
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 18 November 2022
Sec. Infectious Agents and Disease
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.1043596
This article is part of the Research TopicConference Research Topic: 16th International Symposium on SchistosomiasisView all 16 articles
 Silvia Gonçalves Mesquita1,2
Silvia Gonçalves Mesquita1,2 Elena Birgitta Lugli2
Elena Birgitta Lugli2 Giovanni Matera3
Giovanni Matera3 Cristina Toscano Fonseca4
Cristina Toscano Fonseca4 Roberta Lima Caldeira1*
Roberta Lima Caldeira1* Bonnie Webster2*
Bonnie Webster2*Background: Accurate diagnosis followed by timely treatment is an effective strategy for the prevention of complications together with reducing schistosomiasis transmission. Recombinase Polymerase Amplification (RPA) is a simple, rapid, sensitive, and specific isothermal method with low resource needs. This research aimed at the development and optimisation of a real-time (RT) and a lateral flow (LF) RPA assay for the detection of Schistosoma mansoni.
Methodology: Recombinase Polymerase Amplification reactions were performed at full- (as recommended) and half-volumes (to reduce costs), with RT or LF detection systems targeting the S. mansoni mitochondrial minisatellite region. The specificity was assessed using gDNA from other Schistosoma species, helminths co-endemic with S. mansoni, human stool, and urine, and Biomphalaria snail hosts. The analytical sensitivity was evaluated using serial dilutions of gDNA, synthetic copies of the target, and single eggs. The ability of both assays to detect the S. mansoni DNA in human urine and stool samples was also tested. The long-term stability of the RT-RPA reagents was evaluated by storing the reaction components in different temperature conditions for up to 3 weeks.
Results: The RT- and the LF-RPA (SmMIT- and SmMIT-LF-RPA, respectively) presented similar results when used full- and half-volumes, thus the latter was followed in all experiments. The SmMIT-RPA was 100% specific to S. mansoni, able to detect a single egg, with a limit of detection (LOD) of down to 1 fg of gDNA and one synthetic copy of the target. The assay was able to detect S. mansoni DNA from stool containing 1 egg/g and in spiked urine at a concentration of 10 fg/μl. SmMIT-RPA reagents were stable for up to 3 weeks when kept at 19°C, and 2 weeks when stored at 27°C. The SmMIT-LF-RPA cross-reacted with Clinostomidae, presented the LOD of 10 fg and one synthetic copy of the target, being able to detect a single egg and 1 egg/g in a stool sample. The LOD in spiked urine samples was 10 pg/μl.
Conclusion: The half-volume SmMIT-RPA is a promising method to be used in the field. It is specific, sensitive, robust, and tolerant to inhibitors, with a long-term stability of the reaction components and the real-time visualisation of results.
Schistosomiasis is a neglected tropical disease associated with poverty and low sanitation conditions, causing more than 240 million cases worldwide and 1.4 million disability-adjusted life years (DALYs). It is estimated that 779 million people currently live at risk of infection in tropical and subtropical regions (Kyu et al., 2018; Panzner, 2022; World Health Organization, 2022). The clinical manifestations of the disease can be urogenital or intestinal depending on the species that is causing the infection. Schistosoma mansoni is the species that causes intestinal disease in Africa and the Americas. The infection occurs when people have contact with watercourses contaminated with cercariae shed by Biomphalaria snails. The cercariae actively penetrate human skin, losing their tail, and migrating as schistosomula via blood vessels until establishing infection within the mesenteric veins as mature adult worms. After copulation, the female worm laid up to 300 eggs per day that can be released into the environment through the host’s faeces or become trapped in tissues, causing most of the chronic symptoms and complications of the disease (McManus et al., 2018; LoVerde, 2019; Nelwan, 2019). Nonspecific symptoms may occur in the early stage of the infection, such as fever, headache, fatigue, and myalgia and it is known as Katayama syndrome. Chronic schistosomiasis often produces gastrointestinal symptoms including diarrhea and abdominal pain, as well as hepatosplenic symptoms due to eggs lodged in the liver, e.g., fibrosis and portal hypertension (Colley et al., 2014). Less commonly, complications associated with ectopic migration of eggs can be observed in the brain and spinal cord (Vale et al., 2012).
The WHO Guidelines on Control and Elimination of Human Schistosomiasis were recently published. WASH (clean water, sanitation, and hygiene) and environmental interventions are highly recommended together with preventive chemotherapy (PC) by the mass drug administration (MDA) of Praziquantel targeting selected areas and groups (Lo et al., 2022; World Health Organization, 2022). MDA success is conditional to the precise assessment of schistosomiasis prevalence that will determine the appropriate strategy to be used (Utzinger et al., 2015). Therefore, estimating the true prevalence of schistosomiasis has a significant impact on the control and elimination measures (Turner et al., 2017).
The Kato-Katz (KK) technique is the method recommended by the WHO for the diagnosis of human intestinal schistosomiasis and it consists of the microscopic visualisation of eggs in the stool. This test is highly specific, cost-effective, and simple to perform, not needing much technological equipment other than the optic microscope (Katz et al., 1972). It has been extensively used for epidemiological surveys presenting a satisfactory performance in high prevalence settings. However, the KK’s sensitivity varies depending on the period of infection, daily fluctuation of egg excretion, uneven distribution of eggs in the stool, endemicity, and/or co-endemicity of the area (Bärenbold et al., 2017; Cavalcanti et al., 2019; Diego et al., 2021; Ogongo et al., 2022). These limitations are mainly observed in moderate and low endemic areas, where 25–30% of positive cases can be missed (Berhe et al., 2004; Enk et al., 2008; McManus et al., 2018). Schistosomiasis prevalence and intensity of infection has decreased in many endemic regions over the past years, especially due to MDA and WASH improvements (Katz, 2018; Brasil, 2021; Lo et al., 2022). Since the occurrence of light infections is becoming more frequent, the development and implementation of new diagnostic tools are highly needed (Utzinger et al., 2015).
Antigen tests based on the detection of circulating anodic and cathodic antigens (CAA and CCA, respectively) can be used for the indirect detection of S. mansoni using urine and serum samples. The detection of antigens can be performed using two types of lateral flow assays named POC-CCA (commercially available) and UCP-LF-CAA. Both assays are more sensitive than KK, in particular, the UCP-LF-CAA, which is the most sensitive and specific antigen test currently available (Sousa et al., 2019; Assare et al., 2021). However, there are limitations related to these tests. CCA detection does not work for urogenital schistosomiasis. It has been shown to give false-positive results (Graeff-Teixeira et al., 2021) with performance issues recently reported related to different kit batches (Viana et al., 2019) and with complicated interpretation of trace results (Coelho et al., 2016). Although the UCP-LF-CAA assay is very promising and covers all Schistosoma species (Corstjens et al., 2014), it currently needs bespoke laboratory based equipment with 24 h needed to obtain results (Sousa et al., 2019).
Molecular PCR-based methods have been extensively used for schistosomiasis detection due to the high sensitivity, specificity, and accuracy when compared to the KK technique (Pontes et al., 2003; Gomes et al., 2006, 2010; Cnops et al., 2012; Meurs et al., 2015; Frickmann et al., 2021; Siqueira et al., 2021). Despite having great advantages, the use of PCR-based methods is limited by the elevated cost and the need for advanced technological equipment and laboratory infrastructure, hampering large-scale implementation in endemic settings (Diego et al., 2021; Panzner, 2022).
Isothermal molecular methods stand out as promising alternatives to PCR for use at the point-of-care (POC)/point-of-need (PON). The loop-mediated isothermal amplification (LAMP) and the recombinase polymerase amplification (RPA) are the most common approaches used, providing fast and sensitive diagnosis and feasible in the field as they have low resource needs (Lobato and O’Sullivan, 2018; Li et al., 2021). Several LAMP assays have been developed for the detection of S. mansoni in both human and snail hosts over the past years, with favorable results (Abbasi et al., 2010; Hamburger et al., 2013; Fernández-Soto et al., 2014; Gandasegui et al., 2016, 2018; Caldeira et al., 2017; Mwangi et al., 2018; García-Bernalt Diego et al., 2019; Price et al., 2019; Mesquita et al., 2021). RPA was described in 2006 (Piepenburg et al., 2006) and since then it has been used mostly for the detection of Schistosoma haematobium (Rosser et al., 2015; Rostron et al., 2019; Archer et al., 2020, 2022; Frimpong et al., 2021) and Schistosoma japonicum (Sun et al., 2016; Xing et al., 2017; Guo et al., 2021; Deng et al., 2022), with only one study focused on S. mansoni targeting the ribosomal DNA (rDNA) regions 28S and the internal transcribed spacer (ITS; Poulton and Webster, 2018). Although this work represented a first and important step for the use of RPA for S. mansoni diagnosis, the lateral flow approaches lacked specificity with cross-reactivity with S. haematobium and Schistosoma bovis observed. One of the benefits of molecular based approaches is that they can be designed for different DNA biomarkers, allowing assays to be optimised to achieve high levels of sensitivity and specificity (Wang and Hu, 2014; Blasco-Costa et al., 2016). Once the isothermal molecular platform, such as LAMP and RPA, has been established and proved to work in the required settings then the molecular assays can be tailored to the need and sample type. This versatility of molecular platforms presents many cross-cutting opportunities and financial value.
Schistosoma mansoni molecular detection generally relies on stool samples, but urine, serum, and saliva can also be used for that purpose due to the presence of cell-free DNA (cfDNA; LoVerde, 2019). Each type of sample used as a source for DNA has its particularities. Stool samples are widely utilised as the source of both cfDNA and DNA from eggs, the latter often attached to a bead-beating and/or freezing step to facilitate egg disruption and DNA release (Pomari et al., 2019; Barda et al., 2020). Despite stool samples being non-invasive, they are inconvenient and require community sensitisation to ensure the collection of samples (Turner et al., 2017). Conversely, cfDNA presents great advantages as some bodily fluids such as urine are non-invasive, convenient, and usually easier to process, not requiring additional steps for sample preparation (Weerakoon and McManus, 2016). Sample type and sample preparation also have an effect on the downstream molecular assay to be used/tested. For example, PCR based approaches typically need samples that have been processed to remove inhibitors while isothermal assays are more tolerant to such inhibitors (Lobato and O’Sullivan, 2018). However, sample preparation and DNA extraction are currently among the factors that limit the use of molecular-based diagnostics in resource-poor settings and at the POC/PON due to the equipment requirements, costs, and time needs.
