 Ling Xin
Ling Xin Xiaogang Xu
Xiaogang Xu Qingyu Shi1,2
Qingyu Shi1,2 Fupin Hu
Fupin Hu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 08 July 2022
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.900185
This article is part of the Research Topic Horizontal Gene Transfer Mediated Bacterial Antibiotic Resistance, Vol II View all 11 articles
Enterococci are one of the main causes of gastrointestinal tract infections in the healthcare system and can develop resistance to fosfomycin through plasmid or chromosomally encoded fosfomycin resistance genes. To investigate the mechanisms of fosfomycin resistance, a total of 4,414 clinical isolates of non-replicated clinical enterococci collected from 62 hospitals in 26 provinces or cities in China were tested. Antibiotic susceptibility testing, detection of fosfomycin resistance genes, and cloning of the fosX gene were done. The PFGE, MLST, qRT-PCR, and next genome sequencing were carried out. The results revealed that the fosfomycin-resistant rate of enterococci was 3.5% (153/4,414), and the major resistance mechanism was fosX (101/153) and fosB (52/153) genes. The fosX gene could increase 4- fold fosfomycin MIC in Enterococcus faecium BM4105RF transformants, and the results of PFGE showed the 101 E. faecium carrying fosX were grouped into 48 pulse types. The multilocus sequence typing identified ST555 as the vast majority of STs, mostly distributed in Shanghai, China. Furthermore, the fosX gene expression was strongly related to the fosfomycin-resistant levels of enterococci. The present study was the first to describe the high prevalence presence of the fosX gene in E. faecium from China.
Enterococci are significant opportunistic nosocomial pathogens of the gastrointestinal tract, occupying second place in the detection rate of Gram-positive bacteria (Fillgrove et al., 2003). Currently, clinically limited antibiotics are available for the treatment of enterococcal infections (Scortti et al., 2018). As a phosphoric acid antibiotic agent, fosfomycin showed broad-spectrum antibacterial activity and acted as the first step in the synthesis of bacterial peptidoglycan (Fillgrove et al., 2003; Chen et al., 2019; Zhang et al., 2020). With the widespread use of fosfomycin, the emergence of enterococci has become a major concern (Fillgrove et al., 2003; Scortti et al., 2018).
Fosfomycin resistance mechanisms have been proposed, including inherent the acquisition of chromosomal mutations and plasmid-encoded fosfomycin-modifying enzymes. Regarding the mechanisms of chromosomal mutations, it supported the replacement position of cysteine in the active site of the UDP-N-acetylglucosamine-3-enolpruvyltransferase (MurA), such as Borrelia burgdorferi, Mycobacterium tuberculosis, etc. (Fillgrove et al., 2003; Clinical and Laboratory Standards Institute, 2020). Moreover, fosfomycin resistance is also mediated by reduced antimicrobial uptake by chromosomal mutations in the glpT and uhpT genes (Clinical and Laboratory Standards Institute, 2020). Different fosfomycin-modifying enzymes had been described, including metalloenzymes (FosA, FosB, and FosX) (Murray et al., 1990; Bernat et al., 1997; Xu et al., 2013; Chen et al., 2014) and fosfomycin kinases (FomA, FomB, and FosC) (Mendoza et al., 1980; Bernat et al., 1997; Sahni et al., 2013).
Fillgrove et al. (2003) found a related subfamily enzyme named FosX with 30–35% similarity to the sequences of FosA and FosB proteins in the microbial genome sequence database. Scortti et al. (2018) reported that the FosX-mediated resistance is epistatically suppressed by two members of the PrfA virulence regulon, hpt and prfA, which upon activation by host signals induce increased fosfomycin influx into the bacteria cell. Chen et al. (2019) found that the aminoglycosides resistance gene rmtB may be co-disseminated with blaKPC–2 and fosA3 genes through plasmid, which results in fosfomycin-resistance isolates difficult-to-treat pathogen due to limited treatment options. Zhang et al. (2020) reported that the coexistence of van series genes and fos genes simultaneously can greatly improve the transfer efficiency in Enterococcus faecium.
At present, most of the previous studies investigated the mechanism of fosfomycin resistance among Gram-negative bacteria, and only limited information about the resistance mechanism of Gram-positive, particularly enterococci, is available. In the published articles, fosA, fosC, and fosX genes were detected in gram-negative bacteria, while fosB genes were detected in gram-positive bacteria. In this study, we collected 4,414 clinical isolates of enterococci collected from 62 hospitals in 26 provinces or cities across China from 2017 to 2020 and aimed to survey the prevalence of fosfomycin resistance and the associated fosB, fosX, and murA genetics in clinical isolates of enterococci in China.
The bacteria in this research were collected from 62 hospitals in 26 provinces or cities in China from 2017 to 2020. All 4,414 non-duplicated enterococci strains were collected by the CHINET surveillance system, including 2,316 Enterococcus faecium strains and 2,098 Enterococcus faecalis isolates, stored in 40% glycerol broth and frozen at −80°C. The hospitals in the CHINET surveillance system collect bacteria every year, and the staff numbers in hospitals randomly collect bacteria in the process. The distribution of 4,414 isolates was 1,222 in 2017, 1,411 in 2018, 1,277 in 2019, and 504 in 2020. All strains isolated from children and adults accounted for 1.6 and 98.4%, respectively. Among them, the main sources of specimens were urine (64.4%), bile (12.2%), pleural and ascites (12.2%), blood (8.5%), wounds (6.6%), and respiratory tract specimens (5.3%). E. faecalis ATCC 29212 was used as a quality control strain.
The antimicrobial susceptibility to fosfomycin was determined using the agar dilution method by Mueller-Hinton agar (MHA) supplemented with 25 mg/L glucose-6-phosphate (G6P) and the results were interpreted by the Clinical and Laboratory Standards Institute (CLSI) M100 31th Edition guideline. To interpret the susceptibility of E. faecium to fosfomycin, values corresponded to the breakpoints of the urinary tract infection E. faecalis in CLSI (sensitivity ≤ 64 mg/L; resistant ≥ 256 mg/L) (Clinical and Laboratory Standards Institute, 2020).
The fosfomycin resistance genes were detected and confirmed by polymerase chain reaction (PCR) using the primers listed in Table 1. The PCR products were sequenced by the MAP BIOTECH company (Shanghai, China) and the sequences were aligned by BLAST1.
