 Xianbo Jia
Xianbo Jia Ke Zhao2†
Ke Zhao2† Jichen Chen
Jichen Chen- 1Institute of Soil and Fertilizer, Fujian Academy of Agricultural Sciences, Fujian Key Laboratory of Plant Nutrition and Fertilizer, Fuzhou, China
- 2College of Resources and Environment, Fujian Agriculture and Forestry University, Fuzhou, China
- 3College of Life Sciences, Fujian Agriculture and Forestry University, Fuzhou, China
Prodigiosin is a promising secondary metabolite mainly produced by Serratia marcescens. The production of prodigiosin by S. marcescens is regulated by different kinds of regulatory systems, including the EnvZ/OmpR system. In this study, we demonstrated that the regulatory factor OmpR positively regulated prodigiosin production in S. marcescens FZSF02 by directly binding to the promoter region of the prodigiosin biosynthesis cluster with a lacZ reporter assay and electrophoretic mobility shift assay (EMSA). The binding sequence with the pig promoter was identified by a DNase I footprinting assay. We further demonstrate that OmpR regulates its own expression by directly binding to the promoter region of envZ/ompR. For the first time, the regulatory mechanism of prodigiosin production by the transcriptional factor OmpR was revealed.
Introduction
Prodigiosin, an important secondary metabolite produced by S. marcescens and some other bacteria, is of particular interest for its potential applications, including various pharmacological activities, food colorants, and potential sunscreens (Stankovic et al., 2014; Darshan and Manonmani, 2015). In the genus Serratia, biosynthesis of prodigiosin is regulated not only by external factors, including temperature, pH and medium composition (Paul et al., 2020) but also by various genes (Williamson et al., 2006). Nearly 30 genes have been reported to be involved in prodigiosin biosynthesis in S. marcescens (Pan et al., 2021), and more studies should be carried out to search for new regulatory genes and uncover the complex regulatory mechanisms of this secondary metabolite.
The two-component system is a family of signal transduction proteins reported to be present in all types of life (Stock et al., 1999; Scharf, 2010; Papon and Stock, 2019). In bacteria, the classical two-component system consists of a sensor protein and a regulatory protein (Yuchuan et al., 2019). Sensor proteins respond to chemical or physical signals by phosphorylating regulatory proteins, and phosphorylated regulatory proteins can regulate the expression levels of downstream genes by binding to certain DNA sequences (Groisman, 2016). Two-component systems regulate many processes of bacteria, such as adaptation to environmental changes: osmolarity (Boyce et al., 2016), temperature (Dhiman et al., 2015; Najnin et al., 2016), oxygen (Dixon, 1998; Wright et al., 2018), regulation of developmental pathways, and behaviors, such as sporulation (Sarwar and Garza, 2015), biofilm formation (Lai et al., 2005), quorum sensing (Kruppa et al., 2004), regulation of secondary metabolite biosynthesis (Sola-Landa et al., 2003), virulence (Beier and Gross, 2006), and antibiotic resistance (Bhagirath et al., 2019; Tierney and Rather, 2019). Biosynthesis of prodigiosin was also regulated by different types of two component systems in various Serratia strains, including PigQ/W and PhoB/PhoR in Serratia 39,006 (Fineran et al., 2005; Gristwood et al., 2009); EepR/EepS in S. marcescens CMS376, S. marcescens K904, and S. marcescens Nima (Stella et al., 2015); RssB/RssA in S. marcescens CH-1 (Horng et al., 2010); and CpxR/A in S. marcescens FS14 (Qiu et al., 2021).
The two component system EnvZ/OmpR is an important signal transduction system in bacteria responding to various environmental stress and growth conditions (Qin et al., 2001). We have previously demonstrated that when envZ or ompR was knocked out, S. marcescens FZSF02 lost its prodigiosin biosynthesis ability (Jia et al., 2021), and OmpR was also recently found to control prodigiosin biosynthesis in S. marcescens JNB5-1 (Pan et al., 2022). However, the regulatory mechanism of the two-component EnvZ/OmpR system on prodigiosin production is still unknown.
In this study, with LacZ-reporter studies and an electrophoretic mobility shift assay (EMSA), we demonstrated that OmpR positively regulated prodigiosin biosynthesis by directly binding to the promoter region of the prodigiosin biosynthesis gene cluster. We also found that OmpR can regulate its own expression level by binding the promoter of the EnvZ/OmpR genes.
Materials and methods
Bacterial strains, plasmids, and culture conditions
The bacterial strains and plasmids used in this study are listed in Supplementary Tables S1, S2. Serratia marcescens FZSF02 and its related mutants were incubated in lysogeny broth (LB) at 28°C and 180 rpm. E. coli DH5a and E. coli Rosetta (DE3) were cultured in LB medium at 37°C and 220 rpm. The final concentrations of antibiotics used in this study were as follows: 100 mg/L ampicillin, 100 mg/L kanamycin, and 50 mg/L chloramphenicol.