Moving from morbidity control to elimination as a public health problem requires more sensitive and specific diagnostic tests, especially to verify interruption of transmission, by detecting the infection in humans and snails (World Health Organization, 2022). Generally, as the demand for the novel test increase, its cost tends to decrease. Also, the cost-effectiveness of more accurate tests usually outweigh the actual cost of the test and the economic cost of the disease (Turner et al., 2017). For instance, it is estimated that in Brazil schistosomiasis generates annually a financial burden of nearly 41 million USD. More than 90% of that is related to indirect costs (e.g., loss of productivity and wages due to sick leave, hospitalisation, and premature deaths) that could be avoided by accurate diagnosis of infected people and timely treatment (Nascimento et al., 2019).
In this research, we developed and evaluated the performance of a real-time and a lateral flow RPA assay targeting the mitochondrial minisatellite region of S. mansoni to evaluate the diagnosis of the infection in humans and snails, especially in endemic areas where resources are limited.
For analytical sensitivity and specificity testing, genomic DNA (gDNA) from S. mansoni and other Schistosoma species (Schistosoma haematobium, Schistosoma curassoni, and Schistosoma bovis) were obtained from the Schistosomiasis Collection at The Natural History Museum (SCAN; Emery et al., 2012). Further analytical specificity was evaluated using gDNA from other non-Schistosoma samples including the intermediate hosts Biomphalaria glabrata, Biomphalaria tenagophila, Biomphalaria straminea, and trematodes commonly found infecting Biomphalaria snails belonging to the families Clinostomidae, Echinostomatidae, Notocotylidae, Spirorchiidae, and Strigeidae, all obtained from the Medical Malacology Collection at René Rachou Institute, Fiocruz Minas (Fiocruz-CMM) via the Trematodes Biology Laboratory from the Federal University of Minas Gerais (UFMG). The Helminthology and Medical Malacology Laboratory (HMM) from Fiocruz Minas provided gDNA from helminths co-endemic with S. mansoni including Ascaris lumbricoides, Ancylostomidae, Enterobius vermicularis, Trichuris trichiura, and Fasciola hepatica. Clinical stool samples were obtained under the Ethical Committee of Calabria Region approval (#108, 27 April 2017) and provided by the University “Magna Graecia” of Catanzaro. The data associated to all the specimens used in this study is provided on Supplementary material 1.
Recombinase Polymerase Amplification primers and the internal probe (TIB MolBio-Berlin, Germany) were designed targeting the S. mansoni mitochondrial minisatellite DNA region (GenBank accession number: L27240) following the guidelines from TwistDx™ (Cambridge, United Kingdom). To prevent the formation of primer-dimers, a phosphothioate backbone was added to the reverse primers for both the LF and RT assays (El Wahed et al., 2021) and the position of the 6-FAM and BHQ1 was reversed (compared to the design guidelines) within the RT probe. The primers were tested in-silico using BLAST (Altschul et al., 1990) to check the possibility of cross-reactivity. All primers and probes are described in Table 1 and shown in Figure 1.

Table 1. Mitochondrial primer and probe sequences designed for the LF-and RT-RPA.

Figure 1. Position of the Recombinase Polymerase Amplification (RPA) primers and probe within the Mitochondrion Schistosoma mansoni minisatellite DNA region. Bold sequences represent the primer sites and sequence in italics represents the probe site.
The RT SmMIT-RPA was performed using the TwistAmp® Exo Kit (TwistDX, Cambridge, United Kingdom). RPA reactions were set up as recommended by the manufacturer. Reactions were run in volumes of 50 μl (as recommended) or 25 μl (half-reactions; Supplementary material 2A). For the 50 μl reaction, a master mix was prepared containing the RPA rehydration buffer, water (if needed), forward and reverse primers, and the RT probe. This was then added to the lyophilised RPA pellet and homogenised by pipetting. The magnesium acetate (MgAc) was added to the lid of the RPA tube. Lastly, the DNA was added to the reaction tube. For the 25 μl reaction, a master mix was prepared and added to the lyophilised RPA pellet as above, and once homogenised it was split into two new tubes. The MgAc was added to the lid of each tube and finally, the DNA was added to the reaction. A positive (1 ng of S. mansoni gDNA) and negative (water) controls were included in all the runs. For both protocols, after the DNA addition, the tubes were quickly (~2 s) centrifuged, mixed (by inversion), then quickly (~2 s) centrifuged again. The centrifugation step mixes the MgAc with the other components which starts the RPA reaction. The tubes were incubated at 42°C for 20 min using the portable fluorometer AmpliFire (Douglas Scientific, Alexandria, MN, United States), with a manual mix after 4 min of incubation. Results could be seen in real-time via the device’s touchscreen and exported as an excel file for analysis of the raw data. Samples were considered positive if the amplification curve, normalised by the background level during the initial 4 min of the reaction, crossed the threshold of 346 relative fluorescence units (RFU). To determine the threshold value, we calculated the mean RFU observed in the first 4 min of the reaction of all non-S. mansoni samples used during the standardisation phase. The threshold was then set by three standard deviations of the calculated means, which was 346 RFU. All the protocol steps and the amount of each reagent are detailed in Figure 2 and Supplementary material 2A.
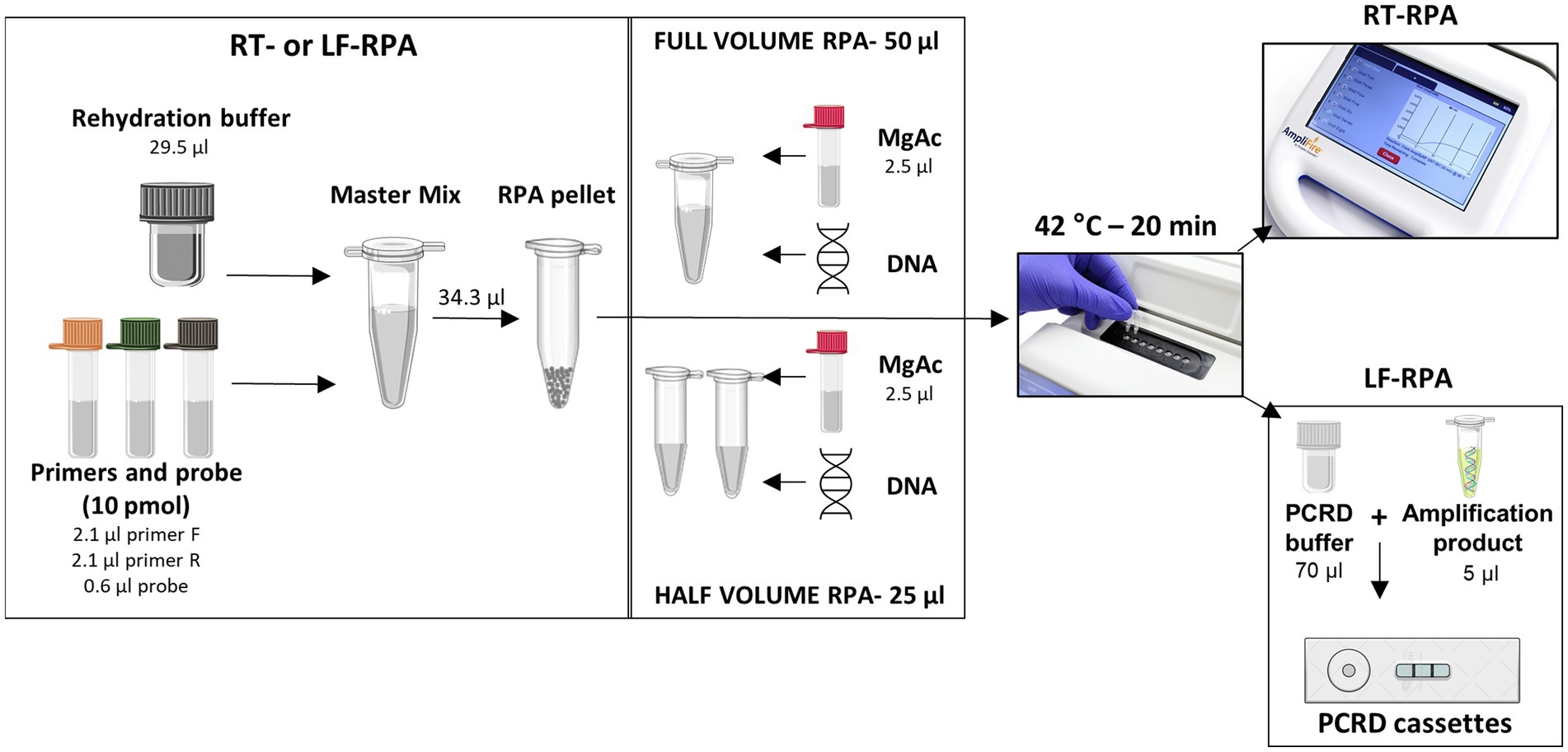
Figure 2. Set up of each RPA protocol used in this study. Images used in this figure were obtained at Agdia, Bioicons, and Mind the Graph websites.
The SmMIT-LF-RPA was performed using the TwistAmp® Nfo Kit (TwistDX, Cambridge, United Kingdom). The reaction was set up as described above and detailed in Figure 2 and Supplementary material 2B. A positive (1 ng of S. mansoni gDNA) and negative (water) controls were included in all the runs. After incubation, 5 μl of the amplification product were added to a 0.5 ml tube together with 70 μl of the PCRD extraction buffer. The mixture was then added to PCRD lateral flow cassettes. Results were observed after 10 min and any changes in the results after that period were not acknowledged. Positive samples presented two lines (both test and control lines) while just the control line was present for negative samples.
The LOD of the assays was evaluated using the samples as described below.
