 Hiran Kanti Santra
Hiran Kanti Santra Debdulal Banerjee
Debdulal Banerjee
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 26 January 2023
Sec. Microbiotechnology
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.1064055
This article is part of the Research Topic Biotechnological Applications of Endophytes in Agriculture, Environment and Industry View all 11 articles
Endophytes play a vital role in plant growth under biotic and abiotic stress conditions. In the present investigation, a Galactose-Rich Heteropolysaccharide (GRH) with a molecular weight of 2.98 × 105 Da was isolated from endophytic Mucor sp. HELF2, a symbiont of the East Indian screw tree Helicteres isora. OVAT (One Variable at A Time) experiment coupled with RSM (Response Surface Methodology) study exhibited 1.5-fold enhanced GRH production (20.10 g L−1) in supplemented potato dextrose broth at a pH of 7.05 after 7.5 days of fermentation in 26°C. GRH has alleviated drought stress (polyethylene glycol induced) in rice seedlings (Oryza sativa ssp. indica MTU 7093 swarna) by improving its physicochemical parameters. It has been revealed that spray with a 50-ppm dosage of GRH exhibited an improvement of 1.58, 2.38, 3, and 4 times in relative water contents and fresh weight of the tissues, root length, and shoot length of the rice seedlings, respectively “in comparison to the control”. Moreover, the soluble sugars, prolines, and chlorophyll contents of the treated rice seedlings were increased upto 3.5 (0.7 ± 0.05 mg/g fresh weight), 3.89 (0.57 ± 0.03 mg/g fresh weight), and 2.32 (1,119 ± 70.8 μg/gm of fresh weight) fold respectively, whereas malondialdehyde contents decreased up to 6 times. The enzymatic antioxidant parameters like peroxidase and superoxide dismutase and catalase activity of the 50 ppm GRH treated seedlings were found to be elevated 1.8 (720 ± 53 unit/gm/min fresh weight), 1.34 (75.34 ± 4.8 unit/gm/min fresh weight), and up to 3 (100 ppm treatment for catalase – 54.78 ± 2.91 unit/gm/min fresh weight) fold, respectively. In this context, the present outcomes contribute to the development of novel strategies to ameliorate drought stress and could fortify the agro-economy of India.
The foundation of the global food economy is agriculture, and in a nation like India, Gross Domestic Production (GDP) is heavily reliant on the agrarian model. Crop loss due to biotic and abiotic stressors is a widespread issue that requires effective management techniques to keep the agroecosystem in good shape. Biotic stress-related issues can be addressed with a variety of chemical formulations, such as pesticides, herbicides, fungicides, and biological techniques, such as biological control agents (BCA) and plant growth promotors (PGPs), but abiotic stress management strategies have received scant attention. As a result, issues with salt and drought stress are severely impeding crop development and yield, with dryness being the most detrimental. Agriculture production is significantly declining globally (Chen et al., 2017). The problem is getting worse as a result of cases of global warming and water scarcity. The agrarian economy is struggling with production-related problems as well as severe financial constraints (Vurukonda et al., 2016). According to reports, to fulfil this ambitious goal by 2050, food production must increase by up to 60 to 110 percent. Drought-related challenges must be immediately resolved (Naumann et al., 2018; Dey et al., 2019; Paglia and Parker, 2021). Therefore, it is urgent to discover a new, long-lasting solution to this global dilemma (Coleman-Derr and Tringe, 2014). One of the initial answers to that problem is to create stress-resistant varieties, but doing so takes time, is rigorous, species-specific, and is expensive (Santra and Banerjee, 2022a). One approach might be to cultivate crops on reclaimed drought-affected land while using foliar plant growth-promoting/stress-resisting chemicals. Rhizobacteria that promote plant growth have already been evaluated for this purpose, and less-studied endophyte or endophytic fungal or bacterial polysaccharides are currently showing promise in this field (Chen et al., 2017; Sun et al., 2020a,b).
Endophytes are ubiquitous in occurrence and are procured from nearly all plants and plant parts studied to date across the globe (Coleman-Derr and Tringe, 2014; Chatterjee et al., 2022). The symbionts of plant tissues known as endophytes help the plant grow and give tolerance in challenging conditions. Due to their horizontal gene transfer as a result of the co-evolution of the host and microorganisms, they share crucial genes of essential metabolomes (Santra et al., 2022; Santra and Banerjee, 2022b; Santra and Banerjee, 2022c). They have a reputation for being bioactive chemical mines that are simple to access using contemporary biotechnological methods. Plant growth-promoting endophytic bacteria and fungi reside on different internal plant tissues and organs, i.e., in stems, roots, flowers, leaves, fruits, and seeds. Endophytes have recently drawn attention because they are an effective tool for teaching plants to tolerate lethal abiotic stressors like drought, salt, and heavy metal toxicity (Cherif et al., 2015; Mesa et al., 2015; Pinedo et al., 2015; Constantin et al., 2019; Moghaddam et al., 2021) through adopting various mechanisms. The two most prevalent and important methods of building stress resistance are decreasing the levels of the key gaseous hormone ethylene through the activity of ACC (1-aminocyclopropane−1-carboxylate) deaminase and increasing the content of prolines in the tissues (Blaha et al., 2006; Gamalero and Bernard, 2012; Marasco et al., 2012). In addition to these, accumulation of siderophores and osmolytes, increased antioxidant and photosynthetic rates, synthesis of phytohormones and organic acids, and emission of volatile organic compounds are other important mechanisms used by endophytes to increase host plant abiotic stress tolerance (Tiwari et al., 2016; Vurukonda et al., 2016). Recent reports include that microbial symbionts or fungal endophytes are the co-evolution partners of green plants and promote habitat-specific stress tolerance in host plants (; Rodriguez and Redman, 2008; Redman et al., 2011). A special type of long-chain polymeric secondary metabolite called exopolysaccharides (EPS) from endophytic sources holds immense agricultural utility especially in ameliorating drought and salt stress (Nadeem et al., 2014; Rolli et al., 2015).
In the current study, exopolysaccharide was extracted from the endophytic fungus Mucor sp. HELF2 (isolated from Helicteres isora flowers). The EPS was galactose-rich heteropolysaccharide (GRH) in composition. GRH was found to be effective in reducing drought stress conditions when applied to the foliar parts of the rice seedling Oryza sativa ssp. indica MTU 7093 swarna. GRH production by HELF2 was optimised by adopting statistical modelling using Minitab and the predicted model led to an enhancement of 1.5 times exopolysaccharide (GRH) production under optimised fermentation conditions. The application of 50 ppm GRH was discovered to be the most efficient dosage, and the physical/biochemical traits of the treated plants were discovered to be higher than those of the untreated ones. Root and shoot length, fresh weight, enzymatic antioxidant profiles, and proline contents were improved remarkably after treatment. The membrane damage caused by lipid peroxidation was also minimised when GRH was applied in vivo. MDA content was reduced and SOD, CAT, and POD values were elevated. The current study illuminates the agricultural potential of endophytic exopolysaccharide, which has the potential to expand the field of sustainable development and improve the agro-economy of our nation’s indigenous population.
Mucor sp. HELF2 was isolated as an endophyte from the flower of an ethnomedicinally valuable plant Helicteres isora collected from forests in the East Singbhum district, Jharkhand, West Bengal, India, and stored, maintained on PDA slants and Petri plates at 4 ± 2°C and 25 ± 2°C, respectively. In brief, plant parts were thoroughly washed by running tap water for 5 min, sodium hypochlorite (2–10%) for 2 min, and hydrogen peroxide (2%) for 1 min, respectively, and explants were incubated on water agar plates at 27°C on biological oxygen demand incubator for endophyte isolation. Water agar plates were supplemented with antibiotics (streptomycin and tetracycline- 50 mg L−1) to avoid bacterial endophytes. The effectiveness of this sterilisation and isolation process was cross-checked by the explant imprintation technique described by Schulz et al. (1993). In brief, the aliquots used for explant sterilisation were spread on a water agar medium and incubated under the same conditions. After, 3–5 days of incubation, fungal hyphae emerged from the tissues and they were transferred to PDA plates for optimum growth (Schulz et al., 1993). Emerging fungal hyphal tips were transferred to PDA (Potato Dextrose Agar) medium and morphology (both macroscopic and microscopic) of the fungal isolate was recorded using light (Primo Star, Zeiss, Germany) and stereo microscope (Stemi 508, Zeiss, Germany).
The organism was identified by rDNA-based molecular technique as there was no reproductive structure produced by the endophytic fungi even in a medium with carnated leaves. In brief, genomic DNA of the fungal isolate was obtained (using DNeasy Plant Minikit-Qiagen, Germany) and a polymerase chain reaction was performed using the two universal primers named ITS1 (5’-TCCGTAGGTGAACCTTGCGG-3′) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3′; Laich and Andrade, 2016; Landum et al., 2016). The PCR products were separated using 1% agarose gel in 1X TAE buffer (90 mM Tris-acetate and 2 nM EDTA, pH 8.0), stained with ethidium bromide (0.5 μg mL−1), and documented using BIO-RAD Gel Doc EZ imager version 5.1 (United States). PCR products were sent for direct bi-directional sequencing using ABI 3730xl Genetic Analyzer (Applied Biosystems, United States) to Bioserve Biotechnologies (India) Pvt. Ltd., A Repro Cell Company, Hyderabad, India. The obtained consensus sequence of 620 bp was used for further study. Sequences were submitted to GenBank and were compared to the GenBank database using BLAST. Fifteen sequences along with HELF2 were selected and aligned using the multiple alignment software program Clustal W and the phylogenetic tree was prepared using MEGA 11 (Tamura et al., 2021).
Endophytic fungi were grown in different 250 mL Erlenmeyer flasks with 50 mL potato dextrose broth in a shaker incubator at 120 rpm for 8 days. An initial medium pH of 6 and a medium temperature of 28°C were maintained.
To detect the optimum culture conditions for the maximum production of GRH, fungi were grown in varying fermentation times (4–10 days), then in different medium pH (5.5–7.9), and then at varying incubation temperatures (20–30°C) in separate Erlenmeyer flasks with separate PDB medium. To find out the requirement of additional nutrients for maximum GRH production and mycelial growth, various carbon sources (5 g%, w/v of fructose, glucose, maltose, starch, rhamnose, raffinose, glycerol), various organic and inorganic nitrogen sources (0.4 g% w/v of peptone, ammonium nitrate, urea, ammonium chloride, glycine and yeast extract) in different Erlenmeyer flasks were used with PDB as the basal medium. After the finalisation of the additional carbon and nitrogen sources, their optimum concentration was confirmed by using different concentrations of these products on a PDB medium and the respective biomass and GRH amounts were calculated. A variety of ionic salts (0.1 g%, w/v of MgCl2, FeCl3, KCl, NaCl) and phosphate sources (0.1 g%, w/v including NaH2PO4, K2HPO4, KH2PO4) were analysed separately to detect their role in fungal biomass and GRH production (Mahapatra and Banerjee, 2013, 2016).
To detect the O2 requirement, fungi were grown with different medium volumes in 250 mL Erlenmeyer flasks. Headspace volume, medium volume, total volume, and medium depth in flask culture were measured for the indirect measurement of the organism’s O2 requirement (Wonglumsom et al., 2000).
Further optimisation was performed with the RSM (Response surface methodology). The investigational design was a Box–Behnken experimental setup with the four most important factors obtained from the OVAT system. The four independent factors had three different levels (−1, 0, and + 1) each for the experiment. GRH production was set to a second-order polynomial equation by the means of multiple regression techniques. The model involving the most significant factors was derived. The system performance follows the subsequent second-order polynomial equation: Y = β0+ ΣβiXi + ΣβijXiXj + ΣβiiX2i, where Y is the predicted response or dependent variable, xi and xj are independent factors, β0 is the intercept of the regression equation, βi is the linear coefficient, βii is the quadratic coefficient and βij is the interaction coefficient (Mahapatra and Banerjee, 2013, 2016).
Fungal biomass was separated from the culture extract by centrifugation at 10,000 rpm. Mycelial biomass was dried at 55°C for 24 h and weighed. The supernatant was concentrated in a rotary evaporator under low pressure at 40°C. Chilled absolute ethanol was added to the concentrated supernatant (5:1 v/v), mixed thoroughly, and kept for 24 h at freezing conditions finally, the recovery of viscous precipitate was done by centrifugation at 10,000 rpm for 10 min. The recovered polysaccharide was dialyzed in a cellulose membrane (MW cut off 10,000) against distilled water for 24 h. It was tested for sugar and protein contents following the methods of Dubois et al. (1956) and Lowry (1951) with glucose and bovine serum albumin as the standard. Obtained EPS solution was concentrated in a rotary evaporator under low pressure at 40°C for characterisation.
