 Kai-Yue Luo1,2,3
Kai-Yue Luo1,2,3 Chang-Lin Zhao1,2,3,4*
Chang-Lin Zhao1,2,3,4*- 1Key Laboratory for Forest Resources Conservation and Utilization in the Southwest Mountains of China, Ministry of Education, Southwest Forestry University, Kunming, China
- 2Yunnan Key Laboratory of Plateau Wetland Conservation, Restoration and Ecological Services, Southwest Forestry University, Kunming, China
- 3College of Biodiversity Conservation, Southwest Forestry University, Kunming, China
- 4Yunnan Key Laboratory for Fungal Diversity and Green Development, Kunming Institute of Botany, Chinese Academy of Science, Kunming, China
Dead wood-associated fungi play an important role in wood degradation and the recycling of organic matter in the forest ecological system. Xenasmataceae is a cosmopolitan group of wood-rotting fungi that grows on tropical, subtropical, temperate, and boreal vegetation. In this study, a new fungal order, Xenasmatales, is introduced based on both morphology and multigene phylogeny to accommodate Xenasmataceae. According to the internal transcribed spacer and nuclear large subunit (ITS+nLSU) and nLSU-only analyses of 13 orders, Xenasmatales formed a single lineage and then grouped with orders Atheliales, Boletales, and Hymenochaetales. The ITS dataset revealed that the new taxon Xenasmatella nigroidea clustered into Xenasmatella and was closely grouped with Xenasmatella vaga. In the present study, Xenasmatella nigroidea collected from Southern China is proposed as a new taxon, based on a combination of morphology and phylogeny. Additionally, a key to the Xenasmatella worldwide is provided.
Introduction
Among eukaryotic microorganisms, wood-decaying fungi interact positively with dead wood, playing a fundamental ecological role as decomposers of plants in the fungal tree of life (James et al., 2020). Wood-associated fungi are cosmopolitan and rich in diversity since they grow on tropical, subtropical, temperate, and boreal vegetation (Gilbertson and Ryvarden, 1987; Núñez and Ryvarden, 2001; Bernicchia and Gorjón, 2010; Dai, 2012; Ryvarden and Melo, 2014; Dai et al., 2015, 2021; Wu et al., 2020).
Xenasmataceae Oberw., a typical wood-associated fungal group mainly distributed in the tropics was discovered by Oberwinkler (1966), and typified by Xenasma Donk. Three genera, namely, Xenasma, Xenasmatella Oberw., and Xenosperma Oberw., have been accommodated in this family, however, higher-level classification of the order has not been designated. The tenth edition of the Dictionary of the Fungi showed that Xenasmataceae belongs to Polyporales Gäum., and consists of three genera (Kirk et al., 2008). MycoBank indicates that Xenasmataceae has a higher classification within Polyporales, although the Index Fungorum shows that Xenasmataceae belongs to the order Russulales.
High phylogenetic diversity among corticioid homobasidiomycetes suggests a close relationship among Radulomyces M.P. Christ., Xenasmatella, and Coronicium J. Erikss. and Ryvarden. Xenasma pseudotsugae (Burt) J. Erikss. nested into the euagarics clade, in which it grouped with Coronicium and Radulomyces. The three taxa of Radulomyces grouped together with Phlebiella pseudotsugae (Burt) K.H. Larss. and Hjortstam and Coronicium alboglaucum (Bourdot and Galzin) Jülich, and were composed of a rather confusing group with no obvious morphological features or ecological specialization to tie these three genera together (Larsson et al., 2004). The classification of corticioid fungi with 50 putative families from published preliminary analyses and phylogenies of sequence data showed that three species of Xenasmatella formed a single lineage with strong support within the unplaced Phlebiella family, in which this clade was unclaimed to any orders (Larsson, 2007). A higher-level phylogenetic classification of the Kingdom Fungi revealed that the Phlebiella clade and Jaapia clade do not show affinities within any orders (Hibbett et al., 2007). An outline of all genera of Basidiomycota with combined SSU, ITS, LSU, tef1, rpb1, and rpb2 datasets showed that Xenasmatella was assigned to Xenasmataceae within the order Russulales (He et al., 2019). Therefore, there is debate on the classification at the order level for the Xenasmataceae.
Recently, Xenasmatella has been studied deeply on the basis of morphology and phylogeny. Phlebiella P. Karst. was deemed to have not been legitimately published previously, and the name Xenasmatella was accepted (Duhem, 2010; Larsson et al., 2020; Maekawa, 2021). Molecular systematics involving Xenasmatella was carried out recently. On the basis of morphological and molecular identification, Zong et al. (2021) studied the sequences of 27 fungal specimens representing 24 species between the Xenasmatella clade and related orders; and the Xenasmatella clade formed a single lineage and three new species, namely, X. rhizomorpha C.L. Zhao, X. tenuis C.L. Zhao, and X. xinpingensis C.L. Zhao. Both the MycoBank database (http://www.MycoBank.org) and Index Fungorum (http://www.indexfungorum.org, accessed on June 20, 2022) have recorded 41 specific and infraspecific names in Xenasmatella. To date, the number of Xenasmatella species accepted worldwide has reached 25 (Oberwinkler, 1966; Stalpers, 1996; Hjortstam and Ryvarden, 2005; Bernicchia and Gorjón, 2010; Duhem, 2010; Larsson et al., 2020; Maekawa, 2021), of which, nine species have been found in China (Dai et al., 2004; Dai, 2011; Huang et al., 2019; Zong and Zhao, 2021; Zong et al., 2021).
In the present study, we verified the taxonomy and phylogeny of Xenasmataceae. In addition, we analyzed the species diversity of Xenasmataceae and constructed a phylogeny to the order level of this family on the basis of large subunit nuclear ribosomal RNA gene (nLSU) sequences, the internal transcribed spacer (ITS) regions, and ITS+nLSU analyses. Based on both morphology and phylogeny, we propose a new fungal order, Xenasmatales and a new species, Xenasmatella nigroidea. A key to the 25 accepted species of Xenasmatella worldwide is also provided.
The accepted species list
Xenasma Donk (1957).
1. Xenasma Aculeatum C.E. Gómez (1972).
2. Xenasma Amylosporum Parmasto (1968).
3. Xenasma Longicystidiatum Boidin and Gilles (2000).
4. Xenasma Parvisporum Pouzar (1982).
5. Xenasma Praeteritum (H.S. Jacks.) Donk (1957).
6. Xenasma Pruinosum (Pat.) Donk (1957).
7. Xenasma Pulverulentum (H.S. Jacks.) Donk (1957).
8. Xenasma Rimicola (P. Karst.) Donk (1957).
9. Xenasma Subclematidis S.S. Rattan (1977).
10. Xenasma Tulasnelloideum (Höhn. and Litsch.) Donk (1957).
11. Xenasma Vassilievae Parmasto (1965).
Xenasmatella Oberwinkler (1966).
1. Xenasmatella Ailaoshanensis C.L. Zhao ex C.L. Zhao and T.K. Zong (2021).
2. Xenasmatella Alnicola (Bourdot and Galzin) K.H. Larss. and Ryvarden (2020).
3. Xenasmatella Ardosiaca (Bourdot and Galzin) Stalpers (1996).
4. Xenasmatella Athelioidea (N. Maek.) N. Maek. (2021).
5. Xenasmatella Bicornis (Boidin and Gilles) Piatek (2005).
6. Xenasmatella Borealis (K.H. Larss. and Hjortstam) Duhem (2010).
7. Xenasmatella Caricis-Pendulae (P. Roberts) Duhem (2010).
8. Xenasmatella Christiansenii (Parmasto) Stalpers (1996).
9. Xenasmatella Cinnamomea (Burds. and Nakasone) Stalpers (1996).
10. Xenasmatella Fibrillosa (Hallenb.) Stalpers (1996).
11. Xenasmatella Globigera (Hjortstam and Ryvarden) Duhem (2010).
12. Xenasmatella Gossypina (C.L. Zhao) G. Gruhn and Trichies (2021).
13. Xenasmatella Inopinata (H.S. Jacks.) Hjortstam and Ryvarden (1979).
14. Xenasmatella Insperata (H.S. Jacks.) Jülich (1979).
15. Xenasmatella Nasti Boidin and Gilles ex Stalpers (1996).
16. Xenasmatella Odontioidea Ryvarden and Liberta (1978).
17. Xenasmatella Palmicola (Hjortstam and Ryvarden) Duhem (2010).
18. Xenasmatella Rhizomorpha C.L. Zhao (2021).
19. Xenasmatella Romellii Hjortstam (1983).
20. Xenasmatella Sanguinescens Svrček (1973).
21. Xenasmatella Subflavidogrisea (Litsch.) Oberw. ex Jülich (1979).
22. Xenasmatella Tenuis C.L. Zhao (2021).
23. Xenasmatella Vaga (Fr.) Stalpers (1996).
24. Xenasmatella Wuliangshanensis (C.L. Zhao) G. Gruhn and Trichies (2021).
25. Xenasmatella Xinpingensis C.L. Zhao (2021).
Xenosperma Oberw. (1966).
