 Pamela Monaco
Pamela Monaco Gino Naclerio
Gino Naclerio Antonietta Mello
Antonietta Mello Antonio Bucci
Antonio Bucci
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Microbiol. , 06 October 2022
Sec. Microbial Symbioses
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.1017089
Among the hypogeous ectomycorrhizal fungi, the white truffle Tuber magnatum Picco is the species of greatest interest, both from an ecological and economic point of view. The increasing market demand of the precious white truffle along with the fall in its natural production led to a growing interest in cultivation techniques and encouraged truffle growers and researchers to deeper investigate factors that could affect and improve T. magnatum productivity. In this context, microbial communities play a central role. Indeed, in the last few years, the hypothesis of a potential link between microbial community composition and truffle orchard productivity is arousing a greater attention. Moreover, since the value of the prized T. magnatum can vary in relation to its provenience, the need to define a reliable tracking system is also emerging and bacteria appear to be a promising tool. Accordingly, the present mini-review summarises the knowledge currently available on T. magnatum microbial communities, focusing on the role of truffle-associated bacteria and highlighting similarities and differences between samples of different origin, to address the following issues: (i) Is there a correlation between microbial taxa and truffle ground productivity? (ii) Can bacteria actually be used as markers of T. magnatum geographic origin? The identification of microorganisms able to promote T. magnatum formation may represent an important advance in the field of truffle farming. Similarly, the detection of bacterial taxa that can be used as markers of T. magnatum origin could have a considerable impact on truffle industry and trade, even at local scale.
Most of terrestrial plant roots are colonised by mycorrhizal fungi; indeed, it is estimated that more than 90% of all plant species and over 6,000 fungal species in the Glomeromycotina, Ascomycotina, and Basidiomycotina subdivisions are involved in the formation of mycorrhizas, symbiotic associations between fungi and plant roots (Bonfante and Anca, 2009; Bonfante and Genre, 2010). Mycorrhizal fungi play a key role in plant ecosystems since they improve the nutrient status of the host plant, providing minerals and increasing water absorption from the soil, and confer resistance to stress and disease (Bonfante and Genre, 2010; Mello and Balestrini, 2018; Lanfranco and Bonfante, 2022). Moreover, they allow an efficient horizontal transfer of nutrients through the development of the so-called wood-wide web, an extensive hyphal network in the soil that connects different plants (Bonfante and Genre, 2010). At the same time, for its growth and reproduction the fungus needs the host plant, which provides carbon compounds (Mello and Balestrini, 2018). Based on the taxonomic position of plant and fungal partners and anatomical traits, mycorrhizas are commonly divided into two main categories: ectomycorrhizas and endomycorrhizas, depending on whether the fungus colonises the intercellular spaces or develops inside root cells.
A well-known example of ectomycorrhizal fungi is given by truffles (Tuber spp.), hypogeous fungi whose fruit bodies sequester the spores and develop underground, bringing benefits to the forest ecosystems and to the host plants (Trappe and Claridge, 2010; Monaco et al., 2020a). In addition to their ecological role, some truffle species (as for instance Tuber magnatum Picco, T. melanosporum Vittad., T. borchii Vittad., and T. aestivum Vittad.) are of considerable economic and commercial importance. Moreover, truffle hunting and collection practise has recently been recognised as Intangible Cultural Heritage of Humanity1 due to its naturalistic, cultural, and anthropological relevance.
Among the more than 180 Tuber species currently known, T. magnatum Picco, the so-called Italian white truffle, is the species of greatest interest and with the highest economic value (Bonito et al., 2010, 2013; Vita et al., 2015; Benucci and Bonito, 2016). Indeed, in the final stage of its complex life cycle, this ectomycorrhizal ascomycete of the Pezizales order produces edible fruiting bodies that can be considered one of the most expensive foods in the world, reaching a cost of thousands of euros per kilogram (Riccioni et al., 2016; Patel et al., 2017; Daba et al., 2019; Laruccia et al., 2020; Monaco et al., 2021a). Multiple factors contribute to the exorbitant prices of T. magnatum ascomata. On one side, their valuable culinary properties and unique organoleptic qualities ensure that market demand and truffle trade are constantly expanding. On the other side, the annual production of T. magnatum results generally insufficient to satisfy these needs (Bach et al., 2021), because of the restricted distribution range, limited seasonal availability, and difficulties in cultivation (Marjanović et al., 2015; Belfiori et al., 2020; Zambonelli et al., 2021). The indiscriminate collection, climate change, deforestation, and wildfires are added to this (Perlińska-Lenart et al., 2020; Marozzi et al., 2022), putting T. magnatum at risk of extinction. In this context, the full understanding of the factors that influence the life cycle of the precious white truffle and promote the development of fruiting bodies is of fundamental importance, also because this species is considered an indicator of a healthy environment. Among these factors, bacterial communities play a central role (Amicucci et al., 2018; Monaco et al., 2020a; Sillo et al., 2022). Accordingly, in the present mini-review, we summarised the knowledge currently available on T. magnatum associated bacterial communities, highlighting similarities and differences between samples of different origin, to address the following issues: (i) Is there a correlation between microbial taxa and truffle ground productivity?/Are there microbial taxa related to a higher truffle production? (ii) Can bacteria actually be used as markers of T. magnatum geographic origin?
