
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 12 January 2022
Sec. Microbiotechnology
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.803490
This article is part of the Research TopicGreen Microbial Technology: From Microbial cell factories to Green industryView all 6 articles
 Jia-jun Huang1,2†
Jia-jun Huang1,2† Tao Wei1,2†
Tao Wei1,2† Zhi-wei Ye1,2
Zhi-wei Ye1,2 Qian-wang Zheng1,2Bing-hua Jiang3Wen-feng Han1,2An-qi Ye1,2Pei-yun Han1,2
Qian-wang Zheng1,2Bing-hua Jiang3Wen-feng Han1,2An-qi Ye1,2Pei-yun Han1,2 Li-qiong Guo1,2*
Li-qiong Guo1,2* Jun-fang Lin1,2*
Jun-fang Lin1,2*
Given the rapid development of genome mining in this decade, the substrate channel of paclitaxel might be identified in the near future. A robust microbial cell factory with gene dbat, encoding a key rate-limiting enzyme 10-deacetylbaccatin III-10-O-transferase (DBAT) in paclitaxel biosynthesis to synthesize the precursor baccatin III, will lay out a promising foundation for paclitaxel de novo synthesis. Here, we integrated gene dbat into the wild-type Escherichia coli BW25113 to construct strain BWD01. Yet, it was relatively unstable in baccatin III synthesis. Mutant gene dbatS189V with improved thermostability was screened out from a semi-rational mutation library of DBAT. When it was over-expressed in an engineered strain N05 with improved acetyl-CoA generation, combined with carbon source optimization of fermentation engineering, the production level of baccatin III was significantly increased. Using this combination, integrated strain N05S01 with mutant dbatS189V achieved a 10.50-fold increase in baccatin III production compared with original strain BWD01. Our findings suggest that the combination of protein engineering and metabolic engineering will become a promising strategy for paclitaxel production.
Baccatin III is a precursor to paclitaxel (Taxol), a billion-dollar drug that is well known and recommended for treatment in more than 20 types of cancer (Wani et al., 1971; Cragg et al., 1993; Thornburg et al., 2017). Besides, this compound has been used to exert immunomodulatory activities on the major histocompatibility complex (MHC)-restricted antigen presentation (Lee et al., 2011), to slow the progression of various types of tumors (Lee et al., 2014), and to ameliorate bleomycin-induced pulmonary fibrosis (Nie et al., 2019). Baccatin III can be synthesized from 10-deacetylbaccatin III (10-DAB) by a one-step acyl transfer reaction of 10-deacetylbaccatin III 10-O-transferase (DBAT) (Walker and Croteau, 1999). Loncaric et al. (2006) verified the regiospecificity of DBAT by identifying the molecular structures of the products of 10-DAB and different coenzyme A (CoA) donors engaged in catalysis by this enzyme. The use of the robust DBAT and sufficient acetyl-CoA in a microbial cell factory provides an efficient way to produce baccatin III and avoids the low atom economy of its chemical synthesis (Danishefsky et al., 1996; Walker and Croteau, 2001).
DBAT is broadly distributed in plants (i.e., Taxus spp.) and endophytic fungi (Sah et al., 2019). Heterologous production of this eukaryotic protein in Escherichia coli may facilitate a higher yield of baccatin III because of (i) its unparalleled short generation times (Bentebibel et al., 2005; Loncaric et al., 2006; Malik et al., 2011), (ii) its higher cell intensity (Lee, 1996; Liu X. et al., 2018), and (iii) the use of a simpler required medium (Bonfill et al., 2007; Wang et al., 2016). Also, benefiting from the easy genetic manipulation tool of E. coli, a rapid gene integration process to further construct an engineered strain as a microbial cell factory is accessible (Wei et al., 2016). Li et al. (2017) expressed DBAT from six species of Taxus spp. in E. coli and discovered that the DBAT from T. wallichiana var. mairei showed the highest activity for 10-DAB. Although DBAT is highly regiospecific, these plant-derived genes generally have low thermal stability. Sadykhov et al. (2006) compared the thermal stability of formate dehydrogenases (FDH) from microorganisms and plants and found an inactivation rate constant for FDH from Glycine max (soybean) that was 10,000-fold higher than that for FDG from Pseudomonas sp. 101 at the same temperature (Alexandra et al., 1999). Thus, there is an urgent need to explore the thermostability of plant-origin DBATs when these are expressed in vivo in E. coli.
Semi-rational design by site-directed mutagenesis is a combination of directed evolution and rational design. Directed evolution is a useful tool to improve enzyme properties (Powell et al., 2001; Zhu and Zhang, 2017). However, it typically requires screening rare positive hits from huge volumes of mutants (Chica et al., 2005; Huang et al., 2019). The use of rational design can greatly decrease the size of the mutant libraries (e.g., down to hundreds of mutants), thus increasing the efficiency of the identification of desired mutants (Zhang et al., 2015; de Souza et al., 2016; Li et al., 2019). For example, Hotspot Wizard 3.0 (Sumbalova et al., 2018) is a web server for automatically established design of mutations and smart libraries in protein engineering, which is based on the amino acid frequency and evolutionary information from three databases1. Chen et al. (2018) applied homologous modeling, molecular docking, and the Hotspot Wizard 3.0 server to construct an L-rhamnose isomerase mutant with a 134.1% increase in relative activity when acting on D-allulose. Pongpamorn et al. (2019) developed a G513Y variant of flavin-dependent monooxygenase with an activity half-life that was 72-fold (50°C) and 160-fold (45°C) longer than the wild-type enzyme when assessed via the FireProt calculations (Musil et al., 2017) of the Hotspot Wizard 3.0 server. Semi-rational design is a suitable tool to investigate the thermostability of DBATs.
The natural substrates to generate baccatin III are 10-DAB and acetyl-CoA. Acetyl-CoA, a central metabolite, is involved in various biological processes (Krivoruchko et al., 2015) and acts as the acetyl donor in the biosynthesis of various acetyl chemicals (Lian et al., 2014). Modification of the acetyl-CoA metabolic pathway can provide efficient and rapid access to these metabolites (Liu et al., 2019; Zhang et al., 2019). Huang et al. (2018) constructed E. coli engineered strains to efficiently generate succinate from acetate via enhanced shunting of acetyl-CoA to succinate. Mainguet et al. (2013) reshaped a nonnative route for a glyoxylate shunt to allow the synthesis of four-carbon tricarboxylic acid (TCA) cycle intermediates from acetyl-CoA in E. coli, which can increase the carbon yield of acetate and biofuels from many carbon sources. Zhang et al. (2019) constructed E. coli metabolic engineered strains for efficient supply of acetyl-CoA from different carbon sources. Besides, studies suggest that when the products’ synthetic pathway involves metabolic intermediates, using glycerol as the fermentation carbon source is beneficial for products generation (Islam et al., 2017; Strucko et al., 2018; Yu et al., 2020). Hence, strengthening the accumulation and generation of acetyl-CoA by metabolic engineering or fermentation engineering is accessible and might provide significant amounts of acetyl donors to produce baccatin III.
In this study, two key factors, namely, DBAT activity and acetyl-CoA supplementation, were optimized to improve the production of baccatin III. The best single mutant of DBAT was screened out from a library of hotspot site-saturation mutants based on FireProt calculations with the Hotspot Wizard 3.0 server and the molecular mechanism was identified by thermostability investigation and molecular docking simulation analysis. Based on this, the engineered E. coli strain N05 with highly efficient supplied acetyl-CoA was used as a host and cultivated with an optimized carbon source to enhance the in vivo synthesis of baccatin III. The successful construction of an engineered E. coli strain N05S01 that yields high amounts of baccatin III provides a rapid and highly economical tool and lays out a foundation for the future industrial production of paclitaxel.
Strains and plasmids used in this study are listed in Table 1. The primers are listed in Supplementary Table 1. Strain E. coli N05, provided by Prof. Tao Yong of the Chinese Academy of Sciences, and E. coli BW25113 provided by Prof. Liu Jianzhong of Sun Yat-sen University, were used as the parental strain for gene integration and baccatin III production. PrimeSTAR® Max DNA Polymerase, used for site-directed mutagenesis, was purchased from Takara Biomedical Technology Co., Ltd. (Dalian, China). Restriction enzymes and T4 DNA ligase were purchased from Thermo Fisher Scientific Co., Ltd. (Beijing, China). 10-DAB and baccatin III were purchased from J&K Scientific Co., Ltd. (Beijing, China). The HiPure Gel Pure DNA Micro Kit was acquired from Magen Co., Ltd. (Guangzhou, China). Acetyl-CoA was purchased from Sigma–Aldrich Inc. (St. Louis, MO, United States). A microorganism acetyl coenzyme A (ACA) enzyme-linked immunosorbent assay (ELISA) kit was obtained from Chundu Biotechnology Co., Ltd. (Wuhan, China). The general chemical reagents used in this study were obtained from standard suppliers.

Table 1. Strains and plasmids used in this study.
Gene dbat was amplified from the vector pET-32a-DBAT using the primers (see Supplementary Table 1) dbat_optF/rbs_dbat_optF/dbat_R to optimize the first 50 bp of gene dbat according to E. coli codon preference and adding RBS sequence in front of the optimized gene dbat. The method and PCR systems of gene integration are referred to Huang et al. (2014). The rbs_dbat fragment was cloned into the BamHI/SalI sites of pHKTT5b to obtain pHKTT5b-dbato. The pHKTT5b-dbato was transformed into E. coli BW25113 and the vector integration was achieved with the help of the helper plasmid pAH69. The plasmid pCP20 was used to remove the FLP recognition target-flanked resistant marker in E. coli BW25113 (Lim et al., 2016). The engineered strain BWD01 integrating PT5 promoter and gene dbat in E. coli BW25113 was inoculated to a 250-mL shake-flask containing 50 mL fresh Terrific Broth (TB) medium (see Supplementary Table 2). 10-DAB with a final concentration of 80 μM was added as the substrate, and glycerol in TB medium acted as a carbon source. The bacteria were incubated at 150 rpm for 48 h at 37, 25, and 20°C, respectively.