Serial 10-fold dilutions from 1 ng to 1 fg of S. mansoni gDNA were prepared from an adult worm extracted using the DNeasy Blood & Tissue kit (Qiagen, Hilden, Germany) at an original concentration of 3.8 ng/μl, measured by Nanodrop Spectrophotometer (ThermoFisher, Massachusetts, United States). Synthetic copies of the target region were obtained (gBlocks, IDT, Newark, New Jersey, United States) and diluted from 1 × 105 copies/μl down to 1 copy/μl. The analytical sensitivity of SmMIT-RPA was calculated by repeating 10 times the reaction using gDNA dilutions and three times using synthetic copies dilutions.
Single eggs of S. mansoni were provided by the Snail Schistosome Resource (SSR, Natural History Museum, United Kingdom-https://www.nhm.ac.uk/our-science/our-work/sustainability/schistosome-snail-resource.html) via the NIAID Schistosomiasis Resource Center (SRC, Biomedical Research Institute, United States-https://www.afbr-bri.org/schistosomiasis/). Individual eggs were isolated from a pool of eggs by capturing each one with a micropipette under a stereomicroscope. The DNA was then extracted using the SwiftX™ DNA kit (Xpedite Diagnostics, Germany) following protocol 1 from the manufacturer, with and without the heating step (Supplementary material 3). In brief, the protocol consists of the addition of buffer DL (50 μl) and magnetic beads (7.5 μl) to the tubes containing the single egg and incubate at room temperature (~19°C) or heated (95°C) for 5 min, followed by magnetic separation of the supernatant, which contains the extracted DNA. The direct addition of whole single eggs, with no DNA extraction procedure, in fresh (live eggs) and frozen conditions was also tested.
For both the LF and RT assays, the specificity was assessed against gDNA from other organisms that may be present in clinical and field samples. This included host DNA (snail host DNA and DNA from human urine and stool). gDNA from the snail hosts B. glabrata, B. straminea, and B. tenagophila were tested together with other trematodes that are commonly found infecting Biomphalaria snails in the neotropical region (Clinostomidae, Echinostomatidae, Notocotylidae, Spirorchiidae, and Strigeidae). Helminths of medical importance that are often co-endemic with S. mansoni (A. lumbricoides, Ancylostomidae, E. vermicularis, F. hepatica, and T. trichiura) were tested together with other commonly occurring Schistosoma species (S. bovis, S. curassoni, S. haematobium).
Stool samples that had been previously collected and characterised were used. One sample collected in Guinea Bissau, negative by both Kato-Katz and PCR methods, and one sample from Colombia, positive by Kato-Katz (1 egg/g) and by PCR. gDNA from these samples was extracted using the QIAamp DNA Stool Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol.
Seven aliquots of 100 μl of urine from a non-infected donor were spiked with different concentrations of S. mansoni gDNA with final concentrations within the samples ranging between 1 ng/μl and 1 fg/μl. Two aliquots of 100 μl of urine from the same donor were spiked with a pool of gDNA from medically important helminths (A. lumbricoides, Ancylostomidae, E. vermicularis, F. hepatica, and T. trichiura), with and without S. mansoni DNA. One aliquot of 100 μl of urine without the addition of any DNA was also used. The urine samples were filtered using Whatman® qualitative filters paper grade 3:6 μm (Sigma-Aldrich, St. Louis, Missouri, United States). The filters were dried at room temperature and 6 mm holes were made using a hole puncher. The paper holes were used for the DNA extraction using the QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany) and following the protocol described by Lodh et al. (2017).
Aiming to simulate point-of-need settings, we evaluated alternative storage conditions of the RT primers, probe, and kit reagents for up to 3 weeks. The reaction components were stored protected from light, at 19 and 27°C, mixed or separate. Reactions were conducted on days 0, 1, 2, and 3, and on weeks 1, 2, and 3, so we could evaluate the efficiency of the reaction over time.
Primers and probes targeting 184 bp of the mitochondrial S. mansoni minisatellite DNA region were manually designed as detailed in Table 1. In-silico specificity, based on BLAST (Altschul et al., 1990), was 100% specific for S mansoni.
Both real-time and lateral flow assays were successfully performed using full (50 μl) and half (25 μl) reaction volumes, keeping the final concentrations of each reaction component as in the original protocol (Table 2; Supplementary material 4). For that reason, the adapted protocol was followed in all experiments performed and the results presented from here on in were obtained by using half reaction volumes (25 μl), as detailed in Figure 2.
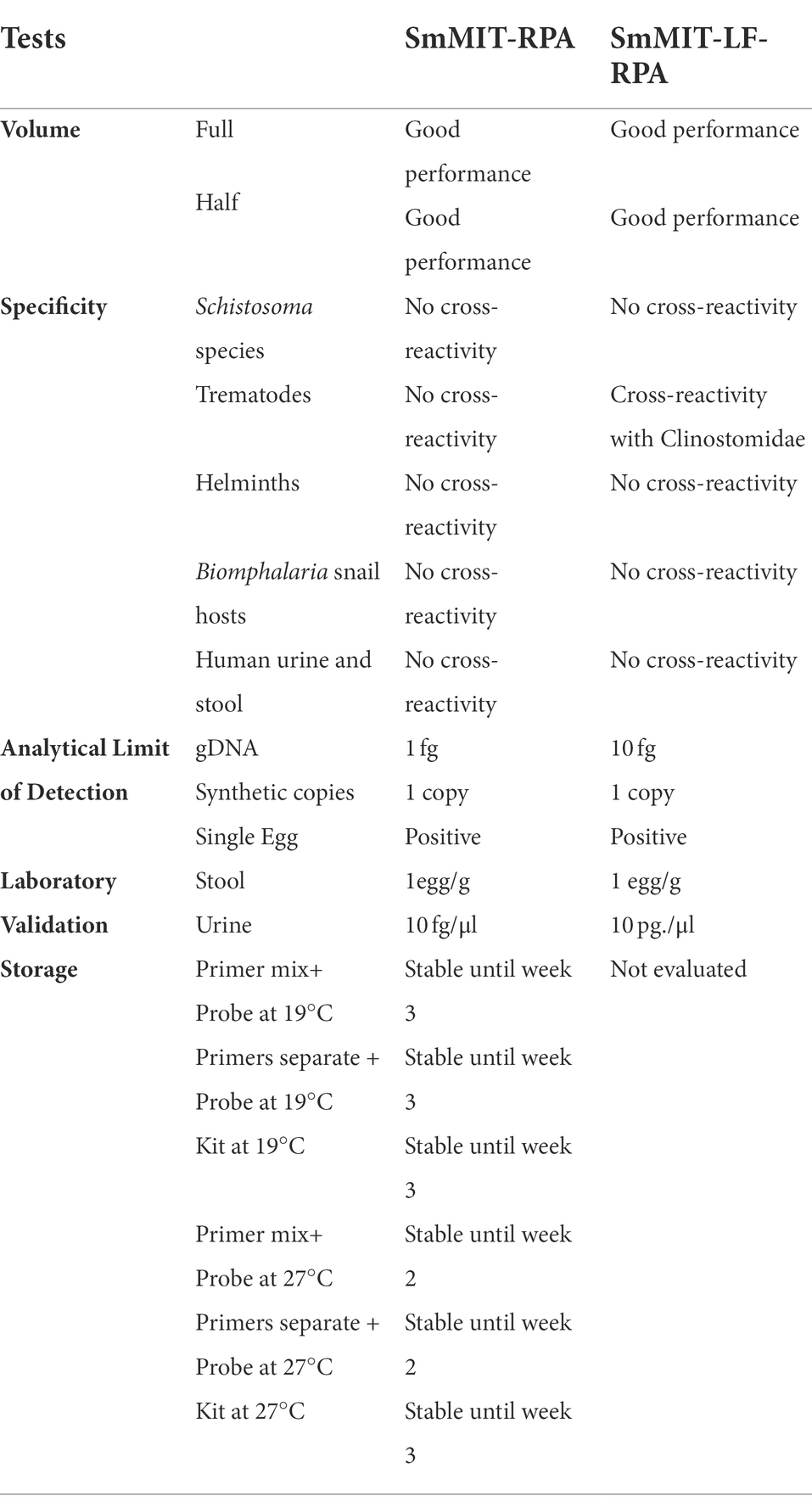
Table 2. The performance of SmMIT-and SmMIT-LF-RPA assays.
The SmMIT-RPA presented a high limit of detection being able to detect down to 1 fg of S. mansoni gDNA, one synthetic copy of the target, and a single S. mansoni egg in all conditions evaluated being: (i) DNA extracted with heated incubation; (ii) DNA extracted without heated incubation; (iii) fresh non-extracted egg; and (iv) frozen non-extracted egg (Figure 3; Table 2).

Figure 3. Limit of detection of the SmMIT-RPA. The assay was able to detect down to (A) 1 fg of the gDNA; (B) One synthetic copy of the target; (C) Single eggs extracted with the SwiftX™ DNA kit with or without the heating step, and single crude eggs, collected fresh and frozen (no DNA extraction procedure used). All graphs are displayed with the background baseline fluorescence subtracted. Legend: RFU-relative fluorescence units; ng-nanogram; pg-picogram, fg-femtogram; −ve-negative control (water).
The SmMIT-RPA assay was specific to S. mansoni with no cross-reactivity observed with snail or human DNA, DNA from other trematodes (including other Schistosoma species), and other helminths of medical importance (Figure 4; Table 2).
