
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 07 November 2022
Sec. Evolutionary and Genomic Microbiology
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.1033675
This article is part of the Research Topic Metagenomic Surveillance and Epidemiology of Clinically Opportunistic Pathogens View all 6 articles
 Romário Alves Rodrigues1*†
Romário Alves Rodrigues1*† Lucas José Luduverio Pizauro2,3†
Lucas José Luduverio Pizauro2,3† Alessandro de Mello Varani3
Alessandro de Mello Varani3 Camila Chioda de Almeida1
Camila Chioda de Almeida1 Saura Rodrigues Silva3Marita Vedovelli Cardozo4Janet I. MacInnes5
Saura Rodrigues Silva3Marita Vedovelli Cardozo4Janet I. MacInnes5 Andrew M. Kropinski5,6Poliana de Castro Melo2Fernando Antonio Ávila1
Andrew M. Kropinski5,6Poliana de Castro Melo2Fernando Antonio Ávila1Staphylococcus aureus causes nosocomial and intramammary infections in humans and cattle, respectively. A large number of virulence factors are thought to play important roles in the pathogenesis of this bacterium. Currently, genome-wide and data-analysis studies are being used to better understand its epidemiology. In this study, we conducted a genome wide comparison and phylogenomic analyses of S. aureus to find specific virulence patterns associated with clinical and subclinical mastitis strains in cattle and compare them with those of human origin. The presence/absence of key virulence factors such as adhesin, biofilm, antimicrobial resistance, and toxin genes, as well as the phylogeny and sequence type of the isolates were evaluated. A total of 248 genomes (27 clinical mastitis, 43 subclinical mastitis, 21 milk, 53 skin-related abscesses, 49 skin infections, and 55 pus from cellulitis) isolated from 32 countries were evaluated. We found that the cflA, fnbA, ebpS, spa, sdrC, coa, emp, vWF, atl, sasH, sasA, and sasF adhesion genes, as well as the aur, hglA, hglB, and hglC toxin genes were highly associated in clinical mastitis strains. The strains had diverse genetic origins (72 protein A and 48 sequence types with ST97, ST8 and ST152 being frequent in isolates from clinical mastitis, abscess, and skin infection, respectively). Further, our phylogenomic analyses suggested that zoonotic and/or zooanthroponotic transmission may have occurred. These findings contribute to a better understanding of S. aureus epidemiology and the relationships between adhesion mechanisms, biofilm formation, antimicrobial resistance, and toxins and could aid in the development of improved vaccines and strain genotyping methods.
Staphylococcus aureus is a commensal microorganism that can cause several important diseases in humans and animals. It is considered a facultative intracellular pathogen that is responsible for recurrent infections (Watkins and Unnikrishnan, 2020). In humans, its primary entry points are damaged skin and mucosa, which can lead to skin infections, septicemia, endocarditis, and abscesses (Lalaouna et al., 2018). In cattle, intramammary S. aureus infections (clinical and subclinical mastitis) spread primarily via milking machines, milkers’ hands, and vectors such as flies (Zadoks et al., 2011).
Virulence factors are important in the development, duration, and severity of S. aureus related illnesses since they are often involved in host defense evasion (Wójcik-Bojek et al., 2022). Adhesins play a role in cell attachment, invasion, and biofilm formation (Kerro Dego, 2020), thus allowing for increased persistence and protection against antimicrobial factors in the host (Gajewska and Chajęcka-Wierzchowska, 2020). The production of toxins such as hemolysins, leukotoxins, proteases, and other enzymes can cause direct host cell injury (Otto, 2014). S. aureus can also carry a large number of resistance genes which pose a serious economic threat to the dairy sector and to public health (Igbinosa et al., 2016).
The contamination of food products by S. aureus can be aided by virulence factors that mediate the interaction of pathogen with the host and the environment (Gajewska and Chajęcka-Wierzchowska, 2020; Bencardino et al., 2021). When a dairy herd has clinical mastitis (animals with obvious disease) or subclinical mastitis (animals with no obvious signs) milk contamination and discard rates increase, productivity decreases, antimicrobial treatment costs increase, and animals may have to be culled prematurely (Exel et al., 2022). Zoonotic transmission of S. aureus has been widely documented (Patel et al., 2021) and characterization of sequence types (STs) helps to elucidate strain relationships and epidemiology, as well as gene transfer amongst strains from different hosts (Bruce et al., 2022).
Despite the importance of S. aureus in human and veterinary medicine there is a dearth of studies on gene profile, strain-to-strain relationships, disease-host relationships, and host-pathogen interactions (Pizauro et al., 2021). In the current study we used genome wide analyses for genome comparison to better understand strain-to-strain relationships, disease-host relationships, and host-pathogen interactions. Comparative genomics and phylogenomic analyses were used to characterize the adhesion, biofilm, toxin, and antimicrobial resistance genes of S. aureus isolated from human and cattle samples.
The National Center for Biotechnology Information database (NCBI; Clark et al., 2016) was searched for S. aureus genomes of bovine (Bos taurus) and human (Homo sapiens) origin. For this search, we used “organism” from the “Available Facets” section of the NCBI’s web platform Sequence Set Browser, and then the microorganism “Staphylococcus aureus” from the “Top Organisms” section. The S. aureus genomes from Bos taurus were analyzed, in addition to raw milk/bulk milk, and the condition (clinical mastitis, subclinical mastitis) was described as a stage of the disease in the BioSample description or in scholarly publications. S. aureus genomes of isolates from Homo sapiens that had abscess, skin infection, or pus listed as the source of isolation in the BioSample description or academic publications were then obtained; in addition, other sources of isolation such as surgical site, skin, and wound secretion were accepted for the group “skin infections.” Genome sequences deposited from June 2020 until January 2021 were analyzed.
S. aureus genomes were annotated using Rapid Annotation using Subsystem Technology (RAST; Aziz et al., 2008). The identification of genes related to adhesion and biofilm formation were classified based on the results of the RAST platform and the Virulence Factor of Pathogenic Bacteria (VFDB) reference database (Chen et al., 2005). The antimicrobial resistance genes were annotated using ResFinder 4.1 and toxin genes with VirulenceFinder 2.0, both from the Center of Genomic Epidemiology (Bortolaia et al., 2020).
To normalize the analyses, all genomes were annotated using Prokka 1.14.6 (Seemann, 2014). The computational tool Roary 3.13.0 was used to infer the pangenome (Page et al., 2015). The Roary tool was also used to calculate pangenome size and reveal orthologous genes.
The phylogenomic concatenated matrix was created with PRANK (Löytynoja, 2014) using codon-aware alignment composed of 247 genes shared across all analyzed Staphylococcus spp. genomes. The phylogenetic tree was constructed using the maximum likelihood (ML). The ML tree was calculated using IQ-Tree 2 (Minh et al., 2020) with the best-of-fit model GTR + F + R6 according to AIC criteria (Akaike, 1974), with the software tool ModelFinder (Kalyaanamoorthy et al., 2017). Clade support was estimated using the ultrafast bootstrap (UFBoot) and SH-aLRT algorithms (Hoang et al., 2018) with 1.000 replicates. Tree rooting was based on the Escherichia coli genomes (GenBank access numbers: CP058682, NZ CP037943, NZ CP027390, NZ AP018808, and NZ AP018808) and tree drawing was done using Interactive tree of life (iTOL) v6 (Letunic and Bork, 2016).
Multilocus sequence types were determined in silico using the MLST 2.0 online platform (Larsen et al., 2012), and spa typing was done using spaTyper version 1.0 (Bartels et al., 2014), both from the Center of Genomic Epidemiology.
Statistical analyzes were performed using the R program 4.1.2 (R Core Team, 2017). The Fisher’s exact test was used to analyze possible associations between the presence of each of the virulence genes (gene by gene) in each type of disease evaluated, using a contingency table considering “1” as presence and “0” as absence and a significant value of p < 0.05. Multinomial logistic regression was done with all genes that showed a significant value in Fisher’s test to assess the significance of association between the types of condition, considering raw milk samples as the reference, and the origin of the isolates being the response variable, and the genes, the explanatory variable. Using a modification of the method of Åvall-Jääskeläinen et al. (2021) the multinomial logistic regression model was then analyzed by Wald’s Z test and the value of p values were obtained by the pnorm() function of the nnet package. Genes with p < 0.05 were considered significant for the determination of the model in each disease (Dalgaard, 2008).
Multinational logistic regression model used was:
β0c represents the medium value for the category c of the response variableβ1c Screened virulence genes, represent the fixed effects of the Virulence genes category with c as r = 1,---,36;β2c MLST, represent the fixed effects of the variable MLST category with c as j = 1---,26.c = 1,--5, represent the 5 logit functions that will be used when the Raw milk is used as a baseline for comparison of each source.
Evaluation between the association of the presence of adhesion and biofilm genes and the origin of isolation was done using the Spearman correlation test (Dalgaard, 2008). The correlation coefficients and the gene frequency by origin were presented in heat maps.
The treeWAS R package https://github.com/caitiecollins/treeWAS (Collins and Didelot, 2018) was used to apply a phylogenetic tree-based approach to the genes and genome studied. For these analyses, a genetic dataset with a matrix containing binary genetic data (gene presence/absence), a phenotypic variable (isolates isolation source as a continuous numeric value) and the phylogenetic tree consisted of the 247 genes that were shared among all isolates was used. These findings were further assessed with the multinomial logistic regression.
The genomes of 248 independent strains of S. aureus from six different sources: diseased cattle (27 clinical mastitis and 43 subclinical mastitis cases); diseased human (53 skin-related abscesses, 49 skin infections, and 55 pus from cellulitis cases) and healthy controls (21 raw milk/bulk milk) were evaluated. The isolates came from 32 countries, i.e., Argentina (1), Australia (5), Bahamas (1), Belgium (4), Brazil (8), Canada (1), Chile (7), China (9), Colombia (2), Cuba (4), Denmark (1), Ecuador (3), Finland (4), Ghana (13), India (5), Ireland (2), Italy (4), Japan (1), Lebanon (7), Malaysia (1), Netherlands (7), Norway (1), Russia (32), South Sudan (1), Spain (1), Suriname (1), Switzerland (24), Taiwan (52), Thailand (1), Turkey (7), United Kingdom (20) and United States (18) (Figure 1).
