
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 28 September 2022
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 13 - 2022 | https://doi.org/10.3389/fmicb.2022.1000787
This article is part of the Research TopicWomen in Antimicrobials, Resistance and Chemotherapy: 2022View all 6 articles
 Javier E. Cañada-García1
Javier E. Cañada-García1 Natalin Grippo1,2Eva Ramírez de Arellano1Verónica Bautista1Noelia Lara1Ana María Navarro1Teresa Cabezas3Nora Mariela Martínez-Ramírez4Silvia García-Cobos1Jorge Calvo5,6
Natalin Grippo1,2Eva Ramírez de Arellano1Verónica Bautista1Noelia Lara1Ana María Navarro1Teresa Cabezas3Nora Mariela Martínez-Ramírez4Silvia García-Cobos1Jorge Calvo5,6 Emilia Cercenado7,8
Emilia Cercenado7,8 Belén Aracil1,5
Belén Aracil1,5 María Pérez-Vázquez1,5*†
María Pérez-Vázquez1,5*† Jesús Oteo-Iglesias1,5†the Spanish IMP Study Group
Jesús Oteo-Iglesias1,5†the Spanish IMP Study GroupObjectives: Little is known about IMP-producing Enterobacterales (IMP-Ent) in Europe. We analyzed at genomic and phenotypic level IMP-Ent isolates circulating in Spain in a 9-year period.
Materials and methods: IMP-Ent isolates submitted to our reference laboratory were included. Antibiotic susceptibility was performed using microdilution method (EUCAST), and IMP-carbapenemase activity was measured with carbapenemase inhibitors, the β-CARBA method, the modified Hodge test (MHT), and the modified carbapenemase inhibition method (mCIM). All isolates collected were sequenced for high-resolution single-nucleotide polymorphism (SNP) typing, core genome multilocus sequence typing (cgMLST), and resistome analysis.
Results: Fifty IMP-Ent isolates, collected from 19 hospitals in 13 Spanish provinces, were detected: Klebsiella pneumoniae (IMP-Kpn) (24; 48%), Enterobacter roggenkampii (13; 26%), Enterobacter hormaechei (8, 16%), Klebsiella oxytoca (two; 4%), Enterobacter asburiae (one, 2%), Serratia marcescens (one; 2%) and Escherichia coli (one; 2%). All isolates were positive by the MHT and β-CARBA tests; 48 (96%) were mCIM positive; 12 (24%) and 26 (52%) displayed positive inhibition with dipicolinic (meropenem) and EDTA (ertapenem), respectively. Five IMP-carbapenemase types were identified: IMP-8 (22; 44%), IMP-22 (17; 34%), IMP-13 (7; 14%), IMP-28 (two; 4%), and IMP-15 (two; 4%), predominating IMP-8 in K. pneumoniae and IMP-22 in E. roggenkampii. IMP-28 was exclusively identified in K. oxytoca and IMP-15 in E. hormaechei. Predominant STs were ST405 (29.2%), ST15 (25%) and ST464 (20.8%) in IMP-Kpn; ST96 (100%) in E. roggenkampii and ST182 (62.5%) in E. hormachei. Colistin and amikacin were the most active non-carbapenem antibiotics against IMP-Ent.
Conclusion: IMP-Ent isolates remain infrequent in Spain, although in recent years have been circulating causing nosocomial outbreaks, being IMP-8-producing K. pneumoniae and IMP-22-producing E. roggenkampii the most frequently detected in this study. Inhibition with EDTA or dipicolinic acid presented false negative results in some IMP-producing strains. Active microbiological and molecular surveillance is essential for a better comprehension and control of IMP-Ent dissemination.
Carbapenemase-producing Enterobacterales (CPE) have emerged as a major global health threat, mainly due to spread of OXA-48, KPC and NDM carbapenemases (Tzouvelekis et al., 2012). In contrast, imipenemase (IMP)-type carbapenemases, although common in Pseudomonas aeruginosa, are infrequently detected and thus poorly understood in Enterobacterales species (Zhao and Hu, 2011). Worldwide, Asia and Oceania display the highest prevalence of IMP-producing carbapenem-resistant Enterobacterales (IMP-Ent; Zhao and Hu, 2011; Matsumura et al., 2017; Han et al., 2020); which are scarce in Europe. There is a lack of information on the molecular characterization by whole genome sequencing (WGS) of a large collection of IMP-producing Enterobacterales. The European Survey on Carbapenemase-Producing Enterobacteriaceae (EuSCAPE) reported only three IMP-producing Klebsiella pneumoniae isolates out of 684 (0.4%) carbapenemase-encoding organisms (David et al., 2019).
Moreover, many rapid commercially available tests for carbapenemase identification (e.g., immunochromatography, PCR) do not detect blaIMP genes, or include only specific types (e.g., blaIMP-1) that are not necessarily the most prevalent. Further, due to their low frequency, a detailed understanding of IMP-Ent behaviour in phenotypic tests for carbapenemase characterization, such as the modified Hodge test (MHT) test, colorimetric assays, the carbapenemase inhibition method, and use of specific inhibitors, is lacking. Thus, there is substantial risk for under-detection of IMP-Ent.
The aim of this multicenter nation-wide study was to provide knowledge about the IMP-Ent isolates, characterizing their molecular epidemiology and resistance mechanisms (including WGS analysis) and evaluating different phenotypic methods for carbapenemases detection in these bacteria in Spain.
This study was performed by the unrestricted and non-mandatory national Spanish Antibiotic Resistance Surveillance Programme, operated by our official public health Institute (Instituto de Salud Carlos III). We included IMP-Ent isolates submitted to our reference lab from January 2012 to December 2021, analyzing only the first isolate obtained from each patient. Initial assays were performed at each participating hospital using standard microbiological methods. CPE isolates were identified according to the European Committee on Antimicrobial Susceptibility Testing (EUCAST)-established meropenem cut-off values for CPE (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2017) and phenotypic confirmation of carbapenemase production was verified using at least one EUCAST-recommended method (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2017).
Antibiotic susceptibility testing was performed by micro-dilution (DKMGN panel, Thermo Fisher Scientific, Waltham, MA, United States) and interpreted according to EUCAST breakpoints (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2017, 2022). Carbapenemase activity of IMP-Ent was tested using meropenem plus dipicolinic acid, phenyl-boronic acid, and cloxacillin inhibitors (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2017) (Rosco Diagnostica A/S, Taastrup, Denmark), and ertapenem plus 10 μl of EDTA inhibitor; the β-CARBA colorimetric detection method (Bio-Rad, Marnes-la-Coquette, France); the MHT, with meropenem discs containing 600 mg cloxacillin; and the modified carbapenemase inhibition method (mCIM) (Pierce et al., 2017).
Genomic library preparation and sequence analysis were conducted as described (Pérez-Vázquez et al., 2019). Raw sequence data were submitted to the European Nucleotide Archive (PRJEB54568). Quality of short reads was assessed using FASTQC, and assembly into contigs was performed with Unicycler 0.4.8 (Wick et al., 2017). Quality of the assembly was assessed with QUAST1. Prokka v1.14-beta (Seemann, 2014) was used for automatic de novo assembly annotation. Mash v2.0 (Ondov et al., 2016) was used to analyze the similarity of each sequence to the genomes in RefSeq bacterial database;2 finally, we report the species and subspecies if assigned as the top matched with the smallest distance. For all top matched sequences with Mash, ANI distances against reference genomes of the main subspecies were calculated with OthoANIu toll (Yoon et al., 2017); the identification was considered the one of the reference in the pair with highest ANI value applying the threshold described by Sutton et al. (2018). Mash distances and ANI results are reported in Supplementary Table S1. All the Klebsiella species and subspecies were confirmed with Kleborate.3
Prokka v.1.14 (Seemann, 2014) was used to annotate de novo assemblies, these annotated assemblies were used as input for Roary v3.13.0 (Page et al., 2015). For Klebsiella pneumoniae and Enterobacter cloacae complex alignments of 3.589 and 686 core genes (present in ≥ 99% of isolates) comprising 3.533.275 bp and 672.747 bp were generated, respectively. Variable positions of the alignment were extracted with Snp sites (Page et al., 2016) in both species (64.568 and 67.619 SNPs, respectively), finally pairwise distances were calculated with MegaX v10.0.5 (Kumar et al., 2018).
