
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 25 April 2023
Sec. Food Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1112709
This article is part of the Research Topic Innovative Microbial Technologies for Future and Sustainable Food Science View all 12 articles
 Ying Chen1†
Ying Chen1† Shou-Yue Xia2†
Shou-Yue Xia2† Fu-Xia Ru2†
Fu-Xia Ru2† Jun-Jie Feng2†
Jun-Jie Feng2† Ji Tao2
Ji Tao2 Zhi-Yuan Wei2Xiu Li3Chengjia Qian4
Zhi-Yuan Wei2Xiu Li3Chengjia Qian4 Qiong Lin1*
Qiong Lin1* Jian-Huan Chen2*
Jian-Huan Chen2*Purpose: Helicobacter pylori (HP) infection is an identified risk factor for pediatric chronic gastritis (PCG), but its impact on gastric juice microbiota (GJM) remains to be further elucidated in PCG. This study aimed to analyze and compare the microbial communities and microbial interactive networks of GJM in PCG that clinically tested positive and negative for HP (HP+ and HP−, respectively).
Methods: A total of 45 PCG patients aged from 6 to 16 years were recruited, including 20 HP+ and 25 HP− patients tested by culture and rapid urease test. Gastric juice samples were collected from these PCG patients and subjected to high-throughput amplicon sequencing and subsequent analysis of 16S rRNA genes.
Results: While no significant change in alpha diversity, significant differences in beta diversity were observed between HP+ and HP− PCG. At the genus level, Streptococcus, Helicobacter, and Granulicatella were significantly enriched in HP+ PCG, whereas Campylobacter and Absconditabacteriales (SR1) were significantly enriched in HP− PCG. Network analysis showed that Streptococcus was the only genus positively correlated with Helicobacter (r = 0.497) in the GJM network of overall PCG. Moreover, compared to HP− PCG, HP+ PCG showed a reduction in microbial network connectivity in GJM. Netshift analysis identified driver microbes including Streptococcus and other four genera, which substantially contributed to the GJM network transition from HP− PCG to HP+ PCG. Furthermore, Predicted GJM function analysis indicated up-regulated pathways related to the metabolism of nucleotides, carbohydrates, and L-Lysine, the urea cycle, as well as endotoxin peptidoglycan biosynthesis and maturation in HP+ PCG.
Conclusion: GJM in HP+ PCG exhibited dramatically altered beta diversity, taxonomic structure, and function, with reduced microbial network connectivity, which could be involved in the disease etiology.
Chronic gastritis is a common health problem in children (Sipponen and Maaroos, 2015). Helicobacter pylori (HP) infection is an important cause of chronic gastritis, which occurs generally in childhood (Sabbagh et al., 2019), and is associated with peptic ulcer, gastric atrophy, intestinal metaplasia, and gastric cancer. It is important to study the impact of HP for the understanding of the etiology of pediatric chronic gastritis (PCG).
HP plays a pivotal role in the gastric microbiota. The stomach is a special area in the digestive tract, with gastric acid secretion constituting its unique ecological environment and characteristic microbial community (Caguazango, 2020). HP could promote inflammation in the gastric mucosa, and leads to changes in the gastric mucosa microbiota (GMM) in PCG (Zheng et al., 2021). In addition, the gastric juice microbiota (GJM) is typically affected by various factors such as diet (Martinsen et al., 2019), which could result in greater variation compared to GMM. The diversity of GJM has been reported to be higher than that of GMM. Sung et al. found that the relative abundance of Actinobacteria, Bacteroidetes, and Firmicutes was relatively higher in GJM than in GMM (Sung et al., 2016). Besides, FLéJOU et al. found that non-HP bacteria could have explicit effects on gastritis (Fléjou et al., 1990), and Streptococcus showed a higher relative abundance in HP− cancer patients. (Sohn et al., 2017). It is also noteworthy that the collection of gastric juice is less invasive compared with gastric mucosal biopsy.
In the current study, we recruited a cohort of PCG and analyzed the impact of HP abundance on the microbial communities and co-occurrence networks of GJM.
This study was approved by the Medical Ethics Committee of Jiangnan University and conducted in accordance with the Declaration of Helsinki. A total of 45 PCG were recruited in the current study, including 27 boys and 18 girls, with an average age of 9.03 ± 0.93 years old. Informed consent was obtained from all of the participants after explanation of the nature of the study. Gastritis diagnosis was made based on endoscopic observation and pathological features of gastric atrophy. Exclusion criteria included treatment with antibiotics or proton pump inhibitors (PPIs) within 6 months; serious mental or organ damage; or poor patient compliance. HP infection status of these PCG patients was tested using gastric mucosal biopsy and rapid urease tests, and confirmed with HP culture on brain heart infusion agar plate supplemented with blood, inoculated with gastric mucosal biopsy tissue homogenate in a microaerophilic environment (85% N2, 10% CO2, 5% O2) at 37°C for 3 days.
Gastric juice samples were collected from all participants with sterilized 1.5-mL tubes containing ethanol on ice, and stored at −80°C until analysis. The V4 hypervariable region of the 16S rRNA gene was amplified from the gastric fluid samples with V4-specific primers 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). Amplicons were checked using the 2% agarose gel, and purified using GeneJET Gel Extraction Kit (Thermo Fisher Scientific, Waltham). Constructed libraries were sequenced on an Ion S5XL sequencer (Thermo Fisher Scientific, Waltham) with a single-end 400-bp read length configuration following standard protocols provided by the manufacturer.
The 16S rRNA gene sequencing reads were analyzed using the QIIME2 (version 2020.11.0) analysis pipeline as previously described (Bolyen et al., 2019; Wei et al., 2022). In brief, low-quality and chimeric sequences were filtered using DADA2. Amplicon sequence variant (ASV) tables at 100% sequence similarity were generated. Taxonomy classification was assigned to ASVs using q2-feature-classifier and the SILVA database (release r132) at a 99% similarity cutoff (Glockner et al., 2017). Microbiota diversity was analyzed using QIIME2: alpha diversity metrics including Pielou’s evenness, the Chao1, Shannon and Simpson’s indices, and beta diversity including weighted/unweighted UniFrac distances, and Bray-Curtis distances followed by non-metric multidimensional scaling (NMDS) analysis and Principal Coordinate Analysis (PCoA). The linear discriminant analysis (LDA) effect size (LEfSe) algorithm was used to identify group-enriched taxa (Segata et al., 2011). Phylogenetic cladograms were drawn using GraPhlAn (version 1.1.3). PICRUSt2 was used to predict microbiota function based on the 16S rRNA sequencing data (Douglas et al., 2020).