GRH was purified by gel chromatographic technique using a Sepharose-6B gel filtration column (65 × 2 cm) and average molecular weight was determined following the methods of Mahapatra and Banerjee (2013). The dried polysaccharide was subjected to characterisation using GC–MS with some pre-treatments (Proestos et al., 2006). Using a water bath at 70°C for 15 min, 100 mg of dried exopolysaccharide was combined with 1 mL MeOH, 20 μl ribitol (which serves as an internal standard), and 20 μl nor-leucine. The entire mixture was then centrifuged for 5 min at 10,000 rpm, and the supernatant was immediately dried and dissolved in 20 μl of methoxy-amine HCL for 120 min at 37°C. The final 1 μl of the derivatized EPS sample was loaded onto the GC–MS for analysis of monosaccharide composition after 40 μl of TMS (Trimethyl siloxane) had been added. The instrument was set up with a 30 m × 0.25 mm DB-5 Ultra Inert column. With a split ratio of 25:1, the inlet temperature was 230°C, and the MS transfer line temperature was 250°C. A constant flow mode was used for the column’s flow, which was 1.3 mL min−1 with an average linear starting velocity of 39 cm sec−1. Helium was used as the carrier gas, and ZB − 1701 served as the guard column. The program was isothermal, holding at 70°C for 5 min, increasing by 10°C per minute to 180°C, holding for 2 min, increasing by 10°C per minute again to 220°C, holding for 1 min, and finally ramping up by 2.5°C per minute. Up to 265°C with a 1-min hold, then a ramp up to 285°C with a 1-min hold, and finally a climb of 1°C per min up to 290°C with a 0.6-min hold. The mass spectrum was obtained in scan mode from 40 to 650 amu with a detection threshold of 100 ion counts while the detector was in positive ion mode. Appropriate configurations (D-dextrorotatory and L-laevorotatory form) of the sugars were identified by matching them with the NIST library. To detect the sugar linkages, procedures of Ciucanu and Kerek (1984) and Das et al. (2009) were adopted using the GC–MS equipment.
Initially, healthy and disease-free rice seeds (O. sativa ssp. indica MTU-7093 Swarna) were surface sterilised with a series of surface disinfectants: sodium hypochlorite (2.5%) for 20 min, deionised double-distilled water (3–5 times thoroughly) and then soaked in water for germination followed by storing at 22°C for 72–96 h. Uniformly seeds were transferred to a hydroponics box supplemented with Hoagland solution and were replaced at an interval of 3–4 days (Chen et al., 2011). Seedlings reaching an age of 15–20 days were divided into four separate groups with the control group (un-inoculated water), 20 ppm, 50 ppm, and 100 ppm GRH application, respectively, at a frequency of 3 times a day (morning, noon, and afternoon) for 45 days. Simultaneously, 20% polyethylene glycol (PEG)-6,000 (for 7 days) was mixed with a hydroponic solution as a drought-inducing component, and all the biochemical tests were performed from drought-induced seedlings. The fresh weight of rice seedlings was measured and leaves were stored at −20°C for further biochemical estimations.
The relative water content (RWC) of the treated and control plants were calculated in percentage following the method of Arndt et al. (2015). Fresh leaves were plucked and fresh tissue weight (FW) was measured, then immersed in a 50 mL tube with distilled water, and placed in the dark at 4°C for 20 h. Further, the leaves were dried with filter paper and again weighed for turgid weight (TW) calculation. Lastly, the same leaves were incubated at 80°C for a period of 72 h and dry weight (DW) was measured immediately. Relative water content was calculated by the formula RWC (%) = (FW−DW)/ (TW−DW).
The chlorophyll content (mg g−1 of fresh weight) of the fresh leaves was measured according to the modified formula of Lichtenthaler and Wellburn (1983). Firstly, fresh leaves (0.5 g fresh weight) were split into small pieces and immediately dissolved in 50 mL methanol (80% v/v), covered with black paper, or kept in dark conditions for 24–36 h at 28–30°C. Centrifugation was performed and the supernatant was estimated (645 nm and 653 nm) for chlorophyll contents. Chlorophyll content (mg L−1 FW) = 8.05 A653 + 20.29 A645.
To calculate the proline contents of the seedling methods proposed by Bates et al. (1973) were adopted. Leaves (0.5 g fresh weight) were split into small pieces and put in a test tube. Further treatment was done by mixing with 5 mL of 3% sulfosalicylic acid, incubated in a water bath for 10 min and 2 mL of the supernatant was mixed thoroughly with 2 mL of acetic acid, and 3 mL of 2.5% ninhydrin. Finally, the mixture was incubated in the water bath for a time period of 40 min−1 h and extracted using 4 mL methylbenzene, optical density was measured at 520 nm and compared with a proline standard curve.
Soluble sugar contents (in terms of mg g−1 fresh weight) were measured by the method of Watanabe et al. (2000). Fresh leaves (0.2 g fresh weight) were crushed in 80% v/v ethanol (10 mL) and centrifuged at 8000 g for 10 min at 4°C. We mixed 1 mL of supernatant thoroughly with 3 mL of anthrone reagent followed by heating at 100°C for 10–12 min, which was stopped by rapid cooling them on the ice. Finally, at 620 nm absorbance was estimated using glucose as a standard.
Malondialdehyde content (nmol g−1 fresh weight) was reported according to the method of Del Buono et al. (2011) 0.5 gm of fresh leaves were homogenised in 5% (w/v) trichloroacetic acid (TCA), centrifuged at 12000 g for a time period of approximately 15–20 min and then the supernatant was mixed with 5 mL of 0.5% thiobarbituric acid (TBA)-prepared with 20% TCA followed by incubation for 25 min and cooling at 100°C and room temperature, respectively. Finally, after centrifugation (7,500 g for 5 min), the supernatant was measured for its absorbance at 450, 532, and 600 nm. The amount of MDA was calculated by the following formula MDA content (nmol g−1) = 6.45 (A532-A600)−0.56A450.
The methods of Lei et al. (2015) were followed for the measurement of peroxidase activity. The system contained multiple chemicals; 2.9 mL of 0.05 M phosphate buffer, 0.5 mL of 2% H2O2, 0.1 mL of 2% guaiacol, and also 0.1 mL of crude enzyme extract followed by the absorbance measurement at 470 nm. Lastly, POD activity was calculated as an amount of guaiacol oxidised per minute in nanomoles per minute per mg of protein. At the end of the reaction, the absorbance was measured at 470 nm. POD activity was defined as the amount of guaiacol oxidised per minute, and was expressed as nanomoles per minute per mg of protein.
Catalase activity was calculated following the protocols of Lei et al. (2015). O.1 mL H2O2 (2%) and 2 mL phosphate buffer (50 mM-pH 7.0) were mixed and the whole reaction was initiated by the addition of 0.1 mL of crude enzyme extract. Finally, the catalase activity was measured (at 240 nm) in terms of the decrease of values of H2O2 per minute, as nanomoles/min/gm of protein.
Superoxide dismutase activity was assayed following the protocols of Lei et al. (2015). The whole system contained a series of valuable freshly prepared reagents; 1.5 mL of 0.05 M phosphate buffer, 0.3 mL of 130 mM methionine solution, 0.3 mL of 750 μM nitroblue tetrazolium solution, 0.3 mL of 100 μM EDTA -Na2 solution, 0.3 mL of 20 μM lactochrome solution, 0.5 mL of distilled water and finally 0.1 mL of crude enzyme extract. The complete reaction was initiated at 4000 Lx of illumination for a constant 20 min with no interruption. The control set comprises the same set of reagents and illumination but with no crude enzyme extract, rather replaced with a phosphate buffer. The third setup of control contains only phosphate buffer followed by incubation in dark conditions for the same time period of 20 min. Finally, after the completion of the reaction, the absorbance was estimated at 560 nm of wavelength. One unit of SOD activity was defined as the amount of enzyme which inhibits NBT reduction by 50%, also the results were expressed as unit/mg protein.
All experiments were performed in triplicate and the results are presented as means ± standard errors (SE). Data were analysed by Prism GraphPad version 9.2.0 (332) software (San Diego, California, United States). BBD experiments were done in Minitab (version 20.2).
The organism had enormously intertwining hyphae and septate hyaline whitish mycelium, and it lacked any sexual or asexual reproductive structures (Figures 1A–C). rDNA sequence data of the isolate was deposited in GenBank (ON146358). A BLAST search of the earlier existing database indicates a close genetic connection with other species of Mucor and the evolutionary history of the endophytic fungal isolate HELF2 was included using the neighbour-joining method (Saitou and Nei, 1987). The most appropriate phylogenetic tree with a total branch length of 0.00649585 is represented in Figure 1D. The tree was constructed to scale with branch lengths in similar units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances between the species were analysed using the maximum composite likelihood method (Tamura et al., 2021) and were in the units of the number of base substitutions per site. Gaps and missing data were removed from the dataset. There was a total of 620 nucleotides in the final dataset.
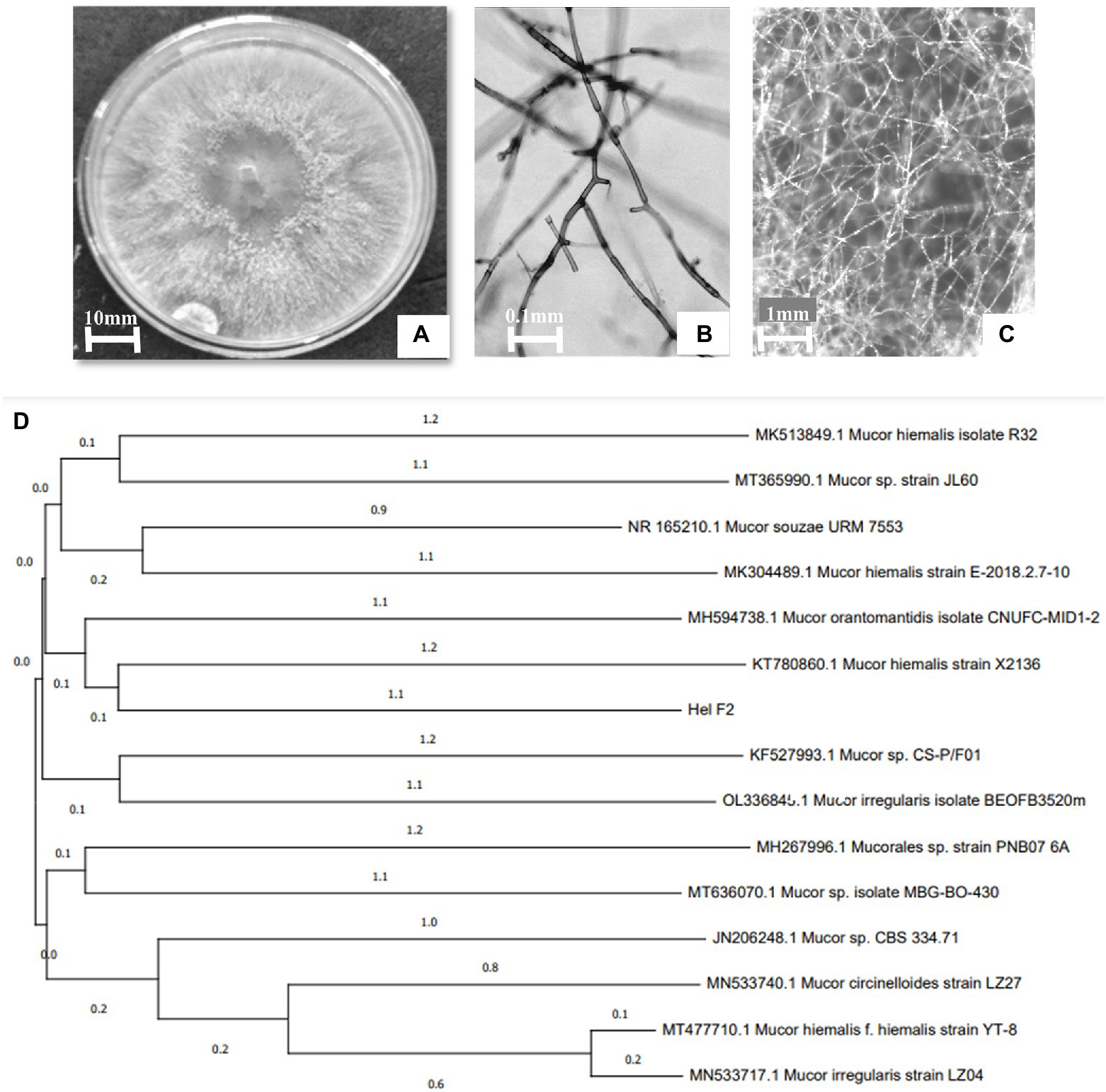
Figure 1. (A) 8-day old culture of HELF2 grown on PDA medium. Sterile hyphal aggregation without any sexual or asexual reproductive structures seen under a light (B) and stereo (C) microscope. (D) Phylogenetic tree shows the relationship of endophytic Mucor sp. HELF2 with other Mucor sp. strains.