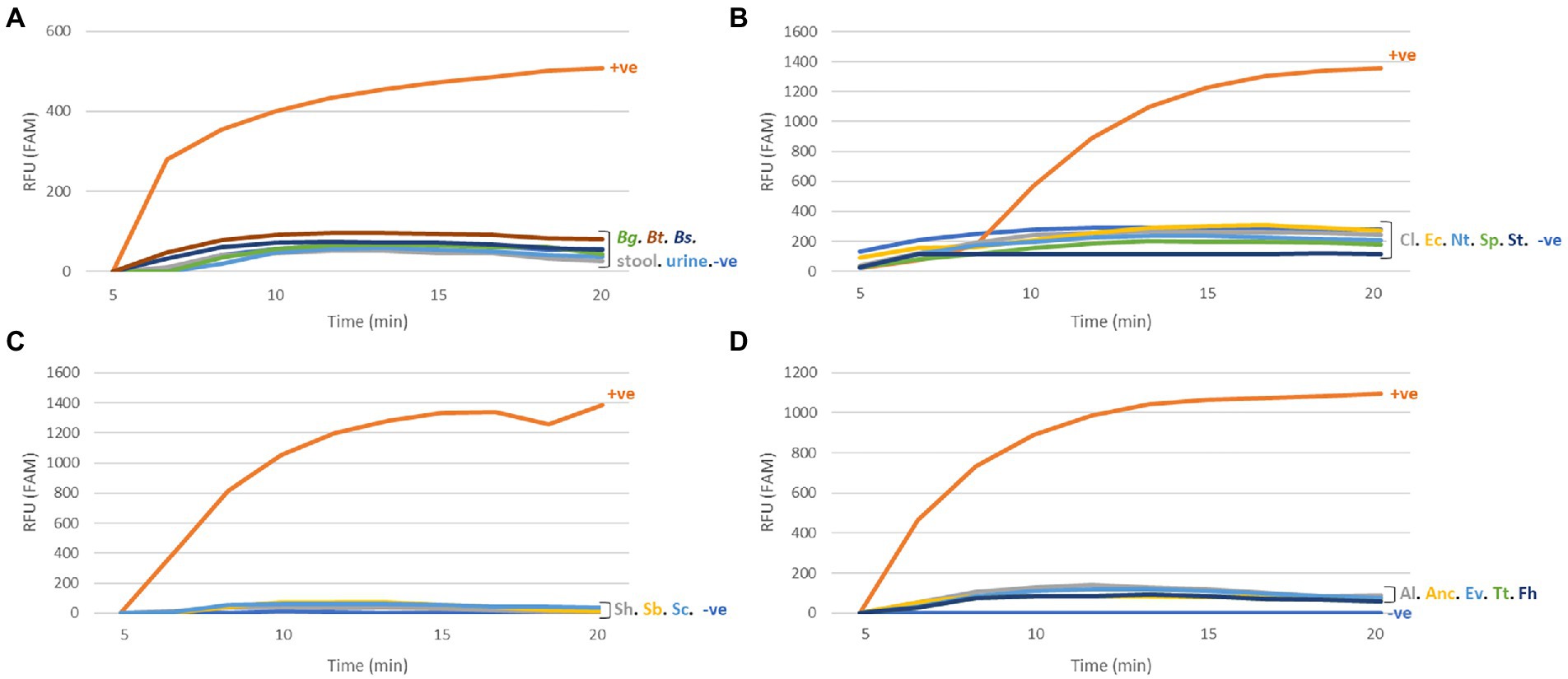
Figure 4. Specificity of the SmMIT-RPA. The assay was specific to Schistosoma mansoni with no cross-reactivity against (A) The host species; (B) Trematodes; (C) Schistosoma species; (D) Helminths of medical importance. All graphs are displayed with the background baseline fluorescence subtracted. Legend: RFU-relative fluorescence units; +ve-positive control (1 ng of S. mansoni gDNA); −ve-negative control (water); Bg, B. glabrata; Bt, B. tenagophila; Bs, B. straminea; Cl, Clinostomidae; Ec, Echinostomatidae; Nt, Notocotylidae; Sp, Spirorchiidae; St, Strigeidae; Sh, S. haematobium; Sb, S. bovis; Sc, S. curassoni; Al, A. lumbricoides; Anc, Ancylostomidae; Ev, E. vermicularis; Fh, F. hepatica; and Tt, T. trichiura.
The SmMIT-RPA presented a good performance when used on biological samples. Schistosoma mansoni DNA was detected in the positive stool sample known to contain 1 egg/g of stool (previously detected by the KK technique), and urine samples spiked with S. mansoni DNA at a final concentration of 10 fg/μl (Table 2; Supplementary material 5A) indicating high tolerance to molecular assay inhibitors present in urine (e.g., urea).
Our results showed that storing the primers and probe mixed together affected the assay performance giving false positive results. Keeping just the two primers mixed together or separately did not affect the RPA performance. However, the storage temperature influenced the results. Storing the primers and probe at 27°C reduced their longevity to 2 weeks, 1 week less than when stored at 19°C. The lyophilised RPA pellet, rehydration buffer, and MgAc produced consistent results after being stored for 3 weeks at both temperature conditions (19 and 27°C; Table 2). These results indicate that the SmMIT-RPA is a promising method able to produce robust results without the need for consistent cold chain for up to 3 weeks.
The SmMIT-LF-RPA detected down to one synthetic copy of the target region and one single egg either crude (frozen) or extracted with the SwiftX™ DNA kit (original protocol, i.e., with the heating step). The limit of detection of the assay using serial dilutions of the gDNA was 10 fg, 10-fold less sensitive than the SmMIT-RPA (Figure 5; Table 2).

Figure 5. Limit of detection of the SmMIT-LF-RPA. The assay was able to detect down to 10 fg of the gDNA, one copy of the synthetic target DNA, and one single egg with or without DNA extraction. Legend: ng, nanogram; pg, picogram, fg, femtogram; +ve, positive control (1 ng of Schistosoma mansoni gDNA); and −ve, negative control (water).
The SmMIT-LF-RPA assay showed high specificity for S. mansoni with no cross-reactivity against B. glabrata, B. straminea, B. tenagophila, human urine, and stool; nor was there cross-reactivity to other co-endemic human helminths and other trematodes belonging to the families Echinostomatidae, Notocotylidae, Spirorchiidae, Strigeidae, and three other Schistosoma species. However, cross-reactivity was observed with a trematode from the Clinostomidae family (Figure 6; Table 2).
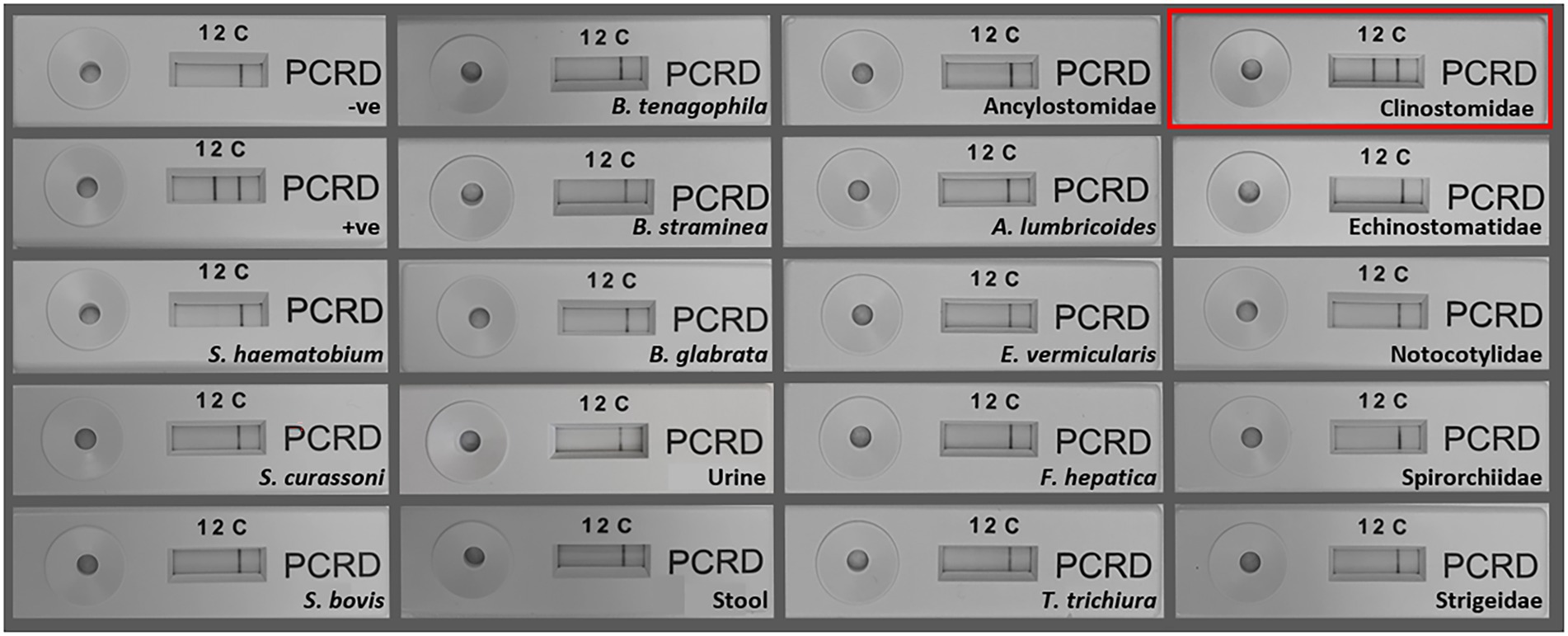
Figure 6. Specificity of the SmMIT-LF-RPA. The assay was specific to Schistosoma mansoni with no cross-reactivity with S. haematobium, S. bovis, S. curassoni, human stool, human urine B. glabrata, B. straminea, B. tenagophila, A. lumbricoides, Ancylostomidae, E. vermicularis, T. trichiura, F. hepatica, and trematodes belonging to the families, Echinostomatidae, Notocotylidae, Spirorchiidae, and Strigeidae. Cross-reactivity was observed for a Clinostomidae trematode (highlighted in red). Legend: +ve-positive control (1 ng of S. mansoni gDNA); −ve-negative control (water).
A loss in LF sensitivity was observed when spiked urine samples were analysed. SmMIT-LF-RPA was able to detect 10 pg/μl of S. mansoni DNA, a 1,000-fold decrease when compared to the SmMIT-RPA. The assay was able to detect the presence of the S. mansoni DNA in the positive stool sample (1 egg/g by the KK technique; Table 2; Supplementary material 5B).
