 Tie Zhao
Tie Zhao Chunhui Wang3
Chunhui Wang3 Jianguo Wu
Jianguo Wu Qiwei Zhang
Qiwei Zhang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 12 May 2022
Sec. Virology
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.863802
This article is part of the Research TopicAdvances in Emerging Coronavirus Identification and Tracing MethodsView all 21 articles
Background: Coronavirus disease 2019 (COVID-19) is a global pandemic. Previous studies have reported dyslipidemia in patients with COVID-19. Herein, we conducted a retrospective study and a bioinformatics analysis to evaluate the essential data of the lipid profile as well as the possible mechanism in patients with COVID-19.
Methods: First of all, the retrospective study included three cohorts: patients with COVID-19, a healthy population, and patients with chronic obstructive pulmonary disease (COPD). For each subject, serum lipid profiles in the biochemical data were compared, including triglycerides (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C). Furthermore, bioinformatics analyses were performed for exploring the biological or immunological mechanisms.
Results: In line with the biochemical data of the three cohorts, the statistical result displayed that patients with COVID-19 were more likely to have lower levels of TC and HDL-C as compared with healthy individuals. The differential proteins associated with COVID-19 are involved in the lipid pathway and can target and regulate cytokines and immune cells. Additionally, a heatmap revealed that severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infections were possibly involved in lipid metabolic reprogramming. The viral proteins, such as spike (S) and non-structural protein 2 (Nsp2) of SARS-CoV-2, may be involved in metabolic reprogramming.
Conclusion: The metabolic reprogramming after SARS-CoV-2 infections is probably associated with the immune and clinical phenotype of patients. Hence, metabolic reprogramming may be targeted for developing antivirals against COVID-19.
The infections of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) led to the coronavirus disease 2019 (COVID-19) pandemic and have been a threat to public health across the world. There are 464,809,377 confirmed cases of COVID-19, including 6,062,536 deaths as of March 20, 2022 (World Health Organization [WHO], 2022). Public health and social and economic growth have been enormously influenced by the COVID-19 pandemic. SARS-CoV-2 contains four structural proteins, [envelope (E), membrane (M), nucleocapsid (N), and spike (S)], 16 non-structural proteins (Nsp1 to Nsp16), and eight accessory proteins (orf3a, orf6, orf7a, orf7b, orf8, orf9b, orf9c, and orf10). These proteins are involved in the viral life cycle and viral interaction with the host. Although the interaction between SARS-CoV-2 and the host is a moot point, the scientific community steadily gained an understanding of pathogenesis in the past.
Although the interactions between the immune system and lipid metabolism during SARS-CoV-2 infections remain unclear, the new development of tumor and metabolism study can provide new ideas and methods for the influence of SARS-CoV-2 infections. Interestingly, lipids are involved in viral pathogenesis and the pathophysiology of viral disease (Nie et al., 2020). Lipids not only constitute virus envelope but also involve viral replication and invasion. The composition of viruses and cells always includes lipids involved in membrane fusion and replication during the entry and the release from the host cell membrane. Previous studies (Wei et al., 2020; Jin et al., 2021; Mahat et al., 2021) have shown that lipid profiles, such as the total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C), in patients with COVID-19 are significantly altered. TC increases in the cell membrane, which benefits the virus entry to the host cells and the membrane fusion (Theken et al., 2021). The alterations of lipid profile in patients with COVID-19 seem to be proportional to the clinical phenotype and might be a target for risk evaluation. In addition, TC can regulate T-cell-mediated immune response and constitute T-cell receptors (TCRs) as a critical regulator, directly or indirectly (Bietz et al., 2017; Puleston et al., 2020). A previous study about lymphocytic choriomeningitis virus (LCMV) also showed that the clearance of the LCMV was significantly delayed in hypercholesterolemic mice, and LCMV-specific CD8+ T cells were suppressed (Ludewig et al., 2001). Cholesterol accumulation reduced the activation of CD8α- dendritic cells, thereby impairing Th1 cell responses while enhancing Th2 cell responses (Kim et al., 2021). Other evidence from oncology has demonstrated that multiple lipid species can be sensed by innate immune cells including macrophages and dendritic cells. Dyslipidemia is a critical regulator of adaptive immunity, which in turn can regulate adaptive immune cells (Kim et al., 2021).
However, the concentration of lipid profiles in patients with COVID-19 was reported with variable values (Gao et al., 2020; Hu et al., 2020; Lei et al., 2020; Malik et al., 2021). A likely explanation is that the genetic phenotypes and underlying diseases are significantly different among patients with COVID-19. To extend the existing evidence regarding the relationship between COVID-19 and lipid profile, a retrospective study and mechanism exploration by bioinformatics analyses were performed. We did extensive research about the actual relation between viral pathogenesis and lipid alteration through existing data. We attempted to elucidate the correlation between lipid profile and immunoreaction among patients with COVID-19, including lipid metabolism and profile, for example, dyslipidemia mechanism, cytokines, and T-cell-mediated immune response.
This retrospective study included three cohorts, 25 COVID-19 cases, 25 cases of the healthly examination population (control group, CG), and 25 cases with chronic obstructive pulmonary disease (COPD), recruited from the Huizhou Central People’s Hospital. COPD and CG never went through a previous infection with COVID-19 or received the vaccination. The patients were diagnosed with COVID-19 in light of the World Health Organization (WHO) guidelines.1 The nasopharyngeal swabs of patients with COVID-19 were collected for diagnosis. The laboratory-confirmed patient was defined as a positive result on the real-time reverse-transcriptase polymerase chain reaction (RT-PCR) assay of nasopharyngeal swab specimens. These cases were well balanced for gender, age, and primary disease. All the COVID-19 symptoms were mild, and no severe cases appeared. This study was performed according to the principles of the Declaration of Helsinki and approved by the Huizhou Central People’s Hospital following its guidelines for the protection of individual privacy.
For three cohorts, serum lipid profiles of patients with COVID-19, patients with COPD, and the healthy examination population were tested by biochemical methods (Roche Cobas 8000), including triglycerides (TG), TC HDL-C, and LDL-C. The sera of patients with COVID-19 were collected on admission.
The data based on the initial screening are retrieved mainly from the National Center for Biotechnology Information. Proteomic and lipidomic data from the sera/plasma of patients with COVID-19 were acquired from the early studies (Shen et al., 2020; Wu et al., 2020) and GEO datasets (GEO accession number: GSE157103). R was used to screen for differential proteins and lipids (R Core Team, 2018). Glycolysis pathway data were acquired from Caccuri et al. (2021). Profiles of serum cytokines and chemokines in patients with COVID-19 were acquired from Zawawi et al. (2021). SARS-CoV-2 S, E, Nsp15, Nsp16 (Sharma et al., 2021), and Nsp2 (Davies et al., 2020) proteins were determined and compared with the host transcriptomic responses to key viral genes.
The Venn diagram was generated based on the datasets (Venn, 2022). Gene Ontology (GO) [involving biological process (BP), cell component (CC), molecular function (MF), and Kyoto Encyclopedia of Genes and Genomes (KEGG)] were utilized to analyze the expected signaling pathways and corresponding functions of differential proteins via the package of R or platform of Enrichr (Chen et al., 2013; R Core Team, 2018). The heatmap was also generated by the package of R (R Core Team, 2018). The proteins network was constructed via the STRING dataset (Szklarczyk et al., 2021). Lipidmap was produced by the analysis of KEGG (Fahy et al., 2007). CytoHubba, an app of Cytoscape, was screened for hub genes (Shannon et al., 2003). The immune cell infiltration analysis was performed by GEPIA2021 (Li et al., 2021).