Mucor sp. HELF2 was grown on a 250 mL Erlenmeyer flask in submerged condition for 10 days and the highest production of fungal GRH and biomass was detected after 8 days of fermentation (Table 1). A temperature of 26°C and a medium pH of 7.1 was found to be the most suitable one for GRH production. Glucose and peptone at a concentration of 11 g L−1 and 5.5 g L−1 were found to be the most appropriate ones for maximum GRH yield (Table 1). NaCl at a concentration of 0.1 g L−1 was the most effective salt (or source of metal ions) for GRH production. The detailed effect of different parameters on biomass and GRH production is summarised in Table 1.
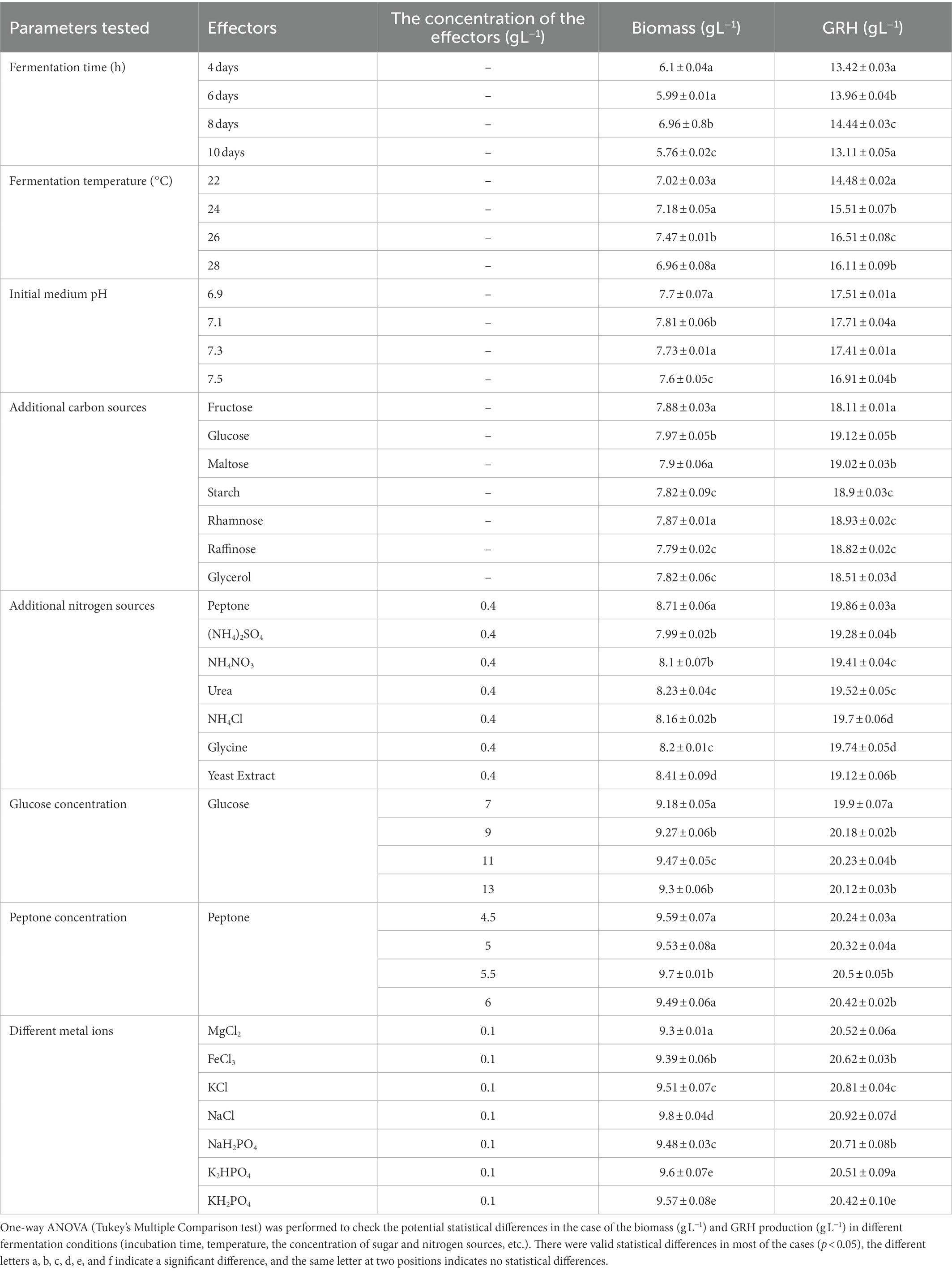
Table 1. Effect of different fermentation influencing physical conditions and chemical supplements, on biomass and Galactose Rich Heteropolysaccharide production by endophytic fungi Mucor sp. HELF2 in submerged fermentation conditions.
The amount of dissolved oxygen in the fermentation medium affects EPS production. It depends on the medium volume, the headspace of the medium, and the medium depth. A medium volume of 75 mL in a 320 mL Erlenmeyer flask with 245 mL of headspace volume and 2.1 cm of medium depth and 2.53 cm of surface area was found to be the criteria for optimum GRH production (Table 2).
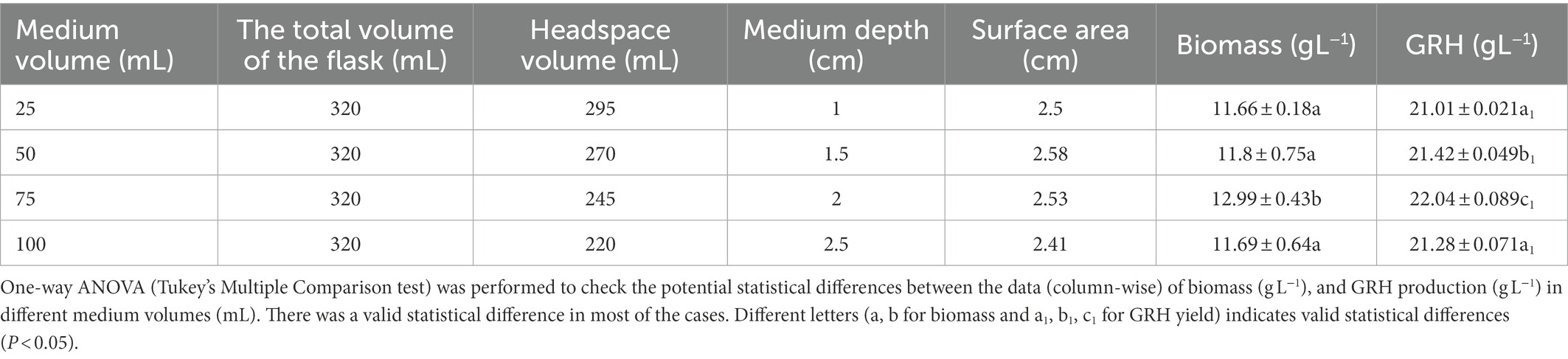
Table 2. Effect of medium volume, headspace volume, and medium depth on the dissolved oxygen level in the fermentation medium and their effect on GRH and biomass production.
After OVAT optimisation RSM was adopted using a three-level Box Behnken Design. The most important four factors (glucose concentration, peptone concentration, medium pH, and fermentation time) with five replicates at the center points were established as a model for analysis of GRH production. The experimental design with variable predicted and measured values of GRH was presented in Table 3. Maximum GRH production was noted at the five replicated center points. The predicted response Y for GRH production by Mucor sp. HELF2 was described as coded factors in the following equation YGRH = 5.9810–0.168024x1–0.39671x2-0.09103x3-0.37144x4-0.25686x1x2–0.03737x1x3 + 0.09369x1x4–0.0367 x2x3 + 0.32736 x2x4–0.0337 x3x4−0.65663 x21-0.6395 x22−0.50414 x23–0.85426 x24. Here YGRH is the predicted GRH yield and x1, x2, x3, and x4 are the four coded factors of glucose concentration, peptone concentration, medium pH, and fermentation time (day) respectively. A regression analysis with detailed statistical data related to the experiment is presented in Table 4. The F-test data of 1994.082 proved that the model was significant. The adjusted determinant coefficient (R2 Adj) was found to be 0.9998 which represents that there is a high degree of correlation between the experimental and predicted values and there is more than 99% variation in response that could be predicted by second-order polynomial prediction equation. Adeq precision was reported to be 111.901 which indicates that the model is appropriate. The lack of fit F-value of 5.782 and p-value (p < 0.0001) was not at all valuable to the pure error and the fitness of the model was perfect. The high degree of precision and uniformity of the investigational outcomes were proved by the p value of lack of fit- 0.2830 (> 0.05) and a p value of probability (>F less than 0.05). The other linear and quadratic effects of glucose concentration, urea concentration, M-pH, and fermentation time were also significant (p < 0.0001). Finally, three-dimensional response surface plots and contour plots were constructed by Minitab (20.2) for a clear understanding of the effects of the parameters on GRH production (Figure 2).
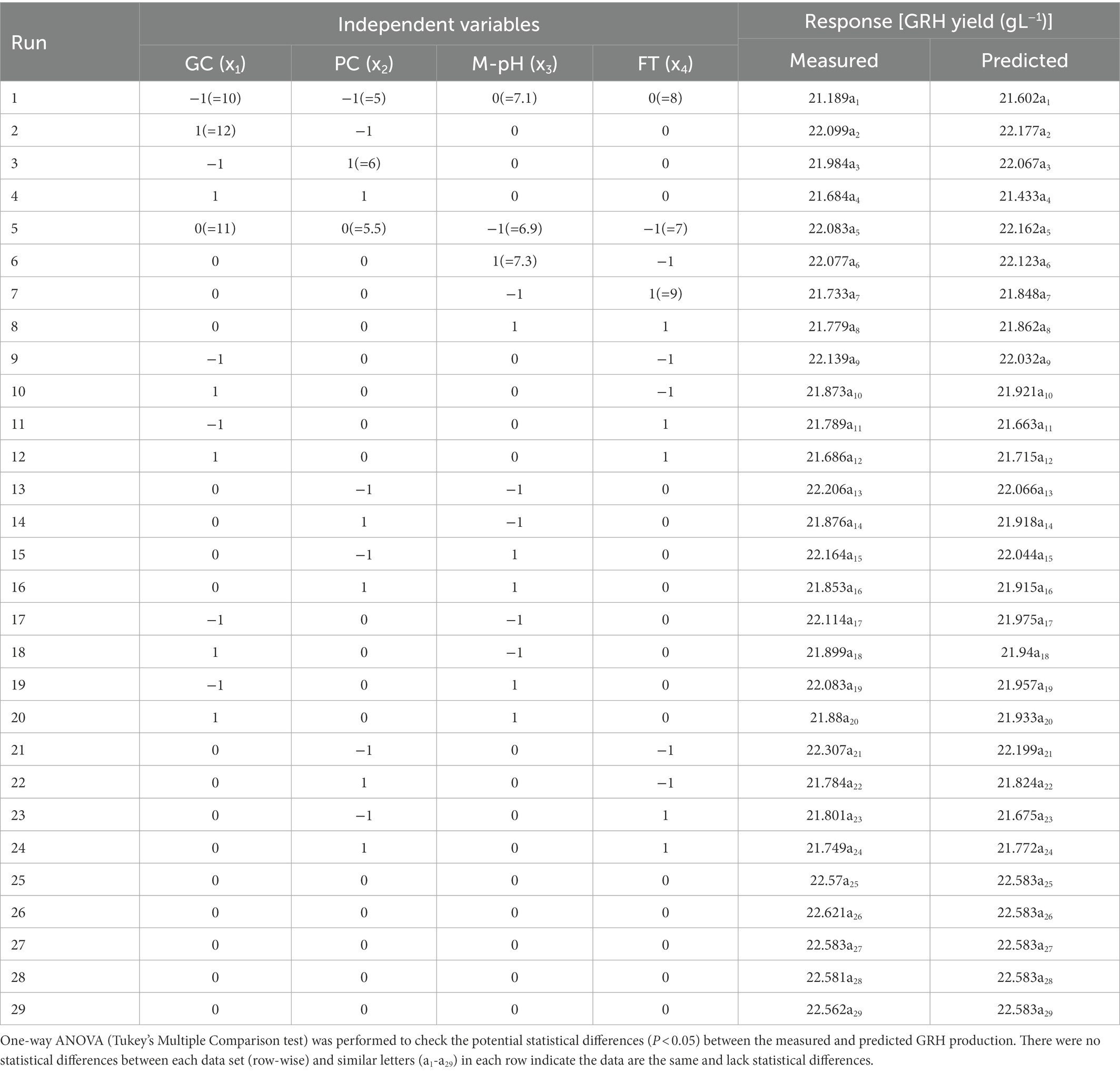
Table 3. Experimental design and outcomes of the Box–Behnken Design (BBD) for optimisation of the GRH production from Mucor sp. HELF2.