1. Xenosperma Hexagonosporum Boidin and Gilles (1989).
2. Xenosperma Ludibundum (D.P. Rogers and Liberta) Oberw. ex Jülich (1979).
3. Xenosperma Murrillii Gilb. and M. Blackw. (1987).
4. Xenosperma Pravum Boidin and Gilles (1989).
Materials and methods
Sample collection and herbarium specimen preparation
Fresh fruit bodies of fungi growing on the stumps of angiosperms were collected from Honghe, Yunnan Province, P.R. China. The samples were photographed in situ, and macroscopic details were recorded. Field photographs were taken by a Jianeng 80D camera. All photographs were focus stacked and merged using Helicon Focus software. Once the macroscopic details were recorded, the specimens were transported to a field station where they were dried on an electronic food dryer at 45°C. Once dried, the specimens were labeled and sealed in envelopes and plastic bags. The dried specimens were deposited in the herbarium of the Southwest Forestry University (SWFC), Kunming, Yunnan Province, P.R. China.
Morphology
The macromorphological descriptions were based on field notes and photos captured in the field and laboratory. The color, texture, taste, and odor of fruit bodies were mostly based on the authors' field trip investigations. Rayner (1970) and Petersen (1996) were used for the color terms. All materials were examined under a Nikon 80i microscope. Drawings were made with the aid of a drawing tube. The measurements and drawings were made from slide preparations stained with cotton blue (0.1 mg aniline blue dissolved in 60 g pure lactic acid), melzer's reagent (1.5 g potassium iodide, 0.5 g crystalline iodine, 22 g chloral hydrate, and aq. dest. 20 ml), and 5% potassium hydroxide. Spores were measured from the sections of the tubes; and when presenting spore size data, 5% of the measurements excluded from each end of the range are shown in parentheses (Wu et al., 2022). The following abbreviations were used: KOH = 5% potassium hydroxide water solution, CB = cotton clue, CB– = acyanophilous, IKI = Melzer's reagent, IKI– = both inamyloid and indextrinoid, L = means spore length (arithmetic average for all spores), W = means spore width (arithmetic average for all spores), Q = variation in the L/W ratios between the specimens studied, and n = a/b (number of spores (a) measured from given number (b) of specimens).
Molecular phylogeny
The CTAB rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd., Beijing, P.R. China) was used to obtain genomic DNA from the dried specimens following the manufacturer's instructions (Zhao and Wu, 2017). The nuclear ribosomal ITS region was amplified with the primers ITS5 and ITS4 (White et al., 1990). The nuclear nLSU region was amplified with the primer pairs LR0R and LR7 (http://lutzonilab.org/nuclear-ribosomal-dna/, accessed on September 12, 2021). The PCR procedure used for ITS was as follows: initial denaturation at 95°C for 3 min, followed by 35 cycles at 94°C for 40 s, 58°C for 45 s, and 72°C for 1 min, and a final extension of 72°C for 10 min. The PCR procedure used for nLSU was as follows: initial denaturation at 94°C for 1 min, followed by 35 cycles at 94°C for 30 s, 48°C for 1 min, and 72°C for 1.5 min, and a final extension of 72°C for 10 min. The PCR products were purified and sequenced at Kunming Tsingke Biological Technology Limited Company (Yunnan Province, P.R. China). All the newly generated sequences were deposited in the National Center for Biotechnology Information (NCBI) GenBank (https://www.ncbi.nlm.nih.gov/genbank/, accessed on September 12, 2021) (Table 1).
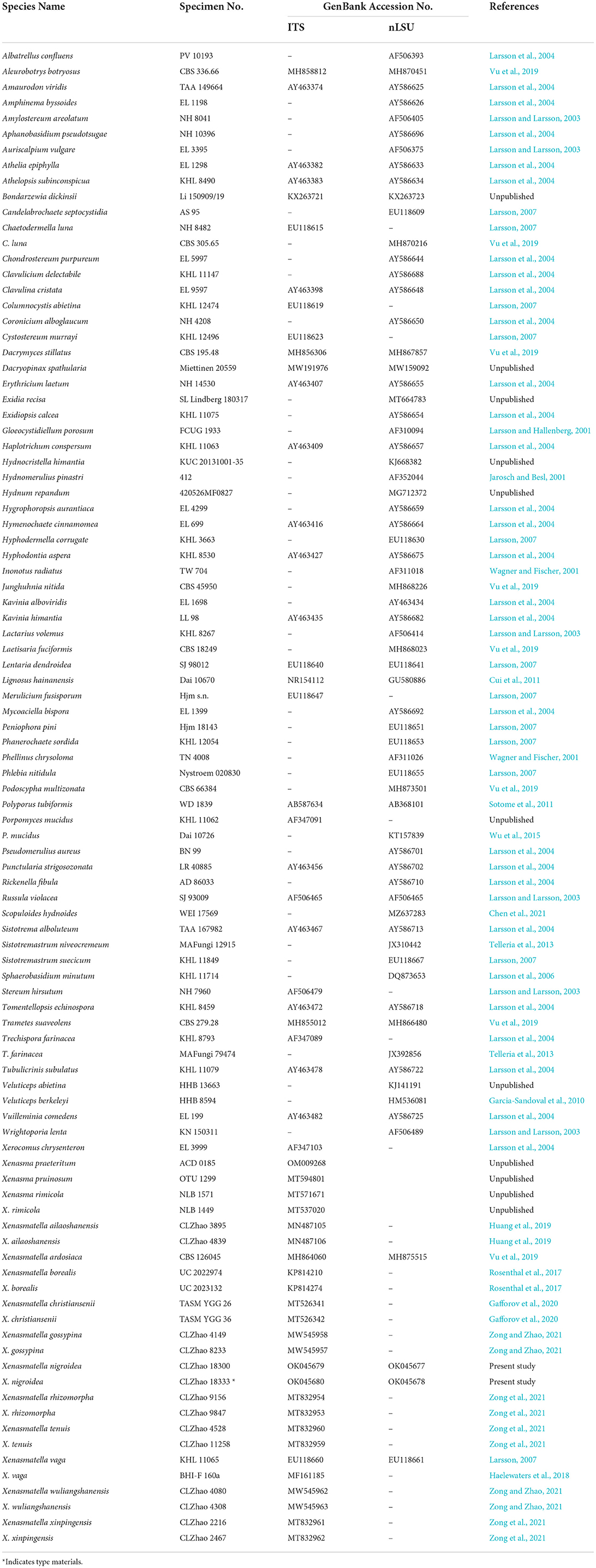
Table 1. The list of species, specimens, and GenBank accession numbers of sequences used in this study.
The sequences and alignment were adjusted manually using AliView version 1.27 (Larsson, 2014). The datasets were aligned with Mesquite version 3.51. The ITS+nLSU dataset and the nLSU-only sequence dataset were used to position a new order, Xenasmatales, and the ITS-only dataset was used to position a new species among the Xenasmatella-related taxa. Sequences of Dacrymyces stillatus and Dacryopinax spathularia retrieved from GenBank were used as the outgroup for the ITS+nLSU sequences (Figure 1) (He et al., 2019); sequences of Exidia recisa and Exidiopsis calcea retrieved from GenBank were used as the outgroup for the nLSU sequences (Figure 2) (Larsson, 2007); and the sequence of Trametes suaveolens was used as the outgroup for the ITS-only sequences (Figure 3) (Zong and Zhao, 2021).