Values of serum lipid were shown as the mean (M) ± standard deviation (SD). The comparison for the lipid of three cohorts was made by one-way ANOVA using the SPSS 24.0 (IBM Corp.). The least significant difference (LSD) was further compared to show any significant difference between the two groups. Graphic plotting was generated using GraphPad Prism 8 (GraphPad Software, Inc.).
The study included 75 cases, which consisted of 25 COVID-19 patients positive for SARS-CoV-2 RNA, 25 cases diagnosed with COPD that all the history and symptoms supported, and 25 healthy people. The mean ages of CG, patients with COVID-19, and patients with COPD were 51 ±16.2, 47 ±15.4, and 54 ±17.6 years, respectively. The patients with COVID-19 did not use any statins according to medication guidelines. Other cohorts were similar. In all cases, previously diagnosed metabolic diseases (obesity, hypertension, and diabetes) were not incorporated based on self-report.
We found that there were significant differences in TG (F = 3.506, P < 0.05), TC (F = 17.123, P < 0.0001), and HDL-C (F = 21.473, P < 0.0001) levels between the three cohorts, but no significant difference was observed in LDL-C (F = 0.97, P > 0.05). In line with the biochemical data of the three cohorts, the statistical result displayed that the patients with COVID-19 were more likely to have a lower level of TC (P < 0.001) and HDL-C (P < 0.001) as compared with the healthy examination population. Patients with COPD had similar results (P < 0.001) (Figure 1).
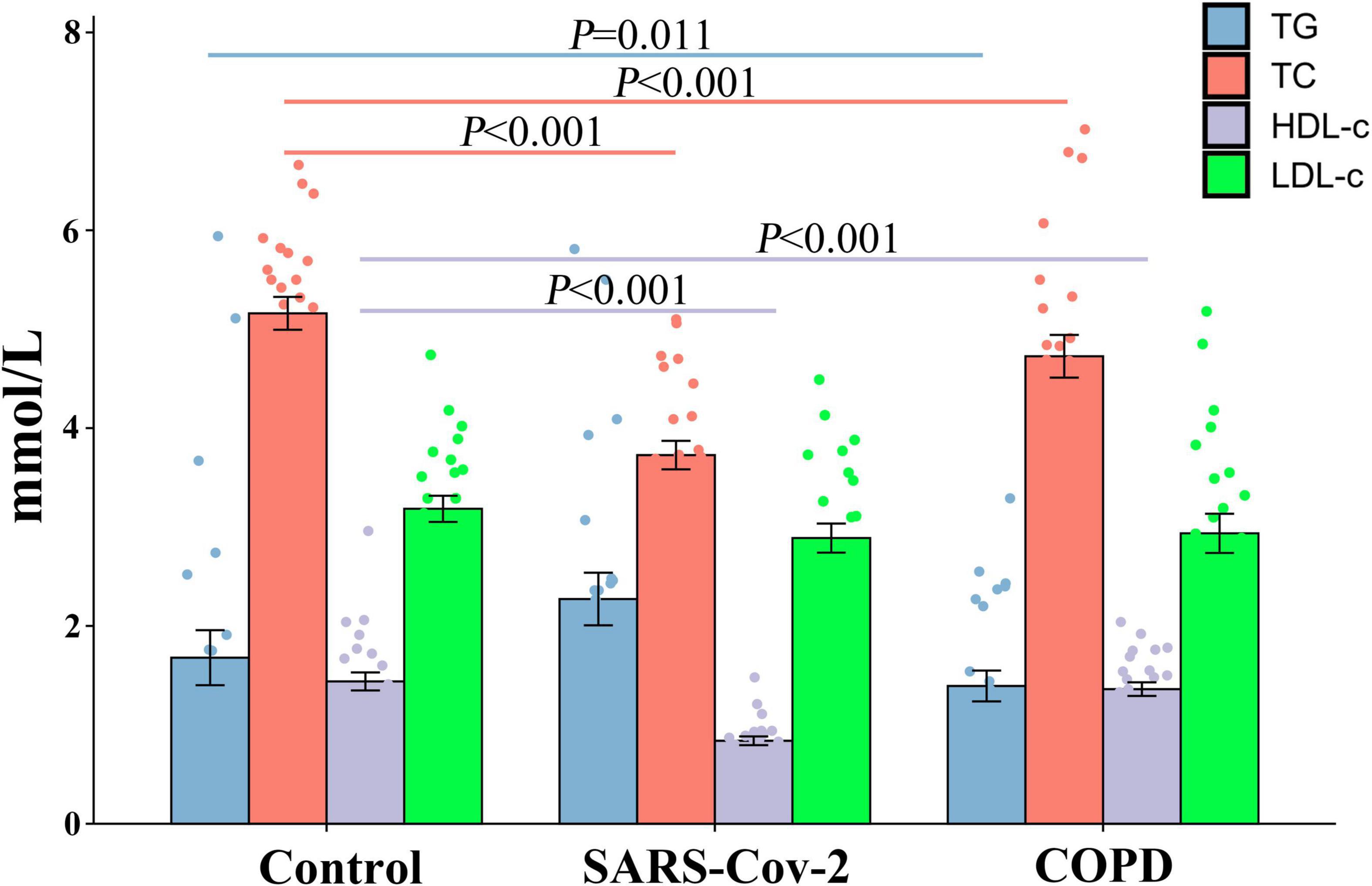
Figure 1. Comparison of the lipid level between the COVID-19, COPD patients, and healthy population. In COVID-19 and COPD patients, TG and HDL-C expression was reduced, but not for TC and LDL-C.
Differential proteins in patients with COVID-19 versus CG were acquired in GEO datasets. To further study the role of differential proteins, GO and KEGG signaling pathway analysis indicated that some proteins were involved in lipid pathways (Supplementary Table 1), such as the PPAR signaling pathway, cholesterol metabolism, fatty acid biosynthesis, positive regulation of cholesterol esterification (GO:0010873), and high-density lipoprotein particle remodeling (GO:0034375). The proteome of the sera/plasma of patients with COVID-19 showed 21 common proteins (Figure 2A), which are ORM1, ITIH3, ALB, SAA2, PGLYRP2, APOA1, NID1, GSN, CPN2, LGALS3BP, AGT, LCP1, C2, CLEC3B, ITIH4, APOM, CRTAC1, APOA2, ORM2, AHSG, and GPLD1. These 21 proteins had similar enrichment results that were associated with lipid pathways (Figure 2B and Supplementary Table 2). The lipid pathway included cholesterol metabolism, the PPAR signaling pathway, fat digestion and absorption, positive regulation of cholesterol esterification (GO:0010873), regulation of cholesterol esterification (GO:0010872), and so on. Conversely, lipid composition in patients with COVID-19 did not correspond with what other studies showed (data not shown). The enrichment analysis of lipidome in patients with COVID-19 showed that the blood lipid of humans was mainly involved in sphingolipid metabolism, cholesterol metabolism, fat digestion and absorption, and the sphingolipid signaling pathway (Supplementary Table 3).
