 Gabriela Flores-Vargas
Gabriela Flores-Vargas Jordyn Bergsveinson
Jordyn Bergsveinson John R. Lawrence2
John R. Lawrence2 Darren R. Korber
Darren R. Korber
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 13 December 2021
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.766242
This article is part of the Research Topic Biofilms: Multi-species Community Interactions View all 6 articles
Characterizing the response of microbial communities to a range of antibiotic concentrations is one of the strategies used to understand the impact of antibiotic resistance. Many studies have described the occurrence and prevalence of antibiotic resistance in microbial communities from reservoirs such as hospitals, sewage, and farm feedlots, where bacteria are often exposed to high and/or constant concentrations of antibiotics. Outside of these sources, antibiotics generally occur at lower, sub-minimum inhibitory concentrations (sub-MICs). The constant exposure to low concentrations of antibiotics may serve as a chemical “cue” that drives development of antibiotic resistance. Low concentrations of antibiotics have not yet been broadly described in reservoirs outside of the aforementioned environments, nor is the transfer and dissemination of antibiotic resistant bacteria and genes within natural microbial communities fully understood. This review will thus focus on low antibiotic-concentration environmental reservoirs and mechanisms that are important in the dissemination of antibiotic resistance to help identify key knowledge gaps concerning the environmental resistome.
Antibiotic resistance is a well-recognized major threat to global public health. Many studies have surveyed antibiotic resistant bacterial strains or their corresponding genes in hotspots or reservoirs of antimicrobial resistance such as hospitals, waste-water treatment plants (WWTP), sewage systems, and animal feeding operations, where antibiotics are commonly found at high concentrations (Bengtsson-Palme et al., 2018; Burcham et al., 2019; Kraemer et al., 2019; Maestre-Carballa et al., 2019; Xiang et al., 2020). In particular, attempts have been made to correlate the concentration of antibiotics with the occurrence of antibiotic resistant bacterial strains or genes in these hotspots (Rodriguez-Mozaz et al., 2015; Pärnänen et al., 2019).
The complete set of resistance-related genes in a particular system is termed the environmental resistome, and the determination of its risk has been proposed by Martínez et al. (2015), where antibiotic resistance genes (ARGs) are ranked depending on the associated public health risks they might pose (Bürgmann et al., 2018; Asante and Osei Sekyere, 2019). It is known that constant exposure of microbial communities to higher concentrations of antibiotics exerts a selective pressure on the environmental resistome (Ebmeyer et al., 2021), and thus it is also expected that constant exposure to “low-level” or sub-inhibitory concentrations of antibiotics may similarly play important roles in driving antibiotic resistance in environmental microbial communities such as freshwater biofilms and agriculture-impacted environments (Rodriguez-Mozaz et al., 2015; Abe et al., 2020).
Agricultural practices such as irrigation, soil fertilization and animal production are major sources of antibiotic resistance transmission to soil and water resistomes (Manaia et al., 2020) since intensive agriculture operations are more likely to carry ARGs, antibiotic-resistant bacteria (ARB) and run-off chemicals. Even though antibiotics from agriculture run-off can be found at sub-inhibitory concentrations in receiving water bodies or downstream systems, constant exposure over long time periods will contribute to selective pressure for antibiotic resistance in environmental microbial communities such as freshwater biofilms (Andersson and Hughes, 2012; Manaia et al., 2020; Sasikaladevi et al., 2020).
One often overlooked dimension of biofilms is their virome, or the complete assemblage of viral particles that exist within a particular biofilm consortium. The ecological roles of viruses in the spread of antibiotic resistance are not yet fully understood (Bekliz et al., 2019; Palermo et al., 2019). Though it is known that bacteriophage can spread antibiotic resistance or virulence genes via transduction, the role of bacterial viruses as potential vectors for environmental antibiotic resistance dissemination is still an ongoing debate (Parsley et al., 2010; Enault et al., 2017; Petrovich et al., 2020). Overall, more studies are required to resolve the complexity of environmental resistomes in pursuit of minimizing the spread of antibiotic resistance (Quinlan et al., 2011; Kraemer et al., 2019; White and Hughes, 2019).