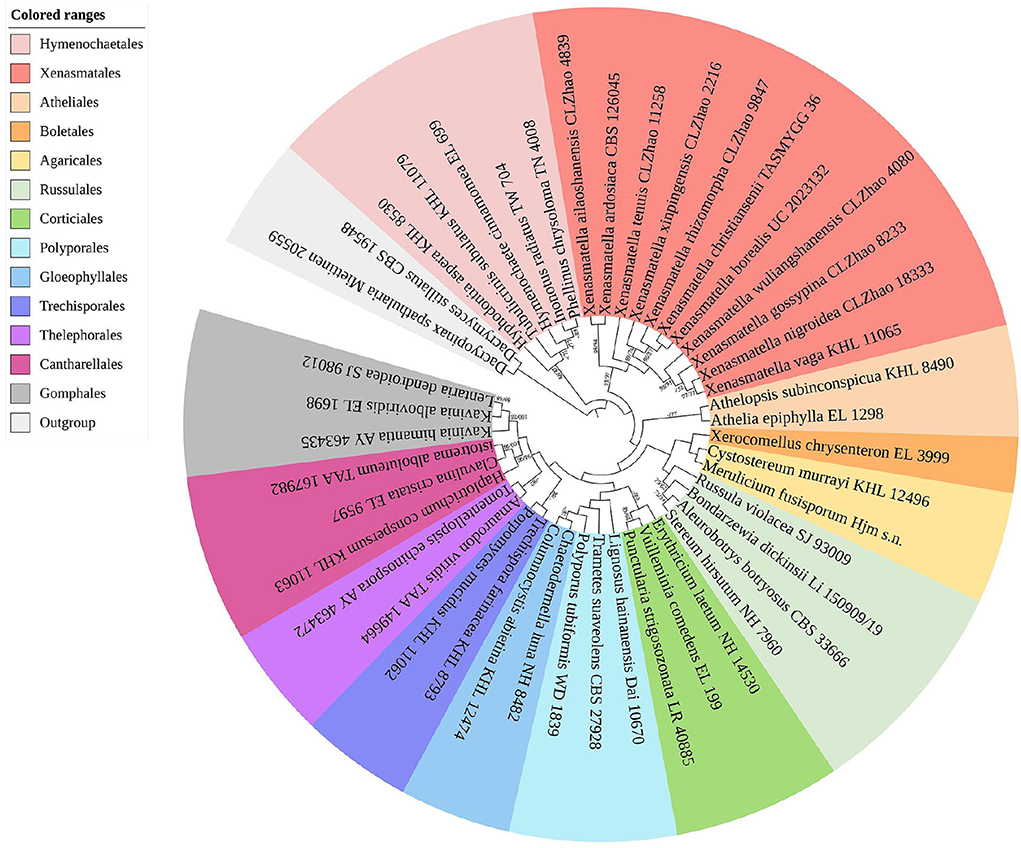
Figure 1. A maximum parsimony strict consensus tree illustrating the phylogeny of the new order Xenasmatales and related order in the class Agaricomycetes based on ITS+nLSU sequences. The orders represented by each color are indicated in the upper left of the phylogenetic tree. Branches are labeled with a maximum likelihood bootstrap value ≥ 70%, and a parsimony bootstrap value ≥ 50, respectively.

Figure 2. A maximum parsimony strict consensus tree illustrating the phylogeny of the new order Xenasmatales and related order in the class Agaricomycetes based on nLSU sequences. The orders represented by each color are indicated in the upper left of the phylogenetic tree. Branches are labeled with a maximum likelihood bootstrap value ≥ 70%, a parsimony bootstrap value ≥ 50%, and Bayesian posterior probabilities ≥ 0.95, respectively.
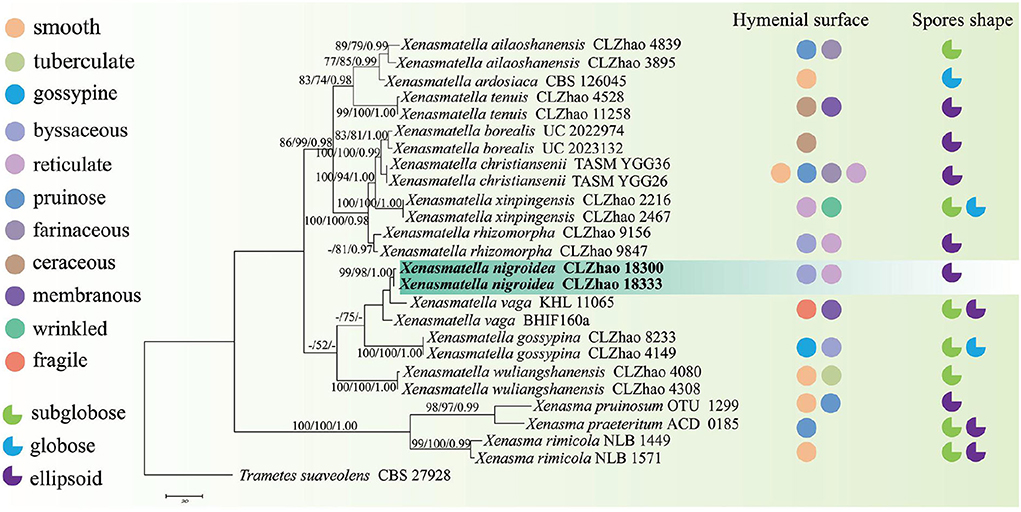
Figure 3. A maximum parsimony strict consensus tree illustrating the phylogeny of a new species and related species in Xenasmatella and Xenasma based on ITS sequences. Branches are labeled with a maximum likelihood bootstrap value ≥ 70%, a parsimony bootstrap value ≥ 50%, and Bayesian posterior probabilities ≥ 0.95, respectively. The new species are in bold.
The three combined datasets were analyzed using maximum parsimony (MP), maximum likelihood (ML), and Bayesian inference (BI), according to Zhao and Wu (2017), and the tree was constructed using PAUP* version 4.0b10 (Swofford, 2002). All characters were equally weighted and gaps were treated as missing data. Trees were inferred using the heuristic search option with TBR branch swapping and 1,000 random sequence additions. Max-trees were set to 5,000, branches of zero length were collapsed, and all parsimonious trees were saved. Clade robustness was assessed using the bootstrap (BT) analysis with 1,000 replicates (Felsenstein, 1985). Descriptive tree statistics—tree length (TL), consistency index (CI), retention index (RI), rescaled consistency index (RC), and homoplasy index (HI)—were calculated for each maximum parsimonious tree generated. In addition, multiple sequence alignment was analyzed using ML in RAxML-HPC2 through the Cipres Science Gateway (Miller et al., 2012). Branch support (BS) for ML analysis was determined by 1,000 bootstrap replicates.
MrModeltest 2.3 (Nylander, 2004) was used to determine the best-fit evolution model for each dataset of BI, which was performed using MrBayes 3.2.7a with a GTR+I+G model of DNA substitution and a gamma distribution rate variation across sites (Ronquist et al., 2012). A total of 4 Markov chains were run for 2 runs from random starting trees for 1 million generations for the ITS+nLSU dataset (Figure 1), 1.4 million generations for the nLSU-only sequences (Figure 2), and 0.5 million generations for the ITS-only sequences (Figure 3), with trees and parameters sampled every 1,000 generations. The first one-fourth of all generations was discarded as a burn-in. The majority rule consensus tree of all remaining trees was calculated. Branches were considered significantly supported if they received a maximum likelihood bootstrap value (BS) ≥70%, a maximum parsimony bootstrap value (BT) ≥70%, or Bayesian posterior probabilities (BPP) ≥0.95.
Results
Phylogenetic analyses
The ITS+nLSU dataset (Figure 1) included sequences from 45 fungal specimens representing 45 species. The dataset had an aligned length of 3,095 characters, of which 1,910 characters are constant, 353 are variable and parsimony uninformative, and 832 are parsimony informative. Maximum parsimony analysis yielded 45 equally parsimonious trees (TL = 3,984, CI = 0.4666, HI = 0.5334, RI = 0.3909, and RC = 0.1824). The best model was GTR+I+G [lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1)]. Bayesian and ML analyses showed a topology similar to that of MP analysis with split frequencies equal to 0.009126 (BI), and the effective sample size (ESS) across the two runs is double that of the average ESS (avg ESS) = 250.5.
The ITS+nLSU rDNA gene regions (Figure 1) were based on 13 orders, namely, Agaricales Underw., Atheliales Jülich, Boletales E.J. Gilbert, Cantharellales Gäum., Corticiales K.H. Larss., Gloeophyllales Thorn, Gomphales Jülich, Hymenochaetales Oberw., Polyporales, Russulales, Thelephorales Corner ex Oberw., Trechisporales, and Xenasmatales, while Xenasmatella was separated from the other orders.