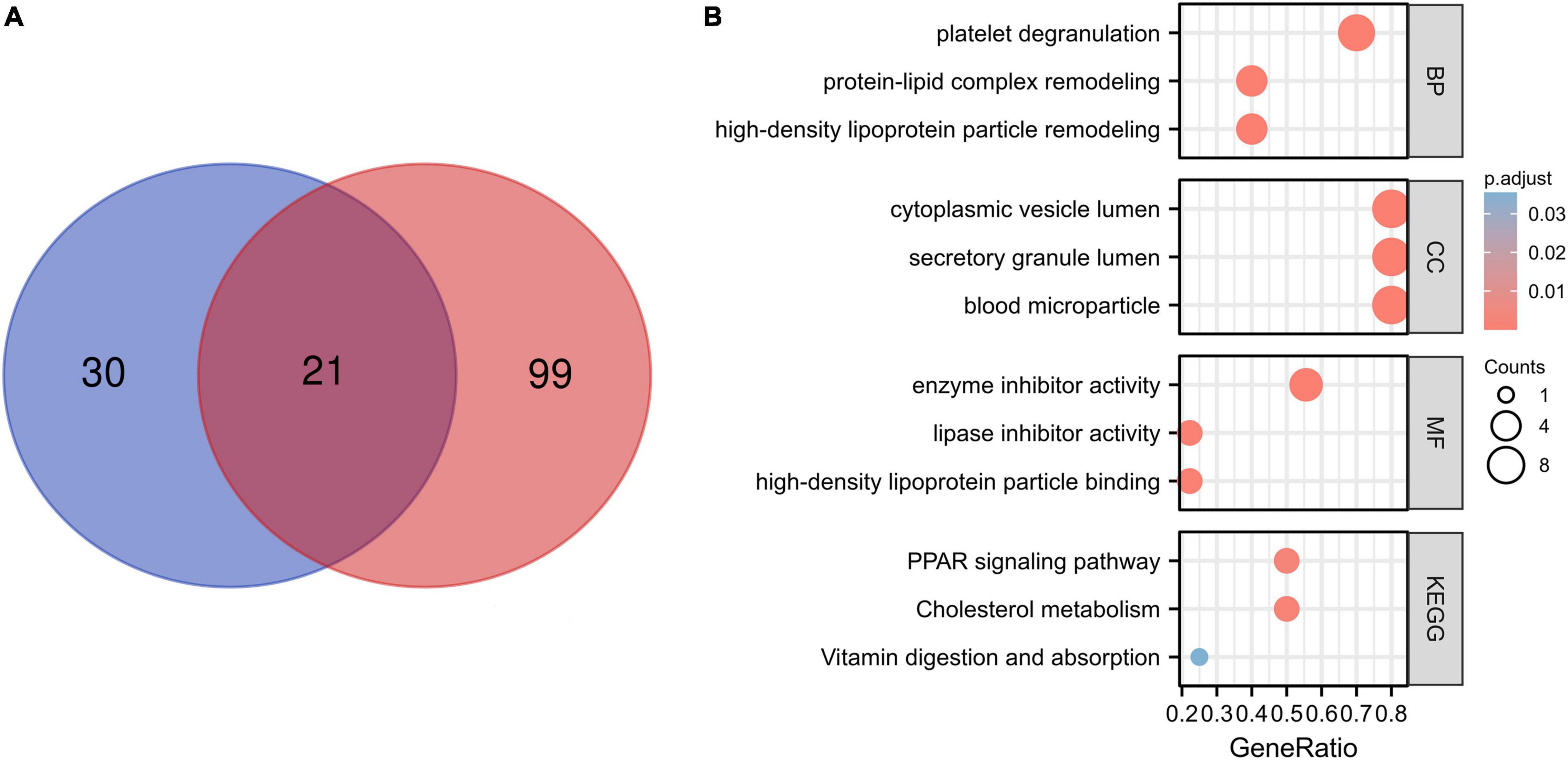
Figure 2. Proteome of the patients with COVID-19 and enrichment analysis. (A) A total of 21 common proteins were identified according to the proteome of the sera and plasma from patients with COVID-19 by Venn diagram. The blue color shows the proteome of sera of patients with COVID-19, and the red color shows plasma proteome associated with COVID-19. (B) A total of 21 common proteins involved in lipid pathway by the enrichment analysis, for example, protein–lipid complex remodeling and high-density lipoprotein particle remodeling.
The heatmap analysis showed the expression and increment of LDHA, GAPDH, and PKM post-SARS-CoV-2 infection; UV-inactivated SARS-CoV-2 can increase LDHA (Figures 3A,B). The GO and KEGG signaling pathway analysis indicated that the differential proteins of SARS-CoV-2 in endothelial cells were involved in glycolysis (Figure 3C). The lipid pathway and glycolysis occurred showed the potential for metabolic reprogramming post-SARS-CoV-2 infection (Figures 3D–F).
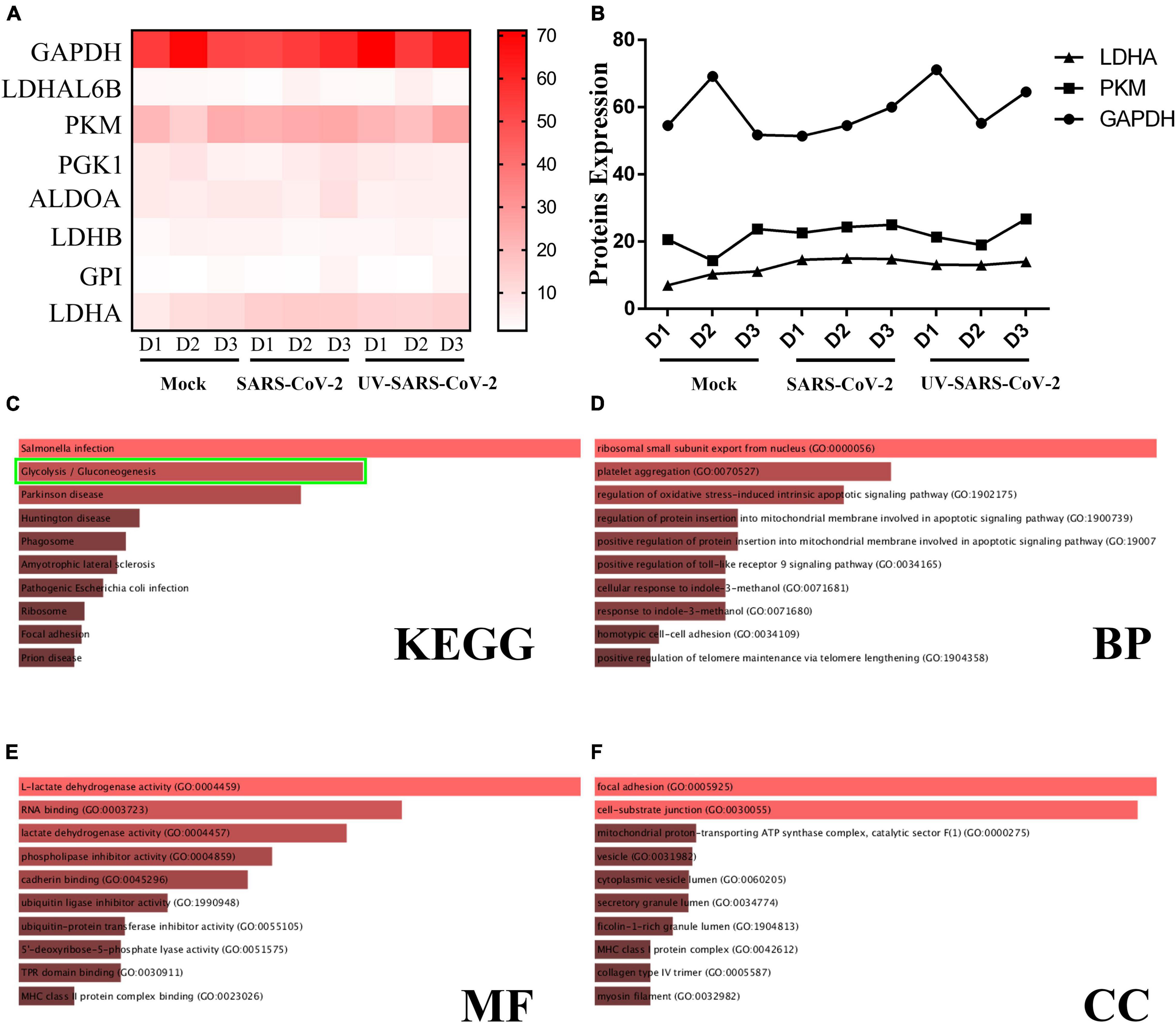
Figure 3. SARS-CoV-2 infection upregulates the glycolysis pathway. (A) A heatmap was generated from the SARS-CoV-2 infection and UV-inactivated SARS-CoV-2 treatment. In both groups, LDHA level was increased. (B) The expression of LDHA, PKM, and GAPDH was significantly different among the healthy population, SARS-CoV-2, and UV-SARS-CoV-2 groups. (C) The KEGG analysis revealed the SARS-CoV-2 infection and protein correlation involved in the glycolysis pathway (the green bounding box). (D–F) The BP, MF, CC analysis revealed the SARS-CoV-2 infection involved in the oxidative stress, the lactate dehydrogenase activity, et al.