Interactions between fungi and bacteria have long been studied in mycology (Waksman, 1927). Some bacterial species are beneficial to fungi, promoting the establishment of mycorrhizas and fruiting body development (Aspray et al., 2006; Frey-Klett et al., 2007, 2011), others can be responsible for fungal pathogenicity, may control sporulation (Partida-Martinez and Hertweck, 2005; Benucci and Bonito, 2016) or have a detrimental effect on mycelium development (Barbieri et al., 2005).
These interactions also concern Tuber species, which are in close contact with microorganisms throughout their life cycle. Indeed, truffles harbour and interact with complex microbial communities of bacteria, yeasts, and filamentous fungi, as well as viruses (Stielow and Menzel, 2010; Splivallo et al., 2015, 2019; Vahdatzadeh et al., 2015, 2019; Benucci and Bonito, 2016; Ratti et al., 2016). A combination of culture-dependent and independent methods and different techniques (from denaturing gradient gel electrophoresis to high-throughput sequencing) has been employed to investigate the truffle microbiota (Mello et al., 2013; Vahdatzadeh et al., 2015). Considerable differences emerged depending on the method of analysis used (Bonfante and Anca, 2009); whilst molecular techniques allow to describe more accurately the bacterial communities as a whole, culture-dependent methods have some limitations since microorganisms that can be cultivated in the laboratory represent only a small fraction of the total diversity existing in nature (Stewart, 2012; Perlińska-Lenart et al., 2020).
Bacteria can heavily colonise truffle ascocarps, both the inner tissues (gleba) and the surface (peridium), reaching a density from millions to billions of cells per gram (dry weight; Reale et al., 2009; Splivallo et al., 2015, 2019; Vahdatzadeh et al., 2015, 2019). Peridium and gleba seem to attract specific bacterial genera (Barbieri et al., 2016), which appear to be selected from the soil communities during the early stage of truffle formation. In fact, it is believed that soil bacteria colonise Tuber primordia before the differentiation of ascocarpic tissues occurs, when the primordium is directly in contact with soil. Subsequently, after the differentiation of the peridium, bacteria would be trapped in the gleba, partly protected from soil exchanges by the peridium that, on the contrary, remains in contact with ground throughout the ascocarp development (Antony-Babu et al., 2014; Monaco et al., 2020a; Vita et al., 2020).
Multiple factors can influence the composition of the truffle-associated microbial communities. Several studies highlighted variations in the microbiota structure related to the different Tuber species, life cycle stage of the fungus (e.g., mycorrhizas vs. fruiting bodies), ascocarp maturation, tissue specificity (gleba vs. peridium), storage period, collection site, harvesting season, and environmental conditions (Vahdatzadeh et al., 2015; Monaco et al., 2021b; Niimi et al., 2021a; Sillo et al., 2022). Therefore, Tuber species provide diverse microhabitats hosting complex and changeable microbial communities, involved in numerous functions. In particular, bacteria—the third partner of the symbiosis between Tuber and its host plant—seem to play a central role in the complex biological processes of signalling and nutrient exchanges involving hyphae, ectomycorrhizas, and ascocarps (Barbieri et al., 2016). In exchange for water and nutrients, they produce biostimulants (e.g., phytohormones and specific amino acids), promote the growth of mycelium and ectomycorrhiza formation (Sbrana et al., 2002; Frey-Klett et al., 2007), and participate in the development and maturation of truffle fruiting bodies (Mello et al., 2010; Antony-Babu et al., 2014; Amicucci et al., 2018). Bacteria may improve fungal nutrition (of both primordium and developed ascocarp) by enhancing the availability of some elements (N, P, micronutrients) through their nitrogen fixing activity, chemical transformation, phosphate solubilisation, and chelating compound production (Pavić et al., 2011, 2013). Moreover, some bacterial taxa could be involved in spore germination and in the opening of asci and ascospore release thanks to their cellulolytic and chitinolytic activities (Gazzanelli et al., 1999; Pavić et al., 2011); others inhibit/counteract the growth of pathogens and contaminating fungi by producing antimicrobial substances. Microorganisms are also partly responsible for truffle aroma since they synthesise sulphur volatile compounds that, besides determining the organoleptic properties of fruiting bodies, attract mammals. In this way, bacteria take indirectly part in the dissemination of truffle spores, confirming their key role in the life cycle of the fungus (Splivallo et al., 2011, 2015; Splivallo and Ebeler, 2015).
To date, there are about a dozen papers describing the microbial communities associated with the precious white truffle T. magnatum Picco. Most studies have focused on bacteria, whilst fungi have been little investigated, probably because of the difficulties related to the massive presence of Tuber DNA that can interfere and hinder the amplification, sequencing and detection of “exogenous fungi” from the gleba (Niimi et al., 2021a; Marozzi et al., 2022).