The nLSU-alone dataset (Figure 2) included sequences from 58 fungal specimens representing 58 species. The dataset had an aligned length of 1,343 characters, of which 726 characters are constant, 176 are variable and parsimony-uninformative, and 441 are parsimony-informative. Maximum parsimony analysis yielded 3 equally parsimonious trees (TL = 2,864, CI = 0.3209, HI = 0.6791, RI = 0.4476, and RC = 0.1436). The best model for the ITS dataset estimated and applied in the Bayesian analysis was GTR+I+G [lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1)]. The Bayesian and ML analyses resulted in a topology similar to that of MP analysis with split frequencies equal to 0.009830 (BI), and the effective sample size (ESS) across the two runs is double that of the average ESS (avg ESS) = 402.
The nLSU regions (Figure 2) were based on 13 orders, namely, Agaricales, Atheliales, Boletales, Cantharellales, Corticiales, Gloeophyllales, Gomphales, Hymenochaetales, Polyporales, Russulales, Thelephorales, Trechisporales, and Xenasmatales, while Xenasmatella was separated from the other orders.
The ITS-alone dataset (Figure 3) included sequences from 26 fungal specimens representing 15 species belonging to Xenasma and Xenasmatella. The dataset had an aligned length of 598 characters, of which 267 characters are constant, 74 are variable and parsimony-uninformative, and 257 are parsimony-informative. Maximum parsimony analysis yielded 1 equally parsimonious tree (TL = 629, CI = 0.7329, HI = 0.2671, RI = 0.8301, and RC = 0.6084). The best model for the ITS dataset estimated and applied in the Bayesian analysis was GTR+I+G [lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1)]. The Bayesian and ML analyses resulted in a topology similar to MP analysis with split frequencies equal to 0.007632 (BI), and the effective sample size (ESS) across the two runs is double that of the average ESS (avg ESS) = 300.5.
In the ITS sequence analysis (Figure 3), a previously undescribed species was grouped into Xenasmatella with a sister group to X. vaga (Fr.) Stalpers.
Taxonomy
Xenasmatales K.Y. Luo and C.L. Zhao, ord. nov.
MycoBank no.: MB 842882
Type family: Xenasmataceae Oberw.
Basidiomata resupinate. Hyphal systems are monomitic, generative hyphae with clamp connections. Basidia pleural. Basidiospores are colorless.
Xenasmataceae Oberw., Sydowia 19(1–6): 25 (1966).
MycoBank no.: MB 81527
Type genus: Xenasma Donk
Basidiomata resupinate, ceraceous to geletinous. Hyphal systems are monomitic, generative hyphae with clamp connections. Basidia pleural usually with 4 sterigmata and a basal clamp connection. Basidiospores are colorless.
Xenasma Donk, Fungus, Wageningen 27: 25 (1957).
MycoBank no.: MB 18755
Type species: Xenasma rimicola (P. Karst.) Donk.
Basidiomata resupinate, adnate, are ceraceous to gelatinous when fresh, membranaceous when dry, and have a hymenophore smooth. Hyphal system are monomitic, generative hyphae with clamp connections. Cystidia and cystidioles are present. Basidia are cylindrical to subclavate, pleural, usually with 4 sterigmata and a basal clamp connection. Basidiospores are globose to cylindrical, colorless, thin-walled, warted to striate, non-amyloid, and weakly dextrinoid.
Xenosperma Oberw., Sydowia 19(1–6): 45 (1966).
MycoBank no.: MB 18759
Type species: Xenosperma ludibundum (D.P. Rogers and Liberta) Oberw.
Basidiomata resupinate, closely adnate to the substratum, are gelatinous when fresh and pruinose when dry. Hyphal systems are monomitic, generative hyphae with clamp connections. Cystidia are absent. Basidia pleural, usually with 2–4 sterigmata and a basal clamp connection. Basidiospores are angular, colorless, thin-walled, tetrahedral, with some protuberances, IKI–, and CB–.
Xenasmatella Oberw., Sydowia 19(1–6): 28 (1966).
MycoBank no.: MB 18756
Type species: Xenasmatella subflavidogrisea (Litsch.) Oberw. ex Jülich.
Basidiomata resupinate with a gelatinous. Hyphal system with clamped generative hyphae. Cystidia are absent. Basidia pleural, usually with 4 sterigmata and a basal clamp connection. Basidiospores are hyaline, thin-walled, warted, IKI–, and CB–.
Xenasmatella nigroidea K.Y. Luo and C.L. Zhao, sp. nov.
MycoBank no.: MB 842470, Figures 4, 5.

Figure 4. Basidiomata of Xenasmatella nigroidea (holotype). Bars: (A) 1 cm; (B) 1 mm.
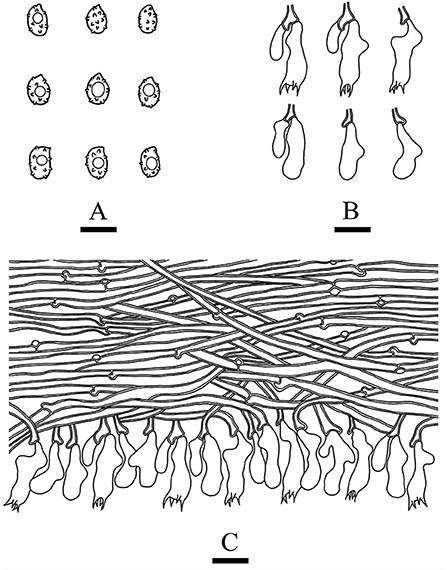
Figure 5. Microscopic structures of Xenasmatella nigroidea (drawn from the holotype). (A) Basidiospores. (B) Basidia and basidioles. (C) A section of hymenium. Bars: (A) 5 μm; (B,C) 10 μm.
Holotype—China. Yunnan Province, Honghe, Pingbian County, Daweishan National Nature Reserve, GPS coordinates 23°42′ N, 103°32′ E, altitude 1,500 m asl., on angiosperm stump, leg. C.L. Zhao, August 3, 2019, CLZhao 18333 (SWFC).
Etymology—nigroidea (Lat.): refers to the black hymenial surface.
Basidiomata: Basidiomata are annuals, resupinate, thin, very hard to separate from substrate, odorless or tasteless when fresh, grayish when fresh, gray to black and brittle when dry, up to 7.5 cm long, 3.5 cm wide, 70–150 μm thick. Hymenial is surface smooth, and byssaceous to reticulate under the lens. Sterile margin indistinct, black, up to 1 mm wide.
Hyphal system: monomitic, generative hyphae with clamp connections, thick-walled, unbranched, 2.5–4 μm in diameter, IKI–, CB–, and tissues unchanged in KOH.
Hymenium: cystidia and cystidioles are absent; basidia are pleural, clavate, with 4 sterigmata and a basal clamp connection, 12.0–18.0 × 4.5–6 μm; basidioles are shaped similar to basidia but slightly smaller.
Basidiospores: ellipsoid, colorless, thin-walled, warted throughout, asperulate with blunt spines up to 0.2 μm long, with one oil drop inside, IKI–, CB–, 3.5–4.5 × 2.5–3.5 μm, L = 4.07 μm, W = 2.87 μm, Q = 1.38–1.45 (n = 60/2).
Type of rot: White rot.
Additional specimen examined: CHINA, Yunnan Province, Honghe, Pingbian County, Daweishan National Nature Reserve, GPS coordinates 23°40′ N, 103°31′ E, altitude 1,500 m asl., on the angiosperm stump, leg. C.L. Zhao, August 3, 2019, CLZhao 18300 (SWFC).