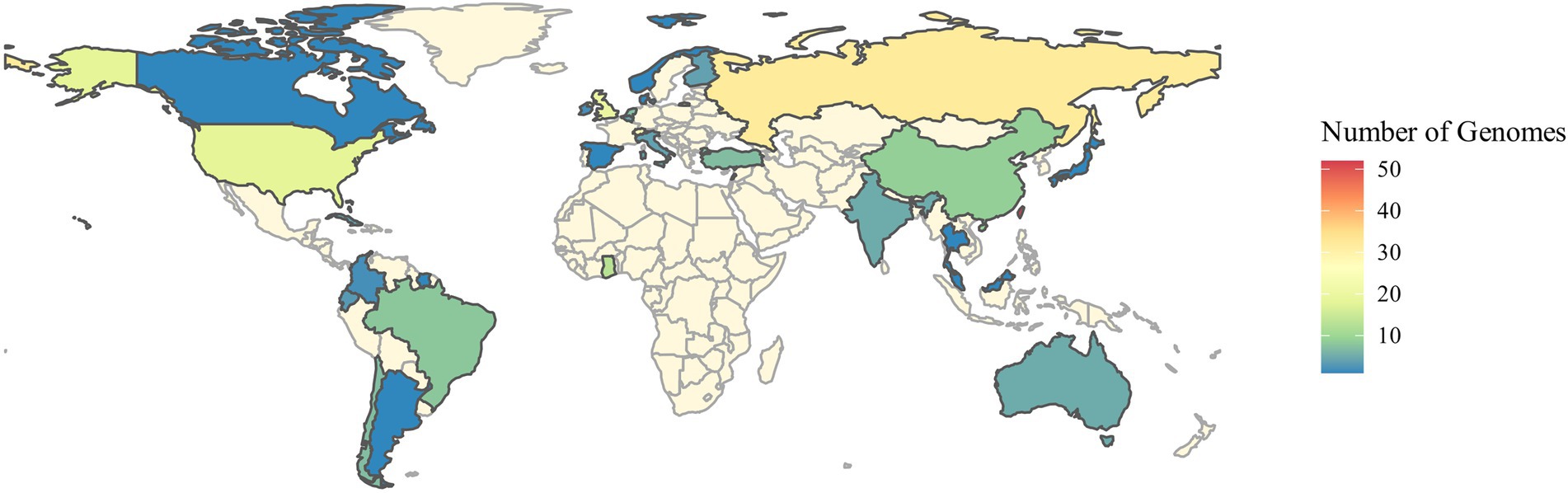
Figure 1. Spatial distribution of the isolates according to the information presented in the NCBI GenBank.
Based on assessment of these 248 S. aureus genomes, the pangenome analysis revealed 15,011 accessory genes and 873 genes belonging to the central genome. Five hundred and ninety-four genes were classified as soft-core genes (i.e., shared between 95 and 98% of the identified genomes); 1,842 as shell genes (shared between 15 and 95% of the genomes) and 11,702 as cloud genes (shared by less than 15% of the analyzed isolates). Central genome genes were associated with translation, ribosomal structure and biogenesis (9.86%), amino acid transport and metabolism (9.05%), transcription (8.25%), energy production and conversion (8.25%); many others were of unknown function (27.16%; Figure 2). The increase in the number of single genes was inversely related to the number of new genes revealed for this species (Figure 3).
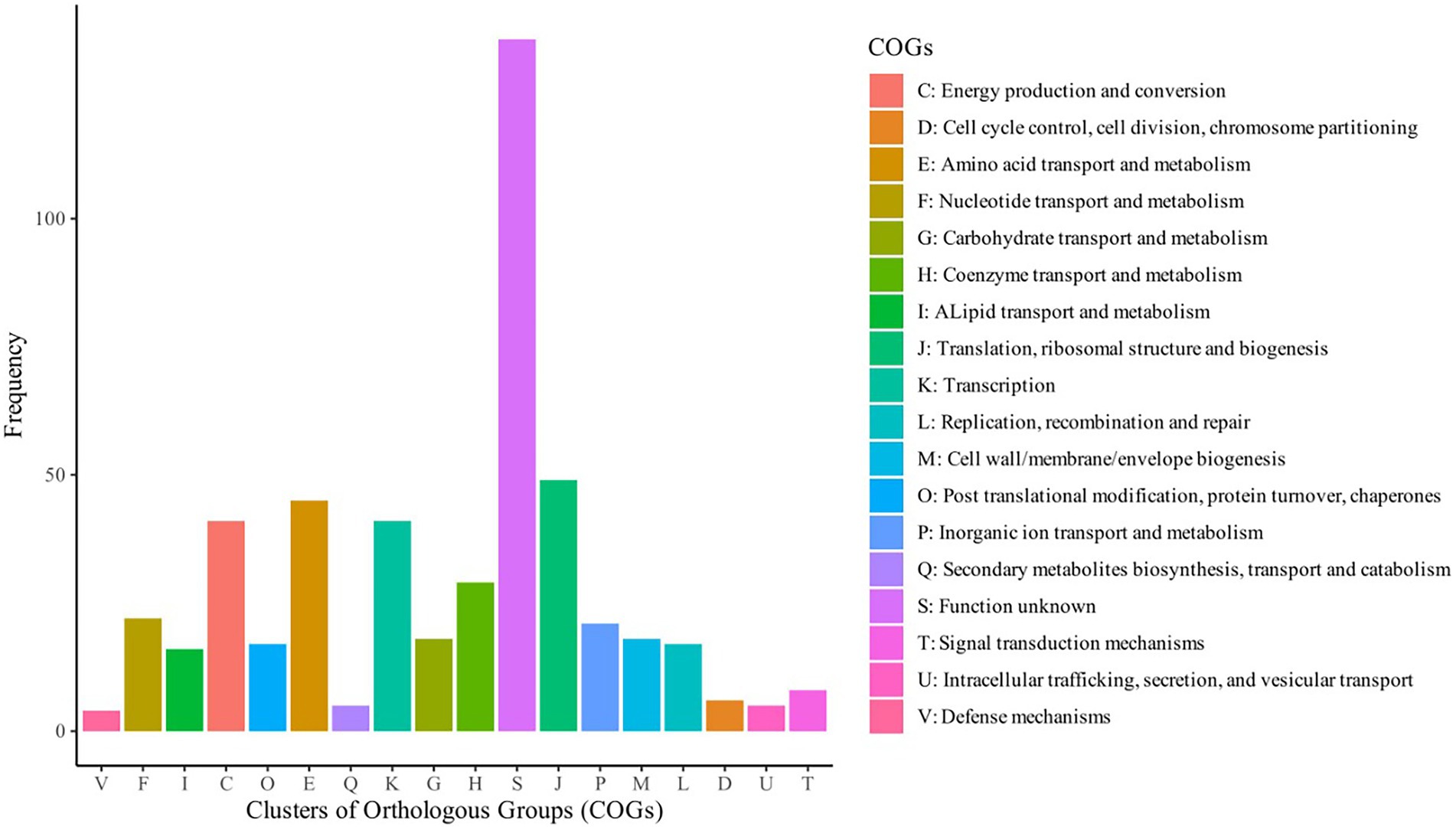
Figure 2. Category of Clusters of Orthologous Groups (COG) of genes central to the genomes of 248 Staphylococcus aureus isolated from humans (abscesses, skin infections and pus) and from cattle (clinical mastitis, subclinical mastitis and raw milk).