There was 1 mL of fermentation culture pipetted and centrifuged at 12,000 rpm for 5 min to separate the cell sedimentation and the medium supernatants. The baccatin III in the supernatants was analyzed using a high-performance liquid chromatography (HPLC) system [LC-2000, Techcomp (China) Ltd.] equipped with an ultraviolet (UV) detector (LC-2030) and a Diamosil C18 (2) column (250 mm × 4.6 mm, 5 μm). The mobile phase was 50% acetonitrile (A)-50% water (B), with a flow rate of 1 ml/min, at 28°C, and the UV detection wavelength was 227 nm. All assays were carried out in triplicate.
The pET-32a-DBAT was transformed into E. coli BL21 (DE3), then the recombinant was incubated in 50 ml fresh TB medium containing ampicillin (100 μg/ml) and incubated at 37°C with mixing at 200 rpm for 2–3 h until a spectrophotometer registered an optical density at 600 nm (OD600) of approximately 0.6–0.8. IPTG was added at a final concentration of 0.02 mM to induce gene expression, and the cell cultures were incubated for an additional 15 h at 20°C and 120 rpm. The following operations were performed on ice or at 4°C unless otherwise stated. After induction, cells were harvested by centrifugation (5,000 rpm, 4°C, 10 min) and washed twice with phosphate-buffered saline (PBS) [137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, and 2 mM KH2PO4, pH7.4]. The precipitates were stored at 4°C and were used for expression assays of DBAT and whole-cell catalysis.
The precipitates were resuspended in 10 ml lysis buffer [PBS buffer (pH 7.4), 1 mM PMSF, and 1 mM DTT], then sonicated for 12 min in an ice bath with 25% intermittent power (4 s on, 6 s off). The sample was centrifuged at 8,000 rpm at 4°C for 15 min to remove the cellular debris. The cell supernatants were filtered through a 0.22 μm membrane for further purification by using Ni-NTA agarose resin (Qiagen) according to the manufacturer’s manual. The crude extract, purified protein, and the cellular debris were detected by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (12%) and Coomassie blue staining, respectively.
The precipitates were resuspended in 2 mL of PBS, adjusted to an OD600 of 15. The cell mixtures were incubated in the presence of 80 μM 10-DAB and 2 mM glucose (acted as carbon source) at different temperatures (18°C–37°C) with 150 rpm shaking for 1 h to investigate the enzymatic function of DBAT. The reaction mixture was centrifuged at maximum speed for 10 min and the supernatant was analyzed by HPLC. For experimental controls, whole-cell biosynthesis (Wu and Li, 2018) of baccatin III without adding glucose was performed under the same conditions as already described. All assays were carried out in triplicate.
A three-dimensional (3D) structure model of DBAT was predicted by homology modeling via MODELLER 9.18 in our previous work (Lin et al., 2018). The model was uploaded to the Hotspot Wizard 3.0 online server, where the mutation hotspots and amino acid information in the solvent channel were predicted. Single-point mutations were determined by the Hotspot Wizard 3.0 server and generated via site-directed mutagenesis (details in Supplementary Material). The pairs of forward and reverse primers used are listed in Supplementary Table 3, with mutated bases in italics.
Wild-type DBAT (WT) strain and all mutant strains were induced in TB medium and sonicated as described. The crude extraction of all samples was detected by SDS-PAGE (12%) followed by Coomassie blue staining, and the expression level results were analyzed by the image analysis software Image J2 (Carvajal-Vergara et al., 2010; Liu Q. et al., 2018).
The best mutant from the library screening (DBATbest) and the WT strain were chosen for thermal stability analysis. The induction and expression were the same as described. The supernatants were used for the thermal stability study. The crude extraction of WT and the best mutant were incubated at 25°C for 0, 0.25, 0.5, 0.75, 1, 2, 3, 4, and 5 h, respectively. The assay mixture, which contained a certain amount of crude enzyme, 80 μM 10-DAB, 1 mM acetyl-CoA, and 1 mM Mg2+ in 50 mM PBS (pH 7.0), was allowed to react at 22°C for 30 min. The reaction was terminated by adding to the mixture 500 μl methanol and analysis was proceeded by HPLC. Relative conversion values were expressed as percentages relative to the conversion measured at the initial state (25°C, 0 h).
In silico studies were performed to analyze the difference between the skeleton of the mutation models and the WT model. Molecular docking studies were used to compare the conformation of the complex of DBATbest with two substrates, 10-DAB and acetyl-CoA, and the complex of WT with the same substrates. Visual analysis used PyMOL software to investigate the change in the difference in the number of hydrogen bonds between both substrates and target proteins and the binding energy (a parameter showing stability) (Satish et al., 2018) of these two complexes.
The DBATbest gene was integrated into E. coli BW25113 via the same method as described, the engineered strain named BWS01. Fermentation of strain BWS01 was performed under the same condition as strain BWD01. Compared to the bioconversion capacity of producing baccatin III by engineered strains with different integrated genes to produce baccatin III.
Escherichia coli strain N05 was reported as an engineered strain with high acetyl-CoA production. The DBATbest gene was integrated into E. coli N05 via the same method as described, the engineered strain named N05S01. Fermentation of strain N05S01 was performed under the same condition as strain BWD01 and strain BWS01. Finally, to further improve the production of baccatin III, the concentration of glycerol in the TB medium was increased. After fermentation, baccatin III yields in culture supernatant were detected by HPLC as described. All assays were carried out in triplicate.
Engineered strain BWD01 was constructed with an integrant expression of dbat gene controlled by T5 promoter in E. coli BW25113 under the cooperation of the integration vector pHKTT5b, the helper plasmid pAH69, and pCP20. The supernatants of strain BWD01 fermented at 37°C showed no baccatin III in an HPLC analysis. However, the yield showed 4.68 ± 0.21 μM when the fermentation temperature was reduced to 25°C. Once the temperature was decreased to 20°C, the yield could reach 15.79 ± 0.63 μM. Results showed that the integrant expression of gene dbat in E. coli was relatively thermally sensitive.
Semi-rational design is a reliable strategy to improve the thermal stability of DBAT, and it is necessary to construct a heterologous plasmid expression system for investigating DBAT’s activity. DBAT expressed in E. coli BL21 (DE3) using the expression vector pET-32a partially formed inclusions. The protein purification showed that the purified protein located at a protein size of about 70 kDa was the 6 × His fusion target protein with a theoretical molecular weight of 67 kDa, which indicated the successful expression of DBAT in E. coli (Supplementary Figure 1). The recombinant cell pellet without sonication treatment was used for whole-cell biosynthesis of baccatin III. Using glucose as a carbon source substrate with 10-DAB addition, baccatin III can be detected in the supernatants after the reaction. Supplementary Figure 2 showed that without the participation of glucose, no product could be detected in the supernatant of the whole-cell system. It speculated that DBAT had little capacity to pull acetyl-CoA from the stable metabolic process. Results implied that glucose may act as an “activator” in this bioconversion to achieve the in vivo synthesis of baccatin III (Figure 1). The relative conversion of baccatin III presented a bell-shaped curve from 18 to 35°C and reached the maximum at 22°C (Supplementary Figure 3). The conversion of baccatin III at 25°C was the largest change, and it exactly decreased to 50%. When the temperature exceeds 35°C, no product could be detected. Therefore, 25°C, which allowed DBAT to present a medium level of conversion, was used as the subsequent screening temperature of the mutation library.
