
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 09 February 2023
Sec. Terrestrial Microbiology
Volume 14 - 2023 | https://doi.org/10.3389/fmicb.2023.1101773
This article is part of the Research Topic Soil Microbiome Community and Functional Succession Mechanism Driven by Different Factors in Agricultural Ecology View all 43 articles
 Jingjing Wang1,2*
Jingjing Wang1,2* Siqi Zhao1,2
Siqi Zhao1,2 Song Xu1,2
Song Xu1,2 Wei Zhao1,2
Wei Zhao1,2 Xiaoxia Zhang1,2
Xiaoxia Zhang1,2 Yu Lei1,2,3Huanhuan Zhai1,2,3
Yu Lei1,2,3Huanhuan Zhai1,2,3 Zhiyong Huang1,2*
Zhiyong Huang1,2*Microbial inoculation with plant growth-promoting microorganisms (PGPMs) is one of the most promising technologies to solve the current global challenges. Co-inoculants is more efficient and stable than mono-inoculants. However, the growth promoting mechanism of co-inoculants in complex soil system is still poorly understood. In this study, the effects on rice, soil and the microbiome of the mono-inoculant Bacillus velezensis FH-1 (F) and Brevundimonas diminuta NYM3 (N) and the co-inoculant FN obtained in previous works were compared. Correlation analysis and PLS-PM were used to explore the primary mechanism of different inoculants promoting rice growth. We hypothesized that inoculants promoted plant growth (i) by themselves, (ii) by improving soil nutrient availability or (iii) by regulating the rhizosphere microbiome in complex soil system. We also assumed that different inoculants had different ways of promoting plant growth. The results showed that FN significantly promoted rice growth and nitrogen absorption and slightly increased soil total nitrogen and microbial network complexity compared with F, N and the control (CK). B. velezensis FH-1 and B. diminuta NYM3 interfered with each other’s colonization in FN. FN increased the complexity of the microbial network compared to F and N. The bacterial community of FN was quite different from CK and N, while the fungal community was not significantly different from other treatments. The species and functions enriched or inhibited by FN are part of F. The correlation analysis and PLS-PM results showed that inoculants (F/N/FN) promoted the growth of rice mainly by regulating the rhizosphere microbiome rather than by themselves or by improving soil nutrient availability. Co-inoculant FN promotes rice growth specifically by enhancing microbial nitrification function through enriching related species compared with F or N. This may provide theoretical guidance for the construction and application of co-inoculants in the future.
Microbial inoculation with plant growth-promoting microorganisms (PGPMs) is one of the most promising technologies to solve the current global challenges of increasing food demand, human population, environmental pollution, land degradation and extreme climate (Backer et al., 2018; Singh et al., 2019, 2020). Currently, the global market for microbial inoculants is proliferating (Waltz, 2017; Basu et al., 2021). Despite excellent prospects, poor stability of mono-inoculation hinder the large-scale implementation of microbial inoculants in mainstream agriculture (Awasthi, 2019).
Co-inoculation with two or more PGPMs is more efficient and stable than a traditional microbial mono-inoculant due to several benefits provided by biodiversity (Hu et al., 2017; Kaminsky et al., 2019; Santoyo et al., 2021; Khan, 2022). Co-inoculation of rhizobia and plant growth-promoting bacteria (PGPB) are common. Compared with inoculation of rhizobia alone, co-inoculation of rhizobia and PGPB would enhance nodulation and N2-fixing efficiency by producing larger nodules, thus promoting soybean crop growth (Fox et al., 2011; Zeffa et al., 2020). Co-inoculation of PGPBs or co-inoculation of PGPB and arbuscular mycorrhizal fungi (AMF) has also been widely reported to promote plant growth compared with single inoculation (Hu et al., 2017; Emmanuel and Babalola, 2020; Moreira et al., 2020). For example, co-inoculation of Bacillus and Pseudomonas promoted the development of cucumber, tomato, wheat, and other plants (Ansari and Ahmad, 2019; He et al., 2019; Sun et al., 2021). Co-inoculation with Bacillus and AMF promoted the growth of wheat, tobacco, turmeric (Curcuma longa), and other plants (Begum et al., 2022; Rehman et al., 2022; Sarathambal et al., 2022).
Similar to PGPM, co-inoculants also promote plant growth by improving soil nutrient availability, secreting hormones, changing the soil microbial community, and antagonizing (Olenska et al., 2020; Kong and Liu, 2022; Luo et al., 2022). Some studies have shown that co-inoculants can promote the absorption of nutrients by crops, improve soil enzyme activity, and/or change the structure of the microbial community while promoting the growth of crops (Hu et al., 2017; Moreira et al., 2020; Neemisha et al., 2022; Sarathambal et al., 2022). However, most of these are in vitro studies, and the primary mechanism by which inoculants promotes crop growth in the complex soil systems is still unclear.
Some studies have deeply analyzed the cooperation among species in co-inoculants and found that they mainly interact beneficially through metabolites. For example, Bacillus may promote the nitrogen fixation of Bradyrhizobium mainly by secreting hormones and other substances (Sibponkrung et al., 2020). Bacillus stimulated resident rhizosphere Pseudomonas for plant health through metabolic interactions (Sun et al., 2021). Fructose exuded by the AMF (Rhizophagus irregularis) stimulated the phosphatase activity of phosphate solubilizing bacteria (PSB) (Rahnella aquatilis), simultaneously stimulating the processes involved in phosphorus uptake by the AMF (Zhang et al., 2018). However, the interactions among species in co-inoculants in soil are still poorly understood.
In this study, the effects of the mono-inoculant Bacillus velezensis FH-1 (F) and Brevundimonas diminuta NYM3 (N), and co-inoculant FN obtained in previous works were compared. Correlation analysis and PLS-PM were used to explore the primary mechanism of different inoculants promoting rice growth. We hypothesized that inoculants promoted plant growth (i) by themselves, (ii) by improving soil nutrient availability or (iii) by regulating the rhizosphere microbiome in complex soil system. If inoculants promoted plant growth by themselves, plant should be closely related to the number of the inoculants. If inoculants promoted plant growth by improving soil nutrient availability, plant should be closely related to the soil available nutrient (such as N, P, K, Fe). If inoculants promoted plant growth by regulating the rhizosphere microbiome, plant should be closely related to the microbial diversity or some species. We also assumed that different inoculants had different ways of promoting plant growth. This study may provide theoretical guidance for the construction and application of co-inoculants in the future.
The microbial co-inoculants FN are composed of Bacillus sp. FH-1 and Brevundimonas sp. NYM3, which were obtained in previous works (Zhao et al., 2020). 16S rDNA sequence analysis using primers 27F/1492R was performed to further identify the Bacillus sp. FH-1 and Brevundimonas sp. NYM3. The GenBank accession numbers for the full-length 16S rRNA genes of Bacillus FH-1 and Brevundimonas NYM-3 were OM780304 and OM780305, respectively. The sequences were aligned with BLAST, and phylogenetic trees were constructed using the neighbor-joining method provided in MEGA version 5.0 with a bootstrap value of 1,000 replicates.
For scanning electron microscopy (SEM), Bacillus FH-1 and Brevundimonas NYM-3 at the exponential phase were harvested and washed three times with phosphate-buffered saline (PBS) buffer (pH = 7.2). The samples were fixed for 2 h in 2.5% glutaraldehyde and postfixed for 1 h with 1% osmium tetroxide. The samples were dehydrated with ethanol and dried in an Automated Critical Point Dryer (Leica EM CPD300). Then, the samples were coated with platinum and observed under a scanning microscope (Hitachi SU8010).
The interactions between Bacillus sp. FH-1 and Brevundimonas sp. NYM3 was tested using modified dual culture plate assay (Oszust and Frąc, 2020; Anith et al., 2021). Bacillus sp. FH-1 and Brevundimonas sp. NYM3 were cultured in LB liquid medium at 37°C for 24 h. Bacterial cells were harvested by centrifugation at 10,000 rpm g for 1 min and resuspended in sterile water to an optical density of 1.00 at 600 nm. To study the antagonism between the two bacteria, 5 μl of Bacillus sp. FH-1 and 5 μl of Brevundimonas sp. NYM3 were placed on a LB plate at a distance of about 0.5 cm from each other. Because the expansion of bacterial colony is slow, the distance between the two bacteria is close. As controls, another 5 μl of Bacillus sp. FH-1 and 5 μl of Brevundimonas sp. NYM3 were also placed on the LB plate at a distance of about 1.5 cm from others. The experiment was set up in triplicates (n = 3). The plates were incubated at 37°C for 5–7 days. If the diameter of a bacterial colony is inhibited, it means that it is antagonized by another bacteria.
A modified agar well diffusion method was also used to evaluate the interactions between Bacillus sp. FH-1 and Brevundimonas sp. NYM3 (Lin and Pan, 2019; Ji et al., 2021). Bacillus sp. FH-1 and Brevundimonas sp. NYM3 were cultured in LB liquid medium at 37°C for 24 h. 100 μl of Bacillus sp. FH-1 or Brevundimonas sp. NYM3 was spread evenly on LB agar plates. Then, 5 μl of Brevundimonas sp. NYM3 or Bacillus sp. FH-1 was inoculated on LB agar plates. All the plates were cultured at 37°C for 2–3 days. If there is an inhibition zone around the inoculated bacteria, it indicates that the inoculated bacteria antagonize the spreader and vice versa.
Bacillus FH-1 and Brevundimonas NYM-3 were grown at 30°C for 72 h in LB medium on a rotary shaker (180 rpm). The bacterial number was count with a microscope. The bacterial broth was diluted to 1 × 108 CFU/ml with tap water.
Soil (pH 7.69, organic matter 17.80 g/kg, total N 3.00 g/kg, available N 37.33 mg/kg, total P 0.39 g/kg, available P 9.57 mg/kg, total K 8.87 g/kg and available K 61.84 mg/kg) was collected from the upper 30 cm of a weed field in the Airport economic area, Tianjin, China. The sampled soil was air dried and mixed thoroughly, followed by a sieving step (0.5-cm mesh) to remove plant debris. Thirteen rice seeds (Nei 5 You 8,015 Hybrid rice, Zhejiang Agricultural Science and Technology Seed Industry Co., Ltd., Zhejiang, China) were sown in each plastic pot (diameter 8 cm, height 10 cm) containing 240 g of soil. After 5 days of sowing, 11 rice seedlings with the same growth were kept. Then, pot soils were drenched with 30 ml of the prepared inoculums or equivalent water. There were four treatments: (i) soil drenched with Bacillus FH-1 (F), (ii) soil drenched with Brevundimonas NYM-3 (N), (iii) soil drenched with equal proportions of Bacillus FH-1 and Brevundimonas NYM-3 (FN), and (iv) soil drenched with water (CK). Nine replications of each treatment were set up during the whole experimental period. Pots were placed randomly in a growth chamber (CIMO, Shanghai, China) with 75% relative humidity and 16-h light. Before seedling emergence, the temperature was controlled at 30°C. Then, the temperature was set at 28°C day/24°C night for one leaf stage, 28°C day/25°C night for two-leaf stage, and 28°C day/22°C night for other stages. The pots were watered 30 ml every 48 h, and the position of the rice pots was randomly changed.
At 16 days after sowing, six replications of each treatment were randomly chosen (a total of 24 samples) for further analysis. Plants of each pot were harvested and carefully separated into roots and shoots to determine the growth parameters, including length, fresh weight, and dry weight, using a ruler and an electronic balance (Mettler Toledo, Shanghai, China), respectively. Meanwhile, rhizosphere soil samples of each treatment were collected and stored at 4°C and −80°C.
The soil pH, total organic carbon, total nitrogen, total phosphorus, total potassium, available nitrogen, available phosphorus, and available potassium were determined by Suzhou Comin Biotechnology Co., Ltd., Suzhou, China.
Soil metagenomic DNA was isolated from 24 soil samples by the PowerSoil DNA isolation kit (MO BIO Laboratories, Inc., Carlsbad, CA, United States) according to the manufacturer’s instructions. DNA purity and concentration were monitored by 1% agarose gels and NanoDrop ND-2000 spectrophotometry (NanoDrop Technologies, Wilmington, DE, United States), respectively.
Quantification of the copy number of bacteria and fungi was performed using a real-time PCR assay. Real-time PCR experiments were conducted in a 7500 Fast Real-Time PCR System (Applied Biosystem, Foster City, CA, United States). Bacterial-specific primers (338F 5’-ACTCCTACGGGAGGCAGCAG-3′ and 518R 5’-ATTACCGCGGCTGCTGG-3′) and fungal-specific primers (ITS1 5′-TCCGTAGGTGAACCTGCGG-3′ and 5.8S 5′-CGCTGCGTTCTTCATCG-3′) were used. Each PCR was performed in a total reaction volume of 20 μl, which consisted of using 10 μl SYBR Select Master Mix (Applied Biosystem, Foster City, CA, United States), 1 μl each primer, 1 μl template DNA and 7 μl ddH2O. The final two-step cycling program included a 10-min initial preincubation at 95°C followed by 40 cycles of 95°C for 15 s and 60°C for 1 min.