Construction of the in-frame deletion mutant and complementary strains
FZSF02∆ompR and FZSF02∆envZ were constructed in our previous study (Jia et al., 2021). FZSF02∆envZ∆ompR was also constructed with the homologous recombination method (Jia et al., 2021). Complementary strains were constructed with the plasmid pRK415 as we have reported in our previous study (Jia et al., 2021). pRK415-ompR, pRK415-envZ and pRK415-∆envZ∆ompR were transformed into FZSF02∆ompR, FZSF02∆envZ, and FZSF02∆envZ∆ompR, respectively, to construct the complementary strains. The primers used in these experiments are listed in Supplementary Table S2.
lacZ reporter assays
The pig promoter and ompR promoter were inserted upstream of lacZ in the plasmid pTOPO-lacZ-Cmr to construct the plasmids pTOPO-Pigpro-lacZ-Cmr and pTOPO-OmpRpro-lacZ-Cmr. The primers used in this experiment are listed in Supplementary Table S2. The constructed plasmids with the Pig promoter and ompR promoter were transformed into the wild-type strain S. marcescens FZSF02 and the ompR-knockout strain FZSF02∆ompR, respectively. The plasmids pTOPO-Pigpro-Cmr and pTOPO-ompRpro-Cmr were also transformed into S. marcescens FZSF02 and FZSF02∆ompR as controls. For liquid β-galactosidase assays, constructed strains were cultured at 28°C and 180 rpm for 16 h, and enzyme activities were measured in sonicated extracts according to the method described by Pardee et al. (1959).
Electrophoretic mobility shift assay
The coding sequence of ompR was cloned into pEASY®-Blunt E2 (TransGen Biotech, Beijing, China). The plasmid pEASY®- Blunt E2-ompR was transformed into E. coli Rosetta (DE3). The DNA fragments containing the pig promoter (406 bp) and ompR promoter (257 bp) were cloned into the pTOPO-Blunt simple vector (Aidlab, China), respectively, and then promoter probes were obtained through polymerase chain reaction (PCR) with primers M13F and M13R labeled with Cy5.5 at the 5′ end. The Pigpro probe and ompRpro probe were 556 and 407 bp, respectively. Electrophoretic mobility shift assay (EMSA) was carried out with an EMSA/Gel-Shift kit (Beyotime, Shanghai, China). The purified probe and protein were mixed with EMSA/Gel-Shift binding buffer (5×), and a total of 10 μl of the reaction system was supplied with distilled water and incubated for 30 min at 28°C. 6% native PAGE was prepared as the kit protocol described, and the reaction mixture was loaded onto the PAGE. Electrophoresis was performed at 60 V for 3 h in 0.5× TBE buffer, and the gels were exposed to an Odyssey CLx (LI-COR® Biosciences).
DNase I footprinting assay
The pig promoter probe was prepared by polymerase chain reaction (PCR) with primers M13F and M13R (labeled with Hex). The DNase I foot-printing assay was carried out as described by Shi et al. (2017).
Results
OmpR activates the transcription level of the pig gene cluster
We have previously demonstrated that gene deletion of ompR (GenBank: QJU42212.1) or envZ (GenBank: QJU42211.1) would result in loss of prodigiosin producing ability in S. marcescens FZSF02; transcription levels of pigA also down-regulated significantly when ompR or envZ was knocked out assayed by qPCR (Jia et al., 2021). The effect of OmpR and EnvZ on prodigiosin-producing ability was further demonstrated by knockout ompR and envZ, double knockout of envZ and ompR in this study; gene deletion strains all lost prodigiosin-producing ability, prodigiosin-producing ability restored in complementary strain (Figures 1A–C). OmpR was very conserved in amino acid sequences among many Gram-negative bacteria, such as Rouxiella aceris, Ewingella americana, Yersinia enterocolitica, Citrobacter youngae, Hafnia psychrotolerans, Escherichia coli and Klebsiella pneumoniae (Figure 1D). Many studies have demonstrated that OmpR influences a wide variety of cellular processes in E. coli, Salmonella sp. and Shigella sp., as well as pathogenic species of Yersinia sp. (Jaworska et al., 2021), but the influence of secondary metabolite biosynthesis by OmpR has rarely been reported. We have previously demonstrated that OmpR can positively regulate the production of the secondary metabolite prodigiosin in S. marcescens FZSF02, but the regulatory mechanism was unknown.