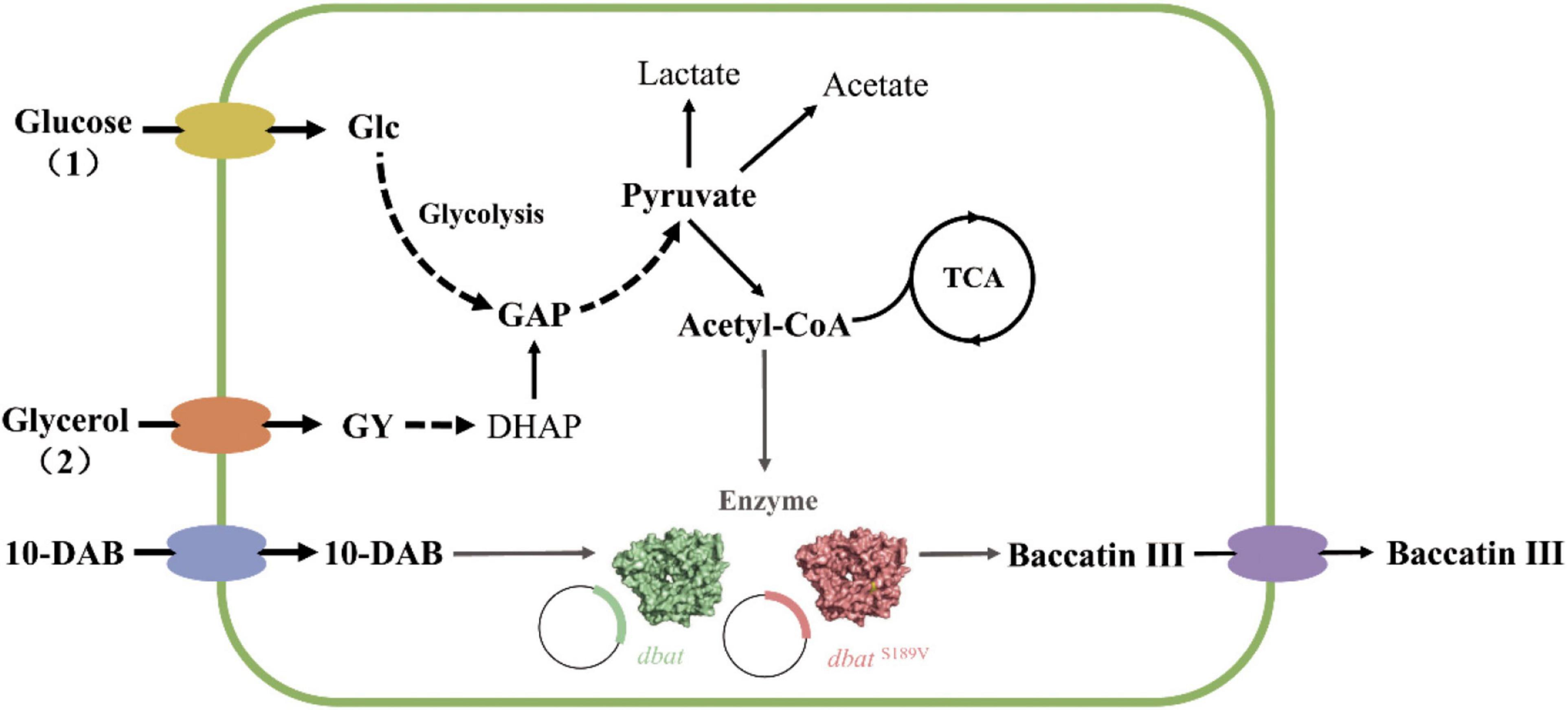
Figure 1. Process for whole-cell bioconversion of baccatin III by wild-type DBAT (WT) and DBATS189V in E. coli. (1) Producing baccatin III using glucose as a carbon source. Glucose is metabolized to produce pyruvate, which in turn, produces acetyl-CoA, which provides acetyl groups for 10-DAB to produce baccatin III. (2) Producing baccatin III using glycerol as a carbon source. Glycerol is metabolized to produce DHAP, then metabolized by glycolysis to produce pyruvate, which in turn, produces acetyl-CoA, which provides acetyl groups for 10-DAB, and produces baccatin III. The dotted arrow indicates that the multi-step reaction process is omitted in this metabolic pathway diagram. The gray arrow points to the direction of the enzymatic reaction to produce baccatin III. DBAT and DBATS189V were overexpressed in E. coli by recombinant plasmids. Glc glucose, GY glycerol; GAP, glyceraldehyde-3-phosphate; DHAP, dihydroxyacetone phosphate; 10-DAB, 10-deacetylbaccatin III; TAC, tricarboxylic acid cycle; DBAT, 10-deacetylbaccatin III-10-O-transferase.
The binding pocket and solvent channel of DBAT were predicted by the Hotspot Wizard 3.0 server (Supplementary Figure 4). There were 13 mutational “hotspots” (Pro37, Val39, Asn42, Ile43, Ser122, His162, His123, Glu124, Ser159, Leu168, Gly171, Ile175, and Ser189) in the solvent channel (Figure 2) chosen for site-directed mutagenesis. As the residue His162 was proved to be the key catalytic site of DBAT in our previous study (You et al., 2018). When the residue His162 was mutated to other amino acids, DBAT will completely lose its activity. Similar results were obtained in other studies (Li et al., 2017). Thus, the residue His162 was excluded in this study. Mutational landscapes, amino acid frequency, and the evolutionary information of these amino acid residues based on data from three major databases to provide mutation-selection were also analyzed by the server (see Supplementary Table 4). Each mutation acted on the production of baccatin III differently. Peak areas were converted to product concentrations based on external standard calibrations of baccatin III (Supplementary Figure 5). The yield of baccatin III obtained by the WT enzyme whole-cell catalytic reaction was used as the 100% whole-cell conversion rate of baccatin III. Compared with the yield of baccatin III, the mutant’s relative whole-cell conversion rate was obtained. Results showed that DBATG171C and DBATS159R had no measurable product in the whole-cell bioconversion of 10-DAB at 25°C (Figure 3A), while other mutations, such as DBATP37T, DBATV39A, DBATN42F, and DBATH123L, were negatively impacted on bioconversion. On the other hand, compared with that of WT, the bioconversion efficiency of DBATS189L and DBATS189V was relatively 3.15 and 3.81 times higher. Results suggested that residue Ser189 was probably the critical site for thermal stability improvement. To screen out the optimal point mutation of the Ser189 site, a new round of saturation mutation was performed. Results showed that DBATS189V still exhibited the best bioconversion ability of 10-DAB and DBATS189C was the second best with a 3.31-fold increase (Figure 3B). When S189 was mutated to leucine and isoleucine, the yields of baccatin III were also improved (higher than twofold). The remaining mutants were less effective in the catalytic activity.
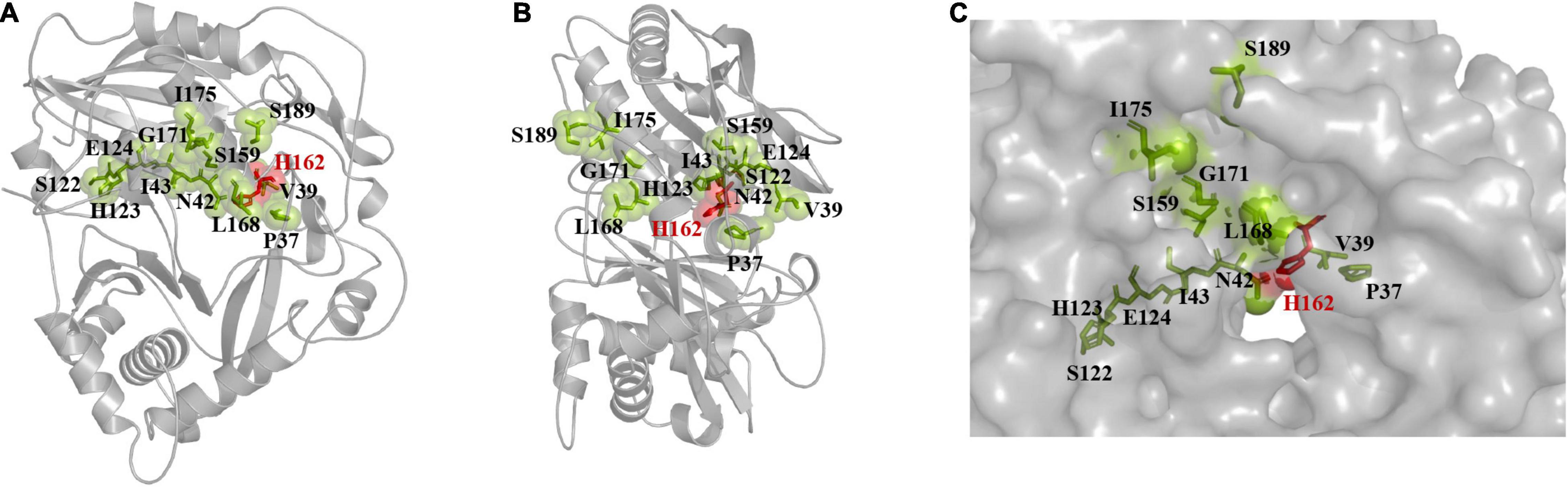
Figure 2. “Hotspots” distribution was predicted by Hotspot Wizard 3.0. The center residue His162 is displayed as dots style and shown in red. The others are displayed as spheres styles and shown in green by PyMOL. (A) The front face of the model is displayed as cartoon style. (B) The side face of the model is displayed as cartoon style. (C) The distribution of residues near the solvent channel, the model is displayed as surface style.
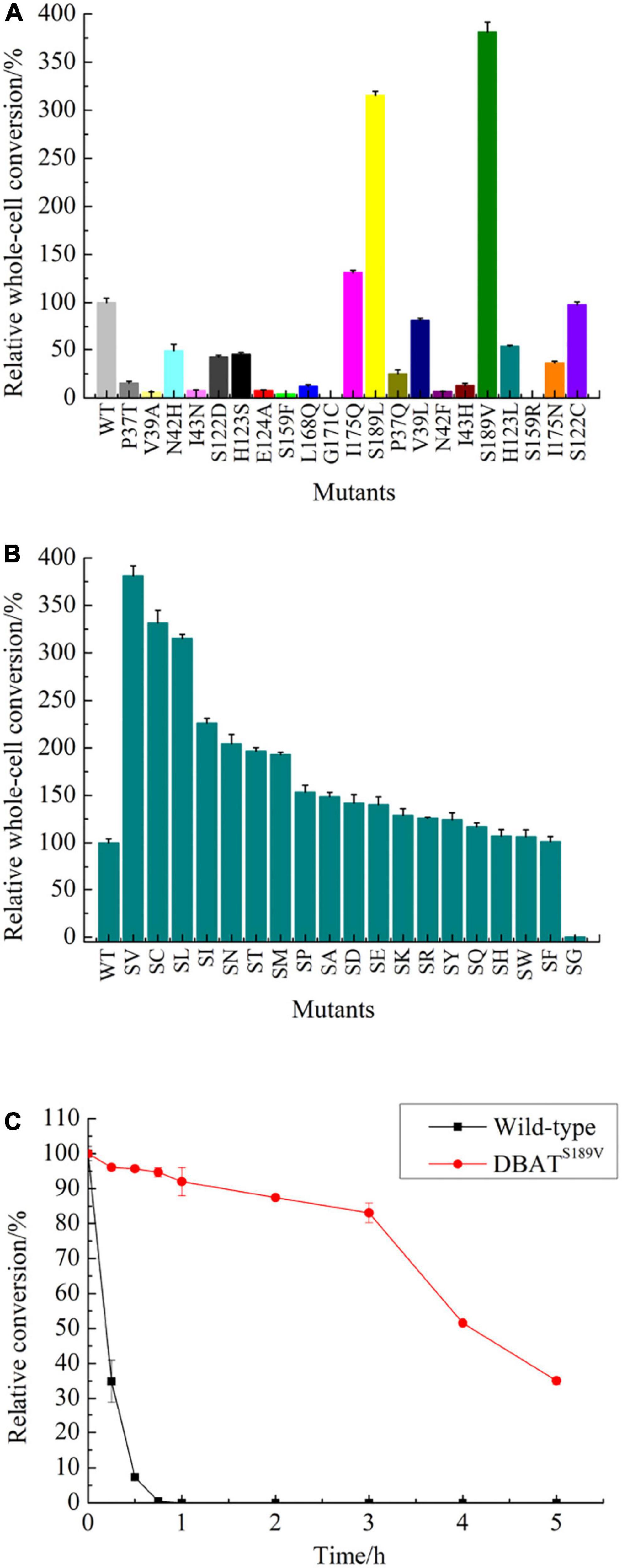
Figure 3. Two rounds of mutation library screening results and thermal stability analysis results. (A) Screening results for the “hotspot” mutants by whole-cell bioconversion. (B) Screening results for site-saturated mutations by whole-cell bioconversion. (C) Thermal stability of the crude enzymes of DBATS189V and WT.