Standards for real-time PCR assays were prepared as described elsewhere (Wang et al., 2017). Briefly, the specific 16S rRNA gene of Brevundimonas sp. NYM-3 and the specific ITS gene of Trichoderma longibrachiatum MF-1 were PCR-amplified from extracted DNA with the primers. The PCR products were cloned into a T vector (GoldTopo, Tianjin, China). Plasmids used as standards for quantitative analyses were extracted from the correct insert clones of each target gene using a Mini Plasmid Kit (TIANGEN, Beijing, China). The concentration of plasmid DNA was determined on a NanoDrop (NanoDrop-1,000, Thermo Scientific, United States), and the copy numbers of the target genes were calculated directly from the concentration of the extracted plasmid DNA. Tenfold serial dilutions of each known copy number of the plasmid DNA were subjected to a real-time PCR assay in triplicate to generate an external standard curve.
The bacterial hypervariable regions (V4-V5) of the 16S rRNA genes and the fungal hypervariable regions (ITS2) of the ITS genes were amplified using primers 515F (5′- GTGYCAGCMGCCGCGGTAA - 3′) - 926R (5′- CCGYCAATTYMTTTRAGTTT - 3′) and fITS7F (5′ - GTGAATCATCGAATCTTTGAA - 3′) - ITS4R (5′ - TCCTCCGCTTATTGATATGC - 3′), respectively. PCR products were purified and then sequenced using the MiSeq platform at Novogene Co., Ltd. (Tianjin, China). The raw sequence data have been deposited into the NCBI Sequence Read Archive under accession PRJNA804354. Raw data were processed and analyzed (NMDS, Adonis, LEfSe analysis, function prediction, and so on) using BMKCloud1.
All statistical analyses were performed using R (version 3.1.1). The effects of microbial inoculants on rice, soil, microbial quality, abundance, and α-diversity were evaluated by Tukey’s HSD test. We used all genera to construct the network with the “Hmisc” package in R and Gephi (Wang et al., 2021). The package “pheatmap” and Spearman correlation analysis were used to evaluate the relationships between microorganisms, rice, and soil. Partial least squares path models (PLS-PMs) were used to assess the effects of microbial inoculants, microbial diversity, and soil on rice.
The phylogenetic trees showed that FH-1 is B. velezensis and NYM-3 is B. diminuta (Supplementary Figure S1). B. velezensis was the conspecific species integrating B. amyloliquefaciens subsp. plantarum and B. methylotrophicus (Rabbee et al., 2019). The scanning electron microscope (SEM) images demonstrated that the size of FH-1 is approximately 2,300 nm × 700 nm and NYM-3 is approximately 1,400 nm × 450 nm (Supplementary Figure S1). Dual culture plate assay and agar well diffusion method both revealed that FH-1 inhibited the growth of NYM-3 cells (Supplementary Figure S2).
Rice pot experiments showed that the fresh weight, dry weight, and height of rice seedlings were significantly increased by the three microbial inoculations (F/N/FN) compared with CK (Figures 1A–E). The fresh weight, dry weight, and height of rice seedlings (both shoot and root) in FN were significantly higher than those in F and N. This indicated that FN was more effective than F or N in promoting rice growth.
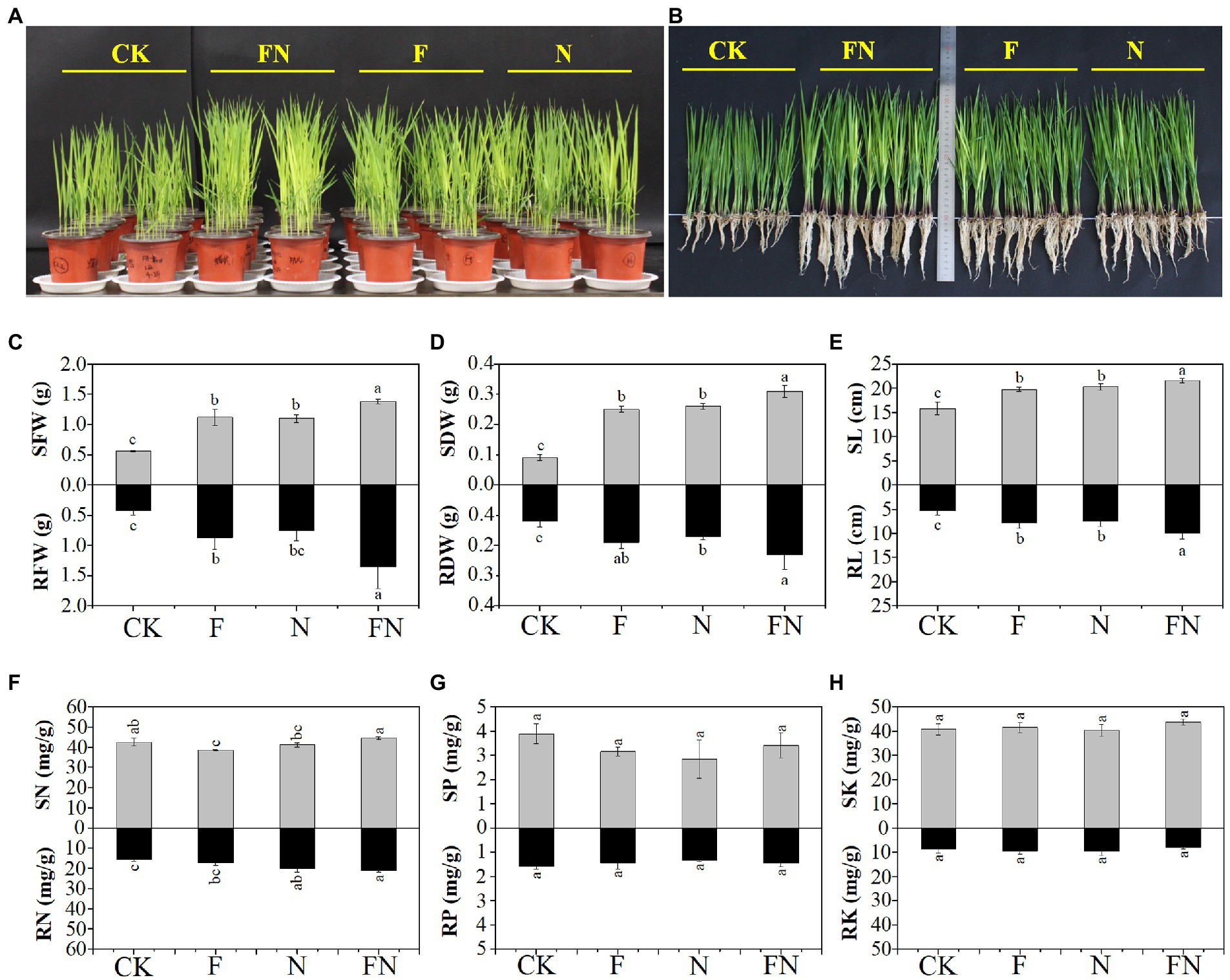
Figure 1. The effects of different microbial inoculants on rice. (A) The rice pot experiments; (B) photos showing 16-day-old rice plants; (C) rice shoot (S) and root (R) fresh weight (FW); (D) rice shoot (S) and root (R) dry weight (DW); (E) rice shoot (S) and root (R) length (L); (F) Rice shoot (S) and root (R) nitrogen concentration (N); (G) rice shoot (S) and root (R) phosphorus concentration (P); (H) rice shoot (S) and root (R) potassium concentration (K); CK, non-inoculated; F, inoculated with Bacillus velezensis FH-1; N, inoculated with Brevundimonas diminuta NYM-3; FN, inoculated with B. velezensis FH-1 and B. diminuta NYM-3. Data followed by the different lowercase letters are significantly different at p ≤ 0.05.
The nitrogen concentration of rice shoots in FN was significantly higher than that in F and N. The nitrogen concentration of rice roots in FN was significantly higher than that in CK and F. There was no significant difference in phosphorus and potassium concentrations in rice seedlings among the different treatments (Figures 1F–H).
Soil pH, total nitrogen, and available potassium differed significantly among the different treatments (Table 1). The soil pH was significantly higher in all three microbial inoculations (F/N/FN) than in CK. Soil total nitrogen was significantly higher in FN than in N. Soil available potassium was significantly higher in N than in F and FN.
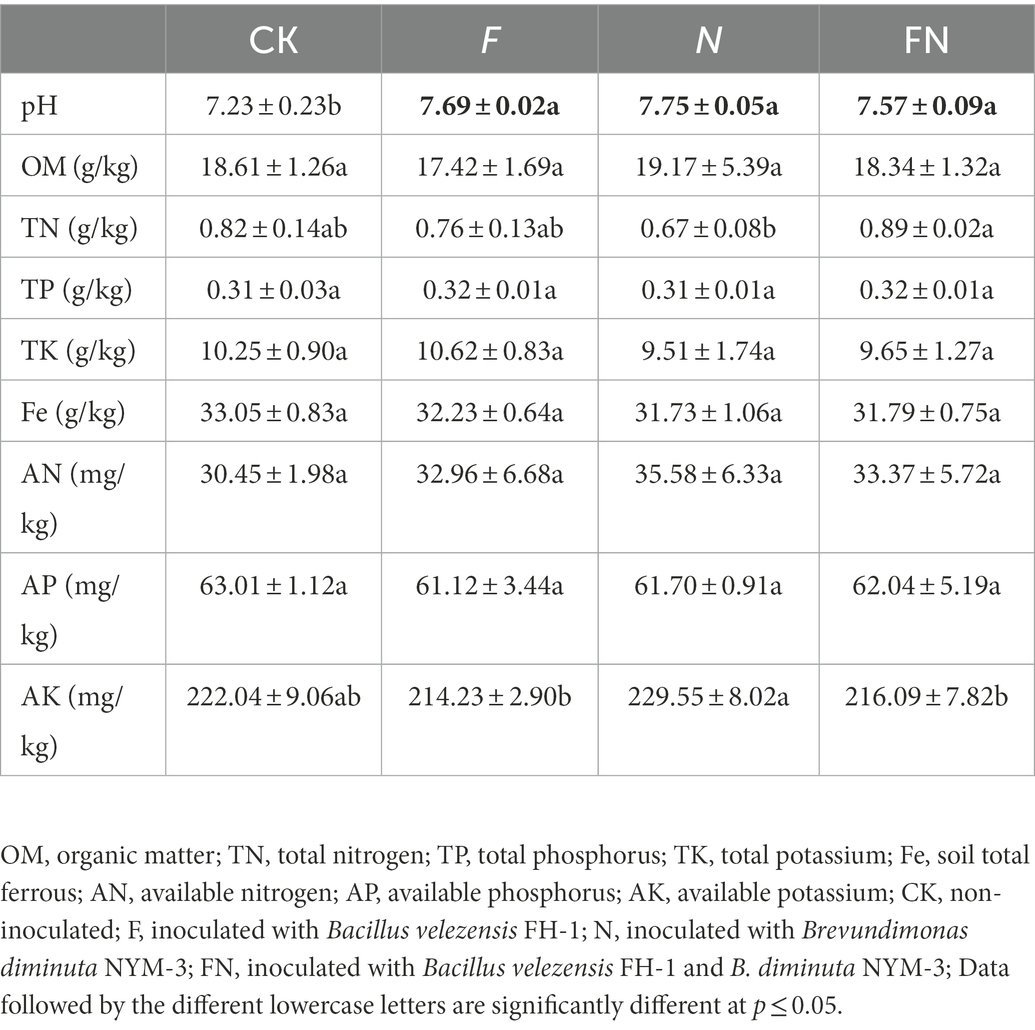
Table 1. The effects of different microbial inoculants on soil properties.
The quantitative real-time PCR results showed that the number of rhizosphere bacteria and fungi in F and N was significantly higher than that in CK (Figure 2A). The number of bacteria and fungi in FN was slightly higher than that in CK but slightly lower than that in F and N. The number of fungi was significantly higher than that of bacteria in CK and FN.