ASVs with average relative abundance >0.1% of the microbiome were subjected to correlation analysis of their occurrence patterns (Barberan et al., 2012; Cardinale et al., 2015). The SparCC algorithm was used to estimate the correlations among gut microbes(Friedman and Alm, 2012). 1,000 bootstrap replicates were applied to calculate the pseudo p-values, and correlations with |correlation coefficient |(r)| > 0.2 and p < 0.01 were considered significant. For each genus with significant SparCC correlations, its degree was calculated as an indicator of its weight in the network by summing up its edges. The SparCC network was further constructed using Cytoscape 3.9.1 (Shannon et al., 2003). In addition, NetShift 2 was used to evaluate potential driver microbes as described (Kuntal et al., 2019), and to calculate Neighbor shift (NESH) scores to quantify increased interactions in the network shift.
The demographic and clinical characteristics of 45 PGC patients were summarized in Table 1. Endoscopic findings showed a significantly higher proportion of gastritis with additional clinical conditions (GAC) including duodenitis, bile reflux, duodenal ulcer, and gastric ulcer in HP+ PCG than in HP− PCG (OR = 4.97, 95% CI: 0.97–34.69, p = 0.040). No significant difference was found in age, gender, and other clinical features between HP+ and HP− PCG (all p > 0.05).
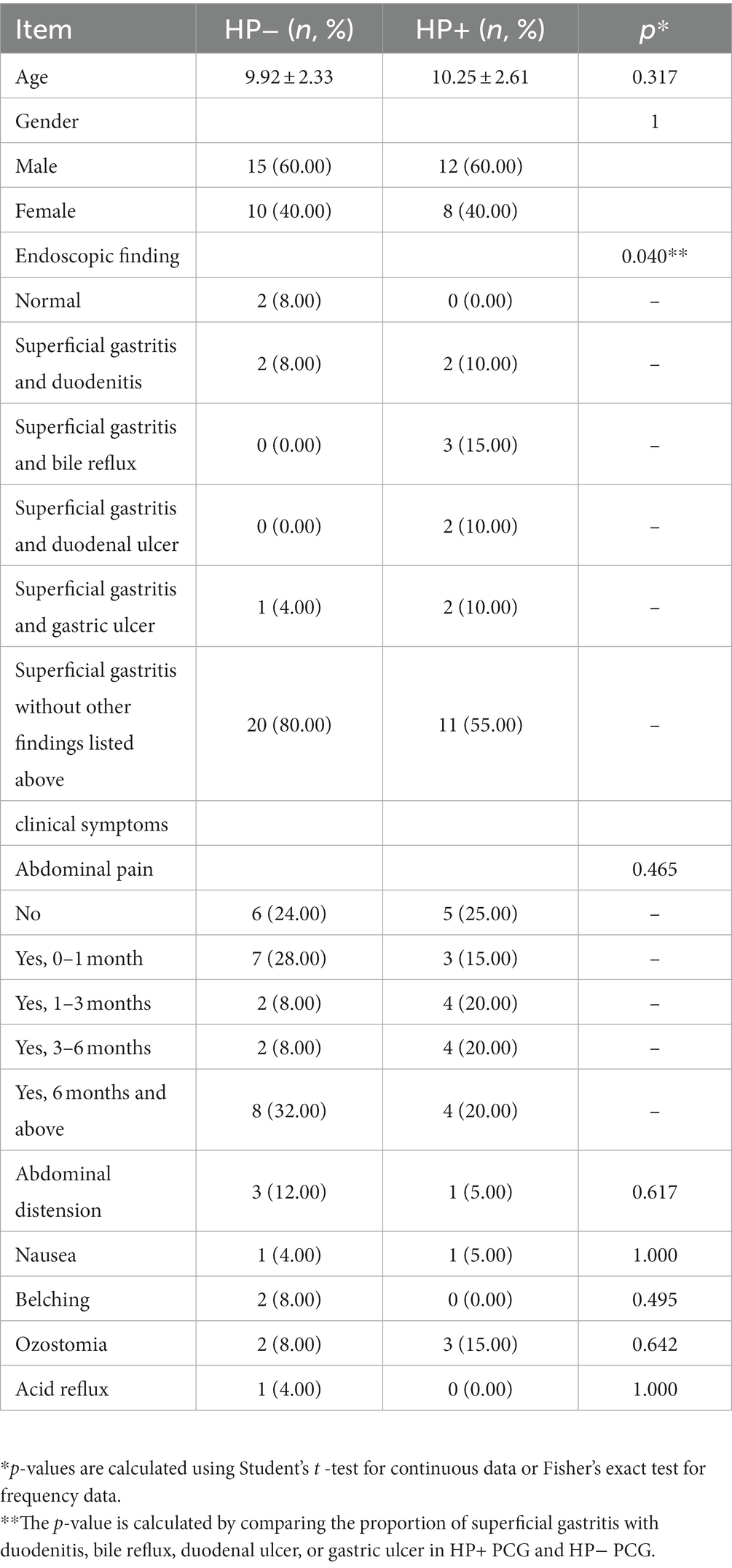
Table 1. Demographic and clinical characteristics of HP− and HP+ PCG in the current study.
The results of the rarefaction curve shown in Supplementary Figure S1 indicated that our current study’s sequencing data could sufficiently capture the majority of the microbial taxa. In addition, alpha diversity results showed no significant difference (all p > 0.05) in the Shannon, Simpson, evenness, and Chao1 (Figures 1A–D) indices between the two groups. Nevertheless, PCoA and NMDS results based on Bray-Curtis distances suggested a significant difference in beta diversity between HP+ and HP− PCG (PERMANOVA p = 0.007 and 0.007 respectively), although the two groups might not be completely separated (Figures 1E,F).