Biofilms are ubiquitous throughout nature and may be found attached to any submerged surface or solid-liquid interface, such as stones, wood, shells, sediment, or on other living aquatic organisms (Lawrence et al., 2012, 2015). Occurring at the base of aquatic food chains, biofilms represent a key microbial compartment that serves vital roles for ecosystem functioning due to their biomass and diversity. The biofilm’s multiple trophic levels are formed by primary producers and heterotrophic prokaryotes and eukaryotes which commonly include bacteria, algae, archaea, fungi and Protista (Villanueva et al., 2011; Proia et al., 2013; Lawrence et al., 2015).
The structure of biofilms is highly variable and influenced by myriad factors, but typically characterized by microcolonies of bacterial cells encased in an extracellular polymeric substance (EPS) matrix and separated from other microcolonies by interstitial voids (water channels) (Caldwell et al., 1992, 1993; Lewandowski, 2000; Donlan et al., 2002). Besides polysaccharides, the biofilm EPS matrix also includes proteins, lipids, extracellular DNA and even inorganic solids (Flemming and Wingender, 2010). It can account for 50–90% of the total organic carbon of biofilms and some consider it to be the primary matrix material of the biofilm (Flemming and Wingender, 2010; Fulaz et al., 2019).
The biofilm “lifestyle” offers multiple potential benefits to member microorganisms, including enhanced nutrient uptake, limited diffusion of potentially deleterious compounds by the EPS matrix, stable positioning in optimal locations, beneficial interactions between complementary neighbors, and creation of chemically stable microniches (Lawrence et al., 2012, 2015; Orazi and O’Toole, 2019). The fast-growing and sessile nature of the microbial communities, as well as their dependence on environmental physical and chemical conditions, make them ideally suited as indicators of disturbances in river systems, from tracking increases in temperature to the presence of inorganic pollutant chemicals and materials from the surrounding environment (Romaní, 2009; Ramanan et al., 2016; Tu et al., 2020). Because of their autotrophic and heterotrophic metabolism, biofilm communities are major sites for the uptake, storage and transformation of fluvially-dissolved organic matter and nutrients, thus playing a major functional role in stream ecosystems (Romaní et al., 2004; Bott et al., 2006).
In addition to the EPS matrix providing structural rigidity and protection, environmental factors such as water flow velocity, availability and types of nutrients, temperature, suspended solids, and light availability contribute to shaping the three-dimensional structure (often termed “architecture”) of biofilm microcolonies and the abundance and nature of inhabiting bacteria (Lawrence et al., 1991, 2004, 2015; Mangalappalli-Illathu et al., 2008a; Wingender and Flemming, 2011; Tu et al., 2020), as well as the extent and thickness of biofilm development. The development of the biofilm may further be influenced by the growth of algae, cyanobacteria, protozoa and the EPS matrix, particularly the latter, as it has been shown to strongly impact the development of the biofilm’s microenvironmental conditions (Romaní et al., 2004, 2014; Flemming and Wingender, 2010).
Bacterial cells within biofilm communities have the capacity to respond, when challenged with stressors like antibiotic discharge (Schroeder et al., 2017), using various mechanisms. Importantly, pathogenic virulence factors and antibiotic or stress resistance factors share features that are involved in efflux pumps, porins, cell-wall alterations and two-component sensory systems that activate or repress the expression of genes or gene operons (Beceiro et al., 2013; Schroeder et al., 2017). ARGs are often co-located with pathogen virulence factors in biofilm-producing organisms, thereby aiding in the selection for, and spread of, antibiotic resistance in pathogenic organisms (Schroeder et al., 2017) and often leading to multiple drug resistance (MDR).
ARGs, which comprise the “resistome” of a given metagenome dataset (Maestre-Carballa et al., 2019) proliferate via horizontal gene transfer (HGT). The “mobilome,” or all gene elements related to HGT, is defined as all detectable horizontal mobile genetic elements (MGE) within a given metagenomic dataset (Slizovskiy et al., 2020) and includes plasmids, transposons, insertion sequences, bacteriophage, integrons and extracellular DNA (e-DNA). MGEs can be transferred between distantly related bacteria, including different phyla, and therefore do not tend to carry genes essential for cell metabolism or survival (Wellington et al., 2013). Recently, Partridge et al. (2018) published an extensive review on MGEs in clinically relevant species and their association with antimicrobial resistance, which we recommend readers to review.