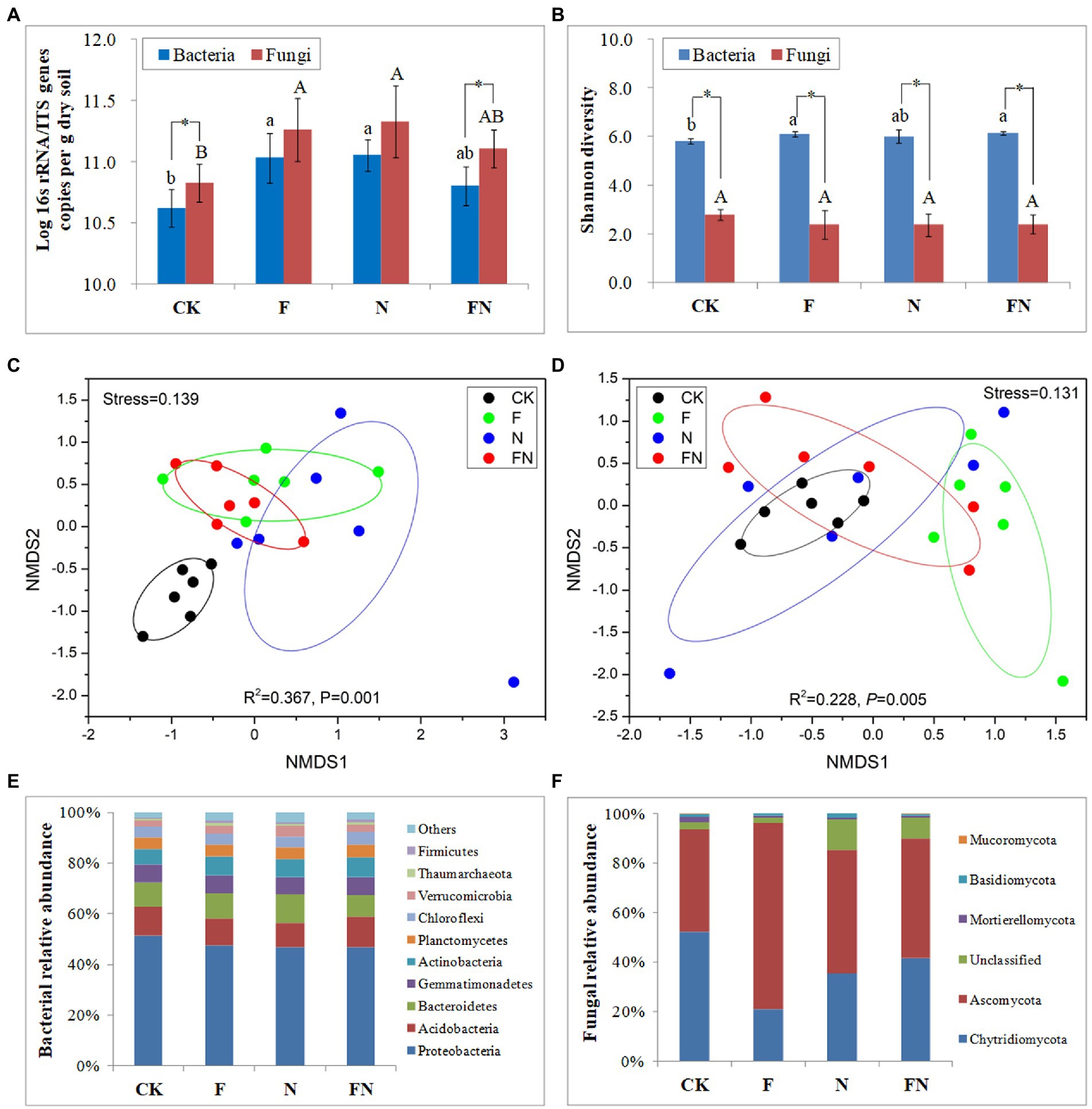
Figure 2. The effects of different microbial inoculants on rhizosphere microbiome. (A) Microbial quantity; (B) microbial α diversity; (C) bacterial β diversity; (D) fungal β diversity; (E) bacterial composition; (F) fungal composition. Beta diversity was revealed by NMDS using bray curtis. CK, non-inoculated; F, inoculated with B. velezensis FH-1; N, inoculated with B. diminuta NYM-3; FN, inoculated with B. velezensis FH-1 and B. diminuta NYM-3. Data followed by the different letters are significantly different at p ≤ 0.05. Significance levels are indicated by *(p < 0.05), **(p < 0.01) and ***(p < 0.001).
Microbial α-diversity was characterized by the Shannon index (Figure 2B). The bacterial α diversity was significantly higher than the fungal α diversity in all treatments. The bacterial α diversity in FN and F was significantly higher than that in CK. There was no significant difference in fungal α-diversity between the different treatments.
The stress in the NMDS was less than 0.2, which indicated that the result had certain reliability. The NMDS and PERMANOVA (Adonis) results showed significant differences in both bacterial (R2 = 0.367, p = 0.001) and fungal (R2 = 0.228, p = 0.005) β-diversity among the different treatments (Figures 2C,D). The bacterial communities of the inoculations (F/N/FN) were separated from CK (p < 0.05) (Figure 2C; Table 2). The fungal community of F was separated from CK (p < 0.05) (Figure 2D; Table 2). All bacterial and fungal communities of FN overlapped with those of F and N.
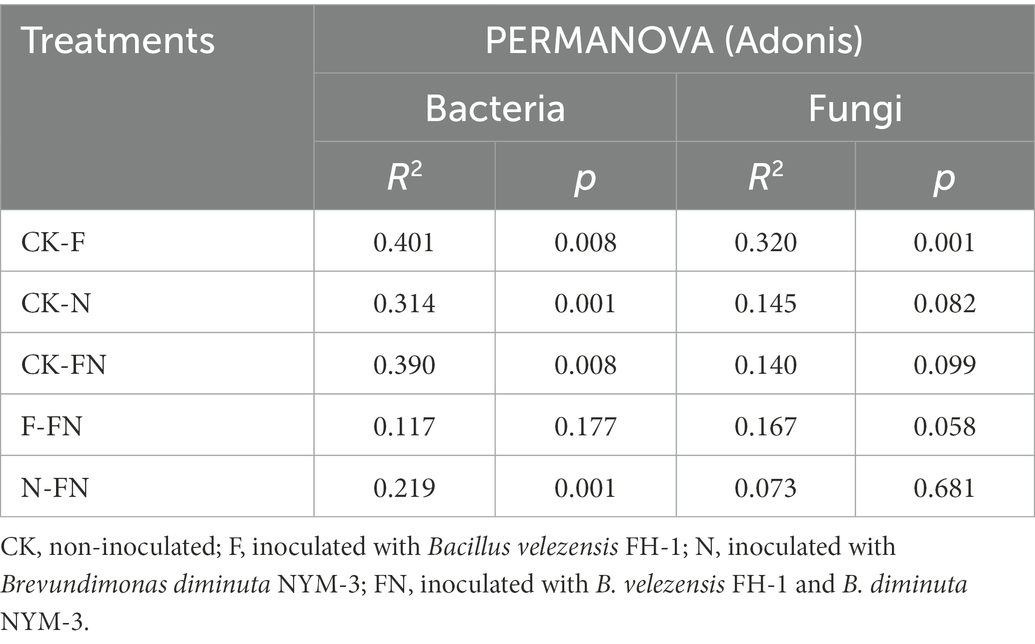
Table 2. Differences in microbial β diversity among different treatments.
Rice rhizosphere bacteria mainly consisted of Proteobacteria, Acidobacteria, Bacteroidetes, Gemmatimonadetes, Actinobacteria, and Planctomycetes. The relative abundance of Verrucomicrobia was significantly higher in the N treatment than in the other treatments. There were more Thaumarchaeota and Firmicutes in F and FN than in CK (Figure 2E; Supplementary Table S1).
Rice rhizosphere fungi are mainly composed of Ascomycota and Chytridiomycota. The relative abundance of Ascomycota was significantly higher in the F treatment than in the other treatments. The relative abundance of Chytridiomycota was significantly higher in CK and FN than in F. The relative abundance of Mortierellomycota was significantly higher in inoculations (F/N/FN) than in CK (Figure 2F; Supplementary Table S1).
Local BLAST (sequence similarity >99%) was used to estimate the colonization of B. velezensis FH-1 and B. diminuta NYM-3 in each treatment. This method only uses part of the 16S rRNA sequence to identify species is not accurate, and will be affected by indigenous bacteria. However, the colonization of inoculants can be inferred from the comparison between inoculated and uninoculated treatments. The results showed that the relative abundance and number (bacterial number × the relative abundance) of B. velezensis FH-1 did not differ significantly among the different treatments (Figure 3). However, the relative abundance and number of B. velezensis FH-1 were higher in F and FN than in CK and N, indicating that FH-1 may have weakly colonized F and FN. The relative abundance and number of B. velezensis FH-1 in FN were lower than those in F, suggesting that B. diminuta NYM-3 may have hindered the colonization of FH-1.
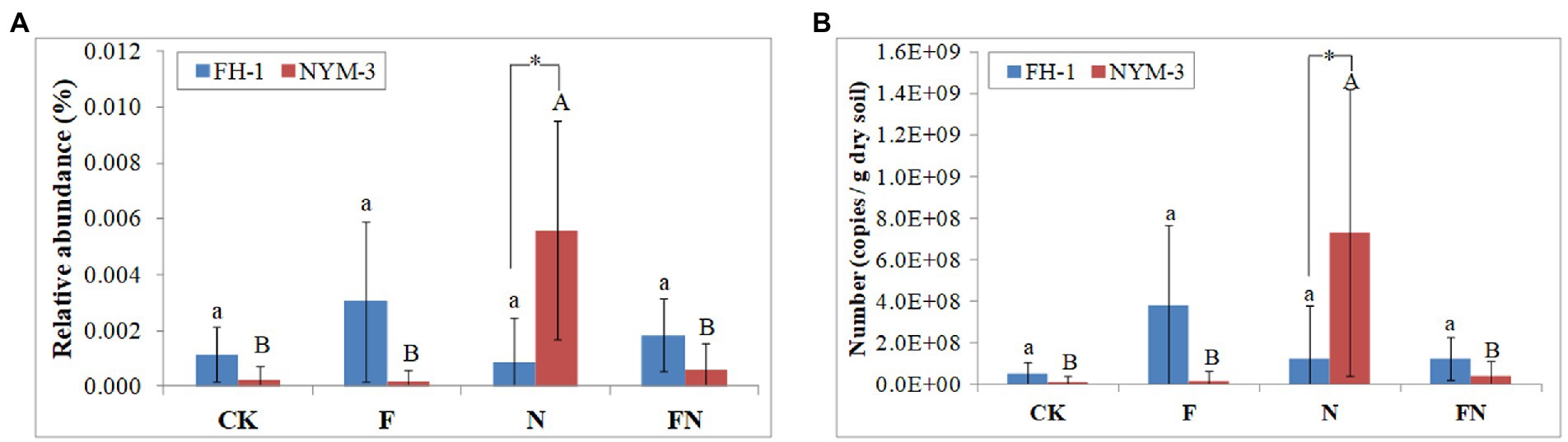
Figure 3. The relative abundance (A) and number (B) of B. velezensis and B. diminuta NYM-3 in rice rhizosphere soil. CK, non-inoculated; F, inoculated with B. velezensis FH-1; N, inoculated with B. diminuta NYM-3; FN, inoculated with B. velezensis FH-1 and B. diminuta NYM-3. Data followed by the different letters are significantly different at p ≤ 0.05 among different treatments.
The relative abundance and number of B. diminuta NYM-3 were significantly higher in N than in other treatments, suggesting that NYM-3 efficiently colonized N. The relative abundance and number of B. diminuta NYM-3 were slightly higher in FN than in CK and F, suggesting that B. diminuta NYM-3 weakly colonized FN (Figure 3). The relative abundance and number of B. diminuta NYM-3 in FN were significantly lower than those in N, suggesting that B. velezensis FH-1 also hindered the colonization of B. diminuta NYM-3. The presence of B. velezensis FH-1 and B. diminuta NYM-3 in all treatments indicated that they might be indigenous bacteria.
To further characterize the effect of the microbial inoculants on the rhizosphere microbiome, we assessed the cooccurrence network patterns of microbial communities compared to CK at the genus level based on a strong (Spearman’s r > 0.6) and significant (p < 0.05) correlation. The results showed that FN had higher edges, the ratio of negative correlations, average degree, and graph density and lower positive correlations, average path length, and modularity than F or N (Figure 4; Table 3). Higher edges, ratio of positive correlations, average degree, average path length, network diameter, graph density, and modularity and a lower ratio of negative correlations and average clustering coefficient in N than in F. A higher average degree represents a greater network complexity. This indicated that FN had the highest network complexity, while F had the lowest network complexity. There were 19 genera in F and FN, while only 6 genera in N interacted with Bacillus. Eleven genera in F and 1 genus in N are the same as those contained in FN. There were 3 genera directly interacted with Brevundimonas in N and FN, and only one genus was the same (Supplementary Figure S3; Supplementary Table S2). This indicated that the interaction of specific taxa was affected by inoculants.