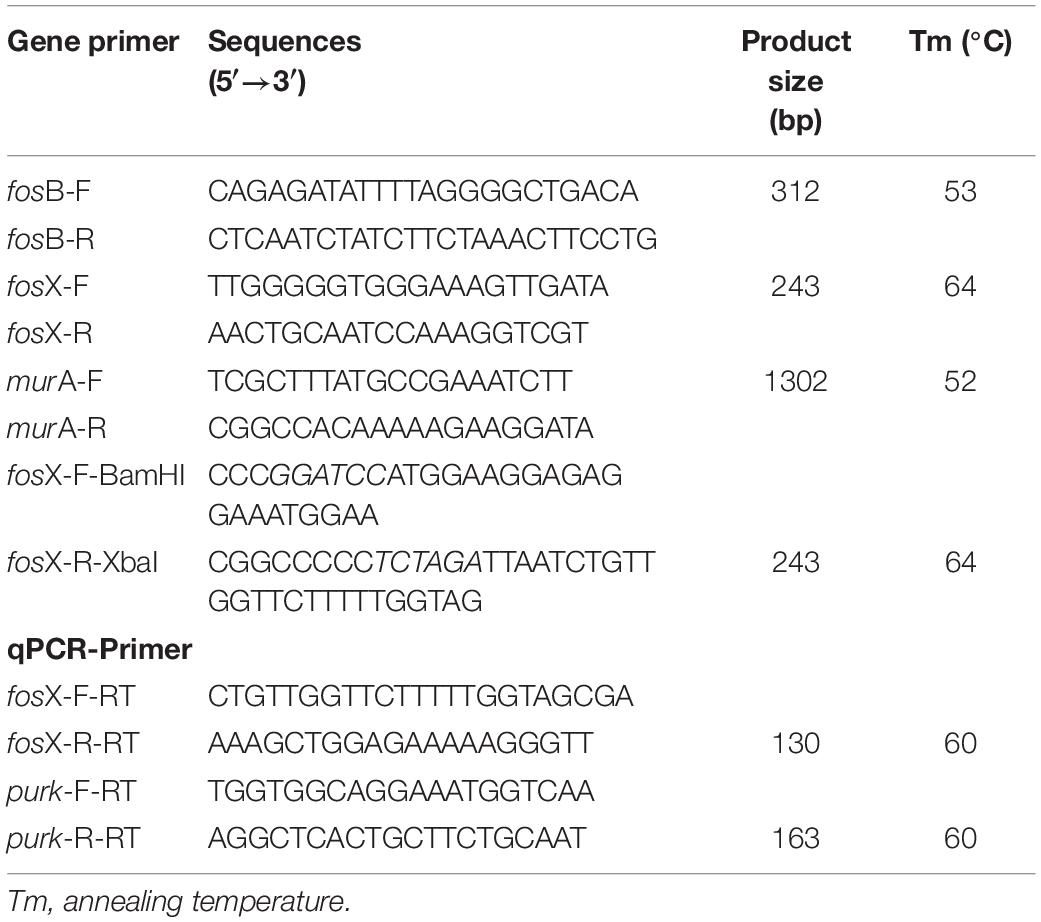
Table 1. PCR primers of genes.
To further elucidate the function of fosX, we selected the fosX fragment from a fosfomycin-resistant E. faecium (MIC = 512 mg/L) for cloning and cloned it into the shuttle vector plasmid pIB166. The recombinant plasmid pIB166_fosX was transformed into Escherichia coli DH5α competent cells, selected with chloramphenicol (5 mg/L). Later, the resulting plasmids were electroporated from E. coli DH5α into E. faecium BM4105RF competent cells, selected with rifampin (50 mg/L) and chloramphenicol (5 mg/L). The presence of fosX in E. coli and E. faecium transformants were confirmed by PCR and sequenced by the MAP BIOTECH company (Shanghai, China).
A pulsed-field gel electrophoresis analysis was performed using a CHEF mapper system (Bio-Rad, United States) (Murray et al., 1990). Agarose gel blocks were lysed with proteinase K at 20 mg/ml and digested with SmaI. The digested DNA was subjected to electrophoresis at 6 V/cm, 14°C, in a 1.0% agarose gel with pulse times of 1 to 20 s for 20 h. The banding patterns were interpreted using the criteria devised by Tenover et al. (Chen et al., 2014).
Genomic DNAs of enterococci strains were subjected to whole-genome sequencing using Illumina (Illumina, San Diego, CA, United States) short-read sequencing (150 bp paired-end reads). To ensure the quality of information analysis, the raw data is filtered using FASTX-Toolkit software2, assembled by the velvet V1.2.03 software3, and used glimmer software4 to predict genes. The functional annotation of genes was confirmed by searching the NCBI’s nr library5. Isolates were screened using the following seven housekeeping genes: adenylate kinase (adk), ATP synthase-alpha subunit (atpA), D-alanine ligase (ddl), glyceraldehyde-3-phosphate dehydrogenase (gyd), glucose-6-phosphate dehydrogenase (gdh), phosphoribosylaminoimidazole carboxylase ATPase subunit (purK), and phosphate ATP-binding cassette transporter (pstS). The alleles and sequence types (STs) were determined via the MLST database6. The phylogenetic tree was performed by MEGA_X and iTOL7.
A single colony was selected from the blood agar plate and added to brain heart infusion broth and incubated for 18 h at 37°C with shaking at 180 rpm. The bacterial culture was centrifuged at 12,000 rpm for 5 min to collect the supernatant. Total RNA was extracted using a Life Real kit (Life Real, Hangzhou, China) according to the manufacturer’s instructions. Thereafter, the RNAs were reverse transcribed to cDNA for qRT-PCR analysis in the light of the manufacturer’s manual of a commercial cDNA synthesis kit with a gDNA eraser (Takara, Dalian, China). The qPCR was performed on a Life Tech-ViiA7 qPCR multiplex reactions System (Life Technologies, United States) with specific primers (Table 1) for the PrimeScriptTM RT reagent Kit (Takara, Dalian, China). The relative expression levels of the fosX gene were normalized to the purK reference gene. The quantification of the target genes was analyzed using the comparative threshold cycle 2–△△Ct method (Zhang et al., 2020). All experiments were repeated in triplicate independently. The MIC range of the strains carrying the fosX gene to fosfomycin was 512–4,096 mg/L. Among them, the MICs of fosfomycin of HS2166, HS570, HS2069 and HS1440 were 512 mg/L, 1,024 mg/L, 2,048 mg/L, and 4,096 mg/L. The relative expression of the fosX gene of HS570, HS2069, and HS1440 isolates was normalized to that of the HS2166 strain.