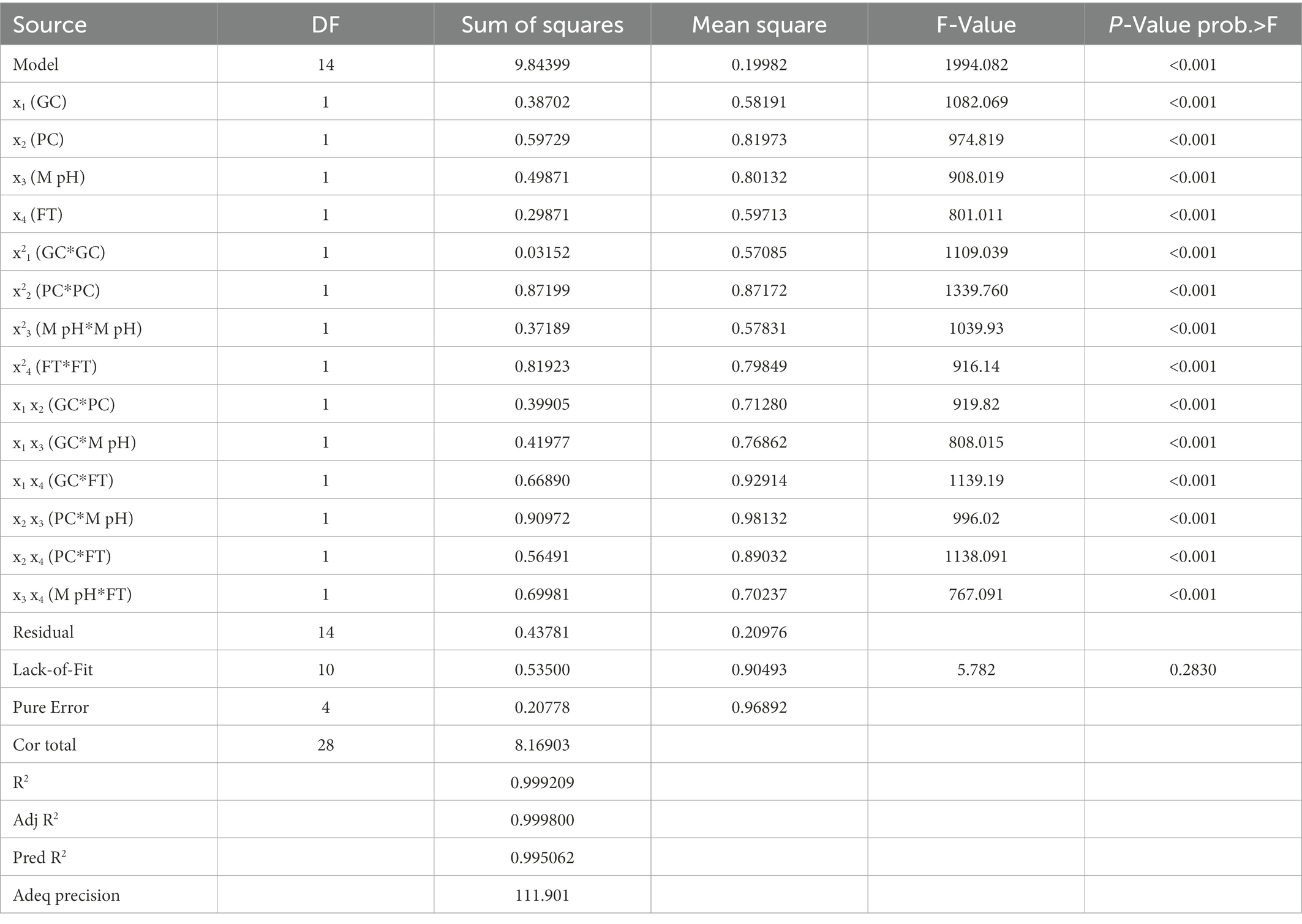
Table 4. ANOVA for response surface quadratic regression model of Galactose Rich Heteropolysaccharide production by endophytic fungi Mucor sp. HELF2.
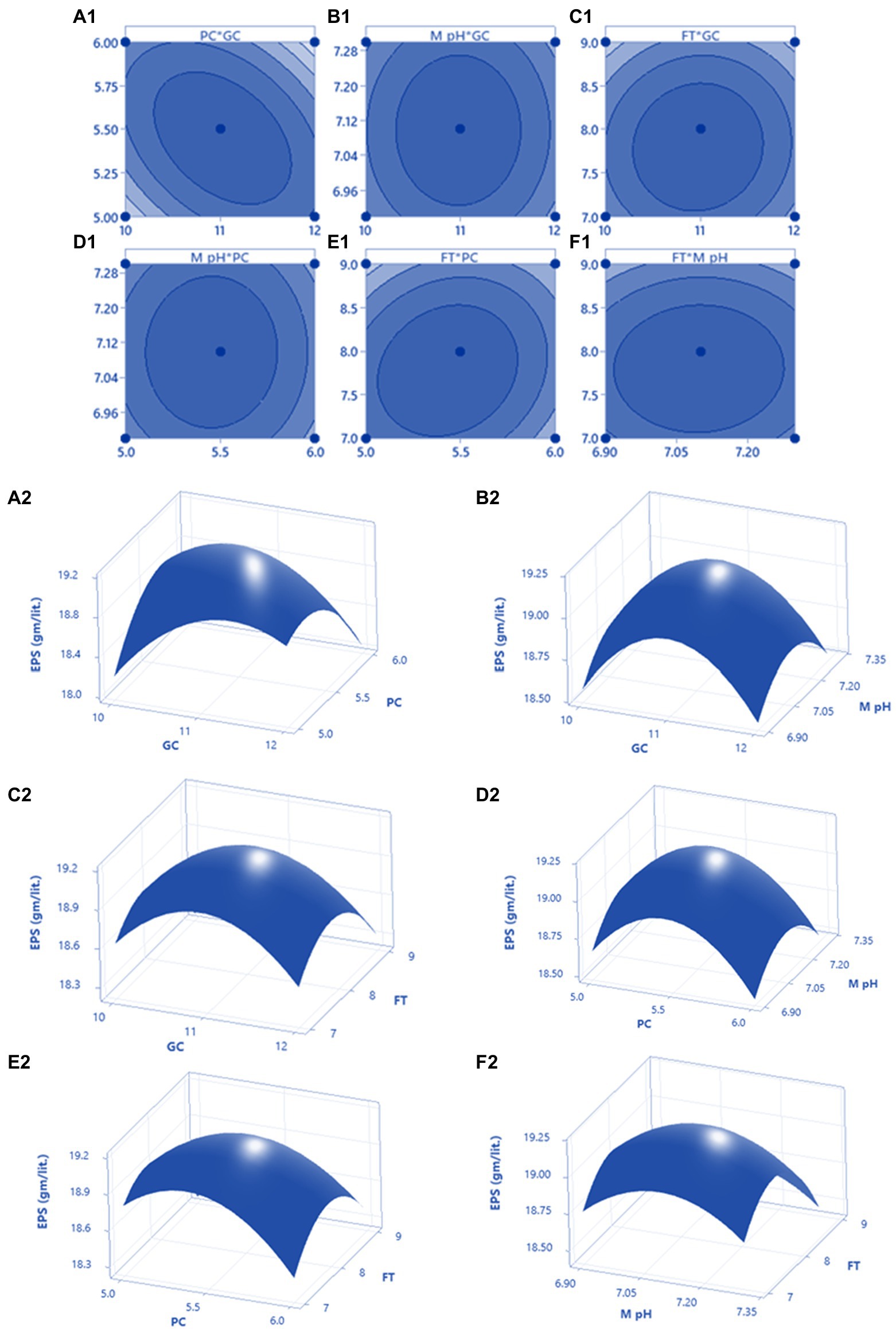
Figure 2. The 3D plot with 2D projection and contour plot showing the most important interactions of factors in RSM optimization of GRH production by HELF2. (A1,A2) between peptone concentration (PC) vs. glucose concentration (GC) at fermentation time (FT) 8 days and medium pH 7.1 (MpH); (B1,B2) between MpH (7.1) and GC (11 g L−1) at FT (8 days) and PC (5.5 g L−1); (C1,C2) between FT (8 days) and GC (11 g L−1) at PC (5.5 g L−1) and MpH (7.1); (D1,D2) between MpH (7.1) and PC (5.5 g L−1) at FT (8 days) and GC (11 g L−1); (E1,E2) between FT (8 days) and PC (5.5 g L−1) at MpH (7.1) and GC (11 g L−1); (F1,F2) between FT (8 days) and MpH (7.1) at GC (11 g L−1) and PC (5.5 g L−1).
The model predicted a maximum response of 20.10 g L−1 GRH yield when the necessary components are 5.2 g L−1 of peptone, 10.5 g L−1 of glucose, 7.05 MpH, and 180 (7.5 days) h of fermentation time. These predictions were authenticated by performing laboratory experiments in flask culture by triplicate with an outcome of 19.951 ± 0.091 g L−1 of GRH.
Exopolysaccharide produced by Endophytic Mucor sp. HELF2 was precipitated by applying chilled ethanol and crude EPS was then dialysed, and purified by gel filtration chromatography with a Sepharose-6B column. One major fraction obtained was eluted between 29 and 42 tubes (Figure 3C) and the colorimetric test confirms the absence of proteins in those fractions. The fraction was further investigated for monosaccharide analysis. The molecular weight of the homogeneous EPS was calculated from a calibration curve of standard dextran as ~2.98 × 105 Da (Figure 3D). Monosaccharide analysis of the derivatised EPS samples showed the occurrence of galactose, fucose, and glucose in a 13:2:1 ratio with D, L, and D configuration, respectively (Table 5). Each repeating unit of the fraction contained 13 galactose, two fucoses, and one glucose, which indicates that the studied EPS contained approximately 104 repeating units. We, therefore, considered that the Galactose Rich Heteropolysaccharide (GRH) could have been produced by endophytic Mucor sp. HELF2. FT-IR analysis of the EPS sample revealed the occurrence of strong absorption peaks at particular wavelengths of 3400.71, 2950.89, 1651.56, 1489.73 which represents C-H, O-H, C-O asymmetric stretching respectively, which indicates the basic characteristics and purity of the carbohydrate moiety. Figures 3A,B represent the FT-IR spectrum and GC–MS spectrum of the crude and derivatised EPS, respectively.
