Schrempf, D., Woodhams, M. D., Von Haeseler, A., et al. (2020). IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37, 1530–1534. doi: 10.1093/molbev/msaa015
Misas-Villamil, J. C., van der Hoorn, R. A. L., and Doehlemann, G. (2016). Papain-like cysteine proteases as hubs in plant immunity. New Phytol. 212, 902–907. doi: 10.1111/nph.14117
Mistry, J., Chuguransky, S., Williams, L., Qureshi, M., Salazar, G. A., Sonnhammer, E. L. L., et al. (2021). Pfam: the protein families database in 2021. Nucleic Acids Res. 49, D412–D419. doi: 10.1093/nar/gkaa913
Morrison, L. S., Goundry, A., Faria, M. S., Tetley, L., Eschenlauer, S. C., Westrop, G. D., et al. (2012). Ecotin-like serine peptidase inhibitor ISP1 of Leishmania major plays a role in flagellar pocket dynamics and promastigote differentiation. Cell. Microbiol. 14, 1271–1286. doi: 10.1111/j.1462-5822.2012.01798.x
Myint, S. L., Zlatkov, N., Aung, K. M., Toh, E., Sjöström, A., Nadeem, A., et al. (2021). Ecotin and LamB in Escherichia coli influence the susceptibility to type VI secretion-mediated interbacterial competition and killing by vibrio cholerae. Biochim. Biophys. Acta - Gen. Subj. 1865:129912. doi: 10.1016/j.bbagen.2021.129912
Nagy, Z. A., Szakács, D., Boros, E., Héja, D., Vígh, E., Sándor, N., et al. (2019). Ecotin, a microbial inhibitor of serine proteases, blocks multiple complement dependent and independent microbicidal activities of human serum. PLoS Pathog. 15:e1008232. doi: 10.1371/journal.ppat.1008232
Ondov, B. D., Bergman, N. H., and Phillippy, A. M. (2011). Interactive metagenomic visualization in a web browser. BMC Bioinformatics 12:385. doi: 10.1186/1471-2105-12-385
Pál, G., Szilágyi, L., and Gráf, L. (1996). Stable monomeric form of an originally dimeric serine proteinase inhibitor, ecotin, was constructed via site directed mutagenesis. FEBS Lett. 385, 165–170. doi: 10.1016/0014-5793(96)00376-6
Peña, M. S., Cabral, G. C., Fotoran, W. L., Perez, K. R., and Stolf, B. S. (2017). Metacaspase-binding peptide inhibits heat shock-induced death in Leishmania (L.) amazonensis. Cell Death Dis. 8:e2645. doi: 10.1038/cddis.2017.59
Perona, J. J., Tsu, C. A., Craik, C. S., and Fletterick, R. J. (1997). Crystal structure of an ecotin-collagenase complex suggests a model for recognition and cleavage of the collagen triple helix. Biochemistry 36, 5381–5392. doi: 10.1021/bi9617522
Rajan, S., and Saiman, L. (2002). Pulmonary infections in patients with cystic fibrosis. Semin. Respir. Infect. 17, 47–56. doi: 10.1053/srin.2002.31690
Rawlings, N. D., Barrett, A. J., Thomas, P. D., Huang, X., Bateman, A., and Finn, R. D. (2018). The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 46, D624–D632. doi: 10.1093/nar/gkx1134
Ryan, C. A., and Pearce, G. (2003). Systemins: A functionally defined family of peptide signals that regulate defensive genes in Solanaceae species. Proc. Natl. Acad. Sci. U. S. A. 100, 14577–14580. doi: 10.1073/pnas.1934788100
Salimiyan Rizi, K., Ghazvini, K., and Farsiani, H. (2019). Clinical and pathogenesis overview of Enterobacter infections. Rev. Clin. Med. 6, 146–154.