Sequence types (STs) were calculated according to multilocus sequence typing (MLST) schemes of the Institut Pasteur for K. pneumoniae4 and E. cloacae complex using Ariba v2.6.2 (Hunt et al., 2017). Core-genome MLST (cgMLST) consisting of 2.538 targets for K. pneumoniae a provided by SeqSphere+3.5.0 (Ridom, Germany), was performed. For global E. cloacae complex, Enterobacter hormaechei and Enterobacter roggenkampii trees ad hoc schemes were created using the MLST+ target definer with the default parameters and a reference sequence: Enterobacter cloacae subsp. cloacae ATCC 13047 (accession no NC_014121), E. hormaechei (accession no NC_021046) and E. roggenkampii (accession no NZ_CP022148.1). A total of 38, 24 and 26 NCBI RefSeq genomes for global E. cloacae complex, E. hormaechei and E. roggenkampii, respectively, were used as query genomes to validate in a pairwise comparison using BLAST. Genes selected in the cgMLST were common in all query genomes and they presented a percentage of identity higher that 90%. The final cgMLST schemes consisted in 631 targets genes and 4.322 accessory genes for E. cloacae complex, 2.123 and 1.798 for E. hormaechei and 2.466 and 1.951 for E. roggenkampii. The average percentage of good targets included in the cgMLST was 99.3, 98.9 and 99.7% for E. cloacae complex, E. hormaechei and E. roggenkampii, respectively.
Antibiotic resistance genes were analyzed by SRST2 (Inouye et al., 2014) using the ARG-ANNOT database (Gupta et al., 2014) and ResFinder (CGE server)5, with ID thresholds of 100% for β-lactamase variants (except for chromosomal AmpC genes), and 98% for the other resistance genes. The K-locus and virulence genes were characterized in IMP-producing K. pneumoniae isolates using Kleborate.6 The presence of ybt, clb, and iuc was used to assign a virulence score, as described (Lam et al., 2021).
During the study period, 50 non-duplicated IMP-Ent isolates from 19 Spanish hospitals in 13 provinces were submitted to the reference laboratory (Table 1), which corresponded to 0.4% of the 12,038 carbapenemase-producing Enterobacterales isolates detected in this laboratory. Most of IMP-Ent isolates were from men (64%) and people over 65 years old (52%). No temporal significant trend was detected in the proportion of IMP-Ent among total CPE identified by our laboratory by year, which ranged between 2.1% in 2012 and 0.2% in 2015.
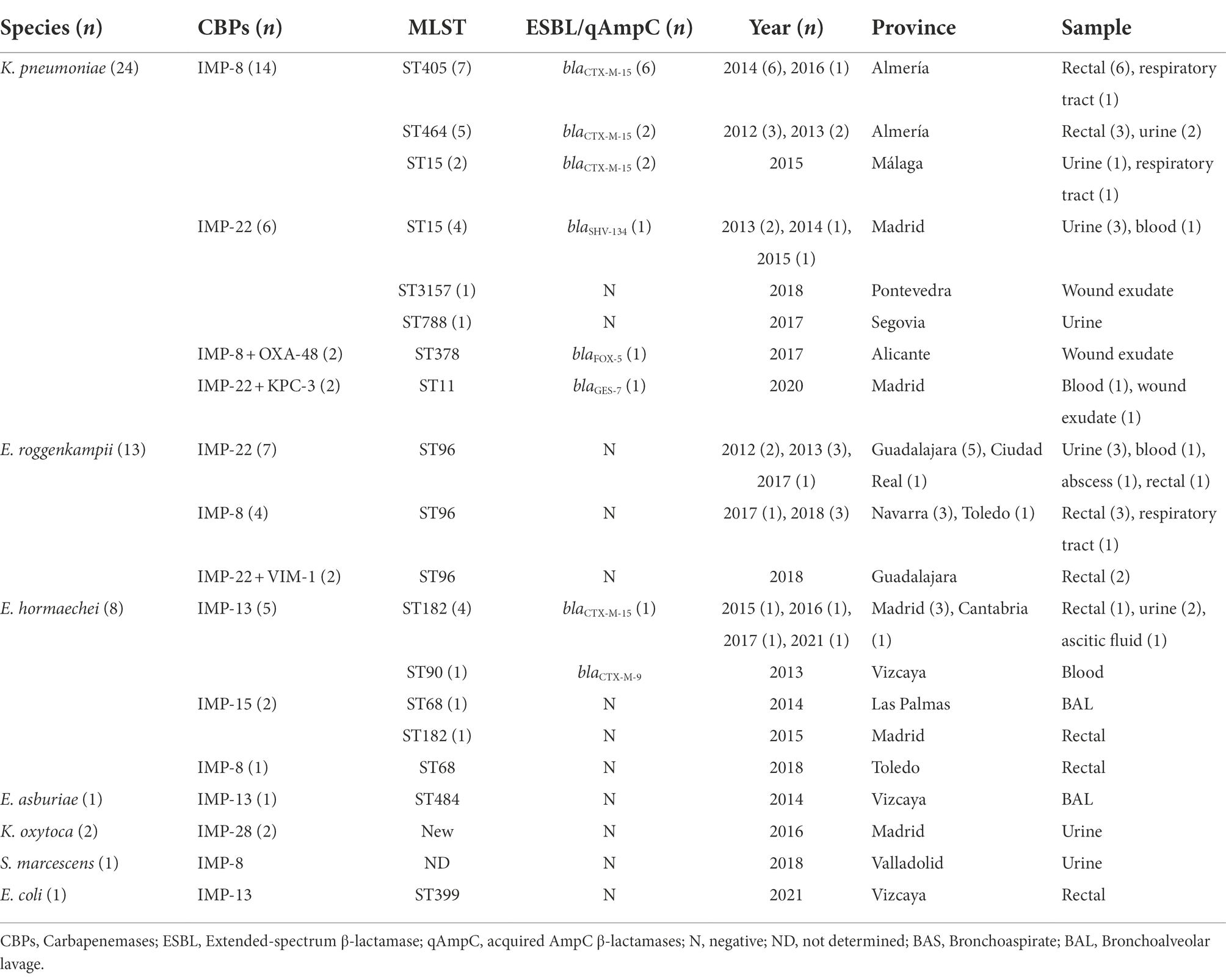
Table 1. Main microbiological and epidemiological features of the 50 IMP-producing Enterobacterales isolates included in this study.
Of the 50 isolates, 32 (64%) produced clinical infections, including 16 (32%) urinary tract infections, 5 (10%) respiratory tract infections, 4 (8%) bacteraemia cases, and 7 (14%) other infections; the remaining 18 (36%) were from rectal samples.