To evaluate the difference in antibiotic resistance of enterococci carrying the fosX gene with different levels of fosfomycin resistance, we extracted DNA from the HS2166, HS570, HS2069, and HS1440 strains and performed next genome sequencing according to the precious method (Xu et al., 2013). Raw sequences were filtered using FASTX-Toolkit (see text footnote 2) and assembled into contigs using the velvet V1.2.03 program. The Comprehensive Antibiotic Database8 was used to identify antibiotic resistance genes. The replicon sequences were identified by PlasmidFinder 2.19. The Virulence Factors Database10 was used to identify virulence genes.
The study protocol was approved by the Institutional Review Board of Huashan Hospital, Fudan University (no. 2020-040).
Among the 4,414 isolates, the rate of fosfomycin-resistant enterococci was 3.5% (153/4414). Fosfomycin was very active against enterococci (MIC50 and MIC90 were both 50 mg/L; the median MIC value was 50 mg/L). Of 153 fosfomycin-resistant clinical isolates, E. faecium and E. faecalis accounted for 77.1% (117/153) and 23.5% (36/153), respectively. The MIC range of fosfomycin against 153 fosfomycin-resistant strains was 512–4,096 mg/L, of which MIC50 and MIC90 were 2,048 and 4,096 mg/L, and the median MIC value was 2,048 mg/L. The PCR results showed that the strains carrying the fosB and fosX genes were 33.8% (52/153) and 65.6% (101/153) in the current study, respectively.
The fosX gene was cloned into E. faecium BM4105RF by cloning experiment, and the results revealed that the fosfomycin MICs of E. faecium BM4105RF transformant carrying fosX was 128 mg/L while the recipient and donor strains had fosfomycin MICs of 32 and 2,048 mg/L, indicating that fosX could be the expression in E. faecium. According to the results of PFGE, the fosX carrying E. faecium isolates were grouped into 48 pulse types. The top five pulse types were named ENT-27, ENT-15, ENT-29, ENT-8 and ENT-30, and the percentage were 8.9% (9/101), 5.9% (6/101), 4.9% (5/101), 4% (4/101), and 4% (4/101). The 101 fosX carrying E. faecium isolates in this study were categorized into fourteen different STs (ST555, ST6, ST17, ST38, ST78, ST 80, ST 172, ST195, ST252, ST323, ST393, ST400, ST921, and one novel ST). The top five STs were ST555, ST78, ST80, ST17, and ST252, and the percentage were 57.4% (58/101), 12.8% (13/101), 10.9% (11/101), 4% (4/101), and 3% (3/101). The minimum spanning tree analysis showed that seven groups were formed by enterococci STs by MEGA_X and iTOL analysis. The MIC of ST555, ST78, ST17, and ST252 E. faecium to Fosfomycin could reach 4,096 mg/L. 94% (95/101) of E. faecium belonged to the CC17 clonal complex, and the MIC of E. faecium to Fosfomycin can reach 4,096 mg/L. The fosfomycin MIC of 6% (6/101) of the E. faecium belonging to the non-CC17 clonal complex could reach 2,048 mg/L. In terms of geographical distribution, the distribution of 58 isolates of ST555 E. faecium was 44 in Shanghai, five strains in Beijing, two strains in Anhui, two strains in Hubei, two strains in Tianjin, one strain in Henan, one strain in Gansu, and one strain in Jiangsu province. Four ST17 E. faecium isolates were only distributed in Shanghai, and other types were scattered. The results of PFGE, MLST, and phylogenetic tree were summarized in supplement material and Figure 1.
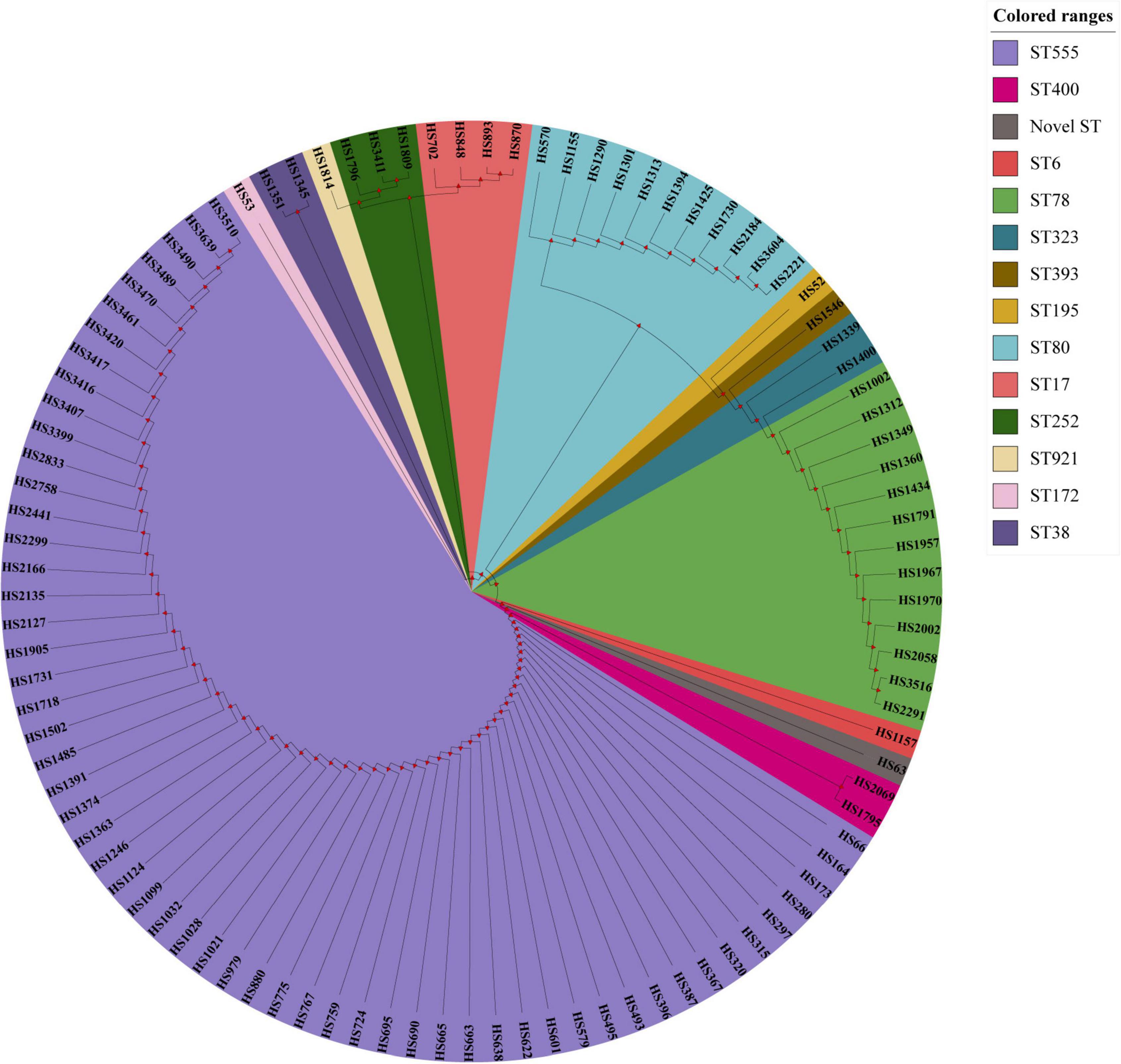
Figure 1. The MLST results of 101 E. faecium carrying fosX gene.
