
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 09 March 2022
Sec. Microbiotechnology
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.807014
 Yan-Duo Wang1†
Yan-Duo Wang1† Jian Yang2†
Jian Yang2† Qi Li1Yuan-Yuan Li1Xiang-Mei Tan1Si-Yang Yao3Shu-Bin Niu3
Qi Li1Yuan-Yuan Li1Xiang-Mei Tan1Si-Yang Yao3Shu-Bin Niu3 Hui Deng4
Hui Deng4 Lan-Ping Guo2*
Lan-Ping Guo2* Gang Ding1*
Gang Ding1*Seco-sativene sesquiterpenoids are an important member of phytotoxins and plant growth regulators isolated from a narrow spectrum of fungi. In this report, eight seco-sativene sesquiterpenoids (1–8) were first analyzed using the UPLC-Q-TOF-MS/MS technique in positive mode, from which their mass fragmentation pathways were suggested. McLafferty rearrangement, 1,3-rearrangement, and neutral losses were considered to be the main fragmentation patterns for the [M+1]+ ions of 1–8. According to the structural features (of different substitutes at C-1, C-2, and C-13) in compounds 1–8, five subtypes (A–E) of seco-sativene were suggested, from which subtypes A, B/D, and E possessed the diagnostic daughter ions at m/z 175, 189, and 203, respectively, whereas subtype C had the characteristic daughter ion at m/z 187 in the UPLC-Q-TOF-MS/MS profiles. Based on the fragmentation patterns of 1–8, several known compounds (1–8) and two new analogues (9 and 10) were detected in the extract of plant pathogen fungus Bipolaris sorokiniana based on UPLC-Q-TOF-MS/MS analysis, of which 1, 2, 9, and 10 were then isolated and elucidated by NMR spectra. The UPLC-Q-TOF-MS/MS spectra of these two new compounds (9 and 10) were consistent with the fragmentation mechanisms of 1–8. Compound 1 displayed moderate antioxidant activities with IC50 of 0.90 and 1.97 mM for DPPH and ABTS+ scavenging capacity, respectively. The results demonstrated that seco-sativene sesquiterpenoids with the same subtypes possessed the same diagnostic daughter ions in the UPLC-Q-TOF-MS/MS profiles, which could contribute to structural characterization of seco-sativene sesquiterpenoids. Our results also further supported that UPLC-Q-TOF-MS/MS is a powerful and sensitive tool for dereplication and detection of new analogues from crude extracts of different biological origins.
Seco-sativenes are a member of sesquiterpenoids possessing a unique bicyclo[3.2.1]octane ring system and different substitutions including glycosylation, methylation, and acylation; different heterocyclic rings such as lactone, furan, and pyran ring; and diverse oxygenation sites (hydroxylation) on the core skeleton, increasing the chemical diversity. The structural differences of seco-sativenes mainly lie in the diverse substituents at C-1, C-2, and C-13 (Li et al., 2020a). From the structural features, it is implied that seco-sativenes come from a sesquiterpene pathway but not from a direct farnesyl pyrophosphate cyclization product. Rearrangement and oxidative cleavage reactions might play a pivotal role in the biosynthetic pathway, which is supported by the isolation of different precursors and intermediates (Mayo et al., 1962a,1965; Mayo and Williams, 1965; Li et al., 2020a,b). Fungus B. sorokiniana is known for producing a variety of secondary metabolites, with sesterterpene, cyclic peptides, and sesquiterpenoids as the most representative classes (Nihashi et al., 2002; Ali et al., 2016; Qader et al., 2017; Phan et al., 2019). Qader et al. (2017) isolated three seco-sativene sesquiterpenoids from B. sorokiniana, in which helminthosporal acid and helminthosporol displayed a strong phytotoxic effect on lettuce seed germination and toxicity against brine shrimps, and helminthosporal acid also showed antifungal activity. Phan et al. (2019) isolated and elucidated 12 seco-sativene sesquiterpenoids including a new seco-sativene sesquiterpenoid and three new sativene analogues from B. sorokiniana, in which helminthosporic acid and dihydroprehelminthosporol displayed weak necrotic activity against wheat leaves and helminthosporol showed an inhibitory effect on seed germination. Seco-sativene analogues displayed strong phytotoxic effects on cereals and gramineous plants, (Ludwig et al., 1956; Ludwig, 1957; Mayo et al., 1961, 1962b,1963; Spencer, 1965; Katsumi et al., 1967; Taniguchi and White, 1967; White and Taniguchi, 1972; Pena-Rodriguez et al., 1988; Pena-Rodriguez and Chilton, 1989; Qader et al., 2017; Phan et al., 2019) whereas others possessed plant-growth-promoting biological activities to rice, lettuce, cucumber, and wheat seedlings (Briggs, 1966; Hashimoto et al., 1967; Nukina et al., 1975; Pena-Rodriguez and Chilton, 1989; Miyazaki et al., 2017, 2018; Qader et al., 2017). In addition, some seco-sativene sesquiterpenoids also possessed antifungal, cytotoxic, and toxic effects, and other analogues could inhibit the growth of the malaria-causing protozoan of Plasmodium falciparum and exhibited certain anti-NO production activities (Li et al., 2020b). The novel core skeleton and diverse biological activities attracted us to chemically investigate this unique member of sesquiterpenoids. Recently, a series of new seco-sativene sesquiterpenoids were isolated from the endophytic fungus Cochliobolus sativus (teleomorph: Bipolaris sorokiniana) inhabiting in a desert plant, Artemisia desertorum, and their structures were mainly determined by NMR experiments, X-ray diffraction, and high-resolution mass analysis. Helminthosporic acid (2) could promote plant leaf growth, whereas cochliobolin F, helminthosporic acid (2), drechslerine B (8), and helminthosporal acid displayed strong phytotoxic effects on corn leaves (Li et al., 2020a). However, the traditional isolation method was used as the main technique for the isolation of seco-sativene sesquiterpenoids, (Ramos et al., 2019) which precluded discovery of new/novel analogues of seco-sativene sesquiterpenoids. Thus, efficient approaches for mining novel structures of seco-sativene sesquiterpenoids are urgent.
Mass spectrometry, especially tandem mass spectrometry, has been one of the most important physicochemical approaches for the characterization of secondary metabolites due to its rapidity and sensitivity (Jin et al., 2018; Liang et al., 2018; Conceição et al., 2020; Scupinari et al., 2020). Molecular weight and formula are often inconclusive for metabolite identification; however, fragmentation patterns represent a specific feature for a certain structural class. Chen et al. (2018) applied neutral loss scan in QqQ-MS and molecular formula calculation in UPLC-Q-TOF-MS to detect amorfrutin analogues, which provided the idea of detection and structural dereplication in the complex crude extract. Yang et al. (2017) used UPLC-Q-TOF-MS/MS coupled with neutral loss scan and diagnostic ions to analyze the secondary metabolites of Schisandra chinensis. Ahad et al. (2020) combined UPLC-Q-TOF-MS with SCX-SPE to achieve the enrichment and structural identification of the same skeleton metabolites. Thus, much evidence demonstrated that fragmentation patterns coupled with UPLC-Q-TOF-MS/MS analysis were an efficient and convenient tool for the detection and dereplication of similar metabolites.
A series of seco-sativene sesquiterpenoids (1–8, Figure 1) were isolated and elucidated in our previous work (Li et al., 2020a). According to the structural features (of different groups at C-1, C-2, and C-13), five subtypes of seco-sativenes were suggested (subtypes A-E) (Figure 1). Interestingly, each subtype of the structure has the same diagnostic daughter ions in the mass spectrometric profile, which could provide a reliable approach to analyze structures of seco-sativenes and target potent new analogues. To date, no investigations about electrospray tandem mass/mass of seco-sativenes sesquiterpenoids were reported. The potential application prospect and unique skeleton of seco-sativenes prompted us to investigate the mass spectrometric cleavage mechanisms of this unique member of sesquiterpenoids.
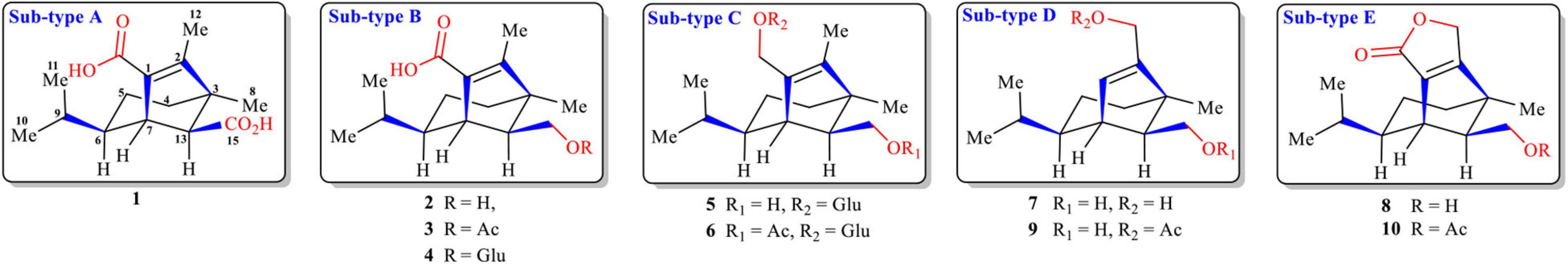
Figure 1. Structures of seco-sativene sesquiterpenoids (1–10).
In this report, the UPLC-Q-TOF-MS/MS fragmentation rules of seco-sativene sesquiterpenoids (1–8) were presented; some known and new seco-sativene sesquiterpenoids were detected from the extract of the plant pathogen fungus Bipolaris sorokiniana based on UPLC-Q-TOF-MS/MS analysis. Two new (9 and 10) and two known (1 and 2) seco-sativene sesquiterpenoids were then isolated and elucidated by HR-ESI-MS and NMR spectra, and the antioxidant activities of these seco-sativene sesquiterpenoids (1, 2, 9, and 10) were assessed.