The IMP-producing isolates included 24 (48%) strains of K. pneumoniae “sensu stricto”; 22 (44%) E. cloacae complex belonging to E. roggenkampii (13; 26%), E. hormaechei (8, 16%, five belonged to E. hormaechei subsp xiangfangensis, two to E. hormaechei subsp. steigerwaltii and one to E. hormaechei subsp. oharae), and Enterobacter asburiae (one, 2%) species; two (4%) Klebsiella oxytoca, one (2%) Serratia marcescens, and one (2%) Escherichia coli (Table 1; Supplementary Table S1). Outside of Asia and Oceania, little is known about IMP-Ent, and most isolates are from single cases or small outbreaks (Zhao and Hu, 2011; Matsumura et al., 2017; García-Castillo et al., 2018). A recently published collection of 935 carbapenem-resistant Enterobacterales from China contained 16 IMP-producing isolates (Han et al., 2020). Thus, our collection of IMP-Ent from Spain can provide valuable insights into the biology of European isolates. In addition, there are very few descriptions in the literature of carbapenemase-producing E. roggenkampii (Mateos et al., 2021; Umeda et al., 2021), and only one associated with IMP carbapenemases (Umeda et al., 2021).
Characterization of β-lactamases genes by DNA sequencing showed five blaIMP genes encoding IMP-8 (22, 44%), IMP-22 (17, 34%), IMP-13 (7, 14%), IMP-28 (2, 4%), and IMP-15 (2, 4%) (Table 1). IMP-8 predominated in K. pneumoniae (16/22; 72.7%), whereas IMP-22 was most prevalent in E. roggenkampii (9/13; 69.2%) isolates. IMP-28 was only found in K. oxytoca (Table 1). An additional carbapenemase gene was observed in six isolates: two K. pneumoniae with IMP-8 plus OXA-48 of ST378, two K. pneumoniae with IMP-22 plus KPC-3 of ST11, and two E. roggenkampii with IMP-22 plus VIM-1 (Table 1).
IMP-8 is the type most frequently described in Enterobacterales worldwide, mainly in Asian countries such as Taiwan, where several outbreaks have been reported (Wang et al., 2015; Matsumura et al., 2017). However, nosocomial outbreaks of IMP-8-producing Enterobacterales have also occurred in other geographical regions, including those mediated by K. oxytoca in Spain (Vergara-López et al., 2013) and E. coli in Argentina (Elena et al., 2018); a specific search in PubMed did not provide any previous description of IMP-8 in S. marcescens. Conversely, IMP-22 has rarely been described, occurring mainly in Spain in K. pneumoniae and E. coli isolates (Miró et al., 2013; Ortega et al., 2016). Interhospital and community dissemination of IMP-22-producing K. pneumoniae has been described recently in the North of Portugal (Gonçalves et al., 2021). IMP-28 was first described in a K. oxytoca isolate in 2009 (Pérez-Llarena et al., 2012), from the same hospital in Madrid where the IMP-28-producing isolates of this study were collected 7 years later (2016). To the best of our knowledge, no previous cases of IMP-13 in E. coli have been reported; being an enzyme predominantly described in P. aeruginosa (Santella et al., 2010).
Frequencies of carbapenem susceptibility in K. pneumoniae and E. cloacae complex isolates were 66.6% (16/24) and 22.7% (5/22) for imipenem, respectively, and 62.5% (15/24) and 31.8% (7/22) for meropenem, respectively, with all susceptible isolates showing minimum inhibitory concentrations (MICs) of 1–2 mg/l (Table 2). Meropenem and imipenem MICs in non-susceptible isolates ranged 4–16 mg/l, being 4 mg/l the most frequent MIC (Table 3).
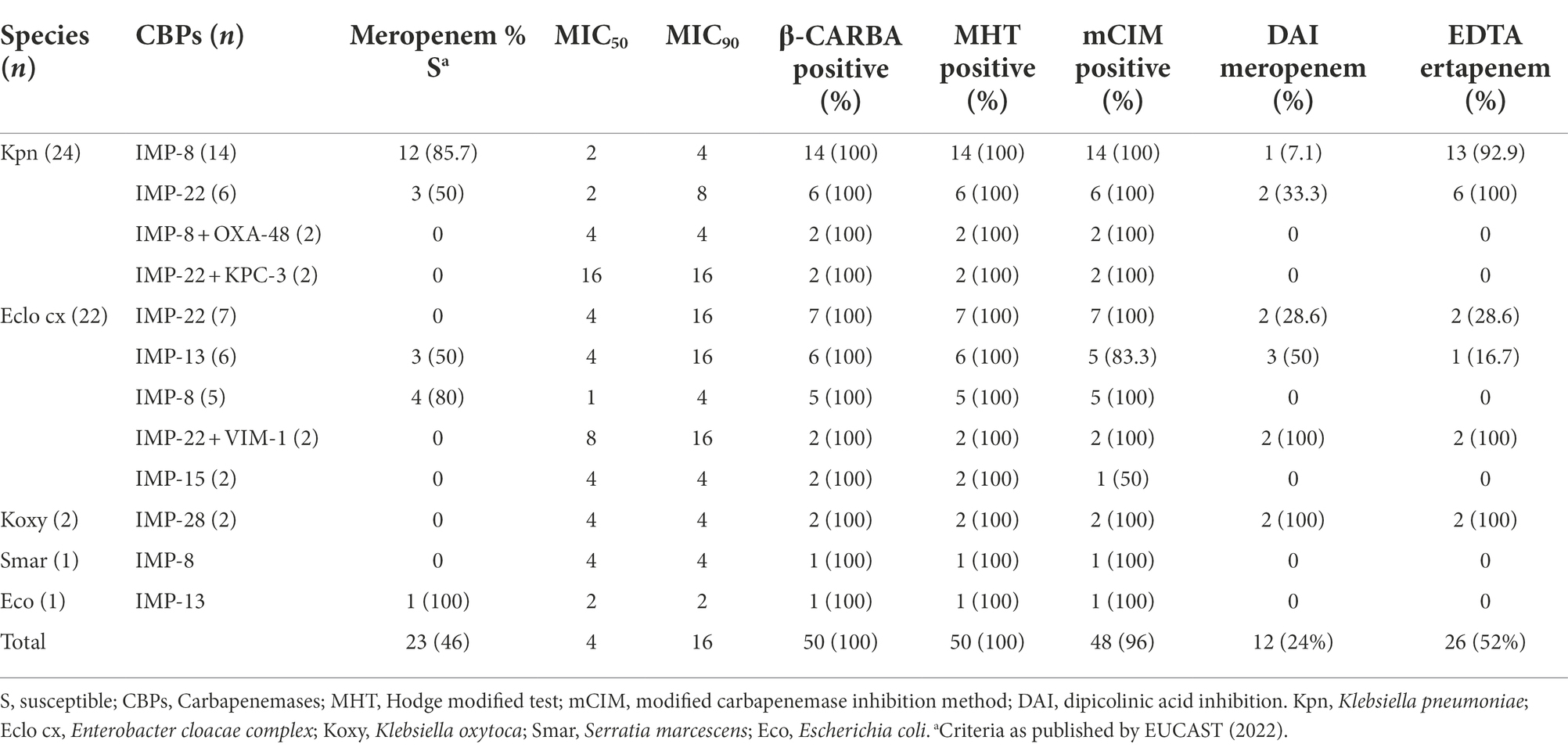
Table 2. Phenotypic features for detection of IMP production in IMP-producing Enterobacterales isolates.