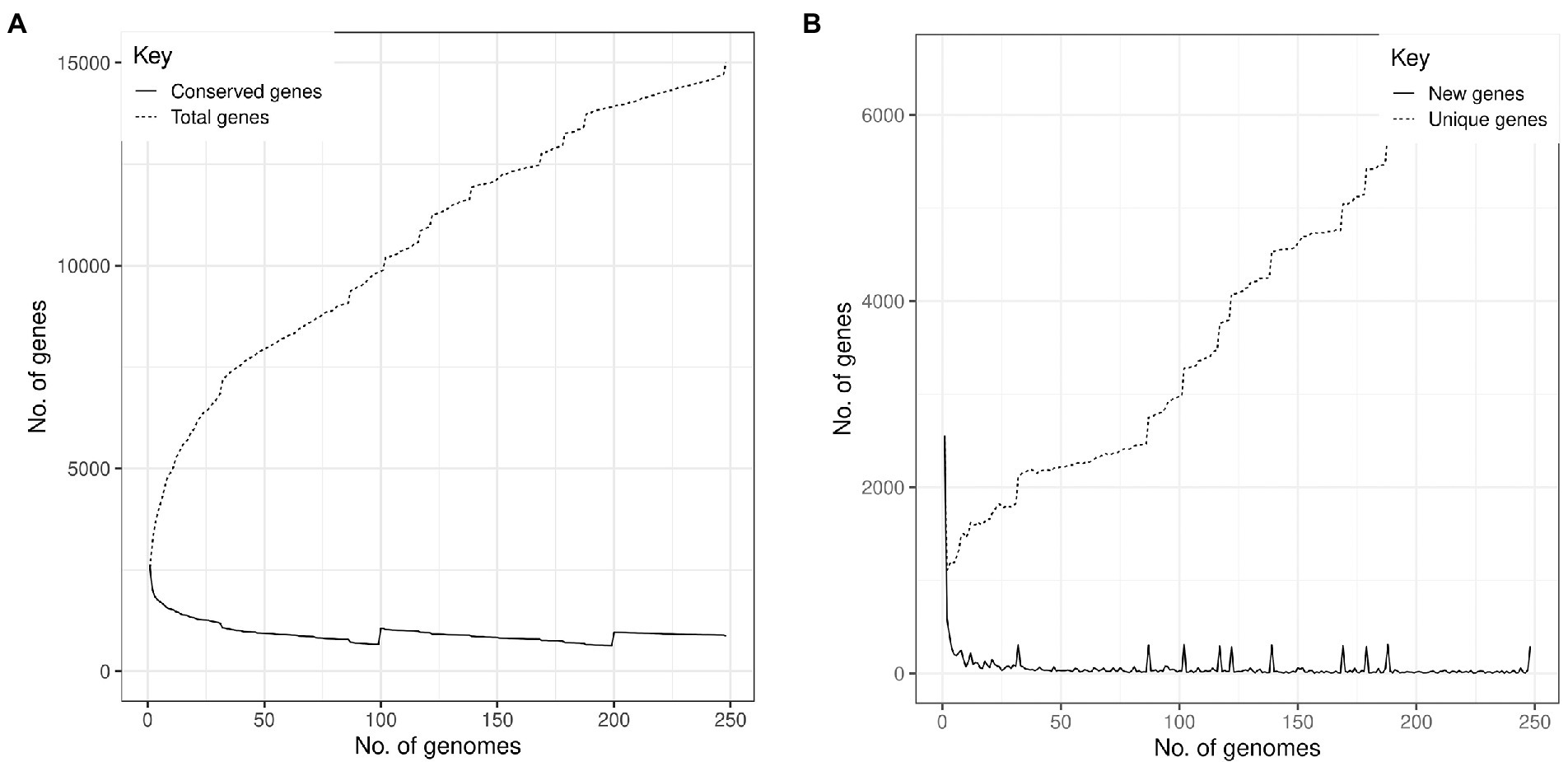
Figure 3. Staphylococcus aureus pangenome profile isolated from humans (abscesses, skin infections, and pus) and from cattle (clinical mastitis, subclinical mastitis, and raw milk). (A) The number of genes as a function of the number of genomes. The solid line indicates the number of conserved genes in the core genome and the dashed line the total number of genes. (B) The number of new pangenome genes as a function of the number of analyzed genomes. The solid line indicates the number of new genes and the dashed line, the total number of unique genes.
Of the 248 S. aureus genomes studied, 70 were bovine disease isolates, 157 were from humans with skin associated disease, and 21 were from raw milk. Annotation using RAST identified 26 adhesin genes (clfA, clfB, cna, fnbA, fnbB, ebpS, spa, sdrC, sdrD, sdrE, coa, eap, emp, efb, vWbp, atl, aap, pls, sasG, sasH, sasA, sasC, sasD, sasF, sasI, sasK) and nine biofilm formation genes (icaA, icaB, icaC, icaD, icaR, rbf, tcaR, sarA and sigB). Thirty antimicrobial resistance genes (adD, aac(6′)-aph(2″), aadE, ant(9)-Ia, aph(3′)-III, blaTEM-116, blaZ, cat (pC233), dfrG, erm(A), erm(B), erm(C), fosB4, fosB6, fusB, fusC, lnu (A), lnu (B), Isa (E), mecA, mecA1, mph(C), msr(A), mup(A), sal (A), str, tet(K), tet(M), vga(A)V and qacD) were identified using ResFinder and 43 toxin genes (i.e., aur, splA, splB, splE, ACME, sak, scn, edinB, hlgA, hlgB, hlgC, lukD, lukE, lukF-PV, lukS-PV, sea, seb, sec, sec3, sed, seg, seh, sei, sej, sel, sek, sem, sen, seo, sep, seq, seu, ser and tst) were identified using VirulenceFinder (Supplementary Material 1).
Adhesin genes clfA, clfB, fnbA, ebpS, spa, coa, eap, emp, vWbp, atl, sasH, sasA, sasC, and sasF genes were observed at high frequencies (> 90%) in isolates from all sources while the cna, aap and pls genes were observed in fewer strains (~20%). Differences in the frequencies of some genes were seen (e.g., clinical mastitis genomes had a lower frequency, approximately 83%) of the icaA, icaB, icaC and icaD biofilm genes and biofilm regulator genes rbf, tcaR, sarA and sigB while in the other groups, these genes were present in >90% of the strains evaluated. There was a high prevalence (>90%) of the fnbB gene in skin and pus infection isolates and of the sdrD and sasG genes in abscesses and pus strains; both of which also presented similar frequencies in the bovine isolates. The sdrE and efb genes were less prevalent (< 90%) in isolates from clinical mastitis and skin infection cases; the sdrC gene in skin infection isolates, and sasD in clinical mastitis, raw milk, and pus isolates (Figure 4; Supplementary Material 2).
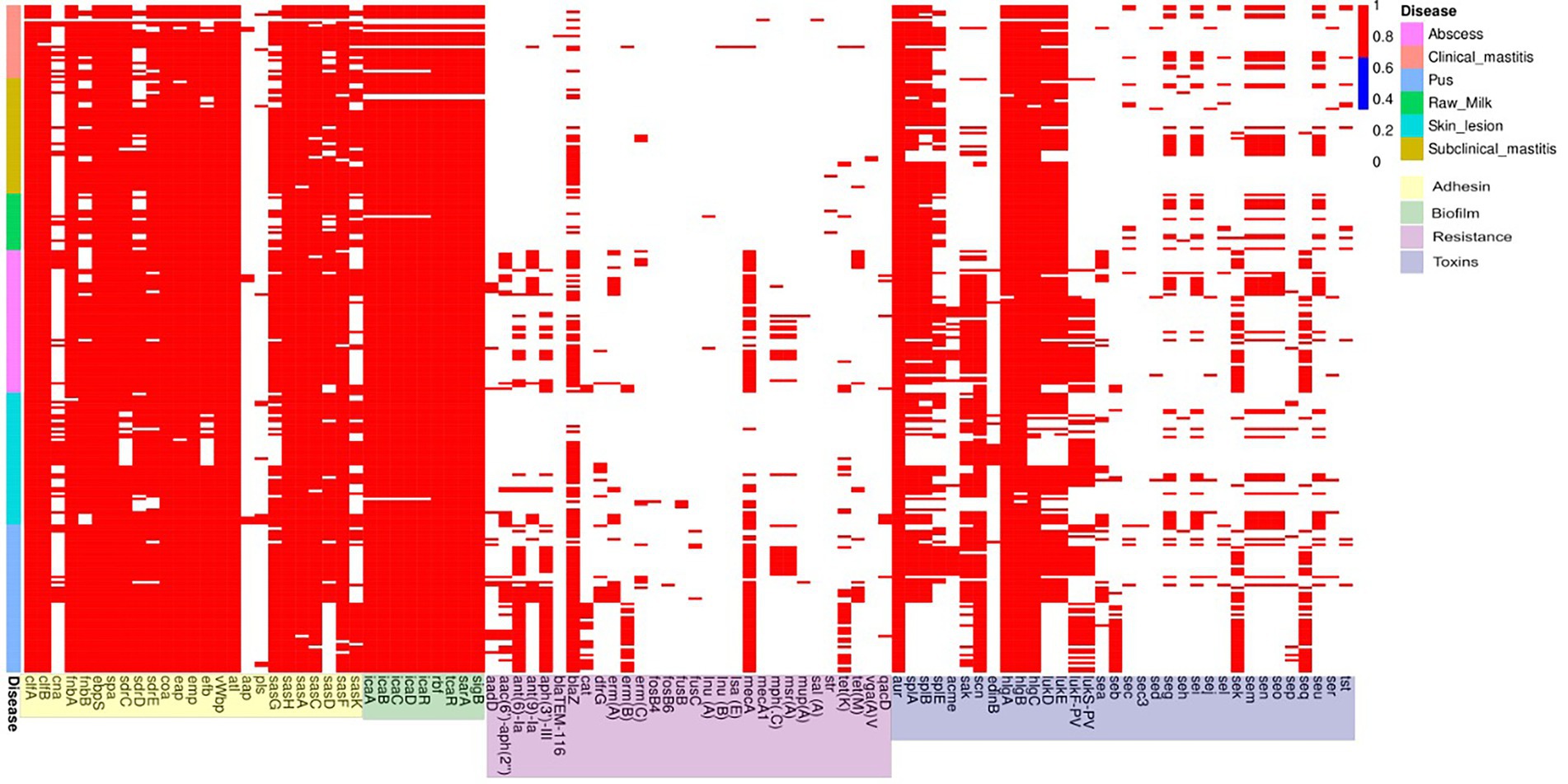
Figure 4. Binary frequency graph of adhesion, biofilm, antimicrobial resistance, and toxin gene frequencies in S. aureus from clinical and subclinical bovine mastitis cases and raw milk, and from human abscesses, skin infections, and pus.
The antimicrobial resistance genes aadD, aac(6′)/aph(2″), aadE, ant(9)-Ia, aph(3′)-III, cat(pC233), dfrG, erm(A), erm(B), mecA, mph(C), msr(A), tet(K) and tet(M) were most prevalent in human isolates and only seen sporadically in bovine isolates. The blaTEM-116 erm(C), fosB4, fosB6, fusB, fusC, lnu(A), lnu(B), lsa(E), mecA1, mup(A), sal(A), str, vga(A)V and qacD genes were observed sporadically in all groups. The blaZ gene was present (albeit unevenly) in all groups while the mecA gene was detected only in bovine isolates; however, both were prevalent (> 90%) in pus isolates (Figure 4). The aur, hlgA, hlgB and hlgC toxin genes were detected in >90% of the isolates of the all groups; the prevalence of the splA gene in the clinical mastitis isolates and raw milk, splB in raw milk, and scn in pus isolates was also >90%. The lukD and lukE genes were also prevalent in isolates from raw milk and abscesses, and as well as lukF-PV and lukS-PV, they were equivalent to each other in almost all groups evaluated (Figure 4).