Optical rotations were measured on a 241 polarimeter (PerkinElmer, Waltham, United States). UV-2102 (Unico, Shanghai, China) was used to record UV data. IR spectra were recorded on an FTIR-8400S spectrophotometer (Shimadzu, Kyoto, Japan). NMR data were acquired on a Bruker 500 spectrometer using solvent signal (CDCl3; δH 7.26/δC 77.6) as reference. Sephadex LH-20 and silica gel were purchased from Pharmacia (Biotech, Sweden) and Shanghai Titan Scientific Co., Ltd. (Shanghai, China), respectively. Semi-preparative HPLC separation was performed on a SEP LC-52 with an MWD UV detector (Separation (Beijing) Technology Co Ltd., Beijing, China) packed with a YMC-Pack ODS-A column. HR-ESI-MS spectra were analyzed using an ESI-Q-TOF-MS (Waters Xevo G2-XS QTof, United States).
Eight seco-sativene sesquiterpenoids were analyzed using a UPLC-Q-TOF-MS/MS system (Waters, United States). Chromatographic analysis was carried out with a Waters ACQUITY UPLC-PDA system equipped with an analytical reverse-phase C-18 column (2.1 × 100 mm, 1.7 μm, ACQUITY BEH, Waters, United States) with an absorbance range of 200 to 400 nm. The column temperature was maintained at 40°C. As the mobile phase, 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B) were used. The gradient conditions were as follows: 0–2 min, 35% B; 2–17 min, 35–98% B; 17–19 min, 98% B; and 19.1–21 min, 35% B. The flow rate from the UPLC system into the ESI-Q-TOF-MS detector was 0.3 ml/min. The auto-injected volume was 0.3 μl. Time-of-flight MS detection was performed with the Xevo G2-XS QTof system (Waters) combined with an ESI source in positive ion scan mode. The desolvation temperature was set at 450°C with desolvation gas flow at 900 L/h, and the source temperature was 80°C. The lock mass in all analyses was leucine-enkephalin [(M+H)+ = 556.2771], used at a concentration of 200 μl/ml and infused at a flow rate of 10 L/min. Raw data were acquired using the centroid mode, and the mass range was set from m/z 100 to 1,000. The capillary voltage was set at 2.5 kV with 30 V of sample cone voltage. The collision energy was set as 6 eV for low-energy scan and a ramp from 30 to 50 eV for high-energy scan. The instrument was controlled by MassLynx 4.1 software.
The strain of Bipolaris sorokiniana (strain number: ACCC36805) was isolated from the seed of wheat and provided by the Chinese Academy of Agricultural Sciences. The fungus was grown on PDA (potato dextrose agar) plates at 25°C for 10 days. Then the fresh mycelium was inoculated into the autoclaving sterilized solid medium with the formula of rice (60.0 g) and distilled water (80 ml) in Fernbach flasks (500 ml) for further fermentation at 25°C for 30 days.
The fermented rice substrate was extracted with EtOAc three times, and the solvent was evaporated to dryness under vacuum to afford 200 g of crude extract. The original extract was fractionated on a silica gel column using petroleum ether-acetone (1:0–0:1) progressively to give five fractions (Fr. 1 to Fr. 5). Fr. 2 (8.0 g) was separated on a silica gel column to obtain eight fractions (Fr. 2.1 to Fr. 2.8). Fr. 4 (27.8 g) was separated on a silica gel column to obtain five parts (Fr. 4.1 to Fr. 4.5). Fr. 4.1 was separated on a silica gel column and RP-HPLC (0–5 min 80% MeOH in H2O, 5–25 min 80–100% MeOH in H2O, 5 ml/min) to obtain Fr. 4.1.1 (13.8 mg, tR = 10.5 min). Fr. 4.1.1 (13.8 mg) was purified by semi-preparative HPLC (0–5 min 70% acetonitrile in H2O, 5–25 min 70–90% acetonitrile in H2O, 2 ml/min) to obtain compound 10 (1.8 mg, tR = 15.2 min). Fr. 4.2 (5.6 mg) was purified by semi-preparative HPLC (60% acetonitrile in H2O, 2.5 ml/min) to obtain compound 9 (2.3 mg, tR = 23.5 min). Fr. 4.3 (3.5 g) was purified by semi-preparative HPLC (0–25 min 80–100% MeOH in H2O, 5 ml/min) to obtain compound 2 (230.1 mg, tR = 10.3 min). Fr. 4.4 (4.02 g) was separated on a silica gel column, Sephadex LH-20 (dichloromethane:methanol = 1:1 v/v) and semi-preparative HPLC to obtain four fractions (Fr. 4.4.1–Fr. 4.4.4). Fr. 4.4.1 (24.7 mg) was purified by semi-preparative HPLC (75% MeOH in H2O, 2 ml/min) to obtain 1 (10.0 mg, 21.6 min).
Colorless oil: -18 (c 0.1, MeOH); UV (MeOH) λmax (log ε) 215 (2.98) nm; IR (neat) νmax 3,420, 2,931, 1,742, 1,456, 1,367, 1,235, 1,031 cm–1; for 1H NMR and 13C NMR data, see Table 1; positive HR-ESI-MS: m/z 267.1957 [calcd. for C16H27O3 [M+H]+, 267.1960].
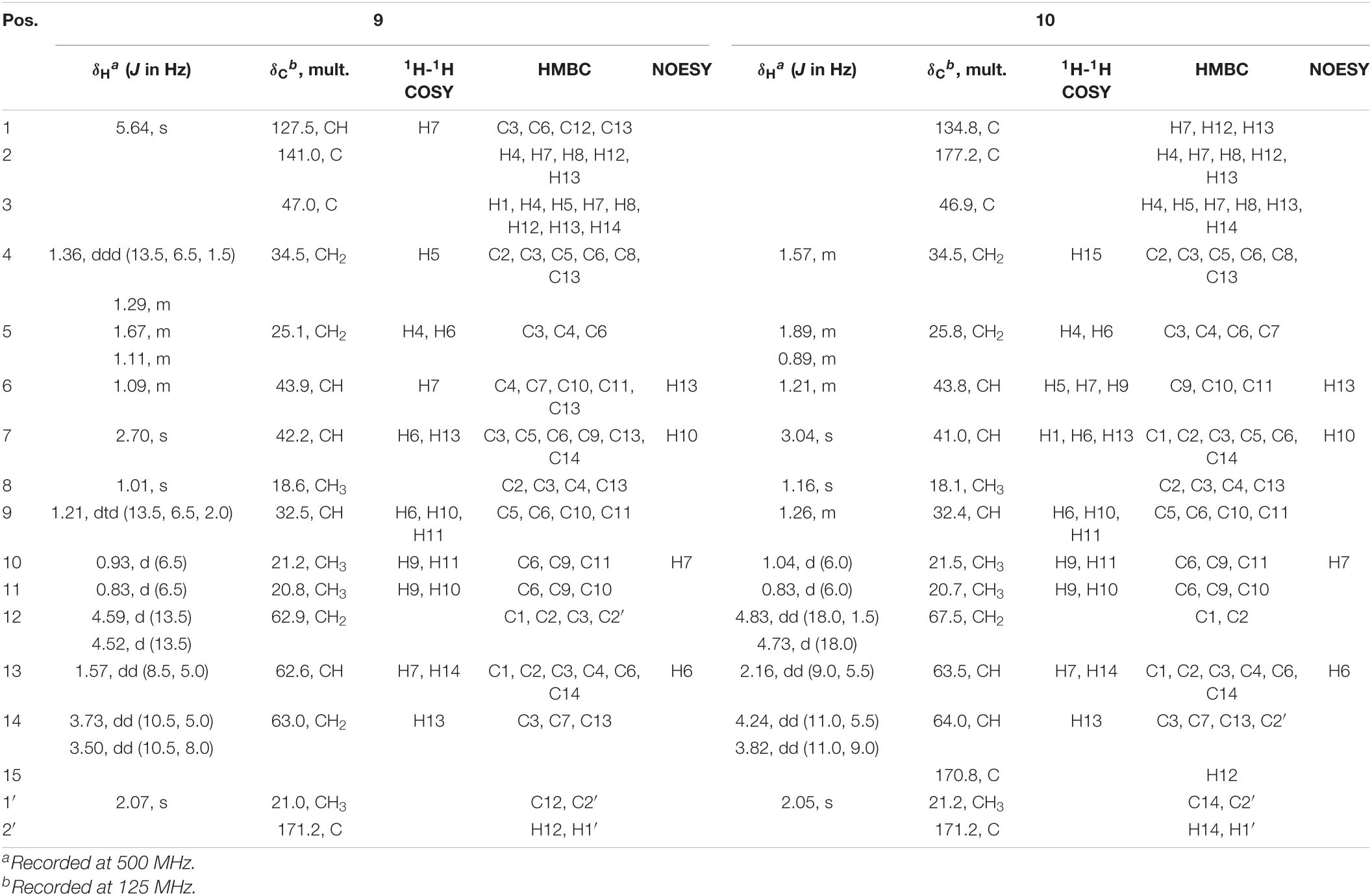
Table 1. NMR spectroscopic data of compounds 9 and 10 in CDCl3.
White powder: -16 (c 0.1, MeOH); UV (MeOH) λmax (log ε) 205 (2.84), 234 (2.97) nm; IR (neat) νmax 2,958, 1,747, 1,645, 1,456, 1,367, 1,338, 1,234, 1,031 cm–1; for 1H NMR and 13C NMR data, see Table 1; positive HR-ESI-MS: m/z 293.1683 [calcd. for C17H25O4 [M+H]+, 293.1675].
Take 15 μl of compounds 1, 2, 9, and 10 with a concentration of 10 mM/L and a serial dilution of seven times, and then mix with DPPH solution. After 30 min, the remaining amount of the DPPH radical was measured spectrophotometrically at 517 nm. In this test, for comparison, VC was considered as the positive control, and ethanol was considered as the negative control.
The clearance rate E is E = [1 – (AS – A0)/(AC – A0)] × 100%, where A0 is the absorbance of the water, AC is the absorbance of ethanol solution, and AS is the absorbance after adding the sample solution. The IC50 value was processed by GraphPad Prism 8.
Take 15 μl of compounds 1, 2, 9, and 10 with a concentration of 10 mM/L and a serial dilution of seven times, and then mix with ABTS+ solution. After 6 min, the remaining amount of the ABTS+ radical was measured spectrophotometrically at 405 nm. In this test, for comparison VC served as the positive control, and ethanol served as the negative control.