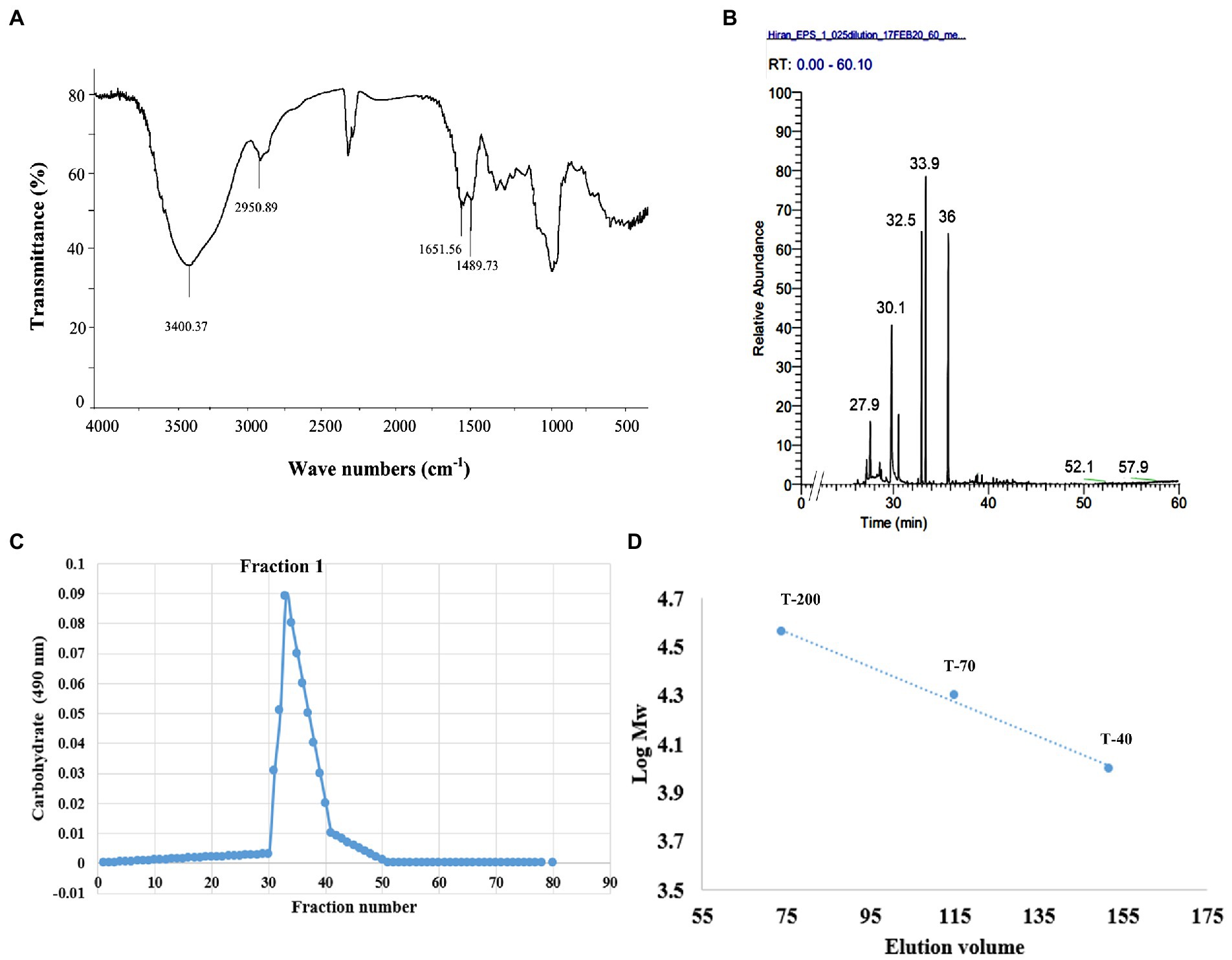
Figure 3. (A) FT-IR spectra of crude HELF2 GRH showing the necessary functional groups present in the sample. (B) GC–MS chromatogram of the derivatized fungal GRH showing the different peaks of monosaccharide compositions at different retention times. (C) Elution profile of the polysaccharide showing the occurrence of dominant fraction (F1) confirmed by carbohydrate test. (D) Standard curve of dextran needed for the detection of sugar concentration in the polysaccharide sample.
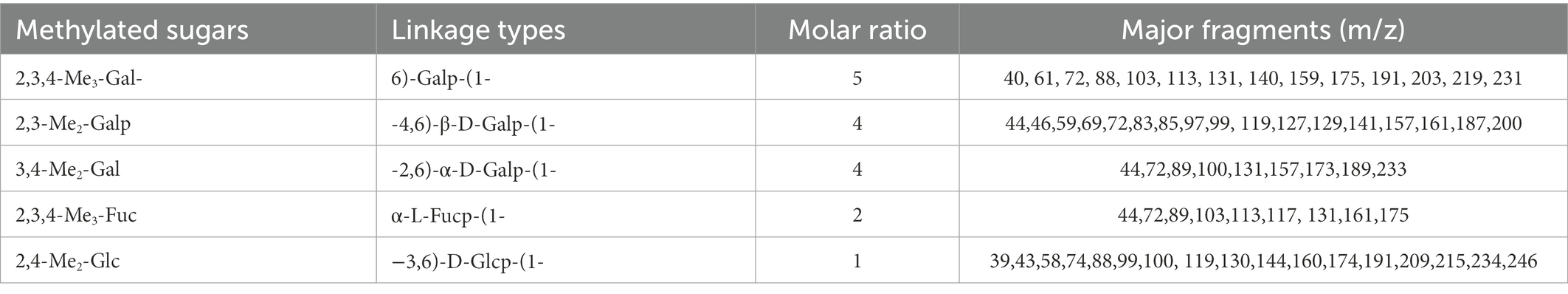
Table 5. Monosaccharide units present in the Galactose Rich Heteropolysaccharide (synthesised by endophytic fungi Mucor sp. HELF2) are represented here with their respective sugar linkages and molar ratios.
GRH-sprayed rice seedlings were found to be much healthier, and more vigorous in terms of their fresh weight and relative water contents in comparison to the control (only drought-inducing agent-PEG treated). The control plants were characterised by low growth, chlorosis, and wilting of leaves. The rice seedlings exhibited maximum growth promotion after continuous 14 days of GRH treatment. The relative water content and fresh weight of the 50 ppm GRH treated plants were found to be higher than the plants of the control set, and plants treated with 20 ppm and 100 ppm GRH dosage. There was a 1.31, 2.38, and 1.74-time improvement in the fresh weight of seedlings in the 20, 50, and 100 ppm GRH treated plants compared to the control. The relative water contents were also increased by 1.14, 1.58, and 1.26 times in 20, 50, and 100 ppm GRH treated plants than the control one. There was a 3, and 4 times increase in root length and shoot length of the treated (50 ppm GRH) plant, respectively, compared to the control. The fresh weight of the seedlings were found to be improved after the GRH treatment and the 50-ppm GRH application was also found to be the most effective in comparison to the control. Table 6 represents the improved physical characteristics of the treated seedlings. Figure 4 represents the treated (20, 50, and 100 ppm) and control rice seedlings, showing their physical changes.
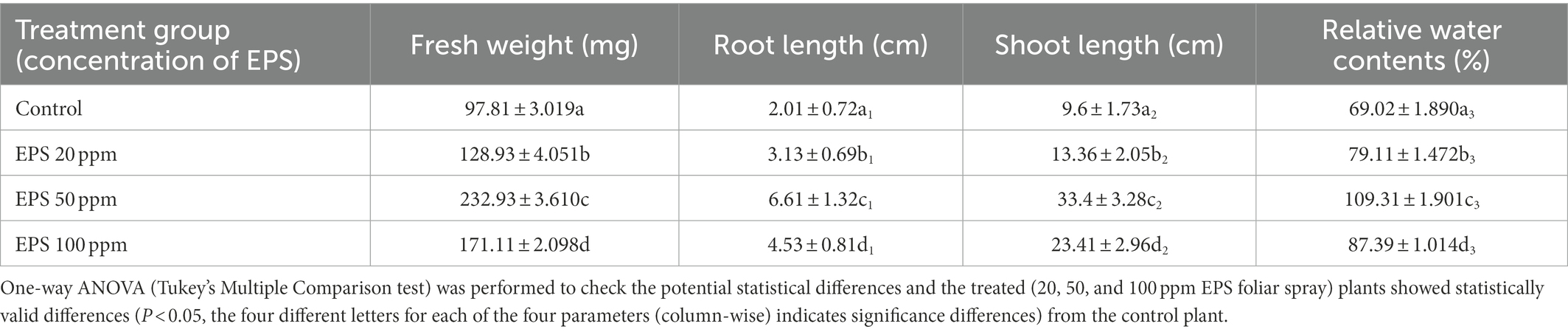
Table 6. Different physical parameters (fresh weight, root length, and shoot length) of GRH-treated and untreated drought-faced rice seedlings are represented here.

Figure 4. Phenotypes of rice seedlings Oryza sativa ssp. indica MTU 7093 Swarna under drought stress (induced by PEG treatment) sprayed with 20, 50, and 100 ppm EPS of Mucor sp. HELF2 endophyte.
Not only physical but also biochemical characteristics were improved in the case of GRH-treated plants even after severe drought situations. The chlorophyll contents of the treated plants were found to be more elevated (2.32 times higher for 50 ppm GRH treated one) than the untreated control (Figure 5A). The proline is a potent indicator of plant stress and higher proline contents indicate higher resistance towards stress and better adaptation to that stressful situation. Here the GRH-treated plant shows higher accumulations (approximately 3.89 times higher for 50 ppm GRH treatment) of proline contents than the control ones (Figure 5B). The presence of increased soluble sugar content in the plant tissues also indicates the higher survival ability of plants in drought-stress situations. In this experiment, we found almost 3.5 times higher accumulation of soluble sugar contents in 50 ppm GRH-treated plants in comparison to the only PEG-treated one (Figure 5C). On the other hand, the MDA (Malondialdehyde) content is found to be correlated with lipid peroxidation and membrane damage. Higher MDA content in the plant tissues indicates the detrimental situation induced by the stress factors. In the case of treated seedlings, there was a sharp six time decrease in MDA contents compared to the control (Figure 6A). Other enzymatic antioxidative parameters were found to be also elevated after GRH treatment even in extreme drought situations. SOD, CAT, and POD activities increased up to 1.44, 2.09, and 1.79 times, respectively, in the case of GRH-treated rice seedlings compared to those treated only with PEG (Figures 6B–D).
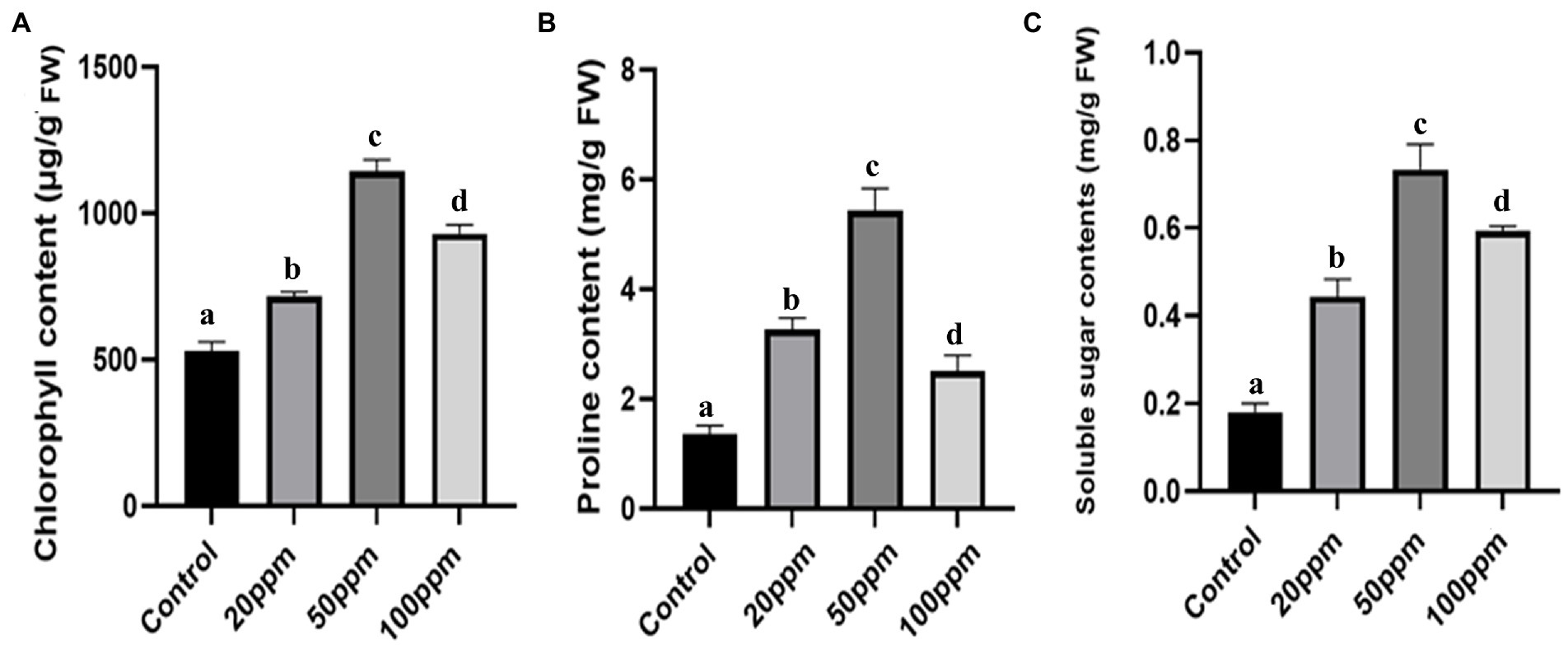
Figure 5. The effect of GRH foliar spray (20, 50, and 100 ppm) on chlorophyll content (A), soluble sugar content (B), proline content (C) of O. sativa ssp. indica MTU 7093 swarna in comparison to control. Values on the graphs are the means ± Standard error (SE) of three replicates. Tukey’s multiple comparison test was performed. The letters a, b, c, and d indicate significant differences compared to the control plant (At, p < 0.05).