Due to the presence of fosX in E. faecium with different fosfomycin MICs, we determined the relationship of fosX expression levels of different fosfomycin MICs of fosX carrying fosfomycin-resistant E. faecium. The fosfomycin MIC of four E. faecium, named HS2166, HS570, HS2069, and HS1440 were 512, 1,024, 2,048, and 4,096 mg/L. Interestingly, fosX expression of fosfomycin-resistant strain (HS2166, MIC = 512 mg/L) was lower than that of HS570, HS2069, and HS1440 isolates which showed fosfomycin MICs 1,024, 2,048, and 4,096 mg/L, and the fosX relative expression of HS570 and HS2069 strains showed no significant difference (Figure 2). Overall, the qPCR results showed that with the enhancement of the resistance to fosfomycin of E. faecium, the relative expression level of the fosX gene increased correspondingly.
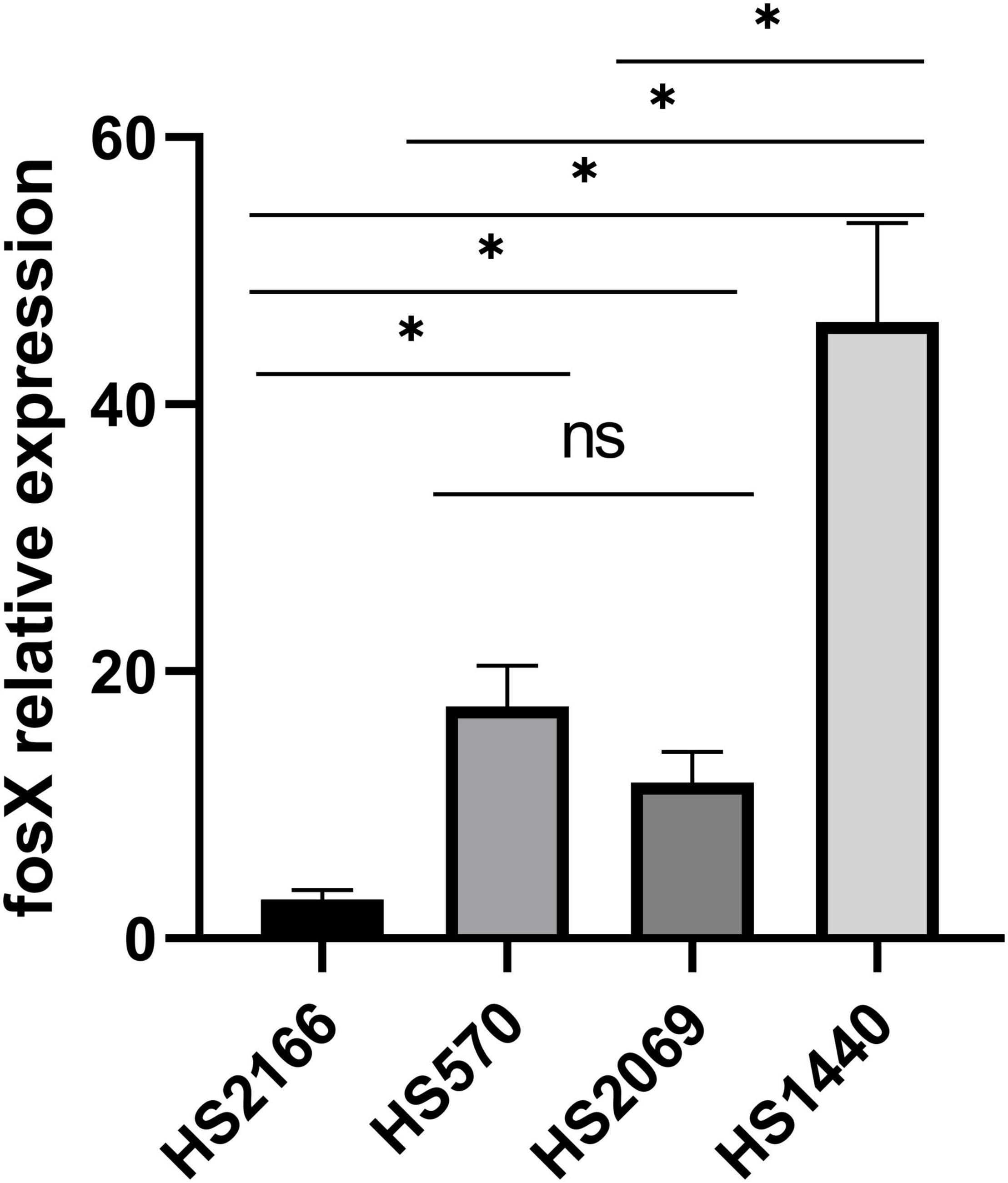
Figure 2. The relative expression of fosX gene in HS2166, HS570, HS2069, and HS1440.