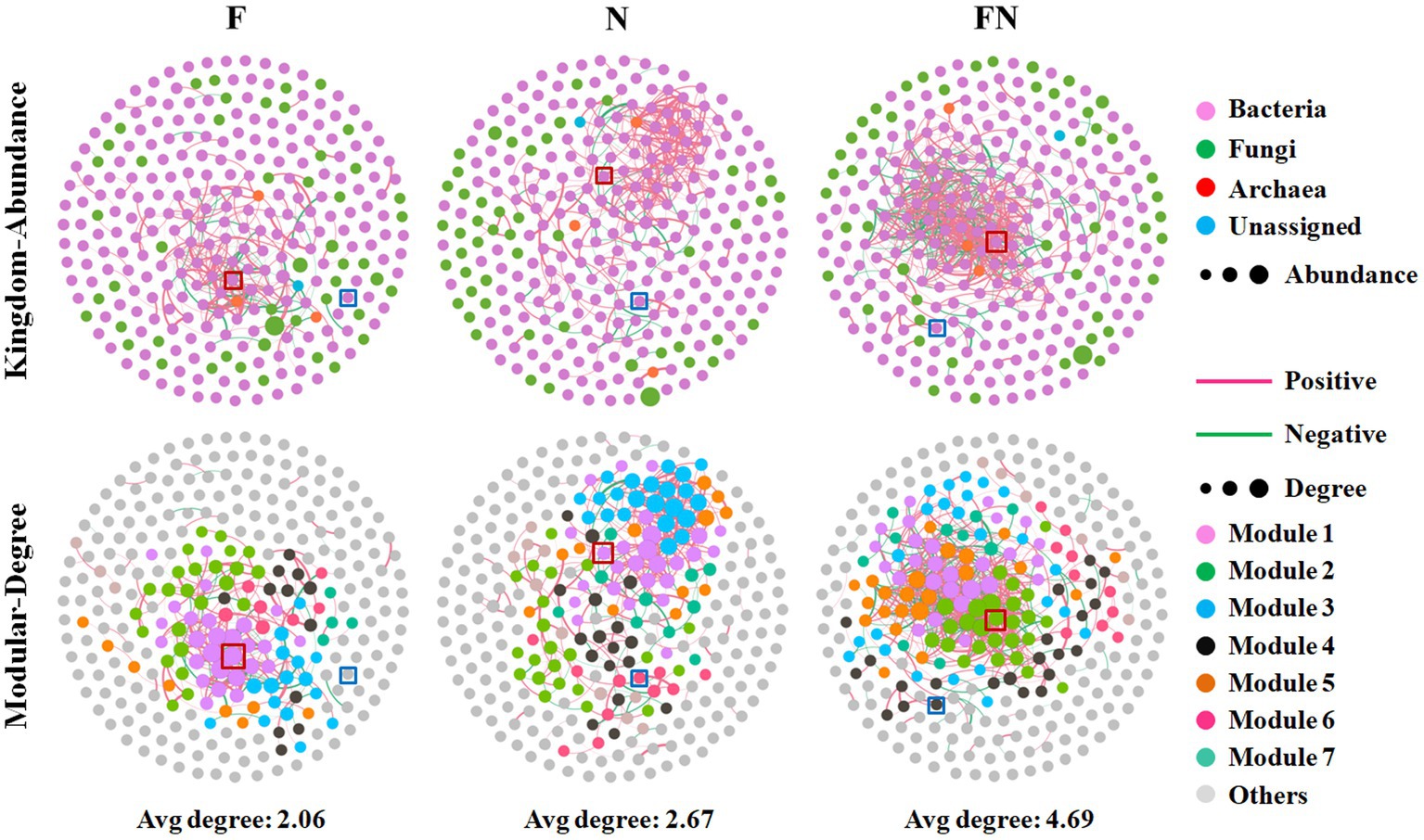
Figure 4. Microbial networks among different microbial inoculants. CK, non-inoculated; F, inoculated with B. velezensis FH-1; N, inoculated with B. diminuta NYM-3; FN, inoculated with B. velezensis FH-1 and B. diminuta NYM-3. Bacillus (n37) is labeled by red square, Brevundimonas (n39) is labeled by blue square.
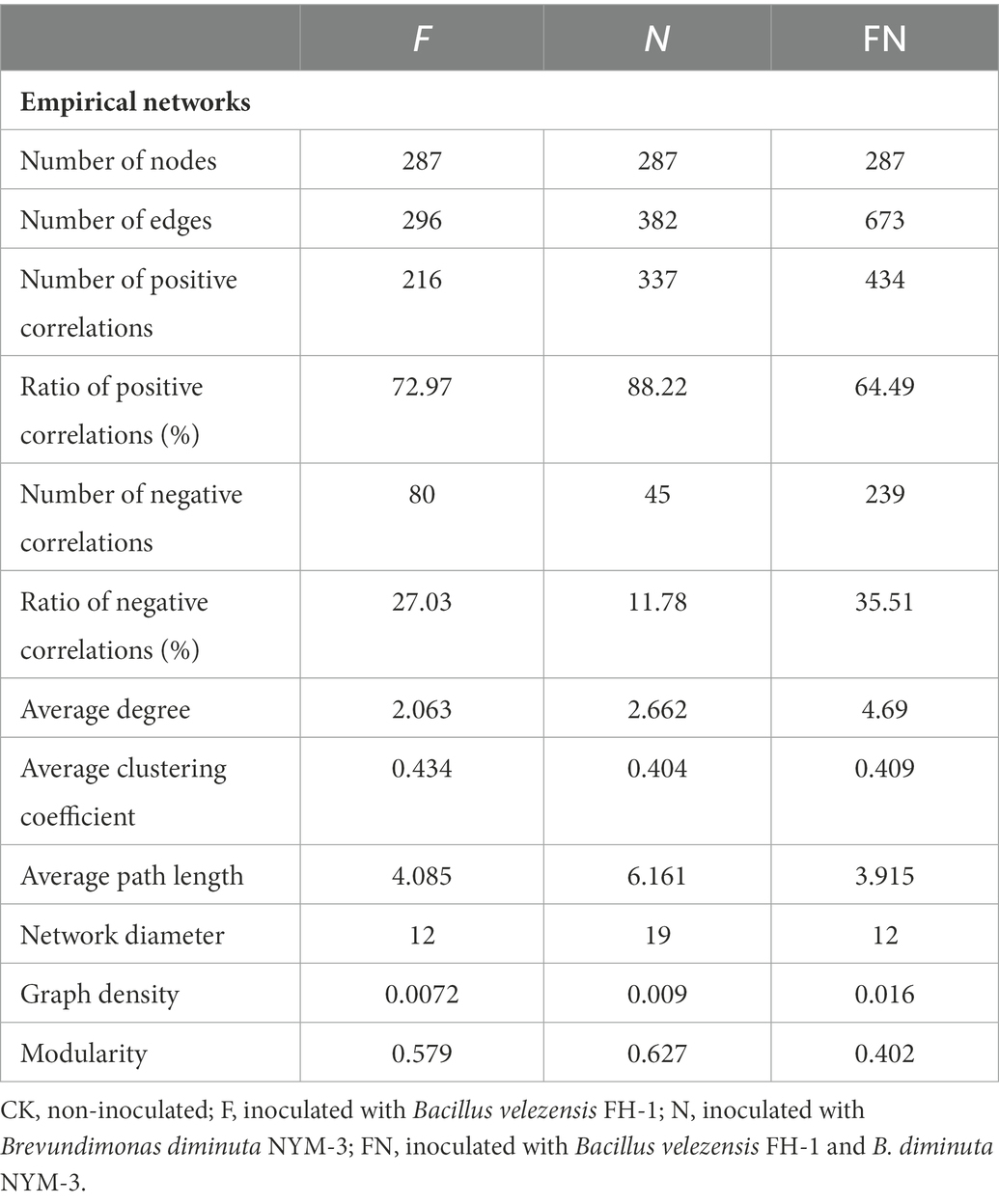
Table 3. Topological properties of rhizosphere bacterial networks obtained from different microbial inoculants treatments.
LEfSe analysis of bacteria showed that all inoculation treatments (F, N, and FN) significantly enriched uncultured_bacterium_g_Pseudomonas compared to the CK (Figure 5A; Supplementary Figure S4A). Pseudomonadales, Pseudomonadaceae, and Pseudomonas were enriched by N compared to the CK. All inoculation treatments (F, N, and FN) significantly inhibited Alphaproteobacteria, Sphingomonadales, Sphingomonadaceae, Sphingomonas, Sphingomonas flava, uncultured_bacterium_g_Sphingomonas, Xanthomonadales, Xanthomonadaceae, Lysobacter, and uncultured_bacterium_g_Lysobacter compared to CK. Both F and FN inhibited Proteobacteria compared to CK. There was no significant difference among the three inoculums (F/N/FN) in the relative abundance of common enriched or inhibited species (Supplementary Figure S5).
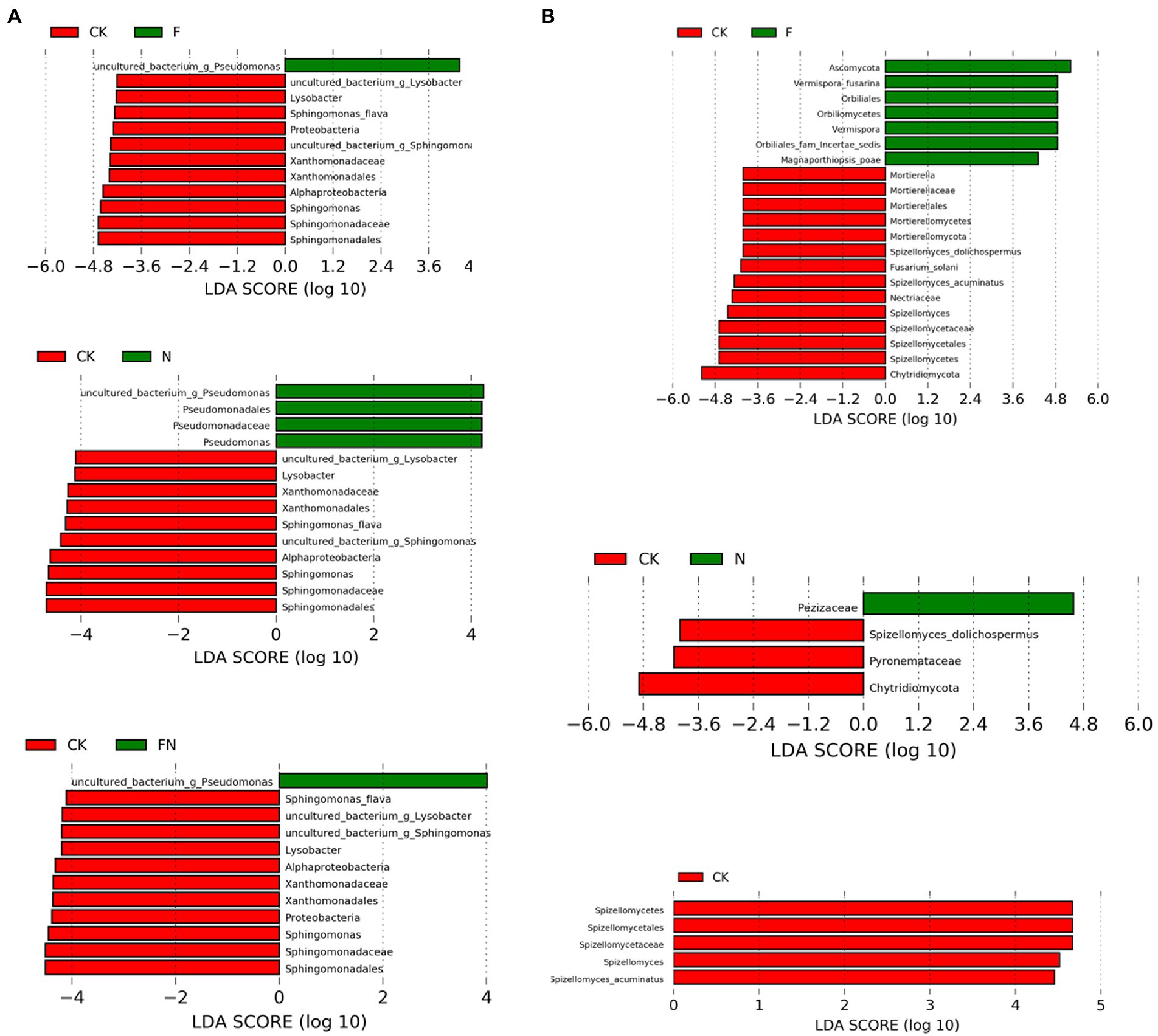
Figure 5. Cladograms of LEfSe analysis of different microbial inoculants on bacterial (A) and fungal (B) community. CK, non-inoculated; F, inoculated with B. velezensis FH-1; N, inoculated with Brevundimonas sp. NYM-3; FN, inoculated with B. velezensis FH-1 and Brevundimonas sp. NYM-3.