Bacterial communities associated with T. magnatum ascocarps were described for the first time by Barbieri et al. (2007), who reported important differences in the composition of the microbiota depending on the analysis method (Table 1). Indeed, whilst most of the isolated strains were fluorescent pseudomonads belonging to the γ-Proteobacteria class, 16S rDNA clone library sequencing and fluorescence in situ hybridization (FISH) showed a predominance of α-Proteobacteria, mainly represented by members of Sinorhizobium/Ensifer and Rhizobium/Agrobacterium groups, as well as Bradyrhizobium species (e.g., B. elkanii). Subsequent studies based on high-throughput sequencing techniques confirmed these results, highlighting that the bacterial communities associated with white truffle ascomata from diverse countries (Italy, Croatia, Serbia, and Hungary) were dominated by α-Proteobacteria and, in particular, by Bradyrhizobium species, regardless of their provenience and maturation degree (Monaco et al., 2021b; Niimi et al., 2021a,b; Marozzi et al., 2022). However, a different outcome emerged from the analysis of some T. magnatum collected in Tuscany (Central Italy), with gleba communities characterised by a prevalence of γ-Proteobacteria and Pedobacter, Burkholderia, Pseudomonas, and Flavobacterium as dominant taxa (Sillo et al., 2022).
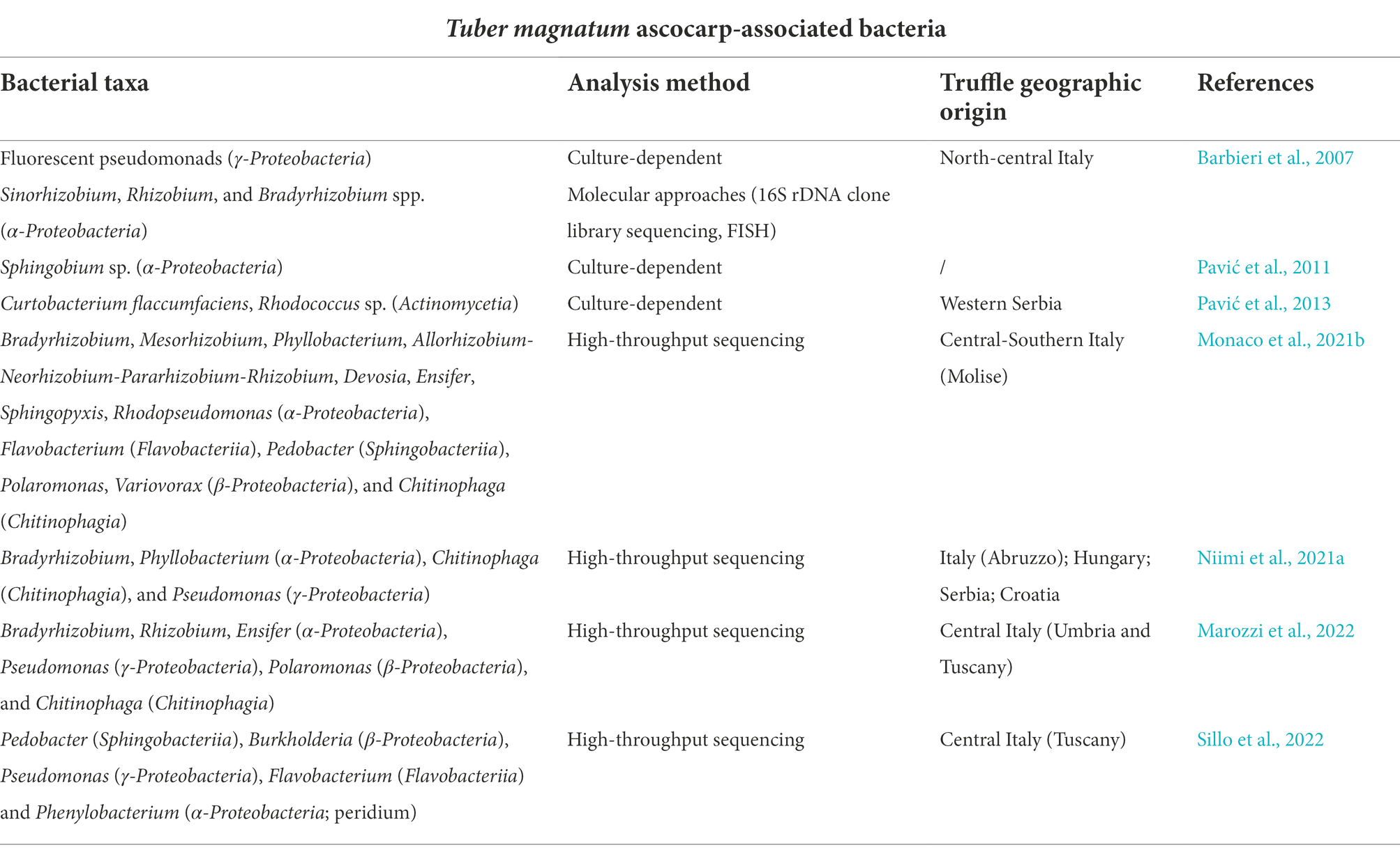
Table 1. Main bacterial taxa associated with Tuber magnatum ascocarps reported in literature.