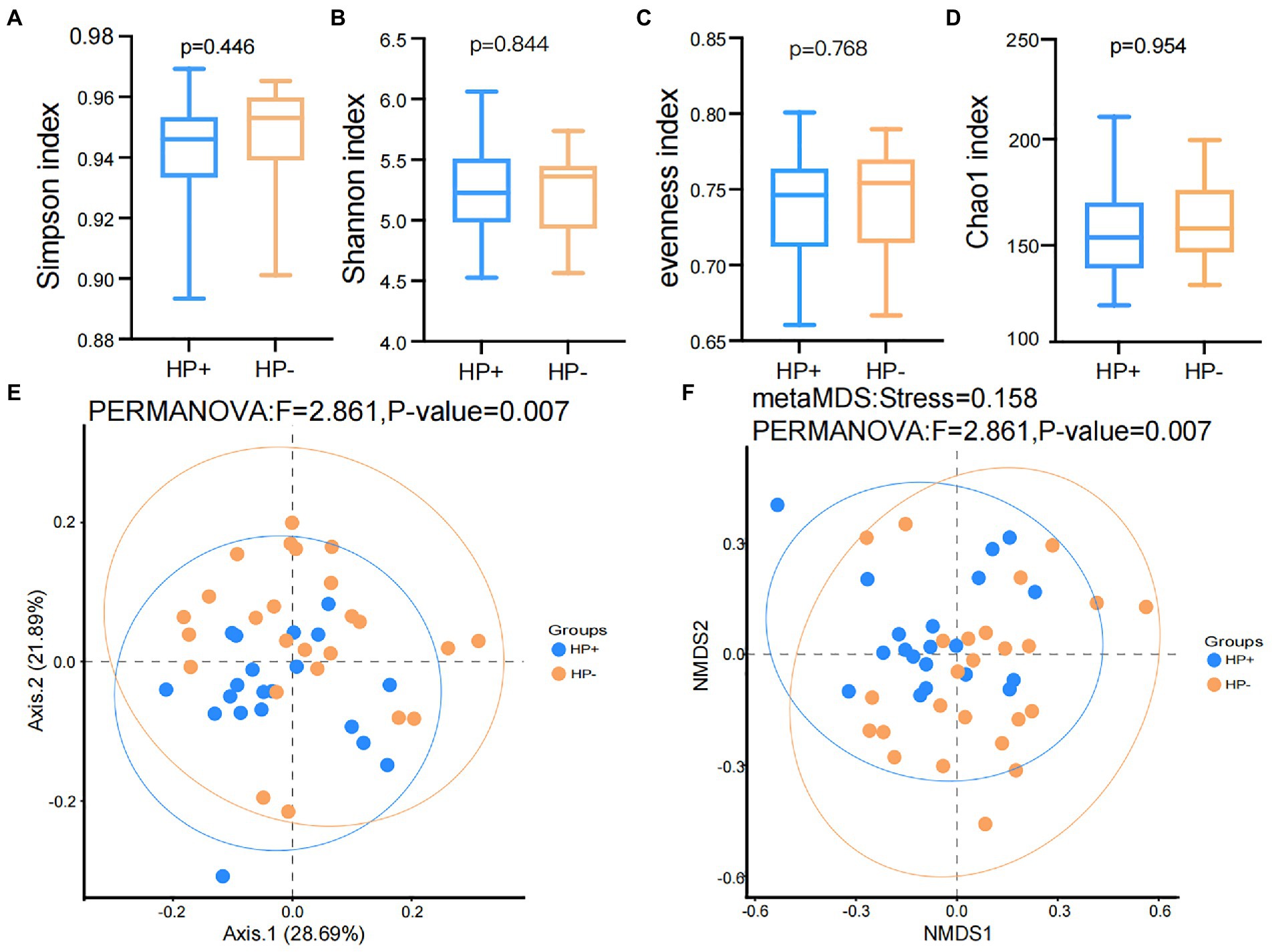
Figure 1. Comparison of Alpha and beta diversities between HP− and HP+ PCG. (A) Simpson index. (B) Shannon index. (C) Eveness index. (D) Chao1 index. (E) PCoA based on Bray-Curtis distances. (F) NMDS on Bray-Curtis distances.
GJM in both HP+ PCG and HP− PCG patients was dominated by 8 phyla, including Firmicutes, Bacteroidetes, Proteobacteria, Fusobacteriota, Actinobacteriota, Campilobacterota, Patescibacteria, and Spirochaetota (Figure 2A). Prevotellaceae, Streptococcaceae, Neisseriaceae, Pasteurellaceae, Fusobacteriaceae, Carnobacteriaceae, Veillonellaceae, Micrococcaceae, Porphyromonadaceae, and Gemellaceae were the top 10 families, and Prevotella, Streptococcus, Alloprevotella, Neisseria, Fusobacterium, Haemophilus, Granulicatella, Rothia, Porphyromonas, and Veillonella were the top 10 genera in PCG GJM (Figures 2B,C). LEfSe analysis was then applied to identify the most relevant taxa responsible for differences between the two groups. As shown in the cladogram in Figure 3A, at the phylum level, Firmicutes and Campilobacterota were enriched in HP+ PCG. At the family level, Streptococcaceae, Helicobacteraceae, and Carnobacteriaceae were enriched in HP+ PCG, whereas Campylobacteraceae and Absconditabacteriales (SR1) were enriched in HP− PCG (Figure 3B). At the genus level, Streptococcus, Helicobacter, and Granulicatella were enriched in HP+ PCG, whereas Campylobacter and Absconditabacteriales (SR1) were enriched in HP− PCG (Figure 3C and Supplementary Figure S2).