Due to its importance in clinical infections and tendency to form biofilms, Pseudomonas aeruginosa has often been used as a model for the study of antibiotic resistance and tolerance in biofilms. Therefore, most of the molecular mechanisms of antibiotic resistance known today are based on in vitro studies first benchmarked using P. aeruginosa biofilms (Brooun et al., 2000; Hall and Mah, 2017). In addition, Gram-negative bacteria are intrinsically more resistant to antibiotics (i.e., vancomycin) than Gram-positives, due to the relative impermeability of the Gram-negative cells’ outer membrane (Hall and Mah, 2017). Conjugation, presumably the principal route of HGT in sessile bacterial communities, has been documented many times in hospital studies (Lerminiaux and Cameron, 2019; Abe et al., 2020). Higher rates of e-DNA transformation also occurs in biofilms, with this e-DNA being concentrated within the biofilm EPS (Madsen et al., 2012). For example, Lehmann et al. (2016) recorded the prevalence of MGE in freshwater biofilms after sub-minimum inhibitory concentrations (sub-MIC) exposure to treated sewage effluent (2.5 ppm), and determined that biofilms which were exposed to sewage effluent had significantly higher prevalence of Class 1 integrons (Table 1). Transfer of resistance elements may also be carried out via transduction by phage, though this mechanism is notably underexplored in biofilm communities.
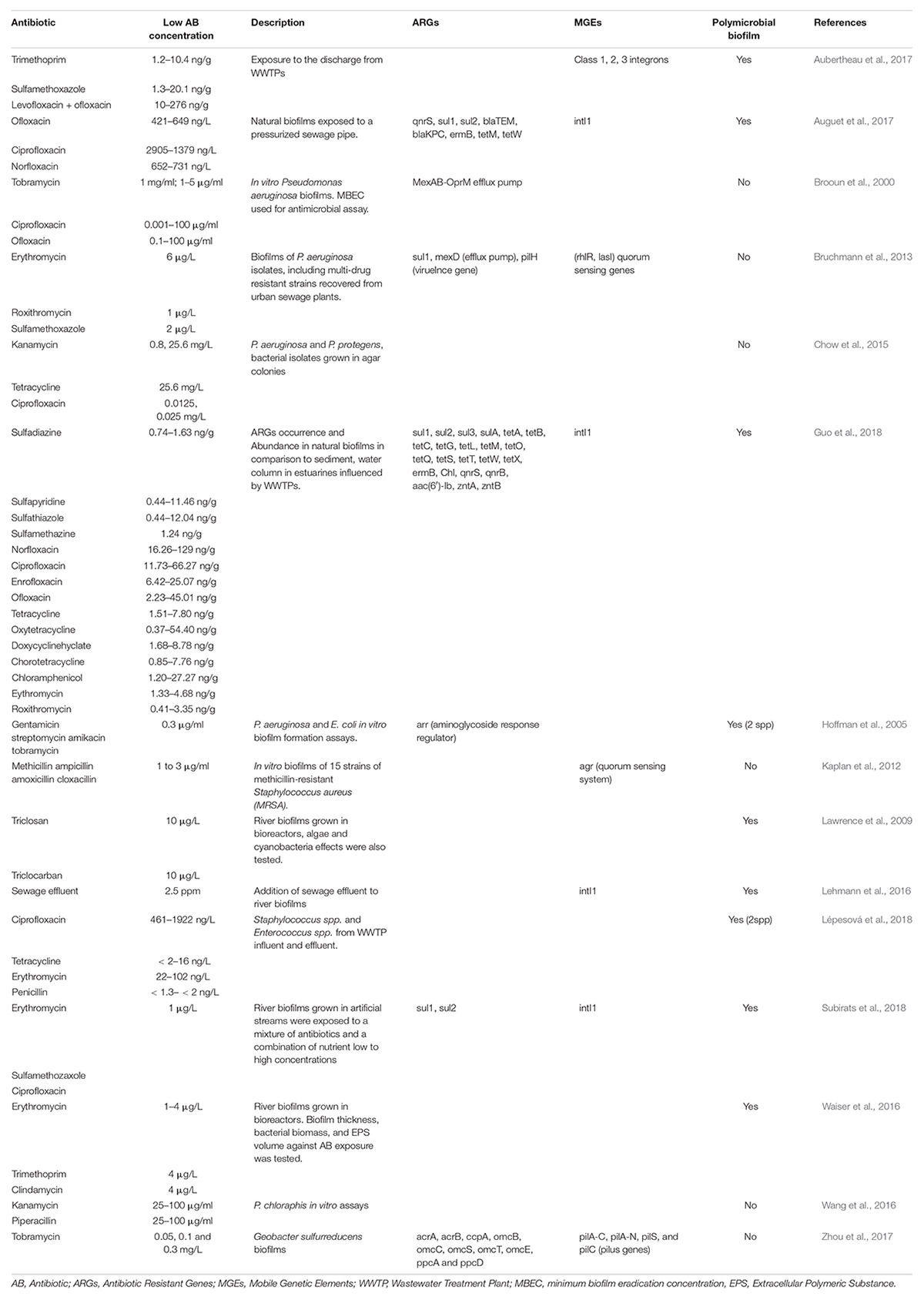
Table 1. Examples of biofilms exposed to low antibiotic concentrations.