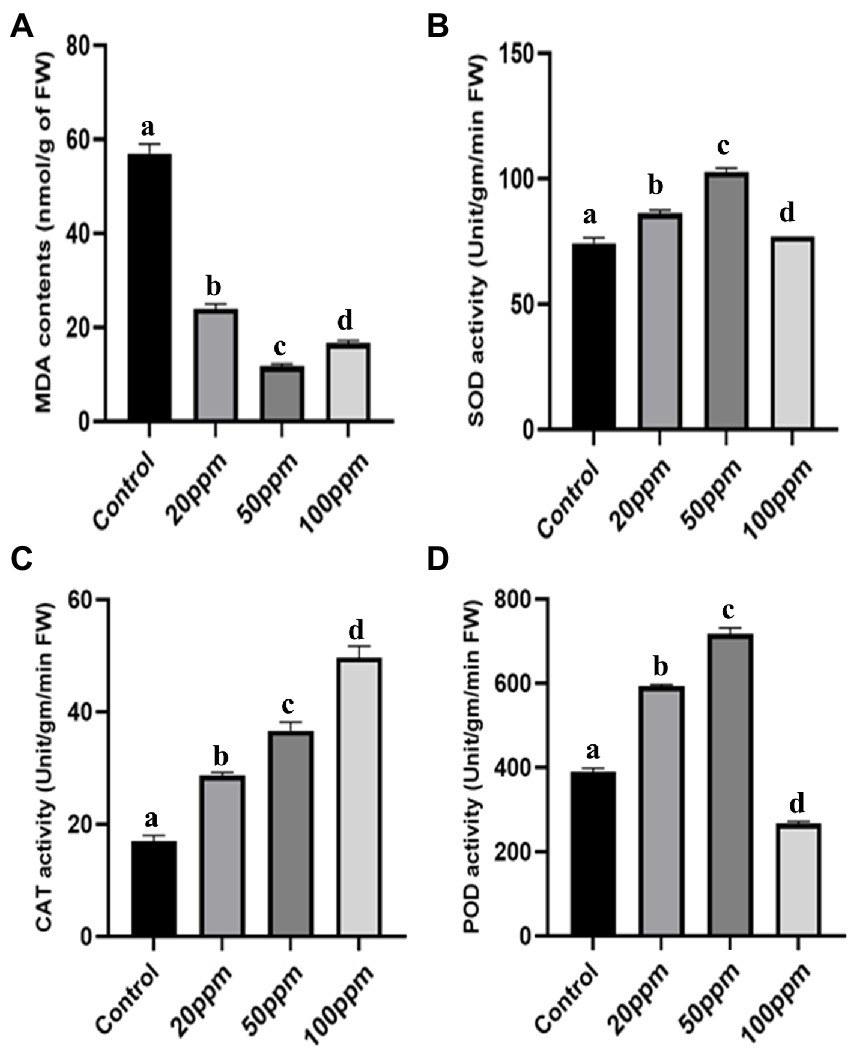
Figure 6. The effect of GRH foliar spray (20, 50, and 100 ppm) on malondialdehyde (MDA) contents (A), peroxidase (POD) activity (B), catalase (CAT) activity (C), superoxide dismutase (SOD) activity (D) of O. sativa ssp. indica MTU 7093 swarna in comparison to control. Values on the graphs are the means ± Standard error (SE) of three replicates. Tukey’s multiple comparison test was performed. The letters a, b, c, and d indicate significant differences compared to the control plant (At, p < 0.05).
Agriculture is seen as the most important and crucial sector of the global economy, and it significantly affects our GDP (Gross Domestic Production). The increased explosion in population in recent years has increased the demands for global agricultural output or food production by 60–100% by the end of 2050 to meet these growing needs, but the main obstacles are the lack of suitable fertile croplands and the rising instances of soil desertification due to insufficient precipitation, random evaporation, and a lack of freshwater resources, among other factors (Naumann et al., 2018; Dey et al., 2019; Paglia and Parker, 2021). Therefore, the primary requirement for a successful solution is the restoration of land or the development of salt or drought stress varieties. The development of drought-tolerant plants could temporarily meet the world’s food demand and protect crop plants, but the situation becomes severe when drought conditions (like the 2011–17 California drought and the 1997–99 Melbourne Millennium drought) occur on large scales around the globe. The food supply chain is hampered, and even the forest environment is impacted (Allen et al., 2010). Therefore, it is strongly recommended that deep ecological techniques that use non-toxic, natural substances be developed to address these issues. Exopolysaccharides produced by microbes, especially endophytes could have a significant impact (Chen et al., 2017). Even in situations with salt and drought challenges, endophytic fungi and bacteria are well known for their ability to promote plant growth (Azad and Kaminskyj, 2016; Bibi et al., 2019; Ali et al., 2021; Gupta et al., 2021). There are many reports on how microorganisms (both endophytes and rhizospheric) can reduce abiotic stress (Hammami et al., 2016), and endophytes often play an osmoprotective role in maintaining good water chemistry (managing Na+/K+ balance) within cells (Jha et al., 2011; Abdelaziz et al., 2017). Previous research has been undertaken, examining the role of endophytic fungi and bacteria in reducing the effects of salt stress in rice, maize, soybean, quinoa, barley, and barrel medic, as well as in the model plant Arabidopsis thaliana through endogenous hormone (abscisic acid) mediated morphological, biochemical (through ion balancing), and antioxidant defence-related pathways (Baltruschat et al., 2008; Bagheri et al., 2013; Jogawat et al., 2013; Li et al., 2017; Shahzad et al., 2017; Asaf et al., 2018; Fan et al., 2020; Ali et al., 2022; González-Teuber et al., 2022). Exo-polysaccharides and gamma-polyglutamic acid are also discovered to be the most useful compounds released by plant growth-promoting microbes and have exceptional biotic and abiotic stress tolerance (Livingston et al., 2009; Lei et al., 2017). Here, drought stress ameliorating properties of GRH was evaluated on rice plants. Due to their widespread popularity around the world, rice seedlings (Oryza sativa ssp. indica MTU 7093 Swarna- Indian subcontinental cultivar) were taken into consideration for their studies on drought relief. It is a very demanding staple food, especially in China, India, and Japan (Uga et al., 2013; Zhu, 2016), and has greater irrigation water needs (Bouman et al., 2005, 2007). According to recent studies, around forty-two million hectares of rice farming face significant challenges because of a lack of water. To formulate an appropriate response, we reported on how Galactose Rich Heteropolysaccharide reduces the effects of drought stress on rice plants. Chen et al. (2017) and Santra and Banerjee (2022a) both found that the application of direct endophyte and EPS produced from endophyte alleviated salt and drought stress in wheat and rice plants, respectively. Due to their high polymeric configurations, effective water-holding capacities, and strong affinity to create bio-films or similar sorts of aggregations, polysaccharides are thought to have significant crop resistance (against both biotic and abiotic) enhancers and plant growth promotors (Muley et al., 2019). Chitosan, β-D-glucan, and other microbial polysaccharides have been found to have growth-stimulating and systemic disease resistance-inducing characteristics on cash crops such as Solanum lycopersicum, Hordeum vulgare, Solanum tuberosum, Saccharum officinarum, Gossypium herbaceum, and Glycine max (Uyen, 2014; Gandra et al., 2016; Blainski et al., 2018; Zhang et al., 2020).
The physical and biochemical traits of seedlings treated with GRH significantly improved. Proline levels and soluble sugar characteristics were also found to be altered, improving the stress-tolerating enzyme profile and enabling the plant to adapt to dry circumstances more successfully. The lower levels of MDA suggest that lipid peroxidation has significantly decreased and that membrane damage has become less frequent. The best concentration of EPS (i.e., Galactose rich heteropolysaccharide- GRH) for controlling the stressful condition was 50 ppm. Lower treatment doses are not sufficient to cause a noticeable change in the plantlets, whereas greater concentrations of GRH are likely to have a negative effect on the health of the plant. The findings of Sun et al. (2020b) are consistent with our findings because the 50 ppm EPS application was also determined to be the best-fitting one in that instance. Through a rise in endogenous ABA levels, polysaccharide treatments affect the stomatal physiology of the test plants’ leaves and cause partial stomatal closure, which minimises water evaporation. To improve the internal water levels of the tissues, which are essential for the plant’s proper growth and metabolism, these bioactive compounds function as anti-transpirant agents (Vishwakarma et al., 2017). Treatments with GRH increase the relative water contents of the tissues in this instance as well. The uniform build-up of rigid and highly water-soluble osmolytes (such as sugars, amino acids, and prolines) throughout the plant tissues, which provides subcellular stability and integrity, is another mechanism by which exopolysaccharide-mediated drought stress relief works (Hare et al., 1998; Krishnan et al., 2008). Increased levels of osmolyte aggregation raise osmotic pressure, which in turn induces higher water intake and insignificant water outflow. This keeps the cells’ critical osmotic pressure constant needed for optimum cell growth and division (Kaur and Asthir, 2015). The proline and soluble sugar levels (osmolytes) are increased by 50 ppm of GRH treatment in the current study as well, balancing the ideal subcellular environment for a healthy water weight required for cell growth. Proline is found to be an important osmo-regulator, and its exogenous administration increases hosts’ resistance to abiotic stress (Yoshiba et al., 1997; Ben Ahmed et al., 2010). For abiotically challenged plants, exogenous administration of water-soluble polysaccharides also demonstrates a similar response and causes a significant rise in proline levels (Yu et al., 2017; Zou et al., 2018). Thus, in this instance, GRH functions as a biological elicitor or priming agent that activates the cascades of biochemical processes required for water balance and antioxidant defence—ROS scavenging. Thus, as seen in the cases of rice, parsley, and tobacco, polysaccharides generate faster activation of transcription factors leading to the expression of defence-related genes, increasing the alleviation of drought stress (Conrath et al., 2002, Ortmann et al., 2006, Bozsoki et al., 2017). In drought-stricken areas, microbial exopolysaccharide enhances plant development by up- and down-regulating the expression of proline synthase and proline dehydrogenase, respectively (Sun et al., 2020a). By increasing SOD, POD, and CAT levels and fostering the effective operation of cellular biochemical machinery, which is essential for the survival of the plant, osmolytes also effectively eliminate harmful free radicals (reactive oxygen species) under drought stress (Sun et al., 2020b). The three key members of the antioxidant system SOD, CAT, and POD act in an integrated approach, where SOD acts as the first line of defence and converts superoxide free radicals to H2O2, which is further catalysed into water and oxygen by CAT and POD (Das and Roychoudhury, 2014). Last but not least, the MDA concentrations decrease, reducing the peroxidation of membrane proteins and lipids (Fu et al., 2010; Miller et al., 2010). Spraying potato and wheat with chitosan, polysaccharides from Ganoderma lucidum, Lactobacillus plantarum, and Pantoea agglomerans, respectively, activates the antioxidant defence cascades (Ortmann et al., 2006; Blainski et al., 2018; Muley et al., 2019; Zhang et al., 2020). Here, foliar GRH spray applied at a dosage of 50 ppm enhanced the SOD, POD, and CAT levels while concurrently lowering the MDA contents.
The bio-active GRH produced by Mucor sp. HELF2 was a polymer of D-galactose, L-fucose, and D-glucose (molar ratio—13:2:1) with a molecular weight of 2.98 × 105 Da. Galacto-rhamnan and beta-glucan exopolysaccharides with molecular weights of 1.87 × 105 and 2 × 105 Da were found in endophytic Fusarium sp. SD5 and Pestalotiopsis sp. BC55, respectively, according to Mahapatra and Banerjee’s reports from 2013 and 2016. Polysaccharides from edible mushroom Termitomyces heimii, and Meripilus giganteus also represents a similar type of monosaccharide compositions of L-fucose, D-galactose, and D-glucose, etc. (Maity et al., 2017, 2020). The EPS manufacturing process was optimised for carbon, nitrogen sources, a medium pH, and fermentation temperature to produce the greatest quantity of polysaccharides. The appropriate oxygen demand was also considered. The optimisation data makes it possible to quickly and affordably obtain the polysaccharides in large quantities. The results of the current inquiry on the optimization of GRH production broadly concur with those of Mahapatra and Banerjee (2013, 2016).
The present study examined endophytic exopolysaccharides (GRH) from an ecologically valuable plant and checked the drought tolerance action of the GRH on rice plants. Finally, varying concentrations of fungal EPS were used to reduce the effects of drought stress. Our research clarifies the idea of creating rice types resistant to drought through the external application of EPS, which supports environmentally friendly farming methods. This study provides the first evidence of the use of endophytic Mucor sp. HELF2 produced D-galactose-rich heteropolysaccharide to reduce drought stress in rice seedlings.