Wild-type and all mutants were sonicated, and the supernatants were examined by SDS-PAGE (12%) followed by staining with Coomassie blue (Supplementary Figure 6). Located in the position of the mutant protein according to the WT target protein, and compared their protein expression levels roughly. No significant difference was shown in the expression levels of mutants and WT evaluated by Image J software (Supplementary Figure 7). The highest relative expression was found with DBATG171C and the lowest with DBATS159R, both were negative mutants with unmeasurable products. DBATS189V, the most effective mutant in biotransformation, was only 88.09% of the WT. Results suggested that the soluble expression level might not be the key factor in the semi-rational evolution of DBAT.
Thermal stability results showed that DBATS189V could still retain 80% of its activity when placed at 25°C for 3 h. Its t1/2 value (half-life period) at 25°C was about 245 min. In contrast, the t1/2 value of WT was only about 10 min (Figure 3C), and its activity was lost, and the product was undetectable after 1 h. The thermal stability analysis results showed that the half-life of mutant S189V at 25°C was 24.5 times longer than that of WT.
As a result of molecular docking, it could also be seen that two substrates, namely, 10-DAB and acetyl-CoA, entered the target protein from both sides, with mutual binding of the target protein in the solvent channel (Figures 4A, 5A). On one hand, it was found that the binding energy of the mutational complex S189V_10-DAB_AcCoA (S189V docking with acetyl-CoA and 10-DAB) was lower than that of the wild-type complex WT_10-DAB_AcCoA (WT docking with acetyl-CoA and 10-DAB), which was reflected in its improved stability (Table 2). On the other hand, the number of hydrogen bonds formed between substrates and DBATS189V (Figures 4B–E) was more than that between substrates and WT (Figures 5B–E). All the results further indicated that the conversion from serine to valine improved the structural and thermal stability of DBAT and consequently increased production.
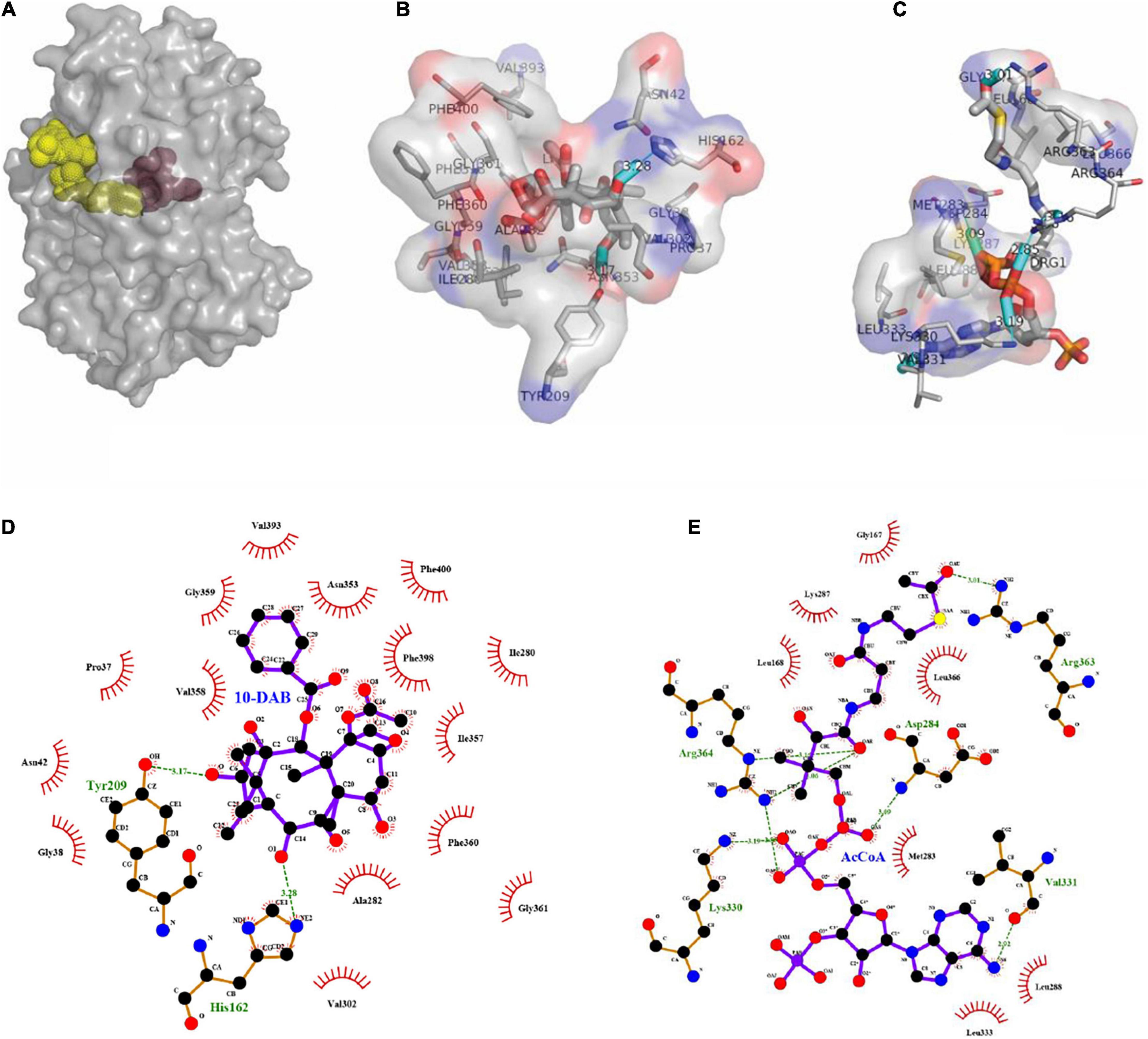
Figure 4. Molecular docking results of DBATS198V and two substrates. The complex was named S189V_10-DAB_AcCoA. (A) Panoramic conformation of S189V_10-DAB_AcCoA. (B) Interaction between 10-DAB and DBATS189V in 3D-model. (C) Interaction between acetyl-CoA and DBATS189V in 3D-model. (D) The hydrogen bonds were shown as green dotted lines between 10-DAB and DBATS189V. (E) The hydrogen bonds were shown as green dotted lines between acetyl-CoA and DBATS189V.
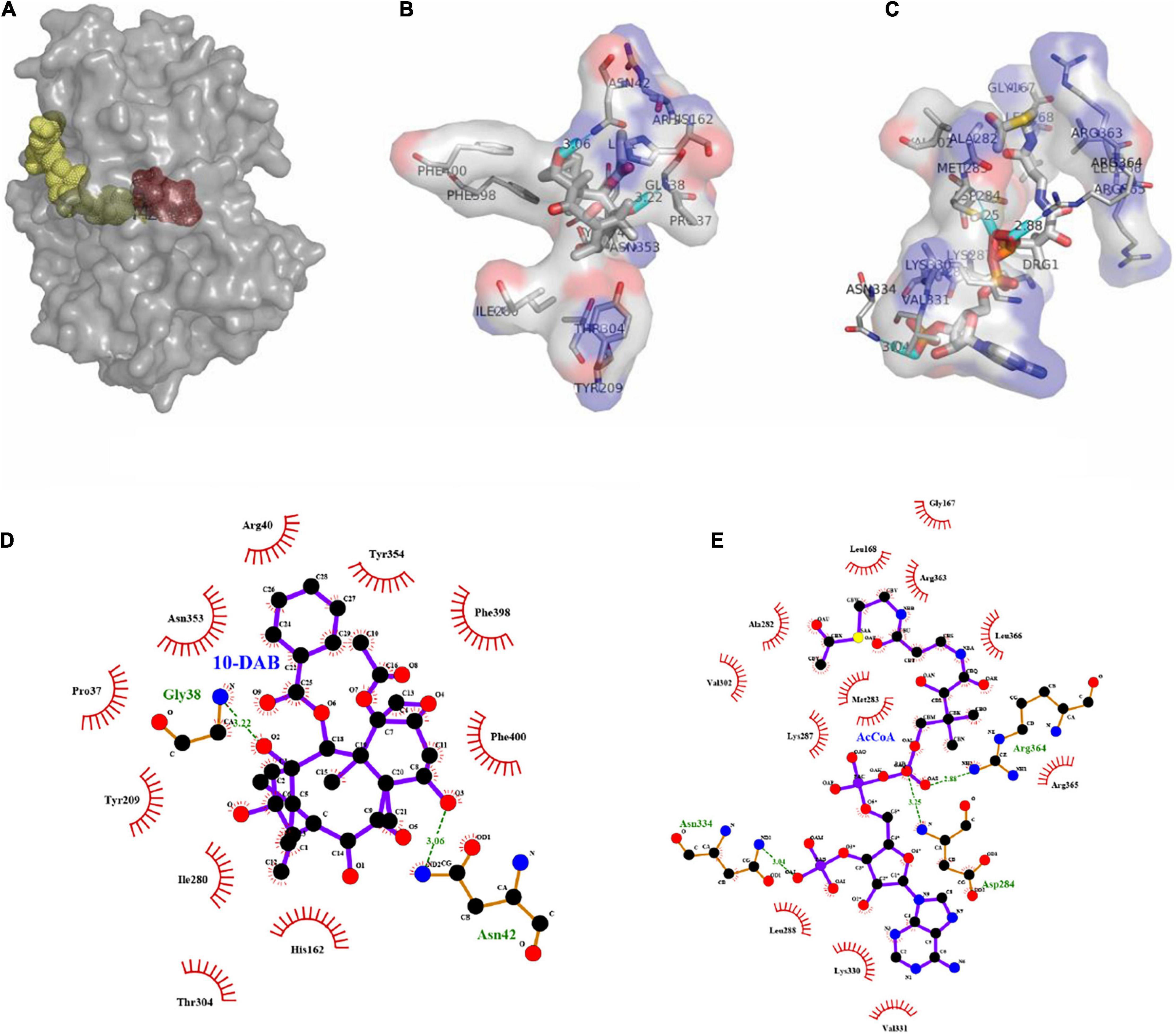
Figure 5. Molecular docking results of WT and two substrates. The complex was named WT_10-DAB_AcCoA. (A) Panoramic conformation of WT_10-DAB_AcCoA. (B) Interaction between 10-DAB and WT in 3D-model. (C) Interaction between acetyl-CoA and WT in 3D-model. (D) The hydrogen bonds were shown as green dotted lines between 10-DAB and WT. (E) The hydrogen bonds were shown as green dotted lines between acetyl-CoA and WT.