In the last few years, the hypothesis of a potential relationship between microbial community composition and truffle orchard productivity is arousing increasing interest. Mello et al. (2010) first analysed the soil fungal and bacterial communities within a natural truffle ground of T. magnatum in Montemagno (Piedmont, Northern-Italy) in relation to its productive niches, with the aim of identifying potential “productivity markers.” They found that no specific fungal populations could be associated with productive/unproductive sites, even if Mortierella genus and Fusarium oxysporum appeared more abundant in productive soils. On the contrary, a potential link between the presence of T. magnatum ascocarps and the γ-Proteobacterium Moraxella osloensis was observed, indicating this bacterium as a promising marker of truffle productivity. A more recent study highlighted that microorganisms appear to be a better indicator of the truffle ground productive potential than chemical parameters, such as pH and phosphorus concentration. A decrease in bacterial diversity from T. magnatum unproductive to productive soils has been detected with prokaryotes belonging to Nitrososphaerales and Gemmatales orders almost exclusively present in unproductive lands (Sillo et al., 2022). However, in natural conditions, different soil microbial communities can sustain white truffle production as proved by the significative differences observed in the structure of the fungal and bacterial communities between T. magnatum productive sites (Marozzi et al., 2022). Fruiting body communities showed a lower diversity compared to the surrounding bulk soil, with a further reduction from peridium to gleba (Sillo et al., 2022). Indeed, the truffle microbial communities converge on few selected taxa (mainly represented by Mortierella for fungi, Bradyrhizobium, Rhizobium, Pseudomonas, Ensifer, Polaromonas, Pedobacter, Chitinophaga, and Phyllobacterium for bacteria), which form the “core microbiota” of T. magnatum ascocarps (Monaco et al., 2021b; Niimi et al., 2021a; Marozzi et al., 2022). Factors driving this selection have not yet been fully understood (Vahdatzadeh et al., 2015). A selective pressure on microorganisms related to specific potential functions and an evolutionary adaptation of some bacterial taxa to the genus Tuber could explain the reduction of diversity and the exclusive presence of certain microbial groups inside the fruiting bodies (Marozzi et al., 2022; Sillo et al., 2022). Therefore, the constant occurrence of defined bacterial genera in T. magnatum ascocarps with different geographical origin confirms/demonstrates their central role in truffle ecology and life cycle. On the other hand, since several biotic and abiotic factors drive the composition of microbial communities, “variable taxa” can complete the truffle microbiota contributing to determine the differences observed even at local scale (Monaco et al., 2021b). For example, the environmental conditions of the hypogeous habitat can affect (promoting or not) the bacterial colonisation of ascocarps and lead to changes in the microbiota structure, as reported by Amicucci et al. (2018) for pigment-producing bacteria that, in response to specific environmental stimuli, could release secondary metabolites (e.g., carotenoids), which seem to be responsible for the chromatic alteration (reddish patches) often observed within T. magnatum ascomata.
In the last years, the increasing market demand of the precious white truffle along with the fall in its natural production led to a growing interest in cultivation techniques (Bach et al., 2021) and encouraged truffle growers and researchers to deeper investigate factors that could affect and improve T. magnatum productivity. In this context, microbial communities deserve particular attention, since they are of undoubted importance for the ecology and life cycle of the fungus. Interestingly, a potential link between microbial taxa and truffle ground productivity was observed and specific bacteria (e.g., Moraxella osloensis) have been identified as promising T. magnatum “productivity markers” (Mello et al., 2010; Sillo et al., 2022). However, to date, studies on this subject are still very few and several aspects related to the role of bacteria in truffle biology should be properly investigated (Splivallo et al., 2019; Monaco et al., 2021b). For example, since the truffle mycelium can influence the composition and activity of soil microbial communities (Napoli et al., 2010; Mello et al., 2013; Marozzi et al., 2022), it should be clarified whether the taxa retrieved in “productive soils” actually contribute to a higher truffle production or, on the contrary, their occurrence is a consequence of T. magnatum presence. Another important open question concerns the nitrogen-fixing activity and the resulting involvement of bacteria in truffle nutrition, development, and maturation. Indeed, as previously described, potential nitrogen-fixing bacteria (mainly represented by Bradyrhizobium and Rhizobium genera) usually occur in Tuber ascomata, regardless of their maturity level (Barbieri et al., 2007). The bacterial ability to modify nutrient availability in the soil could be of fundamental importance for the fungus, especially during the early stages of ascocarp formation (Barbieri et al., 2007; Vahdatzadeh et al., 2015; Monaco et al., 2020a). In 2010, Barbieri and colleagues (Barbieri et al., 2010) evaluated the nitrogenase activity and demonstrated for the first time the expression of nifH genes from Bradyrhizobia in T. magnatum ascomata. Nevertheless, to date there is no coherent direct evidence to support that bacterial nitrogen fixation occurs during truffle development and it remains to be proved that nitrogen fixed by bacteria within truffle ascocarps is subsequently really transferred to the fungus (Vahdatzadeh et al., 2015; Marozzi et al., 2022).