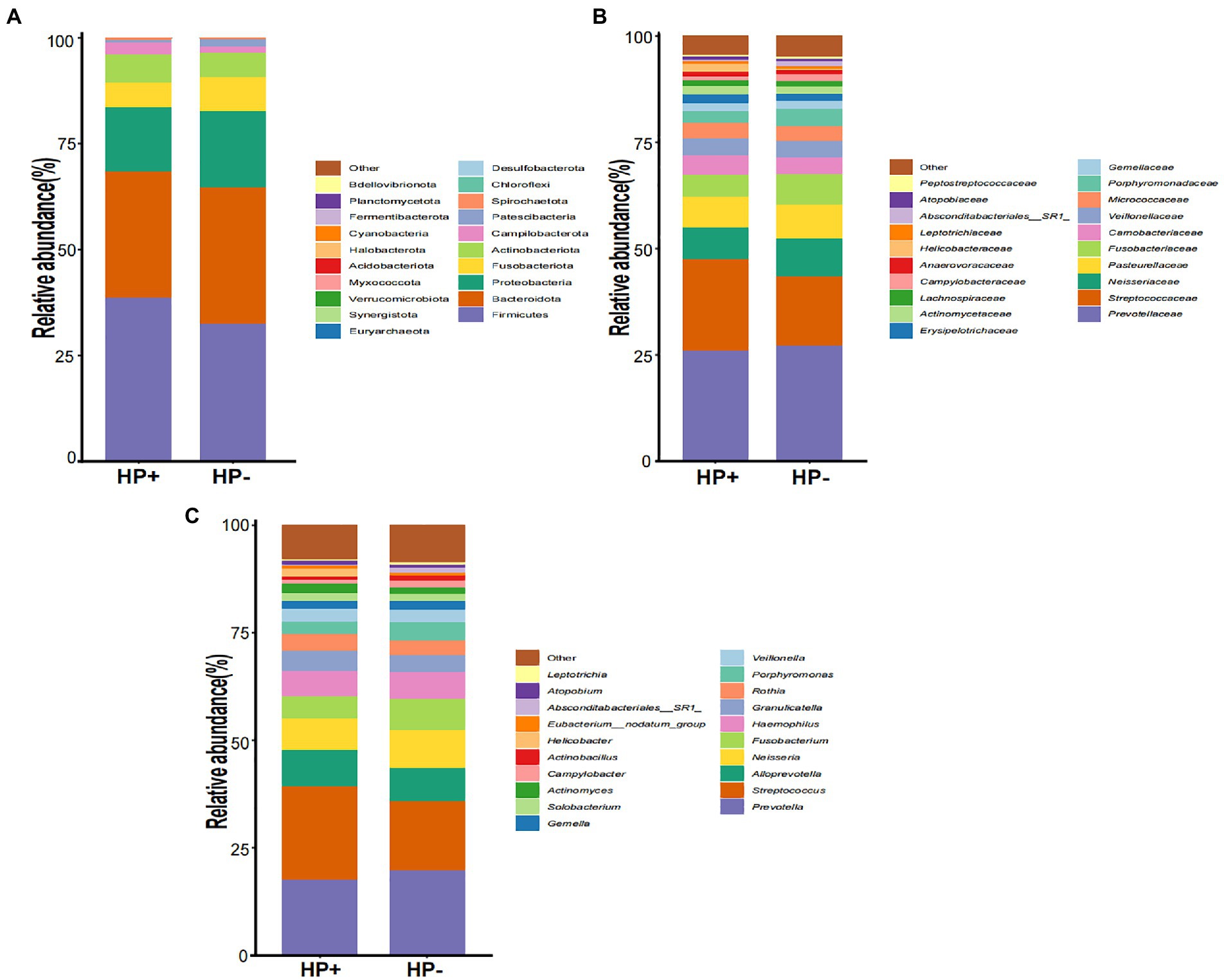
Figure 2. The microbial abundance of GJM in HP− and HP+ PCG. Microbial abundance is shown at the (A) phylum level, (B) family level, (C) and the genus level.
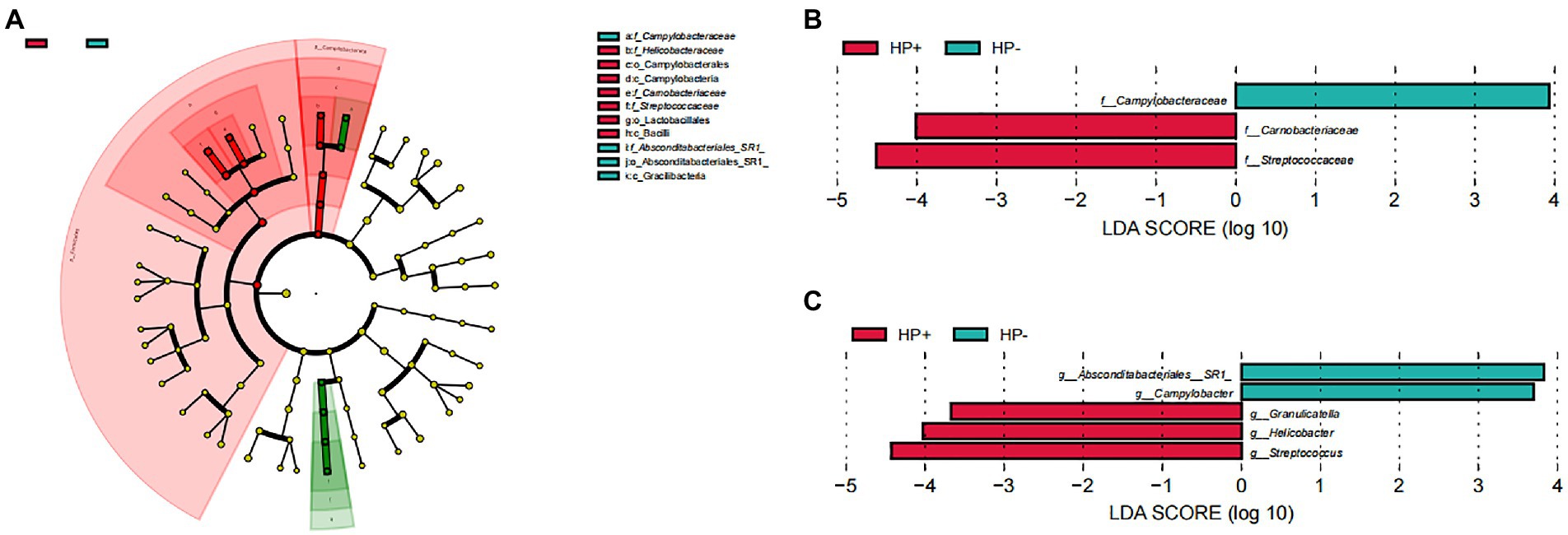
Figure 3. LEfSe analysis of GJM in HP− and HP+ PCG. Phylogenetic cladograms (A), and barplots at (B) the family, and (C) Genus levels are shown.
In addition, as GAC was dramatically increased in HP+ PCG, we further divided the HP+ PCG patients into a group of gastritis only (GO) and a group of GAC according to the endoscopic findings, and compared GJM between the two groups. Our results showed that the genus Absconditabacteriales (SR1) was enriched in GO, whereas genera including Prevotella, Megashaera, and Actinomyces were enriched in GAC (Figure 4).

Figure 4. LEfSe analysis of GJM in GO and GAC subgroups of HP+ PCG.