Sensitive, specific, simple, and rapid diagnostic methods are essential to reach the World Health Organization (WHO) target of eliminating schistosomiasis as a public health problem (prevalence of heavy infections lower than 1%) by 2030 (WHO World Health Organization, 2022). To this end, Recombinase Polymerase Amplification (RPA) is an isothermal amplification method that presents great potential. It has been piloted for detection of urogenital schistosomiasis caused by Schistosoma haematobium (Rosser et al., 2015; Rostron et al., 2019; Archer et al., 2020; Frimpong et al., 2021) and intestinal schistosomiasis caused by Schistosoma japonicum (Sun et al., 2016; Xing et al., 2017; Guo et al., 2021; Deng et al., 2022), with promising results for its use in the field at the point-of-need (PON). Only one study has explored the development of a RPA assay to detect Schistosoma mansoni DNA, and this was done using the lateral flow RPA. The assay’s molecular targets were the 28S and ITS rDNA regions and although analytical sensitivity was good, the assays were found to be non-specific to S. mansoni with cross-reactivity observed with other Schistosoma species, namely S. haematobium and S. bovis (Poulton and Webster, 2018). The development of a RT-RPA targeting these ribosomal regions may be advantageous for a genus-specific assay. The genus or species level diagnostic specificity need for schistosomiasis, will depend on the diagnostic use case, with species level specificity detailed as a priority within the WHO schistosomiasis diagnostic target product profile (World Health Organization, 2021). In the present study, real-time fluorescence-based (RT) and lateral flow (LF) RPA assays were developed targeting the mitochondrial minisatellite region (termed here as SmMIT-and SmMIT-LF-RPA, respectively RPA) with higher sensitivity and specificity to S. mansoni, compared to the assays developed by Poulton and Webster (2018), with further potential for its use in endemic settings.
The developed SmMIT-RPA assay was 100% specific for S. mansoni in this study, proving to be a good alternative for the detection of S. mansoni infections in both human and snail hosts, without any cross-reactivity with other Schistosoma species and trematodes tested. It will be important to further test this specificity in terms of the genetic diversity of S. mansoni geographical strains, to make sure that specificity does not limit its ability to detect S. mansoni from all endemic areas (Webster et al., 2013). This is particularly relevant as the mitochondrial minisatellite target, used here, may be prone to high mutation rates due to its mitochondrial origins. It would also be relevant to check for specificity and cross-reactivity within co-infections, where inter-species interactions can lead to the production of hybrid offspring, e.g., S. mansoni/S. haematobium parthenogenic hybrids that have been observed to occur in co-endemic areas (Huyse et al., 2013). As the target used here is a mitochondrial target, only the maternal line of the interactions will be detected, however such hybrids are never found without the presence of the original species and so a specific diagnostic, such as the SmMIT-RPA, will still provide a diagnosis. The SmMIT-RPA assay should be further tested on Schistosoma rodhaini, the sister species to S. mansoni to check its ability to distinguish between S. mansoni, S. rodhaini and S. mansoni/S. rodhaini hybrids. This is particularly important for snail xenomonitoring of Biomphalaria snails, as S. rodhaini and S. mansoni/S. rodhaini are not implicated in human infections (Rey et al., 2021).
It was expected that the SmMIT-LF-RPA would show the same specificity as the primers and probes designed for both assays were very similar, except for the specific modifications required for each approach. However, cross-reactivity with cercariae belonging to the Clinostomidae family was observed only in the SmMIT-LF-RPA. Trematodes in the family Clinostomidae belong to the superfamily Schistosomatoidea together with the family Schistosomatidae, to which S. mansoni belongs. The close phylogenetic relationship between both families may reflect genomic similarities. RPA is known to be highly specific, but it can also tolerate the presence of a few nucleotide mismatches within the primer and probe regions, which can lead to false-positive results (Lobato and O’Sullivan, 2018), hence the need for rigorous specificity testing. Currently, there is no data available in public databases for the mitochondrial minisatellite region of non-schistosome trematodes, limiting the in-silico evaluation of the primers’ specificity. Species belonging to the Clinostomidae family have been reported parasitising Biomphalaria snails in Brazil, including the hosts species (B. glabrata, B. straminea, and B. tenagophila; Mesquita et al, 2020; Sousa et al., 2022). These trematodes are parasites of birds with fish being the second intermediate host. Human infections are rare and accidental due to the ingestion of raw fish. These infections have been reported in Asia and currently do not represent risks for human health in endemic areas for schistosomiasis (Lee et al., 2017; Kim et al., 2019). However, this cross-reactivity should be taken into account by the local malacological surveillance and schistosomiasis control program to measure the benefits and risks of using the SmMIT-LF-RPA for snail xenomonitoring. Further modifications of the primers and probe could be carried out to prevent the cross-reactivity with this non-Schistosoma trematode.
The SmMIT-RPA assay presented a high limit of detection (LOD) being able to detect down to 1 fg of S. mansoni DNA, with an analytical sensitivity of 20% at this level, but 100% for 1 ng. A reason for the low percentage of analytical sensitivity at low concentrations of S. mansoni DNA could be due to crowding agents in the RPA assay components. The crowding agents have an important role acting in the formation of the primer-recombinase complex (Piepenburg et al., 2006). However, they can also influence the reaction performance when there are low copies of the target due to its viscosity, which may be the reason for inconsistent results when low amounts of DNA were added to the reaction (Lobato and O’Sullivan, 2018). Moreover, analytical sensitivity does not always correspond to diagnostic sensitivity due to the nature of the starting material. When synthetic copies of the target were used, the LOD was 1 copy with an analytical sensitivity of 100%. This may be due to the higher purity of commercial samples compared to the gDNA dilutions obtained from adult worms’ extracts. In comparison to the LOD of the SmMIT-RPA (down to 1 fg), the SmMIT-LF-RPA presented a 10-fold reduction (down to 10 fg) in the assay’s analytical sensitivity. The difference in sensitivity between LF- and RT-RPA approaches has previously been reported for the assays developed for S. haematobium, with the LF assay able to detect 100 fg of the gDNA while the RT was able to detect down to 1 fg (Rosser et al., 2015; Rostron et al., 2019).
Both RPAs were able to detect the presence of DNA extracted using the SwiftX™ DNA kit from single S. mansoni eggs. This kit consists of the same components and steps as the discontinued Speed Xtract Nucleic Acid Kit (Qiagen, Hilden, Germany) and is a simple, fast, and efficient extraction method that requires few laboratory resources (e.g., pipettes and magnetic rack). A modification in the original protocol from the manufacturer was evaluated and the incubation of the sample was tested at both 95°C (as recommended) and at room temperature (~19°C), with the latter allowing to dispense with the requirement for a heating block that may rely on electricity. Both extraction conditions presented positive results. In addition, whole eggs (frozen and fresh) added directly to the reaction mix also produced positive results. This detection of S. mansoni DNA from single crude eggs reduces the requirements for sample preparation making both SmMIT-RPA and SmMIT-LF-RPA assays even more field-friendly while also reducing costs. However, clinical samples may present additional complications related to processing, egg disruption, and removal of inhibitors particularly related to stool samples. This has been demonstrated for other molecular assays where bead-beating and/or freezing prior to DNA extraction has been shown to increase DNA yields, improving the performance of molecular diagnostics (Pomari et al., 2019; Barda et al., 2020). The use of egg disruption strategies coupled with different DNA extraction methods needs to be further tested for RPA-based assays, particularly for clinical samples.
Recombinase Polymerase Amplification has been tested on multiple types of samples, e.g., urine, stool, blood, bodily fluids, and animal and plant products among others (Daher et al., 2016; Lobato and O’Sullivan, 2018). Moreover, the tolerance of the RPA reaction components to known PCR inhibitors found in clinical samples has been demonstrated in previous studies (Archer et al., 2022), including the direct addition of crude urine into the reaction mix (Rosser et al., 2015). The present study showed that both the LF-and RT-RPA approaches were able to detect S. mansoni DNA in clinical and spiked samples (stool and urine, respectively). Stool samples are the most used biological material for the molecular detection of S. mansoni, as it is the source of eggs and cfDNA. Promisingly, both LF-and RT-RPAs developed in this study were able to detect the infection in a stool sample having 1 egg/g that has previously been characterised as positive sample by KK and PCR. Several authors have also reported that urine samples can be used as the source of cfDNA of S. mansoni (Lodh et al., 2017; Fernández-Soto et al., 2019; Diab et al., 2021; Allam, 2022). In this study, urine samples were spiked with different amounts of gDNA to simulate the detection of cfDNA in clinical samples. As with testing of the gDNA standards, a difference was observed in the sensitivity of the SmMIT-RPA and SmMIT-LF-RPA, with the LF assay being less sensitive when using spiked urine samples. A 10-fold decrease in analytical sensitivity was observed for the LF-versus the RT-RPA assays using gDNA, whereas when the DNA was incorporated into the urine, there was a further loss of sensitivity for the LF assay (1,000-fold decrease). This may result from the RT-RPA having a higher tolerance to the presence of the inhibitors than the LF-RPA. However, additional studies should be conducted with larger numbers of clinical samples to investigate the effect of inhibitors on RPA outcome and whether they need to be removed.
The long-term stability of the lyophilised RT-RPA pellet and reagents has been previously evaluated by Chandu et al. (2016), proving the robustness of the reagents kept at −15–8°C for 1 year, and at 22–28°C for up to 6 months. Our findings confirm the stability of the reagents plus the primers and probe for 3 weeks when stored at 19°C and up to 2 weeks when stored at 27°C, agreeing with results previously obtained by Lillis et al., (2016). Infra-structure limitations in endemic areas may hamper the use of molecular methods. For this reason, the ability to work without dependence of cold chain storage facilitates the use of the SmMIT-RPA in these contexts. The longevity of all reagents for a period longer than 3 weeks or at a temperature higher than 27°C, as well as the impact of humidity, should be evaluated in the future.