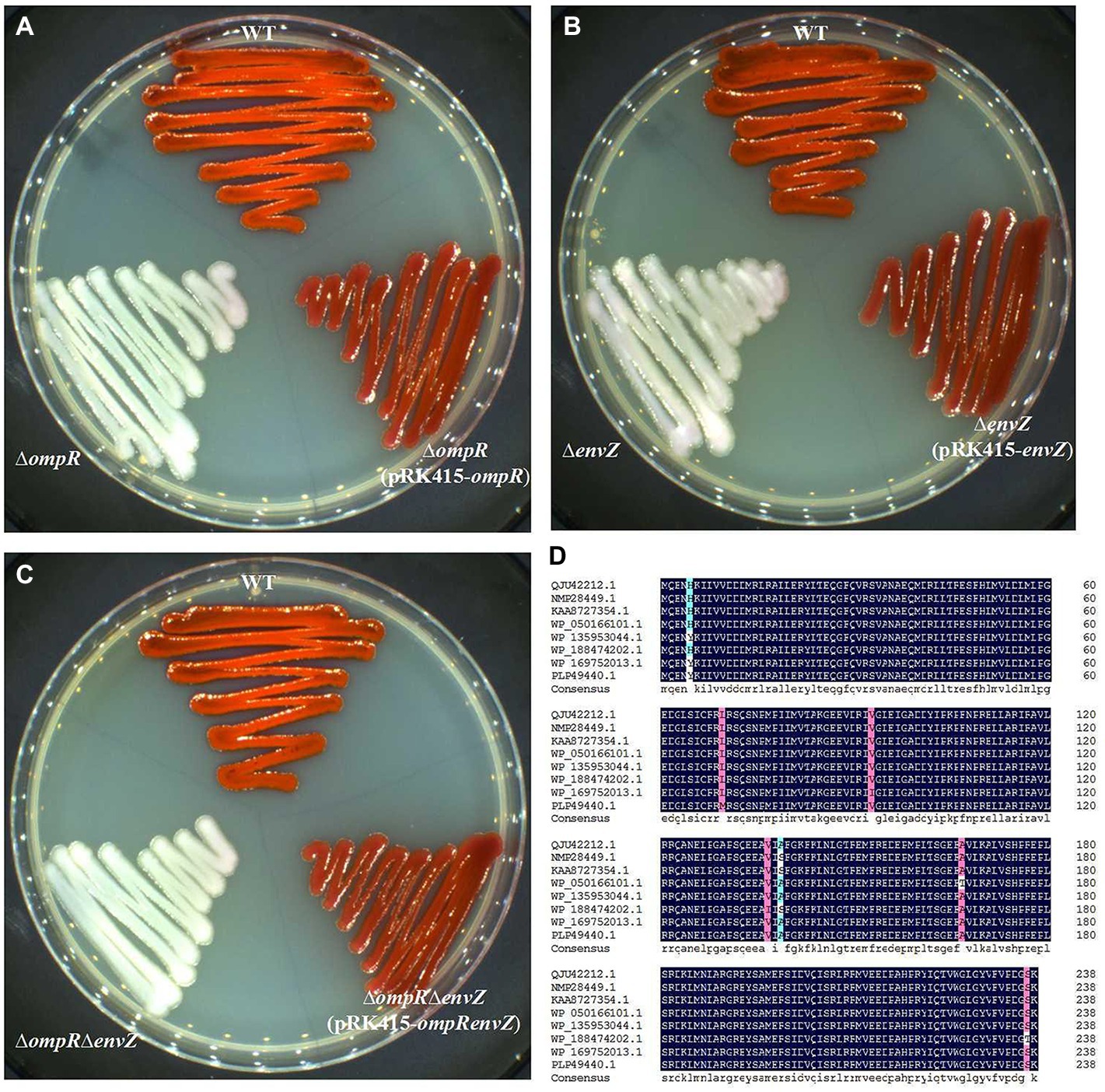
Figure 1. EnvZ/OmpR two-component system and its effect on the prodigiosin-producing ability of S. marcescens FZSF02. (A) Prodigiosin-producing ability of strain WT (wild-type strain of S. marcescens FZSF02), and strain ∆ompR (in frame deletion of ompR of FZSF02), complementary strain ∆ompR (pRK415-ompR). (B) Prodigiosin-producing ability of strain WT (wild-type strain of S. marcescens FZSF02), ∆envZ (in frame deletion of envZ of FZSF02) and complementary strain ∆envZ (pRK415-envZ). (C) Prodigiosin-producing ability of strain WT (wild-type strain of S. marcescens FZSF02), strain ∆ompR∆envZ (in frame deletion of envZ and ompR of FZSF02) and complementary strain ∆ompR∆envZ (pRK415-ompRenvZ). All strains were incubated on LB agar plates at 27°C for 48 h. (D) Multiple sequence alignment of OmpR homologies to analyze its high conservation. Sequences chosen for this analysis were from S. marcescens FZSF02 (QJU42212.1), R. aceris (NMP28449.1), E. americana (KAA8727354.1), Y. enterocolitica (WP_050166101.1), C. youngae (WP_135953044.1), H. psychrotolerans (WP_188474202.1), E. coli (WP_169752013.1), and K. pneumoniae (PLP49440.1).
To further test whether the regulation of prodigiosin biosynthesis by OmpR is at the transcriptional level, the β-galactosidase activity of FZSF02 and FZSF02∆ompR was assayed when the lacZ gene was under control by the pig cluster promoter. The results showed that the β-galactosidase activity of FZSF02∆ompR (pTOPO-Pigpro-lacZ-Cmr) decreased by 88.5% compared with that of the wild-type strain FZSF02 WT (pTOPO-Pigpro-lacZ-Cmr; Figure 2A), and almost no β-galactosidase activity was tested in the control group of WT (pTOPO-Pigpro-Cmr) and ∆ompR (pTOPO-Pigpro-Cmr; Figure 2A). This result indicates that OmpR directly or indirectly activates the transcription level of the pig gene cluster, and influences prodigiosin synthesis in strain FZSF02.

Figure 2. OmpR regulates the prodigiosin-producing ability of FZSF02 directly. (A) Analysis of β-galactosidase activity of ∆ompR (pTOPO-Pigpro-lacZ-Cmr) and WT (pTOPO-Pigpro-lacZ-Cmr) harboring the Pigpro-lacZ reporter fusion. WT (pTOPO-Pigpro-Cmr) and ∆ompR (pTOPO-Pigpro-Cmr) were constructed as controls. (B) EMSA for OmpR protein binding to the promoter of the pig cluster. (C) Analysis of β-galactosidase activity of ∆ompR (pTOPO-ompRpro-lacZ-Cmr) and WT (pTOPO-ompRpro-lacZ-Cmr) harboring the ompRpro-lacZ reporter fusion. WT (pTOPO-ompRpro-Cmr) and ∆ompR (pTOPO-ompRpro-Cmr) were constructed as controls. (D) EMSA for assay OmpR protein binding ability to the promoter of envZ/ompR.