To explore the interaction among microbes in GJM, we conducted a correlation analysis using the SparCC algorithm and the genus abundance of genera with average relative abundance ≥ 0.1%, and constructed microbial co-occurrence networks with cutoff set as |r| > = 0.2 and p < 0.01 in overall PCG, HP− PCG, and HP+ PCG. In the GJM network of overall PCG, Streptococcus, the most enriched genus in HP+ PCG, was found to be the only genus positively correlated with Helicobacter (r = 0.497) (Figure 5A). It was also positively correlated with Granulicatella, Gemella, Actinomyces, Rothia, Atopobium, and Megasphaera, (r = 0.746, 0.598, 0.504, 0.497, 0.484, 0.483, and 0.465, respectively), and negatively correlated with Absconditabacteriales (SR1), (r = −0.402). No significant correlation between other microbes and Helicobacter was found in GJM networks of either HP+ PCG or HP− PCG. Notably, lower network connectivity was found in HP+ PCG compared to that in HP− PCG or overall PCG (Figures 5B,C, and Supplementary Tables S1–3). Similarly, Actinomyces and Prevotella 6 were the dominant members in overall PCG (Figure 5B). In HP+ PCG, Streptococcus showed positive correlations with Rothia (r = 0.716), Megasphaera (r = 0.708), and Gemella (r = 0.679), (Figure 5C). In HP− PCG, Streptococcus showed positive correlations with Granulicatella (r = 0.783) and Gemella (r = 0.621). Furthermore, Streptococcus was identified as a driver microbe responsible for the microbial changes between HP+ and HP− PCG in Netshift analysis (Table 2, Figure 5D). Additional driver microbes identified by Netshift analysis included Capnocytophaga, Actinomyces, Neisseria, and Megasphaera.
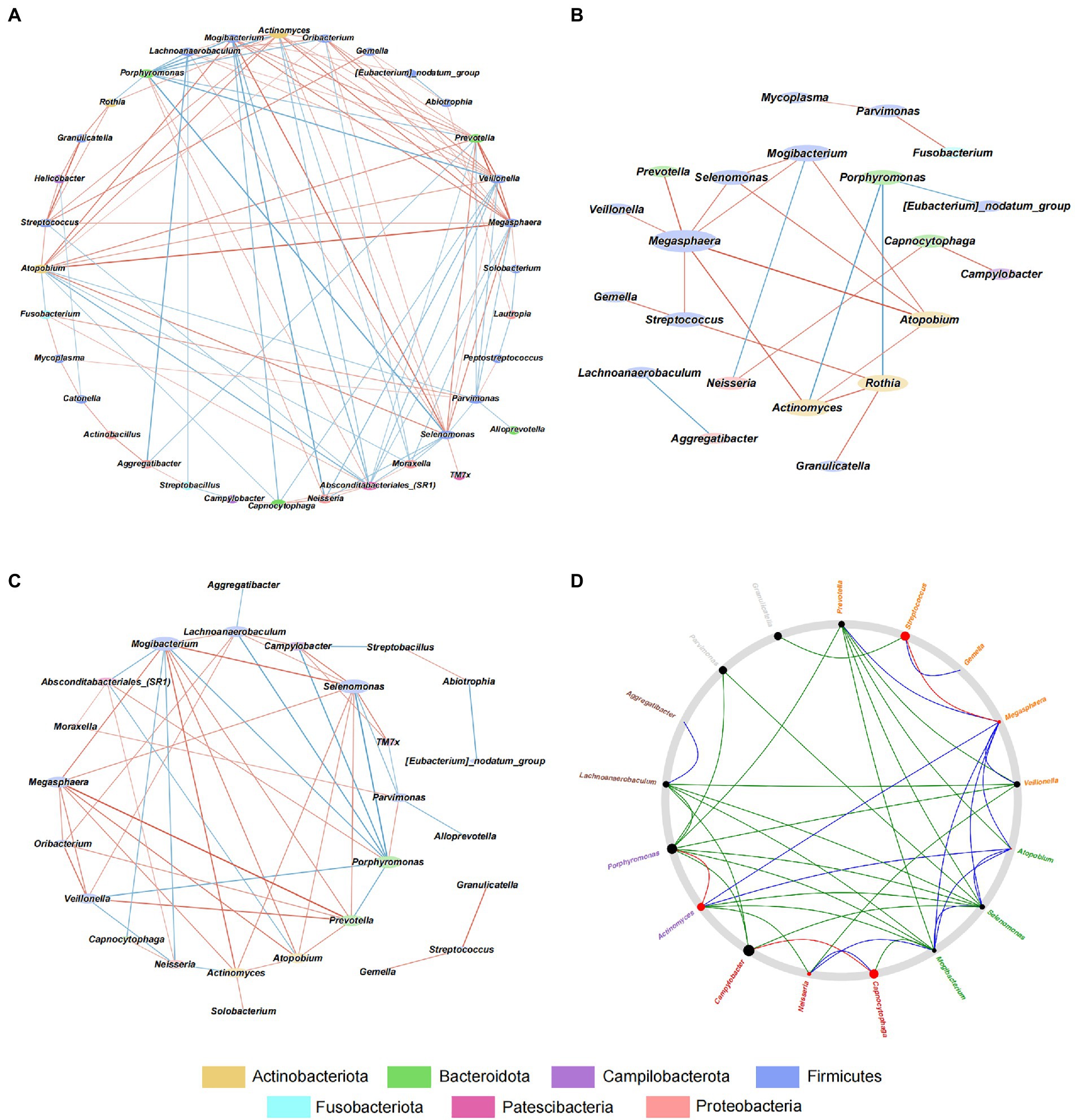
Figure 5. Interactive Network in HP−, HP+, and overall PCG at the genus level. (A) Interactive network analysis of HP− PCG. (B) Interactive network analysis of overall PCG. (C) Interactive Network analysis of HP+ PCG. (D) Netshift analysis of GJM between HP− and HP+ PCG.

Table 2. Genera with the top five NESH score in Netshift analysis.
PICRUSt2 predicted a dramatical difference in GJM function, with a total of 35 Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways differentially abundant between HP+ and HP− PCG (Figure 6). The 30 pathways significantly upregulated in HP+ PCG were mainly related to the metabolism of nucleotides, carbohydrates, and L-Lysine, as well as endotoxin peptidoglycan biosynthesis and maturation. In addition, the urea cycle was also significantly upregulated in HP+ PCG. In contrast, pathways enriched in HP− PCG were related to the biosynthesis of thiamin diphosphate, CMP-3-deoxy-D-manno-octulosonat, flavin, and lipid IVA which was an intermediate in the biosynthetic pathway of lipid A and lipopolysaccharide (LPS).