Even though RPA presents many advantages in terms of performance and accuracy, the costs of the assay might be a limiting factor considering its application in endemic settings, where usually there is limited availability of financial resources. In general, a qPCR reaction costs 1.5 USD per sample (not including the cost of DNA extraction and the thermocycler; Archer et al., 2022), whereas the KK technique, which is the recommended diagnostic test by WHO and local health authorities, costs approximately 0.1–0.3 USD per sample (not including the cost of microscope and personnel; Speich et al., 2010). The cost of a full-volume SmMIT-RPA per sample is 6.98 USD meanwhile the half-volume reaction cost is 3.49 USD. The SmMIT-LF-RPA has a higher cost per sample given the price of the cassettes, being 9.08 USD for each full-volume reaction, and 5.84 USD, for the half-volume reaction. The half-volume SmMIT-RPA is the most cost-effective protocol among the ones assessed in this study. Though it is 2.3 times more expensive than qPCR, the SmMIT-RPA is simpler and faster, the results are easier to interpret, and it is more field-friendly. The costs of each RPA assay are detailed in Supplementary material 6 and does not include the costs of DNA extractions as well as the cost of the fluorometer and other equipment needed (e.g., pipette, centrifuge, and vortex). The findings from this study represent important progress on price reduction by using half of the reaction volume without compromising good and consistent results for the detection of S. mansoni. The use of a smaller reaction volume, than that recommended by the manufacturer, has been tested before aiming at the elimination of the mixing step during incubation (Lillis et al., 2016), and this should be tested further with this SmMIT-RPA assay. Further optimisation of the assay and large-scale use may reduce the cost per reaction. Moreover, the cost-benefits of implementing a more sensitive and accurate diagnostic test should be considered. Accurate diagnosis enables timely treatment, the prevention of long-term complications and financial losses due to lack of productivity and sick leaves, as well as reducing the potential of emergence of new transmission foci (Turner et al., 2017). Thus, the impact of using a more expensive diagnostic test will be smaller in the long run given the benefits of its implementation.
The SmMIT-RPA presented a better performance overall when compared to the SmMIT-LF-RPA. Besides being more specific and sensitive, the SmMIT-RPA is advantageous as real-time visualisation of results does not require opening the reaction tubes after DNA amplification. RPA can amplify up to 104-fold the target in 10 min (Daher et al., 2016) and assays requiring end-point visualisation of the result can be prone to cross-contamination among the samples. Therefore, until a closed system is developed and available for the LF assay, there will be a risk of contamination when this approach is used.
Determining the precise prevalence of schistosomiasis in a specific area is conditional to a sensitive, specific, reproducible, and accessible diagnostic method. However, diagnostics improvements are very much needed for detection of light infections in low prevalence settings, and for the verification of transmission interruption (Ogongo et al., 2022; World Health Organization, 2022). If good diagnostic tools are not available, the true prevalence of a specific area may be underestimated limiting the efficiency of schistosomiasis control programs (Panzner, 2022). The promising results obtained in this study suggest that the SmMIT-RPA may allow for a more accurate and rapid diagnosis of schistosomiasis and therefore may influence the decision-making processes involved in determining appropriate destination of public funding aimed at the elimination of schistosomiasis as a public health problem by 2030. Additional analysis will be conducted in order to validate the use of this assay for the detection of S. mansoni in clinical and field samples from endemic areas.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
BW, CF, RC, and SM: conceptualization. BW, RC, and SM: methodology and project administration. BW and SM: validation and data curation. SM: formal analysis, writing—original draft, and visualization. BW, EL, and SM: investigation. BW, GM, and RC: resources. BW, CF, EL, GM, and RC: writing—review and editing. BW, CF, and RC: supervision. BW, CF, and SM: funding acquisition. All authors contributed to the article and approved the submitted version.
This research has been financed by the Royal Society of Tropical Medicine and Hygiene (RSTMH) via the Small Grant Programme and was partially supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES - Finance Code 001; SM is funded by CAPES via the PrINT programme, and Vice-Presidência de Educação, Informação e Comunicação (VPEIC-Fiocruz); CF is funded by CNPq Fellowship (grant number 303131/2018-7); and support was also provided via the Natural History Museum’s Departmental Investment Fund and Conselho Nacional de Desenvolvimento Científico/Programa de Excelência em Pesquisa—Pesquisa e Ensaios Clínicos (PROEP/PEC; 420685/2017-0).
The authors would like to thank the Wellcome Trust funded Schistosome and Snail Resource (SSR; Wellcome Trust Biomedical Resource Grant 221368/Z/20/Z 2021-2026), NIAID Schistosomiasis Resource Center (SRC; NIH-NIAID Contract HHSN272201700014I), Schistosomiasis Collection at The Natural History Museum (SCAN), Medical Malacology Collection (Fiocruz-CMM), Helminthology, Medical Malacology Research Group from Fiocruz Minas (HMM), Trematodes Biology Laboratory from the Federal University of Minas Gerais (UFMG), and University “Magna Graecia” of Catanzaro for providing the samples used in this study; Agdia for the authorisation for using the images available on their website to create Figure 2; Bioicons (bioicons.com), Mind the Graph (mindthegraph.com), and SMART-Servier Medical Art (smart.servier.com) for the images used to create Figures 2 and Supplementary material 3; and Andy Wende and Ludovic Ebert from Xpedite Diagnostics for their assistance on the designing of DNA extraction assays.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1043596/full#supplementary-material
Abbasi, I., King, C. H., Muchiri, E. M., and Hamburger, J. (2010). Detection of Schistosoma mansoni and Schistosoma haematobium DNA by loop-mediated isothermal amplification: identification of infected snails from early prepatency. Am. J. Trop. Med. Hyg. 83, 427–432. doi: 10.4269/ajtmh.2010.09-0764
Allam, A. F. (2022). Performance of loop-mediated isothermal amplification (LAMP) for detection of Schistosoma mansoni infection compared with Kato–Katz and real-time PCR. J. Helminthol. 96:e28. doi: 10.1017/S0022149X22000153
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Archer, J., Barksby, R., Pennance, T., Rostron, P., Bakar, F., Knopp, S., et al. (2020). Analytical and clinical assessment of a portable, isothermal Recombinase polymerase amplification (RPA) assay for the molecular diagnosis of urogenital schistosomiasis. Molecules 25, 1–15. doi: 10.3390/molecules25184175
Archer, J., Patwary, F. K., Sturt, A. S., Webb, E. L., Phiri, C. R., Mweene, T., et al. (2022). Validation of the isothermal Schistosoma haematobium Recombinase polymerase amplification (RPA) assay, coupled with simplified sample preparation, for diagnosing female genital schistosomiasis using cervicovaginal lavage and vaginal self-swab samples. PLoS Negl. Trop. Dis. 16:e0010276. doi: 10.1371/journal.pntd.0010276
Assare, R. K., Tra-Bi, M. I., Coulibaly, J. T., Corstjens, P. L. A. M., Ouattara, M., Hurlimann, E., et al. (2021). Accuracy of two circulating antigen tests for the diagnosis and surveillance of Schistosoma mansoni infection in low-endemicity settings of Côte d’Ivoire. Am. J. Trop. Med. Hyg. 105, 677–683. doi: 10.4269/ajtmh.21-0031
Barda, B., Schindler, C., Wampfler, R., Ame, S., Ali, S. M., and Keiser, J. (2020). Comparison of real-time PCR and the Kato-Katz method for the diagnosis of soil-transmitted helminthiasis and assessment of cure in a randomized controlled trial. BMC Microbiol. 20, 1–8. doi: 10.1186/s12866-020-01963-9
Bärenbold, O., Raso, G., Coulibaly, J. T., N’Goran, E. K., Utzinger, J., and Vounatsou, P. (2017). Estimating sensitivity of the Kato-Katz technique for the diagnosis of Schistosoma mansoni and hookworm in relation to infection intensity. PLoS Negl. Trop. Dis. 11, 1–14. doi: 10.1371/journal.pntd.0005953
Berhe, N., Medhin, G., Erko, B., Smith, T., Gedamu, S., Bereded, D., et al. (2004). Variations in helminth fecal egg counts in Kato-Katz thick smears and their implications in assessing infection status with Schistosoma mansoni. Acta Trop. 92, 205–212. doi: 10.1016/j.actatropica.2004.06.011
Blasco-Costa, I., Cutmore, S. C., Miller, T. L., and Nolan, M. J. (2016). Molecular approaches to trematode systematics: ‘best practice’ and implications for future study. Syst. Parasitol. 93, 295–306. doi: 10.1007/s11230-016-9631-2
Brasil, M.D.S. (2021). Boletim Epidemiológio-Doenças Tropicais Negligenciadas. Available at: https://www.gov.br/saude/pt-br/media/pdf/2021/marco/3/boletim_especial_doencas_negligenciadas.pdf
Caldeira, R. L., Jannotti-Passos, L. K., and Dos Santos, C. O. (2017). Use of molecular methods for the rapid mass detection of Schistosoma mansoni (Platyhelminthes: Trematoda) in Biomphalaria spp. (Gastropoda: Planorbidae). J. Trop. Med. 2017, 1–6. doi: 10.1155/2017/8628971
Cavalcanti, M. G., Cunha, A. F. A., and Peralta, J. M. (2019). The advances in molecular and new point-of-care (POC) diagnosis of schistosomiasis pre-and post-praziquantel use-and the pursuit of more reliable approaches for low endemic and non-endemic areas. Front. Immunol. 10, 1–10. doi: 10.3389/fimmu.2019.00858
Chandu, D., Paul, S., Parker, M., Dudin, Y., King-Sitzes, J., Perez, T., et al. (2016). Development of a rapid point-of-use DNA test for the screening of Genuity® roundup ready 2 yield® soybean in seed samples. Biomed. Res. Int. 2016, 1–12. doi: 10.1155/2016/3145921