Schaaf, B., Wieghorst, A., Aries, S. P., Dalhoff, K., and Braun, J. (2000). Neutrophil inflammation and activation in bronchiectasis: comparison with pneumonia and idiopathic pulmonary fibrosis. Respiration 67, 52–59. doi: 10.1159/000029463
Schaller, A., Stintzi, A., and Graff, L. (2012). Subtilases - versatile tools for protein turnover, plant development, and interactions with the environment. Physiol. Plant. 145, 52–66. doi: 10.1111/j.1399-3054.2011.01529.x
Seymour, J. L., Lindquist, R. N., Dennis, M. S., Lazarus, R. A., Moffat, B., Yansura, D., et al. (1994). Ecotin is a potent anticoagulant and reversible tight-binding inhibitor of factor Xa. Biochemistry 33, 3949–3958. doi: 10.1021/bi00179a022
Shadab, M., Jha, B., Asad, M., Deepthi, M., Kamran, M., and Ali, N. (2017). Apoptosis-like cell death in Leishmania donovani treated with KalsomeTM10, a new liposomal amphotericin B. PLoS One 12:e0171306. doi: 10.1371/journal.pone.0171306
Shin, D. H., Song, H. K., Seong, I. S., Lee, C. S., Chung, C. H., and Suh, S. W. (1996). Crystal structure analyses of uncomplexed ecotin in two crystal forms: implications for its function and stability. Protein Sci. 5, 2236–2247. doi: 10.1002/pro.5560051110
Silverman, G. A., Bird, P. I., Carrell, R. W., Church, F. C., Coughlin, P. B., Gettins, P. G. W., et al. (2001). The serpins are an expanding superfamily of structurally similar but functionally diverse proteins. J. Biol. Chem. 276, 33293–33296. doi: 10.1074/jbc.R100016200
Spence, M. A., Mortimer, M. D., Buckle, A. M., Minh, B. Q., and Jackson, C. J. (2021). A comprehensive phylogenetic analysis of the serpin superfamily. Mol. Biol. Evol. 38, 2915–2929. doi: 10.1093/molbev/msab081
Spiers, A. J., Buckling, A., and Rainey, P. B. (2000). The causes of pseudomonas diversity. Microbiology 146, 2345–2350. doi: 10.1099/00221287-146-10-2345
Stanton, B. A., Hampton, T. H., and Ashare, A. (2020). SARS-CoV-2 (COVID-19) and cystic fibrosis. Am. J. Physiol. - Lung Cell. Mol. Physiol. 319, L408–L415. doi: 10.1152/ajplung.00225.2020
Stein, P. E., and Carrell, R. W. (1995). What do dysfunctional serpins tell us about molecular mobility and disease? Nat. Struct. Mol. Biol. 2, 96–113. doi: 10.1038/nsb0295-96
Subramaniam, S., Müller, V. S., Hering, N. A., Mollenkopf, H., Becker, D., Heroven, A. K., et al. (2019). Contribution of the Cpx envelope stress system to metabolism and virulence regulation in salmonella enterica serovar typhimurium. PLoS One 14:e0211584. doi: 10.1371/journal.pone.0211584
Szallies, A., Kubata, B. K., and Duszenko, M. (2002). A metacaspase of Trypanosoma brucei causes loss of respiration competence and clonal death in the yeast Saccharomyces cerevisiae. FEBS Lett. 517, 144–150. doi: 10.1016/S0014-5793(02)02608-X
Tamura, K., Stecher, G., and Kumar, S. (2021). MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. doi: 10.1093/molbev/msab120
Thiel, S., Vorup-Jensen, T., Stover, C. M., Schwaeble, W., Laursen, S. B., Poulsen, K., et al. (1997). A second serine protease associated with mannan-binding lectin that activates complement. Nature 386, 506–510. doi: 10.1038/386506a0
Thomas, E. L., Lehrer, R. I., and Rest, R. F. (1988). Human neutrophil antimicrobial activity. Clin. Infect. Dis. 10, S450–S456. doi: 10.1093/cid/10.Supplement_2.S450
Thomas, C., Nothaft, H., Yadav, R., Fodor, C., Alemka, A., Oni, O., et al. (2020). Characterization of ecotin homologs from campylobacter rectus and campylobacter showae. PLoS One 15:e0244031. doi: 10.1371/journal.pone.0244031
Tian, M., Benedetti, B., and Kamoun, S. (2005). A second Kazal-like protease inhibitor from Phytophthora infestans inhibits and interacts with the apoplastic pathogenesis-related protease P69B of tomato. Plant Physiol. 138, 1785–1793. doi: 10.1104/pp.105.061226
Tian, M., Huitema, E., Da Cunha, L., Torto-Alalibo, T., and Kamoun, S. (2004). A Kazal-like extracellular serine protease inhibitor from Phytophthora infestans targets the tomato pathogenesis-related protease P69B. J. Biol. Chem. 279, 26370–26377. doi: 10.1074/jbc.M400941200
Tracz, D. M., Gilmour, M. W., Mabon, P., Beniac, D. R., Hoang, L., Kibsey, P., et al. (2015). Tatumella saanichensis sp. nov., isolated from a cystic fibrosis patient. Int. J. Syst. Evol. Microbiol. 65, 1959–1966. doi: 10.1099/ijs.0.000207
Tseng, B. S., Reichhardt, C., Merrihew, G. E., Araujo-Hernandez, S. A., Harrison, J. J., MacCoss, M. J., et al. (2018). A biofilm matrix-associated protease inhibitor protects Pseudomonas aeruginosa from proteolytic attack. MBio 9:e00543–18. doi: 10.1128/mBio.00543-18