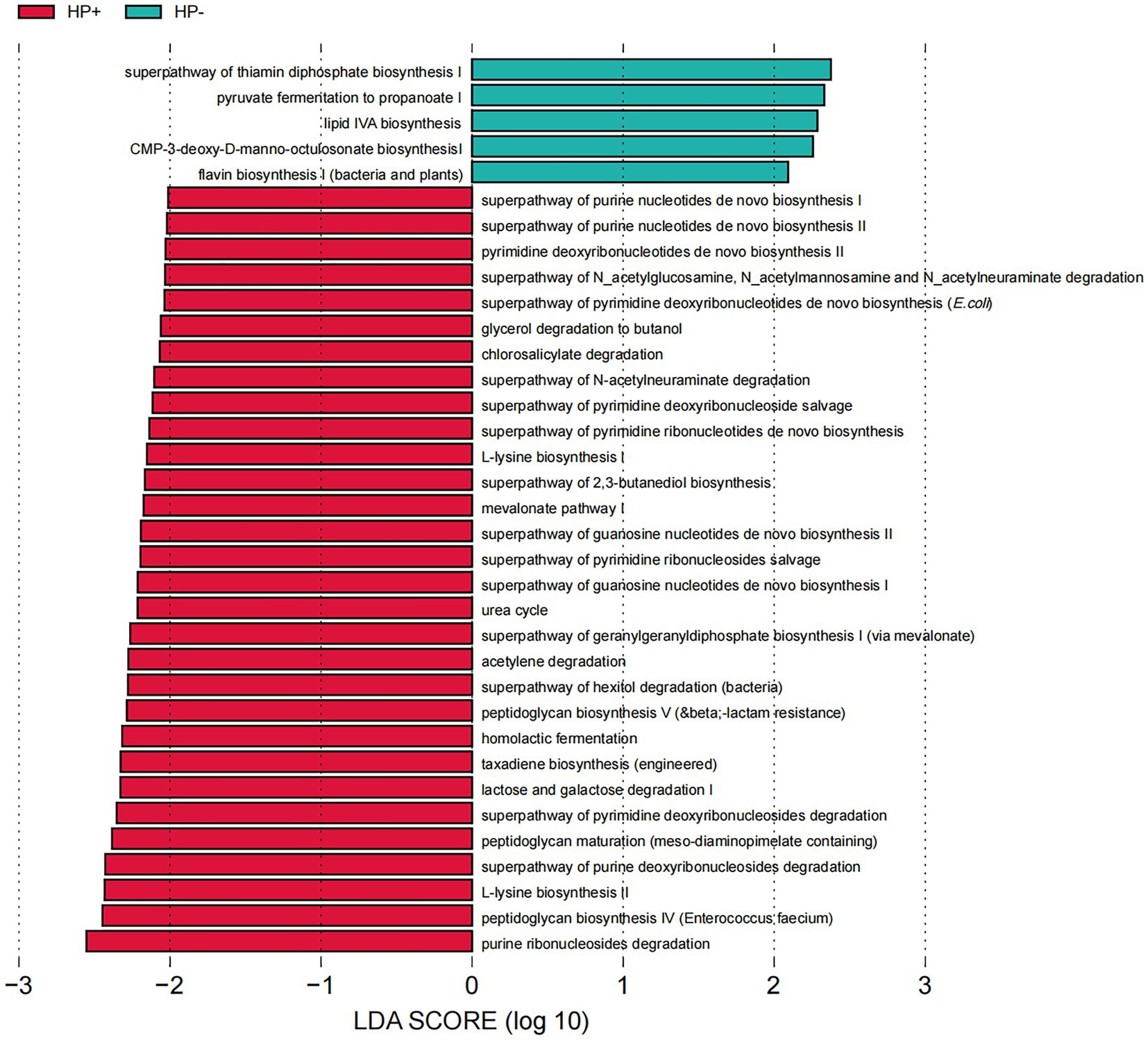
Figure 6. LEfSe analysis of GJM KEGG pathways differentially enriched in HP− and HP+ PCG.
Since Warren and Marshall first cultivated HP from children’s gastric mucosa, after two decades of in-depth research, HP has been identified as a pivotal pathogen of chronic gastritis and peptic ulcer (Kishikawa et al., 2020). More, the influence of HP on the gastric microbiota might contribute to the development of PCG. By analyzing GJM in HP+ and HP− PCG clinically tested by routine methods, our current study showed a substantial impact of HP abundance on GJM composition, microbial interactive network, and function.
By far, most existing studies have focused on the influence of HP on the gastrointestinal microbiota in adults (He et al., 2019; Tao et al., 2020), and there have been only limited studies in children, especially for GJM. Brawner et al. reported that HP infection significantly reduced the alpha diversity of GMM in PCG. Different from such results in GMM, our current study showed no significant difference in alpha diversity of GJM between HP+ and HP− PCG, pointing to different effects of HP on GMM and GJM. Notably, Helicobacter could also be detected in HP−PCG clinically tested via routine methods, yet with an extremely low relative abundance than in HP+ PCG. 16S amplicon sequencing was recently reported with higher sensitivity in detecting HP in the gastric microbiota, by revealing HP existence in some cases that were tested negative for HP via routine methods (Gantuya et al., 2021). Helicobacter is commonly found in human children and non-human primate infants (Wei et al., 2022). Such findings suggested that an extremely low abundance of HP might still exist in samples that test negative by routine clinical methods.
Our results showed a significant difference in beta diversity with dramatic changes in taxonomic structure between HP+ and HP− PCG. The higher abundance of Firmicutes in HP+ PCG was similar to the gut microbiota of asymptomatic children with HP infection (Benavides-Ward et al., 2018). In addition, our study showed that a non-HP bacterium Streptococcus, with a significantly positive correlation with Helicobacter in all PCG, was the most enriched genus in HP+ PCG. Streptococcus is a gram-positive bacterium of spherical shape and has been reported to be involved in several diseases and health issues, such as gastritis (Minalyan et al., 2017). Moreover, our GJM networks revealed that Streptococcus acted as a driver that contributed to the shift of the GJM network from HP− PCG to HP+ PCG. Another non-HP bacterium Granulicatella is associated with atrophic gastritis or intestinal metaplasia in patients following successful HP eradication (Liu et al., 2022). In contrast, Campylobacter and Absconditabacteriales (SR1) were enriched in our HP− PCG patients. Campylobacter infection usually causes diarrhea, fever, and stomach cramps. Absconditabacteriales exhibited significantly lower abundance in the gastric microbiota in HP− Crohn’s disease patients than in control subjects (Ostrowski et al., 2021). Such findings indicated non-HP pathogens could play a potential role in PCG.