The first 10 nodes (subproteins) (APOA1, ALB, AHSG, APOA2, ITIH4, ITIH3, ORM1, GSN, ORM2, and APOM) with the highest values were screened as fibrin clot (clotting cascade) and lipoprotein particle (Figures 4A,B). A protein–protein interaction (PPI) network for the first 10 nodes was constructed using the STRING database. These subproteins may regulate cytokines and chemokines in patients with COVID-19 (Figure 4C).
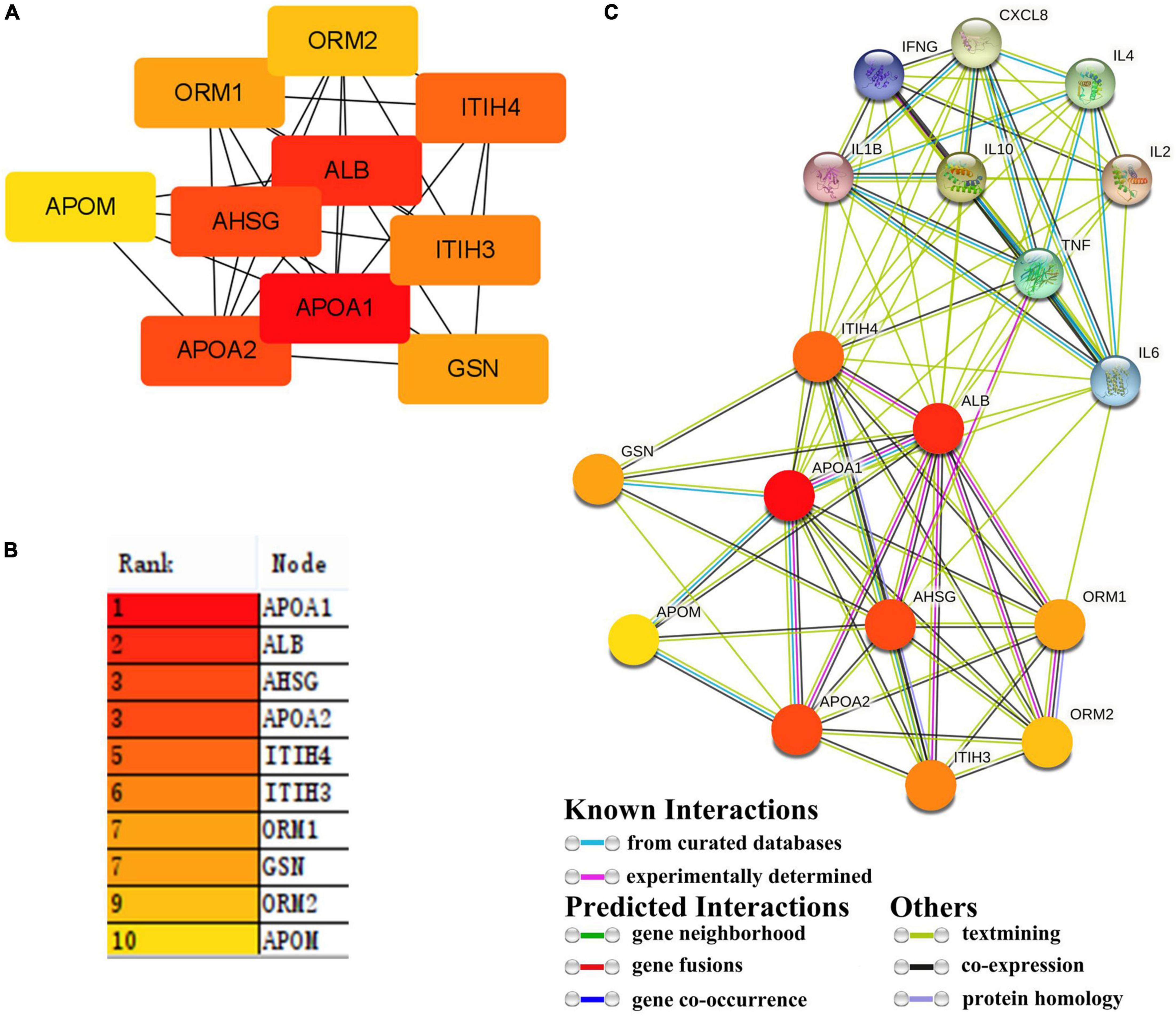
Figure 4. Hub protein in 21 proteins and interaction between hub proteins and cytokines. (A,B) 21 proteins were screened for 10 hub proteins. (C) The interaction between hub proteins and cytokines.
GEPIA2021 analysis further confirmed the correlations between the 10 sub-protein levels and cell types. Regarding the 10 sub-protein expression levels, the analysis of immune infiltration revealed that the CD4+ cell has the highest median value in the lung and the CD8+ cell has the highest median value in blood except GSN. A component analysis of the immune cells showed that CD4+ T cells, CD8+ T cells, and NK cells were significantly related to 10 subproteins (Figure 5).