The clearance rate E is E = [1 – (AS – A0)/(AC – A0)] × 100%, where A0 is the absorbance of the water, AC is the absorbance of ethanol solution, and AS is the absorbance after adding the sample solution. The IC50 value was processed by GraphPad Prism 8.
The protonated parent ion m/z 267, [M+H]+ of cochliobolin A (1) was not observed in the UPLC-Q-TOF-MS/MS spectra, and it might be that this ion could easily lose one molecule of H2O to produce a daughter ion at m/z 249 [M+1-H2O]+. The abundance of ion m/z 249 was the highest in the profile. Thus, the precursor protonic molecular ion [M+1-H2O]+ of 1 was selected for analysis in the UPLC-Q-TOF-MS/MS spectrum. The high-resolution mass and fragment ions together with the elemental constituents of cochliobolin A (1) were listed in Table 2. The fragmentation routes according to ESI-Q-TOF-MS/MS analysis were depicted in Scheme 1, in which typical neutral losses, McLafferty rearrangement, and 1,3-rearrangement were the main fragmentation patterns for the parent ion m/z 267 [M+1]+ (Liang et al., 2018). The daughter ion (m/z 249) was formed from the parent ion (m/z 267) through the McLafferty rearrangement and 1,3 rearrangement by loss of one molecule of H2O (−18). Then, the daughter ion (m/z 221) was produced from the precursor ion (m/z 249) through neutral loss of one molecule of CO (−28). The daughter ion (m/z 231) was yielded from the precursor ion (m/z 249) through the McLafferty rearrangement by neutral loss of one molecule of H2O (−18). The diagnostic daughter ion (m/z 175) might have originated from the precursor ion (m/z 249) by loss of one molecule of HCO2H (−46) (m/z 203) and one molecule of CO (−28) (m/z 175) through the McLafferty rearrangement and neutral loss (Scheme 1 and Supplementary Figure 1).
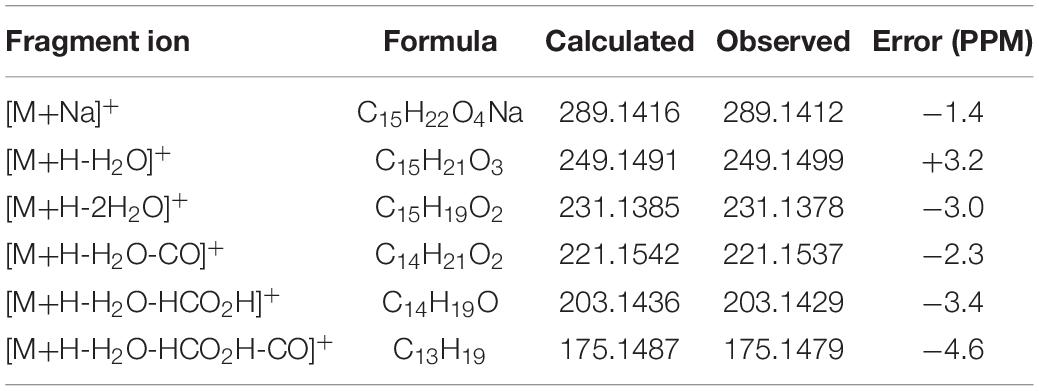
Table 2. Elemental constituents of major product ions from [M+Na]+ for compound 1 (subtype A).
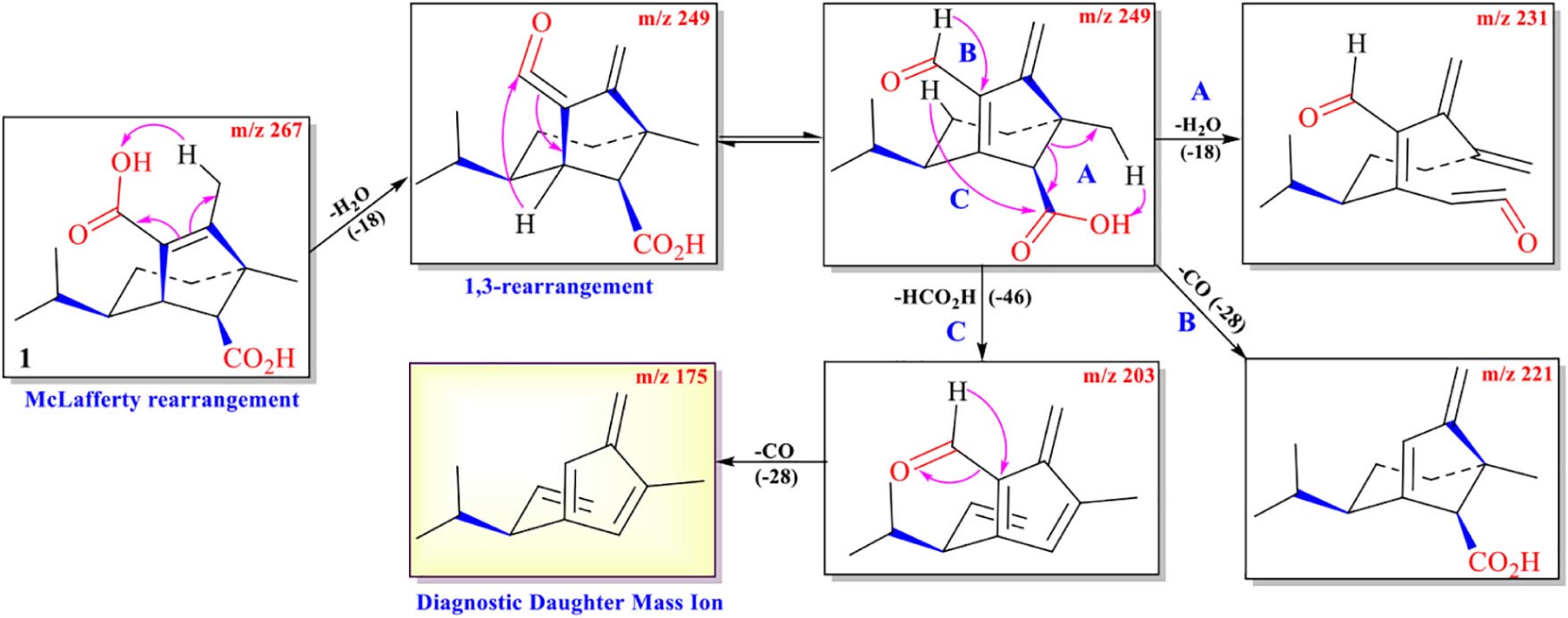
Scheme 1. Possible fragmentation pathway of 1.
The protonated parent ion m/z 253, [M+H]+ of compound 2 was observed in the mass profile with a relatively low abundance, which easily lost one molecule of H2O (−18) to produce the intermediate ion m/z 235 through the McLafferty rearrangement and 1,3-rearrangement or neutral loss of one molecule of H2O (–18) with H-13. Then, the intermediate ion m/z 235 successively lost one molecule of H2O (–18) and one molecule of CO (–28) by neutral loss to form the key diagnostic daughter ion (m/z 189). The high-resolution mass and fragment ions together with the elemental constituents of compound 2 were listed in Table 3. Compounds 3 and 4 had similar fragmentation pathways to that of compound 2 (Scheme 2, Supplementary Figures 2–4, and Supplementary Tables 1, 2).
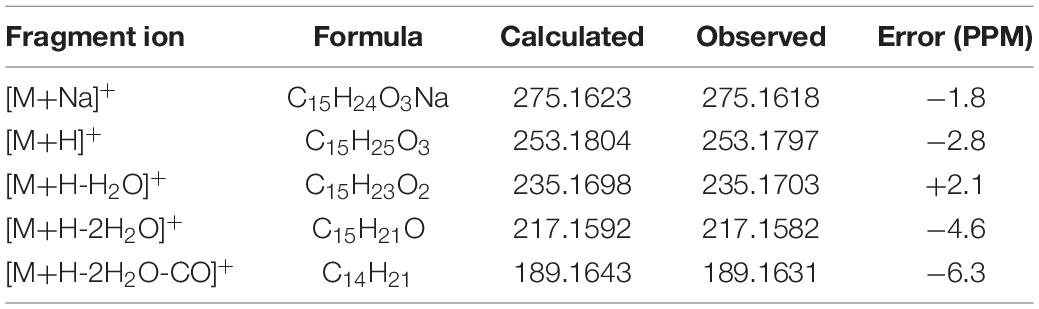
Table 3. Elemental constituents of major product ions from [M+Na]+ for compound 2 (subtype B).
Scheme 2. Possible fragmentation pathways of 2–4.