LEfSe analysis of fungi showed that F enriched Ascomycota, Orbiliomycetes, Orbiliales, Orbiliales_fam_Incertae_sedis, Vermispora, Vermispora fusarina, and Magnaporthiopsis poae compared to CK (Figure 5B; Supplementary Figure S4B). Pezizaceae was enriched by N compared to CK. F and FN significantly inhibited Spizellomycetes, Spizellomycetales, Spizellomycetaceae, Spizellomyces, and Spizellomyces acuminatus. Chytridiomycota and Spizellomyces dolichospermus were inhibited by F and N compared to CK. F also inhibited Nectriaceae, Fusarium solani, Mortierellomycota, Mortierellomycetes, Mortierellales, Mortierellaceae, and Mortierella compared to CK. Pyronemataceae was inhibited by N compared to CK.
Bacterial function prediction (FAPROTAX) was analyzed with OTU abundance >0.1% (Supplementary Figure S6A). The reports showed that 22.09% (345 out of 1,562) of records were assigned to at least one group. Difference analysis results showed that F had significantly higher manganese_oxidation, aromatic_compound_degradation, and predatory_or_exoparasitic and lower chemoheterotrophy and chitinolysis than CK (Figure 6A). N had significantly lower chemoheterotrophy and chitinolysis than CK. FN had significantly higher nitrification, aerobic ammonia oxidation, manganese oxidation, chloroplasts, aerobic nitrite oxidation, and predatory or exoparasitic and lower chemoheterotrophy and chitinolysis than CK.
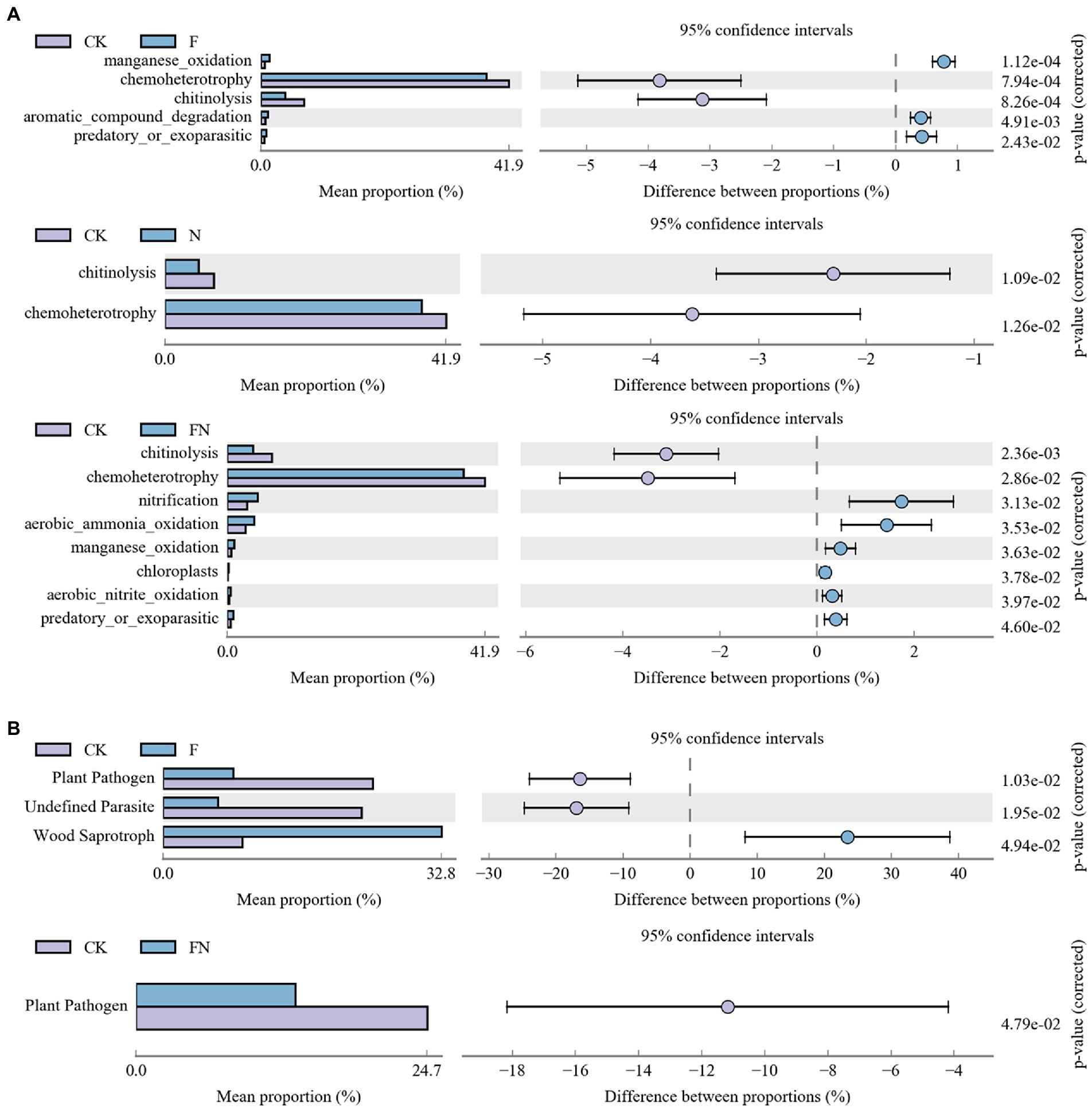
Figure 6. Bacterial (A) and fungal (B) functions affected by different treatments. CK, non-inoculated; F, inoculated with B. velezensis FH-1; N, inoculated with B. diminuta NYM-3; FN, inoculated with B. velezensis FH-1 and B. diminuta NYM-3.
Fungal function prediction (Guild) was also analyzed with OTU abundance >0.1% (Supplementary Figure S6B). The difference analysis results showed that F had a significantly higher wood saprotroph and lower plant pathogens and undefined parasites than CK (Figure 6B). FN had a significantly lower plant pathogen than CK. There was no significant difference between N and CK.
The rice, soil, and microbial variables significantly affected by inoculations (F/N/FN) were selected for the correlation analysis. The results showed that the enriched species Uncultured_g_Pseudomonas and Ascomycota, the microbiome (except fungal NMDS2 and inhibited function), and soil pH were significantly positive, while the inhibited species (except Mortierellomycota, Mortierellomycetes, Mortierellales, Mortierellaceae, Mortierella, Spizellomyces and Spizellomyces acuminatus) and the inhibited functions were significantly negatively correlated with rice height, weight and root nitrogen concentration in F (Supplementary Figure S7A). Soil pH was significantly positively correlated with enriched species (except Uncultured_g_Pseudomonas and Magnaporthiopsis poae), microbiome (only bacterial Shannon diversity, bacterial NMDS2, fungal NMDS1, manganese oxidation, aromatic compound degradation, predatory or exoparasitic and wood daprotroph) while negatively correlated with inhibited bacterial species, inhibited fungal species (only Nectriaceae, Chytridiomycota, spizellomycetes, spizellomycetales, spizellomycetaceae) and inhibited functions in F. Inhibited bacterial taxa, Uncultured_g_Pseudomonas and Chytridiomycota had a significantly negative correlation with microbiome (except fungal NMDS2). Other enriched or inhibited taxa only significantly correlated with some variables of the microbiome in F.
Inoculant B. diminuta, enriched species (except Pezizaceae), microbiome (only bacterial and fungal number and bacterial NMDS1) and soil pH were significantly positively correlated, while the inhibited species and functions were significantly negatively correlated with rice (except shoot nitrogen concentration) in N (Supplementary Figure S7B). Soil pH was significantly positively correlated with the inoculant Brevundiomonas diminuta, enriched species (except Pezizaceae), and microbiome (only bacterial and fungal number and bacterial NMDS1), while it was negatively correlated with inhibited species (except alphaproteobacterial, Xanthomonadales, Xanthomonadaceae, S. dolichospermus, Pyronemataceae) and inhibited functions in N. Most enriched and inhibited bacterial taxa had a significant correlation with the microbiome (except fungal beta diversity) in N.
Enriched species, microbiome (except fungal beta diversity and inhibited functions) and soil pH were significantly positive, while the inhibited bacterial species and inhibited functions were negatively correlated with rice in FN (Supplementary Figure S7C). Soil pH was significantly positively correlated with bacterial Shannon diversity but negatively correlated with inhibited bacterial species (except Xanthomonadales and Xanthomonadaceae) and inhibited bacterial functions in FN. Inhibited bacterial taxa had a significant correlation with the microbiome (except fungal beta diversity) in FN.
Variables significantly related to rice were selected for PLS-PM analysis to explore the contribution of inoculants, microbiome and soil to rice growth promotion. The results showed that all the GoFs in the three PLS-PMs were greater than 0.66 (Figure 7). The GoF index is used to measure the overall quality of a model with acceptable “good” values greater than 0.7 (Sanchez, 2013). All the R2 values in the three PLS-PMs are greater than 0.43. R2 indicates the amount of variance in the endogenous latent variable explained by its independent manifest variables. Values greater than 0.6 can be considered good R2 values (Sanchez, 2013). This indicated that the three models are credible.
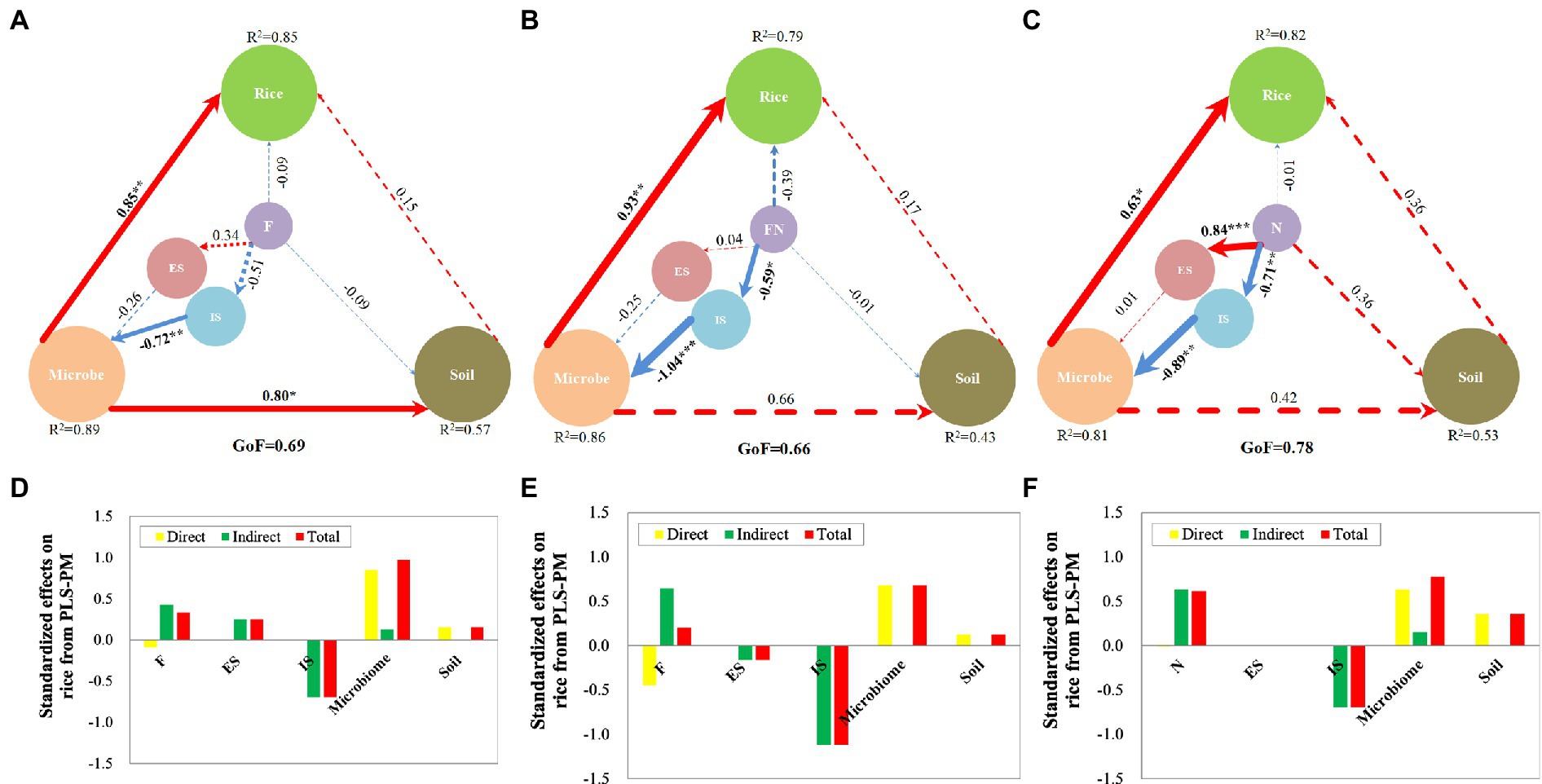
Figure 7. Partial least-squares path model (PLS-PM) in different treatments. (A) PLS-PM of F treatment; (B) PLS-PM of FN treatment; (C) PLS-PM of N treatment; (D) Effects on rice from PLS-PM of F treatment; (E) Effects on rice from PLS-PM of FN treatment; (F) Effects on rice from PLS-PM of N treatment. PLS-PM describing the relationships among microbial inoculants, enriched species (ES), inhibited species (IS), microbiome and soil with respect to rice in different treatments. Larger path coefficients are shown as wider arrows, and red and blue colors indicate positive and negative effects, respectively. Path coefficients and coefficients of determination (R2) were calculated after 999 bootstraps, and significance levels are indicated by *(p < 0.05), **(p < 0.01), and ***(p < 0.001). Models with different structures were assessed using the Goodness of Fit (GoF) statistic, a measure of the overall prediction performance. F, inoculated with B. velezensis FH-1; N, inoculated with B. diminuta NYM-3; FN, inoculated with B. velezensis FH-1 and B. diminuta NYM-3; ES, enriched species; IS, inhibited species.