Table 2. Key parameters of docking complex WT_10-DAB_AcCoA and S189V_10-DAB_AcCoA.
Glycerol was screened out in carbon source optimization from starch, sucrose, glucose, glycerol, lactose, and fructose (Figure 6A, details in Supplementary Material). Fermentation with glycerol (Figure 1) in the same concentration, the conversion rate of baccatin III could reach 41.05 ± 1.06% (increased by 24.3% compared with glucose). Interestingly, the cell density of the fermentation broth also increased and OD600 reached 30.6 (increased by 16.2% compared with glucose) (Figure 6B). The addition of a carbon source was continuously increased, and it was found that when glycerol was added to a concentration of 10 g/L, the conversion rate of baccatin III could reach a maximum of 60.36 ± 1.29%, and the OD600 could reach 32.78 (Figure 6C).
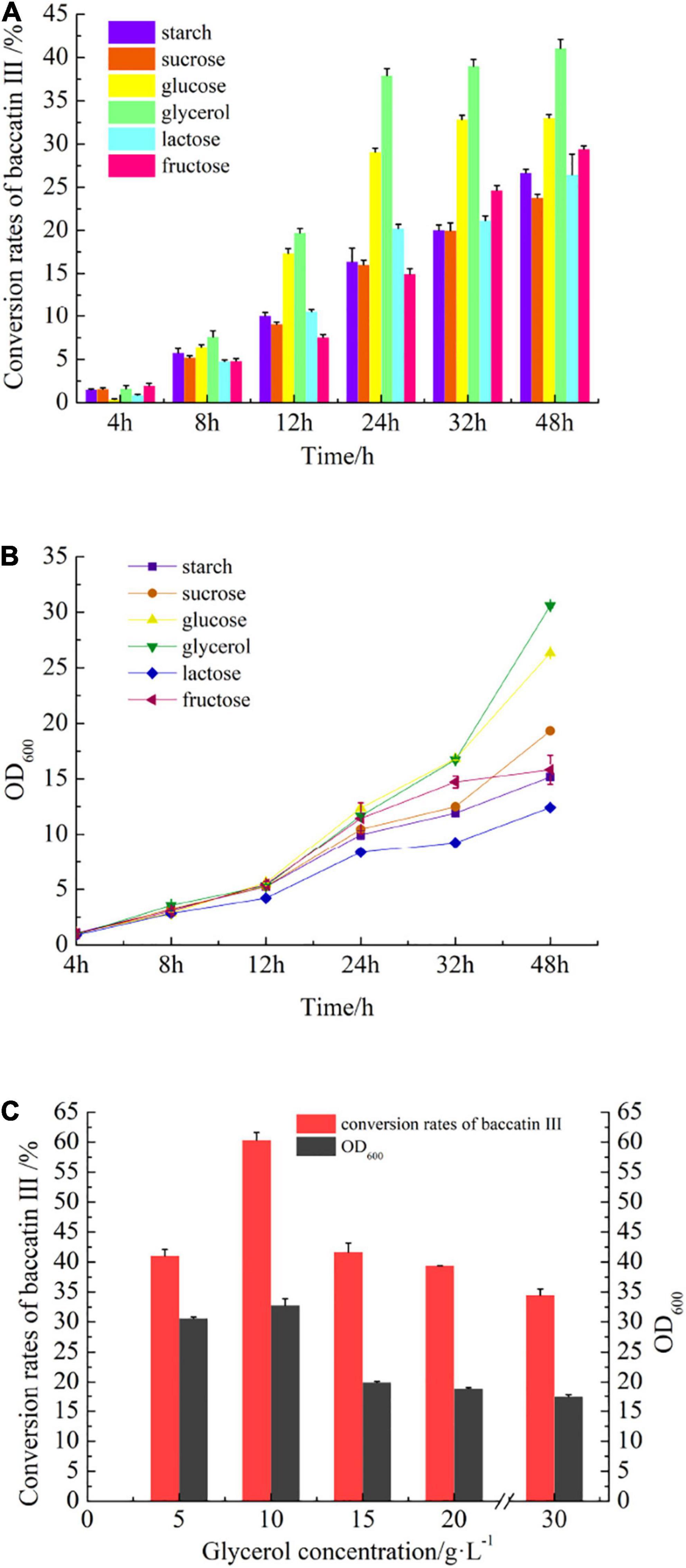
Figure 6. The effect of carbon source optimization on baccatin III production and cell growth during fermentation. (A) The effect of different carbon sources on baccatin III production. (B) The effect of different carbon sources on cell growth. Take OD600 value as the parameter of cell concentration. (C) The effect of glycerol content on baccatin III production and cell growth.
Strain N05 is a metabolically engineered strain that can efficiently accumulate acetyl-CoA (Zhang et al., 2019). The mutant gene dbatS189V was integrated into the chromosomes of E. coli strains BW25113 and N05, respectively. Two engineered strains, BWS01 (BW25113::dbatS189V) and N05S01 (N05::dbatS189V) were successfully constructed. Using 10-DAB with a final concentration of 80 μM, baccatin III production in the culture supernatant of strain N05S01, strain BWS01, and strain BWD01 fermented at 25°C for 48 h in the optimal condition of using glycerol as the carbon source with 10 g/L concentration were detected (Figure 7). The yield of baccatin III of strain N05S01 was 49.10 ± 0.36 μM (28.8 mg/L), which was 10.50 times higher than that of the fermentation product of strain BWD01 (4.68 ± 0.21 μM).
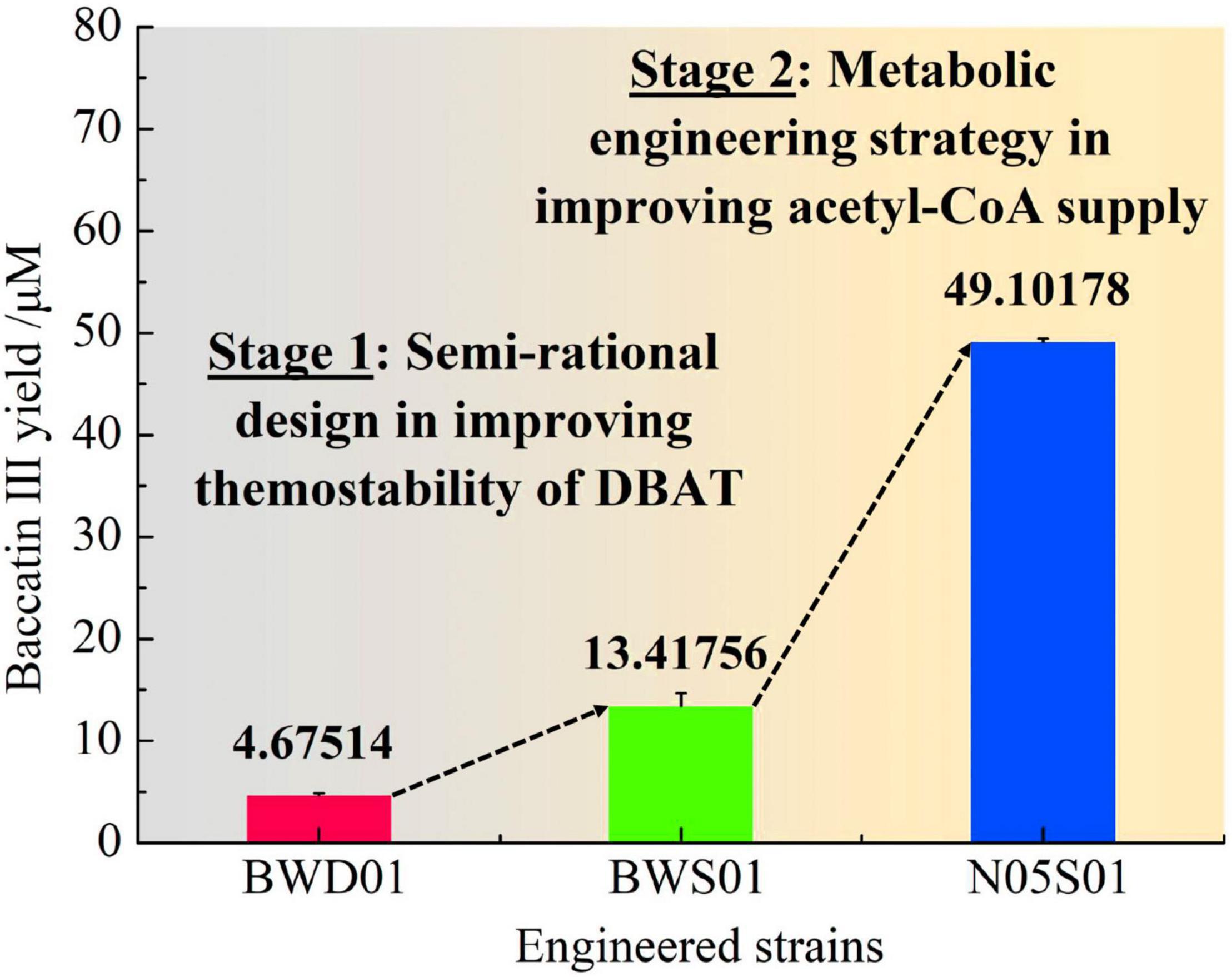
Figure 7. Results of the fermentation of integrated engineered strains to produce baccatin III. Baccatin III production has undergone two stages of improvement. Stage 1, Semi-rational design in improving thermostability of DBAT; Stage 2, Metabolic engineering strategy in improving acetyl-CoA supply.