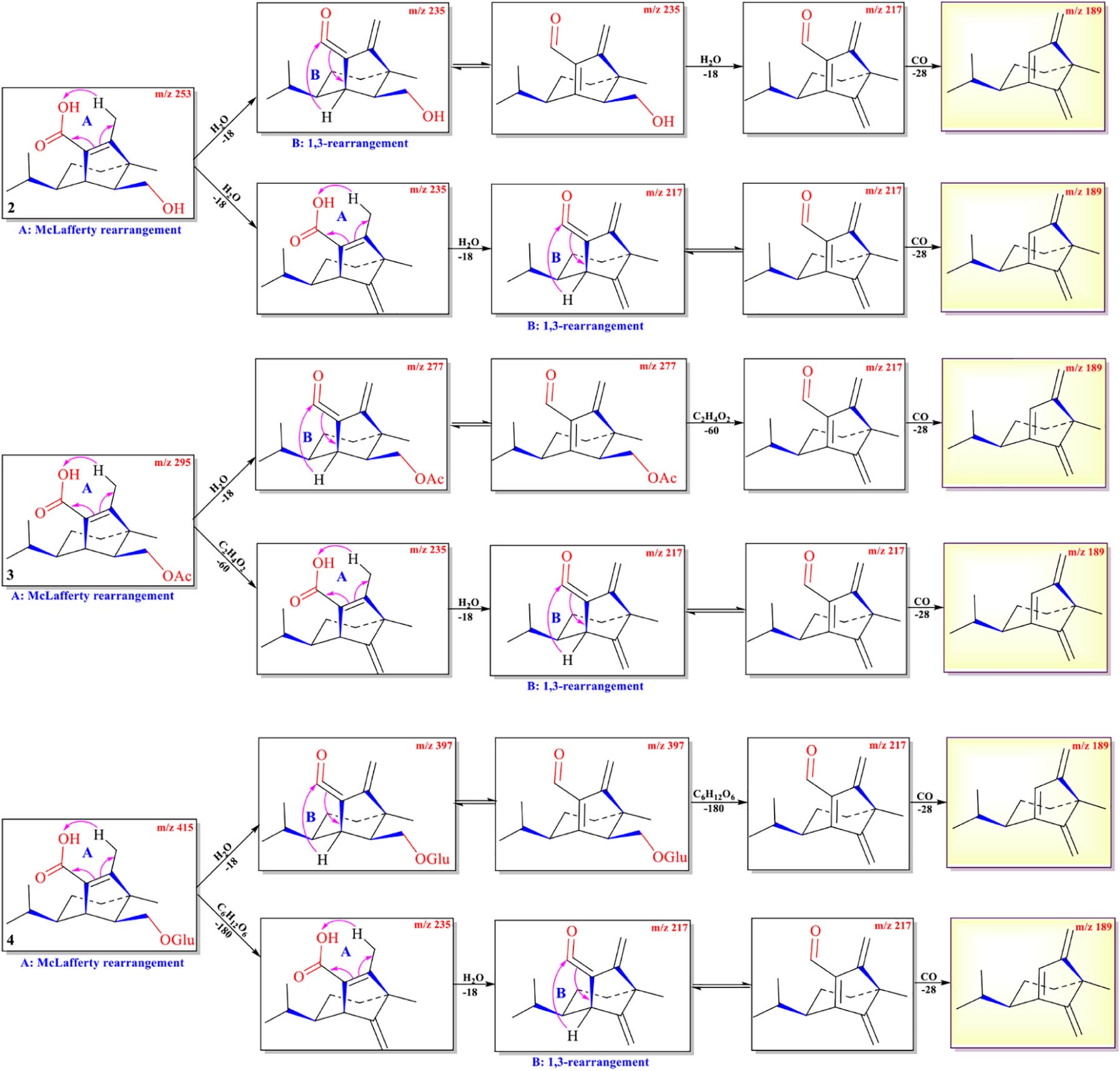
The protonated parent ions of 5 and 6 were m/z 401 [M+H]+ and m/z 443 [M+H]+, respectively, in the UPLC-Q-TOF-MS/MS spectra. These two ions lost one molecule of glucose (–180) by the McLafferty rearrangement to form the intermediate ions m/z 221 and m/z 263, respectively, which yielded the same diagnostic ion m/z 203 through neutral loss of one molecule of H2O (−18) and one molecule of CH3COOH (−60) (Scheme 3 and Supplementary Figures 5, 6). The high-resolution mass and fragment ions together with the elemental constituents of compounds 5 and 6 were listed in Table 4 and Supplementary Table 3.
Scheme 3. Possible fragmentation pathways of 5 and 6.

Table 4. Elemental constituents of major product ions from [M+Na]+ for compound 5 (subtype C).
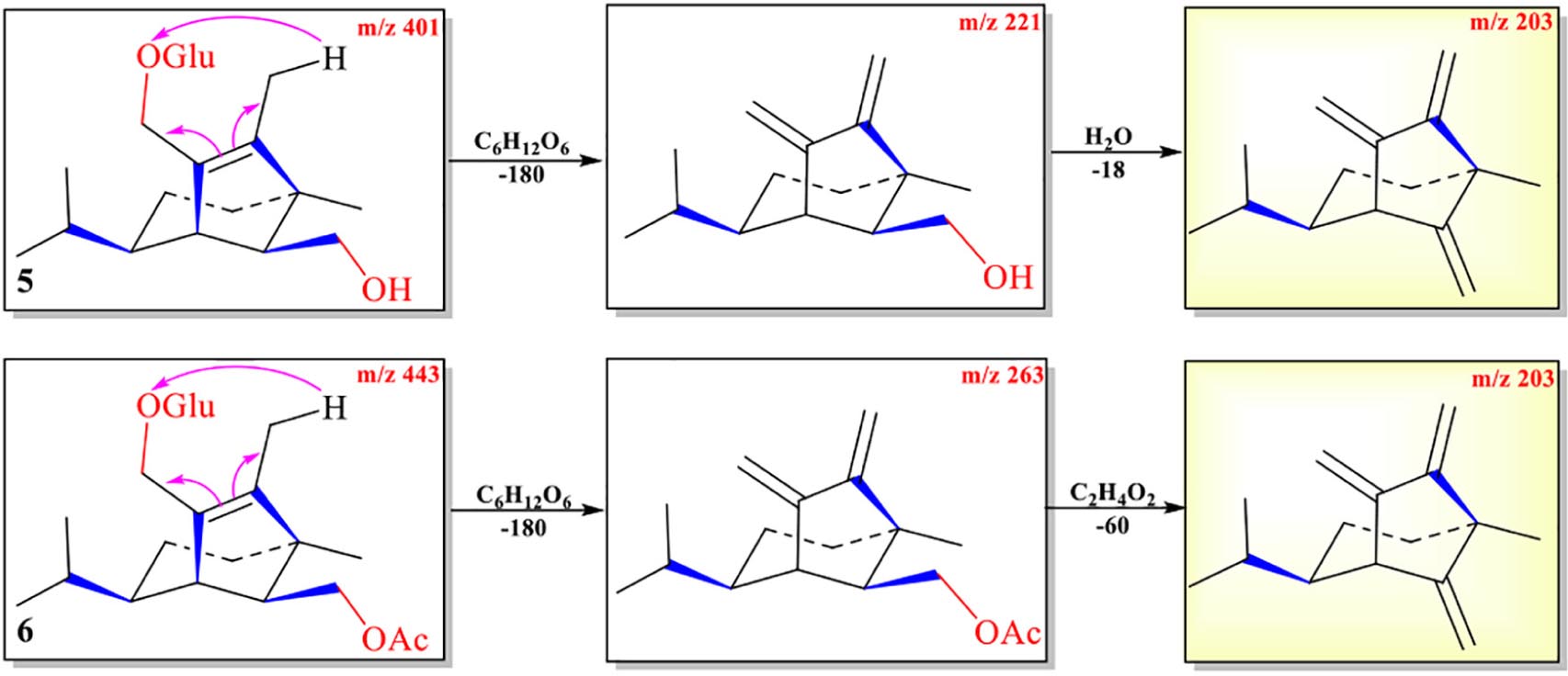
The abundance of protonated parent ion of 7 m/z 225 [M+H]+ was relatively low in the UPLC-Q-TOF-MS/MS spectra. It might be the neutral loss of one molecule of H2O (−18) and one molecule of CH3CO2H (−60) to form an intermediate ion m/z 207. The abundance of the ion (m/z 207) was the highest in the mass profile. The diagnostic ion (m/z 189) was formed from ion m/z 207 by loss of one molecule of H2O (−18) through the McLafferty rearrangement (Scheme 4 and Supplementary Figure 7). The high-resolution mass and fragment ions together with the elemental constituents of compound 7 are listed in Table 5.
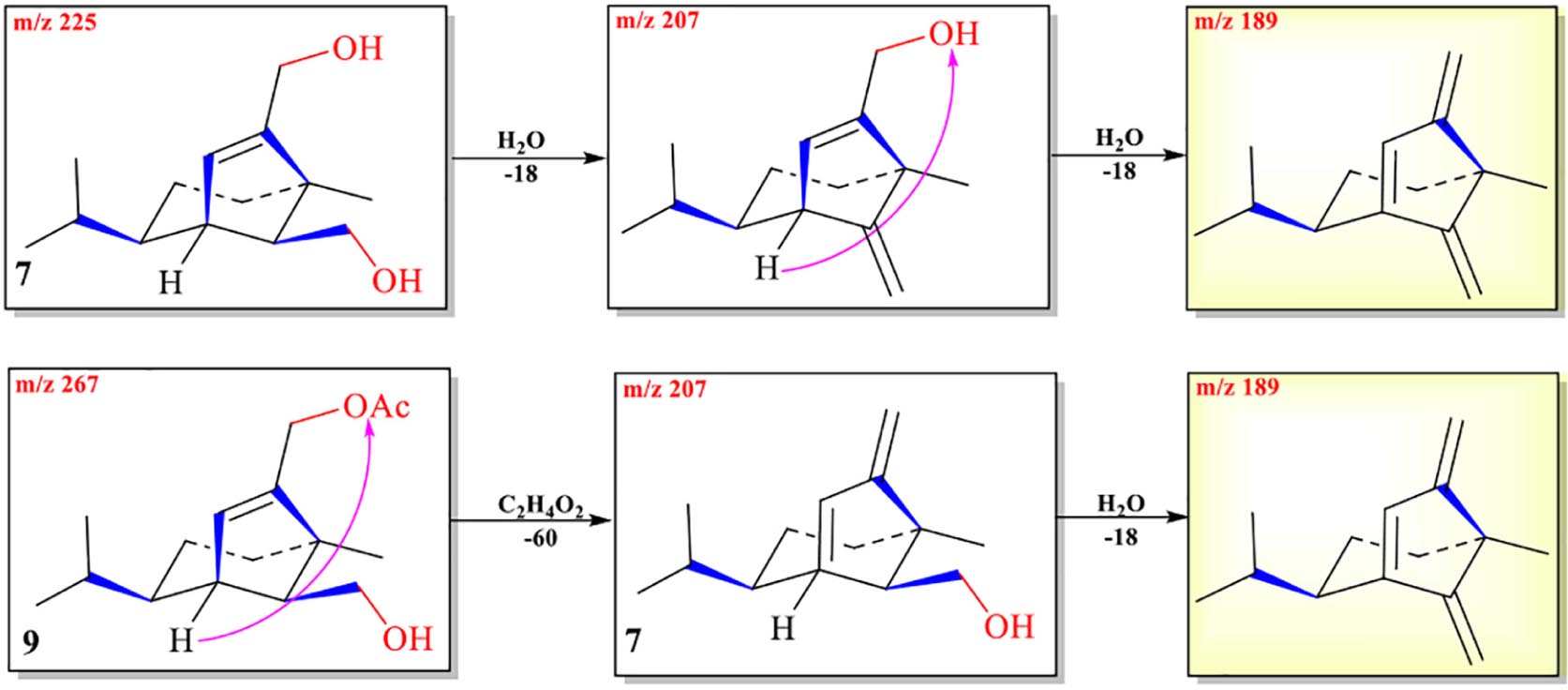
Scheme 4. Possible fragmentation pathways of 7 and 9.