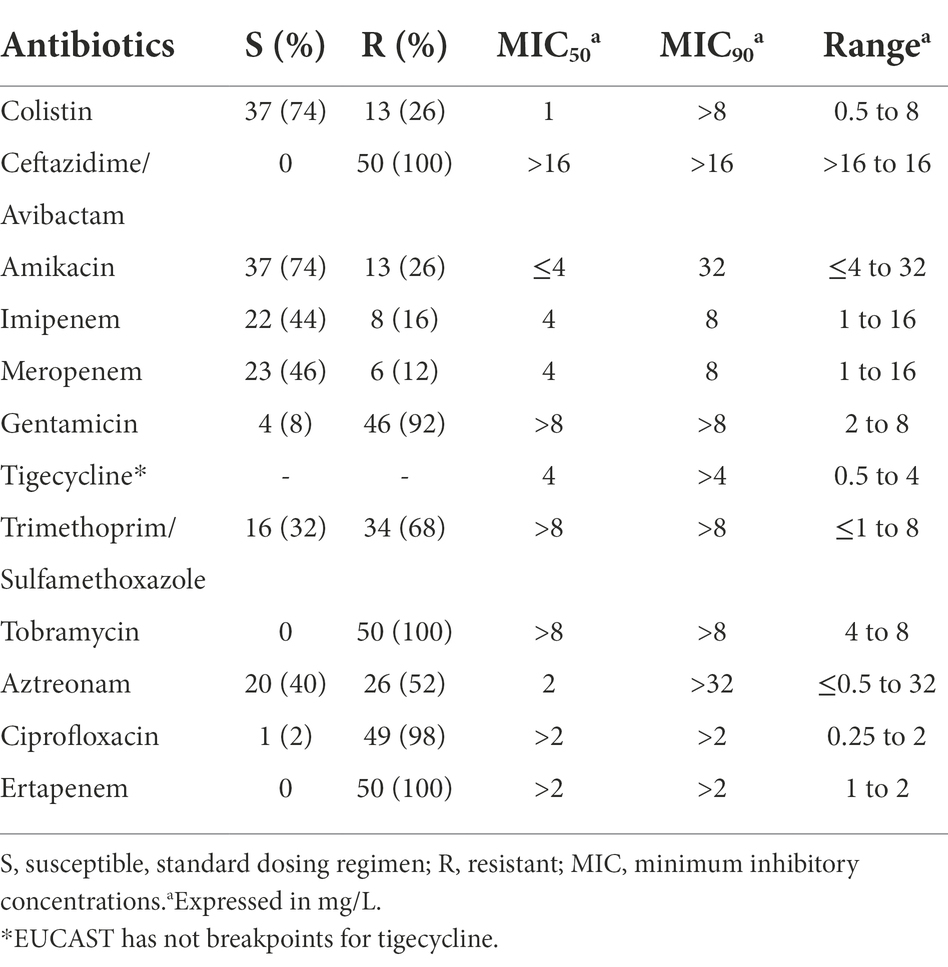
Table 3. Antibiotic susceptibility of 50 IMP-producing Enterobacterales as determined by the microdilution method according to EUCAST clinical breakpoints.
No IMP-Ent isolates were susceptible to ertapenem, more of them (62%) had MICs >2 mg/l. IMP-producing K. pneumoniae isolates showed high meropenem susceptibility rates, 75% (15 of the 20 isolates that produced only IMP carbapenemase) compared to other metallo-β-lactamases producers, such as NDM and VIM producers (Lutgring et al., 2020; Vázquez-Ucha et al., 2021). However, all isolates had MICs for meropenem within screening cut-off values recommended by EUCAST to suspect CPE (European Committee on Antimicrobial Susceptibility Testing [EUCAST], 2017), a key fact to consider for avoiding under-detection of IMP-producing K. pneumoniae.
According to carbapenemase type, four of the 20 (20%) isolates that produced only IMP-8 (14 K. pneumoniae, 4 E. roggenkampii, one E. hormaechei, one S. marcescens) were non-susceptible to meropenem; while 10 of the 13 (76.9%) isolates that produced only IMP-22 (7 E. roggenkampii and 6 K. pneumoniae) were non-susceptible to this antibiotic.
Table 3 shows antibiotic susceptibility of the 50 IMP-producing Enterobacterales. Regarding non-carbapenem antibiotics, the most active antibiotics tested were colistin (74% of susceptibility), amikacin (74%), aztreonam (40%), and trimethoprim/sulfamethoxazole (32%). Eleven of the 13 colistin-resistant isolates were E. roggenkampii. In a recent study with Enterobacterales from companion animals in Japan (Sato et al., 2022), many of them belonging to lineages with human clinical isolates, resistance to colistin was higher in Enterobacter spp. (20.3%) than in K. pneumoniae (1.2%) or E. coli (0%).
We performed different phenotypic tests for carbapenemase characterization and found all isolates were positive by the MHT and β-CARBA methods, regardless of species (Table 2). The mCIM was positive in 48 (96%) isolates, with a mean inhibition zone of 7.3 mm (range: 6–18 mm); two E. hormaechei isolates (IMP-13 and IMP-15) displaying intermediate results (16–18 mm) (Table 2). However, dipicolinic acid inhibition results yielded poor with only 12 (24%) positive isolates: four E. roggenkampii, three E. hormaechei, three K. pneumoniae and two K. oxytoca. The mean difference between inhibition zones observed with meropenem tablets (10 μg) and those obtained with meropenem plus dipicolinic acid tablets was 2.1 mm (range: 0–7 mm). This poor inhibition by dipicolinic acid could result from interference by chromosomal AmpC in E. cloacae complex isolates and, mainly, due to the slight hydrolytic activity of IMP carbapenemases against meropenem in K. pneumoniae (Doyle et al., 2012). However, the inhibition method using ertapenem plus EDTA showed that in 26 isolates (52%; 19 K. pneumoniae, 4 E. roggenkampii, 2 K. oxytoca, and one E. hormaechei), the activity of ertapenem was recovering with EDTA (increase of the inhibition zone ≥5 mm). Although not recommended by EUCAST guidelines, the use of ertapenem plus EDTA seems to be more sensitive for detection of IMP-Ent, mainly in K. pneumoniae isolates (19/24; 79.2%), than meropenem plus dipicolinic.
By WGS, we determined the profile of genes conferring resistance to antimicrobial agents (resistome) (Supplementary Table S1). An average of 11.2 antimicrobial resistance genes were detected in IMP-Ent isolates (range: 4–19), with small differences observed between E. cloacae complex (10.3; range: 7–18) and K. pneumoniae (12.8; range: 7–19) (Supplementary Table S1).
Acquired ESBLs or AmpC genes were detected in 15 (37.5%) IMP-Ent; 14 has ESBLs genes (12 blaCTX-M-15, 1 blaSHV-134 and 1 blaGES-7) and one AmpC acquired genes (blaFOX-5) (Table 1). Chromosomal AmpC cephalosporinases genes were detected in E. hormaechei and E. asburiae (blaACT), and in E. roggenkampii (blaMIR) (Supplementary Table S1). The prevalence of ESBL in IMP-producing strains (28%) was much lower than the 61.3% detected in a recent Spanish study with isolates producing any type of carbapenemase (Cañada-García et al., 2022); although, as in it, CTX-M-15 predominated. The different prevalence of ESBL and carbapenemase co-production according to Enterobacterales species (Arana et al., 2017; Cañada-García et al., 2022), usually higher in K. pneumoniae, was also detected in IMP-producing isolates, with only 11.5% of isolates co-producing IMP and ESBL in other species than K. pneumoniae.
Six E. hormaechei and one E. asburiae carried mcr-9.1 gene, but they were susceptible to colistin, as previously described (Macesic et al., 2021). No mcr genes were detected in colistin resistant isolates (11 E. roggenkampii and two K. pneumoniae). Cluster-dependent colistin resistance has been described in Enterobacter cloacae complex, mainly due to the expression of phoP/phoQ two-component regulatory system (Guérin et al., 2016).
All 14 IMP-producing K. pneumoniae isolates non-susceptible to aztreonam had additional carbapenemase or ESBL/AmpC acquired genes (Table 1). The high prevalence of aztreonam non-susceptibility in E. cloacae complex could be consistent with overproduction of chromosomal AmpC in these species.