Since the dbat gene was successfully cloned over two decades, research on DBAT has mainly focused on (i) cloning and heterologous expression of DBAT from different species (Guo et al., 2007; Han et al., 2014; Sah et al., 2019), (ii) the regional specificity and substrate diversity of DBAT (Loncaric et al., 2006, 2007), and (iii) the exploration of the unnatural substrate (Li et al., 2017; Lin et al., 2018; You et al., 2018, 2019; Huang et al., 2020). However, research on the thermal stability of DBAT has not been reported. In this study, baccatin III cannot be detected in the fermentation supernatant of the dbat gene-integrated strain BWD01 fermented at 37°C. It was found that the induction temperature of DBAT was mainly low (probably 20°C or 16°C) in the studies of the heterologous expression of DBAT in E. coli (Li et al., 2017; Lin et al., 2018). In this study, fermentation temperature increased from 20°C to 25°C and reduced 70.36% of the baccatin III yield. Taken together, improving the thermal stability of DBAT of plant origin might be a considerable path to increase the yield and titer of baccatin III in the E. coli cell factories (Huang et al., 2020).
In the following study, semi-rational evolution was applied to improve the thermal stability of DBAT. S189, which was located at the entrance of the solvent channel as well as the surface of DBAT, was the crucial residue selected by the mutation library in this study. Compared with the WT, the yield of baccatin III produced by mutants DBATS189V, DBATS189C, DBATS189I, and DBATS189L were significantly increased. When the serine at position 189 was mutated to cysteine, the surface structure of mutant DBATS189C changed, and a hydrogen bond was formed between Cys189 and Lys295, which is also located on the surface structure of the enzyme (Supplementary Figure 8) and might enhance the stability of the structure (Sun et al., 2015; Muchiri and Walker, 2017). Therefore, it was speculated that the S189C mutation improves the catalytic efficiency of DBAT by enhancing structural stability. DBATS189V, DBATS189I, and DBATS189L were mutants that represented a transition from a hydrophilic amino acid to a hydrophobic amino acid. Changing S189 into a hydrophobic amino acid was conducive to creating a hydrophobic environment that was more suitable for substrate entry and reaction. Gallardo et al. (2010) reported that increasing the hydrophobic folding of the enzyme near the binding pocket was beneficial in terms of improving its stability. Meanwhile, studies showed that valine located on the surface of the enzyme plays an important role in the stability of protein structure (Sugita et al., 1998; Wu et al., 2009; Weisslocker-Schaetzel et al., 2017). Valine, leucine, and isoleucine presented similar molecular structures and hydrophobic effects on DBAT. Valine is not only a hydrophobic amino acid, but also plays an important role in improving the rigidity of enzymes, and it can also form hydrogen bonds with Lys295 (Supplementary Figures 8E,F), which might explain why DBATS189V has the best catalytic effect.
Enhancing the stability of the enzyme surface and the hydrophobicity of the entrance of the enzyme activity channel have a significant effect on improving the catalytic efficiency of the enzyme. Recently, it was reported that enhancing the subunit interface of enzymes can improve stability more efficiently. Computational interface redesign can be a rapid and powerful strategy for enzyme stabilization (Meng et al., 2020).
Through the optimization of carbon source types, it was found that glycerol was the most suitable carbon source for baccatin III production. Discussed from the perspective of the metabolic process after the carbon source enters the strain, all kinds of carbon sources generate pyruvate through the glycolysis pathway and then form acetyl-CoA to provide raw materials for the reaction. Starch, sucrose, lactose, and fructose all need to produce glucose before entering the glycolysis pathway. Therefore, the path to produce acetyl-CoA is longer than the metabolic path of glucose. However, fructose has a shunt path that can enter the glycolysis pathway through generating fructose 6-phosphate. Thus, the effect of fructose was relatively better than the other three. Glycerol can enter the glycolysis pathway by forming glyceraldehyde 3-phosphate (GAP) in three steps. It was shorter than that from glucose (four steps). Therefore, glycerol became the most suitable carbon source in the pathway to generate acetyl-CoA, glucose and fructose were followed by Figure 6A. Through the optimization of carbon source dosage, it was also found that the supply of intracellular acetyl-CoA was insufficient for DBATS189V (Figure 6C). To investigate the acetyl-CoA accumulation after generating pyruvate from the glycolysis pathway is necessary. Zhang et al. (2019) constructed an engineered E. coli strain that modified the metabolic pathway of the metabolic node substance acetyl-CoA genetically can efficiently be synthesized N-acetylglycine using different carbon sources. Strain N05 has knocked out the gltA gene, a downstream gene of the metabolism of acetyl-CoA, to block acetyl-CoA from entering the tricarboxylic acid cycle, which enhanced the accumulation of acetyl-CoA in the result.
Testing by a microorganism acetyl coenzyme A (ACA) enzyme-linked immunosorbent assay (ELISA) kit (Chundu Biotechnology Co., Ltd.), the content of acetyl-CoA in BL21 (DE3) decreased during the test period (OD600 value varied from 0.6 to 1.6). In contrast, the knockout of gene gltA leads to the increase of acetyl-CoA accumulation in strain N05 during the logarithmic phase (Supplementary Table 5). The shunting of acetyl-CoA flux contributed more to baccatin III production. The engineered strain with gene gltA deletion might establish a better balance between cell growth and baccatin III production.
Currently, the preparation method of baccatin III is mainly through direct extraction from yews. The yield of baccatin III obtained by using plant cell in vitro culture preparation is low (2.4–4.6 mg/L by using immobilized cell culture in shaking flask and 7.8 mg/L by using bioreactor) (Malik et al., 2011). The upgrade of the bioreactor and the addition of different types of elicitors and inducers had produced a corresponding increase in the production of baccatin III by suspension cell culture, but they also take 8 days to 1 month for preparation. Compared to the methods mentioned above, microbial cell factory with integrant thermal stable mutant expression in this study is marker-less, short-period, and highly efficient. It could consume a simple carbon source and require no additional cofactor in fermentation. Considering the high medicinal value of baccatin III and paclitaxel, the engineered strains constructed in this study provided a competitive way for industrial synthesis.
Given that the wild-type DBAT was thermally unstable, semi-rational evolution of DBAT was performed in this study. Bioinformatics methods (e.g., HotSpot Wizard, and Molecular Docking) were carried out and 13 mutation sites were screened. The conversion efficiency of mutant DBATS189V, the best candidate, was higher than that of wild-type DBAT (a 3.81-fold increase). In addition, to meet the increasing demand of acetyl donors from mutant DBAT, E. coli strain N05, which could generate more acetyl-CoA, was used as a host for integrant expression of mutant DBATS189V. The engineered strain N05S01 produced 49.10 ± 0.36 μM baccatin III after fermentation at 25°C for 48 h. It was 10.50 times higher than that of strain BWD01, which had no optimization in thermal stability of DBAT and acetyl-CoA supply. Taken together, this research combined the approaches of protein engineering and metabolic engineering to construct a cell factory with an increased yield of the exogenous products baccatin III. It laid out a valuable foundation for the construction of the paclitaxel cell factory in the coming future.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
J-JH and TW carried out the main work, collected and analyzed the data, and drafted the manuscript. W-FH, A-QY, and P-YH participated in the research. Z-WY, Q-WZ, and B-HJ supervised the work, participated in data analysis, and revised the manuscript. J-FL and L-QG participated in the conception and design of the study, and finalized the manuscript. All authors read and approved the final manuscript.
This work was supported by the Key-Area Research and Development Program of Guangdong Province (Grant Nos. 2014B050505018, 2018B020205003, and 2020B020205001) and National Natural Science Foundation of China (Grant Nos. 31772373 and 32072646).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Tao Yong from the Chinese Academy of Sciences for providing engineered strain E. coli N05, Liu Jianzhong from Sun Yat-sen University for providing plasmid pHKTT5b, pAH69, pCP20, and strain E. coli DH5αλpir and E. coli BW25113, and Huang Rui for project discussions. We also thank Enago (http://www.enago.cn) for English language editing.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.803490/full#supplementary-material
DBAT, 10-deacetylbaccatin III 10-O-transferase; 10-DAB, 10-deacetylbaccatin III; WT, wild-type DBAT; IPTG, isopropyl- β-D -thiogalactopyranoside; ACA ELISA, acetyl-coenzyme A enzyme-linked immunosorbent assay; AcCoA, acetyl-coenzyme A; LB, Luria-Bertani; TB, Terrific Broth; TB-10-gly, TB medium with glycerol at a concentration of 10 g/L; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; HPLC, high-performance liquid chromatography.