Therefore, a more detailed understanding of the relationship between truffles and their environment is required. In order to define a possible link between productive seasons and soil microbial composition, it could be interesting to characterise the microbial communities within truffle sites over time. The identification of microorganisms actually able to promote T. magnatum formation may represent an important advance for the development and/or improvement of the white truffle cultivation, with significant repercussions in the field of truffle farming (Perlińska-Lenart et al., 2020; Monaco et al., 2020a; Marozzi et al., 2022). Besides being potential “productivity markers,” bacteria appear to be also a promising tool for tracing the geographic origin of truffles (Monaco et al., 2021b; Niimi et al., 2021a). Indeed, since the (economic) value of the prized T. magnatum can vary in relation to fruiting body provenience (Sillo et al., 2022), an increasing interest to define a reliable tracking system is emerging. Researchers have so far mainly focused on other aspects, including the analysis of intraspecific genetic variability (Mello et al., 2005; Monaco et al., 2021b), population genetic structure (Rubini et al., 2005; Belfiori et al., 2020), antioxidant compounds (Vita et al., 2018), transcriptomic, proteomic, and volatilomic profiles (Vita et al., 2020), as well as morphological traits, such as peridium thickness (Monaco et al., 2021a). The cost of truffle is directly influenced also by fruiting body aroma, a unique mixture of volatile organic compounds (VOCs) partially synthesised by bacteria. Interestingly, some of these chemicals are common to several Tuber species and might be of mixed truffle and microbial origin, whereas others are species-specific and could derive only from microbes (Vahdatzadeh et al., 2019). In fact, it is known that besides a core microbiota shared between diverse Tuber species, “non-fixed” bacterial taxa complete the composition of the truffle-associated microbial communities and contribute to determine the observed inter- and intra-specific differences (Monaco et al., 2021b). Hence, it is reasonable to search for potential markers of T. magnatum geographic origin among these variable bacteria. Of course, to be a good biomarker, a microbial taxon should have specific features. Firstly, it should be almost constantly present in truffles coming from a particular geographical area and absent in fruit bodies with a diverse provenience or, at least, show significantly different abundance values. A “tracer microorganism” also needs to be well represented within the examined microbial community so that it can be easily detected through the available analysis methods. In addition, the taxonomic level of investigation should be as specific as possible. Niimi et al. (2021a) identified an OTU belonging to the genus Pseudomonas as a potential marker of geographic origin of some Hungarian white truffles. Despite this encouraging result, there are still some difficulties and limitations to take into account: (1) the reduced number of studies and analysed samples that do not allow appropriate comparisons and (2) the small-scale heterogeneity in truffle microbiota composition (Monaco et al., 2021b). Moreover, since environmental factors, such as temperature, humidity, soil properties, microclimatic conditions, and snow cover (Monaco et al., 2020b), significantly affect the structure of microbial communities, even little variations can determine important changes in the truffle microbiota.
In conclusion, although further studies on the role of bacteria, microbiota composition and factors driving the establishment of a specific bacterial signature in T. magnatum are required, the analysis of microbial taxa appears to be a promising tool for the identification of Tuber productivity markers and geographic origin indicators, with a considerable impact on truffle farming, industry, and trade.
AB, AM, and PM: conceptualization. PM: writing–original draft preparation and visualisation. AB, AM, GN, and PM: writing–review and editing. AB and AM: supervision. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Amicucci, A., Barbieri, E., Sparvoli, V., Gioacchini, A. M., Calcabrini, C., Palma, F., et al. (2018). Microbial and pigment profile of the reddish patch occurring within Tuber magnatum ascomata. Fungal Biol. 122, 1134–1141. doi: 10.1016/j.funbio.2018.07.007
Antony-Babu, S., Deveau, A., Van Nostrand, J. D., Zhou, J., Le Tacon, F., Robin, C., et al. (2014). Black truffle-associated bacterial communities during the development and maturation of Tuber melanosporum ascocarps and putative functional roles. Environ. Microbiol. 16, 2831–2847. doi: 10.1111/1462-2920.12294
Aspray, T. J., Frey-Klett, P., Jones, J. E., Whipps, J. M., Garbaye, J., and Bending, G. D. (2006). Mycorrhization helper bacteria: a case of specificity for altering ectomycorrhiza architecture but not ectomycorrhiza formation. Mycorrhiza 16, 533–541. doi: 10.1007/s00572-006-0068-3
Bach, C., Beacco, P., Cammaletti, P., Babel-Chen, Z., Levesque, E., Todesco, F., et al. (2021). First production of Italian white truffle (Tuber magnatum Pico) ascocarps in an orchard outside its natural range distribution in France. Mycorrhiza 31, 383–388. doi: 10.1007/s00572-020-01013-2
Barbieri, E., Ceccaroli, P., Agostini, D., Donati Zeppa, S., Gioacchini, A. M., and Stocchi, V. (2016). “Truffle-associated bacteria: extrapolation from diversity to function”, True truffle (Tuber spp.) in the world. Soil Ecology, Systematics and Biochemistry. eds. A. Zambonelli, M. Iotti, and C. Murat (Cham: Springer), 301–317.