Discussion
There have been debates among mycologists regarding the order level taxonomic status of the Xenasmataceae. Corticioid homobasidiomycetes have a high phylogenetic diversity. Thus, an accurate place for the taxa of Xenasmataceae has not been decided. However, it was only assigned to euagarics clade (Larsson et al., 2004). Later, the Phlebiella family was proposed by Larsson (2007) on the basis of corticioid fungi; however, this group was not placed under any order. Recently, Xenasmataceae was placed under Russulales by He et al. (2019). Zong et al. (2021) studied the specimens and sequences from China and treated this group as Xenasmatella as the phylogenetic datasets showed that this clade does not belong to any order. In the present study (Figure 1), the ITS+nLSU analyses of 13 orders, namely, Agaricales, Atheliales, Boletales, Cantharellales, Corticiales, Gloeophyllales, Gomphales, Hymenochaetales, Polyporales, Russulales, Thelephorales, Trechisporales, and Xenasmatales showed that the taxa of Xenasmataceae form a single lineage with the sequences of Hymenochaetales and Atheliales; and this is similar to the results of Larsson (2007). In the present study (Figure 2), the nLSU analysis showed that the taxa of Xenasmataceae form a single lineage with the sequences of Hymenochaetales and Boletales; and this is similar to the results of Larsson (2007). In the present study (Table 2), we have enumerated morphological differences among the related orders. Therefore, a new fungal order, Xenasmatales, is proposed on the basis of morphological and molecular identification.

Table 2. Morphological characteristics of the relevant orders used in this study.
Phlebiella was not deemed to be a legitimately published genus (Duhem, 2010), and transferring to Xenasmatella was proposed. Later, Larsson et al. (2020) studied corticioid fungi (Basidiomycota and Agaricomycetes) and agreed with Duhem (2010), who suggested accepting the genus Xenasmatella. Recently, several mycologists have suggested the replacement of the invalid genus Phlebiella with Xenasmatella on the basis of morphology and molecular analyses (Maekawa, 2021; Zong et al., 2021).
On the basis of ITS dataset, a previous study showed that nine species of Xenasmatella have been reported, of which 6 new species were found in China, namely, X. ailaoshanensis C.L. Zhao ex C.L. Zhao and T.K. Zong, X. gossypina, X. rhizomorpha, X. tenuis, X. wuliangshanensis, and X. xinpingensis. According to our sequence data, Xenasmatella nigroidea was nested into Xenasmatella with strong statistical support (Figure 3), and formed a sister group with X. vaga. However, X. nigroidea is morphologically distinguished from X. vaga by larger basidiospores (5–5.5 × 4–4.5 μm). In addition, it turns dark red or purplish with KOH (Bernicchia and Gorjón, 2010).
Morphological comparisons of Xenasmatella nigroidea and other species are included in Table 3. Xenasmatella nigroidea is similar to X. christiansenii (Parmasto) Stalpers, X. fibrillosa (Hallenb.) Stalpers, X. gossypina, and X. rhizomorpha C.L. Zhao by having gossypine, byssaceous to reticulate hymenial surface, however, X. christiansenii is distinguished from X. nigroidea by its larger basidiospores (6–7 × 4–4.5 μm) and asperulate with blunt spines (up to 1 μm long; Bernicchia and Gorjón, 2010). Xenasmatella fibrillosa differs from X. nigroidea due to the presence of a white to pale yellowish white hymenial surface and longer basidiospores (4.5–5.5 μm; Bernicchia and Gorjón, 2010). Xenasmatella gossypina can be distinguished from X. nigroidea because it has cotton to flocculent basidiomata with a cream to buff hymenial surface and subglobose to globose basidiospores (Zong and Zhao, 2021). Xenasmatella rhizomorpha is separated from X. nigroidea by the clay-buff to cinnamon hymenial surface and the presence of the rhizomorphs (Zong et al., 2021).
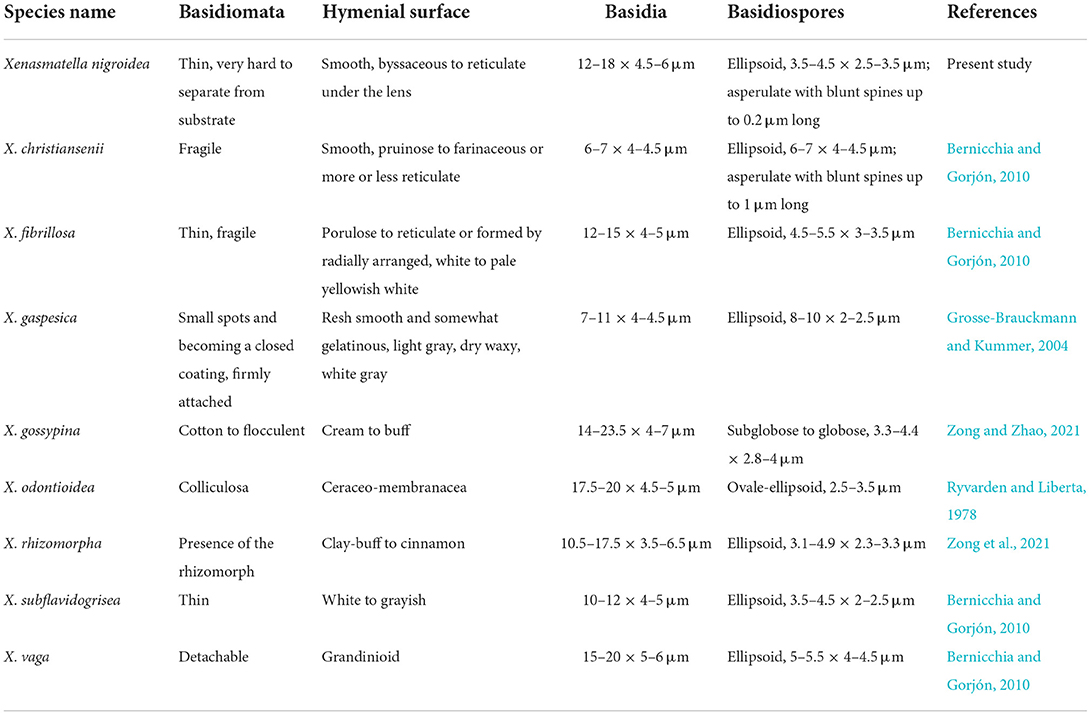
Table 3. Morphological characteristic comparison of Xenasmatella nigroidea and other species.
Xenasmatella nigroidea is similar to X. gaspesica (Liberta) Hjortstam, X. odontioidea Ryvarden & Liberta, X. subflavidogrisea (Litsch.) Oberw. ex Jülich, and X. vaga (Fr.) Stalpers due to the presence of the ellipsoid or narrowly ellipsoid basidiospores. However, X. gaspesica differs from X. nigroidea because it has smaller basidia (7–11 × 4–4.5 μm) and larger basidiospores (8–10 × 2–2.5 μm; Grosse-Brauckmann and Kummer, 2004). Xenasmatella odontioidea can be distinguished from X. nigroidea by its colliculosa hymenial surface and shorter basidiospores (2.5–3.5 μm; Ryvarden and Liberta, 1978). Xenasmatella subflavidogrisea is separated from X. nigroidea due to the presence of a white to grayish hymenial surface, turning dark reddish brown in KOH and narrower basidiospores (2–2.5 μm; Bernicchia and Gorjón, 2010). Xenasmatella vaga differs from X. nigroidea due to its grandinioid hymenial surface and larger basidiospores (5–5.5 × 4–4.5 μm; Bernicchia and Gorjón, 2010).
Based on the geographical distribution in America, Asia, and Europe, and ecological habits, white-rot causing Xenasmataceae have been reported in angiosperms and gymnosperms (Figure 6 and Table 4) (Stalpers, 1996; Dai et al., 2004; Hjortstam and Ryvarden, 2005; Bernicchia and Gorjón, 2010; Duhem, 2010; Dai, 2011; Huang et al., 2019; Larsson et al., 2020; Maekawa, 2021; Zong and Zhao, 2021; Zong et al., 2021). Key to 25 accepted species of Xenasmatella worldwide in Table 5. Many wood-decaying fungi have been recently reported worldwide (Zhu et al., 2019; Angelini et al., 2020; Gafforov et al., 2020; Zhao and Zhao, 2021). According to the results of our study on Xenasmatella, all these fungi can be classified into a new taxon (Figure 3). In addition, this study contributes to the knowledge of the fungal diversity in Asia.