Uren, A. (2000). Identification of Paracaspases and Metacaspases two ancient families of caspase-like proteins, one of which plays a key role in MALT lymphoma. Mol. Cell 6, 961–967. doi: 10.1016/s1097-2765(00)00094-0
Van Der Hoorn, R. A. L., and Klemenčič, M. (2021). Plant proteases: from molecular mechanisms to functions in development and immunity. J. Exp. Bot. 72, 3337–3339. doi: 10.1093/JXB/ERAB129
van Griensven, J., and Diro, E. (2012). Visceral Leishmaniasis. Infect. Dis. Clin. N. Am. 26, 309–322. doi: 10.1016/j.idc.2012.03.005
Vartapetian, A. B., Tuzhikov, A. I., Chichkova, N. V., Taliansky, M., and Wolpert, T. J. (2011). A plant alternative to animal caspases: subtilisin-like proteases. Cell Death Differ. 18, 1289–1297. doi: 10.1038/cdd.2011.49
Verma, S., Das, S., Mandal, A., Ansari, M. Y., Kumari, S., Mansuri, R., et al. (2017). Role of inhibitors of serine peptidases in protecting Leishmania donovani against the hydrolytic peptidases of sand fly midgut. Parasit. Vectors 10:303. doi: 10.1186/s13071-017-2239-9
Verma, S., Mandal, A., Ansari, M. Y., Kumar, A., Abhishek, K., Ghosh, A. K., et al. (2018). Leishmania donovani inhibitor of serine peptidases 2 mediated inhibition of lectin pathway and upregulation of C5aR signaling promote parasite survival inside host. Front. Immunol. 9:0063. doi: 10.3389/fimmu.2018.00063
Vogt, S. L., Scholz, R., Peng, Y., Guest, R. L., Scott, N. E., Woodward, S. E., et al. (2019). Characterization of the Citrobacter rodentium Cpx regulon and its role in host infection. Mol. Microbiol. 111, 700–716. doi: 10.1111/mmi.14182
Wang, G., Chen, H., Xia, Y., Cui, J., Gu, Z., Song, Y., et al. (2013). How are the non-classically secreted bacterial proteins released into the extracellular milieu? Curr. Microbiol. 67, 688–695. doi: 10.1007/s00284-013-0422-6
Wang, S. X., Esmon, C. T., and Fletterick, R. J. (2001). Crystal structure of thrombin-ecotin reveals conformational changes and extended interactions. Biochemistry 40, 10038–10046. doi: 10.1021/bi010712h
Wang, S. X., Hur, E., Sousa, C. A., Brinen, L., Slivka, E. J., and Fletterick, R. J. (2003). The extended interactions and Gla domain of blood coagulation factor Xa. Biochemistry 42, 7959–7966. doi: 10.1021/bi027320a
Waugh, S. M., Harris, J. L., Fletterick, R., and Craik, C. S. (2000). The structure of the pro-apoptotic protease granzyme B reveals the molecular determinants of its specificity. Nat. Struct. Biol. 7, 762–765. doi: 10.1038/78992
Webb, J. S., Thompson, L. S., James, S., Charlton, T., Tolker-Nielsen, T., Koch, B., et al. (2003). Cell death in Pseudomonas aeruginosa biofilm development. J. Bacteriol. 185, 4585–4592. doi: 10.1128/JB.185.15.4585-4592.2003
Weinberg, J. B., Alexander, B. D., Majure, J. M., Williams, L. W., Kim, J. Y., Vandamme, P., et al. (2007). Burkholderia glumae infection in an infant with chronic granulomatous disease. J. Clin. Microbiol. 45, 662–665. doi: 10.1128/JCM.02058-06
Witko-Sarsat, V., Halbwachs-Mecarelli, L., Schuster, A., Nusbaum, P., Ueki, I., Canteloup, S., et al. (1999). Proteinase 3, a potent secretagogue in airways, is present in cystic fibrosis sputum. Am. J. Respir. Cell Mol. Biol. 20, 729–736. doi: 10.1165/ajrcmb.20.4.3371
Yaghi, J., Fattouh, N., Akkawi, C., El Chamy, L., Maroun, R. G., and Khalil, G. (2020). Unusually high prevalence of Cosecretion of ambler class A and B Carbapenemases and nonenzymatic mechanisms in multidrug-resistant clinical isolates of Pseudomonas aeruginosa in Lebanon. Microb. Drug Resist. 26, 150–159. doi: 10.1089/mdr.2019.0040
Yang, S. Q., Wang, C. I., Gillmor, S. A., Fletterick, R. J., and Craik, C. S. (1998). Ecotin: A serine protease inhibitor with two distinct and interacting binding sites. J. Mol. Biol. 279, 945–957. doi: 10.1006/jmbi.1998.1748
Keywords: protease inhibition, virulence, ecotin, host-associated bacteria, serpin
Citation: De Meyer F and Carlier A (2023) Ecotin: A versatile protease inhibitor of bacteria and eukaryotes. Front. Microbiol. 14:1114690. doi: 10.3389/fmicb.2023.1114690
Edited by:
Daniel Yero, Autonomous University of Barcelona, SpainReviewed by:
Christine M. Szymanski, University of Alberta, CanadaNicole Thielens, UMR5075 Institut de Biologie Structurale (IBS), France
Copyright © 2023 De Meyer and Carlier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aurélien Carlier, ✉ YXVyZWxpZW4uY2FybGllckB1Z2VudC5iZQ==