OmpR activates the transcription level of the pig gene cluster by directly binding to its promoter sequence
To study whether OmpR regulates the expression of the pig gene cluster by binding with its promoter directly, EMSA was used to detect the binding ability between OmpR and the pig gene cluster promoter sequence. The results showed that OmpR can bind with the probe prepared with the prodigiosin cluster promoter sequence (Figure 2B). A DNase I footprinting assay showed that the proposed binding sequence of OmpR on the pig promoter was 5′CATTTATTTACATTTAC3′ (Figure 3), which located on −103 bp to −86 bp relative to the A of the ATG start codon of pigA. Pig promoter sequence is between pigA (QJU38817.1) and cueR (QJU38818.1) on the genome of FZSF02.
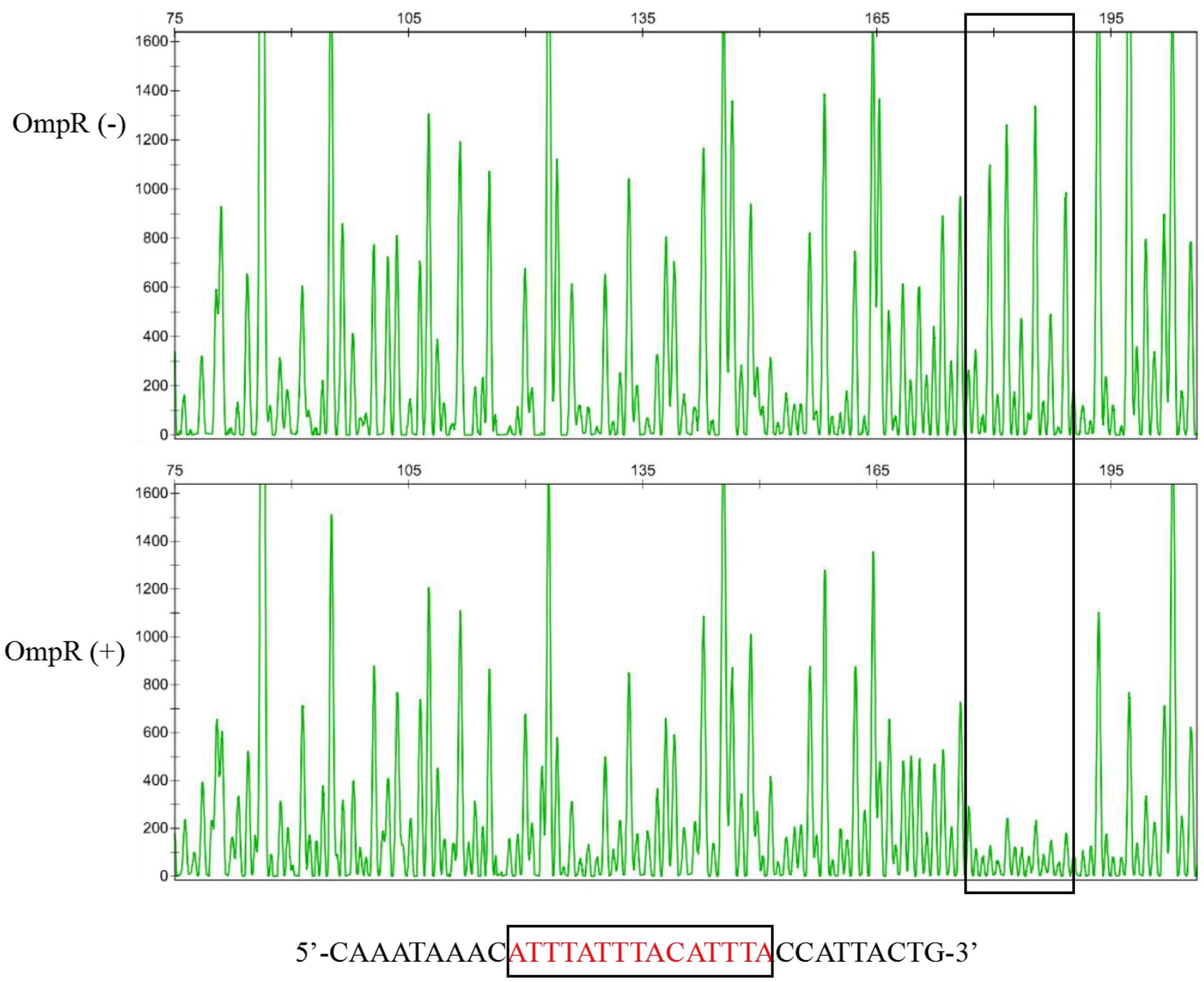
Figure 3. Confirmation of the proposed binding sequence of OmpR with the pig cluster promoter by DNaseI foot-printing assay. The OmpR (−) group indicates that the promoter probe was not incubated with OmpR protein before treatment with DNase I, and the OmpR (+) group indicates that the promoter probe was incubated with OmpR protein before treatment with DNase I.