The four next genome sequencing results of HS2166, HS570, HS2069, and HS1440 strains were listed in Figure 3 and Table 2. The antibiotic resistance genes of the HS2166 strain with fosfomycin MIC of 512 mg/L were aac(6’)-Ii, ermB, dfrG, dfrF, inuA, efmA, and fosX genes. There were aac(6’)-Ii, aph(3’)-IIIa, efmA, ermB, tet(45), tetU, and fosX genes in the HS570 strain with fosfomycin MIC of 1,024 mg/L. The drug resistance genes in the HS2069 strain with fosfomycin MIC of 2,048 mg/L were aac(6’)-Ii, bacA, isa, ermB, and fosX genes. The resistance genes in HS1440 strain with fosfomycin resistance level of 4,096 mg/L were aac(6’)-Ii, ant(6)-Ia, efmA, dfrG, ermB, ermT, tet(45), inuA, and fosX genes. The repUS15 and rep2 plasmids were present in all four strains, and the repIIa plasmid was present in HS2166, HS570, and HS2069. The plasmids in HS1400 isolate with fosfomycin MIC of 4,096 mg/L were repUS12, repUS15, repUS43, rep2, and rep29, and the virulence genes in the HS1400 strain were hasB, hasC, cap8D, and kfiD. Interestingly, no virulence gene was detected in the HS2166 strain with fosfomycin MIC of 512 mg/L. Only one virulence gene named bopD was detected in HS570 isolate with fosfomycin MIC of 1,024 mg/L.
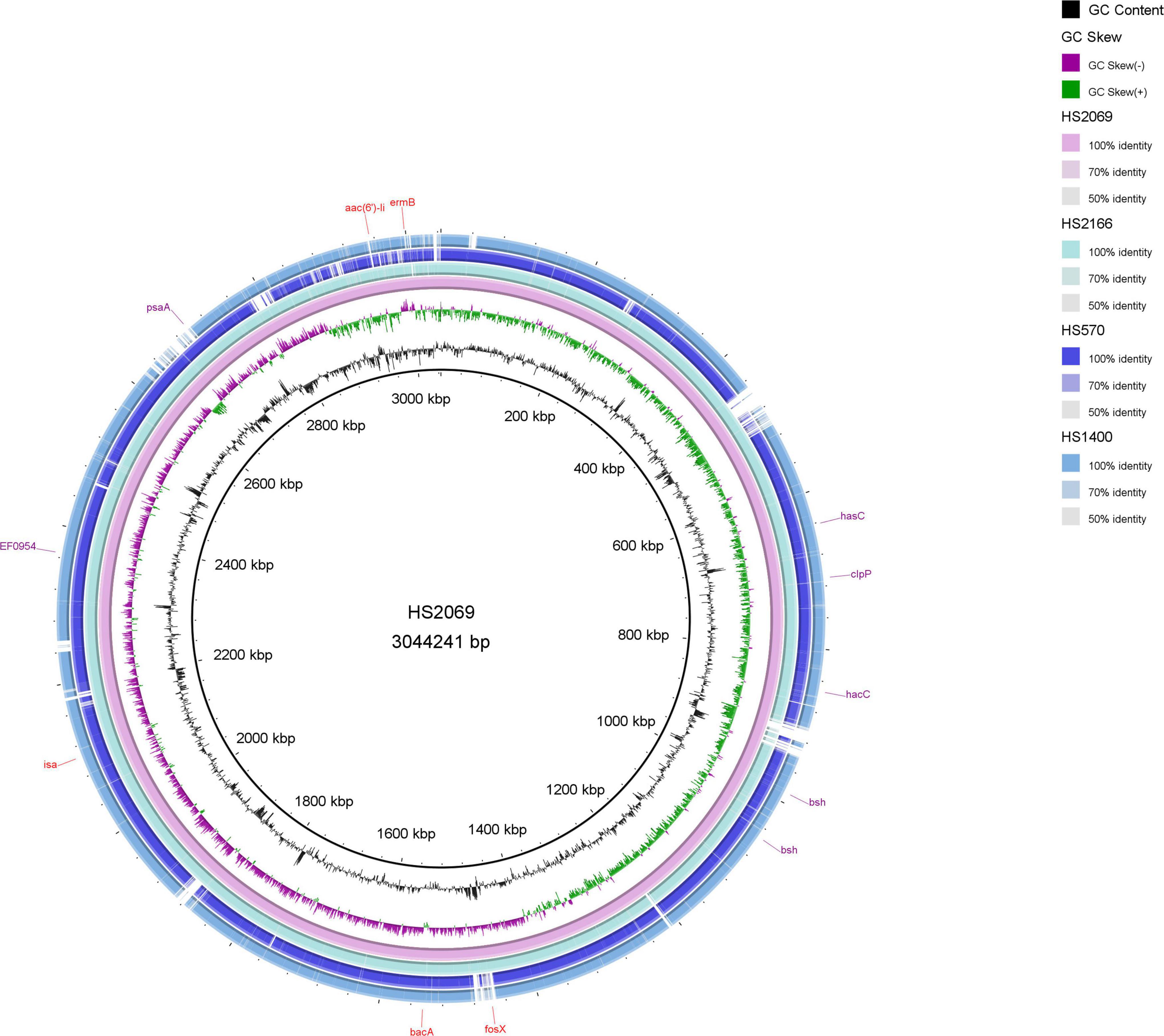
Figure 3. The circle map of HS2069, HS2166, HS570, and HS1440.
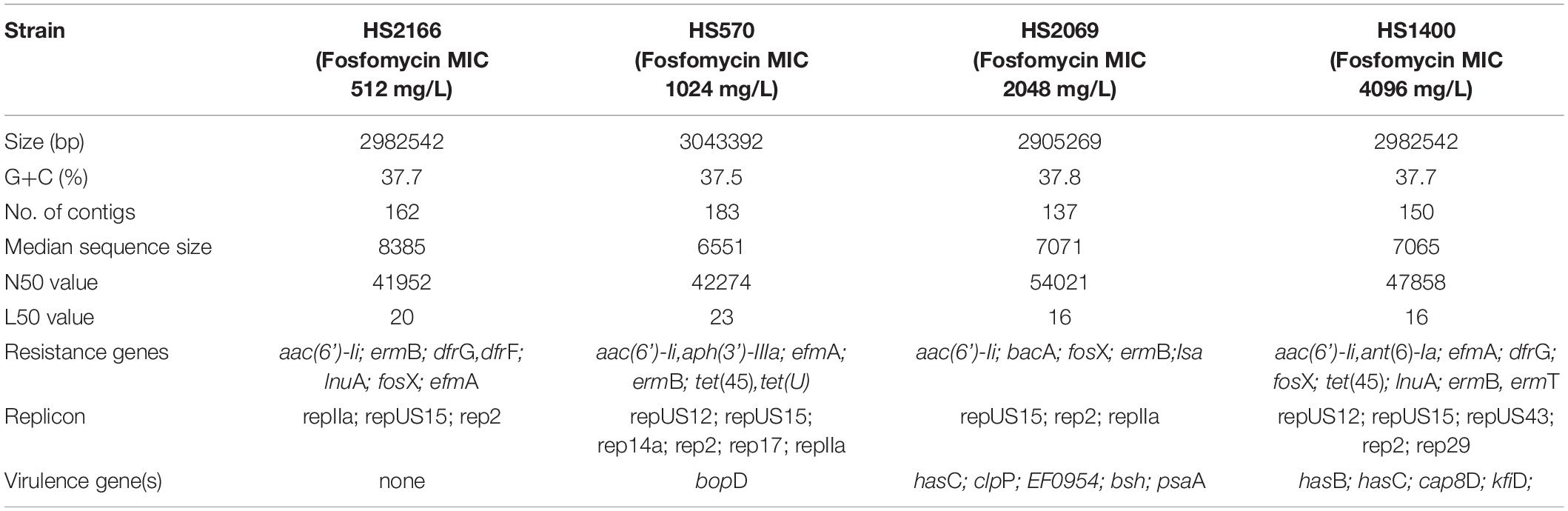
Table 2. The comparative genome sequences in HS2166, HS570, HS2069, and HS1440 strains.