IMP-producing K. pneumoniae isolates belonged to seven capsular polysaccharide cps loci (Supplementary Table S1), mainly KL151 (7; 29.2%), KL53 (5; 20.8%) and KL24 (4; 16.7%). High correlations between K-loci and STs have been previously described (Cañada-García et al., 2022); in this study all KL151 isolates were ST405 and all KL53 were ST464. The two K. pneumoniae isolates causing bacteremia belonged to unrelated KL24 and KL30 cps loci.
A total of eight isolates of K. pneumoniae presented yersiniabactin-encoding locus (ybt), six of them belonged to ST405, and the other two to ST788 and ST3293, respectively. In one ST405 isolate, yersiniabactin, aerobactin (iuc5 linage) and salmochelin (iro5 linage) were detected. Ybt has been previously related to virulence, particularly in pulmonary infection (Lawlor et al., 2007), but this association could not be proved in this study since 6 of the 8 isolates that carried it were implicated in colonizations, and only two were involved in urine infections.
CP-Kpn isolates were grouped into 7 STs (Table 1), being the most prevalent ST405 (n = 7; 29.2%), ST15 (n = 6; 25%) and ST464 (n = 5; 20.8%) which account for 75% of all IMP-producing K. pneumoniae isolates. Full correlation between STs ST405 and ST464 with K-locus KL151 and KL53, respectively, were observed. However, ST15 isolates belonged to two different K-locus (four KL24 and two KL112).
Genome assemblies of the 24 IMP-producing K. pneumoniae isolates included in this study were analyzed using a gene-by-gene approach and the allelic distance from cgMLST was visualized in a minimum spanning tree (Figure 1). Five clusters were detected considering a genetic diversity ≤5 alleles (Figure 1). In only two of those clusters there have been grouped more than three isolates: Cluster 1 consisted of six ST405/IMP-8-produccing isolates submitted from a hospital located in the south of the country (four isolates were identical with the 2.538 alleles cgMLST scheme, and two presented one allele difference), these six isolates differed by an average of 2 SNPs (range 0–3); and Cluster 3 consisted in five ST464/IMP-8-produccing isolates submitted from the same hospital (average allelic distance: 4 alleles), these five isolates differed by an average of five SNPs (range 0–10). The average difference between Cluster 1 and Cluster 3 isolates was 20.489 SNPs and 1.961 alleles.
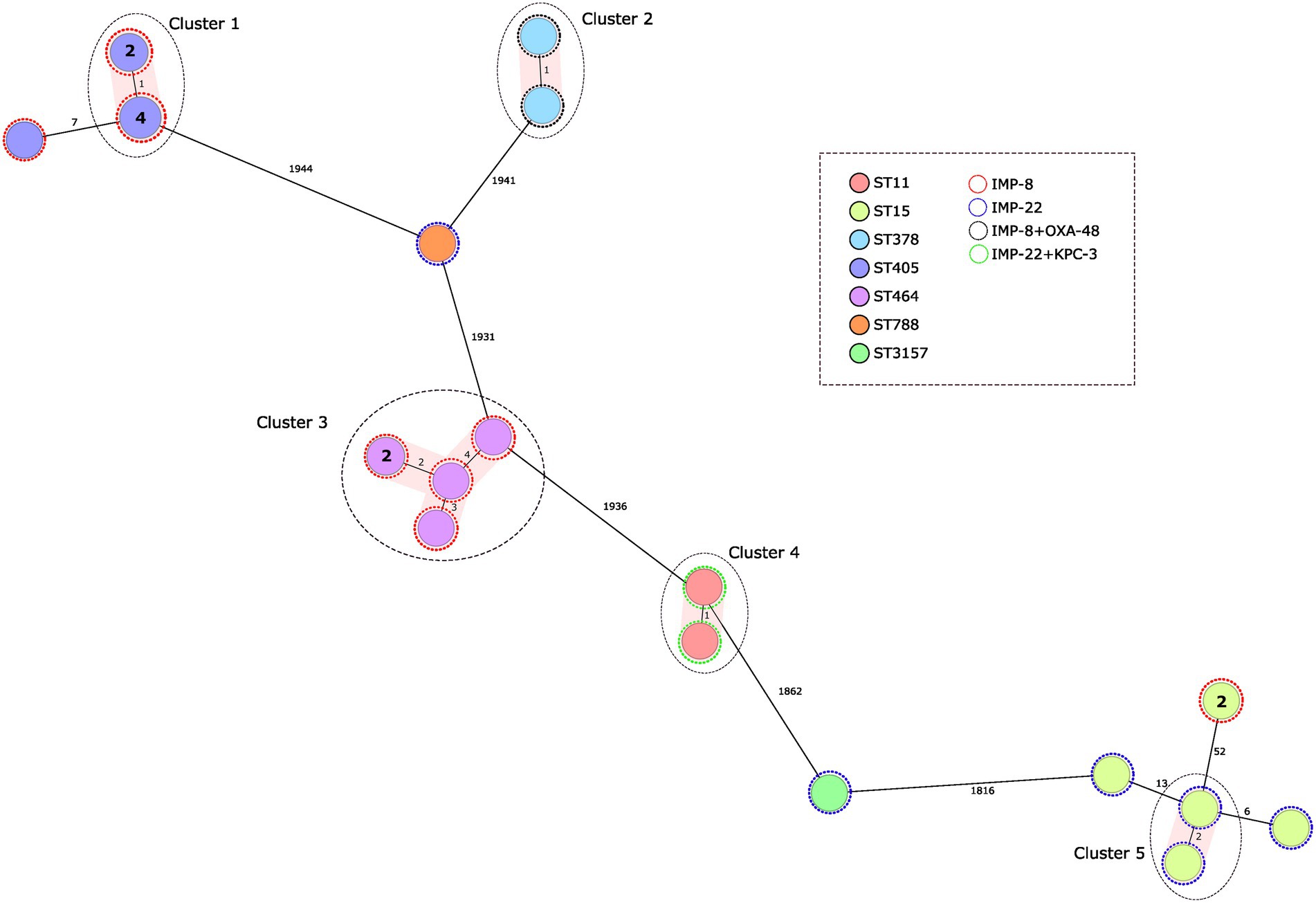
Figure 1. Population structure of IMP-producing Klebsiella pneumoniae: minimum-spanning tree. Distances shown are based on cgMLST of 2.358 genes using the parameter ‘pairwise ignoring missing values’. Fill colors in each circle indicate the MLST type and the color of the dashed line in circles indicates IMP type. Grey ovals represent clusters, applying a threshold of both 5 SNPs and 5 alleles.
K. pneumoniae ST15 represents the only ST detected in two different provinces and encoding different blaIMP genes (blaIMP-8, blaIMP-22). Notably, we did not detect ST14 or ST37, the types found to be most prevalent in an international study analyzing 22 IMP-producing K. pneumoniae isolates, all originating from Asia-Pacific countries (Matsumura et al., 2017). Rather, ST405 and ST15 predominated in our collection. Both have frequently been described in Spain in recent years, although they were associated with other carbapenemases, including OXA-48, for both ST405 and ST15 (Oteo et al., 2015), and VIM-1, for ST15 (Sánchez-Romero et al., 2012). However, other carbapenemases types continue to be prevalent in these STs (Oteo et al., 2015; Cañada-García et al., 2022); the appearance of IMP in different STs according to geographical location could be more related to sporadic transmission by mobile genetic elements between the predominant clones than to a true switch of the ST/carbapenemase type association.