OMPR activates its own expression level
To test whether the autoregulation of OmpR exists in S. marcescens FZSF02, the β-galactosidase activity of FZSF02 and FZSF02∆ompR was assayed when the lacZ gene was first controlled by the ompR promoter. The results showed that the β-galactosidase activity of FZSF02∆ompR (100 U/ml) decreased by 92.3% compared with that of the wild-type strain FZSF02 (1,300 U/ml; Figure 2C). This finding demonstrates that OmpR can activate its own expression level.
OmpR can directly bind to the envZ/ompR promoter
To test whether the activation of OmpR on its own expression is performed by binding with the envZ/ompR promoter, EMAS was used to examine the binding ability between OmpR and the envZ/ompR promoter sequence. The results showed that when the envZ/ompR promoter sequence was used as a probe, OmpR could bind with the labeled probe (Figure 2D). The addition of unlabeled probe can compete with the labeled probe, which further demonstrates the binding ability.
Discussion
Prodigiosin was a kind of bacterial secondary metabolites produced mainly by many S. marcescens strains. Various regulating genes involved in prodigiosin biosynthesis have been found in the past two decades, but new regulators, such as RcsB (Pan et al., 2021), CpxA/R (Qiu et al., 2021) and Fnr (Sun et al., 2021), have still been reported continuously. Research of the these genes may help to uncover the regulatory mechanism behind prodigiosin biosynthesis in S. marcescens.
EnvZ/OmpR is known to control motility (Prüß, 2017), intracellular survival (Du et al., 2022), antibiotic resistance (Ko and Choi, 2022), virulence (Tipton and Rather, 2017), and other characteristics of different bacterial strains, but few studies have reported the role of EnvZ/OmpR in S. marcescens. We have demonstrated previously that mutation of ompR or envZ would cause the loss of prodigiosin producing ability in S. marcescens FZSF02 and confirmed that the two-component system EnvZ/OmpR was a newly found system that can regulate prodigiosin biosynthesis (Jia et al., 2021). In this study, the regulatory function of EnvZ/OmpR on prodigiosin biosynthesis was further confirmed by gene deletion and complementation (Figure 1C). For the EnvZ/OmpR system, OmpR was reported to play the role by binding to the gene promoters and regulating the expression of other genes (Wang et al., 2021). LacZ reporter assays and EMSA assay in this study also showed OmpR regulate prodigiosin biosynthesis by directly binding to the promoter region of pig cluster (Figures 2A,B). The binding region of the OmpR was identified as 5′CATTTATTTACATTTAC3′ (Figure 3) by a DNase I footprinting assay. The binding sequence showed 45% identity to the E. coli consensus sequence (5′TTTTACTTTTGTAACATAT3′; Maeda et al., 1991) and 55% identity to that of Y. enterocolitica (5′ATTTATTGATGGTAACAATT3′; Nieckarz et al., 2020).
Many two-component systems regulating proteins can autoregulate their own expression by binding to their promoters, and this kind of feedback allows the regulatory functions of the system to be more flexible (Groisman, 2016). For the EnvZ/OmpR two-component system, autoregulation differs among different strains; it exists in Salmonella enterica (Bang et al., 2002; Cameron and Dorman, 2012) but not in E. coli (Ochman and Wilson, 1987; Doolittle et al., 1996) and Acinetobacter baumannii (Tipton and Rather, 2017). The results of the LacZ reporter assay (Figure 2C) and EMSA assay (Figure 2D) indicated that OmpR can bind to the promoter region of envZ/ompR and promote the expression of OmpR and EnvZ.
Based on the above results, we proposed that the regulatory mechanism of the two-component EnvZ/OmpR system on prodigisin biosynthesis was probably as follows (Figure 4): Some unknown factors induced the expression of EnvZ and OmpR, and OmpR was then phosphorylated by EnvZ (Cai and Inouye, 2002). Phosphorylated OmpR activated the expression of more EnvZ and OmpR by binding with the envZ/ompR promoter. When the concentration of OmpR reached a certain level, the pig gene cluster promoter persistently bound with OmpR, and genes involved in prodigiosin biosynthesis were highly expressed at the transcriptional level.