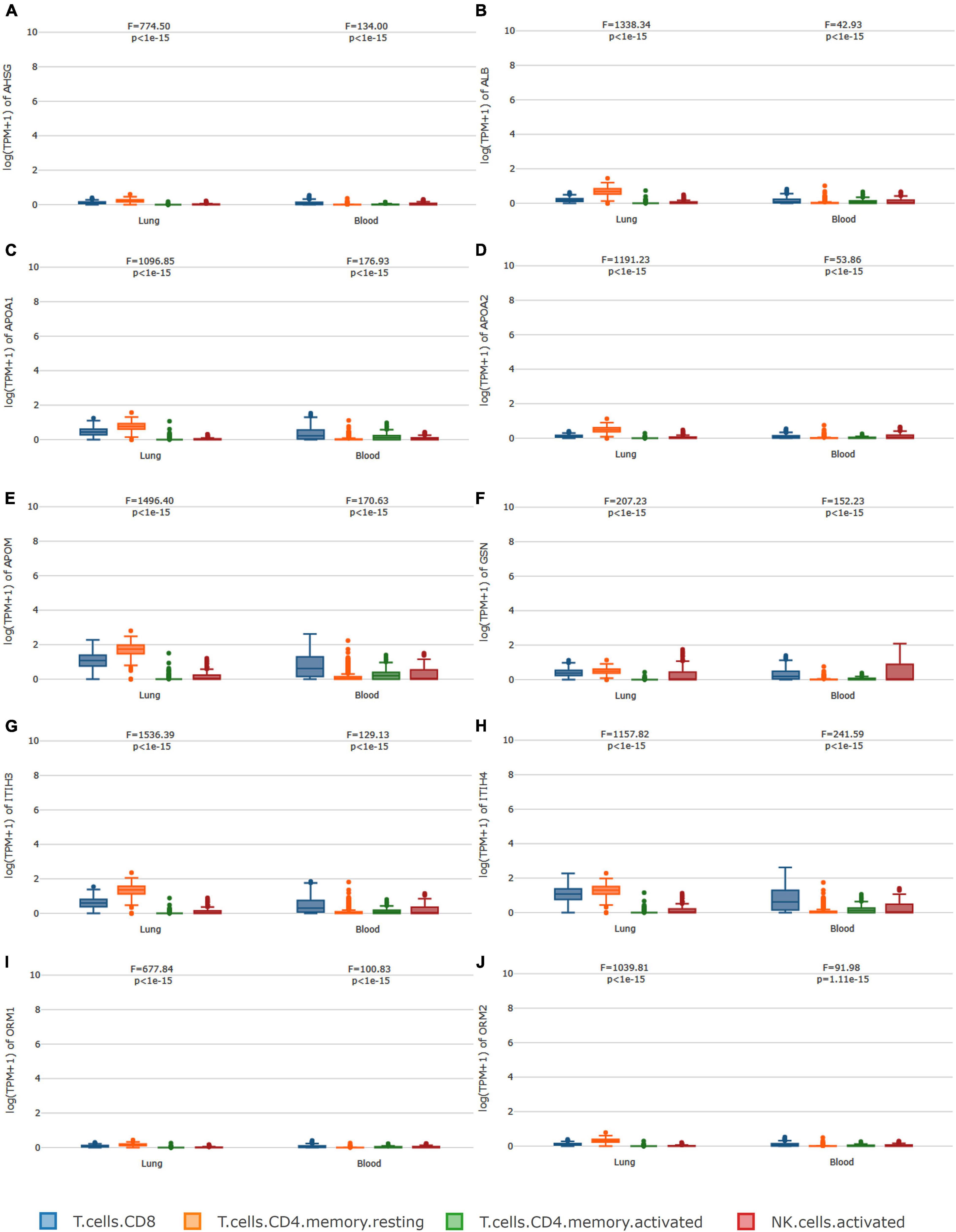
Figure 5. Immune cell type-level expression analysis. In regard to (A–J), the analysis of immune infiltration revealed that the CD4+ cell has the highest median value in the lung, and the CD8+ cell has the highest median value in blood except for GSN. A component analysis of the immune cells showed that CD4+ T cells, CD8+ T cells, and NK cells were significantly related to the 10 subproteins.
The S and Nsp2 proteins may involve metabolic reprogramming (Figure 6), but N, Nsp15, and Nsp16 would not. S1 subunit seems to regulate HSPA1A, HSPA6, HSPA1B, DDIT3, LDHB, HSP90B1, and EIF2AK3; S2 subunit seems to regulate HSPA1B, HSPA1A, HSPA6, and DDIT3; Nsp2 may regulate PLD3, VDAC2, HSPA8, HSPA5, ERLIN1, ERLIN2, and AGPAT2. These proteins are related to glycolysis and lipid pathways.
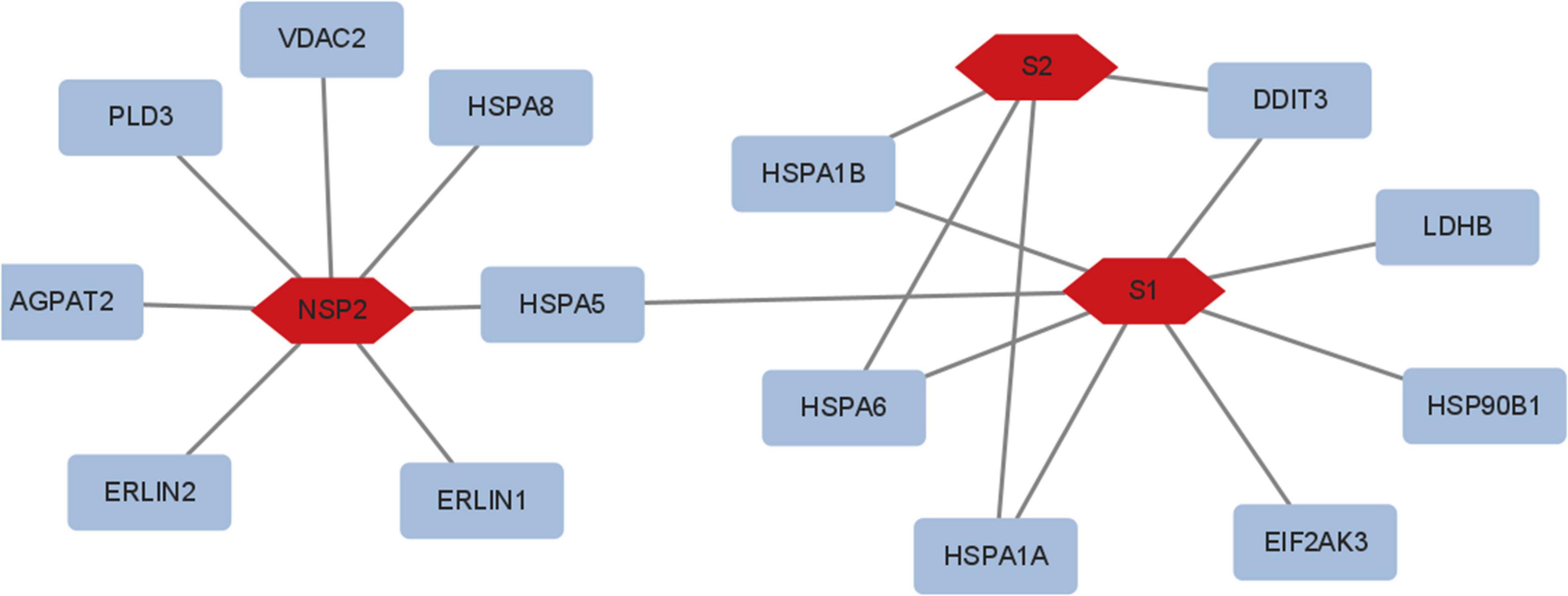
Figure 6. The viral proteins involved in metabolic reprogramming. S1 regulated HSPA1A, HSPA6, HSPA1B, DDIT3, LDHB, HSP90B1, and EIF2AK3; S2 regulated HSPA1B, HSPA1A, HSPA6, and DDIT3; Nsp2 regulated PLD3, VDAC2, HSPA8, HSPA5, ERLIN1, ERLIN2, and AGPAT2. N, Nsp15, and Nsp16 did not modify metabolic reprogramming.
Lipid profile alteration was used as a potential biomarker to aid diagnostics via triggers of viral infection. Our findings indicate that TC and HDL-C were reduced in patients with COVID-19, but TG and LDL-C did not. This finding is consistent with previous studies (Li G. et al., 2020; Li J. et al., 2020; Lv et al., 2020; Sun et al., 2020; Tanaka et al., 2020; Xie et al., 2020; Xue et al., 2020; Zhang B. et al., 2020; Zhang Q. et al., 2020). The serum/plasma concentrations of total TC and HDL-C were significantly lower in patients with COVID-19 with more severe diseases but were not for the TG. Significant changes in host lipidomes were observed in the cases of viral infection with severe disease, which induced changes in host immune function and benefited viral replication. On account of population and deviation, the results are possibly different in LDL-C. Distinctly, the heterogeneity between studies was generally large-to-extreme and multiple studies included small sample sizes. Interestingly, TC and HDL-C levels were associated with the clinical phenotype of SARS-CoV-2 infection. TC and HDL-C have beneficial effects on various pulmonary diseases and other diseases (Nie et al., 2020) and play a key role in modulating both innate and adaptive immune cell responses (Bietz et al., 2017). HDL has a function in inducing an anti-inflammatory or inflammatory profile (Van Lenten et al., 1995; Khovidhunkit et al., 2004). The reduced plasma levels of HDL can be found in patients with infection and sepsis (Wu et al., 2004; Cirstea et al., 2017). Lipid profile alteration is a useful indicator for early warning of the severity of COVID-19 disease (mild or severe) (Nie et al., 2020).