Among IMP-producing E. cloacae complex isolates, 5 different STs were identified; all 13 E. roggenkampii belonged to ST96, E. hormaechei isolates belonged to the ST182 (n = 5), ST68 (n = 2) and ST90 (n = 1), and the E. asburiae isolate belonged to the ST484 (Table 1).
Genome assemblies of the 22 IMP producing E. cloacae complex isolates included in this study were analyzed using a gene-by-gene approach and the allelic distance from cgMLST obtained with the E. cloacae complex, E. hormaechei and E. roggenkampii schemes were visualized in a minimum spanning tree (Figure 2). Only two clusters were detected in E. roggenkampii considering a genetic diversity ≤ 5 alleles in the scheme specific of this specie (Figure 2A). Cluster 1 consisted in four ST96/IMP-8-producing isolates from a hospital in the north of the country (average allelic distance: 3 alleles) and Cluster 2 consisted in three isolates, two were identical and the other presented two allelic differences.
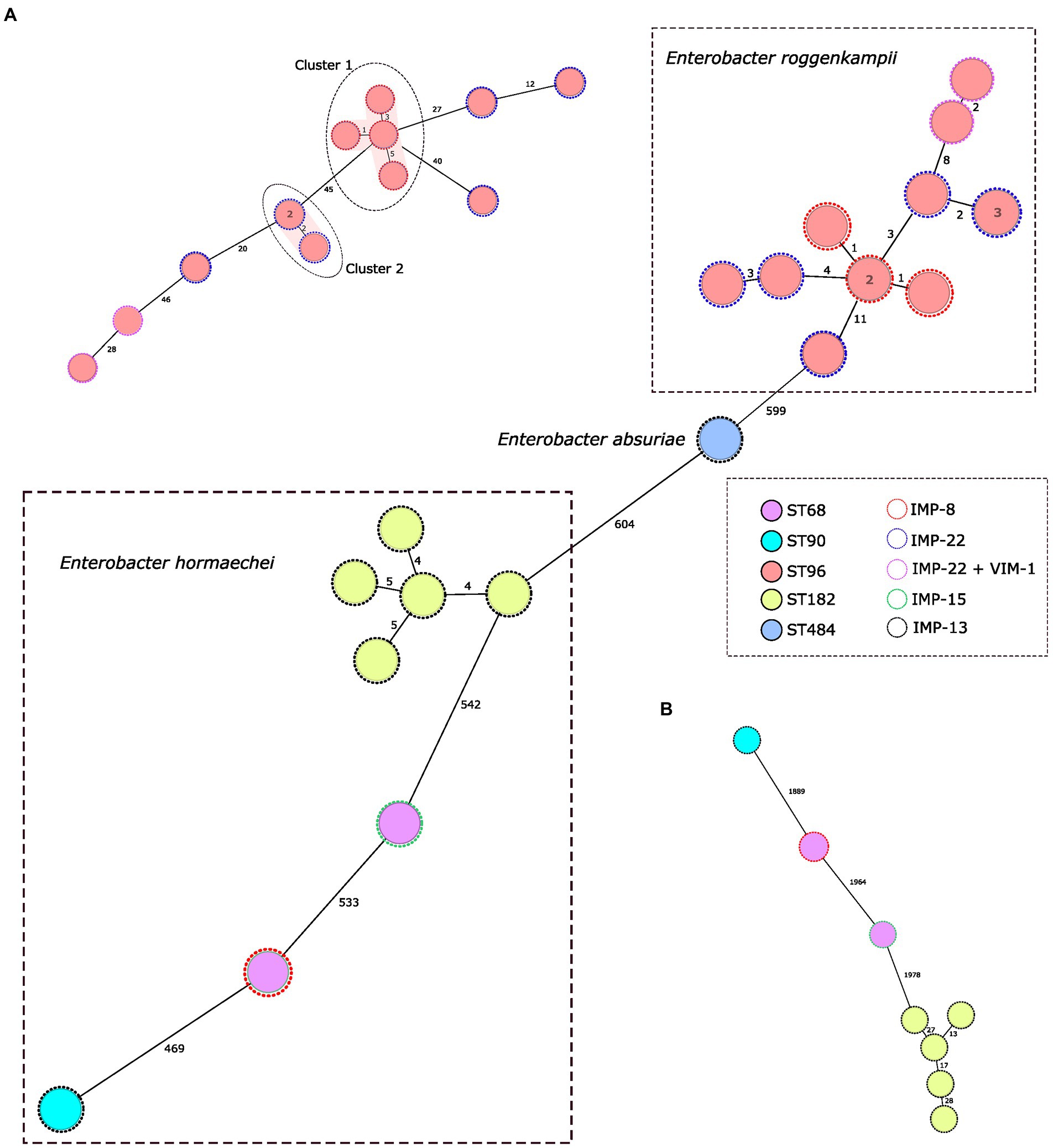
Figure 2. Population structure of IMP-producing Enterobacter cloacae complex: minimum-spanning tree. Distances shown are based on cgMLST of 631 genes using the parameter ‘pairwise ignoring missing values’. Fill colors in each circle indicate the MLST type and the color of the dashed line in circles indicates IMP type. (A,B) represent a minimum-spanning tree for each Enterobacter specie based on specific cgMLST of 2.123 and 2.466 genes, respectively, for Enterobacter hormaechei and Enterobacter roggenkampii. Grey ovals represent clusters, applying a threshold of both 5 SNPs and 5 alleles.
We found that IMP-Ent isolates remain infrequent in Spain, although they have recently been circulating in hospitals from different geographic regions, leading to nosocomial outbreaks. In this study, IMP-8-producing K. pneumoniae and IMP-22-producing E. roggenkampii constitute the most frequent IMP-producing Enterobacterales in Spain. Some specific tests for the diagnosis of MBLs, such as inhibition with EDTA or dipicolinic acid, presented false negative results in some IMP-producing strains. Consequently, this remains a subject of microbiological and epidemiological interest and warrants active surveillance.
Other members of the Spanish IMP Study Group include, Esteban Aznar (Laboratorio BrSalud, San Sebastian de los Reyes, Madrid); José Leiva (Clínica Universitaria de Navarra); Begoña Palop (Hospital Regional de Málaga); José Luis Barrios (Hospital Universitario Cruces, Barakaldo, Vizcaya); Pilar Zamarrón (Hospital Virgen de la Salud, Toledo); Fernando Artiles (Hospital Universitario de Gran Canaria Dr. Negrín, Las Palmas de Gran Canaria); Luis López-Urrutia (Hospital Universitario Río Hortega, Valladolid); Beatriz Orden (Hospital Universitario Puerta de Hierro, Madrid); Susana Hernando (Hospital General de Segovia); and Rafael Carranza (Hospital General La Mancha Centro).
The data presented in the study are deposited in the European Nucleotide Archive (ENA) repository (https://www.ebi.ac.uk/ena), accession number PRJEB54568.
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
MP-V and JO-I conceived, designed and coordinated the study. JC-G, NG, ERA, VB, NL, AN, TC, NM-R, SG-C, JC, EC, BA, and the Spanish IMP Study Group collected isolates and performed the experiments. JC-G, NG, BA, MP-V, and JO-I wrote the manuscript. All authors have read, edited and approved the final manuscript.
This research was supported by grants from the Instituto de Salud Carlos III (numbers PI18CIII/00030 and PI21CIII/00039).
This research was also supported by CIBER-Consorcio Centro de Investigación Biomédica en Red (CB21/13/00095 and CB21/13/000968), Instituto de Salud Carlos III, Ministerio de Ciencia e Innovación and Unión Europea – NextGenerationEU.