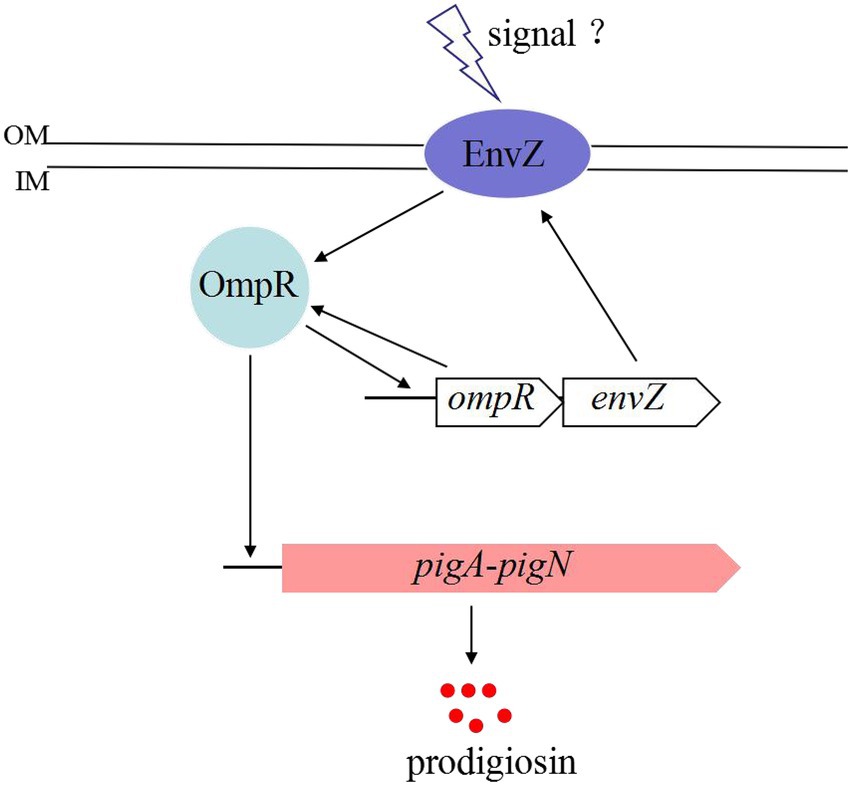
Figure 4. Proposed model of OmpR involvement in the regulatory mechanism of prodigiosin biosynthesis in S. marcescens FZSF02.
Although we have previously found that when ompR was knocked out, S. marcescens FZSF02 lost the prodigiosin biosynthesis ability (Jia et al., 2021), in this study, the proposed regulatory mechanism of OmpR on prodigiosin biosynthesis was demonstrated. EnvZ/OmpR was demonstrated to be a new two-compound system that can directly positively regulate prodigiosin production in S. marcescens FZSF02.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
XJ and JC designed the study, wrote the manuscript, and analyzed the results. XJ, FL, JL, CL, and KZ performed the experiments. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the Exploration Program of Fujian Academy of Agricultural Sciences (ZYTS202217), Scientific Research in the Public Interest of Fujian Province (2020R1025003 and 2021R1025002), Natural Science Foundation of Fujian Province of China (2021J01480), Chinese National Natural Science Foundation (31800068), the Special Program for Extension Research of National Natural Science Foundation of Fujian Academy of Agricultural Sciences (AGY2018-1), and Science and Technology Innovation Team Program of Fujian Academy of Agricultural Sciences (CXTD2021002-3).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1041146/full#supplementary-material
References
Bang, I. S., Audia, J. P., Park, Y. K., and Foster, J. W. (2002). Autoinduction of the ompR response regulator by acid shock and control of the Salmonella enterica acid tolerance response. Mol. Microbiol. 44, 1235–1250. doi: 10.1046/j.1365-2958.2002.02937.x
Beier, D., and Gross, R. (2006). Regulation of bacterial virulence by two-component systems. Curr. Opin. Microbiol. 9, 143–152. doi: 10.1016/j.mib.2006.01.005
Bhagirath, A. Y., Li, Y., Patidar, R., Yerex, K., Ma, X., Kumar, A., et al. (2019). Two component regulatory systems and antibiotic resistance in gram-negative pathogens. Int. J. Mol. Sci. 20:1781. doi: 10.3390/ijms20071781
Boyce, K. J., Cao, C., and Andrianopoulos, A. (2016). Two-component signaling regulates osmotic stress adaptation via SskA and the high-osmolarity glycerol MAPK pathway in the human pathogen Talaromyces marneffei. mSphere 1, e00086–e00015. doi: 10.1128/mSphere.00086-15
Cai, S. J., and Inouye, M. (2002). EnvZ-OmpR interaction and osmoregulation in Escherichia coli. J. Biol. Chem. 277, 24155–24161. doi: 10.1074/jbc.M110715200
Cameron, A. D., and Dorman, C. J. (2012). A fundamental regulatory mechanism operating through OmpR and DNA topology controls expression of salmonella pathogenicity islands SPI-1 and SPI-2. PLoS Genet. 8:e1002615. doi: 10.1371/journal.pgen.1002615
Darshan, N., and Manonmani, H. K. (2015). Prodigiosin and its potential applications. J. Food Sci. Technol. 52, 5393–5407. doi: 10.1007/s13197-015-1740-4
Dhiman, A., Gopalani, M., and Bhatnagar, R. (2015). WalRK two component system of Bacillus anthracis responds to temperature and antibiotic stress. Biochem. Biophys. Res. Commun. 459, 623–628. doi: 10.1016/j.bbrc.2015.02.159
Dixon, R. (1998). The oxygen-responsive NIFL-NIFA complex: a novel two-component regulatory system controlling nitrogenase synthesis in gamma-proteobacteria. Arch. Microbiol. 169, 371–380. doi: 10.1007/s002030050585
Doolittle, R. F., Feng, D. F., Tsang, S., Cho, G., and Little, E. (1996). Determining divergence times of the major kingdoms of living organisms with a protein clock. Science 271, 470–477. doi: 10.1126/science.271.5248.470
Du, Z., Zhang, M., Qin, Y., Zhao, L., Huang, L., Xu, X., et al. (2022). The role and mechanisms of the two-component system EnvZ/OmpR on the intracellular survival of Aeromonas hydrophila. J. Fish Dis. 45, 1609–1621. doi: 10.1111/jfd.13684
Fineran, P. C., Slater, H., Everson, L., Hughes, K., and Salmond, G. P. (2005). Biosynthesis of tripyrrole and beta-lactam secondary metabolites in Serratia: integration of quorum sensing with multiple new regulatory components in the control of prodigiosin and carbapenem antibiotic production. Mol. Microbiol. 56, 1495–1517. doi: 10.1111/j.1365-2958.2005.04660.x