In the present study, rice seedling dehydration stress was lessened by a galactose-rich heteropolysaccharide derived from endophytic Mucor sp. HELF2. The outcome illustrated that treated plants had higher fresh weights, relative water contents, and chlorophyll levels. While the MDA concentration reduced, osmolytes such as soluble sugars, proline, as well as the antioxidant defence enzymes SOD, CAT, and POD, increased. The results support the conclusion that foliar spray of Galactose Rich Heteropolysaccharide efficiently promotes drought resistance in rice plants. GRH production was also optimised by adopting OVAT and RSM techniques and there was a 1.5-fold (20.10 g L−1) enhancement in GRH production in optimised fermentation conditions. The ability of GRH to alleviate the effects of drought stress on rice plants and the high yield of GRH makes it suitable for commercial exploitation. The current investigation’s findings may encourage sustainable farming methods and have an impact on the cultivation of crops in drought-prone areas.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/genbank/, ON146358.
HS designed and performed the experiments, and prepared the draft of the manuscript. DB designed the experiment and finalised the manuscript. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abdelaziz, M. E., Kim, D., Ali, S., Fedoroff, N. V., and Al-Babili, S. (2017). The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 263, 107–115. doi: 10.1016/j.plantsci.2017.07.006
Ali, R., Gul, H., Hamayun, M., Rauf, M., Iqbal, A., Shah, M., et al. (2021). Aspergillus awamori ameliorates the physicochemical characteristics and mineral profile of mung bean under salt stress. Chemical and Biological Technologies in Agriculture 8, 1–13. doi: 10.1186/s40538-021-00208-9
Ali, R., Gul, H., Rauf, M., Arif, M., Hamayun, M., Husna,, et al. (2022). growth-promoting endophytic fungus (Stemphylium lycopersici) ameliorates salt stress tolerance in maize by balancing ionic and metabolic status. Front. Plant Sci. 13:565. doi: 10.3389/fpls.2022.890565
Allen, C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 259, 660–684. doi: 10.1016/j.foreco.2009.09.001
Arndt, S. K., Irawan, A., and Sanders, G. J. (2015). Apoplastic water fraction and rehydration techniques introduce significant errors in measurements of relative water content and osmotic potential in plant leaves. Physiol. Plant. 155, 355–368. doi: 10.1111/ppl.12380
Asaf, L., Sajjad, M. H., Khan, A. L., Waqas, M., Khan, M. A., Jan, R., et al. (2018). Salt tolerance of Glycine max. L induced by endophytic fungus aspergillus flavus CSH1, via regulating its endogenous hormones and antioxidative system. Plant Physiol. Biochem. 128, 13–23. doi: 10.1016/j.plaphy.2018.05.007
Azad, K., and Kaminskyj, S. (2016). A fungal endophyte strategy for mitigating the effect of salt and drought stress on plant growth. Symbiosis 68, 73–78. doi: 10.1007/s13199-015-0370-y
Bagheri, A. A., Saadatmand, S., Niknam, V., Nejadsatari, T., and Babaeizad, V. (2013). Effect of endophytic fungus, Piriformospora indica, on growth and activity of antioxidant enzymes of rice (Oryza sativa L.) under salinity stress. Int. J. Adv. Biol. Biomed. Res. 1, 1337–1350.
Baltruschat, H., Fodor, J., Harrach, B. D., Niemczyk, E., Barna, B., Gullner, G., et al. (2008). Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. New Phytol. 180, 501–510. doi: 10.1111/j.1469-8137.2008.02583.x
Bates, L. S., Waldren, R. P., and Teare, I. D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39, 205–207. doi: 10.1007/BF00018060
Ben Ahmed, C., Ben Rouina, B., Sensoy, S., Boukhriss, M., and Ben Abdullah, F. (2010). Exogenous proline effects on photosynthetic performance and antioxidant defense system of young olive tree. J. Agric. Food Chem. 58, 4216–4222. doi: 10.1021/jf9041479
Bibi, N., Jan, G., Jan, F. G., Hamayun, M., Iqbal, A., Hussain, A., et al. (2019). Cochliobolus sp. acts as a biochemical modulator to alleviate salinity stress in okra plants. Plant Physiol. Biochem. 139, 459–469. doi: 10.1016/j.plaphy.2019.04.019
Blaha, D., Prigent-Combaret, C., Mirza, M. S., and Moënne-Loccoz, Y. (2006). Phylogeny of the 1-aminocyclopropane−1-carboxylic acid deaminase-encoding gene acd S in phytobeneficial and pathogenic Proteobacteria and relation with strain biogeography. FEMS Microbiol. Ecol. 56, 455–470. doi: 10.1111/j.1574-6941.2006.00082.x
Blainski,, da Rocha Neto, A. C., Schimidt, E. C., Voltolini, J. A., Rossi, M. J., di Piero, R. M., et al. (2018). Exopolysaccharides from lactobacillus plantarum induce biochemical and physiological alterations in tomato plant against bacterial spot. Appl. Microbiol. Biotechnol. 102, 4741–4753. doi: 10.1007/s00253-018-8946-0
Bouman, B. A., Humphreys, E., Tuong, T. P., and Barker, R. (2007). Rice and water. Adv. Agron. 92, 187–237. doi: 10.1016/S0065-2113(04)92004-4
Bouman, B. A. M., Peng, S., Castaneda, A. R., and Visperas, R. M. (2005). Yield and water use of irrigated tropical aerobic rice systems. Agric. Water Manag. 74, 87–105. doi: 10.1016/j.agwat.2004.11.007
Bozsoki, Zoltan, Cheng, Jeryl, Feng, Feng, Gysel, Kira, Vinther, Maria, Andersen, Kasper R., et al. “Receptor-mediated chitin perception in legume roots is functionally separable from nod factor perception.” Proceedings of the National Academy of Sciences, 114. (2017). E8118–E8127.
Chatterjee, S., Ghosh, S., and Mandal, N. C. (2022). Potential of an endophytic fungus Alternaria tenuissima PE2 isolated from Psidium guajava L. for the production of bioactive compounds. S. Afr. J. Bot. 150, 658–670. doi: 10.1016/j.sajb.2022.08.016
Chen, Y., Mao, W., Tao, H., Zhu, W., Qi, X., Chen, Y., et al. (2011). Structural characterization and antioxidant properties of an exopolysaccharide produced by the mangrove endophytic fungus aspergillus sp. Y16. Bioresour. Technol. 102, 8179–8184. doi: 10.1016/j.biortech.2011.06.048
Chen, C., Xin, K., Liu, H., Cheng, J., Shen, X., Wang, Y., et al. (2017). Pantoea alhagi, a novel endophytic bacterium with ability to improve growth and drought tolerance in wheat. Sci. Rep. 7, 1–14. doi: 10.1038/srep41564
Cherif, H., Marasco, R., Rolli, E., Ferjani, R., Fusi, M., Soussi, A., et al. (2015). Oasis desert farming selects environment-specific date palm root endophytic communities and cultivable bacteria that promote resistance to drought. Environ. Microbiol. Rep. 7, 668–678. doi: 10.1111/1758-2229.12304
Ciucanu, I., and Kerek, F. (1984). A simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 131, 209–217. doi: 10.1016/0008-6215(84)85242-8
Coleman-Derr, D., and Tringe, S. G. (2014). Building the crops of tomorrow: advantages of symbiont-based approaches to improving abiotic stress tolerance. Front. Microbiol. 5:283. doi: 10.3389/fmicb.2014.00283
Conrath, U., Pieterse, C. M. J., and Brigitte, M.-M. (2002). Priming in plant–pathogen interactions. Trends Plant Sci. 7, 210–216.
Constantin, M. E., Francisco, J. D. L., Babette, V. V., Martijn, R., and Frank, L. W. T. (2019). Endophyte-mediated resistance in tomato to Fusarium oxysporum is independent of ET, JA, and SA. Front. Plant Sci. 10:979.
Das, D., Mondal, S., Maiti, D., Roy, S. K., and Islam, S. S. (2009). Structural characterization of dietary fiber of green chalcumra (Benincasa hispida) fruit by NMR spectroscopic analysis. Nat. Prod. Commun. 4:1934578X0900400. doi: 10.1177/1934578X0900400421
Das, K., and Roychoudhury, A. (2014). Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2:53. doi: 10.3389/fenvs.2014.00053
Del Buono, D., Ioli, G., Nasini, L., and Proietti, P. (2011). A comparative study on the interference of two herbicides in wheat and Italian ryegrass and on their antioxidant activities and detoxification rates. J. Agric. Food Chem. 59, 12109–12115. doi: 10.1021/jf2026555
Dey, R., Lewis, S. C., Arblaster, J. M., and Abram, N. J. (2019). A review of past and projected changes in Australia’s rainfall. Wiley Interdiscip. Rev.: Climate Change 10:e577. doi: 10.1002/wcc.577
Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. T., and Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350–356. doi: 10.1021/ac60111a017
Fan, D., Subramanian, S., and Smith, D. L. (2020). Plant endophytes promote growth and alleviate salt stress in Arabidopsis thaliana. Sci. Rep. 10, 1–18. doi: 10.1038/s41598-020-69713-5
Fu, Q., Liu, C., Ding, N., Lin, Y., and Guo, B. (2010). Ameliorative effects of inoculation with the plant growth-promoting rhizobacterium pseudomonas sp. DW1 on growth of eggplant (Solanum melongena L.) seedlings under salt stress. Agric. Water Manag. 97, 1994–2000. doi: 10.1016/j.agwat.2010.02.003
Gamalero, G.-G. E., and Bernard, R. G. (2012). “Ethylene and abiotic stress tolerance in plants,” in Environmental adaptations and stress tolerance of plants in the era of climate change. (New York, NY: Springer). 395–412.
Gandra, J. R., Oliveira, E. R., Takiya, C. S., Goes, R. H. T. B., Paiva, P. G., Oliveira, K. M. P., et al. (2016). Chitosan improves the chemical composition, microbiological quality, and aerobic stability of sugarcane silage. Anim. Feed Sci. Technol. 214, 44–52. doi: 10.1016/j.anifeedsci.2016.02.020
González-Teuber, M., Contreras, R. A., Zúñiga, G. E., Barrera, D., and Bascuñán-Godoy, L. (2022). Synergistic association with root endophytic fungi improves morpho-physiological and biochemical responses of Chenopodium quinoa to salt stress. Front. Ecol. Evol. 9:787318. doi: 10.3389/fevo.2021.787318
Gupta, S., Schillaci, M., Walker, R., Smith, P., Watt, M., and Roessner, U. (2021). Alleviation of salinity stress in plants by endophytic plant-fungal symbiosis: current knowledge, perspectives and future directions. Plant Soil 461, 219–244. doi: 10.1007/s11104-020-04618-w
Hammami, H., Baptista, P., Martins, F., Gomes, T., Abdelly, C., and Mahmoud, O. M.-B. (2016). Impact of a natural soil salinity gradient on fungal endophytes in wild barley (Hordeum maritimum with.). World J. Microbiol. Biotechnol. 32, 1–11. doi: 10.1007/s11274-016-2142-0
Hare, P. D., Cress, W. A., and Van Staden, J. (1998). Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 21, 535–553. doi: 10.1046/j.1365-3040.1998.00309.x
Jha, Y., Subramanian, R. B., and Patel, S. (2011). Combination of endophytic and rhizospheric plant growth promoting rhizobacteria in Oryza sativa shows higher accumulation of osmoprotectant against saline stress. Acta Physiol. Plant. 33, 797–802. doi: 10.1007/s11738-010-0604-9
Jogawat, A., Saha, S., Bakshi, M., Dayaman, V., Kumar, M., Dua, M., et al. (2013). Piriformospora indica rescues growth diminution of rice seedlings during high salt stress. Plant Signal. Behav. 8:e26891. doi: 10.4161/psb.26891
Kaur, G., and Asthir, B. J. B. P. (2015). Proline: a key player in plant abiotic stress tolerance. Biol. Plant. 59, 609–619. doi: 10.1007/s10535-015-0549-3
Krishnan, N., Dickman, M. B., and Becker, D. F. (2008). Proline modulates the intracellular redox environment and protects mammalian cells against oxidative stress. Free Radic. Biol. Med. 44, 671–681. doi: 10.1016/j.freeradbiomed.2007.10.054
Laich, F., and Andrade, J. (2016). Penicillium pedernalense sp. nov., isolated from whiteleg shrimp heads waste compost. Int. J. Syst. Evol. 66, 4382–4388. doi: 10.1099/ijsem.0.001360
Landum, M. C., Félix, M. R., Alho, J., Garcia, R., Cabrita, M. J., Rei, F., et al. (2016). Antagonistic activity of fungi of Olea europaea L. against Colletotrichum acutatum. Microbiol. Res. 183, 100–108. doi: 10.1016/j.micres.2015.12.001
Lei, P., Pang, X., Feng, X., Li, S., Chi, B., Wang, R., et al. (2017). The microbe-secreted isopeptide poly-γ-glutamic acid induces stress tolerance in Brassica napus L. seedlings by activating crosstalk between H2O2 and Ca2+. Sci. Rep. 7, 1–15. doi: 10.1038/srep41618
Lei, P., Xu, Z., Ding, Y., Tang, B., Zhang, Y., Li, H., et al. (2015). Effect of poly (γ-glutamic acid) on the physiological responses and calcium signaling of rape seedlings (Brassica napus L.) under cold stress. J. Agric. Food Chem. 63, 10399–10406. doi: 10.1021/acs.jafc.5b04523
Li, L., Li, L., Wang, X., Zhu, P., Hongqing, W., and Qi, S. (2017). Plant growth-promoting endophyte Piriformospora indica alleviates salinity stress in Medicago truncatula. Plant Physiol. Biochem. 119, 211–223. doi: 10.1016/j.plaphy.2017.08.029
Lichtenthaler, H. K., and Wellburn, A. R. (1983). Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents, vol. 11, 591–592.