This work was supported by Plan Nacional de I + D + i 2013–2016 and Instituto de Salud Carlos III, Subdirección General de Redes y Centros de Investigación Cooperativa, Ministerio de Economía, Industria y Competitividad, Spanish Network for Research in Infectious Diseases (REIPI RD16CIII/0004/0002 and REIPI RD16/0016/0007) and co-financed by the European Development Regional Fund (EDRF), “A way to achieve Europe,” Operative program Intelligent Growth, 2014–2020.
We thank the Genomics Unit of the Centro Nacional de Microbiología for performing the DNA sequencing.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2022.1000787/full#supplementary-material
1. ^http://quast.bioinf.spbau.ru/
2. ^https://www.ncbi.nlm.nih.gov/refseq/about/prokaryotes/
3. ^https://github.com/katholt/Kleborate
Arana, D. M., Ortega, A., González-Barberá, E., Lara, N., Bautista, V., Gómez-Ruíz, D., et al. (2017). Carbapenem-resistant Citrobacter spp. isolated in Spain from 2013 to 2015 produced a variety of carbapenemases including VIM-1, OXA-48, KPC-2, NDM-1 and VIM-2. J. Antimicrob. Chemother. 72, 3283–3287. doi: 10.1093/jac/dkx325
Cañada-García, J. E., Moure, Z., and Sola-Campoy, P. J. (2022). CARB-ES-19 multicenter study of Klebsiella pneumoniae and Escherichia coli from all Spanish provinces reveals interregional spread of high-risk clones such as ST307/OXA-48 and ST512/KPC-3 13, 1–13. doi: 10.3389/fmicb.2022.918362
David, S., Reuter, S., Harris, S. R., Glasner, C., Feltwell, T., Argimon, S., et al. (2019). Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 4, 1919–1929. doi: 10.1038/s41564-019-0492-8
Doyle, D., Peirano, G., Lascols, C., Lloyd, T., Church, D. L., and Pitouta, J. D. D. (2012). Laboratory detection of Enterobacteriaceae that produce carbapenemases. J. Clin. Microbiol. 50, 3877–3880. doi: 10.1128/JCM.02117-12
Elena, A., Cejas, D., Magariños, F., Jewtuchowicz, V., Facente, A., Gutkind, G., et al. (2018). Spread of clonally related Escherichia coli strains harboring an IncA/C1 plasmid encoding IMP-8 and its recruitment into an unrelated MCR-1-containing isolate. Antimicrob. Agents Chemother. 62, 1–6. doi: 10.1128/AAC.02414-17
European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2017). EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Version 2.0. 1–43. Available at: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Resistance_mechanisms/EUCAST_detection_of_resistance_mechanisms_170711.pdf (Accessed July 6, 2022).
European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2022). Clinical Breakpoints. Available at: https://www.eucast.org/clinical_breakpoints/ (Accessed July 6, 2022).
García-Castillo, M., García-Fernández, S., Gómez-Gil, R., Pitart, C., Oviaño, M., Gracia-Ahufinger, I., et al. (2018). Activity of ceftazidime-avibactam against carbapenemase-producing Enterobacteriaceae from urine specimens obtained during the infection-carbapenem resistance evaluation surveillance trial (iCREST) in Spain. Int. J. Antimicrob. Agents 51, 511–515. doi: 10.1016/j.ijantimicag.2018.01.011
Gonçalves, D., Cecílio, P., Faustino, A., Iglesias, C., Branca, F., Estrada, A., et al. (2021). Intra-and extra-hospital dissemination of IMP-22-producing Klebsiella pneumoniae in northern Portugal: the breach of the hospital frontier toward the community. Front. Microbiol. 12:777054. doi: 10.3389/fmicb.2021.777054
Guérin, F., Isnard, C., Sinel, C., Morand, P., Dhalluin, A., Cattoir, V., et al. (2016). Cluster-dependent colistin hetero-resistance in Enterobacter cloacae complex. J. Antimicrob. Chemother. 71, 3058–3061. doi: 10.1093/jac/dkw260
Gupta, S. K., Padmanabhan, B. R., Diene, S. M., Lopez-Rojas, R., Kempf, M., Landraud, L., et al. (2014). ARG-annot, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 58, 212–220. doi: 10.1128/AAC.01310-13
Han, R., Shi, Q., Wu, S., Yin, D., Peng, M., Dong, D., et al. (2020). Dissemination of Carbapenemases (KPC, NDM, OXA-48, IMP, and VIM) among Carbapenem-resistant Enterobacteriaceae isolated from adult and children patients in China. Front. Cell. Infect. Microbiol. 10:314. doi: 10.3389/fcimb.2020.00314
Hunt, M., Mather, A. E., Sánchez-Busó, L., Page, A. J., Parkhill, J., Keane, J. A., et al. (2017). ARIBA: rapid antimicrobial resistance genotyping directly from sequencing reads. Microb. Genomics 3:e000131. doi: 10.1099/mgen.0.000131
Inouye, M., Dashnow, H., Raven, L. A., Schultz, M. B., Pope, B. J., Tomita, T., et al. (2014). SRST2: rapid genomic surveillance for public health and hospital microbiology labs. Genome Med. 6, 90–16. doi: 10.1186/s13073-014-0090-6
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Lam, M. M. C., Wick, R. R., Watts, S. C., Cerdeira, L. T., Wyres, K. L., and Holt, K. E. (2021). A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat. Commun. 12:4188. doi: 10.1038/s41467-021-24448-3
Lawlor, M. S., O’Connor, C., and Miller, V. L. (2007). Yersiniabactin is a virulence factor for Klebsiella pneumoniae during pulmonary infection. Infect. Immun. 75, 1463–1472. doi: 10.1128/IAI.00372-06
Lutgring, J. D., Balbuena, R., Reese, N., Gilbert, S. E., Ansari, U., Bhatnagar, A., et al. (2020). Antibiotic susceptibility of NDM-producing Enterobacterales collected in the United States in 2017 and 2018. Antimicrob. Agents Chemother. 64, 1–9. doi: 10.1128/AAC.00499-20
Macesic, N., Blakeway, L. V., Stewart, J. D., Hawkey, J., Wyres, K. L., Judd, L. M., et al. (2021). Silent spread of mobile colistin resistance gene mcr-9.1 on IncHI2 ‘superplasmids’ in clinical carbapenem-resistant Enterobacterales. Clin. Microbiol. Infect. 27, 1856.e7–1856.e13. doi: 10.1016/j.cmi.2021.04.020
Mateos, M., Hernández-García, M., Del Campo, R., Martínez-García, L., Gijón, D., Morosini, M. I., et al. (2021). Emergence and persistence over time of carbapenemase-producing enterobacter isolates in a Spanish University Hospital in Madrid, Spain (2005-2018). Microb. Drug Resist. 27, 895–903. doi: 10.1089/mdr.2020.0265
Matsumura, Y., Peirano, G., Motyl, M. R., Adams, M. D., Chen, L., Kreiswirth, B., et al. (2017). Global molecular epidemiology of IMP-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 61, 1–9. doi: 10.1128/AAC.02729-16
Miró, E., Agüero, J., Larrosa, M. N., Fernández, A., Conejo, M. C., Bou, G., et al. (2013). Prevalence and molecular epidemiology of acquired AmpC β-lactamases and carbapenemases in Enterobacteriaceae isolates from 35 hospitals in Spain. Eur. J. Clin. Microbiol. Infect. Dis. 32, 253–259. doi: 10.1007/s10096-012-1737-0