Cnops, L., Tannich, E., Polman, K., Clerinx, J., and Van Esbroeck, M. (2012). Schistosoma real-time PCR as diagnostic tool for international travellers and migrants. Trop. Med. Int. Health 17, 1208–1216. doi: 10.1111/j.1365-3156.2012.03060.x
Coelho, P. M. Z., Siqueira, L. M. V., Grenfell, R. F. Q., Almeida, N. B. F., Katz, N., Almeida, Á., et al. (2016). Improvement of POC-CCA interpretation by using Lyophilization of urine from patients with Schistosoma mansoni low worm burden: towards an elimination of doubts about the concept of trace. PLoS Negl. Trop. Dis. 10, 1–13. doi: 10.1371/journal.pntd.0004778
Colley, D. G., Bustinduy, A. L., Secor, W. E., and King, C. H. (2014). Human schistosomiasis. Lancet 383, 2253–2264. doi: 10.1016/S0140-6736(13)61949-2
Corstjens, P., De Dood, C. J., Kornelis, D., Tjon, K. F. E., Wilson, R. A., Kakiuki, T. M., et al. (2014). Tools for diagnosis, monitoring and screening of Schistosoma infections utilizing lateral-flow based assays and upconverting phosphor labels. Parasitology 141, 1841–1855. doi: 10.1017/S0031182014000626
Daher, R. K., Stewart, G., Boissinot, M., and Bergeron, M. G. (2016). Recombinase polymerase amplification for diagnostic applications. Clin. Chem. 62, 947–958. doi: 10.1373/clinchem.2015.245829
Deng, W., Wang, S., Wang, L., Lv, C., Li, Y., Feng, T., et al. (2022). Laboratory evaluation of a basic Recombinase polymerase amplification (RPA) assay for early detection of Schistosoma japonicum. Pathogens 11:319. doi: 10.3390/pathogens11030319
Diab, R. G., Tolba, M. M., Ghazala, R. A., Abu-Sheasha, G. A., Webster, B. L., and Mady, R. F. (2021). Intestinal schistosomiasis: can a urine sample decide the infection? Parasitol. Int. 80:102201. doi: 10.1016/j.parint.2020.102201
Diego, J. G. B., Fernández-Soto, P., Febrer-Sendra, B., Crego-Vicente, B., and Muro, A. (2021). Loop-mediated isothermal amplification in schistosomiasis. J. Clin. Med. 10, 1–13. doi: 10.3390/jcm10030511
El Wahed, A. A., Patel, P., Maier, M., Pietsch, C., Rüster, D., Böhlken-Fascher, S., et al. (2021). Suitcase lab for rapid detection of SARS-CoV-2 based on Recombinase polymerase amplification assay. Anal. Chem. 93, 2627–2634. doi: 10.1021/acs.analchem.0c04779
Emery, A., Allan, F., Rabone, M., and Rollinson, D. (2012). Schistosomiasis collection at the natural history museum (SCAN). Parasit. Vectors 5:185. doi: 10.1186/1756-3305-5-185
Enk, M. J., Lima, A. C. L., Drummond, S. C., Schall, V. T., and Coelho, P. M. Z. (2008). The effect of the number of stool samples on the observed prevalence and the infection intensity with Schistosoma mansoni among a population in an area of low transmission. Acta Trop. 108, 222–228. doi: 10.1016/j.actatropica.2008.09.016
Fernández-Soto, P., Gandasegui Arahuetes, J., Sánchez Hernández, A., López Abán, J., Vicente Santiago, B., and Muro, A. (2014). A loop-mediated isothermal amplification (LAMP) assay for early detection of Schistosoma mansoni in stool samples: A diagnostic approach in a murine model. PLoS Negl. Trop. Dis. 8:e3126. doi: 10.1371/journal.pntd.0003126
Fernández-Soto, P., Gandasegui, J., Rodríguez, C. C., Pérez-Arellano, J. L., Crego-Vicente, B., Diego, J. G. B., et al. (2019). Detection of Schistosoma mansoni-derived DNA in human urine samples by loop-mediated isothermal amplification (LAMP). PLoS One 14:e0214125. doi: 10.1371/journal.pone.0214125
Frickmann, H., Lunardon, L. M., Hahn, A., Loderstädt, U., Lindner, A. K., Becker, S. L., et al. (2021). Evaluation of a duplex real-time PCR in human serum for simultaneous detection and differentiation of Schistosoma mansoni and Schistosoma haematobium infections – cross-sectional study. Travel Med. Infect. Dis. 41:102035. doi: 10.1016/j.tmaid.2021.102035
Frimpong, M., Kyei-Tuffuor, L., Fondjo, L. A., Ahor, H. S., Adjei-Kusi, P., Maiga-Ascofare, O., et al. (2021). Evaluation of a real-time recombinase polymerase amplification assay for rapid detection of Schistosoma haematobium infection in resource-limited setting. Acta Trop. 216:105847. doi: 10.1016/j.actatropica.2021.105847
Gandasegui, J., Fernández-Soto, P., Hernández-Goenaga, J., López-Abán, J., Vicente, B., and Muro, A. (2016). Biompha-LAMP: A new rapid loop-mediated isothermal amplification assay for detecting Schistosoma mansoni in Biomphalaria glabrata snail host. PLoS Negl. Trop. Dis. 10:e05225. doi: 10.1371/journal.pntd.0005225
Gandasegui, J., Fernández-Soto, P., Muro, A., Simões Barbosa, C., Lopes de Melo, F., Loyo, R., et al. (2018). A field survey using LAMP assay for detection of Schistosoma mansoni in a low-transmission area of schistosomiasis in Umbuzeiro, Brazil: assessment in human and snail samples. PLoS Negl. Trop. Dis. 12, 1–16. doi: 10.1371/journal.pntd.0006314
García-Bernalt Diego, J., Fernández-Soto, P., Crego-Vicente, B., Alonso-Castrillejo, S., Febrer-Sendra, B., Gómez-Sánchez, A., et al. (2019). Progress in loop-mediated isothermal amplification assay for detection of Schistosoma mansoni DNA: towards a ready-to-use test. Sci. Rep. 9, 1–11. doi: 10.1038/s41598-019-51342-2
Gomes, L. I., Marques, L. H. D. S., Enk, M. J., de Oliveira, M. C., Coelho, P. M. Z., and Rabello, A. (2010). Development and evaluation of a sensitive PCR-ELISA system for detection of Schistosoma infection in feces. PLoS Negl. Trop. Dis. 4, 1–8. doi: 10.1371/journal.pntd.0000664
Gomes, A. L. D. V., Melo, F. L., Werkhauser, R. P., and Abath, F. G. C. (2006). Development of a real time polymerase chain reaction for quantitation of Schistosoma mansoni DNA. Mem. Inst. Oswaldo Cruz 101, 133–136. doi: 10.1590/S0074-02762006000900021
Graeff-Teixeira, C., Favero, V., Pascoal, V. F., de Souza, R. P., Rigo, F. D. V., LHD, A., et al. (2021). Low specificity of point-of-care circulating cathodic antigen (POC-CCA) diagnostic test in a non-endemic area for schistosomiasis mansoni in Brazil. Acta Trop. :217. doi: 10.1016/j.actatropica.2021.105863
Guo, Q., Zhou, K., Chen, C., Yue, Y., Shang, Z., Zhou, K., et al. (2021). Development of a Recombinase polymerase amplification assay for Schistosomiasis japonica diagnosis in the experimental mice and domestic goats. Front. Cell. Infect. Microbiol. 11, 1–12. doi: 10.3389/fcimb.2021.791997
Hamburger, J., Abbasi, I., Kariuki, C., Wanjala, A., Mzungu, E., Mungai, P., et al. (2013). Evaluation of loop-mediated isothermal amplification suitable for molecular monitoring of schistosome-infected snails in field laboratories. Am. J. Trop. Med. Hyg. 88, 344–351. doi: 10.4269/ajtmh.2012.12-0208
Huyse, T., Van den Broeck, F., Hellemans, B., Volckaert, F. A. M., and Polman, K. (2013). Hybridisation between the two major African schistosome species of humans. Int. J. Parasitol. 43, 687–689. doi: 10.1016/j.ijpara.2013.04.001
Katz, N. (2018). Inquérito Nacional de Prevalência da Esquistossomose mansoni e Geo-helmintoses. 76.
Katz, N., Chaves, A., and Pellegrino, J. (1972). A simple device for quantitative stool thick-smear technique in schistosomiasis mansoni. Rev. do Inst. Med. Trop. São Paulo 14, 397–400.
Kim, H., Cho, S. W., Oh, H., and Byeon, H. K. (2019). A case of unexpected Clinostomum complanatum infection initially presenting as foreign body in pharynx. Kor. J. Parasitol. 57, 175–177. doi: 10.3347/kjp.2019.57.2.175
Kyu, H. H., Abate, D., Abate, K. H., Abay, S. M., Abbafati, C., Abbasi, N., et al. (2018). Global, regional, and national disability-adjusted life-years (DALYs) for 359 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990-2017: A systematic analysis for the global burden of disease study 2017. Lancet 392, 1859–1922. doi: 10.1016/S0140-6736(18)32335-3
Lee, G. S., Park, S. W., Kim, J., Seo, K. S., You, K. W., Chung, J. H., et al. (2017). A case of endoscopically treated laryngopharyngitis resulting form Clinostomum complanatum infectio. Kor. J. Gastroenterol. 69, 177–180. doi: 10.4166/kjg.2017.69.3.177
Li, H. M., Qin, Z. Q., Bergquist, R., Qian, M. B., Xia, S., Lv, S., et al. (2021). Nucleic acid amplification techniques for the detection of Schistosoma mansoni infection in humans and the intermediate snail host: a structured review and meta-analysis of diagnostic accuracy. Int. J. Infect. Dis. 112, 152–164. doi: 10.1016/j.ijid.2021.08.061
Lillis, L., Siverson, J., Lee, A., Cantera, J., Parker, M., Piepenburg, O., et al. (2016). Factors influencing Recombinase polymerase amplification (RPA) assay outcomes at point of care. Mol. Cell. Probes 30, 74–78. doi: 10.1016/j.mcp.2016.01.009
Lo, N. C., Bezerra, F. S. M., Colley, D. G., Fleming, F. M., Homeida, M., Kabatereine, N., et al. (2022). Review of 2022 WHO guidelines on the control and elimination of schistosomiasis. Lancet Infect. Dis. 3099. doi: 10.1016/S1473-3099(22)00221-3
Lobato, I. M., and O’Sullivan, C. K. (2018). Recombinase polymerase amplification: basics, applications and recent advances. Trends Anal. Chem. 98, 19–35. doi: 10.1016/j.trac.2017.10.015
Lodh, N., Mikita, K., Bosompem, K. M., Anyan, W. K., Quartey, J. K., Otchere, J., et al. (2017). Point of care diagnosis of multiple schistosome parasites: species-specific DNA detection in urine by loop-mediated isothermal amplification (LAMP). Acta Trop. 173, 125–129. doi: 10.1016/j.actatropica.2017.06.015
LoVerde, P. T. (2019). Schistosomiasis. Adv. Exp. Med. Biol. 1154, 45–70. doi: 10.1007/978-3-030-18616-6_3
McManus, D. P., Dunne, D. W., Sacko, M., Utzinger, J., and Vennervald, B. J. (2018). Zhou X-N. Schistosomiasis. Nat. Rev. Dis. Prim. 4:13. doi: 10.1038/s41572-018-0013-8
Mesquita, S. G., FGDS, N., RGC, S., ODS, C., Fonseca, C. T., and Caldeira, R. L. (2021). A loop-mediated isothermal amplification assay for Schistosoma mansoni detection in Biomphalaria spp. from schistosomiasis-endemic areas in Minas Gerais, Brazil. Parasit. Vectors 14, 388–312. doi: 10.1186/s13071-021-04888-y
Mesquita, S. G., Rodrigues-Luiz, G. F., Reis-Cunha, J. L., Cardoso, M. S., De Mendonça, C. L. F., Bueno, L. L., et al. (2020). A multiplex PCR protocol for rapid differential identification of four families of trematodes with medical and veterinary importance transmitted by Biomphalaria Preston. Acta Trop. 2020:211.