Livingston, D. P., Hincha, D. K., and Heyer, A. G. (2009). Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 66, 2007–2023. doi: 10.1007/s00018-009-0002-x
Lowry, O. H. (1951). Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193, 265–275. doi: 10.1016/S0021-9258(19)52451-6
Mahapatra, S., and Banerjee, D. (2013). Optimization of a bioactive exopolysaccharide production from endophytic fusarium solani SD5. Carbohydr. Polym. 97, 627–634. doi: 10.1016/j.carbpol.2013.05.039
Mahapatra, S., and Banerjee, D. (2016). Production and structural elucidation of exopolysaccharide from endophytic Pestalotiopsis sp. BC55. Int. J. Biol. Macromol. 82, 182–191. doi: 10.1016/j.ijbiomac.2015.11.035
Maity, P., Nandi, A. K., Manna, D. K., Pattanayak, M., Sen, I. K., Bhanja, S. K., et al. (2017). Structural characterization and antioxidant activity of a glucan from Meripilus giganteus. Carbohydr. Polym. 157, 1237–1245. doi: 10.1016/j.carbpol.2016.11.006
Maity, P., Nandi, A. K., Pattanayak, M., Manna, D. K., Sen, I. K., Chakraborty, I., et al. (2020). Structural characterization of a heteroglycan from an edible mushroom Termitomyces heimii. Int. J. Biol. Macromol. 151, 305–311. doi: 10.1016/j.ijbiomac.2020.02.120
Marasco, R., Rolli, E., Ettoumi, B., Vigani, G., Mapelli, F., Borin, S., et al. (2012). A drought resistance-promoting microbiome is selected by root system under desert farming. PloS one 7:e48479. doi: 10.1371/journal.pone.0048479
Mesa, J., Mateos-Naranjo, E., Caviedes, M. A., Redondo-Gómez, S., Pajuelo, E., and Rodríguez-Llorente, I. D. (2015). Endophytic cultivable bacteria of the metal bio-accumulator Spartina maritima improve plant growth but not metal uptake in polluted marshes soils. Front. Microbiol. 6:1450. doi: 10.3389/fmicb.2015.01450
Miller, G. A. D., Suzuki, N., Ciftci-Yilmaz, S. U. L. T. A. N., and Mittler, R. O. N. (2010). Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 33, 453–467. doi: 10.1111/j.1365-3040.2009.02041.x
Moghaddam, M. S. H., Naser, S., Jalal, S., and Niloufar, H.-D. (2021). Desert-adapted fungal endophytes induce salinity and drought stress resistance in model crops. Plant Physiol. Biochem. 160, 225–238.
Muley, A. B., Shingote, P. R., Patil, A. P., Dalvi, S. G., and Suprasanna, P. (2019). Gamma radiation degradation of chitosan for application in growth promotion and induction of stress tolerance in potato (Solanum tuberosum L.). Carbohydr. Polym. 210, 289–301. doi: 10.1016/j.carbpol.2019.01.056
Nadeem, S. M., Ahmad, M., Zahir, Z. A., Javaid, A., and Ashraf, M. (2014). The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 32, 429–448. doi: 10.1016/j.biotechadv.2013.12.005
Naumann, G., Alfieri, L., Wyser, K., Mentaschi, L., Betts, R. A., Carrao, H., et al. (2018). Global changes in drought conditions under different levels of warming. Geophys. Res. Lett. 45, 3285–3296. doi: 10.1002/2017GL076521
Ortmann, I., Conrath, U., and Moerschbacher, B. M. (2006). Exopolysaccharides of Pantoea agglomerans have different priming and eliciting activities in suspension-cultured cells of monocots and dicots. FEBS Lett. 580, 4491–4494. doi: 10.1016/j.febslet.2006.07.025
Paglia, E., and Parker, C. (2021). “The intergovernmental panel on climate change: guardian of climate science” in Guardians of public value. ed. A. Boin (Cham: Palgrave Macmillan), 295–321.
Pinedo, I., Ledger, T., Greve, M., and Poupin, M. J. (2015). Burkholderia phytofirmans PsJN induces long-term metabolic and transcriptional changes involved in Arabidopsis thaliana salt tolerance. Front. Plant Sci. 6:466. doi: 10.3389/fpls.2015.00466
Proestos, C., Boziaris, I. S., Nychas, G. J., and Komaitis, M. (2006). Analysis of flavonoids and phenolic acids in Greek aromatic plants: investigation of their antioxidant capacity and antimicrobial activity. Food Chem. 95, 664–671. doi: 10.1016/j.foodchem.2005.01.049
Redman, R. S., Kim, Y. O., Woodward, C. J. D. A., Greer, C., Espino, L., Doty, S. L., et al. (2011). Increased fitness of rice plants to abiotic stress via habitat adapted symbiosis: a strategy for mitigating impacts of climate change. PLoS One 6:e14823. doi: 10.1371/journal.pone.0014823
Rodriguez, R., and Redman, R. (2008). More than 400 million years of evolution and some plants still can’t make it on their own: plant stress tolerance via fungal symbiosis. J. Exp. Bot. 59, 1109–1114. doi: 10.1093/jxb/erm342
Rolli, E., Marasco, R., Vigani, G., Ettoumi, B., Mapelli, F., Deangelis, M. L., et al. (2015). Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 17, 316–331. doi: 10.1111/1462-2920.12439
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Santra, H. K., and Banerjee, D. (2022a). Production, optimization, characterization and drought stress resistance by β-glucan rich heteropolysaccharide from an endophytic fungi Colletotrichum alatae LCS1 isolated from clubmoss (Lycopodium clavatum). Front. Fungal Biol 2:65. doi: 10.3389/ffunb.2021.796010
Santra, H. K., and Banerjee, D. (2022b). Bioactivity study and metabolic profiling of Colletotrichum alatae LCS1, an endophyte of club moss Lycopodium clavatum L. PloS one 17:e0267302. doi: 10.1371/journal.pone.0267302
Santra, H. K., and Banerjee, D. (2022c). Broad-Spectrum antimicrobial action of cell-free culture extracts and volatile organic compounds produced by endophytic fungi Curvularia Eragrostidis. Front. Microbiol. 13:561. doi: 10.3389/fmicb.2022.920561
Santra, H. K., Maity, S., and Banerjee, D. (2022). Production of bioactive compounds with broad spectrum bactericidal action, bio-film inhibition and antilarval potential by the secondary metabolites of the endophytic fungus Cochliobolus sp. APS1 isolated from the Indian medicinal herb Andrographis paniculata. Molecules 27:1459. doi: 10.3390/molecules27051459
Schulz, B., Wanke, U., Draeger, S., and Aust, H-J. (1993). Endophytes from herbaceous plants and shrubs: effectiveness of surface sterilization methods. Mycol. Res. 97, 1447–1450.
Shahzad, R., Khan, A. L., Bilal, S., Waqas, M., Kang, S.-M., and Lee, I.-J. (2017). Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 136, 68–77. doi: 10.1016/j.envexpbot.2017.01.010
Sun, L., Lei, P., Wang, Q., Ma, J., Zhan, Y., Jiang, K., et al. (2020a). The endophyte Pantoea alhagi NX−11 alleviates salt stress damage to rice seedlings by secreting exopolysaccharides. Front. Microbiol. 10:3112. doi: 10.3389/fmicb.2019.03112
Sun, L., Yang, Y., Wang, R., Li, S., Qiu, Y., Lei, P., et al. (2020b). Effects of exopolysaccharide derived from Pantoea alhagi NX−11 on drought resistance of rice and its efficient fermentation preparation. Int. J. Biol. Macromol. 162, 946–955. doi: 10.1016/j.ijbiomac.2020.06.199
Tamura, K., Stecher, G., and Kumar, S. (2021). MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. doi: 10.1093/molbev/msab120
Tiwari, S., Lata, C., Chauhan, P. S., and Nautiyal, C. S. (2016). Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 99, 108–117. doi: 10.1016/j.plaphy.2015.11.001
Uga, Y., Sugimoto, K., Ogawa, S., Rane, J., Ishitani, M., Hara, N., et al. (2013). Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 45, 1097–1102. doi: 10.1038/ng.2725
Uyen, N. H. P. (2014). Radiation degradation of (1→3)-β-D-glucan from yeast with a potential application as a plant growth promoter. Int. J. Biol. Macromol. 69, 165–170. doi: 10.1016/j.ijbiomac.2014.05.041
Vishwakarma, K., Upadhyay, N., Kumar, N., Yadav, G., Singh, J., Mishra, R. K., et al. (2017). Abscisic acid signaling and abiotic stress tolerance in plants: a review on current knowledge and future prospects. Front. Plant Sci. 08:161. doi: 10.3389/fpls.2017.00161
Vurukonda, S. S. K. P., Vardharajula, S., Shrivastava, M., and SkZ, A. (2016). Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 184, 13–24. doi: 10.1016/j.micres.2015.12.003
Watanabe, S., Kojima, K., Ide, Y., and Sasaki, S. (2000). Effects of saline and osmotic stress on proline and sugar accumulation in Populus euphratica in vitro. Plant Cell Tissue Organ Cult. 63, 199–206. doi: 10.1023/A:1010619503680
Wonglumsom, W., Vishnubhatla, A., and Fung, D. Y. C. (2000). Effect of volume of liquid enrichment medium containing oxyrase® on growth of campylobacter jejuni 1. J RAPID METH AUT MIC 8, 111–139. doi: 10.1111/j.1745-4581.2000.tb00354.x
Yoshiba, Y., Kiyosue, T., Nakashima, K., Yamaguchi-Shinozaki, K., and Shinozaki, K. (1997). Regulation of levels of proline as an osmolyte in plants under water stress. Plant Cell Physiol. 38, 1095–1102. doi: 10.1093/oxfordjournals.pcp.a029093
Yu, Z., He, C., Jaime, A., da Silva, T., Zhang, G., Dong, W., et al. (2017). Molecular cloning and functional analysis of DoUGE related to water-soluble polysaccharides from Dendrobium officinale with enhanced abiotic stress tolerance. Plant Cell, Tissue and Organ Culture (PCTOC) 131, 579–599. doi: 10.1007/s11240-017-1308-2
Zhang, X., Aweya, J. J., Huang, Z. X., Kang, Z. Y., Bai, Z. H., Li, K. H., et al. (2020). In vitro fermentation of Gracilaria lemaneiformis sulfated polysaccharides and its Agaro-oligosaccharides by human fecal inocula and its impact on microbiota, Cheong. Carbohydr. Polym. 234:115894. doi: 10.1016/j.carbpol.2020.115894
Zhu, J. K. (2016). Abiotic stress signaling and responses in plants. Cells 167, 313–324. doi: 10.1016/j.cell.2016.08.029
Keywords: optimisation, drought stress alleviation, Mucor sp. HELF2, endophyte, hetero polysaccharide
Citation: Santra HK and Banerjee D (2023) Drought alleviation efficacy of a galactose rich polysaccharide isolated from endophytic Mucor sp. HELF2: A case study on rice plant. Front. Microbiol. 13:1064055. doi: 10.3389/fmicb.2022.1064055
Edited by:
Ravindra Soni, Indira Gandhi Krishi Vishva Vidyalaya, IndiaReviewed by:
Samy Selim, Al Jouf University, Saudi ArabiaCopyright © 2023 Santra and Banerjee. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Debdulal Banerjee, ✉ ZGVidTMzQGdtYWlsLmNvbQ==; ✉ ZGJAbWFpbC52aWR5YXNhZ2FyLmFjLmlu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.