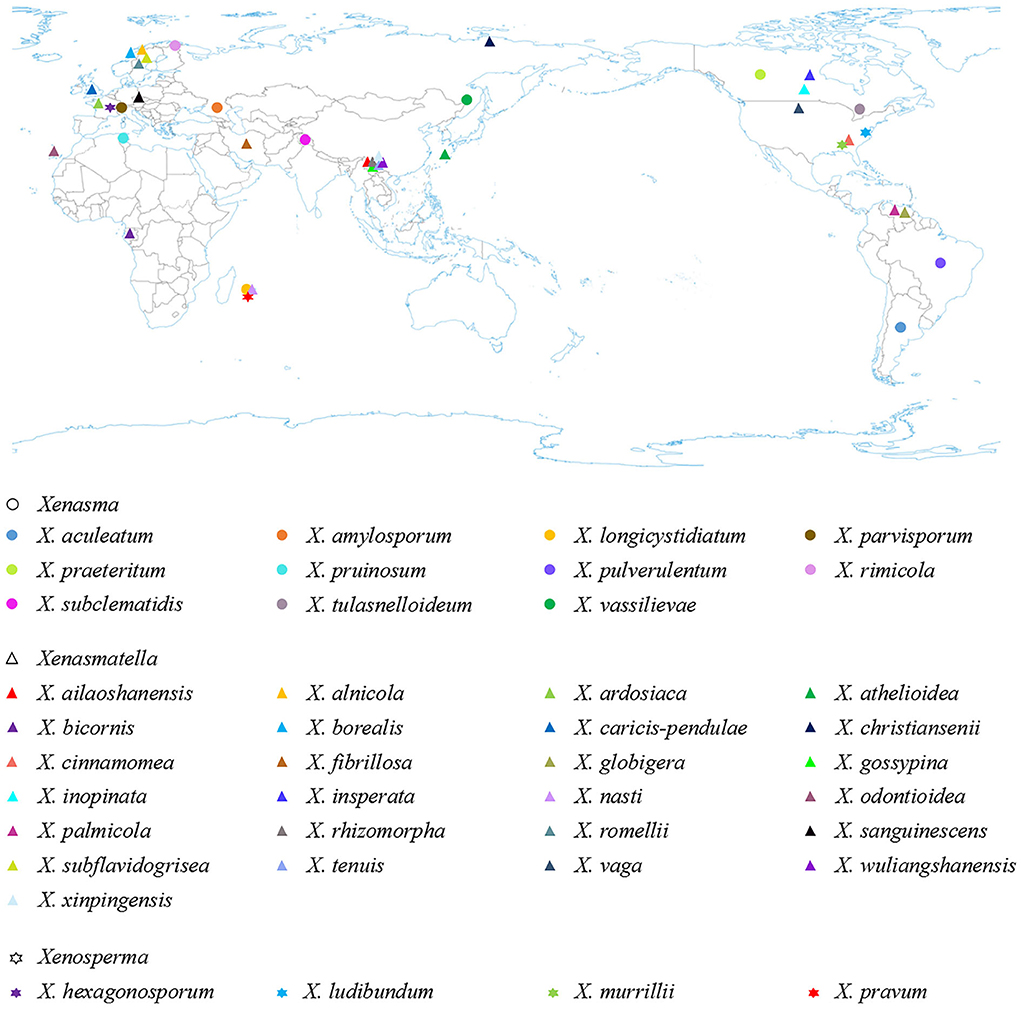
Figure 6. The geographic distribution of Xenasmataceae species (holotype) worldwide.
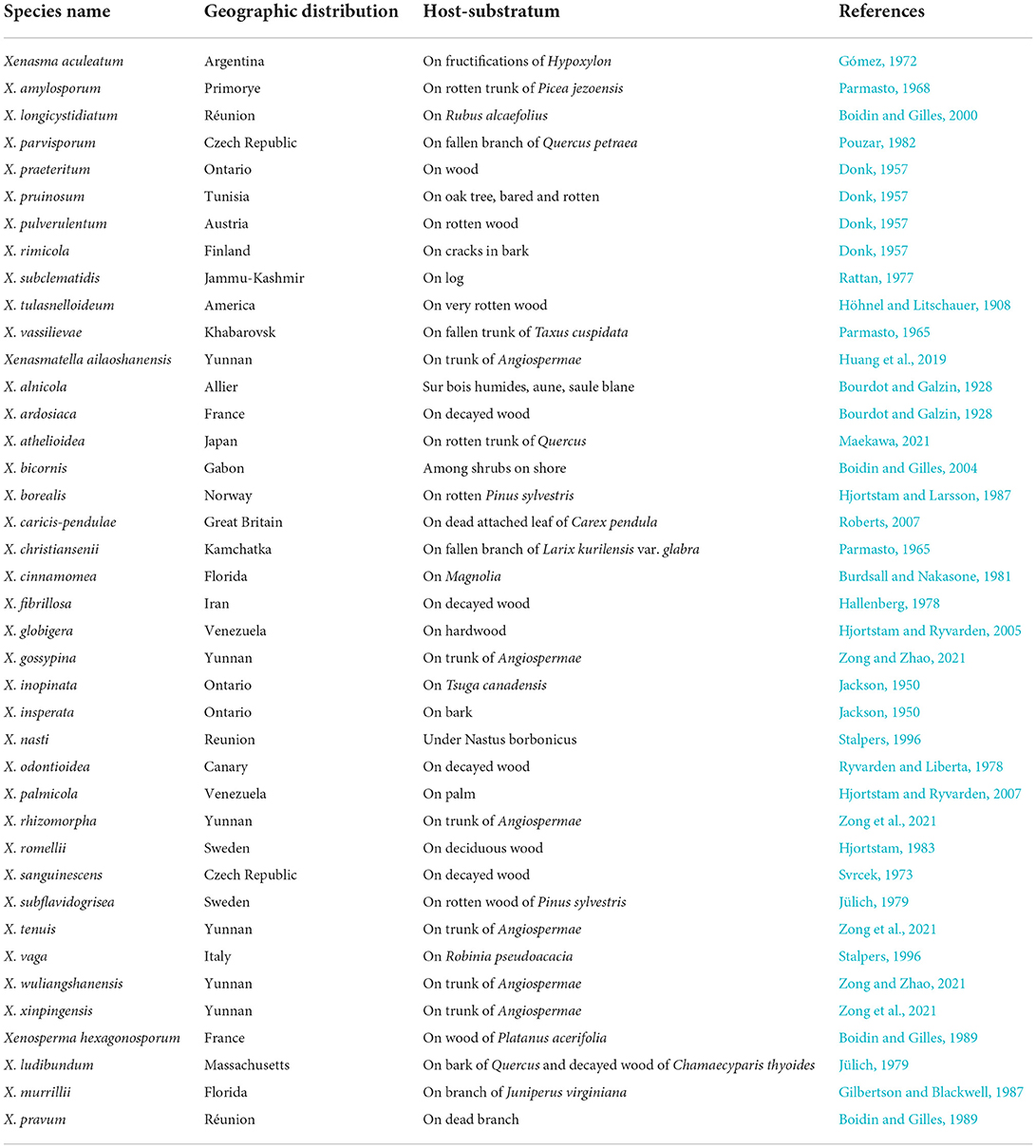
Table 4. The geographic distribution and host-substratum of Xenasmataceae species (holotype).

Table 5. Key to 25 accepted species of Xenasmatella worldwide.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Author contributions
C-LZ: conceptualization, resources, supervision, project administration, and funding acquisition. C-LZ and K-YL: methodology, software, validation, formal analysis, investigation, writing—original draft preparation, writing—review and editing, and visualization. Both authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the National Natural Science Foundation of China (Project No. 32170004, U2102220) to C-LZ, the Yunnan Fundamental Research Project (Grant No. 202001AS070043) to C-LZ, the High-level Talents Program of Yunnan Province (YNQR-QNRC-2018-111) to C-LZ, and the Yunnan Key Laboratory of Plateau Wetland Conservation, Restoration, and Ecological Services (202105AG070002) to K-YL.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Angelini, C., Vizzini, A., Justo, A., Bizzi, A., and Kaya, E. (2020). First report of a neotropical agaric (lepiota spiculata, agaricales, basidiomycota) containing lethal α-amanitin at toxicologically relevant levels. Front. Microbiol. 11, 1833. doi: 10.3389/fmicb.2020.01833
Bernicchia, A., and Gorjón, S. P. (2010). Fungi Europaei 12: Corticiaceae s.l. Alassio: Edizioni Candusso.
Besl, H., and Bresinsky, A. (1997). Chemosystematics of suillaceae and gomphidiaceae (suborder Suillineae). Plant Syst. Evol. 206, 223–242. doi: 10.1007/BF00987949
Boidin, J., and Gilles, G. (1989). Les Corticiés pleurobasidiés (Basidiomycotina) en France. Cryptogamic Bot. 1, 70–79.