Due to the unique mechanisms of action, fosfomycin exhibits significant antimicrobial activity against a broad spectrum of pathogens, including enterococci (Mendoza et al., 1980; Bernat et al., 1997). The fosfomycin resistance rate of enterococci isolated from South Africa was 1% (72/725) (Sahni et al., 2013). Zhang et al. (2020) reported that the fosfomycin resistance rate of enterococci isolated from China was 2.6% (20/761). The fosfomycin resistance rate of enterococci isolated from the United States was 1.3% (12/890) (Guo et al., 2017). In this study, 3.5% (153/4,414) of the enterococci were resistant to fosfomycin.
Little was known about the resistance mechanism of enterococci to fosfomycin in epidemiological research, and the most of genes that have been reported to be plasmid-mediated resistance to fosfomycin were fosB, with a G+C content of 27%, and were mainly distributed in staphylococcus, enterococci, and bacillus (Chen et al., 2014; Fu et al., 2016; Guo et al., 2017; Shinde et al., 2017; Ziwei et al., 2019; Xu et al., 2020). The fosB gene developed resistance to fosfomycin by encoding Mg2+-dependent sulfhydryl transferase, while the fosX was by encoding Mn2+-dependent epoxidase (Roberts et al., 2013). Recently, there are reports that the fosX gene was detected in Acinetobacter baumannii, Klebsiella pneumoniae, and E. faecium (Zhang et al., 2020; Kashefieh et al., 2021; Leite et al., 2021), indicating that the fosX gene is not unique to Gram-positive bacteria. The MICs of fosX-positive fosfomycin-resistant A. baumannii and K. pneumoniae were 128 and ≥ 200 mg/L, respectively (Kashefieh et al., 2021; Leite et al., 2021). Meanwhile, the MIC of fosfomycin-resistant E. faecium carrying the fosX gene was ≥ 512 mg/L (Zhang et al., 2020). In the current study, the MIC range of fosX-positive E. faecium was 512–4,096 mg/L.
Notably, in the report of Zhang et al., fosX, fosB genes, and murA mutation can coexist in fosfomycin-resistant enterococci (Zhang et al., 2020). Mutation of MurA protein exists in A. ba carrying fosX gene with fosfomycin MIC of 128 mg/L (Leite et al., 2021). Unlike previous studies, the fosX gene was exclusively present on fosfomycin-resistant E. faecium. More importantly, this study was the first time to clone the fosX gene in enterococci and proved that the fosX gene can be expressed in E. faecium, which mediates the reduction of the bacteria’s sensitivity to fosfomycin, indicating that these genes might be the primary factors mediating the resistance of E. faecium against fosfomycin.
As previously reported, the PFGE band patterns of 20 fosX-positive fosfomycin-resistant enterococci isolates showed 16 closely related isolates that exhibited ≥ 80% similarity (Zhang et al., 2020). In the current study, the PFGE patterns of 101 enterococci carrying the fosX gene showed 48 related strains that exhibited ≥ 80% similarity. A small number of isolates showed a high degree of homology, indicating that the possibility of genetic correlation between them was extremely small. The most frequent sequence type of enterococci carrying the fosX gene was the ST78 type in the previous studies (Zhang et al., 2020). Differently, ST555 (59/101) was the main clonal type in this study. The CC17 clonal complex E. faecium, which usually carries a virulence island and causes rapid dissemination in hospitals (Mancuso et al., 2021), belongs to a clonal complex closely related to nosocomial infection (Fiore et al., 2019). In this research, 94% (95/101) strains of E. faecium carrying the fosX gene belonged to the CC17 clonal complex, 57.4% (58/101) of the E. faecium belonged to the ST555 type, and most of them were isolated from Shanghai in China. Therefore, monitoring the ST555 type E. faecium is of great significance to curb the spread of antibiotic-resistant enterococci.
It was reported (Zhang et al., 2020) that the susceptibility to fosfomycin of enterococci carrying the fosX gene included resistance and sensitivity. However, the differences were that the enterococci carrying the fosX gene detected in this article were all fosfomycin-resistant strains. In addition, among the fosfomycin-resistant strains, the MIC of the isolates to fosfomycin is related to the expression level of the fosX gene, indicating that the function of the FosX resistance protein needs further verification.