In addition to genetic mechanisms of resistance, high heterogeneity and low permeability of biofilms allow for tolerance mechanisms which include persister cell development, altered cell permeability, and efflux pump regulation (Hall and Mah, 2017; Schroeder et al., 2017). The biofilm matrix and EPS do provide protection against exposure to a sustained stress by sub-MICs of antibiotics, and can offer an adaptive advantage to bacteria within biofilms by limiting the penetration of chemical stressors into deeper regions of the biofilm (Szomolay et al., 2005; Mangalappalli-Illathu et al., 2008b). Bacterial biofilms have also been shown to contain intrinsically resistant persister cells at a level of 1% of the total population that exhibit multi-drug and bactericidal agent tolerance (Lewis, 2005; Gebreyohannes et al., 2019). These dormant or slow-growing cells are generally identical to the rest of the population but display several phenotypic differences, such as resistance to normally lethal concentrations of antibiotics (Stewart and Costerton, 2001; Gebreyohannes et al., 2019; Jackson et al., 2019; Orazi and O’Toole, 2019). In contrast to antibiotic-resistant cells, persister cells exhibit temporary antibiotic-resistant phenotypes which have been found to last as long as 4 weeks (Miyaue et al., 2018).
Recently, two alternate HGT mechanisms facilitated by the biofilm structure were proposed (Abe et al., 2020); membrane vesicles and nanotubes, which are suggested to enable translocation of various cellular materials (possibly including ARGs) between adjacent biofilm bacteria. Extracellular membrane vesicles consist of 20 to 400 nm lipid-bilayer capsules produced and released by both Gram-positive and Gram-negative bacteria. They are ubiquitous in aquatic environments and have been reported in biofilms of P. aeruginosa (Murphy et al., 2014), Escherichia coli (Nakao et al., 2018) and Vibrio cholerae (Altindis et al., 2014). These outer membrane-derived vesicles have the potential to transport DNA, as well as RNA, proteins, metabolites and QS molecules (Abe et al., 2020). Nanotubes are membranous extracellular structures that can elongate to allow direct cell-to-cell contact. Unlike pili used in conjugation, nanotubes can transfer cytoplasmic material such as nutrients and DNA, thus they also have the potential to transfer ARGs to other bacteria. However, there is no current, direct evidence that demonstrates the transfer of ARGs through these two mechanisms in natural biofilms (Abe et al., 2020).
Biofilm cells also engage in QS, a process that enables bacteria to communicate via secreted signaling molecules called autoinducers. This process enables a population of bacteria to regulate gene expression collectively, and therefore control behavior on a community-wide scale. QS autoinducer signaling has also been associated with conjugation, phage induction and transformation (Abe et al., 2020). Antibiotics can act as signals for QS attenuators and there is evidence that QS controls the secretion of virulence factors, and formation of biofilms and conjugation, among other activities (Miller and Bassler, 2001). Vasudevan et al. (2018) describe more than 20 QS inhibitors (i.e., furanone, baicalein, lactonase, etc.) and their synergetic actions with antibiotics. In the absence of QS inhibitors, the biofilm develops a barrier and antibiotics are less able to enter the cell which leads to drug tolerance (Orazi and O’Toole, 2019).