Meurs, L., Brienen, E., Mbow, M., Ochola, E. A., Mboup, S., Karanj, D. M. S., et al. (2015). Is PCR the next reference standard for the diagnosis of Schistosoma in stool? A comparison with microscopy in Senegal and Kenya. PLoS Negl. Trop. Dis. 9:e03959. doi: 10.1371/journal.pntd.0003959
Mwangi, I. N., Agola, E. L., Mugambi, R. M., Shiraho, E. A., and Mkoji, G. M. (2018). Development and evaluation of a loop-mediated isothermal amplification assay for diagnosis of Schistosoma mansoni infection in Fecal samples. J. Parasitol. Res. 2018, 1–7. doi: 10.1155/2018/1267826
Nascimento, G. L., Pegado, H. M., Domingues, A. L. C., De Alencar Ximenes, R. A., Itria, A., Cruz, L. N., et al. (2019). The cost of a disease targeted for elimination in Brazil: the case of schistosomiasis mansoni. Mem. Inst. Oswaldo Cruz 114, 1–9. doi: 10.1590/0074-02760180347
Nelwan, M. L. (2019). Schistosomiasis: Life cycle, diagnosis, and control. Curr. Ther. Res. Clin. Exp. 91, 5–9. doi: 10.1016/j.curtheres.2019.06.001
Ogongo, P., Nyakundi, R. K., Chege, G. K., and Ochola, L. (2022). The road to elimination: Current state of Schistosomiasis research and Progress towards the end game. Front. Immunol. 13, 1–23. doi: 10.3389/fimmu.2022.846108
Panzner, U. (2022). Clinical applications of isothermal diagnosis for human Schistosomiasis. Encyclopedia, 2, 690–704. doi: 10.3390/encyclopedia2020048
Piepenburg, O., Williams, C. H., Stemple, D. L., and Armes, N. A. (2006). DNA detection using recombination proteins. PLoS Biol. 4, 1115–1121. doi: 10.1371/journal.pbio.0040204
Pomari, E., Perandin, F., La Marca, G., and Bisoffi, Z. (2019). Improved detection of DNA Schistosoma haematobium from eggs extracted by bead beating in urine. Parasitol. Res. 118, 683–686. doi: 10.1007/s00436-018-6137-7
Pontes, L. A., Oliveira, M. C., Katz, N., Dias-Neto, E., and Rabello, A. (2003). Comparison of a polymerase chain reaction and the Kato-Katz technique for diagnosing infection with Schistosoma mansoni. Am. J. Trop. Med. Hyg. 68, 652–656. doi: 10.4269/ajtmh.2003.68.652
Poulton, K., and Webster, B. (2018). Development of a lateral flow recombinase polymerase assay for the diagnosis of Schistosoma mansoni infections. Anal. Biochem. 546, 65–71. doi: 10.1016/j.ab.2018.01.031
Price, M., Cyrs, A., Sikasunge, C. S., Mwansa, J., and Lodh, N. (2019). Testing the infection prevalence of Schistosoma mansoni after mass drug administration by comparing sensitivity and specificity of species-specific repeat fragment amplification by PCR and loop-mediated isothermal amplification. Am. J. Trop. Med. Hyg. 101, 78–83. doi: 10.4269/ajtmh.19-0121
Rey, O., Webster, B. L., Huyse, T., Rollinson, D., Van den Broeck, F., Kincaid-Smith, J., et al. (2021). Population genetics of African Schistosoma species. Infect. Genet. Evol. 89:104727. doi: 10.1016/j.meegid.2021.104727
Rosser, A., Rollinson, D., Forrest, M., and Webster, B. L. (2015). Isothermal Recombinase polymerase amplification (RPA) of Schistosoma haematobium DNA and oligochromatographic lateral flow detection. Parasit. Vectors 8, 446–445. doi: 10.1186/s13071-015-1055-3
Rostron, P., Pennance, T., Bakar, F., Rollinson, D., Knopp, S., Allan, F., et al. (2019). Development of a recombinase polymerase amplification (RPA) fluorescence assay for the detection of Schistosoma haematobium. Parasit. Vectors 12, 514–517. doi: 10.1186/s13071-019-3755-6
Siqueira, L. M. V., Senra, C., de Oliveira, A. A., NFDF, C., Gomes, L. I., Rabello, A., et al. (2021). A real-time PCR assay for the diagnosis of intestinal Schistosomiasis and cure assessment after the treatment of individuals with low parasite burden. Front. Immunol. 11, 1–13. doi: 10.3389/fimmu.2020.620417
Sousa, D. G. S., Carvalho-Costa, F. A., Monteiro, K. J. L., Silva, E. L., Castro, E. S., Sousa, R. L. T., et al. (2022). Larval trematodes hosted by Biomphalaria straminea in the Brazilian semiarid region: implications for schistosomiasis control. Trop. Biomed. 39, 1–10. doi: 10.47665/tb.39.1.003
Sousa, M. S., Van Dam, G. J., Pinheiro, M. C. C., De Dood, C. J., Peralta, J. M., Peralta, R. H. S., et al. (2019). Performance of an ultra-sensitive assay targeting the circulating anodic antigen (CAA) for detection of Schistosoma mansoni infection in a low endemic area in Brazil. Front. Immunol. 10:682. doi: 10.3389/fimmu.2019.00682
Speich, B., Knopp, S., Mohammed, K. A., Khamis, I. S., Rinaldi, L., Cringoli, G., et al. (2010). Comparative cost assessment of the Kato-Katz and FLOTAC techniques for soil-transmitted helminth diagnosis in epidemiological surveys. Parasit. Vectors 3, 1–11. doi: 10.1186/1756-3305-3-71
Sun, K., Xing, W., Yu, X., Fu, W., Wang, Y., Zou, M., et al. (2016). Recombinase polymerase amplification combined with a lateral flow dipstick for rapid and visual detection of Schistosoma japonicum. Parasit. Vectors 9, 476–479. doi: 10.1186/s13071-016-1745-5
Turner, H. C., Bettis, A. A., Dunn, J. C., Whitton, J. M., Hollingsworth, T. D., Fleming, F. M., et al. (2017). Economic considerations for moving beyond the Kato-Katz technique for diagnosing intestinal parasites as we move towards elimination. Trends Parasitol. 33, 435–443. doi: 10.1016/j.pt.2017.01.007
Utzinger, J., Becker, S. L., van Lieshout, L., van Dam, G. J., and Knopp, S. (2015). New diagnostic tools in schistosomiasis. Clin. Microbiol. Infect. 21, 529–542. doi: 10.1016/j.cmi.2015.03.014
Vale, T. C., de Sousa-Pereira, S. R., Ribas, J. G. R., and Lambertucci, J. R. (2012). Neuroschistosomiasis mansoni. Neurologist 18, 333–342. doi: 10.1097/NRL.0b013e3182704d1e
Viana, A. G., Gazzinelli-Guimarães, P. H., Castro, V. N. D., Santos, Y. L. D. O. D., Ruas, A. C. L., Bezerra, F. S. D. M., et al. (2019). Discrepancy between batches and impact on the sensitivity of point-of-care circulating cathodic antigen tests for Schistosoma mansoni infection. Acta Trop. 197:105049. doi: 10.1016/j.actatropica.2019.105049
Wang, S., and Hu, W. (2014). Development of “-omics” research in Schistosoma spp. and-omics-based new diagnostic tools for schistosomiasis. Front. Microbiol. 5:313. doi: 10.3389/fmicb.2014.00313
Webster, B. L., Webster, J. P., Gouvras, A. N., Garba, A., Lamine, M. S., Diaw, O. T., et al. (2013). DNA ‘barcoding’ of Schistosoma mansoni across sub-Saharan Africa supports substantial within locality diversity and geographical separation of genotypes. Acta Trop. 128, 250–260. doi: 10.1016/j.actatropica.2012.08.009
Weerakoon, K. G., and McManus, D. P. (2016). Cell-free DNA as a diagnostic tool for human parasitic infections. Trends Parasitol. 32, 378–391. doi: 10.1016/j.pt.2016.01.006
WHO World Health Organization (2020). Ending the neglected to attain the sustainable development goals-A road map for neglected tropical diseases 2021–2030. Geneva. 55
World Health Organization (2022). Diagnostic target product profiles for monitoring, evaluation and surveillance of schistosomiasis control programmes. 28. Available at: https://www.who.int/publications/i/item/9789240031104
World Health Organization (2022). WHO GUIDELINE on control and elimination of human schistosomiasis. 142 p.
Keywords: Schistosomiasis, Schistosoma mansoni, Recombinase Polymerase Amplification, isothermal molecular diagnostics, mitochondrial minisatellite region, real-time RPA, lateral flow RPA
Citation: Mesquita SG, Lugli EB, Matera G, Fonseca CT, Caldeira RL and Webster B (2022) Development of real-time and lateral flow recombinase polymerase amplification assays for rapid detection of Schistosoma mansoni. Front. Microbiol. 13:1043596. doi: 10.3389/fmicb.2022.1043596
Edited by:
Michal Letek, Universidad de León, SpainReviewed by:
Wang Jiangyong, South China Sea Fisheries Research Institute (CAFS), ChinaCopyright © 2022 Mesquita, Lugli, Matera, Fonseca, Caldeira and Webster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bonnie Webster, Yi53ZWJzdGVyQG5obS5hYy51aw==; Roberta Lima Caldeira, cm9iZXJ0YS5jYWxkZWlyYUBmaW9jcnV6LmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.