The results showed that the microbiome contributed more to rice than the soil and inoculants in FN, F and N (Figure 7). The microbiome also contributed more effects on soil than inoculants, especially in F. Inhibited species contributed more effects on the microbiome than enriched species, especially in N and FN. Inoculants had more effects on inhibited species than on enriched species, especially in FN and N, while the opposite was true in N. This indicated that inoculants promoted rice growth mainly by regulating the microbiome.
The manifest variables in FN differed slightly from those in F or N (Table 4). This indicated that the ways in which F, N and FN promoted rice growth were different. B. velezensis FH-1 (F) promoted rice mainly by changing bacterial β diversity (NMDS2), increasing bacterial number, manganese_oxidation, aromatic_compound_degradation, and predatory_or_exoparasitic while reducing chitinolysis function by inhibiting Sphingomonas, Lysobacter, and Nectriaceae and enriching uncultured_g_Pseudomonas and Ascomycota. B. diminuta NYM-3 (N) might promote rice growth mainly by changing bacterial β diversity (NMDS1), increasing bacterial number and reducing chitinolysis function by inhibiting Sphingomonas, Xanthomonadaceae, and Lysobacter and enriching Uncultured_g_Pseudomonas. B. velezensis FH-1 and B. diminuta NYM-3 (FN) might promote rice growth mainly by altering bacterial β diversity (NMDS2), increasing bacterial Shannon diversity, nitrification, aerobic ammonia oxidation, manganese oxidation, chloroplasts, aerobic nitrite oxidation, and predatory or exoparasitic functions while reducing chitinolysis and chemoheterotrophy functions by inhibiting Sphingomonas and Lysobacter and enriching Uncultured_g_Pseudomonas.
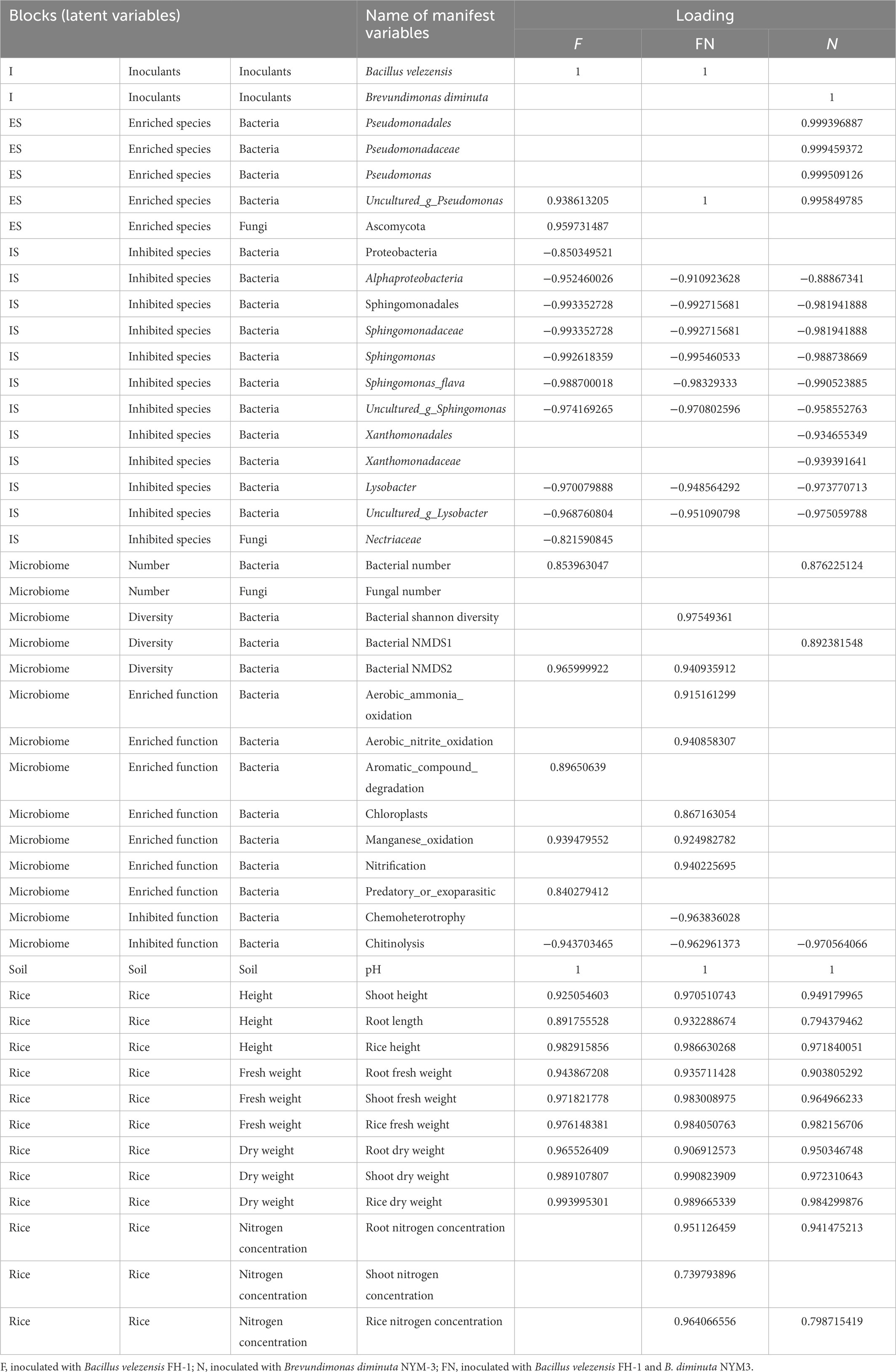
Table 4. Loadings of manifest variables in different PLS-PMs.
B. velezensis (formerly known as Bacillus amyloliquefaciens) is a famous and excellent biopesticide and biofertilizer (Santoyo et al., 2012; Rabbee et al., 2019; Luo et al., 2022). As a commercialized product, it has been successfully used in agriculture for a long time (Wan et al., 2018). Co-inoculation of B. velezensis and some PGPMs (such as Pseudomonas putida, Bradyrhizobium japonicum, Bacillus pumilus, Bacillus licheniformis, Trichoderma harzianum) showed greater promoting effects on crops (tomato, soybean, wheat) than monocultures (He et al., 2019; Oliveira et al., 2019; Sheteiwy et al., 2021). B. diminuta is commonly used for heavy metal remediation, antibiotic degradation and oil degradation (Wang et al., 2016; Liu et al., 2017; Rathi and Yogalakshmi, 2021; Ali et al., 2022). Some works also showed that it could promote the growth of tobacco (Shao et al., 2015). Significant improvement in growth was also observed with co-inoculation of Mesorhizobium sp. and B. diminuta (formerly known as Pseudomonas diminuta) in chickpea compared to single inoculants of Mesorhizobium sp. (Kaur et al., 2015). However, co-inoculation of B. velezensis and B. diminuta to promote crop growth has not been reported. We found that co-inoculation of B. velezensis FH-1 and B. diminuta NYM-3 could significantly promote the growth of riceo-inoculation. Co-inoculation was significantly better than single inoculation, which has great application potential. Our results also showed an antagonism between B. velezensis FH-1 and B. diminuta NYM-3 in vivo and in vitro (Supplementary Figure S2; Figure 3), which has been similarly reported in other literature (Sadiq and Jamil, 2018). This indicated that it may be possible to select species with antagonistic relationship when constructing co-inoculants for natural soil system.
As a famous agent for biofertilizers and biocontrol in agriculture, the plant growth-promoting mechanisms of B. velezensis have been extensively studied (Fan et al., 2018; Rabbee et al., 2019; Luo et al., 2022). It was shown that B. velezensis could promote plant growth by improving soil nutrient availability, secreting hormones and volatile organic compounds (VOCs), changing the soil microbial community and antagonizing pathogens. The plant growth-promoting mechanisms of B. diminuta might be related to the secretion of cytokinin (Shao et al., 2015). However, most of these growth-promoting mechanisms were speculated based on the growth-promoting characteristics of the strains and were not confirmed in pot experiments. Some of these growth-promoting mechanisms have been confirmed in pot experiments, but most of the culture media used are sterilized peat mixtures or sterilized soil rather than complex natural soil (Jiang et al., 2015; Ben Abdallah et al., 2018; Verma and White, 2018; Luo et al., 2022). To develop efficient and stable inoculants for the field, it is necessary to study the growth-promoting mechanism of inoculants in complex natural soil system. In our previous work, the plant growth-promoting characteristics of B. velezensis FH-1 and B. diminuta NYM-3 were investigated. The results showed that both B. velezensis FH-1 and B. diminuta NYM-3 had the ability to fix nitrogen, solubilize phosphate and potassium, and produce siderophores and l-aminocyclopropane-l-carboxylicacid (ACC) deaminase. B. velezensis FH-1 additionally had the ability to antagonize pathogens. B. diminuta NYM-3 additionally had the ability to produce indole-3-acetic acid (IAA) (Zhao et al., 2020). In this study, the results showed that B. velezensis FH-1 and/or B. diminuta NYM-3 promoted the growth of rice mainly by regulating the rhizosphere microbiome rather than by themselves or by improving soil nutrient availability. Our previous study also found that regulating the rhizosphere microbiome may be a meaningful way for B. velezensis FH-1 to promote plant growth (Li et al., 2019; Wang et al., 2021). The regulation of the microbiome as an important growth-promoting pathway of inoculants has been recognized by an increasing number of researchers (Qin et al., 2017; Wang et al., 2017, 2018; Han et al., 2019; Luo et al., 2022). However, the roles of the soil microbiome in promoting growth are mainly based on correlation analysis. More rigorous experiments are still needed to prove the fundamental role of the microbiome in promoting plant growth.
Although all inoculants (F/N/FN) in this study promoted rice growth by regulating the rhizosphere microbiome, different inoculants shaped different microbial structures and functions, resulting in different growth-promoting effects. The better growth promotion effect of co-inoculation was primarily due to the mutual benefit, functional complementarity, or cross-feeding between the two species (Zhang et al., 2016; Figueredo et al., 2017). Few species with antagonistic effects have been reported to promote growth. However, our results showed that FN promotes rice growth mainly by enhancing nitrification function compared with F or N (Figures 1, 6; Table 4; Supplementary Figure S8). The FN increased nitrification (ammonia oxidation and nitrite oxidation) (Figure 6A; Supplementary Figure S8; Supplementary Table S3), soil total nitrogen and available nitrogen (Table 1), rice shoot and root nitrogen contention (Figure 1F) compared with CK. When nitrification was enhanced, ammonium nitrogen was converted into nitrate nitrogen, which may pull nitrogen fixation, increased available nitrogen in soil, and promoted nitrogen absorption by crops (Kuypers et al., 2018; Trivedi et al., 2020). Correlation analysis showed that nitrification and rice dry weight had no significant correlation with soil total nitrogen and available nitrogen (Supplementary Figure S10). Nitrification is significantly related to crop nitrogen content and crop dry weight, and crop nitrogen content is significantly related to crop dry weight (Supplementary Figure S10). This indicated that nitrification promoted rice growth by increasing nitrogen absorption.
Fapprotax analysis showed that nitrification function was mainly contributed by Nitrosospira, Candidatus_Nitrososphaera, uncultured_f_Nitrososphaeraceae, Ellin6067, mle1-7, IS-44, MND1, oc32 and Nitrospira contributions (Supplementary Table S3). The total abundance of these species was also the highest in FN (Supplementary Figure S9). Candidatus_Nitrososphaera and uncultured_bacterium_ f_ Nitrososphaeraceae directly interacted with Bacillus in the cooccurrence network of FN (Supplementary Table S2). This indicated that B. velezensis FH-1 and B. diminuta NYM-3 interfered with each other’s colonization and changed the interaction among species directly or indirectly after co-inoculation, resulting in FN enrichment and inhibition of some species that were enriched and inhibited by F (Supplementary Figure S4). Then, FN enhanced the abundance of species related to nitrification function, thus improving the nitrification activity of soil microorganisms and promoting the absorption of nitrogen nutrients and the growth of rice (Figures 1, 6; Supplementary Figure S10).