Phylogenetic analysis of the S. aureus isolates in this study revealed that the clinical and subclinical cattle isolates formed distinct clades consistent with zoonotic and/or zooanthroponotic transmission (Figure 5). Some STs were more relevant when considered their respective origins? In this regard, ST239 and ST5 isolates were only recovered from humans and ST151 only in bovine isolates. ST8 was more frequent in abscess isolates, with the ST97 in subclinical mastitis, ST59 in pus isolates and ST152 in skin infection. These findings reinforces the epidemiological importance of these STs (Figure 5; Supplementary Material 3).
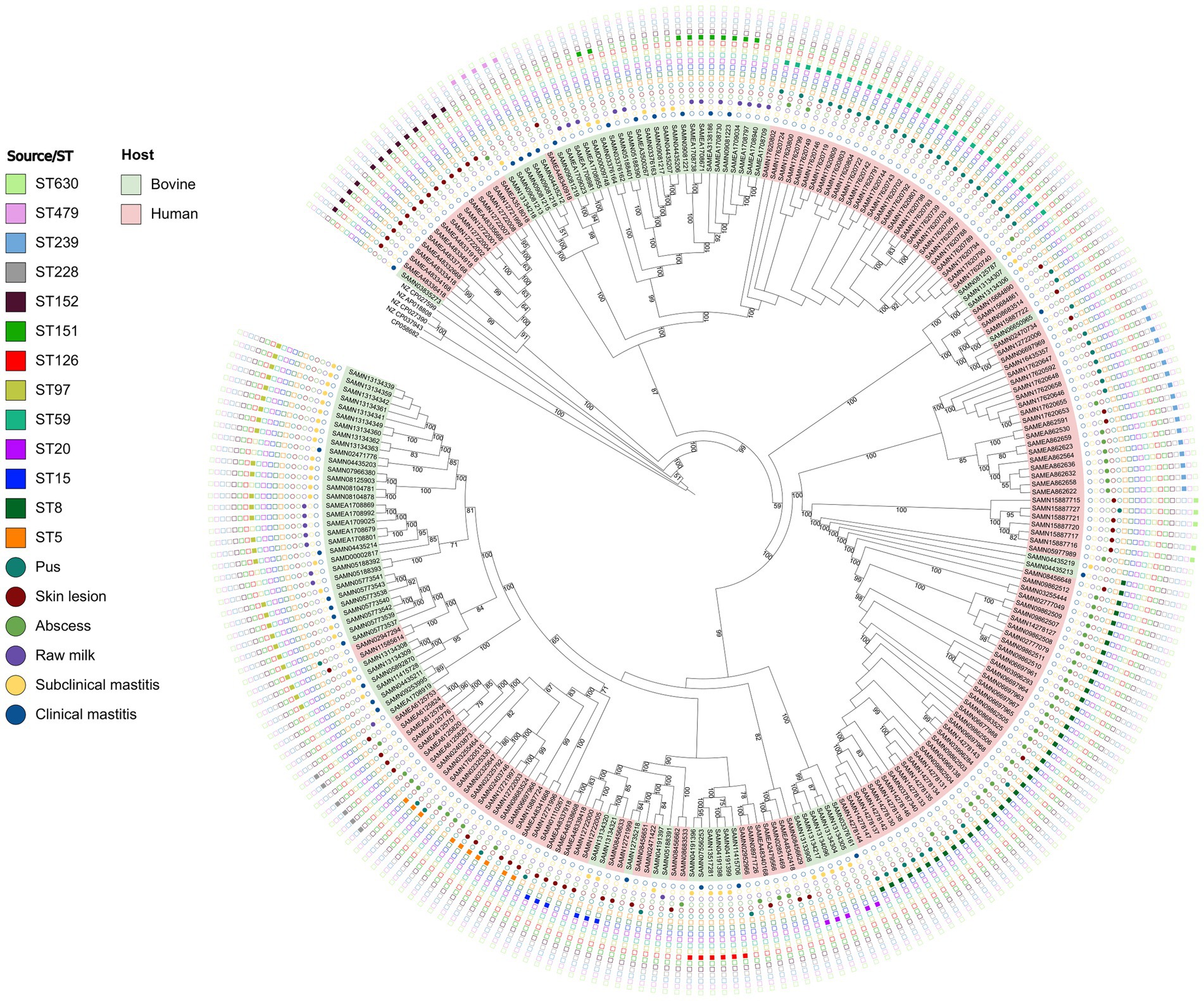
Figure 5. Phylogenomic tree based on 248 genes shared across all analyzed Staphylococcus aureus isolated from bovine clinical mastitis and subclinical mastitis cases and raw milk and from human abscesses, skin, and pus infections.
Forty-eight sequence types (ST) were assigned by MLST analysis. ST8 (15.7%), ST97 (12.1%), ST59 (11.7%), ST239 (6.85%), and ST152 (5.24%) were most prevalent. Eleven isolates could not be typed. The most common protein A (t) types as characterized by spa typing [i.e., t008 (n = 32), t437 (n = 26), t355 (n = 11), and t267 (n = 8)] were categorized according to source of isolation, host, and ST (Supplementary Material 3).
Using the Fisher’s test as a screening step to test the significance of all genes based on their origin it was possible to observe that nine adhesins, nine biofilm, 18 resistance, and 25 toxin genes were significant (p < 0.05; Table 1).

Table 1. Significant gene profile by Fisher’s exact test and by multinomial logistic regression according to the origin of the isolate and the main sequence type.
Multinomial logistic regression analysis showed that most genes had a significant relationship with the origin of the isolate, except for some antimicrobial resistance and toxin genes. The blaZ gene was the only antimicrobial resistance gene did not have a significant association with clinical mastitis isolates and, in conjunction the str, splB and sek genes, with subclinical mastitis isolates. The fusB, fusC, tet(M) and tst genes were not significantly associated with abscess isolates, nor were the aadE and tet(M) genes associated with skin infection isolates, or the aac(6′)-aph(2′), drfG and tet(M) genes to pus isolates. Multinomial logistic regression analysis also showed that there was significant relationships between origin and MLST with clinical mastitis (ST97), abscess (ST8) and skin infection isolates (ST152)., (Table 1; Supplementary Material 3).
With S. aureus isolates from clinical mastitis cases, there was a correlation, close to or equal to 1, between the clfA, fnbA, ebpS, spa, sdrC, coa, emp, vWbP, atl, sasH, sasA, sasF adhesin genes, and the aur, hlgA, hlgB and hlgC toxin genes (Figure 6). As well, there was a close association between the fnbB, sasK and sasG adhesin genes from this source. In the raw milk isolates, a high correlation was observed with the sdrD, sasK, fnbA and splE genes. In human isolates, there was a high correlation of the adhesion genes efb, sdrC and sasG and in skin infections and sasG and sdrE and in abscess isolates. All biofilm formation genes showed a high correlation with S. aureus from clinical and subclinical mastitis cases. In addition, the icaADBC operon and the icaR regulator were highly correlated in raw milk isolates and skin infections (Figure 6; Supplementary Material 4).
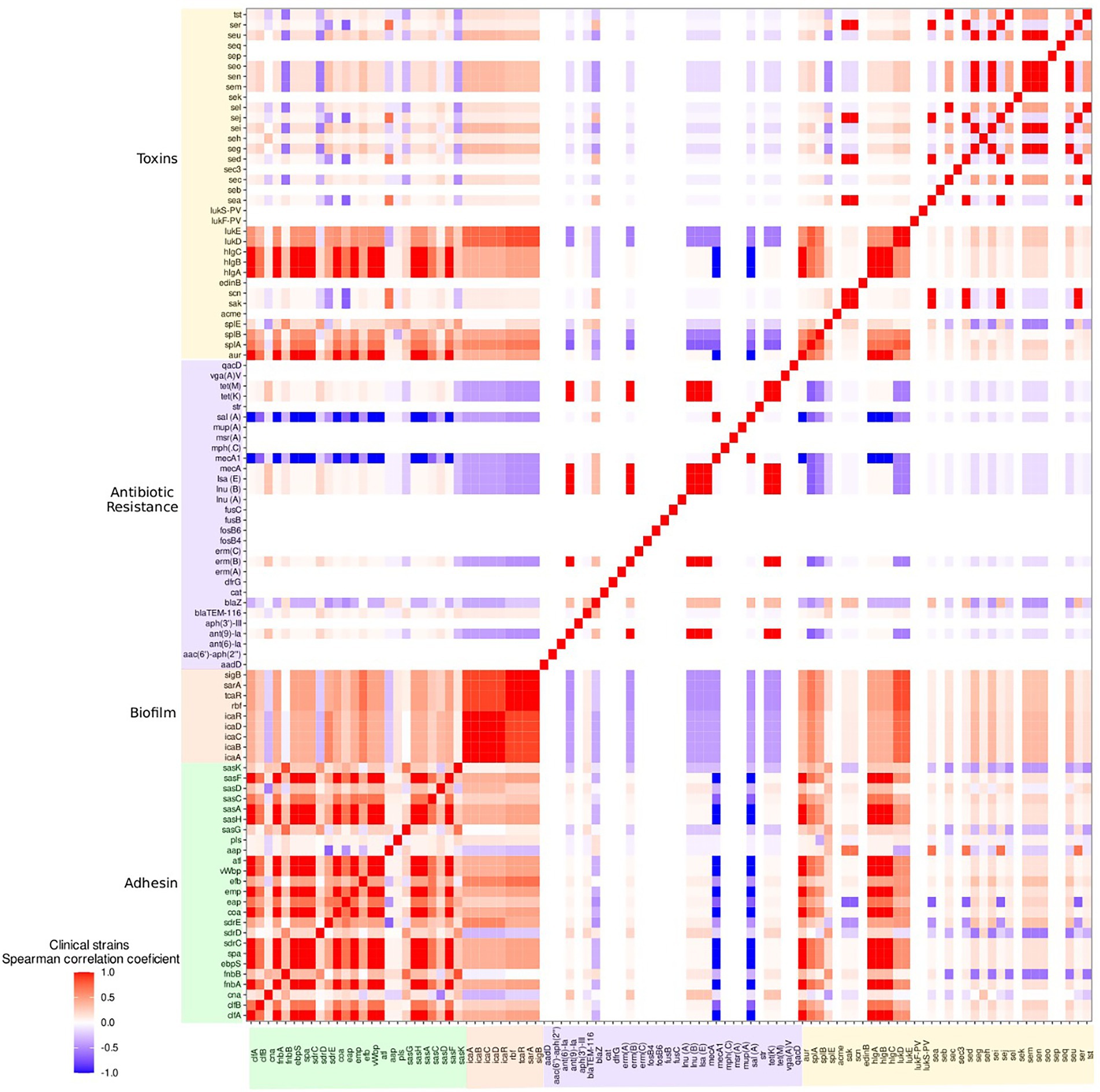
Figure 6. Heat map of the correlation, by the Spearman test, of the presence of S. aureus adhesion, biofilm, toxin, and antimicrobial resistance genes in clinical mastitis isolates.