The change of non-HP pathogens in the stomach may be due to the change in pH value in gastric juice. Rosen et al. reported that acid suppression resulted in the overgrowth of gastric bacteria such as Staphylococcus and Streptococcus (Rosen et al., 2014). In addition, Streptococcus, Actinomyces, Megasphaera, and Granulicatella were significantly increased in patients receiving proton pump inhibitor treatment (Jackson et al., 2016; Takagi et al., 2018).
Recent studies have demonstrated the importance of microbiota networks in understanding microbiota changes in diseases and aging. The reduced network connectivity observed in HP+ PCG suggested a dramatic change in microbial interaction in the group compared to that of HP− PCG, emphasizing the impact of HP on the microbial community. In addition, Streptococcus showed a significant positive correlation with Helicobacter in overall PCG, and contributed to the network shift from HP− to HP+ PCG by interconnecting with multiple hub or driver microbes. Such findings thus highlighted the importance of non-HP pathogens.
Although no significant difference in clinical symptoms such as abdominal pain and belching between the two groups, endoscopic findings showed a significantly higher proportion of PCG with duodenitis, bile reflux, duodenal ulcer, or gastric ulcer. Such findings were in line with previous reports that HP was a cause of duodenitis (Ohkusa et al., 2003), peptic ulcer, and possibly bile reflux (Ladas et al., 1996). Furthermore, our results also indicated GJM might also contribute to the development of duodenitis, bile reflux, and peptic ulcer.
Gastric acid secretion in the stomach constitutes a characteristic microbial community and its function in the gastric juice (Caguazango, 2020), which could be involved in HP infection and the development of PCG. Our PICRUSt2 results implicated a dramatic difference in the GJM function between HP+ and HP− PCG. In line with the positive findings in the clinical rapid urease test, the urea cycle pathway could be significantly upregulated in HP+ PCG compared to HP− PCG. In addition, our findings also indicated that biosynthesis and maturation of PGN were also enhanced in HP+ PCG compared to HP− PCG. PGN is a major cell wall component of Gram-positive bacteria. It is reported that HP cag + strains deliver components of PGN into epithelial cells via the cag secretion system, leading to decreased apoptosis, increased proliferation, and increased cell migration (Suarez et al., 2017). These changes indicated that PGN could play a potentially important role in modulating host inflammatory responses to HP, allowing the bacteria to persist and induce carcinogenic consequences in the gastric niche (Suarez et al., 2017). Interestingly, we also found that L-lysine biosynthesis increased significantly in HP+ PCG. L-lysine could significantly delay and inhibit gastric emptying (Uchida et al., 2017), which might be related to symptoms such as abdominal distension caused by HP.
Due to sample limitations, further studies will be necessary to confirm our findings in other cohorts with larger sample sizes. Nevertheless, the results of our current study demonstrate dramatic differences in GJM between HP+ and HP− PCG in terms of taxonomic structure, microbiota network, and function. GJM analysis could be a less invasive way to monitor and study the gastric microbiota dynamics in PCG.
The raw sequence data presented in the study are deposited in the Genome Sequence Archive in the National Genomics Data Center, China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences (GSA-human), accession number PRJCA014620.
This study was approved by the Ethics Committee of Jiangnan University, and was conducted in accordance with the Declaration of Helsinki. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
YC, S-YX, F-XR, and J-JF performed the data analysis and wrote the manuscript. YC collected the clinical data. JT and Z-YW conducted the experiments. XL and CQ contributed to the discussion. QL and J-HC conceived the project and planned the experiments. All authors contributed to the final manuscript.
This study was supported in part by grants from the Youth Research Program of Wuxi Health Commission (Q202011), Wuxi maternal and child health research project (FYKY202108), and Wuxi Medical Innovation Team (CXTD2021011).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1112709/full#supplementary-material
Barberan, A., Bates, S. T., Casamayor, E. O., and Fierer, N. (2012). Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 6, 343–351. doi: 10.1038/ismej.2011.119
Benavides-Ward, A., Vasquez-Achaya, F., Silva-Caso, W., Aguilar-Luis, M. A., Mazulis, F., Urteaga, N., et al. (2018). Helicobacter pylori and its relationship with variations of gut microbiota in asymptomatic children between 6 and 12 years. BMC. Res. Notes 11:468. doi: 10.1186/s13104-018-3565-5
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Cardinale, M., Grube, M., Erlacher, A., Quehenberger, J., and Berg, G. (2015). Bacterial networks and co-occurrence relationships in the lettuce root microbiota. Environ. Microbiol. 17, 239–252. doi: 10.1111/1462-2920.12686
Caguazango, J. C. (2020). Ecological models of gastric microbiota dysbiosis: Helicobacter pylori and gastric carcinogenesis. Med. Microecol. 3:100010. doi: 10.1016/j.medmic.2020.100010
Douglas, G. M., Maffei, V. J., Zaneveld, J. R., Yurgel, S. N., Brown, J. R., Taylor, C. M., et al. (2020). PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 38, 685–688. doi: 10.1038/s41587-020-0548-6
Fléjou, J. F., Diomandé, I., Molas, G., Goldfain, D., Rotenberg, A., Florent, M., et al. (1990). Human chronic gastritis associated with non-Helicobacter pylori spiral organisms (Gastrospirillum hominis). Four cases and review of the literature. Gastroenterol. Clin. Biol. 14, 806–810.