Inoculants mainly enriched or inhibited species through indirect interactions. Only Bacillus in FN and F directly interacted with the inhibited species Lysobacter. However, other studies showed that in coculture, Bacillus promoted the growth of Lysobacter, but Lysobacter inhibited the growth of Bacillus (Wei et al., 2021). How inoculants enriched or inhibited related taxa still needs further study. The species that directly interacted with Bacillus or Brevundimonas in co-inoculation (FN) and monoinoculation (F or N) were different. There were 10 identical species directly interacting with Bacillus in F and FN. Only 1 identical specie directly interacted with Brevundimonas in N and FN. This indicates that different inoculants will affect the interaction between species, which may lead to the difference in the whole bacterial network, thus resulting in the difference in bacterial structure and function. The species that directly interacted with Bacillus in the rhizosphere soil of rice and cucumber inoculated with B. velezensis FH-1 were also different, which indicated that crop and environmental factors could also significantly affect the interaction between species (Wang et al., 2021). However, the direct interaction between Bacillus and Actinobacteria MB-A2-108 existed in all B. velezensis FH-1 inoculation treatments, indicating that the interaction between species also has a certain robustness. In-depth analysis of the interaction mechanism and influencing factors among species will guide engineering microbiomes.
All inoculations (F/N/FN) enriched Uncultured_g_Pseudomonas and inhibited Sphingomonas flava, Uncultured_g_Sphingomonas and its genus Sphingomonas, family Sphingomonadaceae, order Sphingomonadales and class Alphaproteobacteria and inhibited Uncultured_g_Lysobacter and its genus Lysobacter, family Xanthomonadaceae and order Xanthomonadales (Figure 5; Supplementary Figure S4). These inoculants may regulate these bacteria to form similar bacterial communities (Figure 2C). This implies that the rhizosphere bacterial communities might apply to the Anna Karenina principle that applies to animal and plant microbiomes (Zaneveld et al., 2017; Arnault et al., 2022). That is, “The rhizosphere bacterial communities are more similar in all healthier plants.” In-depth exploration of the rules may provide theoretical guidance for engineering microbiomes. Most members of Pseudomonas are plant growth-promoting bacteria (Costa-Gutierrez et al., 2020; Li et al., 2022). Some studies have also found that inoculation with Bacillus can stimulate the growth of Pseudomonas (Qin et al., 2017; Wan et al., 2018; Sun et al., 2021). Although some Sphingomonas species have been reported to promote plant growth under stress conditions, the more prominent function of Sphingomonas is to remediate environmental contamination (Asaf et al., 2020; Zhou et al., 2022). Lysobacter possesses many lytic enzymes and plays a key role in the degradation of complex macromolecules and plant pathogens present in soil (Brescia et al., 2020; Moon et al., 2021). Functional analysis showed that Sphingomonas, Lysobacter and other inhibited species mainly contributed to chitinolysis and chemoheterotrophy. The reasons for the negative correlation between inhibited species and rice still need to be further analyzed.
Network analysis showed that the potential interaction between bacteria was significantly higher than that between fungi. The interaction between bacteria and fungi was also weaker. These results are supported by other studies (Pan et al., 2021). Bacterial inoculants (F/N/FN) mainly affect fungi through indirect effects. Although inoculants also significantly affected fungal community structure and function, fungal species contributed less to rice growth than bacteria (Supplementary Figure S8; Table 4). F and FN significantly reduced the plant pathogen spizellomycetaceae; although it was also negatively correlated with rice, the correlation was not strong. Some studies have also found that bacteria are more closely related to crop growth than fungi (de Vries et al., 2018).
In conclusion, this study revealed the co-inoculation of antagonistic B. velezensis FH-1 (F) and B. diminuta NYM3 (N) can significantly promote the growth of rice compared with mono-inoculation. Inoculants (F/N/FN) promoted the growth of rice mainly by regulating the rhizosphere microbiome rather than by themselves or by improving soil nutrient availability. FN promoted rice growth specifically by enhancing microbial nitrification function through enriching related species compared with F or N. Overall, the results of this study provide useful information for the construction and application of co-inoculants in the future. However, further investigation is crucial with sterile system and multi-omics to provide more accurate information on the roles of rhizosphere microbiome.
The data presented in the study are deposited in the NCBI repository, accession number PRJNA804354, https://dataview.ncbi.nlm.nih.gov/object/PRJNA804354.
JW led the overall study, contributed to the study design, data collection and interpretation, and wrote the manuscript. SZ contributed to the data collection and data analysis. SX, WZ, and XZ, contributed to the data interpretation and manuscript edits. YL and HZ contributed to the data collection and interpretation. ZH contributed to study design and manuscript edits. All authors contributed to the article and approved the submitted version.
This study was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA28030203); Natural Science Foundation of Tianjin (20JCYBJC01220); Science and Technology Partnership Program, Ministry of Science and Technology of China (KY202001017); Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project (TSBICIP-IJCP-001).
The authors would like to thank all participants who participated in the study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1101773/full#supplementary-material
Ali, A., Li, M., Su, J. F., Li, Y. F., Wang, Z., Bai, Y. H., et al. (2022). Brevundimonas diminuta isolated from mines polluted soil immobilized cadmium (Cd2+) and zinc (Zn2+) through calcium carbonate precipitation: microscopic and spectroscopic investigations. Sci. Total Environ. 813:152668. doi: 10.1016/j.scitotenv.2021.152668
Anith, K. N., Nysanth, N. S., and Natarajan, C. (2021). Novel and rapid agar plate methods for in vitro assessment of bacterial biocontrol isolates’ antagonism against multiple fungal phytopathogens. Lett. Appl. Microbiol. 73, 229–236. doi: 10.1111/lam.13495
Ansari, F. A., and Ahmad, I. (2019). Fluorescent Pseudomonas-Fap2 and Bacillus licheniformis interact positively in biofilm mode enhancing plant growth and photosynthetic attributes. Sci. Rep. 9:4547. doi: 10.1038/s41598-019-40864-4
Arnault, G., Mony, C., and Vandenkoornhuyse, P. (2022). Plant microbiota dysbiosis and the Anna Karenina Principle. Trends Plant Sci. 28, 18–30. doi: 10.1016/j.tplants.2022.08.012
Asaf, S., Numan, M., Khan, A. L., and Al-Harrasi, A. (2020). Sphingomonas: from diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 40, 138–152. doi: 10.1080/07388551.2019.1709793
Awasthi, A. (2019). Field-specific microbial consortia are feasible: a response to Kaminsky et al. Trends Biotechnol. 37, 569–572. doi: 10.1016/j.tibtech.2019.03.002
Backer, R., Rokem, J. S., Ilangumaran, G., Lamont, J., Praslickova, D., Ricci, E., et al. (2018). Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of Biostimulants for sustainable agriculture. Front. Plant Sci. 9:1473. doi: 10.3389/fpls.2018.01473
Basu, A., Prasad, P., Das, S. N., Kalam, S., Sayyed, R. Z., Reddy, M. S., et al. (2021). Plant growth promoting rhizobacteria (PGPR) as green bioinoculants: recent developments, constraints, and prospects. Sustainability 13:1140. doi: 10.3390/su13031140
Begum, N., Wang, L., Ahmad, H., Akhtar, K., Roy, R., Khan, M. I., et al. (2022). Co-inoculation of arbuscular mycorrhizal fungi and the plant growth-promoting rhizobacteria improve growth and photosynthesis in tobacco under drought stress by up-regulating antioxidant and mineral nutrition metabolism. Microb. Ecol. 83, 971–988. doi: 10.1007/s00248-021-01815-7
Ben Abdallah, D., Frikha-Gargouri, O., and Tounsi, S. (2018). Rizhospheric competence, plant growth promotion and biocontrol efficacy of Bacillus amyloliquefaciens subsp plantarum strain 32a. Biol. Control 124, 61–67. doi: 10.1016/j.biocontrol.2018.01.013
Brescia, F., Pertot, I., and Puopolo, G. (2020). “Chapter 16 - Lysobacter” in Beneficial Microbes in Agro-Ecology. eds. N. Amaresan, M. Senthil Kumar, K. Annapurna, K. Kumar, and A. Sankaranarayanan (Academic Press).
Costa-Gutierrez, S. B., Lami, M. J., Caram-Di Santo, M. C., Zenoff, A. M., Vincent, P. A., Molina-Henares, M. A., et al. (2020). Plant growth promotion by Pseudomonas putida Kt2440 under saline stress: role of eptA. Appl. Microbiol. Biotechnol. 104, 4577–4592. doi: 10.1007/s00253-020-10516-z
De Vries, F. T., Griffiths, R. I., Bailey, M., Craig, H., Girlanda, M., Gweon, H. S., et al. (2018). Soil bacterial networks are less stable under drought than fungal networks. Nat. Commun. 9:3033. doi: 10.1038/s41467-018-05516-7
Emmanuel, O. C., and Babalola, O. O. (2020). Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 239:126569. doi: 10.1016/j.micres.2020.126569
Fan, B., Wang, C., Song, X. F., Ding, X. L., Wu, L. M., Wu, H. J., et al. (2018). Bacillus velezensis Fzb42 in 2018: the gram-positive model strain for plant growth promotion and biocontrol. Front. Microbiol. 9:2491. doi: 10.3389/fmicb.2018.02491
Figueredo, M. S., Tonelli, M. L., Ibanez, F., Morla, F., Cerioni, G., Tordable, M. D., et al. (2017). Induced systemic resistance and symbiotic performance of peanut plants challenged with fungal pathogens and co-inoculated with the biocontrol agent Bacillus sp Chep5 and Bradyrhizobium sp Semia6144. Microbiol. Res. 197, 65–73. doi: 10.1016/j.micres.2017.01.002
Fox, S. L., O’hara, G. W., and Brau, L. (2011). Enhanced nodulation and symbiotic effectiveness of Medicago truncatula when co-inoculated with Pseudomonas fluorescens Wsm3457 and Ensifer (Sinorhizobium) medicae Wsm419. Plant Soil 348, 245–254. doi: 10.1007/s11104-011-0959-8
Han, L., Wang, Z., Li, N., Wang, Y., Feng, J., and Zhang, X. (2019). Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community. Appl. Soil Ecol. 136, 55–66. doi: 10.1016/j.apsoil.2018.12.011
He, Y., Pantigoso, H. A., Wu, Z., and Vivanco, J. M. (2019). Co-inoculation of Bacillus sp. and Pseudomonas putida at different development stages acts as a biostimulant to promote growth, yield and nutrient uptake of tomato. J. Appl. Microbiol. 127, 196–207. doi: 10.1111/jam.14273
Hu, J., Wei, Z., Weidner, S., Friman, V.-P., Xu, Y.-C., Shen, Q.-R., et al. (2017). Probiotic Pseudomonas communities enhance plant growth and nutrient assimilation via diversity-mediated ecosystem functioning. Soil Biol. Biochem. 113, 122–129. doi: 10.1016/j.soilbio.2017.05.029
Ji, C., Zhang, M., Kong, Z., Chen, X., Wang, X., Ding, W., et al. (2021). Genomic analysis reveals potential mechanisms underlying promotion of tomato plant growth and antagonism of soilborne pathogens by Bacillus amyloliquefaciens Ba13. Microbiol Spectr 9:e0161521. doi: 10.1128/Spectrum.01615-21
Jiang, C. H., Wu, F., Yu, Z. Y., Xie, P., Ke, H. J., Li, H. W., et al. (2015). Study on screening and antagonistic mechanisms of Bacillus amyloliquefaciens 54 against bacterial fruit blotch (BFB) caused by Acidovorax avenae subsp citrulli. Microbiol. Res. 170, 95–104. doi: 10.1016/j.micres.2014.08.009