Barbieri, E., Ceccaroli, P., Saltarelli, R., Guidi, C., Potenza, L., Basaglia, M., et al. (2010). New evidence for nitrogen fixation within the Italian white truffle Tuber magnatum. Fungal Biol. 114, 936–942. doi: 10.1016/j.funbio.2010.09.001
Barbieri, E., Gioacchini, A. M., Zambonelli, A., Bertini, L., and Stocchi, V. (2005). Determination of microbial volatile organic compounds from Staphylococcus pasteuri against Tuber borchii using solid-phase microextraction and gas chromatography/ion trap mass spectrometry. Rapid Commun. Mass Sp. 19, 3411–3415. doi: 10.1002/rcm.2209
Barbieri, E., Guidi, C., Bertaux, J., Frey-Klett, P., Garbaye, J., Ceccaroli, P., et al. (2007). Occurrence and diversity of bacterial communities in Tuber magnatum during truffle maturation. Environ. Microbiol. 9, 2234–2246. doi: 10.1111/j.1462-2920.2007.01338.x
Belfiori, B., D’Angelo, V., Riccioni, C., Leonardi, M., Paolocci, F., Pacioni, G., et al. (2020). Genetic structure and phylogeography of Tuber magnatum populations. Diversity 12:44. doi: 10.3390/d12020044
Benucci, G. M. N., and Bonito, G. M. (2016). The truffle microbiome: species and geography effects on bacteria associated with fruiting bodies of hypogeous Pezizales. Microb. Ecol. 72, 4–8. doi: 10.1007/s00248-016-0755-3
Bonfante, P., and Anca, I. A. (2009). Plants, mycorrhizal fungi, and bacteria: a network of interactions. Annu. Rev. Microbiol. 63, 363–383. doi: 10.1146/annurev.micro.091208.073504
Bonfante, P., and Genre, A. (2010). Mechanisms underlying beneficial plant-fungus interactions in mycorrhizal symbiosis. Nat. Commun. 1:48. doi: 10.1038/ncomms1046
Bonito, G. M., Gryganskyi, A. P., Trappe, J. M., and Vilgalys, R. (2010). A global meta-analysis of Tuber ITS rDNA sequences: species diversity, host associations and long-distance dispersal. Mol. Ecol. 19, 4994–5008. doi: 10.1111/j.1365-294X.2010.04855.x
Bonito, G. M., Smith, M. E., Nowak, M., Healy, R. A., Guevara, G., Cázares, E., et al. (2013). Historical biogeography and diversification of truffles in the Tuberaceae and their newly identified Southern hemisphere sister lineage. PLoS One 8:e52765. doi: 10.1371/journal.pone.0052765
Daba, G. M., Elkhateeb, W. A., Wen, T. C., and Thomas, P. W. (2019). “The continuous story of truffle-plant interaction,” in Microbiome in Plant Health and Disease. Challenges and Opportunities. eds. V. Kumar, R. Prasad, M. Kumar, and D. K. Choudhary (Singapore: Springer), 375–383.
Frey-Klett, P., Burlinson, P., Deveau, A., Barret, M., Tarkka, M., and Sarniguet, A. (2011). Bacterial-fungal interactions: hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. R. 75, 583–609. doi: 10.1128/mmbr.00020-11
Frey-Klett, P., Garbaye, J., and Tarkka, M. (2007). The mycorrhiza helper bacteria revisited. New Phytol. 176, 22–36. doi: 10.1111/j.1469-8137.2007.02191.x
Gazzanelli, G., Malatesta, M., Pianetti, A., Baffone, W., Stocchi, V., and Citterio, B. (1999). Bacteria associated to fruit bodies of the ecto-mycorrhizal fungus Tuber borchii Vittad. Symbiosis 26, 211–222.
Lanfranco, L., and Bonfante, P. (2022). The need for phosphate: at the root of the mycorrhizal symbiosis. Sci. Bull. 67, 459–460. doi: 10.1016/j.scib.2021.11.018
Laruccia, N., Marletto, V., Leonardi, P., Puliga, F., and Zambonelli, A. (2020). Map of suitability for the spontaneous growth of Tuber magnatum in Emilia-Romagna (Italy). Ital. J. Mycol. 49, 38–53. doi: 10.6092/issn.2531-7342/10832
Marjanović, Ž., Glišić, A., Mutavdžić, D., Saljnikov, E., and Bragato, G. (2015). Ecosystems supporting Tuber magnatum Pico production in Serbia experience specific soil environment seasonality that may facilitate truffle lifecycle completion. Appl. Soil Ecol. 95, 179–190. doi: 10.1016/j.apsoil.2015.05.007
Marozzi, G., Benucci, G. M. N., Turchetti, B., Massaccesi, L., Baciarelli Falini, L., Bonito, G., et al. (2022). Fungal and bacterial diversity in the Tuber magnatum ecosystem and microbiome. Microb. Ecol. doi: 10.1007/s00248-021-01950-1
Mello, A., and Balestrini, R. (2018). Recent insights on biological and ecological aspects of ectomycorrhizal fungi and their interactions. Front. Microbiol. 9:216. doi: 10.3389/fmicb.2018.00216