Energy and metabolites are required for cell survival. SARS-CoV-2 infections lead to a hypoxic microenvironment (Bhattacharya et al., 2021). This process is akin to the tumor microenvironment (TME) that the feature of TME is hypoxic. This promotes the host to compensate for their metabolic profiles to sustain a reprogramming state. The heatmap analysis showed that the expression of LDHA, a protein involved in glycolysis, was increased in SARS-CoV-2 infection. Furthermore, the proteomic data in two studies were analogous (Shen et al., 2020; Wu et al., 2020). The interaction network and enrich analysis revealed that the related pathways of lipid were located in the central node of all patient groups. There were significant similarities in lipid pathways among patients with COVID-19 from different regions. The 21 common proteins in this study supported this view. Enrichment analysis showed that the proteins were mainly involved in cholesterol metabolism, the PPAR signaling pathway, and so on. Anyhow, the hypoxic microenvironment in patients with COVID-19 increases the metabolic reprogramming for local nutrients and oxygen. However, the exact role or influence of metabolism reprogramming in SARS-CoV-2 immune response remains unclear, and lipids may regulate SARS-CoV-2 infection by multiple mechanisms.
Mechanisms derived from the previous study may also shed light on factors contributing to SARS-CoV-2 infectivity, where cholesterol is important either through immune regulator or by mediating signal pathway. The effects of lipids on infection development play a pivotal role. The function of lipids was gradually decrypted, which was used as an alternative source in pathologic conditions (Olsen et al., 2021), and was involved in the virus infection, was involved in transport of cell membrane, and activated intricate signaling pathways related to the immune system (Yu et al., 2021). Lipid metabolism dysfunction in the host has extensive effects on immune cells. The hub proteins were correlated with cytokines and chemokines in patients with COVID-19, and a distinct connection with immune cells was identified. However, the explanations for the lipid phenotype of patients are complex. In addition, individuals with underlying comorbidities (primary disease and metabolic disturbance in patients) will have more dramatic changes such that cholesterol provides a more complicated explanation and elaborate medical regimen. Due to the complex composition of lipids and a dynamically anabolic process, different points-in-time may respond very diversely to changes in lipid metabolism and give rise to ambiguous phenotypes. Enrichment analysis of lipids showed blood lipids of humans mainly involved in the sphingolipid metabolism, the cholesterol metabolism, fat digestion and absorption, and the sphingolipid signaling pathway, which suggested that the pathway was mainly for lipid-controlled biosynthesis or signaling. However, the studies of blood lipid were very heterogeneous. The quantification of blood lipids is still non-determined because of the complex component and much fluctuation of lipid quality and quantity in different space and time.
Interestingly, the lipid levels in patients with COPD changed and compared with the healthy population, but it was similar to patients with COVID-19. In addition, hypoxia is a common characteristic of patients that can change the metabolism (Grieb et al., 2021; Palm and Ekström, 2021). Hence, the patients were artificially ventilated, a procedure that can cause intraoperative complications but also can remit glycolysis or further metabolic reprogramming. So, the external reason was partially confirmed. The glycolysis suppression may be taken as a strategy for COVID-19 therapy and has profound therapeutic implications and significance. On the other hand, the lipid metabolism in patients with COVID-19 as a major altered function is highly similar to infection and sepsis, which is in accordance with a reply for multiple pathogens infection and in modulating inflammatory responses by the lipid moieties. These results indicate that metabolism plays a key role in SARS-CoV-2 pathogenesis and is a possible therapeutic target.
Meanwhile, the data showed that LDHA expression is increased in UV-SARS-CoV-2 infection. Besides anoxia, metabolic reprogramming was induced by the viral proteins as well. Viral structural proteins are involved in such processes. Numerous viruses (Negro, 2010; Funderburg and Mehta, 2016; Melo et al., 2016; Tisoncik-Go et al., 2016; Eisfeld et al., 2017; Kyle et al., 2019), such as Ebola virus, HIV, HBV, HCV, and homologous SARS-CoV (Wu et al., 2017) and MERS (Yan et al., 2019), can dramatically alter the human plasma lipidome. Even in the 12 years since the SARS-CoV infection, lipidome had been significantly changed (Wu et al., 2017). Therefore, viral proteins are involved similarly in metabolic reprogramming.
The viral proteins (Nsp2 and S) also affect lipid synthesis and modification (Díaz, 2020). The S protein of SARS-CoV-2 is a key protein. Numerous studies have confirmed that the S protein binds to ACE2 receptors on the surface of host cells to facilitate viral entry (Du et al., 2009; Walls et al., 2020). The S protein comprises S1 and S2 subunits in the virus replication cycle, binding the host cell receptor or fusing the viral envelope with host cell membranes. S1 plays an important role in protein processing in the endoplasmic reticulum, lipid, atherosclerosis, and so on. S2 was concerned with protein processing in the endoplasmic reticulum, the lipid, and atherosclerosis. Therefore, both S1 and S2 also modify lipid synthesis. Nsp2, a non-structural protein of SARS-CoV-2, disrupts host cell cycle and has similar functionality, which was concerned with protein processing in the endoplasmic reticulum, the lipid, and atherosclerosis. Our analysis revealed that S and Nsp2 proteins are associated with HSPA5, HSPA6, and LDHB in metabolic reprogramming. The hub proteins do not overlap, so SARS-CoV-2 pathogenesis is complicated. These findings also suggest an unknown potential protein inducing lipid synthesis and modification. What is driving the metabolic reprogramming is not clear.
In this study, essential baseline data, such as primary disease and statin use or not, might eliminate the observed heterogeneity. However, the exact timing of the blood collection for lipid profile remains uncertain. However, this can be ignored, as lipid metabolism of SARS-CoV-2 infection is a lengthy process as stated earlier. To eliminate the large between-study heterogeneity, population experiments have been incorporated in this study. This study mainly discussed pivotal lipids (TC and HDL-C) and glycolysis in metabolic reprogramming but did not mention other lipid species, such as the sphingolipids, and their related pathways, or amino acids, organic acids, and nucleotides. In addition, the full impact of metabolic reprogramming in SARS-CoV-2 infection cannot be confirmed by cell culture without pressures imposed by the immune microenvironment.