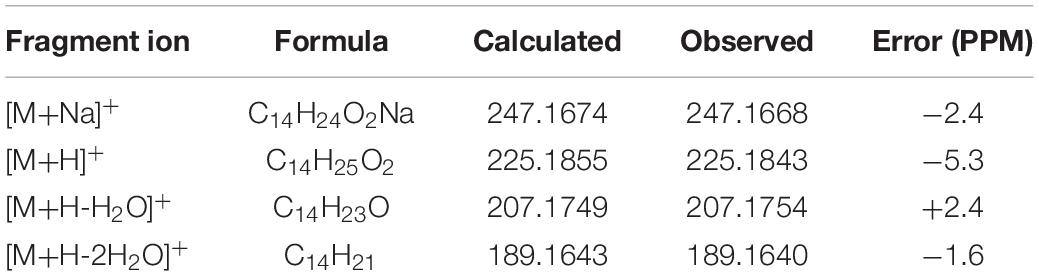
Table 5. Elemental constituents of major product ions from [M+Na]+ for compound 7 (subtype D).
The protonated parent ion of 8 was m/z 251 [M+H]+ in the UPLC-Q-TOF-MS/MS spectra with the highest abundance. The McLafferty rearrangement produced the intermediate ion m/z 233 from m/z 251 by loss of one molecule of H2O (−18). Successive neutral losses of one molecule of H2O (−18) and one molecule of CO (−28) or vice versa yielded the diagnostic daughter ion (m/z 187). The detailed MS analysis is shown in Scheme 5 and Supplementary Figure 8. The high-resolution mass and fragment ions together with the elemental constituents of compound 8 are listed in Table 6.
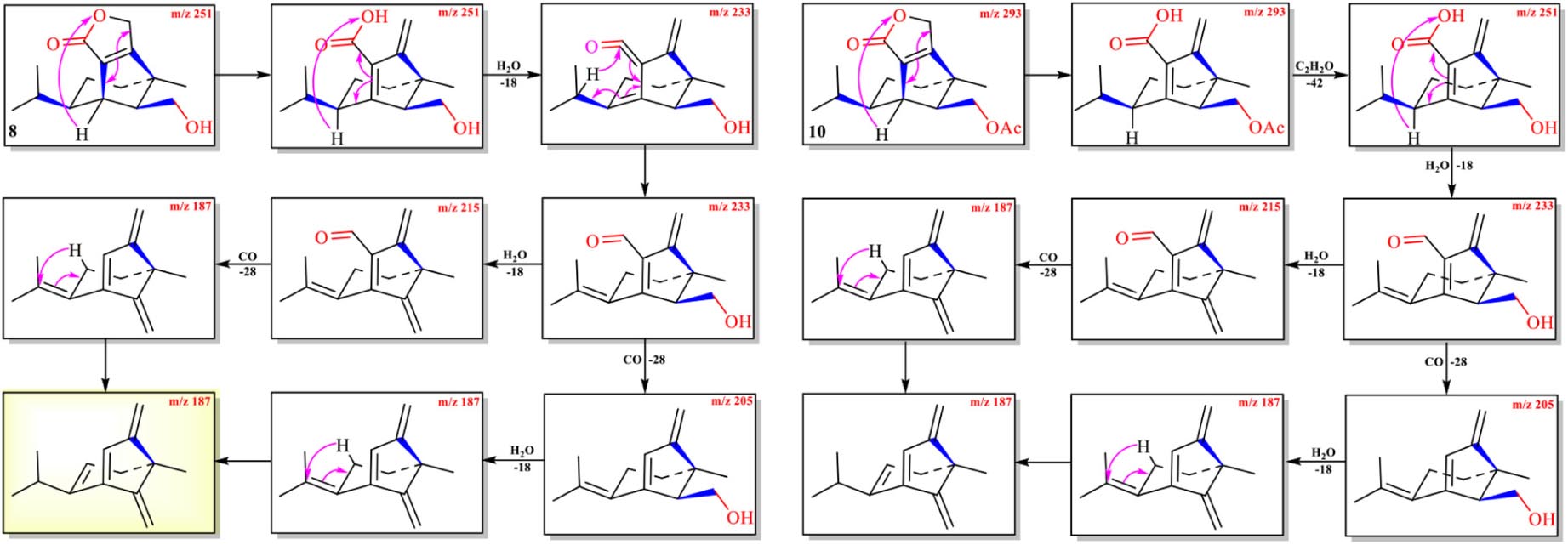
Scheme 5. Possible fragmentation pathways of 8 and 10.
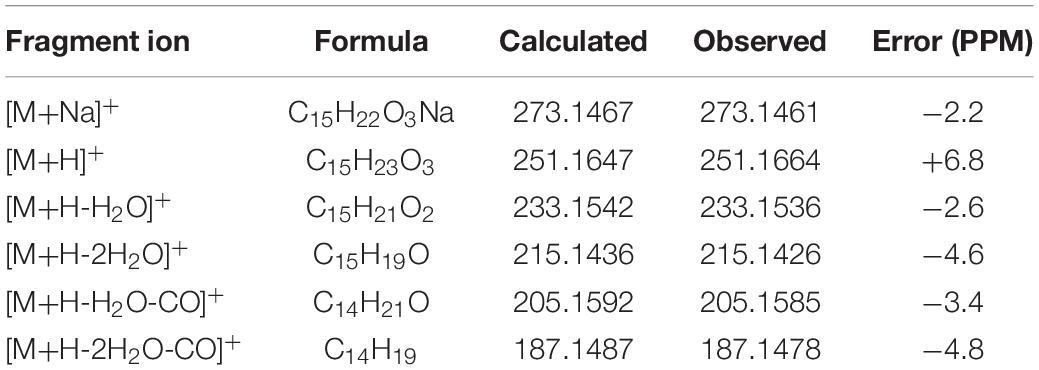
Table 6. Elemental constituents of major product ions from [M+Na]+ for compound 8 (subtype E).
With the UPLC-Q-TOF-MS/MS fragmentation mechanisms of 1–8 in hand, it implied that each subtype seco-sativene sesquiterpenoids had a diagnostic daughter ion in the MS profile (subtype A → m/z 175; subtypes B/D → m/z 189; subtype C → m/z 203; subtype E → m/z 187). Though both subtypes B and D had the same diagnostic daughter ion m/z 189, the last cleavage in the sub-type B was the neutral loss of one molecule of CO (−28), whereas the neutral loss of one molecule of H2O (−18) was the last cleavage in subtype D, which differentiated these two subtypes B and D. Thus, it could give the possible subtype of seco-sativene sesquiterpenoids based on the diagnostic daughter ion from the corresponding ESI-Q-TOF-MS/MS data.
Then, the crude extract of the ethyl acetate fraction of the plant pathogen Bipolaris sorokiniana was then analyzed by UPLC-Q-TOF-MS/MS (Supplementary Figure 11). There were some peaks (compound 8: m/z 251.1664, tR = 3.53 min, λmax = 217 nm; compound 7: m/z 225.1843, tR = 3.56 min, λmax = 206 nm; compound 2: m/z 253.1797, tR = 4.86 min, λmax = 244 nm; compound 1: m/z 249.1548, tR = 4.93 min, λmax = 248 nm; compound 3: m/z 295.1900, tR = 8.24 min, λmax = 243 nm) in the TOF MS profiles possessing typical fragment ions including m/z 175, 187, 189, 203, 205, 207, 215, 217, 231, 233, and 235. However, there are other unidentified compounds (251.1639, tR = 2.85 min; 353.0418, tR = 5.78 min; 237.1855, tR = 5.88 min; 293.1748, tR = 7.03 min; 207.1744, tR = 7.26 min; 235.1693, tR = 7.86 min; 235.1693, tR = 8.68 min; 235.1692, tR = 8.85 min) that had similar cleavage fragments as compounds 1–8, indicating that many known and undescribed analogues existed in the extract.
Compounds 1 (tR = 4.93 min) and 2 (tR = 4.86 min) as the main constituents were isolated from the plant pathogen Bipolaris sorokiniana crude extract.
Two ions (tR = 7.23, 7.01 min) at m/z 267 [M+1]+ (9) and m/z 293 [M+1]+ (10) were detected in the crude extract, and their molecular weights were determined to be C16H26O3 and C17H24O4, respectively (Supplementary Figures 9, 10). The diagnostic ions of 9 and 10 were m/z 189/187 and λmax = 234/215 nm, respectively, implying that structures of 9 and 10 possessed the same subtypes as 7 and 8. The UPLC-Q-TOF-MS/MS fragmentation pathways of these two compounds were nearly the same as those of 7 and 8, except for an additional acetyl group. To confirm this hypothesis, 9 and 10 were isolated from the extract and elucidated by IR, NMR, and HR-ESI-MS spectra. The IR absorption bands at 3,420 cm–1 showed the hydroxyl group in 9, and 2,931 and 1,747 cm–1 revealed the presence of alkyl and ester moieties, respectively, which were also present in that of 10. The 1H-NMR spectrum revealed the similarity of 9/10 with 7/8, except that the chemical shift values of –CH2-12 of 9 and –CH2-14 of 10 were down-fielded in 9/10 (Osterhage et al., 2002) and an additional methyl signal was observed in the 1H-NMR spectrum of 9/10. This implied that the additional acetyl group was connected at C-12 in 9 and C-14 in 10. The key HMBC correlations from –CH2-12/CH2-14 and 1′-Me to C-2′ (δC 171.2 in 9/10) supported the conclusion (Figure 2 and Supplementary Figures 12–29). Thus, the structures of 9 and 10 were determined, and their ESI-Q-TOF-MS/MS fragmentation pathways were consistent with those of 7/8 (Schemes 4, 5 and Supplementary Tables 4, 5).