Boidin, J., and Gilles, G. (2000). Basidiomycètes Aphyllophorales de l'ile de La Reunion. XXI - Suite. Mycotaxon 75, 357–387.
Boidin, J., and Gilles, G. (2004). Homobasidiomycètes Aphyllophorales non porés à basides dominantes à 2 (3) stérigmates. Bull. Trimest. Soc. Mycol. Fr. 119, 1–17.
Bourdot, H., and Galzin, A. (1928). Hyménomycètes de France : Hétérobasidiés. Homobasidiés gymnocarpes / par MM. l'abbé H. Bourdot et A. Galzin. Paris: Bibliothèque nationale de France, département Sciences et techniques, 786.
Burdsall, H. H., and Nakasone, K. K. (1981). New or little known lignicolous aphyllophorales (Basidiomycotina) from southeastern United States. Mycologia 73, 454–476. doi: 10.1080/00275514.1981.12021368
Chen, C. C., Chen, C. Y., and Wu, S. H. (2021). Species diversity, taxonomy and multi-gene phylogeny of phlebioid clade (Phanerochaetaceae, Irpicaceae, Meruliaceae) of polyporales. Fungal Divers. 111, 1–106. doi: 10.1007/s13225-021-00490-w
Cui, B. K., Du, P., and Dai, Y. C. (2011). Three new species of Inonotus (Basidiomycota, Hymenochaetaceae) from China. Mycol. Prog. 10, 107–114. doi: 10.1007/s11557-010-0681-6
Dai, Y. C. (2011). A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 52, 69–79. doi: 10.1007/S10267-010-0068-1
Dai, Y. C. (2012). Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 53, 49–80. doi: 10.1007/s10267-011-0134-3
Dai, Y. C., Cui, B. K., Si, J., He, S. H., Hyde, K. D., Yuan, H. S., et al. (2015). Dynamics of the worldwide number of fungi with emphasis on fungal diversity in China. Mycol. Prog. 14, 62. doi: 10.1007/s11557-015-1084-5
Dai, Y. C., Wei, Y. L., and Zhang, X. Q. (2004). An annotated checklist of non-poroid Aphyllophorales in China. Ann. Bot. Fennici 41, 233–247.
Dai, Y. C., Yang, Z. L., Cui, B. K., Wu, G., Yuan, H. S., Zhou, L. W., et al. (2021). Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 40, 770–805.
Duhem, B. (2010). Deux corticiés nouveaux méditerranéens à spores allantoïdes. Cryptogam. Mycol. 31, 143–152.
Eriksson, J., Hjortstam, K., and Ryvarden, L. (1978). Corticiaceae of North Europe Volume 5: Mycoaciella-Phanerochaete. Oslo: Fungiflora.
Eriksson, J., Hjortstam, K., and Ryvarden, L. (1981). Corticiaceae of North Europe Volume 6: Phlebia-Sarcodontia. Oslo: Fungiflora.
Eriksson, J., Hjortstam, K., and Ryvarden, L. (1984). Corticiaceae of North Europe Volume 7: Schizopora-Suillosporium. Oslo: Fungiflora.
Felsenstein, J. (1985). Confidence intervals on phylogenetics: an approach using bootstrap. Evolution 39, 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Fries, E. (1821–1832). Systema Mycologicum, Sistens Fungorum Ordines, Generaet Species Hucusque Cognitas. Gryphiswaldiae: Ernestus Mauritius.
Fries, E. (1857–1863). Monographia Hymenomycetum Sueciae. Vols. I and II. Leffler, C. A. Uppsala: Nabu Press.
Gafforov, Y., Ordynets, A., Langer, E., Yarasheva, M., de Mello Gugliotta, A., Schigel, D., et al. (2020). Species diversity with comprehensive annotations of wood-inhabiting poroid and corticioid fungi in Uzbekistan. Front. Microbiol. 11, 598321. doi: 10.3389/fmicb.2020.598321
Garcia-Sandoval, R., Wang, Z., Binder, M., and Hibbett, D. S. (2010). Molecular phylogenetics of the Gloeophyllales and relative ages of clades of Agaricomycotina producing a brown rot. Mycologia 103, 510–524. doi: 10.3852/10-209
Gilbertson, R. L., and Blackwell, M. (1987). Notes on wood-rotting fungi on Junipers in the Gulf Coast region. II. Mycotaxon 28, 369–402.
Gilbertson, R. L., and Ryvarden, L. (1987). North American Polypores 1-2. Fungiflora; Oslo: Lubrecht and Cramer Ltd. p. 1–433.
Gómez, C. E. (1972). Xenasma y géneros afines de los alrededores de Buenos Aires (Aphyllophorales). Bol Soc Argent Bot 14, 269–281.
Gonzalez-Avila, A., Contreras-Medina, R., Espinosa, D., and Luna-Vega, I. (2017). Track analysis of the order Gomphales (Fungi: Basidiomycota) in Mexico. Phytotaxa 316, 22–38. doi: 10.11646/phytotaxa.316.1.2
Grosse-Brauckmann, H., and Kummer, V. (2004). Fünf bemerkenswerte Funde corticioider Pilze aus Deutschland. Feddes Repert. 115, 90–101. doi: 10.1002/fedr.200311029
Haelewaters, D., Dirks, A. C., Kappler, L. A., Mitchell, J. K., Quijada, L., Vandegrift, R., et al. (2018). A preliminary checklist of fungi at the Boston Harbor Islands. Northeast. Nat. 25, 45–76. doi: 10.1656/045.025.s904
Hallenberg, N. (1978). Wood-Fungi (Corticiaceae, Coniophoraceae, Lachnocladiaceae, Thelephoraceae) in N. Iran. I. Iran. J. Plant Pathol. 14, 38–87.
He, M. Q., Zhao, R. L., Hyde, K. D., Begerow, D., Kemler, M., Yurkov, A., et al. (2019). Notes, outline and divergence times of Basidiomycota. Fungal Divers. 99, 105–367. doi: 10.1007/s13225-019-00435-4
Hibbett, D. S., Binder, M., Bischoff, J. F., Blackwell, M., Cannon, P. F., Eriksson, O. E., et al. (2007). A higher-level phylogenetic classification of the Fungi. Mycol. Res. 111, 509–547. doi: 10.1016/j.mycres.2007.03.004
Hjortstam, K., and Larsson, K. H. (1987). Additions to Phlebiella (Corticiaceae, Basidiomycetes), with notes on Xenasma and Sistotrema. Mycotaxon 29, 315–319.
Hjortstam, K., and Ryvarden, L. (2005). New taxa and new combinations in tropical corticioid fungi, (Basidiomycotina, Aphyllophorales). Synop. Fungorum 20, 33–41.
Hjortstam, K., and Ryvarden, L. (2007). Studies in corticioid fungi from Venezuela III (Basidiomycotina, Aphyllophorales). Synop. Fungorum 23, 56–107.
Höhnel, F., and Litschauer, V. (1908). Beiträge zur Kenntnis der Corticieen: III. Sitz. K. Akad. Wiss. Math. Naturw. Klasse Abt. I. 117, 1081–1124.
Huang, R. X., Chen, J. Z., Wu, J. R., and Zhao, C. L. (2019). Phlebiella ailaoshanensis sp. nov. (Polyporales, Basidiomycota) described from China. Phytotaxa 419, 105–109. doi: 10.11646/phytotaxa.419.1.8
Jackson, H. S. (1950). Studies of Canadian Thelephoraceae. VII. Some new species of Corticium, section Athele. Can. J. Res. 28, 716–725. doi: 10.1139/cjr50c-045
James, T. Y., Stajich, J. E., Hittinger, C. T., and Rokas, A. (2020). Toward a fully resolved fungal tree of life. Annu. Rev. Microbiol. 74, 291–313. doi: 10.1146/annurev-micro-022020-051835
Jarosch, M. (2001). Zur molekularen systematik der boletales: coniophorineae, paxillineae und suillineae. Bibl. Mycol. 191, 1–158.
Jarosch, M., and Besl, H. (2001). Leucogyrophana, a polyphyletic genus of the order boletales (Basidiomycetes). Plant Biol. 3, 443–448. doi: 10.1055/s-2001-16455
Jülich, W. (1979). Studies in resupinate Basidiomycetes - V. On some new taxa. Persoonia 10, 325–336.
Kirk, P. M., Cannon, P. F., David, J. C., Minter, D. W., and Stalpers, J. A. (2008). Ainsworth and bisby's dictionary of the fungi. 10th ed. Wallingford, Oxon, UK: CAB International Press, 783.