Most studies analyzing the prevalence of ARGs in hotspots of antimicrobial resistance focus on a single species or clinically relevant strains tested in vitro (Høiby et al., 2010; Partridge et al., 2018). While the information extracted from these in vitro experiments is relevant, its noteworthy to remember environmental biofilms are typically formed by myriad species, and the effect after exposure to antibiotic residues or antibiotic-resistant bacteria would likely differ significantly from that of mono-species biofilms. Interactions between the biofilm community and other organisms can range from the biofilm acting as a barrier to allochthonous, incoming organisms, to biofilms serving as a source of diversity which expands membership and adds heterogeneity to key microbial components of the aquatic food chain (Balcázar et al., 2015).
Resistance assays also often simply demonstrate growth in the presence (or absence) of an antibiotic using traditional approaches (e.g., MIC). The ability of sub-MIC levels to act selectively on a population of organisms (i.e., biofilm) is currently not well understood in the context of innate vs. acquired resistance. Innate resistance refers to wildtype genes encoding antibiotic resistance or innate properties of the cell, whereas acquired resistance is conferred following chromosomal mutations or the acquisition of resistance genes through HGT (Fernández and Hancock, 2012; Hall and Mah, 2017; Schroeder et al., 2017). Both mechanisms are known features of biofilm communities, and as such contribute toward the development, maintenance and transmission of ARGs and general resistance (Hall and Mah, 2017). It is clear that more community-based exposure experiments conducted under controlled conditions are needed to provide meaningful data to broaden our understanding of how complex communities respond to stresses induced by very low concentrations of antibiotics.
It is widely accepted that bacteria living in biofilms are more resistant to chemical, physical and mechanical stresses than their planktonic counterparts (Li et al., 2001; Szomolay et al., 2005). The EPS matrix provides protection for the community from chemicals such as antibiotics, antivirals, pharmaceuticals, personal care products, disinfectants, heavy metals, pesticides and drugs (Mangalappalli-Illathu et al., 2008a; Høiby et al., 2010; Balcázar et al., 2015; Guo et al., 2018; Abe et al., 2020). Furthermore, genetic and physiological tolerance mechanisms induced by co-selection of metal and antibiotic resistance factors has been attributed to the sequestration properties of the biofilm matrix and the role of persister cells (Baker-Austin et al., 2006). Indeed, analysis of the occurrence and abundance of ARGs in naturally occurring biofilms in comparison to ARGs found in estuarine sediment and water exposed to antibiotics (sulfonamides, fluoroquinolones, tetracyclines, macrolides, and chloramphenicols) has confirmed that biofilms act as sinks for both types of contaminants (Guo et al., 2018).
Antibiotics in natural environments, including river biofilms, generally migrate from high concentration source reservoirs in the aqueous phase (and decrease with distance from those sources), enter wastewater systems, or directly impact pristine environments (Balcázar et al., 2015; Lehmann et al., 2016; Aubertheau et al., 2017; Grenni et al., 2018). The constant exposure of downstream microbial communities to low concentrations of antibiotics may influence the structure of microbial biofilm communities, and impose selection pressure for antimicrobial resistance, along with other collateral effects on ecosystem diversity (Quinlan et al., 2011; Kraemer et al., 2019; White and Hughes, 2019). Indeed, previous studies have found the total antibiotic concentration in biofilms was 2.3–5.8 times higher (51.7 ng/g–362.4 ng/g) than for sediment samples, with the abundance of ARGs such as intI1, sul1, sul2, aac(6′)-Ib, tetA, tetW was highest in biofilm samples, followed by sediment and water, respectively, (Guo et al., 2018).
It should be noted that recent studies suggest that the dispersal of ARGs or ARB from hotspots, including animal farms, are far more significant contributors to the environmental dissemination of ARGs than is in situ natural environmental selection (Brandt et al., 2015; Karkman et al., 2018; Zhao et al., 2020). Nevertheless, there is currently no consensus on the risk associated with sub-inhibitory concentrations of antibiotics regarding their abilities to select for antibiotic resistance and increase rates of bacterial evolution (Gillings and Stokes, 2012; Karkman et al., 2018; Zhao et al., 2020). Given the frequent domestic use of antibiotics, river biofilm communities are continuously exposed to these stressors. Understanding the ecological impact of long-term exposure of sub-inhibitory concentrations on a natural biofilm requires experimental knowledge of antibiotic concentration, exposure time, biofilm’s spatial distribution, biochemistry of the sorption sites, and the impact on biofilm structure, species abundance, and viability (Balcázar et al., 2015).