Gristwood, T., Fineran, P. C., Everson, L., Williamson, N. R., and Salmond, G. P. (2009). The PhoBR two-component system regulates antibiotic biosynthesis in Serratia in response to phosphate. BMC Microbiol. 9:112. doi: 10.1186/1471-2180-9-112
Groisman, E. A. (2016). Feedback control of two-component regulatory systems. Annu. Rev. Microbiol. 70, 103–124. doi: 10.1146/annurev-micro102215-095331
Horng, Y. T., Chang, K. C., Liu, Y. N., Lai, H. C., and Soo, P. C. (2010). The RssB/RssA two-component system regulates biosynthesis of the tripyrrole antibiotic, prodigiosin, in Serratia marcescens. Int. J. Med. Microbiol. 300, 304–312. doi: 10.1016/j.ijmm.2010.01.003
Jaworska, K., Ludwiczak, M., Murawska, E., Raczkowska, A., and Brzostek, K. (2021). The regulator OmpR in Yersinia enterocolitica participates in iron homeostasis by modulating fur level and affecting the expression of genes involved in iron uptake. Int. J. Mol. Sci. 22:1475. doi: 10.3390/ijms22031475
Jia, X., Liu, F., Zhao, K., Lin, J., Fang, Y., Cai, S., et al. (2021). Identification of essential genes associated with prodigiosin production in Serratia marcescens FZSF02. Front. Microbiol. 12:705853. doi: 10.3389/fmicb.2021.705853
Ko, D., and Choi, S. H. (2022). Mechanistic understanding of antibiotic resistance mediated by EnvZ/OmpR two-component system in salmonella enterica serovar Enteritidis. J. Antimicrob. Chemother. 77, 2419–2428. doi: 10.1093/jac/dkac223
Kruppa, M., Krom, B. P., Chauhan, N., Bambach, A. V., Cihlar, R. L., and Calderone, R. A. (2004). The two-component signal transduction protein Chk1p regulates quorum sensing in Candida albicans. Eukaryot. Cell 3, 1062–1065. doi: 10.1128/EC.3.4.1062-1065.2004
Lai, H. C., Soo, P. C., Wei, J. R., Yi, W. C., Liaw, S. J., Horng, Y. T., et al. (2005). The RssAB two-component signal transduction system in Serratia marcescens regulates swarming motility and cell envelope architecture in response to exogenous saturated fatty acids. J. Bacteriol. 187, 3407–3414. doi: 10.1128/JB.187.10.3407-3414.2005
Maeda, S., Takayanagi, K., Nishimura, Y., Maruyama, T., Sato, K., and Mizuno, T. (1991). Activation of the osmoregulated ompC gene by the OmpR protein in Escherichia coli: a study involving synthetic OmpR-binding sequences. J. Biochem. 110, 324–327. doi: 10.1093/oxfordjournals.jbchem.a123579
Najnin, T., Siddiqui, K. S., Taha, T., Elkaid, N., Kornfeld, G., Curmi, P. M., et al. (2016). Characterization of a temperature-responsive two component regulatory system from the Antarctic archaeon, Methanococcoides burtonii. Sci. Rep. 6:24278. doi: 10.1038/srep24278
Nieckarz, M., Kaczor, P., Jaworska, K., Raczkowska, A., and Brzostek, K. (2020). Urease expression in pathogenic Yersinia enterocolitica strains of bio-serotypes 2/O:9 and 1B/O:8 is differentially regulated by the OmpR regulator. Front. Microbiol. 11:607. doi: 10.3389/fmicb.2020.00607
Ochman, H., and Wilson, A. C. (1987). Evolution in bacteria: evidence for a universal substitution rate in cellular genomes. J. Mol. Evol. 26, 74–86. doi: 10.1007/BF02111283
Pan, X., Tang, M., You, J., Liu, F., Sun, C., Osire, T., et al. (2021). Regulator RcsB controls prodigiosin synthesis and various cellular processes in Serratia marcescens JNB5-1. Appl. Environ. Microbiol. 87, e02052–e02020. doi: 10.1128/AEM.02052-20
Pan, X., You, J., Tang, M., Zhang, X., Xu, M., Yang, T., et al. (2022). Improving prodigiosin production by transcription factor engineering and promoter engineering in Serratia marcescens. Front. Microbiol. 13:977337. doi: 10.3389/fmicb.2022.977337
Papon, N., and Stock, A. M. (2019). Two-component systems. Curr. Biol. 29, R724–R725. doi: 10.1016/j.cub.2019.06.010
Pardee, A. B., Jacob, F., and Monod, J. (1959). The genetic control and cytoplasmic expression of “Inducibility” in the synthesis of β-galactosidase by E. coli. J. Mol. Biol. 1, 165–178. doi: 10.1016/S0022-2836(59)80045-0
Paul, T., Bandyopadhyay, T. K., Mondal, A., Tiwari, O. N., and Bhunia, B. (2020). A comprehensive review on recent trends in production, purification, and applications of prodigiosin. Biomass Conv. Bioref. 12, 1409–1431. doi: 10.1007/s13399-020-00928-2