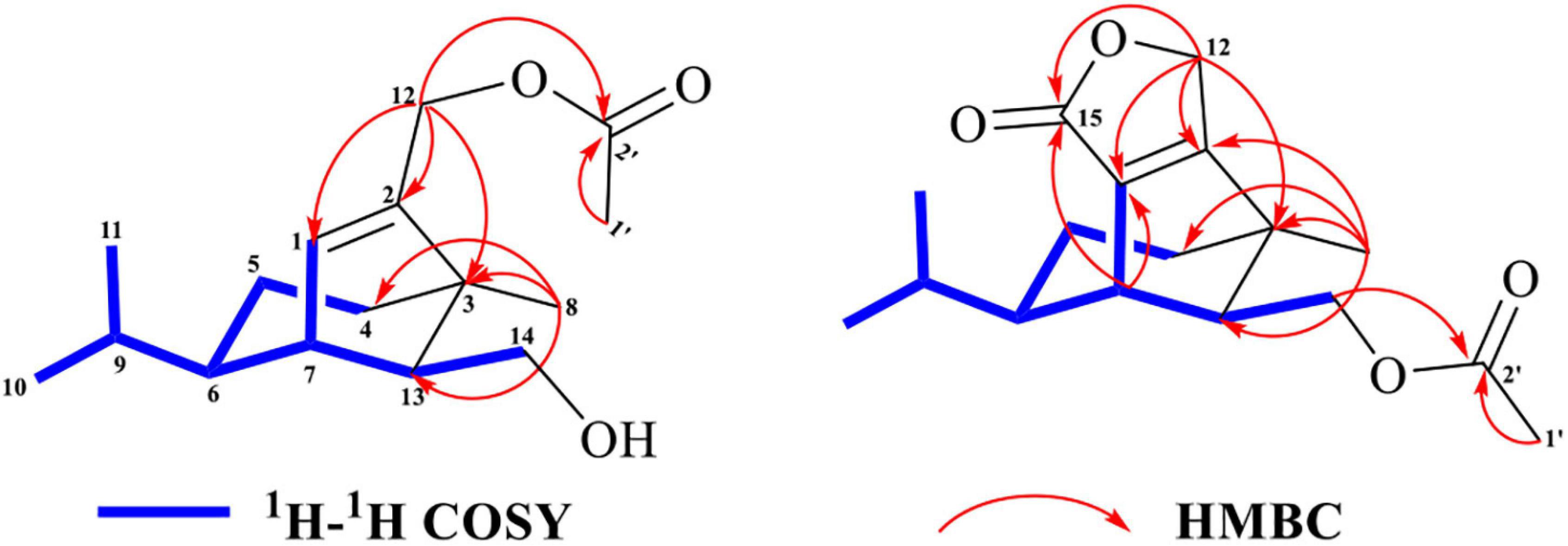
Figure 2. Key 2D-NMR correlations of 9 and 10.
The antioxidant activities of compounds 1 and 2 were evaluated by the DPPH and ABTS+ free radical scavenging test, and the results were presented as IC50 values. The results demonstrated that compound 1 displayed moderate antioxidant activities with IC50 of 0.90 and 1.97 mM for DPPH and ABTS+ scavenging capacity, respectively. However, compound 2 did not show the obvious antioxidant activity. They were measured by comparing the scavenging ability of DPPH free radical and ABTS+ free radical with VC, a well-known potent antioxidant and free radical scavenger with IC50 of 0.14 and 0.42 mM for DPPH and ABTS+ scavenging capacity, respectively (Table 7).

Table 7. DPPH and ABTS+ scavenging capacity of compounds 1, 2, 9, and 10.
This report analyzed the fragmentation patterns of eight representative seco-sativene sesquiterpenoids (1–8) using UPLC-Q-TOF-MS/MS, and McLafferty rearrangement, 1,3-rearrangement, and neural loss (−18, −28) were the main fragmentation patterns. The results indicated that dehydration (−18) occurred easily in seco-sativenes with strong abundance of dehydration peak observed in 1-8, and similar reports were found in other studies (McLafferty and Gohike, 1959; Alén, 1987; Nawamaki and Kuroyanagi, 1996; Sun et al., 2012; Giri et al., 2017; Qian et al., 2018). This may be due to an electron impact inducing fragmentations of alkene monocarboxylic acids to form an active OH ion, which combines an available methyl-hydrogen atom to one lost molecule of H2O (−18) through the McLafferty rearrangement in a six-membered system (Baldas et al., 1969; Alexander et al., 1972). The molecular ion peak [M+H]+ of 1–7 and 9 was not easily observed in the UPLC-Q-TOF-MS/MS profile, whereas [M+Na]+ and [M+H-H2O]+ abundances of these compounds were relatively strong. As there was a characteristic 40 Da difference between [M+Na]+ and [M+H-H2O]+, the molecular ion peak of 1–7 and 9 can be inferred. Compounds 8 and 10 possess a lactone ring at C-1 and C-2, which is different from 1–7 and 9. The special group in 8 and 10 leads to their molecular ion peaks [M+H]+ easily being observed in MS profiles. This might be a key signal for differentiating subtype E from other subtypes. Although the McLafferty rearrangement together with 1,3-rearrangement in alkene monocarboxylic molecules to produce a base peak of dehydration has been reported, this was the first report in seco-sativene sesquiterpenoids mass analysis, which provided a base for the seco-sativenes structural elucidation.
Diagnostic daughter ions of five subtypes of seco-sativene sesquiterpenoids were provided base on UPLC-Q-TOF-MS/MS analysis in this report. Subtypes A, B/D, and E possessed diagnostic daughter ions at m/z 175, 189, and 203, respectively, whereas subtype C showed a characteristic daughter ion at m/z 187 in the UPLC-Q-TOF-MS/MS profiles. The main difference between subtypes B and D was that the last cleavage was the neutral loss of one molecule of CO (−28) in subtype B, not the neutral loss of one molecule of H2O (−18) in subtype D. Diagnostic ions provided signals for the different subtypes of seco-sativenes, and the rearrangement and neutral loss (H2O, CO, and HOAc) in the fragmentation patterns provided the possible groups on the structures of seco-sativene sesquiterpenoids. Thus, the structures of the seco-sativenes could be inferred by fragmentation patterns combined with the diagnostic ions and molecular formula based on the UPLC-Q-TOF-MS/MS profile. This report provides a reliable method for the structural analysis of seco-sativene sesquiterpenoids.
Compounds 1, 2, 9, and 10 and other seco-sativenes are a class of phytotoxins. Compound 1 was previously isolated from C. sativus (teleomorph: B. sorokiniana) without phytotoxicity on corn leaves, but helminthosporal acid with an aldehyde group at C-1 (a carboxyl group at C-1 in 1) possessed strong phytotoxic activity (Li et al., 2020a). Therefore, the aldehyde group in helminthosporal acid might be a potential active group for phytotoxicity. Compound 2 exhibited bidirectional regulation activities. On the one hand, it was the gibberellin-like plant growth regulator, which could promote the growth of plant roots and leaves at low concentrations (Qader et al., 2017; Li et al., 2020a). On the other hand, it showed phytotoxicity on corn leaves (Li et al., 2020a). Compounds 9 and 10 did not show obvious antioxidant activity in this report, and no more activities of 9 and 10 were tested due to limited amounts. Compared with 7 and 8, 9 and 10 possess an extra acetyl group at 12-OH and 5-OH, respectively. Osterhage et al. reported that 7 showed inhibitory activity against tyrosine kinase p56, Microbotryum violaceum, Eurotium repens, and Escherichia coli (Osterhage et al., 2002) and that 8 showed antifungal activity against M. violaceum (Osterhage et al., 2002) and phytotoxicity on corn leaves (Li et al., 2020a). Several reports suggested that 5-CH2OH might be the potent active group of seco-sativenes sesquiterpenoids for their phytotoxicity (Nakajima et al., 1994; Li et al., 2020b). Therefore, 9 might possess phytotoxicity against barley seeds and corn leaves. But their structure–activity relationships (SARs) need to be further studied. At present, the reports on the activities of seco-sativene sesquiterpenoids mainly focused on phytotoxicity and growth-promoting effects with few reports about other activities (Li et al., 2020b). To further explore the medicinal value of seco-sativenes, the antioxidant activity of these compounds (1, 2, 9, and 10) were studied in this report. Only then did compound 1 show moderate activity on DPPH scavenging capacity, which indicated that 13-COOH might be a possible active group, and further biological exploration should be needed in the future.
UPLC-Q-TOF-MS/MS spectrometry has evolved to be a mature and common technique, which is now widely used to analyze secondary metabolites from diverse biological resources. Most of researchers used this technology to identify (new/novel) metabolites or dereplicate in different crude extracts (Jin et al., 2018; Wang et al., 2020). Recently, a molecular networking technique based on (U)HPLC-MS/MS combined with different databases was used in the dereplication and targeting of new natural products from diverse biological resources (Watrous et al., 2012; Yang et al., 2013; Aksenov et al., 2017; Hou et al., 2019; Ramos et al., 2019; Rivera-Chaìvez et al., 2019; Shi et al., 2019; Wu et al., 2019; Chao et al., 2020; Zang et al., 2020; Lei et al., 2021; Lin et al., 2021). The molecular networking technique used the known or new compound as the “seed” to realize the visualization of analogues. In the network, MS data were collected from LC-MS and uploaded to the GNPS database for data processing to produce total molecular network profiles. Every node in the same network represented a compound possessing the same core skeleton. Known or new analogues can be quickly inferred according to molecular weight, molecular formula, and fragmentation patterns based on node analysis through searching different databases or house libraries. The targeted isolation of the seed analogues could be realized by searching the location of the “seed” (Klitgaard et al., 2015; Allard et al., 2016; Trautman and Crawford, 2016; Naman et al., 2017; Olivon et al., 2017a,b; Nothias et al., 2018). When a molecular network is combined with fragmentation patterns, the range of metabolites would be narrowed, and the precision of targeted-isolation-compounds would be improved (He et al., 2021). Thus, molecular networking based on the (U)HPLC-MS/MS technique would provide a more convenient approach for dereplication and targeting-isolation of new seco-sativene sesquiterpenoids in the future.