In conclusion, viral infection induces the alteration of host metabolic reprogramming, which is a remarkable feature. This alteration not only changes the immune and clinical phenotype of patients but is also involved in viral pathogenesis. So the virus–host interaction is figured thoroughly out. Therefore, antivirals may be developed via further study of the metabolic reprogramming mechanism along with the key proteins.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
The studies involving human participants were reviewed and approved by the Huizhou Central People’s Hospital. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
TZ wrote the manuscript and created the figures. CW provided the data of serum of patients with COVID-19. BD and PY edited the manuscript. JW and QZ reviewed the data and revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by grants from the National Key Research and Development Program of China (2018YFE0204503), the National Natural Science Foundation of China (32170139 and 81730061), the Natural Science Foundation of Guangdong Province (2018B030312010, 2021A1515010788, and 2022A1515011190), and the Fundamental Research Funds for the Central Universities (21622101).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.863802/full#supplementary-material
Bhattacharya, S., Agarwal, S., Shrimali, N. M., and Guchhait, P. (2021). Interplay between hypoxia and inflammation contributes to the progression and severity of respiratory viral diseases. Mol. Aspects Med. 81:101000. doi: 10.1016/j.mam.2021.101000
Bietz, A., Zhu, H., Xue, M., and Xu, C. (2017). Cholesterol metabolism in T cells. Front. Immunol. 8:664. doi: 10.3389/fimmu.2017.01664
Caccuri, F., Bugatti, A., Zani, A., De Palma, A., Di Silvestre, D., Manocha, E., et al. (2021). SARS-CoV-2 infection remodels the phenotype and promotes angiogenesis of primary human lung endothelial cells. Microorganisms 9:1438. doi: 10.3390/microorganisms9071438
Chen, E. Y., Tan, C. M., Kou, Y., Duan, Q., Wang, Z., Meirelles, G. V., et al. (2013). Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinformatics 14:128. doi: 10.1186/1471-2105-14-128
Cirstea, M., Walley, K. R., Russell, J. A., Brunham, L. R., Genga, K. R., and Boyd, J. H. (2017). Decreased high-density lipoprotein cholesterol level is an early prognostic marker for organ dysfunction and death in patients with suspected sepsis. J. Crit. Care 38, 289–294. doi: 10.1016/j.jcrc.2016.11.041
Davies, J. P., Almasy, K. M., McDonald, E. F., and Plate, L. (2020). Comparative multiplexed interactomics of SARS-CoV-2 and homologous coronavirus non-structural proteins identifies unique and shared host-cell dependencies. ACS Infect. Dis. 6, 3174–3189. doi: 10.1021/acsinfecdis.0c00500
Díaz, J. (2020). SARS-CoV-2 Molecular Network Structure. Front. Physiol. 11:870. doi: 10.3389/fphys.2020.00870
Du, L., He, Y., Zhou, Y., Liu, S., Zheng, B.-J., and Jiang, S. (2009). The spike protein of SARS-CoV target for vaccine and therapeutic development. Nat. Rev. Microbiol. 7, 226–236. doi: 10.1038/nrmicro2090
Eisfeld, A. J., Halfmann, P. J., Wendler, J. P., Kyle, J. E., Burnum-Johnson, K. E., Peralta, Z., et al. (2017). Multi-Platform ’omics analysis of human ebola virus disease pathogenesis. Cell Host Microbe 22, 817–829. doi: 10.1016/j.chom.2017.10.011
Fahy, E., Sud, M., Cotter, D., and Subramaniam, S. (2007). LIPID MAPS online tools for lipid research. Nucleic Acids Res. 35, 606–612. doi: 10.1093/nar/gkm324
Funderburg, N. T., and Mehta, N. N. (2016). Lipid abnormalities and inflammation in HIV inflection. Curr. HIV/AIDS Rep. 13, 218–225. doi: 10.1007/s11904-016-0321-0
Gao, S., Jiang, F., Jin, W., Shi, Y., Yang, L., Xia, Y., et al. (2020). Risk factors influencing the prognosis of elderly patients infected with COVID-19: a clinical retrospective study in Wuhan, China. Aging 12, 12504–12516. doi: 10.18632/aging.103631
Grieb, P., Swiatkiewicz, M., Prus, K., and Rejdak, K. (2021). Hypoxia may be a determinative factor in COVID-19 progression. Curr. Res. Pharmacol. Drug Discov. 2:100030. doi: 10.1016/j.crphar.2021.100030
Hu, X., Chen, D., Wu, L., He, G., and Ye, W. (2020). Declined serum high density lipoprotein cholesterol is associated with the severity of COVID-19 infection. Clin. Chim. Acta 510, 105–110. doi: 10.1016/j.cca.2020.07.015
Jin, H., He, J., Dong, C., Li, B., Ma, Z., Li, B., et al. (2021). Altered lipid profile is a risk factor for the poor progression of COVID-19: from two retrospective cohorts. Front. Cell. Infect. Microbiol. 11:712530. doi: 10.3389/fcimb.2021.712530
Khovidhunkit, W., Kim, M. S., Memon, R. A., Shigenaga, J. K., Moser, A. H., Feingold, K. R., et al. (2004). Effects of infection and inflammation on lipid and lipoprotein metabolism: mechanisms and consequences to the host. J. Lipid Res. 45, 1169–1196. doi: 10.1194/jlr.R300019-JLR200
Kim, D., Chung, H., Lee, J. E., Kim, J., Hwang, J., and Chung, Y. (2021). Immunologic aspects of dyslipidemia: a critical regulator of adaptive immunity and immune disorders. J. Lipid Atheroscler. 10, 184–201. doi: 10.12997/jla.2021.10.2.184
Kyle, J. E., Burnum-Johnson, K. E., Wendler, J. P., Eisfeld, A. J., Halfmann, P. J., Watanabe, T., et al. (2019). Plasma lipidome reveals critical illness and recovery from human ebola virus disease. Proc. Natl. Acad. Sci. U.S.A. 116, 3919–3928. doi: 10.1073/pnas.1815356116
Lei, P., Zhang, L., Han, P., Zheng, C., Tong, Q., Shang, H., et al. (2020). Liver injury in patients with COVID-19: clinical profiles, CT findings, the correlation of the severity with liver injury. Hepatol. Int. 14, 733–742. doi: 10.1007/s12072-020-10087-1
Li, C., Tang, Z., Zhang, W., Ye, Z., and Liu, F. (2021). GEPIA2021: integrating multiple deconvolution-based analysis into GEPIA. Nucleic Acids Res. 49, W242–W246. doi: 10.1093/nar/gkab418
Li, G., Zhou, C. L., Ba, Y. M., Wang, Y. M., Song, B., Cheng, X. B., et al. (2020). Nutritional risk and therapy for severe and critical COVID-19 patients: a multicenter retrospective observational study. Clin. Nutr. 40, 2154–2161. doi: 10.1016/j.clnu.2020.09.040
Li, J., Li, M., Zheng, S., Li, M., Zhang, M., Sun, M., et al. (2020). Plasma albumin levels predict risk for nonsurvivors in critically ill patients with COVID-19. Biomark. Med. 14, 827–837. doi: 10.2217/bmm-2020-0254
Ludewig, B., Jäggi, M., Dumrese, T., Brduscha-Riem, K., Odermatt, B., Hengartner, H., et al. (2001). Hypercholesterolemia exacerbates virus-induced immunopathologic liver disease via suppression of antiviral cytotoxic T cell responses. J. Immunol. 166, 3369–3376. doi: 10.4049/jimmunol.166.5.3369
Lv, Z., Wang, W., Qiao, B., Cui, X., Feng, Y., Chen, L., et al. (2020). The prognostic value of general laboratory testing in patients with COVID-19. J. Clin. Lab. Anal. 35:e23668. doi: 10.1002/jcla.23668
Mahat, R. K., Rathore, V., Singh, N., Singh, N., Singh, S. K., Shah, R. K., et al. (2021). Lipid profile as an indicator of COVID-19 severity: a systematic review and meta-analysis. Clin. Nutr. ESPEN 45, 91–101. doi: 10.1016/j.clnesp.2021.07.023