Although there is no single assay that is universally used to assess antimicrobial susceptibility in biofilms, plate-based phenotypic assays offer convenient tools for antimicrobial testing [e.g., the minimum biofilm eradication concentration (MBEC) assay]. Biofilm variants of this approach can also be used to determine the minimum inhibitory concentration (MIC) or the minimum biocidal concentration (MBC) (Hall and Mah, 2017; Thieme et al., 2019). The MIC refers to the lowest drug concentration that inhibits visible growth of target bacterial populations, and it is typically measured in planktonic mono-species cultures or on semi-solid agar surfaces (i.e., Kirby-Bauer disc diffusion). Antibiotic concentrations below the MIC are referred to as sub-inhibitory concentrations (sub-MICs), or minimal selective concentrations (MSC) (Gullberg et al., 2011). In surface waters worldwide, concentrations of antibiotics from treated sewage effluents and animal wastes tend to occur in the ng/L to mg/L range (Waiser et al., 2016). Although mg/L concentrations are considered very high, most aquatic systems have antibiotics present at ng/L concentrations (Chow et al., 2021). Furthermore, many antibiotics are excreted from animals and humans in a chemically active and environmentally persistent form, being detected downstream of wastewater treatment plants and adjacent to fields receiving animal manures (Finley et al., 2013).
It is estimated that the MSC for a diverse range of microorganisms ranges between 1/4 and 1/230 of the MIC values (Gullberg et al., 2011). It is also known that antibiotic concentrations 10–100 times lower than MIC values can increase the relative abundance of resistant bacteria and select for their resistance by accelerating the rate of adaptive evolution under in vitro conditions (Gullberg et al., 2011; Friman et al., 2015; Lundström et al., 2016; Danner et al., 2019). For example, sub-MIC antibiotic exposure has been shown to increase genetic diversity in microbial populations via the action of the bacterial SOS response, resulting in an increased mutation rate throughout the genome and via direct mutagenic effects on the DNA (Allen et al., 2010; Kraemer et al., 2019). Sustained MSC exposure can also select for the most resistant species (Romero et al., 2019), thereby altering both structure and function of the biofilm. For instance, the antimicrobials triclosan and triclocarban at concentrations of 10 μg/L for an 8-week period were shown to be highly toxic to autotrophs (algae and cyanobacteria), shifting river biofilms from an autotrophic community to one dominated by heterotrophs (Lawrence et al., 2009).
In WWTP systems, comparisons between influent and effluent biofilms show higher prevalence of antibiotic-resistant bacteria in effluent biofilms (Lépesová et al., 2018). Influent biofilms contained only vancomycin-resistant enterococci, while in effluent biofilms, resistance to 17 antibiotics was observed, with most Enterococci showing multi-drug resistance (MDR). Sub-MIC antibiotic exposures also have demonstrated effects on functional capacity of aquatic biofilms; efforts to profile the responses in biofilms exposed to sub-MICs of antibiotics have been performed downstream of WWTP discharges (Table 1). One such study which profiled the response of P. aeruginosa biofilms (including MDR strains) to sub-MIC concentrations of erythromycin (6 μg/L), roxithromycin (1 μg/L), and sulfamethoxazole (2 μg/L), revealed changes in biofilm dynamics that included biomass formation, spatial structure and expression of specific genes in different P. aeruginosa isolates (Bruchmann et al., 2013). Increases in quorum sensing (QS)-regulating gene activities were also observed after sub-MIC antibiotic exposure, with macrolides (i.e., erythromycin, roxithromycin) specifically upregulating QS-genes (rhlR, lasI). In contrast, the presence of sulfamethoxazole caused upregulation of sul1 and the efflux pump, mexD, confirming the involvement of efflux pumps in sulfonamide resistance (Bruchmann et al., 2013). In many other cases, exposure to low concentrations of antibiotics were associated with mobile genetic element-mediated dissemination of ARGs through HGT (Celli and Trieu-Cuot, 1998; Bruchmann et al., 2013; Finley et al., 2013).