Alexandra, M. R., Galkin, A. G., Kulakova, L. B., Serov, A. E., Savitsky, P. A., Fedorchuk, V. V., et al. (1999). Bacterial formate dehydrogenase. increasing the enzyme thermal stability by hydrophobization of alpha-helices. FEBS Lett. 445, 183–188. doi: 10.1016/S0014-5793(99)00127-1
Bentebibel, S., Moyano, E., Palazon, J., Cusido, R. M., Bonfill, M., Eibl, R., et al. (2005). Effects of immobilization by entrapment in alginate and scale-up on paclitaxel and baccatin III production in cell suspension cultures of Taxus baccata. Biotechnol. Bioeng. 89, 647–655. doi: 10.1002/bit.20321
Bonfill, M., Bentebibel, S., Cusidó, R. M., and Eibl, R. (2007). Paclitaxel and baccatin III production induced by methyl jasmonate in free and immobilized cells of Taxus baccata. Biol. Plantarum 51, 647–652. doi: 10.1007/s10535-007-0137-2
Carvajal-Vergara, X., Sevilla, A., D’Souza, S. L., Ang, Y. S., Schaniel, C., Lee, D. F., et al. (2010). Patient-specific induced pluripotent stem-cell-derived models of LEOPARD syndrome. Nature 465, 808–812. doi: 10.1038/nature09005
Chen, Z., Chen, J., Zhang, W., Zhang, T., Guang, C., and Mu, W. (2018). Improving thermostability and catalytic behavior of L-rhamnose isomerase from Caldicellulosiruptor obsidiansis OB47 toward D-allulose by site-directed mutagenesis. J. Agric. Food Chem. 66, 12017–12024. doi: 10.1021/acs.jafc.8b05107
Chica, R. A., Doucet, N., and Pelletier, J. N. (2005). Semi-rational approaches to engineering enzyme activity: combining the benefits of directed evolution and rational design. Curr. Opin. Biotechnol. 16, 378–384. doi: 10.1016/j.copbio.2005.06.004
Cragg, G. M., Schepartz, S. A., Suffness, M., and Grever, M. R. (1993). The taxol supply crisis. new NCI policies for handling the large-scale production of novel natural product anticancer and anti-HIV agents. J. Nat. Prod. 56, 1657–1668. doi: 10.1021/np50100a001
Danishefsky, S. J., Masters, J. J., and Young, W. B. (1996). Total synthesis of baccatin III and taxol. J. Am. Chem. Soc. 118, 2843–2859.
de Souza, A. R., de Araujo, G. C., Zanphorlin, L. M., Ruller, R., Franco, F. C., Torres, F. A., et al. (2016). Engineering increased thermostability in the GH-10 endo-1,4-β-xylanase from Thermoascus aurantiacus CBMAI 756. Int. J. Biol. Macromol. 93(Pt. A), 20–26. doi: 10.1016/j.ijbiomac.2016.08.056
Gallardo, O., Pastor, F. I., Polaina, J., Diaz, P., Lysek, R., Vogel, P., et al. (2010). Structural insights into the specificity of Xyn10B from Paenibacillus barcinonensis and its improved stability by forced protein evolution. J. Biol. Chem. 285, 2721–2733. doi: 10.1074/jbc.M109.064394
Guo, B., Kai, G., Gong, Y., Jin, H., Wang, Y., Miao, Z., et al. (2007). Molecular cloning and heterologous expression of a 10-deacetylbaccatin III-10-O-acetyltransferase cDNA from taxus x media. Mol. Biol. Rep. 34, 89–95. doi: 10.1007/s11033-006-9018-6
Han, F., Kang, L. Z., Zeng, X. L., Ye, Z. W., Guo, L. Q., and Lin, J. F. (2014). Bioproduction of baccatin III, an advanced precursor of paclitaxel, with transgenic Flammulina velutipes expressing the 10-deacetylbaccatin III-10-O-acetyltransferase gene. J. Sci. Food Agric. 94, 2376–2383. doi: 10.1002/jsfa.6562
Huang, B., Yang, H., Fang, G., Zhang, X., Wu, H., Li, Z., et al. (2018). Central pathway engineering for enhanced succinate biosynthesis from acetate in Escherichia coli. Biotechnol. Bioeng. 115, 943–954. doi: 10.1002/bit.26528
Huang, J. J., Wei, T., Lin, J. F., Guo, L. Q., Han, W. F., Han, P. Y., et al. (2020). High-effective biosynthesis of baccatin III by using the alternative acetyl substrate, N-acetyl-D-glucosamine. J. Appl. Microbiol. 129, 345–355. doi: 10.1111/jam.14620
Huang, M., Chen, Y., and Liu, J. (2014). Chromosomal engineering of Escherichia coli for efficient production of Coenzyme Q10. Chin. J. Chem. Eng. 22, 559–569. doi: 10.1016/s1004-9541(14)60082-3
Huang, R., Chen, H., Upp, D. M., Lewis, J. C., and Zhang, Y.-H. P. J. (2019). A high-throughput method for directed evolution of NAD(P)+-dependent dehydrogenases for the reduction of biomimetic nicotinamide analogues. ACS Catalysis 9, 11709–11719. doi: 10.1021/acscatal.9b03840
Islam, Z. U., Klein, M., Aßkamp, M. R., Ødum, A. S. R., and Nevoigt, E. (2017). A modular metabolic engineering approach for the production of 1,2-propanediol from glycerol by Saccharomyces cerevisiae. Metab. Eng. 44, 223–235. doi: 10.1016/j.ymben.2017.10.002
Krivoruchko, A., Zhang, Y., Siewers, V., Chen, Y., and Nielsen, J. (2015). Microbial acetyl-CoA metabolism and metabolic engineering. Metab. Eng. 28, 28–42. doi: 10.1016/j.ymben.2014.11.009
Lee, S. Y. (1996). High cell-density culture of Escherichia coli. Trends Biotechnol. 14, 98–105. doi: 10.1016/0167-7799(96)80930-9
Lee, Y. H., Lee, Y. R., Kim, K. H., Im, S. A., Song, S., Lee, M. K., et al. (2011). Baccatin III, a synthetic precursor of taxol, enhances MHC-restricted antigen presentation in dendritic cells. Int. Immunopharmacol. 11, 985–991. doi: 10.1016/j.intimp.2011.02.013
Lee, Y. H., Lee, Y. R., Park, C. S., Im, S. A., Song, S., Hong, J. T., et al. (2014). Baccatin III, a precursor for the semisynthesis of paclitaxel, inhibits the accumulation and suppressive activity of myeloid-derived suppressor cells in tumor-bearing mice. Int. Immunopharmacol. 21, 487–493. doi: 10.1016/j.intimp.2014.06.012
Li, B. J., Wang, H., Gong, T., Chen, J. J., Chen, T. J., Yang, J. L., et al. (2017). Improving 10-deacetylbaccatin III-10-β-O-acetyltransferase catalytic fitness for taxol production. Nat. Commun. 8:15544. doi: 10.1038/ncomms15544
Li, T. B., Zhao, F. J., Liu, Z., Jin, Y., Liu, Y., Pei, X. Q., et al. (2019). Structure-guided engineering of ChKRED20 from Chryseobacterium sp. CA49 for asymmetric reduction of aryl ketoesters. Enzyme Microb. Technol. 125, 29–36. doi: 10.1016/j.enzmictec.2019.03.001
Lian, J., Si, T., Nair, N. U., and Zhao, H. (2014). Design and construction of acetyl-CoA overproducing Saccharomyces cerevisiae strains. Metab. Eng. 24, 139–149. doi: 10.1016/j.ymben.2014.05.010
Lim, H. G., Noh, M. H., Jeong, J. H., Park, S., and Jung, G. Y. (2016). Optimum rebalancing of the 3-hydroxypropionic acid production pathway from glycerol in Escherichia coli. ACS Synth. Biol. 5, 1247–1255. doi: 10.1021/acssynbio.5b00303
Lin, S. L., Wei, T., Lin, J. F., Guo, L. Q., Wu, G. P., Wei, J. B., et al. (2018). Bio-production of baccatin III, an important precursor of paclitaxel by a cost-effective approach. Mol. Biotechnol. 60, 492–505. doi: 10.1007/s12033-018-0090-7
Liu, H., Fan, J., Wang, C., Li, C., and Zhou, X. (2019). Enhanced β-amyrin synthesis in Saccharomyces cerevisiae by coupling an optimal acetyl-CoA supply pathway. J. Agric. Food Chem. 67, 3723–3732. doi: 10.1021/acs.jafc.9b00653
Liu, Q., Zhou, J., Yang, T., Zhang, X., Xu, M., and Rao, Z. (2018). Efficient biosynthesis of L-phenylglycine by an engineered Escherichia coli with a tunable multi-enzyme-coordinate expression system. Appl. Microbiol. Biotechnol. 102, 2129–2141. doi: 10.1007/s00253-018-8741-y
Liu, X., Yan, Y., Liu, Y., Mo, T., Wang, X., Song, Y., et al. (2018). Cell culture establishment and regulation of two phenylethanoid glycosides accumulation in cell suspension culture of desert plant Cistanche tubulosa. Plant Cell Tissue Organ Culture 134, 107–118. doi: 10.1007/s11240-018-1404-y
Loncaric, C., Merriweather, E., and Walker, K. D. (2006). Profiling a taxol pathway 10β-acetyltransferase: assessment of the specificity and the production of baccatin III by in vivo acetylation in E. coli. Chem. Biol. 13, 309–317. doi: 10.1016/j.chembiol.2006.01.006
Loncaric, C., Ward, A. F., and Walker, K. D. (2007). Expression of an acetyl-CoA synthase and a CoA-transferase in Escherichia coli to produce modified taxanes in vivo. Biotechnol. J. 2, 266–274. doi: 10.1002/biot.200600194
Mainguet, S. E., Gronenberg, L. S., Wong, S. S., and Liao, J. C. (2013). A reverse glyoxylate shunt to build a non-native route from C4 to C2 in Escherichia coli. Metab. Eng. 19, 116–127. doi: 10.1016/j.ymben.2013.06.004
Malik, S., Cusidó, R. M., Mirjalili, M. H., Moyano, E., Palazón, J., and Bonfill, M. (2011). Production of the anticancer drug taxol in taxus baccata suspension cultures: a review. Proc. Biochem. 46, 23–34. doi: 10.1016/j.procbio.2010.09.004
Meng, Q., Capra, N., Palacio, C. M., Lanfranchi, E., Otzen, M., van Schie, L. Z., et al. (2020). Robust omega-transaminases by computational stabilization of the subunit interface. ACS Catal. 10, 2915–2928. doi: 10.1021/acscatal.9b05223
Muchiri, R., and Walker, K. D. (2017). Paclitaxel biosynthesis: adenylation and thiolation domains of an NRPS TycA PheAT module produce various arylisoserine CoA thioesters. Biochemistry 56, 1415–1425. doi: 10.1021/acs.biochem.6b01188
Musil, M., Stourac, J., Bendl, J., Brezovsky, J., Prokop, Z., Zendulka, J., et al. (2017). FireProt: web server for automated design of thermostable proteins. Nucleic Acids Res. 45, W393–W399. doi: 10.1093/nar/gkx285
Nie, Y., Zhang, D., Qian, F., and Wu, Y. (2019). Baccatin III ameliorates bleomycin-induced pulmonary fibrosis via suppression of TGF-β1 production and TGF-β1-induced fibroblast differentiation. Int. Immunopharmacol. 74:105696. doi: 10.1016/j.intimp.2019.105696
Pongpamorn, P., Watthaisong, P., Pimviriyakul, P., Jaruwat, A., Lawan, N., Chitnumsub, P., et al. (2019). Identification of a hotspot residue for improving the thermostability of a flavin-dependent monooxygenase. Chembiochem 20, 3020–3031. doi: 10.1002/cbic.201900413
Powell, K. A., Ramer, S. W., Del Cardayré, S. B., Stemmer, W. P., and Tobin, M. B. (2001). Directed evolution and biocatalysis. Angew. Chem. Int. Ed. 40, 3948–3959.