Ondov, B. D., Treangen, T. J., Melsted, P., Mallonee, A. B., Bergman, N. H., Koren, S., et al. (2016). Mash: fast genome and metagenome distance estimation using MinHash. Genome Biol. 17, 132–114. doi: 10.1186/s13059-016-0997-x
Ortega, A., Sáez, D., Bautista, V., Fernández-Romero, S., Lara, N., Aracil, B., et al. (2016). Carbapenemase-producing Escherichia coli is becoming more prevalent in Spain mainly because of the polyclonal dissemination of OXA-48. J. Antimicrob. Chemother. 71, 2131–2138. doi: 10.1093/jac/dkw148
Oteo, J., Ortega, A., Bartolomé, R., Bou, G., Conejo, C., Fernández-Martínez, M., et al. (2015). Prospective multicenter study of carbapenemase-producing Enterobacteriaceae from 83 hospitals in Spain reveals high in vitro susceptibility to colistin and meropenem. Antimicrob. Agents Chemother. 59, 3406–3412. doi: 10.1128/AAC.00086-15
Page, A. J., Cummins, C. A., Hunt, M., Wong, V. K., Reuter, S., Holden, M. T. G., et al. (2015). Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics 31, 3691–3693. doi: 10.1093/bioinformatics/btv421
Page, A. J., Taylor, B., Delaney, A. J., Soares, J., Seemann, T., Keane, J. A., et al. (2016). SNP-sites: rapid efficient extraction of SNPs from multi-FASTA alignments. Microb. Genomics 2:e000056. doi: 10.1099/mgen.0.000056
Pérez-Llarena, F. J., Fernández, A., Zamorano, L., Kerff, F., Beceiro, A., Aracil, B., et al. (2012). Characterization of a novel IMP-28 metallo-β-lactamase from a Spanish Klebsiella oxytoca clinical isolate. Antimicrob. Agents Chemother. 56, 4540–4543. doi: 10.1128/AAC.00776-12
Pérez-Vázquez, M., Sola Campoy, P. J., Ortega, A., Bautista, V., Monzón, S., Ruiz-Carrascoso, G., et al. (2019). Emergence of NDM-producing Klebsiella pneumoniae and Escherichia coli in Spain: phylogeny, resistome, virulence and plasmids encoding blaNDM-like genes as determined by WGS. J. Antimicrob. Chemother. 74, 3489–3496. doi: 10.1093/jac/dkz366
Pierce, V. M., Simner, P. J., Lonsway, D. R., Roe-Carpenter, D. E., Johnson, J. K., Brasso, W. B., et al. (2017). Modified carbapenem inactivation method for phenotypic detection of carbapenemase production among Enterobacteriaceae. J. Clin. Microbiol. 55, 2321–2333. doi: 10.1128/JCM.00193-17
Sánchez-Romero, I., Asensio, Á., Oteo, J., Muñoz-Algarra, M., Isidoro, B., Vindel, A., et al. (2012). Nosocomial outbreak of VIM-1-producing Klebsiella pneumoniae isolates of multilocus sequence type 15: molecular basis, clinical risk factors, and outcome. Antimicrob. Agents Chemother. 56, 420–427. doi: 10.1128/AAC.05036-11
Santella, G., Pollini, S., Docquier, J. D., Mereuta, A. I., Gutkind, G., Rossolini, G. M., et al. (2010). Intercontinental dissemination of IMP-13-producing Pseudomonas aeruginosa belonging in sequence type 621. J. Clin. Microbiol. 48, 4342–4343. doi: 10.1128/JCM.00614-10
Sato, T., Harada, K., Usui, M., Yokota, S.-I., and Horiuchi, M. (2022). Colistin susceptibility in companion animal-derived Escherichia coli, Klebsiella spp., and Enterobacter spp. in Japan: frequent isolation of colistin-resistant Enterobacter cloacae complex. Front. Cell. Infect. Microbiol. 12:946841. doi: 10.3389/fcimb.2022.946841
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Sutton, G. G., Brinkac, L. M., Clarke, T. H., and Fouts, D. E. (2018). Enterobacter hormaechei subsp. hoffmannii subsp. nov., Enterobacter hormaechei subsp. xiangfangensis comb. nov., Enterobacter roggenkampii sp. nov., and Enterobacter muelleri is a later heterotypic synonym of Enterobacter asburiae based on computational analysis of sequenced Enterobacter genomes [version 2; referees: 2 approved]. F1000Research 7:521. doi: 10.12688/f1000research.14566.1
Tzouvelekis, L. S., Markogiannakis, A., Psichogiou, M., Tassios, P. T., and Daikos, G. L. (2012). Carbapenemases in Klebsiella pneumoniae and other Enterobacteriaceae: an evolving crisis of global dimensions. Clin. Microbiol. Rev. 25, 682–707. doi: 10.1128/CMR.05035-11
Umeda, K., Nakamura, H., Fukuda, A., Matsumoto, Y., Motooka, D., Nakamura, S., et al. (2021). Genomic characterization of clinical Enterobacter roggenkampii co-harbouring blaIMP-1 and blaGES-5 encoding IncP6 and mcr-9-encoding IncHI2 plasmids isolated in Japan. J. Glob. Antimicrob. Resist. 24, 220–227. doi: 10.1016/j.jgar.2020.11.028
Vázquez-Ucha, J. C., Seoane-Estévez, A., Rodiño-Janeiro, B. K., González-Bardanca, M., Conde-Pérez, K., Martínez-Guitián, M., et al. (2021). Activity of imipenem/relebactam against a Spanish nationwide collection of carbapenemase-producing Enterobacterales. J. Antimicrob. Chemother. 76, 1498–1510. doi: 10.1093/jac/dkab043
Vergara-López, S., Domínguez, M. C., Conejo, M. C., Pascual, Á., and Rodríguez-Baño, J. (2013). Wastewater drainage system as an occult reservoir in a protracted clonal outbreak due to metallo-β-lactamase-producing Klebsiella oxytoca. Clin. Microbiol. Infect. 19, E490–E498. doi: 10.1111/1469-0691.12288
Wang, J. T., Wu, U. I., Lauderdale, T. L. Y., Chen, M. C., Li, S. Y., Hsu, L. Y., et al. (2015). Carbapenem-nonsusceptible Enterobacteriaceae in Taiwan. PLoS One 10, 1–18. doi: 10.1371/journal.pone.0121668
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 13, e1005595–e1005522. doi: 10.1371/journal.pcbi.1005595
Yoon, S. H., Ha, S. M., Lim, J., Kwon, S., and Chun, J. (2017). A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 110, 1281–1286. doi: 10.1007/s10482-017-0844-4
Keywords: antimicrobial resistant bacteria, carbapenem resistance, whole genome sequencing, surveillance, IMP carbapenemase, Enterobacterales
Citation: Cañada-García JE, Grippo N, de Arellano ER, Bautista V, Lara N, Navarro AM, Cabezas T, Martínez-Ramírez NM, García-Cobos S, Calvo J, Cercenado E, Aracil B, Pérez-Vázquez M, Oteo-Iglesias J and the Spanish IMP Study Group (2022) Phenotypic and molecular characterization of IMP-producing Enterobacterales in Spain: Predominance of IMP-8 in Klebsiella pneumoniae and IMP-22 in Enterobacter roggenkampii. Front. Microbiol. 13:1000787. doi: 10.3389/fmicb.2022.1000787
Edited by:
Ana R. Freitas, Cooperativa de Ensino Superior Politécnico e Universitário, PortugalReviewed by:
Teresa G. Ribeiro, University of Porto, PortugalCopyright © 2022 Cañada-García, Grippo, de Arellano, Bautista, Lara, Navarro, Cabezas, Martínez-Ramírez, García-Cobos, Calvo, Cercenado, Aracil, Pérez-Vázquez, Oteo-Iglesias and the Spanish IMP Study Group. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María Pérez-Vázquez, bXBlcmV6dkBpc2NpaWkuZXM=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.