Kaminsky, L. M., Trexler, R. V., Malik, R. J., Hockett, K. L., and Bell, T. H. (2019). The inherent conflicts in developing soil microbial inoculants. Trends Biotechnol. 37, 140–151. doi: 10.1016/j.tibtech.2018.11.011
Kaur, N., Sharma, P., and Sharma, S. (2015). Co-inoculation of Mesorhizobium sp and plant growth promoting rhizobacteria Pseudomonas sp as bio-enhancer and bio-fertilizer in chickpea (Cicer arietinum L.). Legum. Res. 38, 367–374. doi: 10.5958/0976-0571.2015.00099.5
Khan, S. T. (2022). Consortia-based microbial inoculants for sustaining agricultural activities. Appl. Soil Ecol. 176:104503. doi: 10.1016/j.apsoil.2022.104503
Kong, Z. Y., and Liu, H. G. (2022). Modification of rhizosphere microbial communities: a possible mechanism of plant growth promoting rhizobacteria enhancing plant growth and fitness. Front. Plant Sci. 13:920813. doi: 10.3389/fpls.2022.920813
Kuypers, M. M. M., Marchant, H. K., and Kartal, B. (2018). The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 16, 263–276. doi: 10.1038/nrmicro.2018.9
Li, Q., Li, H. C., Yang, Z., Cheng, X., Zhao, Y. C., Qin, L., et al. (2022). Plant growth-promoting rhizobacterium Pseudomonas sp. Cm11 specifically induces lateral roots. New Phytol. 235, 1575–1588. doi: 10.1111/nph.18199
Li, Q., Xu, S., Zhao, W., Yang, R., Zhao, S., Huang, Z., et al. (2019). Rhizosphere microbiome mediated growth-promoting mechanisms of Bacillus amyloliquefaciens Fh-1 on rice. Acta Microbiol Sin. 59, 1–17. doi: 10.13343/j.cnki.wsxb.20190068
Lin, T. H., and Pan, T. M. (2019). Characterization of an antimicrobial substance produced by Lactobacillus plantarum Ntu 102. J. Microbiol. Immunol. Infect. 52, 409–417. doi: 10.1016/j.jmii.2017.08.003
Liu, Y., Chang, H. Q., Li, Z. J., Feng, Y., Cheng, D. M., and Xue, J. M. (2017). Biodegradation of gentamicin by bacterial consortia Amqd4 in synthetic medium and raw gentamicin sewage. Sci. Rep. 7:11004. doi: 10.1038/s41598-017-18286-x
Luo, L., Zhao, C. Z., Wang, E. T., Raza, A., and Yin, C. Y. (2022). Bacillus amyloliquefaciens as an excellent agent for biofertilizer and biocontrol in agriculture: an overview for its mechanisms. Microbiol. Res. 259:127016. doi: 10.1016/j.micres.2022.127016
Moon, J. H., Won, S. J., Maung, C. E. H., Choi, J. H., Choi, S. I., Ajuna, H. B., et al. (2021). The role of Lysobacter antibioticus Hs124 on the control of fall webworm (Hyphantria cunea Drury) and growth promotion of Canadian poplar (Populus canadensis Moench) at Saemangeum reclaimed land in Korea. Microorganisms 9:1580. doi: 10.3390/microorganisms9081580
Moreira, H., Pereira, S. I. A., Vega, A., Castro, P. M. L., and Marques, A. (2020). Synergistic effects of arbuscular mycorrhizal fungi and plant growth-promoting bacteria benefit maize growth under increasing soil salinity. J. Environ. Manag. 257:109982. doi: 10.1016/j.jenvman.2019.109982
Neemisha,, Kumar, A., Sharma, P., Kaur, A., Sharma, S., and Jain, R. (2022). Harnessing rhizobacteria to fulfil inter-linked nutrient dependency on soil and alleviate stresses in plants. J. Appl. Microbiol. 133, 2694–2716. doi: 10.1111/jam.15649
Olenska, E., Malek, W., Wojcik, M., Swiecicka, I., Thijs, S., and Vangronsveld, J. (2020). Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: a methodical review. Sci. Total Environ. 743:140682. doi: 10.1016/j.scitotenv.2020.140682
Oliveira, L. B. G. D., Teixeira Filho, M. C. M., Galindo, F. S., Nogueira, T. A. R., Barco Neto, M., and Buzetti, S. (2019). Formas E tipos de coinoculação na cultura da soja no Cerrado. Rev. Ciências Agrárias 42, 61–70. doi: 10.19084/rca.15828
Oszust, K., and Frąc, M. (2020). Apple pomace microbiome carrying fungal load against phytopathogens – considerations regarding application in agriculture and horticulture. BioResource 15, 945–966. doi: 10.15376/biores.15.1.945-966
Pan, Y. Q., Kang, P., Hu, J. P., and Song, N. P. (2021). Bacterial community demonstrates stronger network connectivity than fungal community in desert-grassland salt marsh. Sci. Total Environ. 798:149118. doi: 10.1016/j.scitotenv.2021.149118
Qin, Y. X., Shang, Q. M., Zhang, Y., Li, P. L., and Chai, Y. R. (2017). Bacillus amyloliquefaciens L-S60 reforms the rhizosphere bacterial community and improves growth conditions in cucumber plug seedling. Front. Microbiol. 8:2620. doi: 10.3389/fmicb.2017.02620
Rabbee, M. F., Ali, M. S., Choi, J., Hwang, B. S., Jeong, S. C., and Baek, K. H. (2019). Bacillus velezensis: a valuable member of bioactive molecules within plant microbiomes. Molecules 24:1046. doi: 10.3390/molecules24061046
Rathi, M., and Yogalakshmi, K. N. (2021). Brevundimonas diminuta Mys6 associated Helianthus annuus L. for enhanced copper phytoremediation. Chemosphere 263:128195. doi: 10.1016/j.chemosphere.2020.128195
Rehman, M. M. U., Zhu, Y., Abrar, M., Khan, W., Wang, W., Iqbal, A., et al. (2022). Moisture- and period-dependent interactive effects of plant growth-promoting rhizobacteria and Am fungus on water use and yield formation in dryland wheat. Plant Soil. doi: 10.1007/s11104-022-05641-9
Sadiq, H., and Jamil, N. (2018). Antagonistic behaviour of organic compounds from Bacillus species and Brevundimonas specie. Pak. J. Pharm. Sci. 31, 919–926.
Santoyo, G., Guzman-Guzman, P., Parra-Cota, F. I., Santos-Villalobos, S. D., Orozco-Mosqueda, M. D., and Glick, B. R. (2021). Plant growth stimulation by microbial consortia. Agronomy 11:219. doi: 10.3390/agronomy11020219
Santoyo, G., Orozco-Mosqueda, M. D., and Govindappa, M. (2012). Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: a review. Biocontrol Sci. Tech. 22, 855–872. doi: 10.1080/09583157.2012.694413
Sarathambal, C., Dinesh, R., Srinivasan, V., Sheeja, T. E., Jeeva, V., and Manzoor, M. (2022). Changes in bacterial diversity and composition in response to co-inoculation of arbuscular mycorrhizae and zinc-solubilizing bacteria in turmeric rhizosphere. Curr. Microbiol. 79:4. doi: 10.1007/s00284-021-02682-8
Shao, L., Liu, K., Li, Z., Zhang, Z., Zhao, W., Yao, L., et al. (2015). Screening, identification and application of the strain Y-P22 producing cytokinins in phyllosphere of tobacco. J. Shandong Agric. Univ. Nat. Sci. 46, 194–197.
Sheteiwy, M. S., Abd Elgawad, H., Xiong, Y. C., Macovei, A., Brestic, M., Skalicky, M., et al. (2021). Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol. Plant. 172, 2153–2169. doi: 10.1111/ppl.13454
Sibponkrung, S., Kondo, T., Tanaka, K., Tittabutr, P., Boonkerd, N., Yoshida, K., et al. (2020). Co-inoculation of Bacillus velezensis strain S141 and Bradyrhizobium strains promotes nodule growth and nitrogen fixation. Microorganisms 8:678. doi: 10.3390/microorganisms8050678
Singh, M., Singh, D., Gupta, A., Pandey, K. D., Singh, P. K., and Kumar, A. (2019). “Plant growth promoting rhizobacteria: application in biofertilizers and biocontrol of phytopathogens” in Pgpr Amelioration in Sustainable Agriculture. eds. A. K. Singh, A. Kumar, and P. K. Singh (Woodhead Publishing).
Singh, B. K., Trivedi, P., Egidi, E., Macdonald, C. A., and Delgado-Baquerizo, M. (2020). Crop microbiome and sustainable agriculture. Nat. Rev. Microbiol. 18, 601–602. doi: 10.1038/s41579-020-00446-y
Sun, X. L., Xu, Z. H., Xie, J. Y., Hesselberg-Thomsen, V., Tan, T. M., Zheng, D. Y., et al. (2021). Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J. 16, 774–787. doi: 10.1038/s41396-021-01125-3
Trivedi, P., Leach, J. E., Tringe, S. G., Sa, T., and Singh, B. K. (2020). Plant-microbiome interactions: from community assembly to plant health. Nat. Rev. Microbiol. 18, 607–621. doi: 10.1038/s41579-020-0412-1
Verma, S. K., and White, J. F. (2018). Indigenous endophytic seed bacteria promote seedling development and defend against fungal disease in browntop millet (Urochloa ramosa L.). J. Appl. Microbiol. 124, 764–778. doi: 10.1111/jam.13673
Waltz, E. (2017). A new crop of microbe startups raises big bucks, takes on the establishment. Nat. Biotechnol. 35, 1120–1122. doi: 10.1038/nbt1217-1120
Wan, T. T., Zhao, H. H., and Wang, W. (2018). Effects of the biocontrol agent Bacillus amyloliquefaciens Sn16-1 on the rhizosphere bacterial community and growth of tomato. J. Phytopathol. 166, 324–332. doi: 10.1111/jph.12690
Wang, J., Li, Q., Xu, S., Zhao, W., Lei, Y., Song, C., et al. (2018). Traits-based integration of multi-species inoculants facilitates shifts of indigenous soil bacterial community. Front. Microbiol. 9:3354. doi: 10.3389/fmicb.2018.03354
Wang, X., Wang, X. J., Liu, M., Zhou, L. J., Gu, Z. L., and Zhao, J. F. (2016). Bioremediation of marine oil pollution by Brevundimonas diminuta: effect of salinity and nutrients. Desalin. Water Treat. 57, 19768–19775. doi: 10.1080/19443994.2015.1106984
Wang, J., Wang, H., Yin, T., Xu, S., Zhao, W., Wang, J., et al. (2017). The persistence and performance of phosphate-solubilizing Gluconacetobacter liquefaciens qzr14 in a cucumber soil. 3 Biotech 7:294. doi: 10.1007/s13205-017-0926-z
Wang, J. J., Xu, S., Yang, R., Zhao, W., Zhu, D., Zhang, X. X., et al. (2021). Bacillus amyloliquefaciens Fh-1 significantly affects cucumber seedlings and the rhizosphere bacterial community but not soil. Sci. Rep. 11:12055. doi: 10.1038/s41598-021-91399-6
Wei, L. F., Yang, J., Ahmed, W., Xiong, X. Y., Liu, Q., Huang, Q., et al. (2021). Unraveling the association between metabolic changes in inter-genus and intra-genus bacteria to mitigate clubroot disease of Chinese cabbage. Agronomy 11:2424. doi: 10.3390/agronomy11122424
Zaneveld, J. R., Mcminds, R., and Vega Thurber, R. (2017). Stress and stability: applying the Anna Karenina principle to animal microbiomes. Nat. Microbiol. 2:17121. doi: 10.1038/nmicrobiol.2017.121
Zeffa, D. M., Fantin, L. H., Koltun, A., De Oliveira, A. L. M., Nunes, M., Canteri, M. G., et al. (2020). Effects of plant growth-promoting rhizobacteria on co-inoculation with Bradyrhizobium in soybean crop: a meta-analysis of studies from 1987 to 2018. PeerJ 8:e7905. doi: 10.7717/peerj.7905
Zhang, L., Feng, G., and Declerck, S. (2018). Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phytate mineralization by a phosphate solubilizing bacterium. ISME J. 12, 2339–2351. doi: 10.1038/s41396-018-0171-4
Zhang, W., Wang, H., Wang, X., Xie, X., Siddikee, M. A., Xu, R., et al. (2016). Enhanced nodulation of peanut when co-inoculated with fungal endophyte Phomopsis liquidambari and Bradyrhizobium. Plant Physiol. Biochem. 98, 1–11. doi: 10.1016/j.plaphy.2015.11.002
Zhao, S., Wang, J., Yang, Z., Li, Q., Yang, R., Zhao, W., et al. (2020). Preparation of microbial compound agents. Microbiol. China 47, 1492–1502. doi: 10.13344/j.microbiol.china.190700
Keywords: PGPM, microbial inoculants, qPCR, diversity, network, LEfSe, FAPROTAX, PLS-PM
Citation: Wang J, Zhao S, Xu S, Zhao W, Zhang X, Lei Y, Zhai H and Huang Z (2023) Co-inoculation of antagonistic Bacillus velezensis FH-1 and Brevundimonas diminuta NYM3 promotes rice growth by regulating the structure and nitrification function of rhizosphere microbiome. Front. Microbiol. 14:1101773. doi: 10.3389/fmicb.2023.1101773
Edited by:
Bruno Tilocca, Magna Græcia University, ItalyReviewed by:
Shrivardhan Dheeman, MVN University, IndiaCopyright © 2023 Wang, Zhao, Xu, Zhao, Zhang, Lei, Zhai and Huang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jingjing Wang, ✉ d2FuZ19qakB0aWIuY2FzLmNu; Zhiyong Huang, ✉ aHVhbmdfenlAdGliLmNhcy5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.