Prüß, B. M. (2017). Involvement of two-component signaling on bacterial motility and biofilm development. J. Bacteriol. 199, e00259–e00217. doi: 10.1128/JB.00259-17
Qin, L., Yoshida, T., and Inouye, M. (2001). The critical role of DNA in the equilibrium between OmpR and phosphorylated OmpR mediated by EnvZ in Escherichia coli. Proc. Natl. Acad. Sci. U. S. A. 98, 908–913. doi: 10.1073/pnas.98.3.908
Qiu, S., Jia, S., Zhang, F., Liu, X., Ran, T., Wang, W., et al. (2021). Two component system CpxR/a regulates the prodigiosin biosynthesis by negative control in Serratia marcescens FS14. Biochem. Biophys. Res. Commun. 579, 136–140. doi: 10.1016/j.bbrc.2021.09.050
Sarwar, Z., and Garza, A. G. (2015). Two-component signal transduction systems that regulate the temporal and spatial expression of Myxococcus xanthus sporulation genes. J. Bacteriol. 198, 377–385. doi: 10.1128/JB.00474-15
Scharf, B. E. (2010). Summary of useful methods for two-component system research. Curr. Opin. Microbiol. 13, 246–252. doi: 10.1016/j.mib.2010.01.006
Shi, K., Fan, X., Qiao, Z., Han, Y., McDermott, T. R., Wang, Q., et al. (2017). Arsenite oxidation regulator AioR regulates bacterial chemotaxis toward arsenite in Agrobacterium tumefaciens GW4. Sci. Rep. 7:43252. doi: 10.1038/srep43252
Sola-Landa, A., Moura, R. S., and Martín, J. F. (2003). The two-component PhoR-PhoP system controls both primary metabolism and secondary metabolite biosynthesis in Streptomyces lividans. Proc. Natl. Acad. Sci. U. S. A. 100, 6133–6138. doi: 10.1073/pnas.0931429100
Stankovic, N., Senerovic, L., Ilic-Tomic, T., Vasiljevic, B., and Nikodinovic-Runic, J. (2014). Properties and applications of undecylprodigiosin and other bacterial prodigiosins. Appl. Microbiol. Biotechnol. 98, 3841–3858. doi: 10.1007/s00253-014-5590-1
Stella, N. A., Lahr, R. M., Brothers, K. M., Kalivoda, E. J., Hunt, K. M., Kwak, D. H., et al. (2015). Serratia marcescens cyclic AMP receptor protein controls transcription of EepR, a novel regulator of antimicrobial secondary metabolites. J. Bacteriol. 197, 2468–2478. doi: 10.1128/JB.00136-15
Stock, A. M., Robinson, V. L., and Goudreau, P. N. (1999). Two-component signal transduction. Annu. Rev. Biochem. 69, 183–215. doi: 10.1146/annurev.biochem.69.1.183
Sun, D., Zhou, X., Liu, C., Zhu, J., Ru, Y., Liu, W., et al. (2021). Fnr negatively regulates prodigiosin synthesis in Serratia sp. ATCC 39006 during aerobic fermentation. Front. Microbiol. 12:734854. doi: 10.3389/fmicb.2021.734854
Tierney, A. R., and Rather, P. N. (2019). Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 14, 533–552. doi: 10.2217/fmb-2019-0002
Tipton, K. A., and Rather, P. N. (2017). An ompR-envZ two-component system ortholog regulates phase variation, osmotic tolerance, motility, and virulence in Acinetobacter baumannii strain AB5075. J. Bacteriol. 199, e00705–e00716. doi: 10.1128/JB.00705-16
Wang, S. T., Kuo, C. J., Huang, C. W., Lee, T. M., Chen, J. W., and Chen, C. S. (2021). OmpR coordinates the expression of virulence factors of Enterohemorrhagic Escherichia coli in the alimentary tract of Caenorhabditis elegans. Mol. Microbiol. 116, 168–183. doi: 10.1111/mmi.14698
Williamson, N. R., Fineran, P. C., Leeper, F. J., and Salmond, G. P. (2006). The biosynthesis and regulation of bacterial prodiginines. Nat. Rev. Microbiol. 4, 887–899. doi: 10.1038/nrmicro1531
Wright, G., Saeki, A., Hikima, T., Nishizono, Y., Hisano, T., Kamaya, M., et al. (2018). Architecture of the complete oxygen-sensing FixL-FixJ two-component signal transduction system. Sci. Signal. 11:eaaq0825. doi: 10.1126/scisignal.aaq0825
Keywords: Serratia marcescens, prodigiosin, OmpR, two-component system, regulatory mechanism
Citation: Jia X, Zhao K, Liu F, Lin J, Lin C and Chen J (2022) Transcriptional factor OmpR positively regulates prodigiosin biosynthesis in Serratia marcescens FZSF02 by binding with the promoter of the prodigiosin cluster. Front. Microbiol. 13:1041146. doi: 10.3389/fmicb.2022.1041146
Edited by:
Harold J. Schreier, University of Maryland, Baltimore County, United StatesCopyright © 2022 Jia, Zhao, Liu, Lin, Lin and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jichen Chen, Y2hlbmppY2hlbjIwMDFAMTYzLmNvbQ==
†These authors have contributed equally to this work