Interestingly, inlet and outlet samples of WWTP systems have demonstrated reductions in the copy numbers of ARGs after water treatment processes (Auguet et al., 2017), with authors quantifying ARGs sul1 and sul2 as the most abundant in both biofilms and water columns, with roughly 1 resistance gene for each 10 copies of 16 s RNA gene. Moreover, significant differences were observed for intI1 (gene encoding integrase of Class I integrons) between biofilms found at sewage outlet (1.60 log) and inlet (1.16 log) locations. Class 1 integrons are often physically linked to multiple resistance determinants for antibiotics, and thus used as a proxy for ARGs of anthropogenic origin (Zhao et al., 2020). These results thus confirm biofilms as a reservoir for mobile genetic elements. Constant exposure to antibiotics over a range of sub-MICs would thus be expected to encourage HGT (Kraemer et al., 2019) and ARB development.
Comparison between resistomes exposed to higher concentrations of antibiotics (e.g., WWTPs) and lower concentrations of antibiotics (e.g., receiving water bodies of agriculture run-off, or other down-gradient systems) require further study to corroborate the transfer and acquisition of antibiotic resistance under these conditions (Mao et al., 2015; Ebmeyer et al., 2021). Additionally, studies on the emergence of MDR and virulence factors should continue to be undertaken to better understand the risk and spread of these factors in environmental resistomes.
In addition to the release of antibiotic residues via the discharge of effluent from WWTPs, agriculture waste products and associated field run-off following application of animal waste as source of fertilizer represents a second major contributor to environmental antibiotic resistance (Topp et al., 2018; Goulas et al., 2020; Henriot et al., 2021). The demand for animal protein has been increasing over the last decade and global trends indicate intensive animal production will continue to increase the occurrence of ARB (Tiseo et al., 2020; Wang et al., 2021). It is estimated that 73% of all antimicrobials sold globally are used to raise animals for human consumption (Van Boeckel et al., 2019). While many countries have legislated tighter controls over antibiotic use (Zhao et al., 2020), others continue to permit prophylactic antibiotic usage in animal operations (Finley et al., 2013; Kraemer et al., 2019).
It is abundantly clear that exposure of microorganisms to veterinary/agricultural antibiotic residues can contribute to the development of ARGs and ARB (Manaia et al., 2020; Wang et al., 2021); however, little is known about the selective pressure that sub-MIC concentrations of antibiotics play on initial microbial colonization processes as well as biofilm development in receiving environments along with the environmental resistome (Yang et al., 2020). The environmental resistome influenced by the agriculture sources is characterized by two processes: extended periods of selective pressure imposed by antibiotic residues, and the subsequent dispersion of ARGs or ARB (Zhao et al., 2020). Dispersion is enhanced by the incomplete biodegradation of antibiotics, as the half-life of antibiotics in manure are estimated to be between 2 and 100 days. Released antibiotics are chemically diverse and degrade at different rates in the environment via sorption, photodegradation oxidation, and biodegradation (Grenni et al., 2018), making it difficult to predict how quickly they degrade and the extent to which they might continue to exert selective pressure for ARB development (O’Neill, 2016; Manyi-Loh et al., 2018).
It is noteworthy that unimpacted environmental resistomes deemed to be “pristine” with respect to anthropogenic activities, are increasingly difficult to find (Hooban et al., 2020). Widespread environmental pollution from antibiotic production facilities or agricultural operations has necessitated the call to monitor wildlife, agriculture, aquaculture systems, along with supposedly unimpacted environmental sites (Khan et al., 2013; Wellington et al., 2013; Thakur and Gray, 2019). In agricultural systems, veterinary drugs reach the surrounding environments, often in a metabolically-active form, through distribution and disposal of livestock manure and urine (Zhao et al., 2020). Manure is a common fertilizer used in farmland soils and crop production, and due to its potential to contribute to ARG dissemination to soils, aquatic systems, including plant microbiomes, it is a practice of ongoing concern (Zhao et al., 2020).