Friedman, J., and Alm, E. J. (2012). Inferring correlation networks from genomic survey data. PLoS Comput. Biol. 8:e1002687. doi: 10.1371/journal.pcbi.1002687
Gantuya, B., El Serag, H. B., Saruuljavkhlan, B., Azzaya, D., Matsumoto, T., Uchida, T., et al. (2021). Advantage of 16S rRNA amplicon sequencing in Helicobacter pylori diagnosis. Helicobacter 26:e12790. doi: 10.1111/hel.12790
Glockner, F. O., Yilmaz, P., Quast, C., Gerken, J., Beccati, A., Ciuprina, A., et al. (2017). 25 years of serving the community with ribosomal RNA gene reference databases and tools. J. Biotechnol. 261, 169–176. doi: 10.1016/j.jbiotec.2017.06.1198
He, C., Peng, C., Wang, H., Ouyang, Y., Zhu, Z., Shu, X., et al. (2019). The eradication of Helicobacter pylori restores rather than disturbs the gastrointestinal microbiota in asymptomatic young adults. Helicobacter 24:e12590. doi: 10.1111/hel.12590
Jackson, M. A., Goodrich, J. K., Maxan, M. E., Freedberg, D. E., Abrams, J. A., Poole, A. C., et al. (2016). Proton pump inhibitors alter the composition of the gut microbiota. Gut 65, 749–756. doi: 10.1136/gutjnl-2015-310861
Kishikawa, H., Ojiro, K., Nakamura, K., Katayama, T., Arahata, K., Takarabe, S., et al. (2020). Previous Helicobacter pylori infection-induced atrophic gastritis: a distinct disease entity in an understudied population without a history of eradication. Helicobacter 25:e12669. doi: 10.1111/hel.12669
Kuntal, B. K., Chandrakar, P., Sadhu, S., and Mande, S. S. (2019). 'NetShift': a methodology for understanding 'driver microbes' from healthy and disease microbiome datasets. ISME J. 13, 442–454. doi: 10.1038/s41396-018-0291-x
Ladas, S. D., Katsogridakis, J., Malamou, H., Giannopoulou, H., Kesse-Elia, M., and Raptis, S. A. (1996). Helicobacter pylori may induce bile reflux: link between H. pylori and bile induced injury to gastric epithelium. Gut 38, 15–18. doi: 10.1136/gut.38.1.15
Liu, D., Zhang, R., Chen, S., Sun, B., and Zhang, K. (2022). Analysis of gastric microbiome reveals three distinctive microbial communities associated with the occurrence of gastric cancer. BMC Microbiol. 22:184. doi: 10.1186/s12866-022-02594-y
Martinsen, T. C., Fossmark, R., and Waldum, H. L. (2019). The phylogeny and biological function of gastric juice-microbiological consequences of removing gastric acid. Int. J. Mol. Sci. 20:6031. doi: 10.3390/ijms20236031
Minalyan, A., Gabrielyan, L., Scott, D., Jacobs, J., and Pisegna, J. R. (2017). The gastric and intestinal microbiome: role of proton pump inhibitors. Curr. Gastroenterol. Rep. 19:42. doi: 10.1007/s11894-017-0577-6
Ohkusa, T., Okayasu, I., Miwa, H., Ohtaka, K., Endo, S., and Sato, N. (2003). Helicobacter pylori infection induces duodenitis and superficial duodenal ulcer in Mongolian gerbils. Gut 52, 797–803. doi: 10.1136/gut.52.6.797
Ostrowski, J., Kulecka, M., Zawada, I., Zeber-Lubecka, N., Paziewska, A., Graca-Pakulska, K., et al. (2021). The gastric microbiota in patients with Crohn's disease; a preliminary study. Sci. Rep. 11:17866. doi: 10.1038/s41598-021-97261-z
Rosen, R., Amirault, J., Liu, H., Mitchell, P., Hu, L., Khatwa, U., et al. (2014). Changes in Gastric and Lung Microflora With Acid Suppression. JAMA Pediatr. 168, 932–937. doi: 10.1001/jamapediatrics.2014.696
Sabbagh, P., Javanian, M., Koppolu, V., Vasigala, V. R., and Ebrahimpour, S. (2019). Helicobacter pylori infection in children: an overview of diagnostic methods. Eur. J. Clin. Microbiol. Infect. Dis. 38, 1035–1045. doi: 10.1007/s10096-019-03502-5
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Genome Biol. 12:R60. doi: 10.1186/gb-2011-12-6-r60
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Sipponen, P., and Maaroos, H. I. (2015). Chronic gastritis. Scand. J. Gastroenterol. 50, 657–667. doi: 10.3109/00365521.2015.1019918
Sohn, S. H., Kim, N., Jo, H. J., Kim, J., Park, J. H., Nam, R. H., et al. (2017). Analysis of gastric body microbiota by pyrosequencing: possible role of bacteria other than Helicobacter pylori in the gastric carcinogenesis. J. Cancer Prev. 22, 115–125. doi: 10.15430/JCP.2017.22.2.115
Suarez, G., Romero-Gallo, J., Sierra, J. C., Piazuelo, M. B., Krishna, U. S., Gomez, M. A., et al. (2017). Genetic manipulation of Helicobacter pylori Virulence Function by Host Carcinogenic Phenotypes. Cancer Res. 77, 2401–2412. doi: 10.1158/0008-5472.CAN-16-2922
Sung, J., Kim, N., Kim, J., Jo, H. J., Park, J. H., Nam, R. H., et al. (2016). Comparison of gastric microbiota between gastric juice and mucosa by next generation sequencing method. J. Cancer Prev. 21, 60–65. doi: 10.15430/JCP.2016.21.1.60
Takagi, T., Naito, Y., Inoue, R., Kashiwagi, S., Uchiyama, K., Mizushima, K., et al. (2018). The influence of long-term use of proton pump inhibitors on the gut microbiota: an age-sex-matched case-control study. J. Clin. Biochem. Nutr. 62, 100–105. doi: 10.3164/jcbn.17-78
Tao, Z. H., Han, J. X., and Fang, J. Y. (2020). Helicobacter pylori infection and eradication: exploring their impacts on the gastrointestinal microbiota. Helicobacter 25:e12754. doi: 10.1111/hel.12754
Uchida, M., Kobayashi, O., and Saito, C. (2017). Correlation Between Gastric Emptying and Gastric Adaptive Relaxation Influenced by Amino Acids. J. Neurogastroenterol Motil. 23, 400–408. doi: 10.5056/jnm16153
Wei, Z. Y., Rao, J. H., Tang, M. T., Zhao, G. A., Li, Q. C., Wu, L. M., et al. (2022). Characterization of changes and driver microbes in gut microbiota during healthy aging using a captive monkey model. Genomics Proteomics Bioinformatics 20, 350–365. doi: 10.1016/j.gpb.2021.09.009
Keywords: gastric microbiota, Helicobacter pylori, pediatric chronic gastritis, network of microbial interaction, Streptococcus
Citation: Chen Y, Xia S-Y, Ru F-X, Feng J-J, Tao J, Wei Z-Y, Li X, Qian C, Lin Q and Chen J-H (2023) Gastric juice microbiota in pediatric chronic gastritis that clinically tested positive and negative for Helicobacter pylori. Front. Microbiol. 14:1112709. doi: 10.3389/fmicb.2023.1112709
Edited by:
Chun Cui, South China University of Technology, ChinaReviewed by:
Chengmin Wang, Guangdong Academy of Sciences, ChinaCopyright © 2023 Chen, Xia, Ru, Feng, Tao, Wei, Li, Qian, Lin and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qiong Lin, bGlucWlvbmc3NkAxNjMuY29t; Jian-Huan Chen, Y2poX2Jpb0Bob3RtYWlsLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.