Larsson, A. (2014). AliView: a fast and lightweight alignment viewer and editor for large data sets. Bioinformatics 30, 3276–3278. doi: 10.1093/bioinformatics/btu531
Larsson, E., and Hallenberg, N. (2001). Species delimitation in the Gloeocystidiellum Porosum-Clavuligerum complex inferred from compatibility studies and nuclear RDNA sequence data. Mycologia 93, 907–914. doi: 10.1080/00275514.2001.12063225
Larsson, E., and Larsson, K. H. (2003). Phylogenetic relationships of Russuloid Basidiomycetes with emphasis on Aphyllophoralean taxa. Mycologia 95, 1037–1065. doi: 10.1080/15572536.2004.11833020
Larsson, K. H. (2007). Re-thinking the classification of corticioid fungi. Mycol. Res. 111, 1040–1063. doi: 10.1016/j.mycres.2007.08.001
Larsson, K. H., Larsson, E., and Koljalg, U. (2004). High phylogenetic diversity among corticioid homobasidiomycetes. Mycol. Res. 108, 983–1002. doi: 10.1017/S0953756204000851
Larsson, K. H., Larsson, E., Ryvarden, L., and Spirin, V. (2020). Some new combinations of corticioid fungi (Basidiomycota. Agaricomycetes). Synop. Fungorum 40, 113–117.
Larsson, K. H., Parmasto, E., Fischer, M., Langer, E., Nakasone, K. K., and Redhead, S. A. (2006). Hymenochaetales: a molecular phylogeny for the hymenochaetoid clade. Mycologia 98, 926–936. doi: 10.1080/15572536.2006.11832622
Maekawa, N. (2021). Taxonomy of corticioid fungi in Japan: present status and future prospects. Mycoscience 62, 345–355. doi: 10.47371/mycosci.2021.10.002
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2012). The CIPRES science gateway: enabling high-impact science for phylogenetics researchers with limited resources. Assoc. Comput. Mach. 39, 1–8. doi: 10.1145/2335755.2335836
Nylander, J. A. A. (2004). MrModeltest v2. Program Distributed by the Author. Uppsala: Evolutionary Biology Centre; Uppsala Univeristy.
Oberwinkler, F. (1966). Primitive Basidiomyceten. Revision einiger Formenkreise von Basidienpilzen mit plastischer Basidie. Sydowia 19, 1–72.
Parmasto, E. (1965). Corticiaceae U.R.S.S. I. Descriptiones taxorum novarum. Combinationes novae. Eesti NSV Tead. Akad. TOIM. 14, 220–233. doi: 10.3176/biol.1965.2.06
Petersen, J. H. (1996). Farvekort. The Danish Mycological Society's Colour-Chart. Greve: Foreningen til Svampekundskabens Fremme.
Piatek, M. (2005). A note on the genus Xenosmatella (Fungi, Basidiomycetes). Polish Bot. J. 50, 11–13.
Rattan, S. S. (1977). The resupinate Aphyllophorales of the North Western Himalayas. Bibl. Mycol. 60, 1–427.
Rayner, R. W. (1970). A Mycological Colour Chart. Commonwealth Mycological Institute, Kew and British Mycological Society, Kew, United Kingdom. p. 1–34.
Roberts, P. J. (2007). Phlebiella caricis-pendulae: A new corticoid fungus from Wales. Synop. Fungorum 22, 25–26.
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Hohna, S., et al. (2012). Mrbayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rosenthal, L. M., Larsson, K. H., Branco, S., Chung, J. A., Glassman, S. I., Liao, H. L., et al. (2017). Survey of corticioid fungi in North American pinaceous forests reveals hyperdi-versity, underpopulated sequence databases, and species that are potentially ectomycorrhizal. Mycologia 109, 115–127. doi: 10.1080/00275514.2017.1281677
Ryvarden, L., and Liberta, A. E. (1978). Contribution to the Aphyllophoralles of the Canary Islands 4. Two new species of Trechispora and Xenmastella. Can. J. Bot. 56, 2617–2619. doi: 10.1139/b78-314
Sotome, K., Hattori, T., and Ota, Y. (2011). Taxonomic study on a threatened polypore, Polyporus pseudobetulinus, and a morphologically similar species, P. subvarius. Mycoscience 52, 319–326. doi: 10.1007/S10267-011-0111-X
Stalpers, J. A. (1996). The aphyllophoraceous fungi II. Keys to the species of the Hericiales. Stud. Mycol. 40, 1–185.
Swofford, D. L. (2002). PAUP*: Phylogenetic Analysis Using Parsimony (*and other methods). Version 4.0b10. Sunderland, Massachusetts: Sinauer Associates.
Telleria, M. T., Melo, I., Duenas, M., Larsson, K. H., and Paz Martin, M. P. (2013). Molecular analyses confirm Brevicellicium in Trechisporales. IMA Fungus 4, 21–28. doi: 10.5598/imafungus.2013.04.01.03
Tobias, W., and Michael, F. (2002). Classification and phylogenetic relationships of Hymenochaete and allied genera of the Hymenochaetales, inferred from rDNA sequence data and nuclear behaviour of vegetative mycelium. Mycol. Prog. 1, 93–104. doi: 10.1007/s11557-006-0008-9
Vu, D., Groenewald, M., de Vries, M., Gehrmann, T., Stielow, B., Eberhardt, U., et al. (2019). Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 92, 135–154. doi: 10.1016/j.simyco.2018.05.001
Wagner, T., and Fischer, M. (2001). Natural groups and a revised system for the European poroid Hymenochaetales (Basidiomycota) supported by nLSU rDNA sequence data. Mycol. Res. 105, 773–782. doi: 10.1017/S0953756201004257
White, T. J., Bruns, T., Lee, S., and Taylor, J. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR protocols: A Guide to Methods And Applications, eds Innis, M. A., Gelfand, D. H., Sninsky, J. J., and White, T. J. (San Diego, CA: Academic Press), 315–322.
Wu, F., Yuan, H. S., Zhou, L. W., Yuan, Y., Cui, B. K., and Dai, Y. C. (2020). Polypore diversity in South China. Mycosystema 39, 653–682.
Wu, F., Yuan, Y., and Zhao, C. L. (2015). Porpomyces submucidus (Hydnodontaceae, Basidiomycota), a new species from tropical China based on morphological and molecular evidence. Phytotaxa 230, 61–68. doi: 10.11646/phytotaxa.230.1.5
Wu, F., Zhou, L. W., Vlasák, J., and Dai, Y. C. (2022). Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 113, 1–192. doi: 10.1007/s13225-021-00496-4
Zhao, C. L., and Wu, Z. Q. (2017). Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 16, 93–100. doi: 10.1007/s11557-016-1259-8
Zhao, W., and Zhao, C. L. (2021). The phylogenetic relationship revealed three new wood-inhabiting fungal species from genus Trechispora. Front. Microbiol. 12, 650195. doi: 10.3389/fmicb.2021.650195
Zhu, L., Song, J., Zhou, J. L., Si, J., and Cui, B. K. (2019). Species diversity, phylogeny, divergence time and biogeography of the genus Sanghuangporus (Basidiomycota). Front. Microbiol. 10, 812. doi: 10.3389/fmicb.2019.00812
Zong, T. K., Wu, J. R., and Zhao, C. L. (2021). Three new Xenasmatella (Polyporales, Basidiomycota) species from China. Phytotaxa 489, 111–120. doi: 10.11646/phytotaxa.489.2.1
Keywords: biodiversity, fungal systematics, ITS, LSU, new taxa, wood-decaying fungi, Xenasmatales, Xenasmatella nigroidea
Citation: Luo K-Y and Zhao C-L (2022) Morphology and multigene phylogeny reveal a new order and a new species of wood-inhabiting basidiomycete fungi (Agaricomycetes). Front. Microbiol. 13:970731. doi: 10.3389/fmicb.2022.970731
Received: 16 June 2022; Accepted: 08 August 2022;
Published: 31 August 2022.
Edited by:
Samantha Chandranath Karunarathna, Qujing Normal University, ChinaReviewed by:
Yusufjon Gafforov, Academy of Science of the Republic of Uzbekistan, UzbekistanKalani Hapuarachchi, Guizhou University, China
Copyright © 2022 Luo and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chang-Lin Zhao, ZnVuZ2lAc3dmdS5lZHUuY24=