Antimicrobial resistance genes ant(9)-Ia, erm(B), mecA, lsa(E), inu(B), tet(M) and tet(K) as well as toxin genes sak, scn, sea, sed, sej and ser were highly correlated in the clinical mastitis group. As well, there was a correlation between the seu, seg, sei, sen, sem and seo genes in all groups. There was a correlation between the sel, sec, and tst gene in all bovine isolates. Further, the sej, sed and ser genes were associated with subclinical mastitis and abscess cases; the seq and sek genes with skin infections; lukF-PV and lukS-PV with subclinical mastitis, skin, and pus infections; lukD and lukE with abscesses and skin infections; splA and splB with abscesses; and the lukD, lukE splA and splB genes with pus isolates (Supplementary Material 4).
TreeWAS analysis indicated that the scn gene was significant considering the Score 1 – “Terminal Score” which measures the measures sample-wide association across the leaves of the phylogenetic tree. The ant(6)-la, lukS-PV, lukF-PV, sak and scn were significant at the Score 2 – “Simultaneous Score” which measures the degree of parallel change in the phenotype and genotype across branches of the tree. None of the genes tested were significant at the Score 3 – “Subsequent Score” which measures the proportion of the tree in which the genotype and phenotype co-exist. These genes were further found to be significant with the multinomial logistic regression, alongside other ones mentioned above (Supplementary Material 5).
Adhesion is a crucial step in S. aureus colonization of the host and occurs before cell penetration, internalization, and chronic infection (Josse et al., 2017). The development of biofilms aids in evasion of host antimicrobial components and facilitates the exchange of genetic material (Idrees et al., 2021). Adhesin and biofilm genes clfA, clfB, fnbA, ebpS, spa, coa, eap, emp, vWbp, atl, sasH, sasA, sasC, sasF, icaR, rbf, tcaR, sarA, sigB and icaADBC were observed in >90% of the isolates. On the other hand, toxin and antimicrobial resistance genes were uncommon consistent with the notion that adhesion and biofilm formation play major roles in the pathogenesis of S. aureus (Petrie et al., 2020). In this regard, the clfA and the clfB genes, reported to promote microbial internalization in bovine mastitis, were present in almost all strains (Ying et al., 2021). Also, the ebpS gene, related to binding to elastin peptides in the host (Downer et al., 2002) was also very common (>95%). Similarly, the fnbA and fnbB genes responsible for adherence to immobilized elastin (Roche et al., 2004) were present in all groups, but fnbB gene with higher frequencies (>90%) in skin and pus infections. The high levels of expression of this gene are associated with the ability of S. aureus to internalize bovine mammary epithelial cells (Pereyra et al., 2016). Further, it was observed that fnbB was associated with sdrD, sasG and sasK in raw milk strains, and with sasC and sasK in clinical mastitis isolates (Supplementary Material 4). This is potentially an important association as the sasG and sdrD genes are reported to be involved in adhesion to epithelial cells (Roche et al., 2003; Abdelbary et al., 2020) and they play a role in cell aggregation and biofilm formation (Schroeder et al., 2009). In addition, our findings reinforce the recent observation that the fnbB gene is one of the main biofilm markers in S. aureus (Kadkhoda et al., 2020) and suggest that the sasK gene may also play an important role as well. The frequency of sdrC, sdrD and sdrE was lower in skin isolates in comparison with the other sources and sdrD was present less frequently in samples obtained from cattle. Thus, the sdr genes may not be important for epithelial tissue colonization and the sdrD gene may not play a key role in the adhesion to mammary gland and the establishment of bovine mastitis; however, further experiments are needed to test this hypothesis in vivo.
The adhesin genes sasC, sasA, sasF and sasH were present in >90% of the isolates from all sources. The sasA gene has been described in gangrenous mastitis and sasF in bovine mastitis isolates (Pizauro et al., 2021) but to date, little is known about sasH (Ythier et al., 2012). The observed lower frequencies (less than 50%) of the cna, pls and aap genes may be associated with the fact that the cna gene expresses a collagen-binding protein that is more often associated with skeletal muscle (Smeltzer and Gillaspy, 2000).The absence of emp and the presence of pls has been reported to be correlated with reduced virulence (Kurlenda et al., 2008), which was inversely observed (low frequency of pls and high of emp; Supplementary Material 2). The Emp and Eap proteins are also reported to be involved in in vivo biofilm formation and their expression is dependent on the icaADBC operon (Johnson et al., 2008). These genes, emp and eap, were observed in >90% of the strains in all sources and therefore, thought to be relevant in abscess formation and staphylococcal persistence. Also, in clinical mastitis isolates, the vWbp and coa genes showed a strongly correlation with each other (Figure 6), which is in agreement with the fact that these genes promote infection through prothrombin activation (Pickering et al., 2021).
The aap gene was present in less than 10% of isolates, whereas the ica locus and the rbf, tcaR, saA and sigB genes were present in >90% of the strains studied, except for clinical mastitis isolates (Supplementary Material 2). These differences may be related to the fact that PIA is the main exopolysaccharide synthesized by the icaADBC operon and that biofilm formation is less important in acute disease than in the maintenance of the strains in the host (Arciola et al., 2015). The association between icaA, icaB, icaC, icaD, and icaR was seen in strains isolated from raw milk and skin infections (Supplementary Material 4). Here, we identified that sasG and fnbB were very common in all groups evaluated (Figure 4) with the former having a frequency > 90% in abscess and pus isolates. Both sasG and fnbB were associated with invasive disease (Rasmussen et al., 2013) and the results showed a low correlation between these genes and clinical mastitis and raw milk isolates, with no association with rbf, tcaR, sarA and sigB genes and icaR (Supplementary Material 4) suggesting the importance of surface proteins in biofilm formation (Vergara-Irigaray et al., 2009; Geoghegan et al., 2010). Previous findings reported a high frequency of association of adhesion (ebpS, atl, pls, sasH and sasF) and biofilm-related genes (icaABCD) in Staphylococcus spp. from clinical mastitis (Pizauro et al., 2021). These results were also observed in the present study in clinical isolates in comparison with other sources, supporting the hypothesis that a more complex adherence and persistence mechanism may be required for S. aureus to be able to infect and persist in the bovine udder. That said, the staphylococci pangenome of clinical and subclinical mastitis isolates is still open so more sampling is required for more comprehensive analyses.
The toxin genes aur, hglA, hglB and hglC toxin genes were the only ones present in >90% in all groups evaluated. These genes were associated with the cflA, fnbA, ebpS, spa, sdrC, coa, emp, vWF, atl, sasH, sasA and sasF adhesin genes in clinical mastitis isolates (Figure 5). Aureolysin (expressed by the aur gene) modifies the adhesion factor CflB and activates other proteases that potentiate the virulence of S. aureus (McAleese et al., 2001). The proteins expressed by the hglA, hglB and hglC genes are bi-component leukotoxins that can form pores in the cell membrane and, consequently, lyse cells (Staali and Colin, 2021). Accordingly, the association of these genes products in clinical mastitis may be important targets for the development of vaccines and therapeutic agents (Ahmad-Mansour et al., 2021). Another association observed was the enterotoxins sec, sei, sen, sem, seo and seu in all evaluated groups, their frequencies were also similar; which have been observed by other studies (Indrawattana et al., 2013; Schwan, 2019; Ren et al., 2020). Also, previous studies have shown that see and sec, sel and tst are frequently present in MRSA strains (Hu et al., 2011), but this relationship was not observed in the current study. An association between the sec, sel and tst genes was seen only in bovine isolates, where the frequency of the mecA gene was low. The importance of these toxin genes in bovine mastitis has been highlighted previously (e.g., Fang et al., 2019). The lukD and lukE genes showed high association with themselves in isolates from abscesses, skin infections and pus; they were also considered significant for all groups in logistic regression but had higher frequencies in bovine isolates (Figure 3; Table 1). The genes lukF-PV and lukS-PV were correlated in all groups that were present, which is consistent with the literature since their products are secreted before joining to form the PVL toxin (Panton-Valentine leukocidin; Kaneko and Kamio, 2004).