Sadykhov, E. G., Serov, A. E., Voinova, N. S., Uglanova, S. V., Petrov, A. S., Alekseeva, A. A., et al. (2006). A comparative study of the thermal stability of formate dehydrogenases from microorganisms and plants. Appl. Biochem. Microbiol. 42, 236–240. doi: 10.1134/s0003683806030021
Sah, B., Subban, K., and Jayabaskaran, C. (2019). Biochemical insights into the recombinant 10-deacetylbaccatin III-10-β-O-acetyltransferase enzyme from the taxol-producing endophytic fungus Lasiodiplodia theobromae. FEMS Microbiol. Lett. 366:72. doi: 10.1093/femsle/fnz072
Satish, L., Millan, S., Sasidharan, V. V., and Sahoo, H. (2018). Molecular level insight into the effect of triethyloctylammonium bromide on the structure, thermal stability, and activity of bovine serum albumin. Int. J. Biol. Macromol. 107(Pt. A), 186–193. doi: 10.1016/j.ijbiomac.2017.08.157
Strucko, T., Zirngibl, K., Pereira, F., Kafkia, E., Mohamed, E. T., Rettel, M., et al. (2018). Laboratory evolution reveals regulatory and metabolic trade-offs of glycerol utilization in Saccharomyces cerevisiae. Metab. Eng. 47, 73–82. doi: 10.1016/j.ymben.2018.03.006
Sugita, Y., Kitao, A., and Go, N. (1998). Computational analysis of thermal stability: effect of Ile→val mutations in human lysozyme. Folding Design 3, 173–181. doi: 10.1016/S1359-0278(98)00025-X
Sumbalova, L., Stourac, J., Martinek, T., Bednar, D., and Damborsky, J. (2018). HotSpot wizard 3.0: web server for automated design of mutations and smart libraries based on sequence input information. Nucleic Acids Res. 46, W356–W362. doi: 10.1093/nar/gky417
Sun, Z., Lonsdale, R., Kong, X. D., Xu, J. H., Zhou, J., and Reetz, M. T. (2015). Reshaping an enzyme binding pocket for enhanced and inverted stereoselectivity: use of smallest amino acid alphabets in directed evolution. Angew Chem. Int. Ed. Engl. 54, 12410–12415. doi: 10.1002/anie.201501809
Thornburg, C. K., Walter, T., and Walker, K. D. (2017). Biocatalysis of a paclitaxel analogue: conversion of baccatin III to N-debenzoyl-N-(2-furoyl)paclitaxel and characterization of an amino phenylpropanoyl CoA transferase. Biochemistry 56, 5920–5930. doi: 10.1021/acs.biochem.7b00912
Walker, K., and Croteau, R. (1999). Molecular cloning of a 10-deacetylbaccatin III-10-O-acetyltransferase cDNA from Taxus and functional expression in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 97, 583–587. doi: 10.1073/pnas.97.2.583
Walker, K., and Croteau, R. (2001). Taxol biosynthetic genes. Phytochemistry 58, 1–7. doi: 10.1016/S0031-9422(01)00160-1
Wang, S., Li, C., Wang, H., Zhong, X., Zhao, J., Tong, Y., et al. (2016). Effect of elicitors, precursors and metabolic inhibitors on paclitaxel production by taxus cuspidata cell culture. J. Forestry Res. 27, 1257–1263. doi: 10.1007/s11676-016-0217-2
Wani, M. C., Taylor, H. L., Wall, M. E., Coggon, P., and Mcphail, A. T. (1971). Plant antitumor agents. VI. The isolation and structure of taxol, a novel antileukemic and antitumor agent from taxus brevifolia. J. Am. Chem. Soc. 93, 2325–2327. doi: 10.1021/ja00738a045
Wei, T., Cheng, B. Y., and Liu, J. Z. (2016). Genome engineering Escherichia coli for L-DOPA overproduction from glucose. Sci. Rep. 6:30080. doi: 10.1038/srep30080
Weisslocker-Schaetzel, M., Lembrouk, M., Santolini, J., and Dorlet, P. (2017). Revisiting the Val/Ile mutation in mammalian and bacterial nitric oxide synthases: a spectroscopic and kinetic study. Biochemistry 56, 748–756. doi: 10.1021/acs.biochem.6b01018
Wu, Q. Y., Li, F., and Wang, X. Y. (2009). Val65 plays an important role in the substrate synergism, structural stability and activity of arginine kinase. Int. J. Biol. Macromol. 45, 393–398. doi: 10.1016/j.ijbiomac.2009.06.016
Wu, S., and Li, Z. (2018). Whole-cell cascade biotransformations for one-pot multistep organic synthesis. Chem. Cat. Chem. 10, 2164–2178. doi: 10.1002/cctc.201701669
You, L. F., Huang, J. J., Lin, S. L., Wei, T., Zheng, Q. W., Jiang, B. H., et al. (2019). In vitro enzymatic synthesis of baccatin III with novel and cheap acetyl donors by the recombinant taxoid 10β-O-acetyltransferase. Bio. Biotransform. 37, 1–7. doi: 10.1080/10242422.2018.1549235
You, L. F., Wei, T., Zheng, Q. W., Lin, J. F., Guo, L. Q., Jiang, B. H., et al. (2018). Activity essential residue analysis of Taxoid 10β-O-acetyltransferase for enzymatic synthesis of baccatin III. Appl. Biochem. Biotechnol. 186, 949–959. doi: 10.1007/s12010-018-2789-0
Yu, Y. H., Pan, H. Y., Guo, L. Q., Lin, J. F., Liao, H. L., and Li, H. Y. (2020). Successful biosynthesis of natural antioxidant ergothioneine in Saccharomyces cerevisiae required only two genes from Grifola frondosa. Microbial. Cell Factories 19:164. doi: 10.1186/s12934-020-01421-1
Zhang, D., Chen, X., Chi, J., Feng, J., Wu, Q., and Zhu, D. (2015). Semi-rational engineering a carbonyl reductase for the enantioselective reduction of β-amino ketones. ACS Catalysis 5, 2452–2457. doi: 10.1021/acscatal.5b00226
Zhang, S., Yang, W., Chen, H., Liu, B., Lin, B., and Tao, Y. (2019). Metabolic engineering for efficient supply of acetyl-CoA from different carbon sources in Escherichia coli. Microb. Cell Fact. 18:130. doi: 10.1186/s12934-019-1177-y
Keywords: baccatin III, 10-deacetylbaccatin III-10-O-transferase, gene integration, semi-rational design, thermostability, acetyl-CoA supplement
Citation: Huang J-j, Wei T, Ye Z-w, Zheng Q-w, Jiang B-h, Han W-f, Ye A-q, Han P-y, Guo L-q and Lin J-f (2022) Microbial Cell Factory of Baccatin III Preparation in Escherichia coli by Increasing DBAT Thermostability and in vivo Acetyl-CoA Supply. Front. Microbiol. 12:803490. doi: 10.3389/fmicb.2021.803490
Received: 28 October 2021; Accepted: 09 December 2021;
Published: 12 January 2022.
Edited by:
Jun Feng, University of Wisconsin–Madison, United StatesReviewed by:
Yuxin Zhao, Iowa State University, United StatesCopyright © 2022 Huang, Wei, Ye, Zheng, Jiang, Han, Ye, Han, Guo and Lin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun-fang Lin, bGluamZAc2NhdS5lZHUuY24=; Li-qiong Guo, Z3VvbHFAc2NhdS5lZHUuY24=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.