In enterococci, the mobile genetic elements could account for 25% of the entire bacterial genome with strong plasticity (Hollenbeck and Rice, 2012). The fosfomycin resistance proteins FosX and FosA in pathogenic microorganisms are related to a catalytically promiscuous progenitor encoded in a phn operon in Mesorhizobium loti (Fillgrove et al., 2007). Chen et al. (2019) found that the fosA3 gene in clinical KPC-producing Klebsiella pneumoniae isolates collected from Zhejiang in China had an identical genetic background, IS26-tetR-cadC-orf1-fosA3-IS26, which is the same as that of the fosA3-positive plasmid pFOS18 in China. No other drug resistance genes have been reported in E. faecium carrying the fosX gene before. Parra-Flores et al. (2021) reported that ampicillin-resistant Listeria monocytogenes strains carried the fosX, tetA, and tetC resistance genes. In this study, fosX, aac(6’)-Ii, and ermB resistance genes co-existed in the genomes of E. faecium strains. Through the expression of the fosX gene was previously reported to be associated with the virulence factors hpy and prfA (Scortti et al., 2018), no such virulence genes were found in this study. The bopD (biofilm formation) virulence gene belongs to the type III secreted protein and exists in the HS570 strain with fosfomycin MIC of 1,024 mg/L. A large number of gram-negative pathogenic bacteria secrete virulence factors via the type III secretion system during infection of host cells (Nogawa et al., 2004). The capsular genotype (hasABC) (Kaczorek et al., 2017), K5-specific UDP-glucose dehydrogenase (kfiD) (Muñoz et al., 1998), caseinolytic peptidase P (clpP) (Jing et al., 2022), bile resistance(bsh) (Parra-Flores et al., 2021), pneumococcal surface adhesion A (psaA) (Hu et al., 2021), and polysaccharide biosynthesis (cap8D) (Ali et al., 2016) were detected from E. faecium in this study. Unfortunately, the effect of virulence factors on fosfomycin-resistant bacteria is unclear and needs to be explored by researchers.
It was worth noting that the previous report of the fosX gene in enterococci had only appeared in Zhejiang, China, which reported that 20 fosX-positive fosfomycin-resistant enterococci were screened out of 790 enterococci isolates. This study was the first time to screen large-scale fosfomycin resistance genes in enterococci isolated from 62 hospitals in 26 provinces or cities in China and found 101 fosfomycin-resistant strains carrying the fosX gene among 4,414 enterococci clinical strains.
In summary, 153 fosfomycin-resistant strains were screened out of 4,414 enterococci clinical isolates, and mechanisms were explored. Our finding described that fosB and fosX genes were the major resistant mechanism of enterococci to fosfomycin. In addition, the fosX gene could increase the resistance of bacteria to fosfomycin in E. faecium bacteria. FosX overexpression was associated with high-level fosfomycin resistance in E. faecium clinical isolates. Moreover, the 101 fosX carrying enterococci showed no obvious clonal transmission by PFGE analysis, and the dominant sequence type in them was ST555. There were 14 groups of 101 E. faecium carrying fosX were formed by E. faecium STs by MEGA-X and iTOL analysis. Therefore, it is necessary to continuously monitor fosfomycin resistance and its mechanisms.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
FH and YG designed the study. LX, QS, and RH collected clinical samples. LX, XX, and JW performed the experiments. LX, YG, and FH analyzed the data and wrote the manuscript. All authors contributed to the article and approved the submitted version.
The publication was supported by Independent Medical Grants from Pfizer, the National Natural Science Foundation of China (81902100 and 81861138052), the National Key Research and Development Program of China (2021YFC2701803), 3-year Action Plan for the Construction of Shanghai Public Health System (GWV-10.2-XD02), and the China Antimicrobial Surveillance Network (Independent Medical Grants from Pfizer, 2018QD100).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
LX would like to thank his supervisor, FH, whose expertise was invaluable in formulating the research questions and methodology. LX would particularly like to acknowledge his team members, YG, XX, QS, RH, and JW, for their wonderful collaboration and patient support.
Ali, L., Spiess, M., Wobser, D., Rodriguez, M., Blum, H. E., and Sakιnç, T. (2016). Identification and functional characterization of the putative polysaccharide biosynthesis protein (CapD) of Enterococcus faecium U0317. Infect. Genet. Evol. 37, 215–224. doi: 10.1016/j.meegid.2015.11.020
Bernat, B. A., Laughlin, L. T., and Armstrong, R. N. (1997). Fosfomycin resistance protein (FosA) is a manganese metalloglutathione transferase related to glyoxalase I and the extradiol dioxygenases. Biochemistry 36, 3050–3055. doi: 10.1021/bi963172a
Chen, C., Xu, X., Qu, T., Yu, Y., Ying, C., Liu, Q., et al. (2014). Prevalence of the fosfomycin-resistance determinant, fosB3, in Enterococcus faecium clinical isolates from China. J. Med. Microbiol. 63, 1484–1489. doi: 10.1099/jmm.0.077701-0
Chen, J., Wang, D., Ding, Y., Zhang, L., and Li, X. (2019). Molecular epidemiology of plasmid-mediated fosfomycin resistance gene determinants in Klebsiella pneumoniae Carbapenemase-producing Klebsiella pneumoniae isolates in China. Microb. Drug Resist. 25, 251–257. doi: 10.1089/mdr.2018.0137
Clinical and Laboratory Standards Institute (2020). Performance Standards for Antimicrobial Susceptibility Testing. M100-S30. Wayne, PA: CLSI.