More antimicrobial resistance genes were observed in human than in bovine isolates. Notably, the aminoglycoside resistance genes (aadD, aac(6′)/aph(2″), aadE, ant(9)-Ia, aph(3′)-III), chloramphenicol [cat(pC233)], tetracyclines [tet(K), tet(M)], macrolides and lincosamines [erm(A), erm(B), mph(C)), msr(A)], trimethoprim (dfrG) and β-Lactams (blaZ, mecA) were detected mostly in human isolates. Nevertheless, bovine strains presented β-lactam (blaZ), macrolide (mphC), macrolide, lincosamide, and streptogramin B (msrA), aminoglycoside (aadD), and tetracycline (tetK), all that have been described before (Pizauro et al., 2019). Previous studies detected high prevalence of aadD genes, aac(6′)/aph(2″), tet(M), msr(A), aph(3′)-III, erm(A) and erm(B) in S. aureus from patients admitted to ICUs (Abiri et al., 2017); as well as the presence of tet(K), tet(M), mph(C), msr(A) and other genes in nosocomial strains have been reported (Lozano et al., 2012). These relationships were also observed in the present study, and the tet(M) gene showed a high correlation with the erm(A), dfrG and ant(9)-Ia genes in pus, skin and abscess isolates with the addition of the cat(pC233) gene. In summary, the results of this study reinforce the growing presence of resistance genes in human strains, while pointing to a relatively low level of resistance genes in strains of bovine origin (Sweileh, 2021). There was an association of ant(9)-Ia, mecA, tet(M) and tet(K), erm(B), lsa(E) and lnu(B) genes, (which confer resistance to aminoglycosides, β-lactams, tetracyclines and macrolides and lincosamides) in the clinical mastitis group (Figure 5). Similarly, the ACME, msr(A) and mph(C) genes in pus isolates showed a low frequency.
The phylogenomic analysis was consistent with zoonotic transmission that the S. aureus is known for (Silva et al., 2022). This could be attributed to the evidence that suggests that some S. aureus strains are adapted to colonize and infect certain host species, whilst other lineages are non-specific (Schmidt et al., 2017). This behavior was highlighted in this study (Figure 5) with a high frequency observed for ST152 in skin lesions, ST 59 in pus suggesting their host specificity. Nevertheless, it was possible to observe that ST97 is predominantly associated with clinical mastitis, subclinical mastitis, and raw milk isolates and ST8 is predominantly associated with abscess and pus which indicates that they may be non-specific. In addition, some strains, from different origins share the same ST (e.g., ST8, ST15, and ST97; Figure 5). Bovine isolates were frequently ST97 and ST151, while most ST8, ST59, ST152, ST239 were from human sources. That said, these sequence types have been associated with both hosts consistent with zoonotic transmission in previous studies (Ndahetuye et al., 2021). The ST59, ST239, ST5, ST228, ST630, ST30, ST80, ST45 and ST88 were often from human cases in the current study. ST59 has been frequently observed in the community and more frequently in food samples (Pang et al., 2020). ST239 is globally disseminated and resistant to many antimicrobials commonly used in hospitals (Wang et al., 2014). As well, ST228, has high transmissibility (Abdelbary et al., 2020), while ST630 has high pathogenicity (Zong et al., 2020) and ST45 responsible for serious invasive diseases (Effelsberg et al., 2020). ST80 and ST88 seem to be mainly associated with the community (Stam-Bolink et al., 2007) and with animals, food and livestock ranchers (Otalu et al., 2018). ST151, ST126, ST479, ST20, ST71, ST133, ST504, ST115, ST425, ST737 and ST3183 were identified only in bovine isolates. ST151 has been frequently reported as bovine lineage common in cases of mastitis, susceptible to the acquisition of vancomycin resistance genes (Guinane et al., 2008); similar to the reported distribution of ST126 in bovine milk (Silva et al., 2016). Mainly described as clinical and subclinical mastitis strains, ST71 belongs to the same lineage as ST97, with more and more isolates demonstrated to be MRSA (Cormican and Keane, 2018), and in the present work they were present in bovine isolates. ST479 was only seen in milk samples (Chenouf et al., 2021); ST115 was consistent with bovine isolates (Smith et al., 2005) and ST3183 was recently identified in raw milk samples (McMillan et al., 2016). ST20 was cited in strains bovines and humans (Aung et al., 2019), and ST133 was cited in several hosts and environmental samples (Roberts et al., 2013). ST425 is currently characterized as a lineage associated with humans and animals (Paterson et al., 2014). Finally, ST737 was reported to be associated with the community, hospital environment (Karbuz et al., 2017) and cattle (Türkyilmaz and Erdem, 2013).
Our findings suggest that the cflA, fnbA, ebpS, spa, sdrC, coa, emp, vWF, atl, sasH, sasA and sasF adhesion genes, as well the aur, hglA, hglB, and hglC toxin genes are associated with clinical mastitis, and thus could be useful for screening tests and as candidates for more effectives vaccines. Further, the surface protein genes sasG and fnbB appear to be necessary for the interaction of S. aureus with cattle. It is also notewothy that the blaZ and mecA genes are frequently present in pus isolates. Finally, some STs have an epidemiological relationship with the type of disease, also showing zoonotic behavior.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
All authors listed have made a substantial, direct, and intellectual contribution to the work, and approved it for publication.
This work was carried out with the support of the Coordination for the Improvement of Higher Education Personnel – Brazil (CAPES) – Financing Code 001.
To the Postgraduate Program in Agricultural Microbiology, Universidade Estadual Paulista (UNESP), Faculty of Agricultural and Veterinary Sciences, Jaboticabal, São Paulo state of Brazil.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1033675/full#supplementary-material
Abdelbary, M. M. H., Feil, E. J., Senn, L., Petignat, C., Prod’hom, G., Schrenzel, J., et al. (2020). Phylogeographical analysis reveals the historic origin, emergence, and evolutionary dynamics of methicillin-resistant Staphylococcus aureus ST228. Front. Microbiol. 11:2063. doi: 10.3389/fmicb.2020.02063
Abiri, P., Akhavan Sepahi, A., Goudarzi, H., and Goudarzi, M. (2017). Distribution of genes encoding toxin, adhesion, and antibacterial resistance among various SCCmec types of methicillin-resistant Staphylococcus aureus isolated from intensive care unit, Tehran, Iran. Jundishapur Journal of Microbiology 10. doi: 10.5812/jjm.14477
Ahmad-Mansour, N., Loubet, P., Pouget, C., Dunyach-Remy, C., Sotto, A., Lavigne, J.-P., et al. (2021). Staphylococcus aureus toxins: an update on their pathogenic properties and potential treatments. Toxins 13:677. doi: 10.3390/toxins13100677
Akaike, H. (1974). A new look at the statistical model identification. IEEE Trans. Automat. Contr. 19, 716–723. doi: 10.1109/TAC.1974.1100705
Arciola, C. R., Campoccia, D., Ravaioli, S., and Montanaro, L. (2015). Polysaccharide intercellular adhesin in biofilm: structural and regulatory aspects. Front. Cell. Infect. Microbiol. 5:7. doi: 10.3389/fcimb.2015.00007
Aung, M. S., Urushibara, N., Kawaguchiya, M., Sumi, A., Shinagawa, M., Takahashi, S., et al. (2019). Clonal diversity and genetic characteristics of methicillin-resistant Staphylococcus aureus isolates from a tertiary care hospital in Japan. Microb. Drug Resist. 25, 1164–1175. doi: 10.1089/mdr.2018.0468
Åvall-Jääskeläinen, S., Koort, J., Simojoki, H., and Taponen, S. (2021). Genomic analysis of Staphylococcus aureus isolates associated with peracute non-gangrenous or gangrenous mastitis and comparison with other mastitis-associated Staphylococcus aureus isolates. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.688819
Aziz, R. K., Bartels, D., Best, A. A., DeJongh, M., Disz, T., Edwards, R. A., et al. (2008). The RAST server: rapid annotations using subsystems technology. BMC Genomics 9:75. doi: 10.1186/1471-2164-9-75
Bartels, M. D., Petersen, A., Worning, P., Nielsen, J. B., Larner-Svensson, H., Johansen, H. K., et al. (2014). Comparing whole-genome sequencing with Sanger sequencing for spa typing of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 52, 4305–4308. doi: 10.1128/JCM.01979-14
Bencardino, D., Amagliani, G., and Brandi, G. (2021). Carriage of Staphylococcus aureus among food handlers: an ongoing challenge in public health. Food Control 130:108362. doi: 10.1016/j.foodcont.2021.108362
Bortolaia, V., Kaas, R. S., Ruppe, E., Roberts, M. C., Schwarz, S., Cattoir, V., et al. (2020). ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 75, 3491–3500. doi: 10.1093/jac/dkaa345
Bruce, S. A., Smith, J. T., Mydosh, J. L., Ball, J., Needle, D. B., Gibson, R., et al. (2022). Shared antibiotic resistance and virulence genes in Staphylococcus aureus from diverse animal hosts. Sci. Rep. 12:4413. doi: 10.1038/s41598-022-08230-z
Chen, L., Yang, J., Yu, J., Yao, Z., Sun, L., Shen, Y., et al. (2005). VFDB: a reference database for bacterial virulence factors. Nucleic Acids Res. 33, D325–D328. doi: 10.1093/nar/gki008
Chenouf, N. S., Mama, O. M., Messaï, C. R., Ruiz-Ripa, L., Fernández-Fernández, R., Carvalho, I., et al. (2021). Detection of methicillin-resistant coagulase-negative staphylococci and PVL/mecA genes in cefoxitin-susceptible Staphylococcus aureus (t044/ST80) from unpasteurized milk sold in stores in Djelfa, Algeria. J. Dairy Sci. 104, 2684–2692. doi: 10.3168/jds.2020-19270
Clark, K., Karsch-Mizrachi, I., Lipman, D. J., Ostell, J., and Sayers, E. W. (2016). GenBank. Nucleic Acids Res. 44, D67–D72. doi: 10.1093/nar/gkv1276
Collins, C., and Didelot, X. (2018). A phylogenetic method to perform genome-wide association studies in microbes that accounts for population structure and recombination. PLoS Comput. Biol. 14:e1005958. doi: 10.1371/journal.pcbi.1005958
R Core Team (2017). R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing.