Eight seco-sativene sesquiterpenoids (1–8) were analyzed using the UPLC-Q-TOF-MS/MS technique in positive mode, from which their possible mass fragmentation patterns were suggested, and neural loss, McLafferty rearrangement, and 1,3-rearrangement were the main clearage patterns. These eight seco-sativene sesquiterpenoids (1–8) were summarized to be five subtypes according to their structural features. Each subtype possessed a diagnostic daughter ion, which, in return, could contribute to the elucidation of seco-sativene sesquiterpenoids. Based on the fragmentation mechanism mentioned above, some analogues including two potentially new ones were detected. Two known (1 and 2) and two new analogues (9 and 10) were then isolated from the extract of the plant pathogen Bipolaris sorokiniana. Their structures were elucidated mainly by NMR spectra and supported based on their UPLC-Q-TOF-MS/MS analysis. The results demonstrated that diagnostic mass ions of seco-sativene sesquiterpenoids in the UPLC-Q-TOF-MS/MS profiles provided a convenient and high-performance approach for structural characterization and also support that UPLC-Q-TOF-MS/MS is a powerful and sensitive tool for dereplication and detection of new analogues in crude extracts. This study will pave the way for the structural analysis and targeting isolation of seco-sativene sesquiterpenoids in different fungal crude extracts.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Y-DW, JY, and GD: experiment and writing – review and editing. Y-DW, JY, Y-YL, X-MT, and QL: data collection. S-YY and S-BN: activity experiment. HD: resources. JY, GD, and L-PG: funding acquisition. GD and Y-DW: writing – original draft preparation. All authors have read and agreed to the published version of the manuscript.
We gratefully acknowledge financial support from the National Natural Science Foundation of China (81891014), the National Key R&D Programme of China (2017YFC1700701), the Key Project at Central Government Level: Establishment of the sustainable use for valuable Chinese Medicine Resources (2060302), and the Fundamental Research Funds for the Central public welfare research institutes (ZZXT201906).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We gratefully acknowledge Jin-wei Ren for kindly helping to measure the NMR spectra.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.807014/full#supplementary-material
Ahad, H., Jin, H., and Liu, Y. (2020). Chemical profiling of spermidines in goji berry by strong cation exchange solid-phase extraction (SCX-SPE) combined with ultrahigh-performance liquid chromatography-quadrupole time-of-flight mass spectrometry (UPLC-Q-TOF/MS/MS). J. Chromatogr. B. 1137, 121923. doi: 10.1016/j.jchromb.2019.121923
Aksenov, A. A., da Silva, R., and Knight, R. (2017). Global chemical analysis of biology by mass spectrometry. Nat. Rev. Chem. 1, 1–20.
Alén, R. (1987). Formation of acid-catalysed dehydration products from xyloisosaccharinic acid. Acta chem. Scand. Ser. B. Org. Chem. Biochem. 41, 76–78.
Alexander, R. G., Bigley, D. B., and Todd, J. F. J. (1972). Mass spectral studies of unsaturated carboxylic acids-I: some cyclic and acyclic βγ−unsaturated acids. Org. Mass Spectrom. 6, 1153–1160.
Ali, L., Khan, A. L., Hussain, J., Al-Harrasi, A., Waqas, M., Kang, S. M., et al. (2016). Sorokiniol: a new enzymes inhibitory metabolite from fungal endophyte Bipolaris sorokiniana LK12. BMC Microbiol. 16:103. doi: 10.1186/s12866-016-0722-7
Allard, P. M., Peìresse, T., Bisson, J., Gindro, K., Marcourt, L., Pham, V. C., et al. (2016). Integration of molecular networking and in-silico MS/MS fragmentation for natural products dereplication. Anal. Chem. 88, 3317–3323. doi: 10.1021/acs.analchem.5b04804
Baldas, J., Porter, Q. N., and Ramsay, C. C. R. (1969). Mass spectrometric studies. V. Benzylidenemalonic acid and related compounds. Aust. J. Chem. 22, 405–422.
Briggs, D. E. (1966). Gibberellin-like activity of helminthosporol and helminthosporic acid. Nature. 210, 418–419.
Chao, R., Hou, X. M., Xu, W. F., Hai, Y., Wei, M. Y., Wang, C. Y., et al. (2020). Targeted Isolation of Asperheptatides from a coral-derived fngus using LC-MS/MS-based molecular networking and antitubercular activities of modified cnnamate derivatives. J. Nat. Prod. 84, 11–19. doi: 10.1021/acs.jnatprod.0c00804
Chen, C., Xue, Y., and Li, Q. M. (2018). Neutral loss scan-based strategy for integrated identification of amorfrutin derivatives, new peroxisome proliferator-activated receptor gamma agonists, from Amorpha Fruticosa by UPLC-QqQ-MS/MS and UPLC-Q-TOF-MS. J. Am. Soc. Mass Spectrom. 29, 685–693. doi: 10.1007/s13361-018-1891-4
Conceição, R. S., Reis, I. M. A., Cerqueira, A. P. M., Perez, C. J., Junior, M. C. D. S., Branco, A., et al. (2020). Rapid structural characterisation of benzylisoquinoline and aporphine alkaloids from Ocotea spixiana acaricide extract by HPTLC-DESI-MSn. Phytochem. Analysis. 31, 711–721. doi: 10.1002/pca.2935
Giri, A., Coutriade, M., and Racaud, A. (2017). Molecular characterization of volatiles and petrochemical base oils by photo-ionization GC× GC-TOF-MS. Anal. Chem. 89, 5395–5403. doi: 10.1021/acs.analchem.7b00124
Hashimoto, T., Sakurai, A., and Tamura, S. (1967). Physiological activities of helminthosporol and helminthosporic acid: I. Effects on growth of intact plants. Plant Cell Physiol. 8, 23–34.
He, Q. F., Wu, Z. L., Li, L., Sun, W. Y., Wang, G. Y., Jiang, R. W., et al. (2021). Discovery of neuritogenic securinega alkaloids from Flueggea suffruticosa by a building blocks-based molecular network strategy. Angew. Chem. Int. Ed. 60, 19609–19613. doi: 10.1002/anie.202103878
Hou, X. M., Li, Y. Y., Shi, Y. W., Fang, Y. W., Chao, R., Gu, Y. C., et al. (2019). Integrating molecular networking and 1H NMR to target the isolation of chrysogeamides from a library of marine-derived Penicillium fungi. J. Org. Chem. 84, 1228–1237. doi: 10.1021/acs.joc.8b02614
Jin, M., Zhang, W., Jiang, H., Du, Y. F., Guo, W., Cao, L., et al. (2018). UPLC-Q-TOF-MS/MS-guided dereplication of Pulsatilla chinensis to identify triterpenoid saponins. Phytochem. Analysis. 29, 516–527. doi: 10.1002/pca.2762
Katsumi, M., Tamura, S., and Sakurai, A. (1967). Gibberellin-like activity of helminthosporol, helminthosporic acid and dihydrohelminthosporic acid in leaf sheath elongation of dwarf 5 mutants of Zea mays. Naturwissenschaften. 54, 96–97.