Fillgrove, K. L., Pakhomova, S., Newcomer, M. E., and Armstrong, R. N. (2003). Mechanistic diversity of fosfomycin resistance in pathogenic microorganisms. J. Am. Chem. Soc. 125, 15730–15731. doi: 10.1021/ja039307z
Fillgrove, K. L., Pakhomova, S., Schaab, M. R., Newcomer, M. E., and Armstrong, R. N. (2007). Structure and mechanism of the genomically encoded fosfomycin resistance protein, FosX, from Listeria monocytogenes. Biochemistry 46, 8110–8120. doi: 10.1021/bi700625p
Fiore, E., Van Tyne, D., and Gilmore, M. S. (2019). Pathogenicity of Enterococci. Microbiol. Spectrum 7:10.1128/microbiolspec.GPP3-0053-2018
Fu, Z., Liu, Y., Chen, C., Guo, Y., Ma, Y., Yang, Y., et al. (2016). Characterization of fosfomycin resistance gene, fosB, in methicillin-resistant Staphylococcus aureus isolates. PLoS One 11:e0154829. doi: 10.1371/journal.pone.0154829
Guo, Y., Tomich, A. D., McElheny, C. L., Cooper, V. S., Tait-Kamradt, A., Wang, M., et al. (2017). High-Level fosfomycin resistance in vancomycin-resistant Enterococcus faecium. Emerg. Infect. Dis. 23, 1902–1904. doi: 10.3201/eid2311.171130
Hollenbeck, B. L., and Rice, L. B. (2012). Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 3, 421–433. doi: 10.4161/viru.21282
Hu, Y., Park, N., Seo, K. S., Park, J. Y., Somarathne, R. P., Olivier, A. K., et al. (2021). Pneumococcal surface adhesion a protein (PsaA) interacts with human Annexin A2 on airway epithelial cells. Virulence 12, 1841–1854. doi: 10.1080/21505594.2021.1947176
Jing, S., Ren, X., Wang, L., Kong, X., Wang, X., Chang, X., et al. (2022). Nepetin reduces virulence factors expression by targeting ClpP against MRSA-induced pneumonia infection. Virulence 13, 578–588. doi: 10.1080/21505594.2022.2051313
Kaczorek, E., Małaczewska, J., Wójcik, R., and Siwicki, A. K. (2017). Biofilm production and other virulence factors in Streptococcus spp. isolated from clinical cases of bovine mastitis in Poland. BMC Vet. Res. 13:398. doi: 10.1186/s12917-017-1322-y
Kashefieh, M., Hosainzadegan, H., Baghbanijavid, S., and Ghotaslou, R. (2021). The molecular epidemiology of resistance to antibiotics among Klebsiella pneumoniae isolates in azerbaijan. Iran. J. Trop. Med. 2021:9195184. doi: 10.1155/2021/9195184
Leite, G. C., Perdigão-Neto, L. V., Ruedas Martins, R. C., Rizek, C., Levin, A. S., and Costa, S. F. (2021). Genetic factors involved in fosfomycin resistance of multidrug-resistant Acinetobacter baumannii. Infect. Genet. Evol. 93:104943. doi: 10.1016/j.meegid.2021.104943
Mancuso, G., Midiri, A., Gerace, E., and Biondo, C. (2021). Bacterial antibiotic resistance: the most critical pathogens. Pathogens (Basel, Switzerland) 10:1310. doi: 10.3390/pathogens10101310
Mendoza, C., Garcia, J. M., Llaneza, J., Mendez, F. J., Hardisson, C., and Ortiz, J. M. (1980). Plasmid-determined resistance to fosfomycin in Serratia marcescens. Antimicrobial Agents Chemotherapy 18, 215–219. doi: 10.1128/AAC.18.2.215
Muñoz, R., García, E., and López, R. (1998). Evidence for horizontal transfer from Streptococcus to Escherichia coli of the kfiD gene encoding the K5-specific UDP-glucose dehydrogenase. J. Mol. Evol. 46, 432–436. doi: 10.1007/pl00006322
Murray, B. E., Singh, K. V., Heath, J. D., Sharma, B. R., and Weinstock, G. M. (1990). Comparison of genomic DNAs of different enterococcal isolates using restriction endonucleases with infrequent recognition sites. J. Clin. Microbiol. 28, 2059–2063. doi: 10.1128/jcm.28.9.2059-2063.1990
Nogawa, H., Kuwae, A., Matsuzawa, T., and Abe, A. (2004). The type III secreted protein BopD in Bordetella bronchiseptica is complexed with BopB for pore formation on the host plasma membrane. J. Bacteriol. 186, 3806–3813. doi: 10.1128/jb.186.12.3806-3813.2004
Parra-Flores, J., Holý, O., Bustamante, F., Lepuschitz, S., Pietzka, A., Contreras-Fernández, A., et al. (2021). Virulence and antibiotic resistance genes in Listeria monocytogenes strains isolated from ready-to-eat foods in chile. Front. Microbiol. 12:796040. doi: 10.3389/fmicb.2021.796040
Roberts, A. A., Sharma, S. V., Strankman, A. W., Duran, S. R., Rawat, M., and Hamilton, C. J. (2013). Mechanistic studies of FosB: a divalent-metal-dependent bacillithiol-S-transferase that mediates fosfomycin resistance in Staphylococcus aureus. Biochem. J. 451, 69–79. doi: 10.1042/BJ20121541
Sahni, R. D., Balaji, V., Varghese, R., John, J., Tansarli, G. S., and Falagas, M. E. (2013). Evaluation of fosfomycin activity against uropathogens in a fosfomycin-naive population in South India: a prospective study. Future Microbiol. 8, 675–680. doi: 10.2217/fmb.13.31
Scortti, M., Han, L., Alvarez, S., Leclercq, A., Moura, A., Lecuit, M., et al. (2018). Epistatic control of intrinsic resistance by virulence genes in Listeria. PLoS Genet. 14:e1007525. doi: 10.1371/journal.pgen.1007525
Shinde, D. B., Sangshetti, J. N., Moloney, M. G., Patil, R. H., and Joshi, S. S. (2017). Mur ligase inhibitors as anti-bacterials: a comprehensive review. Curr. Pharmaceutical Des. 23, 3164–3196. doi: 10.2174/1381612823666170214115048
Xu, W., Chen, T., Wang, H., Zeng, W., Wu, Q., Yu, K., et al. (2020). Molecular mechanisms and epidemiology of fosfomycin resistance in Staphylococcus aureus isolated from patients at a teaching hospital in China. Front. Microbiol. 11:1290. doi: 10.3389/fmicb.2020.01290
Xu, X., Chen, C., Lin, D., Guo, Q., Hu, F., Zhu, D., et al. (2013). The fosfomycin resistance gene fosB3 is located on a transferable, extrachromosomal circular intermediate in clinical Enterococcus faecium isolates. PLoS One 8:e78106. doi: 10.1371/journal.pone.0078106
Zhang, X., Bi, W., Chen, L., Zhang, Y., Fang, R., Cao, J., et al. (2020). Molecular mechanisms and epidemiology of fosfomycin resistance in enterococci isolated from patients at a teaching hospital in China, 2013-2016. J. Global Antimicrobial Resistance 20, 191–196. doi: 10.1016/j.jgar.2019.08.006
Keywords: Enterococci, fosfomycin, resistance, fosX, prevalence
Citation: Xin L, Xu X, Shi Q, Han R, Wang J, Guo Y and Hu F (2022) High Prevalence and Overexpression of Fosfomycin-Resistant Gene fosX in Enterococcus faecium From China. Front. Microbiol. 13:900185. doi: 10.3389/fmicb.2022.900185
Received: 20 March 2022; Accepted: 16 June 2022;
Published: 08 July 2022.
Edited by:
Xingmin Sun, University of South Florida, United StatesReviewed by:
Balkiss Bouhaouala, Pasteur Institute of Tunis, TunisiaCopyright © 2022 Xin, Xu, Shi, Han, Wang, Guo and Hu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yan Guo, Z3VveWFuQGZ1ZGFuLmVkdS5jbg==; Fupin Hu, aHVmdXBpbkBmdWRhbi5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.