Cormican, P., and Keane, O. M. (2018). Complete genome sequences of sequence type 71 (ST71) and ST97 Staphylococcus aureus isolates from bovine milk. Microbiol. Resour. Announc. 7, e00954–e00918. doi: 10.1128/MRA.00954-18
Downer, R., Roche, F., Park, P. W., Mecham, R. P., and Foster, T. J. (2002). The Elastin-binding protein of Staphylococcus aureus (EbpS) is expressed at the cell surface as an integral membrane protein and not as a cell eall-associated protein. J. Biol. Chem. 277, 243–250. doi: 10.1074/jbc.M107621200
Effelsberg, N., Stegger, M., Peitzmann, L., Altinok, O., Coombs, G. W., Pichon, B., et al. (2020). Global epidemiology and evolutionary history of Staphylococcus aureus ST45. J. Clin. Microbiol. 59, e02198–e02120. doi: 10.1128/JCM.02198-20
Exel, C. E., Halasa, T., Koop, G., Steeneveld, W., Lam, T. J. G. M., Benedictus, L., et al. (2022). A stochastic modelling approach to determine the effect of diverse Staphylococcus aureus strains on the economic and epidemiological outcomes of mastitis intervention strategies in dairy cattle. Prev. Vet. Med. 199:105566. doi: 10.1016/j.prevetmed.2021.105566
Fang, R., Cui, J., Cui, T., Guo, H., Ono, H. K., Park, C.-H., et al. (2019). Staphylococcal enterotoxin C is an important virulence factor for mastitis. Toxins 11:E141. doi: 10.3390/toxins11030141
Gajewska, J., and Chajęcka-Wierzchowska, W. (2020). Biofilm formation ability and presence of adhesion genes among coagulase-negative and coagulase-positive staphylococci isolates from raw cow’s milk. Pathogens 9:654. doi: 10.3390/pathogens9080654
Geoghegan, J. A., Corrigan, R. M., Gruszka, D. T., Speziale, P., O’Gara, J. P., Potts, J. R., et al. (2010). Role of surface protein SasG in biofilm formation by Staphylococcus aureus. J. Bacteriol. 192, 5663–5673. doi: 10.1128/JB.00628-10
Guinane, C. M., Sturdevant, D. E., Herron-Olson, L., Otto, M., Smyth, D. S., Villaruz, A. E., et al. (2008). Pathogenomic analysis of the common bovine Staphylococcus aureus clone (ET3): emergence of a virulent subtype with potential risk to public health. J. Infect. Dis. 197, 205–213. doi: 10.1086/524689
Hoang, D. T., Chernomor, O., von Haeseler, A., Minh, B. Q., and Vinh, L. S. (2018). UFBoot2: improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 35, 518–522. doi: 10.1093/molbev/msx281
Hu, D.-L., Maina, E. K., Omoe, K., Inoue, F., Yasujima, M., and Nakane, A. (2011). Superantigenic toxin genes coexist with specific staphylococcal cassette chromosome mec genes in methicillin-resistant Staphylococcus aureus. Tohoku J. Exp. Med. 225, 161–169. doi: 10.1620/tjem.225.161
Idrees, M., Sawant, S., Karodia, N., and Rahman, A. (2021). Staphylococcus aureus biofilm: morphology, genetics, pathogenesis and treatment strategies. Int. J. Environ. Res. Public Health 18:7602. doi: 10.3390/ijerph18147602
Igbinosa, E. O., Beshiru, A., Akporehe, L. U., and Ogofure, A. G. (2016). Detection of methicillin-resistant Staphylococci isolated from food producing animals: a public health implication. Vet. Sci. 3:14. doi: 10.3390/vetsci3030014
Indrawattana, N., Sungkhachat, O., Sookrung, N., Chongsa-nguan, M., Tungtrongchitr, A., Voravuthikunchai, S. P., et al. (2013). Staphylococcus aureus clinical isolates: antibiotic susceptibility, molecular characteristics, and ability to form biofilm. Biomed. Res. Int. 2013:e314654. doi: 10.1155/2013/314654
Johnson, M., Cockayne, A., and Morrissey, J. A. (2008). Iron-regulated biofilm formation in Staphylococcus aureus Newman requires Ica and the secreted protein Emp. Infect. Immun. 76, 1756–1765. doi: 10.1128/IAI.01635-07
Josse, J., Laurent, F., and Diot, A. (2017). Staphylococcal adhesion and host cell invasion: fibronectin-binding and other mechanisms. Front. Microbiol. 8:2433. doi: 10.3389/fmicb.2017.02433
Kadkhoda, H., Ghalavand, Z., Nikmanesh, B., Kodori, M., Houri, H., Taghizadeh Maleki, D., et al. (2020). Characterization of biofilm formation and virulence factors of Staphylococcus aureus isolates from paediatric patients in Tehran, Iran. Iran. J. Basic Med. Sci. 23, 691–698. doi: 10.22038/ijbms.2020.36299.8644
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., von Haeseler, A., and Jermiin, L. S. (2017). ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 14, 587–589. doi: 10.1038/nmeth.4285
Kaneko, J., and Kamio, Y. (2004). Bacterial two-component and hetero-heptameric pore-forming cytolytic toxins: structures, pore-forming mechanism, and organization of the genes. Biosci. Biotechnol. Biochem. 68, 981–1003. doi: 10.1271/bbb.68.981
Karbuz, A., Karahan, Z. C., Aldemir-Kocabaş, B., Tekeli, A., Özdemir, H., Güriz, H., et al. (2017). Evaluation of antimicrobial susceptibilities and virulence factors of Staphylococcus aureus strains isolated from community-acquired and health-care associated pediatric infections. Turk. J. Pediatr. 59, 395–403. doi: 10.24953/turkjped.2017.04.005
Kerro Dego, O. (2020). Bovine mastitis: part I. Anim. Reproduct. Vet. Med. doi: 10.5772/intechopen.93483
Kurlenda, J., Grinholc, M., and Węgrzyn, G. (2008). Presence of cna, emp and pls genes and pathogenicity of methicillin-resistant Staphylococcus aureus strains. World J. Microbiol. Biotechnol. 24, 591–594. doi: 10.1007/s11274-007-9511-7
Lalaouna, D., Desgranges, E., Caldelari, I., and Marzi, S. (2018). “Chapter sixteen - MS2-affinity purification coupled with RNA sequencing approach in the human pathogen Staphylococcus aureus,” in Methods in Enzymology High-Density Sequencing Applications in Microbial Molecular Genetics. ed. A. J. Carpousis (Cambridge, MA: Academic Press), 393–411.
Larsen, M. V., Cosentino, S., Rasmussen, S., Friis, C., Hasman, H., Marvig, R. L., et al. (2012). Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 50, 1355–1361. doi: 10.1128/JCM.06094-11
Letunic, I., and Bork, P. (2016). Interactive tree of life (iTOL) v3: an online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 44, W242–W245. doi: 10.1093/nar/gkw290
Löytynoja, A. (2014). “Phylogeny-aware alignment with PRANK,” in Multiple Sequence Alignment Methods Methods in Molecular Biology., ed. D. J. Russell Totowa, NJ: Humana Press, 155–170.