Mello, A., Ding, G. C., Piceno, Y. M., Napoli, C., Tom, L. M., DeSantis, T. Z., et al. (2013). Truffle brûlés have an impact on the diversity of soil bacterial communities. PLoS One 8:e61945. doi: 10.1371/journal.pone.0061945
Mello, A., Miozzi, L., Vizzini, A., Napoli, C., Kowalchuk, G., and Bonfante, P. (2010). Bacterial and fungal communities associated with Tuber magnatum-productive niches. Plant Biosyst. 144, 323–332. doi: 10.1080/11263500903374724
Mello, A., Murat, C., Vizzini, A., Gavazza, V., and Bonfante, P. (2005). Tuber magnatum Pico, a species of limited geographical distribution: its genetic diversity inside and outside a truffle ground. Environ. Microbiol. 7, 55–65. doi: 10.1111/j.1462-2920.2004.00678.x
Monaco, P., Bucci, A., Naclerio, G., and Mello, A. (2021b). Heterogeneity of the white truffle Tuber magnatum in a limited geographic area of Central-Southern Italy. Environ. Microbiol. Rep. 13, 591–599. doi: 10.1111/1758-2229.12956
Monaco, P., Divino, F., Naclerio, G., and Bucci, A. (2020b). Microbial community analysis with a specific statistical approach after a record breaking snowfall in Southern Italy. Ann. Microbiol. 70:63. doi: 10.1186/s13213-020-01604-6
Monaco, P., Naclerio, G., Bucci, A., and Mello, A. (2021a). Determination of the peridium thickness of Tuber magnatum ascomata from Molise region. Ital. J. Mycol. 50, 92–98. doi: 10.6092/issn.2531-7342/13052
Monaco, P., Toumi, M., Sferra, G., Tóth, E., Naclerio, G., and Bucci, A. (2020a). The bacterial communities of Tuber aestivum: preliminary investigations in Molise region, Southern Italy. Ann. Microbiol. 70:37. doi: 10.1186/s13213-020-01586-5
Napoli, C., Mello, A., Borra, A., Vizzini, A., Sourzat, P., and Bonfante, P. (2010). Tuber melanosporum, when dominant, affects fungal dynamics in truffle grounds. New Phytol. 185, 237–247. doi: 10.1111/j.1469-8137.2009.03053.x
Niimi, J., Deveau, A., and Splivallo, R. (2021a). Geographical-based variations in white truffle Tuber magnatum aroma is explained by quantitative differences in key volatile compounds. New Phytol. 230, 1623–1638. doi: 10.1111/nph.17259
Niimi, J., Deveau, A., and Splivallo, R. (2021b). Aroma and bacterial communities dramatically change with storage of fresh white truffle Tuber magnatum. LWT Food Sci. Technol. 151:112125. doi: 10.1016/j.lwt.2021.112125
Partida-Martinez, L. P., and Hertweck, C. (2005). Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 437, 884–888. doi: 10.1038/nature03997
Patel, S., Rauf, A., Khan, H., Khalid, S., and Mubarak, M. S. (2017). Potential health benefits of natural products derived from truffles: a review. Trends Food Sci. Technol. 70, 1–8. doi: 10.1016/j.tifs.2017.09.009
Pavić, A., Stanković, S., and Marjanović, Ž. (2011). Biochemical characterization of a sphingomonad isolate from the ascocarp of white truffle (Tuber magnatum Pico). Arch. Biol. Sci. 63, 697–704. doi: 10.2298/ABS1103697P
Pavić, A., Stanković, S., Saljnikov, E., Krüger, D., Buscot, F., Tarkka, M., et al. (2013). Actinobacteria may influence white truffle (Tuber magnatum Pico) nutrition, ascocarp degradation and interactions with other soil fungi. Fungal Ecol. 6, 527–538. doi: 10.1016/j.funeco.2013.05.006
Perlińska-Lenart, U., Piłsyk, S., Gryz, E., Turło, J., Hilszczańska, D., and Kruszewska, J. S. (2020). Identification of bacteria and fungi inhabiting fruiting bodies of Burgundy truffle (Tuber aestivum Vittad.). Arch. Microbiol. 202, 2727–2738. doi: 10.1007/s00203-020-02002-x
Ratti, C., Iotti, M., Zambonelli, A., and Terlizzi, F. (2016). “Mycoviruses infecting true truffles,” in True Truffle (Tuber spp.) in the World. Soil Ecology, Systematics and Biochemistry. eds. A. Zambonelli, M. Iotti, and C. Murat (Cham: Springer), 333–349.
Reale, A., Sorrentino, E., Iacumin, L., Tremonte, P., Manzano, M., Maiuro, L., et al. (2009). Irradiation treatments to improve the shelf life of fresh black truffles (truffles preservation by gamma-rays). J. Food Sci. 74, M196–M200. doi: 10.1111/j.1750-3841.2009.01142.x
Riccioni, C., Rubini, A., Belfiori, B., Gregori, G., and Paolocci, F. (2016). “Tuber magnatum: the special one. What makes it so different from the other Tuber spp.?” in True Truffle (Tuber spp.) in the World. Soil Ecology, Systematics and Biochemistry. eds. A. Zambonelli, M. Iotti, and C. Murat (Cham: Springer), 87–103.