Malik, J., Laique, T., Ishaq, U., Ashraf, A., Malik, A., Ali, M., et al. (2021). Effect of COVID-19 on lipid profile and its correlation with acute phase reactants. MedRxiv [Preprint]. doi: 10.1101/2021.04.13.21255142
Melo, C. F., de Oliveira, D. N., Lima, E. O., Guerreiro, T. M., Esteves, C. Z., Beck, R. M., et al. (2016). A Lipidomics approach in the characterization of zika-infected mosquito cells: potential targets for breaking the transmission cycle. PLoS One 11:e0164377. doi: 10.1371/journal.pone.0164377
Negro, F. (2010). Abnormalities of lipid metabolism in hepatitis C virus infection. Gut 59, 1279–1287. doi: 10.1136/gut.2009.192732
Nie, S., Zhao, X., Zhao, K., Zhang, Z., Zhentao, Z., Zhang, Z., et al. (2020). Metabolic disturbances and inflammatory dysfunction predict severity of coronavirus disease 2019 (COVID-19): a retrospective study. MedRxiv [Preprint]. doi: 10.1101/2020.03.24.20042283
Olsen, L., Thum, E., and Rohner, N. (2021). Lipid metabolism in adaptation to extreme nutritional challenges. Dev. Cell 56, 1417–1429. doi: 10.1016/j.devcel.2021.02.024
Palm, A., and Ekström, M. (2021). Hypoxemia severity and survival in ILD and COPD on long-term oxygen therapy-The population-based DISCOVERY study. Respir. Med. 189:106659. doi: 10.1016/j.rmed.2021.106659
Puleston, D. J., Villa, M., and Pearce, E. L. (2020). Ancillary activity: beyond core metabolism in immune cells. Cell Metab. 26, 131–141. doi: 10.1016/j.cmet.2017.06.019
R Core Team (2018). R: A language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Sharma, A., Ong, J. W., Loke, M. F., Chua, E. G., Lee, J. J., Choi, H. W., et al. (2021). Comparative transcriptomic and molecular pathway analyses of HL-CZ human pro-monocytic cells expressing SARS-CoV-2 Spike S1, S2, NP, NSP15 and NSP16 Genes. Microorganisms 9:1193. doi: 10.3390/microorganisms9061193
Shen, B., Yi, X., Sun, Y., Bi, X., Du, J., Zhang, C., et al. (2020). Proteomic and metabolomic characterization of COVID-19 patient sera. Cell 182, 59–72.e15. doi: 10.1016/j.cell.2020.05.032
Sun, J. T., Chen, Z., Nie, P., Ge, H., Shen, L., Yang, F., et al. (2020). Lipid profile features and their associations with disease severity and mortality in patients with COVID-19. Front. Cardiovasc. Med. 7:584987. doi: 10.3389/fcvm.2020.584987
Szklarczyk, D., Gable, A. L., Nastou, K. C., Lyon, D., Kirsch, R., Pyysalo, S., et al. (2021). The STRING database in 2021: customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 49, D605–D612. doi: 10.1093/nar/gkaa1074
Tanaka, S., De Tymowski, C., Assadi, M., Zappella, N., Jean-Baptiste, S., Robert, T., et al. (2020). Lipoprotein concentrations over time in the intensive care unit COVID-19 patients: results from the ApoCOVID study. PLoS One 15:e0239573. doi: 10.1371/journal.pone.0239573
Theken, K. N., Tang, S. Y., Sengupta, S., and FitzGerald, G. A. (2021). The roles of lipids in SARS-CoV-2 viral replication and the host immune response. J. Lipid Res. 62:100129. doi: 10.1016/j.jlr.2021.100129
Tisoncik-Go, J., Gasper, D. J., Kyle, J. E., Eisfeld, A. J., Selinger, C., Hatta, M., et al. (2016). Integrated omics analysis of pathogenic host responses during pandemic H1N1 influenza virus infection: the crucial role of lipid metabolism. Cell Host Microbe 19, 254–266. doi: 10.1016/j.chom.2016.01.002
Van Lenten, B. J., Hama, S. Y., de Beer, F. C., Stafforini, D. M., McIntyre, T. M., Prescott, S. M., et al. (1995). Anti-inflammatory HDL becomes pro-inflammatory during the acute phase response. Loss of protective effect of HDL against LDL oxidation in aortic wall cell cocultures. J. Clin. Investig. 96, 2758–2767. doi: 10.1172/JCI118345
Venn (2022). Calculate and Draw Custom Venn Diagrams. Available online at: http://bioinformatics.psb.ugent.be/webtools/Venn/
Walls, A. C., Park, Y.-J., Tortorici, M. A., Wall, A., McGuire, A. T., and Veesler, D. (2020). Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 181, 281–292. doi: 10.1016/j.cell.2020.02.058
Wei, X., Zeng, W., Su, J., Wan, H., Yu, X., Cao, X., et al. (2020). Hypolipidemia is associated with the severity of COVID-19. J. Clin. Lipidol. 14, 297–304. doi: 10.1016/j.jacl.2020.04.008
Wu, A., Hinds, C. J., and Thiemermann, C. (2004). High-density lipoproteins in sepsis and septic shock: metabolism, actions, and therapeutic applications. Shock 21, 210–221. doi: 10.1097/01.shk.0000111661.09279.82
Wu, D., Shu, T., Yang, X., Song, J. X., Zhang, M., Yao, C., et al. (2020). Plasma metabolomic and lipidomic alterations associated with COVID-19. Natl. Sci. Rev. 7, 1157–1168. doi: 10.1093/nsr/nwaa086
Wu, Q., Zhou, L., Sun, X., Yan, Z., Hu, C., Wu, J., et al. (2017). Altered lipid metabolism in recovered SARS patients twelve years after infection. Sci. Rep. 7:9110. doi: 10.1038/s41598-017-09536-z
Xie, Y., You, Q., Wu, C., Cao, S., Qu, G., Yan, X., et al. (2020). Impact of cardiovascular disease on clinical characteristics and outcomes of coronavirus disease 2019 (COVID-19). Circ. J. 84, 1277–1283. doi: 10.1253/circj.CJ-20-0348
Xue, G., Gan, X., Wu, Z., Xie, D., Xiong, Y., Hua, L., et al. (2020). Novel serological biomarkers for inflammation in predicting disease severity in patients with COVID-19. Int. Immunopharmacol. 89:107065. doi: 10.1016/j.intimp.2020.107065
Yan, B., Chu, H., Yang, D., Sze, K.-H., Lai, P.-M., Yuan, S., et al. (2019). Characterization of the Lipidomic profile of human coronavirus-infected cells: implications for lipid metabolism remodeling upon coronavirus replication. Viruses 11:73. doi: 10.3390/v11010073
Yu, Y., Gao, L., Wang, Y., Xu, B., Maswikiti, E. P., Li, H., et al. (2021). A forgotten corner in cancer immunotherapy: the role of lipids. Front. Oncol. 11:751086. doi: 10.3389/fonc.2021.751086
Zawawi, A., Naser, A. Y., Alwafi, H., and Minshawi, F. (2021). Profile of circulatory cytokines and chemokines in human coronaviruses: a systematic review and meta-analysis. Front. Immunol. 12:666223. doi: 10.3389/fimmu.2021.666223
Zhang, B., Dong, C., Li, S., Song, X., Wei, W., and Liu, L. (2020). Triglyceride to high-density lipoprotein cholesterol ratio is an important determinant of cardiovascular risk and poor prognosis in coronavirus disease-19: a retrospective case series study. Diabetes Metab. Syndr. Obes. 13, 3925–3936. doi: 10.2147/DMSO.S268992
Keywords: COVID-19, lipid, SARS-CoV-2, dyslipidemias, metabolic reprogramming
Citation: Zhao T, Wang C, Duan B, Yang P, Wu J and Zhang Q (2022) Altered Lipid Profile in COVID-19 Patients and Metabolic Reprogramming. Front. Microbiol. 13:863802. doi: 10.3389/fmicb.2022.863802
Received: 27 January 2022; Accepted: 28 March 2022;
Published: 12 May 2022.
Edited by:
Ziyong Sun, Huazhong University of Science and Technology, ChinaReviewed by:
M. Nazmul Hoque, Bangabandhu Sheikh Mujibur Rahman Agricultural University, BangladeshCopyright © 2022 Zhao, Wang, Duan, Yang, Wu and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qiwei Zhang, emhhbmdxd0BqbnUuZWR1LmNu; orcid.org/0000-0002-2770-111X; Jianguo Wu, and1ODk4QGpudS5lZHUuY24=; orcid.org/0000-0002-8326-2895
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.