Klitgaard, A., Nielsen, J. B., and Frandsen, R. J. (2015). Combining stable isotope labeling and molecular networking for biosynthetic pathway characterization. Anal. Chem. 87, 6520–6526. doi: 10.1021/acs.analchem.5b01934
Lei, H., Zhang, Y., Zu, X., Ye, J., Liang, Y., Cheng, T., et al. (2021). Comprehensive profiling of the chemical components and potential markers in raw and processed Cistanche tubulosa by combining ultra-high-performance liquid chromatography coupled with tandem mass spectrometry and MS/MS-based molecular networking. Anal. Bioanal. Chem. 413, 129–139. doi: 10.1007/s00216-020-02983-0
Li, Y. Y., Tan, X. M., Wang, Y. D., Yang, J., Zhang, Y. G., Sun, B. D., et al. (2020a). Bioactive seco-sativene sesquiterpenoids from an Artemisia desertorum endophytic fungus, Cochliobolus sativus. J. Nat. Prod. 85, 1488–1494. doi: 10.1021/acs.jnatprod.9b01148
Li, Y. Y., Tan, X. M., Yang, J., Guo, L. P., and Ding, G. (2020b). Naturally Occurring seco-sativene sesquiterpenoid: chemistry and biology. J. Agric. Food Chem. 68, 9827–9838. doi: 10.1021/acs.jafc.0c04560
Liang, Y., Yan, G. Y., Wu, J. L., Zong, X., Liu, Z., Zhou, H., et al. (2018). Qualitative and quantitative analysis of lipo-alkaloids and fatty acids in Aconitum carmichaelii using LC-MS and GC-MS. Phytochem. Analysis. 29, 398–405. doi: 10.1002/pca.2760
Lin, X., Chai, L., Zhu, H. R., Zhou, Y., Shen, Y., Chen, K. H., et al. (2021). Applying molecular networking for targeted isolation of depsipeptides. RSC Advances 11, 2774–2782. doi: 10.1039/d0ra09388b
Ludwig, R. A. (1957). Toxin production by Helminthosporium sativum and its significance in disease development. Can. J. Bot. 35, 291–303. doi: 10.1139/b57-026
Ludwig, R. A., Clark, R. V., and Julien, J. B. (1956). Studies on the seedling disease of barley caused by Helminthosporium sativum. Can. J. Bot. 34, 653–673. doi: 10.1139/b56-048
Mayo, P. D., and Williams, R. E. (1965). Sativene, parent of the toxin from Helminthosporium sativum. J. Am. Chem. Soc. 87, 3275. doi: 10.1021/ja01092a066
Mayo, P. D., Robinson, J. R., and Spencer, E. Y. (1962a). The biogenesis of helminthosporal. Experientia. 18, 359–360. doi: 10.1007/bf02172248
Mayo, P. D., Spencer, E. Y., and White, R. W. (1961). Helminthosporal, the toxin from Helminthosporium sativum: I. Isolation and characterization. Can. J. Chem. 39, 1608–1612. doi: 10.1139/v61-205
Mayo, P. D., Spencer, E. Y., and White, R. W. (1962b). The constitution of helminthosporal. J. Am. Chem. Soc. 84, 494–495. doi: 10.1021/ja00862a037
Mayo, P. D., Spencer, E. Y., and White, R. W. (1963). Terpenoids: IV. The structure and stereochemistry of helminthosporal. Can. J. Chem. 41, 2996–3004. doi: 10.1139/v63-440
Mayo, P. D., Williams, R. E., and Spencer, E. Y. (1965). Terpenoids: VIII. The immediate precursors of helminthosporal and helminthosporol. Can. J. Chem. 43, 1357–1365. doi: 10.1139/v65-184
McLafferty, F. W., and Gohike, R. S. (1959). Mass spectrometric analysis. Aromatic acids and esters. Anal. Chem. 31, 2076–2082. doi: 10.1021/ac60156a062
Miyazaki, S., Jiang, K., Kobayashi, M., Asami, T., and Nakajima, M. (2017). Helminthosporic acid functions as an agonist for gibberellin receptor. Biosci. Biotechnol. Biochem. 81, 2152–2159. doi: 10.1080/09168451.2017.1381018
Miyazaki, S., Tomita, K., Yamane, H., Kobayashi, M., Asami, T., and Nakajima, M. (2018). Characterization of a helminthosporic acid analog that is a selective agonist of gibberellin receptor. Bioorg. Med. Chem. Lett. 28, 2465–2470. doi: 10.1016/j.bmcl.2018.06.005
Nakajima, H., Isomi, K., and Hamasaki, T. (1994). Sorokinianin: a novel phytotoxin produced by the phytopathogenic fungus Bipolaris sorokiniana. Tetrahedron Lett. 35, 9597–9600. doi: 10.1016/0040-4039(94)88520-6
Naman, C. B., Rattan, R., and Nikoulina, S. E. (2017). Integrating molecular networking and biological assays to target the isolation of a cytotoxic cyclic octapeptide, samoamide A, from an American Samoan marine cyanobacterium. J. Nat. Prod. 80, 625–633. doi: 10.1021/acs.jnatprod.6b00907
Nawamaki, K., and Kuroyanagi, M. (1996). Sesquiterpenoids from Acorus calamus as germination inhibitors. Phytochem. 43, 1175–1182. doi: 10.1016/s0031-9422(96)00401-3
Nihashi, Y., Lim, C. H., Tanaka, C., Miyagawa, H., and Ueno, T. (2002). Phytotoxic sesterterpene, 11-epiterpestacin, from Bipolaris sorokiniana NSDR-011. Biosci. Biotechnol. Biochem. 66, 685–688. doi: 10.1271/bbb.66.685
Nothias, L. F., Nothias-Esposito, M., and da Silva, R. (2018). Bioactivity-based molecular networking for the discovery of drug leads in natural product bioassay-guided fractionation. J. Nat. Prod. 81, 758–767. doi: 10.1021/acs.jnatprod.7b00737
Nukina, M., Hattori, H., and Marumo, S. (1975). Cis-Sativenediol, a plant growth promotor, produced by fungi. J. Am. Chem. Soc. 97, 2542–2543.
Olivon, F., Allard, P. M., Koval, A., Righi, D., Genta-Jouve, G., Neyts, J., et al. (2017a). Bioactive natural products prioritization using massive multi-informational molecular networks. ACS Chem. Biol. 12, 2644–2651. doi: 10.1021/acschembio.7b00413
Olivon, F., Grelier, G., Roussi, F., Litaudon, M., and Touboul, D. (2017b). MZmine 2 data-preprocessing to enhance molecular networking reliability. Anal. Chem. 89, 7836–7840. doi: 10.1021/acs.analchem.7b01563
Osterhage, C., König, G. M., and Höller, U. (2002). Rare sesquiterpenes from the algicolous fungus Drechslera dematioidea. J. Nat. Prod. 65, 306–313. doi: 10.1021/np010092l
Pena-Rodriguez, L. M., and Chilton, W. S. (1989). Victoxinine and prehelminthosporolactone, two minor phytotoxic metabolites produced by Bipolaris sp., a pathogen of Johnson grass. J. Nat. Prod. 52, 899–901. doi: 10.1021/np50064a046
Pena-Rodriguez, L. M., Armingeon, N. A., and Chilton, W. S. (1988). Toxins from weed pathogens, I. Phytotoxins from a bipolaris pathogen of johnson grass. J. Nat. Prod. 51, 821–828. doi: 10.1021/np50059a001
Phan, C. S., Li, H., Kessler, S., Solomon, P. S., Piggott, A. M., and Chooi, Y. H. (2019). Bipolenins K–N: New sesquiterpenoids from the fungal plant pathogen Bipolaris sorokiniana. Beilstein. J. Org. Chem. 15, 2020–2028. doi: 10.3762/bjoc.15.198
Qader, M. M., Kumar, N. S., Jayasinghe, L., Araya, H., and Fujimoto, Y. (2017). Bioactive sesquiterpenes from an endophytic fungus Bipolaris sorokiniana isolated from a popular medicinal plant Costus speciosus. Mycology 8, 17–20.
Qian,H., Yu, F. J., and Lu, D. Y. (2018). Identification of poliumoside metabolites in rat plasma, urine, bile, and intestinal bacteria with UPLC/Q-TOF-MS. Chin. J. Nat. Med. 16, 871–880. doi: 10.1016/S1875-5364(18)30129-8
Ramos, A. E. F., Evanno, L., Poupon, E., Champy, P., and Beniddir, M. A. (2019). Natural products targeting strategies involving molecular networking: different manners, one goal. Nat. Prod. Rep. 36, 960–980. doi: 10.1039/c9np00006b
Rivera-Chaìvez, J., Zacatenco-Abarca, J., and Morales-Jimeìnez, J. (2019). Cuautepestalorin, a 7, 8-dihydrochromene–oxoisochromane adduct bearing a hexacyclic scaffold from Pestalotiopsis sp. IQ-011. Org. lett. 21, 3558–3562. doi: 10.1021/acs.orglett.9b00962
Scupinari, T., Mannochio Russo, H., and Sabino Ferrari, A. B. (2020). Crotalaria spectabilis as a source of pyrrolizidine alkaloids and phenolic compounds: HPLC-MS/MS dereplication and monocrotaline quantification of seed and leaf extracts. Phytochem. Analysis. 31, 747–755. doi: 10.1002/pca.2938
Shi, Y., Gu, R., Li, Y., Wang, X., Ren, W., and Li, X. (2019). Exploring novel herbicidin analogues by transcriptional regulator overexpression and MS/MS molecular networking. Microb. Cell Factories 18, 1–17. doi: 10.1186/s12934-019-1225-7
Spencer, E. J. (1965). World review of pest control. Bulletin of the Entomological Society of America 4, 75.
Sun, B. S., Xu, M. Y., and Li, Z. (2012). UPLC-Q-TOF-MS/MS analysis for steaming times-dependent profiling of steamed Panax quinquefolius and its ginsenosides transformations induced by repetitious steaming. J. Ginseng. Res. 36, 277. doi: 10.5142/jgr.2012.36.3.277
Taniguchi, E., and White, G. A. (1967). Site of action of the phytotoxin, helminthosporal. Biochem. Biophys. Res. Commun. 28, 879–885. doi: 10.1016/0006-291x(67)90060-5
Trautman, E., and Crawford, M. (2016). Linking biosynthetic gene clusters to their metabolites via pathway-targeted molecular networking. Curr. Top. Med. Chem. 16, 1705–1716. doi: 10.2174/1568026616666151012111046
Wang, F., Huang, S., Chen, Q., Hu, Z., Li, Z., Zheng, P., et al. (2020). Chemical characterisation and quantification of the major constituents in the Chinese herbal formula Jian-Pi-Yi-Shen pill by UPLC-Q-TOF-MS/MS and HPLC-QQQ-MS/MS. Phytochem. Anal. 31, 915–929. doi: 10.1002/pca.2963
Watrous, J., Roach, P., and Alexandrov, T. (2012). Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA. 109, E1743–E1752. doi: 10.1073/pnas.1203689109
White, G., and Taniguchi, E. (1972). The mode of action of helminthosporal. II. Effect on the permeability of plant cell membranes. Can. J. Bot. 50, 1415–1420.
Wu, C., van der Heul, H. U., and Melnik, A. V. (2019). Lugdunomycin, an Angucyline-drived molecule with unprecedented chemistry architecture. Angew. Chem. Int. Ed. 131, 2835–2840. doi: 10.1002/anie.201814581
Yang, J. Y., Sanchez, L. M., and Rath, C. M. (2013). Molecular networking as a dereplication strategy. J. Nat. Prod. 76, 1686–1699.
Yang, S., Shan, L., Luo, H., Sheng, X., Du, J., and Li, Y. (2017). Rapid classification and identification of chemical components of schisandra chinensis by uplc-q-tof/ms combined with data post-processing. Molecules. 22, 1778. doi: 10.3390/molecules22101778
Keywords: Bipolaris sorokiniana, seco-sativene sesquiterpenoids, McLafferty rearrangement, NMR analysis, antioxidant activity
Citation: Wang Y-D, Yang J, Li Q, Li Y-Y, Tan X-M, Yao S-Y, Niu S-B, Deng H, Guo L-P and Ding G (2022) UPLC-Q-TOF-MS/MS Analysis of Seco-Sativene Sesquiterpenoids to Detect New and Bioactive Analogues From Plant Pathogen Bipolaris sorokiniana. Front. Microbiol. 13:807014. doi: 10.3389/fmicb.2022.807014
Received: 03 November 2021; Accepted: 27 January 2022;
Published: 09 March 2022.
Edited by:
Mukesh Kumar Awasthi, Northwest A&F University, ChinaReviewed by:
Hayssam M. Ali, King Saud University, Saudi ArabiaCopyright © 2022 Wang, Yang, Li, Li, Tan, Yao, Niu, Deng, Guo and Ding. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lan-Ping Guo, Z2xwMDFAMTI2LmNvbQ==; Gang Ding, ZGdmeWhjaGluYUAxNjMuY29t, Z2RpbmdAaW1wbGFkLmFjLmNu
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.