Rubini, A., Paolocci, F., Riccioni, C., Vendramin, G. G., and Arcioni, S. (2005). Genetic and phylogeographic structures of the symbiotic fungus Tuber magnatum. Appl. Environ. Microbiol. 71, 6584–6589. doi: 10.1128/aem.71.11.6584-6589.2005
Sbrana, C., Agnolucci, M., Bedini, S., Lepera, A., Toffanin, A., Giovannetti, M., et al. (2002). Diversity of culturable bacterial populations associated to Tuber borchii ectomycorrhizas and their activity on T. borchii mycelial growth. FEMS Microbiol. Lett. 211, 195–201. doi: 10.1111/j.1574-6968.2002.tb11224.x
Sillo, F., Vergine, M., Luvisi, A., Calvo, A., Petruzzelli, G., Balestrini, R., et al. (2022). Bacterial communities in the fruiting bodies and background soils of the white truffle Tuber magnatum. Front. Microbiol. 13:864434. doi: 10.3389/fmicb.2022.864434
Splivallo, R., Deveau, A., Valdez, N., Kirchhoff, N., Frey-Klett, P., and Karlovsky, P. (2015). Bacteria associated with truffle-fruiting bodies contribute to truffle aroma. Environ. Microbiol. 17, 2647–2660. doi: 10.1111/1462-2920.12521
Splivallo, R., and Ebeler, S. E. (2015). Sulfur volatiles of microbial origin are key contributors to human-sensed truffle aroma. Appl. Microbiol. Biotechnol. 99, 2583–2592. doi: 10.1007/s00253-014-6360-9
Splivallo, R., Ottonello, S., Mello, A., and Karlovsky, P. (2011). Truffle volatiles: from chemical ecology to aroma biosynthesis. New Phytol. 189, 688–699. doi: 10.1111/j.1469-8137.2010.03523.x
Splivallo, R., Vahdatzadeh, M., Maciá-Vicente, J. G., Molinier, V., Peter, M., Egli, S., et al. (2019). Orchard conditions and fruiting body characteristics drive the microbiome of the black truffle Tuber aestivum. Front. Microbiol. 10:1437. doi: 10.3389/fmicb.2019.01437
Stewart, E. J. (2012). Growing unculturable bacteria. J. Bacteriol. 194, 4151–4160. doi: 10.1128/JB.00345-12
Stielow, B., and Menzel, W. (2010). Complete nucleotide sequence of TaV1, a novel totivirus isolated from a black truffle ascocarp (Tuber aestivum Vittad.). Arch. Virol. 155, 2075–2078. doi: 10.1007/s00705-010-0824-8
Trappe, J. M., and Claridge, A. W. (2010). The hidden life of truffles. Sci. Am. 302, 78–84. doi: 10.1038/scientificamerican0410-78
Vahdatzadeh, M., Deveau, A., and Splivallo, R. (2015). The role of the microbiome of truffles in aroma formation: a meta-analysis approach. Appl. Environ. Microbiol. 81, 6946–6952. doi: 10.1128/AEM.01098-15
Vahdatzadeh, M., Deveau, A., and Splivallo, R. (2019). Are bacteria responsible for aroma deterioration upon storage of the black truffle Tuber aestivum: a microbiome and volatilome study. Food Microbiol. 84:103251. doi: 10.1016/j.fm.2019.103251
Vita, F., Franchina, F. A., Taiti, C., Locato, V., Pennazza, G., Santonico, M., et al. (2018). Environmental conditions influence the biochemical properties of the fruiting bodies of Tuber magnatum Pico. Sci. Rep. 8:7243. doi: 10.1038/s41598-018-25520-7
Vita, F., Giuntoli, B., Bertolini, E., Taiti, C., Marone, E., D’Ambrosio, C., et al. (2020). Tuberomics: a molecular profiling for the adaption of edible fungi (Tuber magnatum Pico) to different natural environments. BMC Genomics 21:90. doi: 10.1186/s12864-020-6522-3
Vita, F., Taiti, C., Pompeiano, A., Bazihizina, N., Lucarotti, V., Mancuso, S., et al. (2015). Volatile organic compounds in truffle (Tuber magnatum Pico): comparison of samples from different regions of Italy and from different seasons. Sci. Rep. 5:12629. doi: 10.1038/srep12629
Zambonelli, A., Iotti, M., Puliga, F., and Hall, I. R. (2021). “Enhancing white truffle (Tuber magnatum Picco and Tuber borchii Vittad.) cultivation through biotechnology innovation,” in Advances in Plant Breeding Strategies: Vegetable Crops. eds. J. M. Al-Khayri, S. M. Jain, and D. V. Johnson (Cham: Springer), 505–532.
Keywords: ectomycorrhizal fungi, truffle, Tuber magnatum, microbial communities, bacteria
Citation: Monaco P, Naclerio G, Mello A and Bucci A (2022) Role and potentialities of bacteria associated with Tuber magnatum: A mini-review. Front. Microbiol. 13:1017089. doi: 10.3389/fmicb.2022.1017089
Edited by:
Loredana Canfora, Council for Agricultural and Economics Research (CREA), ItalyReviewed by:
Alessandra Zambonelli, University of Bologna, ItalyCopyright © 2022 Monaco, Naclerio, Mello and Bucci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pamela Monaco, cGFtZWxhLm1vbmFjb0B1bmltb2wuaXQ=
†These authors have contributed equally to this work and share senior authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.