Lozano, C., Rezusta, A., Gómez, P., Gómez-Sanz, E., Báez, N., Martin-Saco, G., et al. (2012). High prevalence of spa types associated with the clonal lineage CC398 among tetracycline-resistant methicillin-resistant Staphylococcus aureus strains in a Spanish hospital. J. Antimicrob. Chemother. 67, 330–334. doi: 10.1093/jac/dkr497
McAleese, F. M., Walsh, E. J., Sieprawska, M., Potempa, J., and Foster, T. J. (2001). Loss of clumping factor B fibrinogen binding activity by Staphylococcus aureus involves cessation of transcription, shedding and cleavage by metalloprotease. J. Biol. Chem. 276, 29969–29978. doi: 10.1074/jbc.M102389200
McMillan, K., Moore, S. C., McAuley, C. M., Fegan, N., and Fox, E. M. (2016). Characterization of Staphylococcus aureus isolates from raw milk sources in Victoria, Australia. BMC Microbiol. 16:169. doi: 10.1186/s12866-016-0789-1
Minh, B. Q., Schmidt, H. A., Chernomor, O., Schrempf, D., Woodhams, M. D., von Haeseler, A., et al. (2020). IQ-TREE 2: new models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 37, 1530–1534. doi: 10.1093/molbev/msaa015
Ndahetuye, J. B., Leijon, M., Båge, R., Artursson, K., and Persson, Y. (2021). Genetic characterization of Staphylococcus aureus from subclinical mastitis cases in dairy cows in Rwanda. Front. Vet. Sci. 8:751229. doi: 10.3389/fvets.2021.751229
Otalu, O. J., Kwaga, J. K. P., Okolocha, E. C., Islam, M. Z., and Moodley, A. (2018). High genetic similarity of MRSA ST88 isolated from pigs and humans in Kogi state, Nigeria. Front. Microbiol. 9:3098. doi: 10.3389/fmicb.2018.03098
Otto, M. (2014). Staphylococcus aureus toxins. Curr. Opin. Microbiol. 17, 32–37. doi: 10.1016/j.mib.2013.11.004
Page, A. J., Cummins, C. A., Hunt, M., Wong, V. K., Reuter, S., Holden, M. T. G., et al. (2015). Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics 31, 3691–3693. doi: 10.1093/bioinformatics/btv421
Pang, R., Wu, S., Zhang, F., Huang, J., Wu, H., Zhang, J., et al. (2020). The genomic context for the evolution and transmission of community-associated Staphylococcus aureus ST59 through the food chain. Front. Microbiol. 11:422. doi: 10.3389/fmicb.2020.00422
Patel, K., Godden, S. M., Royster, E. E., Crooker, B. A., Johnson, T. J., and Smith, E. A. (2021). Prevalence, antibiotic resistance, virulence and genetic diversity of Staphylococcus aureus isolated from bulk tank milk samples of U.S. dairy herds. BMC Genomics 22:367. doi: 10.1186/s12864-021-07603-4
Paterson, G. K., Harrison, E. M., and Holmes, M. A. (2014). The emergence of mecC methicillin-resistant Staphylococcus aureus. Trends Microbiol. 22, 42–47. doi: 10.1016/j.tim.2013.11.003
Pereyra, E. A. L., Picech, F., Renna, M. S., Baravalle, C., Andreotti, C. S., Russi, R., et al. (2016). Detection of Staphylococcus aureus adhesion and biofilm-producing genes and their expression during internalization in bovine mammary epithelial cells. Vet. Microbiol. 183, 69–77. doi: 10.1016/j.vetmic.2015.12.002
Petrie, L. E., Leonard, A. C., Murphy, J., and Cox, G. (2020). Development and validation of a high-throughput whole cell assay to investigate Staphylococcus aureus adhesion to host ligands. J. Biol. Chem. 295, 16700–16712. doi: 10.1074/jbc.RA120.015360
Pickering, A. C., Yebra, G., Gong, X., Goncheva, M. I., Wee, B. A., MacFadyen, A. C., et al. (2021). Evolutionary and functional analysis of coagulase positivity among the Staphylococci 6, e00381–e00321. doi: 10.1128/mSphere.00381-21
Pizauro, L. J. L., de Almeida, C. C., Gohari, I. M., MacInnes, J. I., Zafalon, L. F., Kropinski, A. M., et al. (2019). Complete genome sequences of 11 Staphylococcus sp. strains isolated from buffalo milk and milkers’ hands. Microbiol. Resour. Announc. 8, e01264–e01219. doi: 10.1128/MRA.01264-19
Pizauro, L. J. L., de Almeida, C. C., Silva, S. R., MacInnes, J. I., Kropinski, A. M., Zafalon, L. F., et al. (2021). Genomic comparisons and phylogenetic analysis of mastitis-related staphylococci with a focus on adhesion, biofilm, and related regulatory genes. Sci. Rep. 11:17392. doi: 10.1038/s41598-021-96842-2
Rasmussen, G., Monecke, S., Ehricht, R., and Söderquist, B. (2013). Prevalence of clonal complexes and virulence genes among commensal and invasive Staphylococcus aureus isolates in Sweden. PLoS One 8:e77477. doi: 10.1371/journal.pone.0077477
Ren, Q., Liao, G., Wu, Z., Lv, J., and Chen, W. (2020). Prevalence and characterization of Staphylococcus aureus isolates from subclinical bovine mastitis in southern Xinjiang, China. J. Dairy Sci. 103, 3368–3380. doi: 10.3168/jds.2019-17420
Roberts, M. C., Soge, O. O., and No, D. (2013). Comparison of multi-drug resistant environmental methicillin-resistant Staphylococcus aureus isolated from recreational beaches and high touch surfaces in built environments. Front. Microbiol. 4:74. doi: 10.3389/fmicb.2013.00074
Roche, F. M., Downer, R., Keane, F., Speziale, P., Park, P. W., and Foster, T. J. (2004). The N-terminal A domain of fibronectin-binding proteins A and B promotes adhesion of Staphylococcus aureus to elastin. J. Biol. Chem. 279, 38433–38440. doi: 10.1074/jbc.M402122200
Roche, F. M., Meehan, M., and Foster, T. J. (2003). The Staphylococcus aureus surface protein SasG and its homologues promote bacterial adherence to human desquamated nasal epithelial cells. Microbiology 149, 2759–2767. doi: 10.1099/mic.0.26412-0
Schmidt, T., Kock, M. M., and Ehlers, M. M. (2017). Molecular characterization of Staphylococcus aureus isolated from bovine mastitis and close human contacts in south African dairy herds: genetic diversity and inter-species host transmission. Front. Microbiol. 8:511. doi: 10.3389/fmicb.2017.00511
Schroeder, K., Jularic, M., Horsburgh, S. M., Hirschhausen, N., Neumann, C., Bertling, A., et al. (2009). Molecular characterization of a novel Staphylococcus aureus surface protein (SasC) involved in cell aggregation and biofilm accumulation. PLoS One 4:e7567. doi: 10.1371/journal.pone.0007567
Schwan, W. R. (2019). Staphylococcus aureus toxins: armaments for a significant pathogen. Toxins 11:457. doi: 10.3390/toxins11080457
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Silva, V., Alfarela, C., Caniça, M., Manageiro, V., Nóvoa, M., Leiva, B., et al. (2022). A one health approach molecular analysis of Staphylococcus aureus reveals distinct lineages in isolates from Miranda donkeys (Equus asinus) and their handlers. Antibiotics 11:374. doi: 10.3390/antibiotics11030374
Silva, D. M., da Silva, M. P., Vidigal, P. M. P., Barcelos, R. M., Klein, R. C., Aguilar, A. P., et al. (2016). Draft genome sequences of Staphylococcus aureus strains isolated from bubclinical bovine mastitis in Brazil. Genome Announc. 4, e01594–e01515. doi: 10.1128/genomeA.01594-15
Smeltzer, M. S., and Gillaspy, A. F. (2000). Molecular pathogenesis of Staphylcoccal Osteomyelitis. Poult. Sci. 79, 1042–1049. doi: 10.1093/ps/79.7.1042
Smith, E. M., Green, L. E., Medley, G. F., Bird, H. E., Fox, L. K., Schukken, Y. H., et al. (2005). Multilocus sequence typing of intercontinental bovine Staphylococcus aureus isolates. J. Clin. Microbiol. 43, 4737–4743. doi: 10.1128/JCM.43.9.4737-4743.2005
Staali, L., and Colin, D. A. (2021). Bi-component HlgC/HlgB and HlgA/HlgB γ-hemolysins from S. aureus: modulation of Ca2+ channels activity through a differential mechanism. Toxicon 201, 74–85. doi: 10.1016/j.toxicon.2021.08.007
Stam-Bolink, E. M., Mithoe, D., Baas, W. H., Arends, J. P., and Möller, A. V. M. (2007). Spread of a methicillin-resistant Staphylococcus aureus ST80 strain in the community of the northern Netherlands. Eur. J. Clin. Microbiol. Infect. Dis. 26, 723–727. doi: 10.1007/s10096-007-0352-y
Sweileh, W. M. (2021). Global research publications on irrational use of antimicrobials: call for more research to contain antimicrobial resistance. Glob. Health 17:94. doi: 10.1186/s12992-021-00754-9
Türkyilmaz, S., and Erdem, Z. (2013). Molecular typing of methicillin resistant Staphylococcus aureus strains isolated from cows and farm workers. Vet. Fak. Derg. 19, 963–968. doi: 10.9775/kvfd.2013.9177
Vergara-Irigaray, M., Valle, J., Merino, N., Latasa, C., García, B., De Los Mozos, I. R., et al. (2009). Relevant role of fibronectin-binding proteins in Staphylococcus aureus biofilm-associated foreign-body infections. Infect. Immun. 77, 3978–3991. doi: 10.1128/IAI.00616-09
Wang, Z., Zhou, H., Wang, H., Chen, H., Leung, K. K., Tsui, S., et al. (2014). Comparative genomics of methicillin-resistant Staphylococcus aureus ST239: distinct geographical variants in Beijing and Hong Kong. BMC Genomics 15:529. doi: 10.1186/1471-2164-15-529
Watkins, K. E., and Unnikrishnan, M. (2020). “Chapter three – evasion of host defenses by intracellular Staphylococcus aureus,” in Advances in Applied Microbiology. eds. G. M. Gadd and S. Sariaslani (Oxford: Academic Press), 105–141.
Wójcik-Bojek, U., Różalska, B., and Sadowska, B. (2022). Staphylococcus aureus—a known opponent against host defense mechanisms and vaccine development—do we still have a chance to win? Int. J. Mol. Sci. 23:948. doi: 10.3390/ijms23020948
Ying, Y.-T., Ren, W.-J., Tan, X., Yang, J., Liu, R., and Du, A.-F. (2021). Annexin A2-mediated internalization of Staphylococcus aureus into bovine mammary epithelial cells requires its interaction with clumping factor B. Microorganisms 9:2090. doi: 10.3390/microorganisms9102090
Ythier, M., Resch, G., Waridel, P., Panchaud, A., Gfeller, A., Majcherczyk, P., et al. (2012). Proteomic and transcriptomic profiling of Staphylococcus aureus surface LPXTG-proteins: correlation with agr genotypes and adherence phenotypes. Mol. Cell. Proteom. 11, 1123–1139. doi: 10.1074/mcp.M111.014191
Zadoks, R. N., Middleton, J. R., McDougall, S., Katholm, J., and Schukken, Y. H. (2011). Molecular epidemiology of mastitis pathogens of dairy cattle and comparative relevance to humans. J. Mammary Gland Biol. Neoplasia 16, 357–372. doi: 10.1007/s10911-011-9236-y
Keywords: biofilm formation, public health, virulence genes, adhesins, phylogeny, antimicrobial resistance
Citation: Rodrigues RA, Pizauro LJL, Varani AM, de Almeida CC, Silva SR, Cardozo MV, MacInnes JI, Kropinski AM, Melo PC and Ávila FA (2022) Comparative genomics study of Staphylococcus aureus isolated from cattle and humans reveals virulence patterns exclusively associated with bovine clinical mastitis strains. Front. Microbiol. 13:1033675. doi: 10.3389/fmicb.2022.1033675
Edited by:
Ben Pascoe, University of Oxford, United KingdomReviewed by:
Evangelos Mourkas, University of Oxford, United KingdomCopyright © 2022 Rodrigues, Pizauro, Varani, de Almeida, Silva, Cardozo, MacInnes, Kropinski, Melo and Ávila. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Romário Alves Rodrigues, cm9tYXJpby5hLnJvZHJpZ3Vlc0B1bmVzcC5icg==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.