With regard to agriculture and aquaculture practices, 51 antibiotics have been reported as being in use according to the World Health Organization (WHO, 2017), including the six most-common classes of antibiotics: tetracyclines, quinolones, aminoglycosides, polymyxins, macrolides, penicillins, and sulfonamides (Chekabab et al., 2021). It is important to note that antibiotic usage for humans and animals often occurs within similar drug classes (e.g., gentamicin/aminoglycoside, ampicillin/β-lactams, ciprofloxacin/fluoroquinolones, trimethoprim/antifolate, and tetracycline) (Finley et al., 2013; EUCAST, 2019); thus, the potential for cross-resistance to those drugs used in either setting exists. For instance, in soil microbiomes exposed to manure (Han et al., 2018), 163 ARGs were identified in untreated soils whereas 245, 230, and 245 ARGs were detected in soils treated with manure from poultry, swine and cattle, respectively. Of the total ARGs, the highest abundance was found to encode β-lactamases (19.6%), multi-drug resistance (18.8%) and macrolides (15.4%).
Tetracyclines and macrolides have historically seen extensive use in agriculture (Ebmeyer et al., 2021) and ARGs conferring resistance to these antibiotics are among the most prevalent (Chekabab et al., 2020, 2021). In China, most of the veterinary antibiotics consist of tetracyclines and sulfonamides (Zhou et al., 2020). Manure, soil and water samples of sixteen animal farms in Southeastern China were sampled to determine the abundance of ARGs (Wang et al., 2016); 22 ARGs were found to confer resistance to five major classes of antibiotics including tetracyclines, sulfonamides, quinolones, aminoglycosides, and macrolides. The author’s results showed that the spread of the agriculture-impacted resistome was dominated by sul genes, which were the most extensive, followed by tet and erm genes.
Metals can also act as co-selective agents driving the enrichment of genetic elements which contain resistance genes to both metals and ARGs (Baker-Austin et al., 2006; Zhao et al., 2020). The co-selection of metals and ARGs has been recorded in agriculture-impacted resistomes (Zhao et al., 2020). In some agricultural settings, Cu, As, and Zn continues to be used as supplementary feed additives for animal growth promotion and disease prevention (Poole, 2017).
Opportunistic bacterial pathogens such as Aeromonas spp., Citrobacter spp., Enterobacter spp., Mycobacterium spp., Legionella spp., Staphylococcus spp., Enterococcus spp., P. aeruginosa, and K. pneumoniae naturally occur in aquatic and soil environments and can persist and grow in biofilms of drinking water systems (Schwartz et al., 2003; Obst et al., 2006; Wingender and Flemming, 2011; Ahmad et al., 2021). Agricultural practices, such as irrigation, as well as drinking water distribution systems in which biofilms are known to form, have also been suggested to act as reservoirs for pathogenic microorganisms (Wang et al., 2021). As biofilms develop, resistant bacteria can be released to continue their life cycle as planktonic cells, and subsequently colonize, or disperse to, other ecosystems carrying their ARG compliment. Additionally, pathogenic bacteria such as P. aeruginosa and Legionella pneumophila that are resistant to an antibiotic can survive sewage treatments and migrate downstream to eventually be included in a biofilm (Wingender and Flemming, 2011). Most bacterial populations inhabiting freshwater biofilms are non-pathogenic. However, non-pathogenic bacteria have a higher chance of interacting with pathogenic bacteria in high-density systems such as biofilms (Wingender and Flemming, 2011).
Veterinary antibiotic usage over the last several years have been reduced worldwide as result of greater awareness of the threat of antibiotic resistance (Kraemer et al., 2019); however, agricultural antibiotic use still remains a major source of environmental antibiotic pollution (Wang et al., 2021). Overall, the effects of sub-MIC antibiotics outside clinical settings have not yet been as fully explored nor monitored possibly due to detection difficulties pose by low effective concentrations. However, recent studies support the premise that acquisition of resistance can occur at sub-MIC antibiotic concentrations (Marti et al., 2014; Chow et al., 2015, 2021). Clearly, more experimental data is required to further comprehend the risk of exposed to low dosages of antimicrobials, as well as the response interactions from the entire microbial community and not just that of individual bacteria.
Over the past decade, studies of the ecology bacterial viruses (bacteriophage) have become more feasible thanks to the increased accessibility of high-throughput sequencing methods. However, a clear understanding of the ecological role of bacteriophage in aquatic environments, including freshwater biofilms, is still lacking (Battin et al., 2016; Bekliz et al., 2019; Palermo et al., 2019). This lack of knowledge of environmental phage is compounded by the inability to characterize the majority of bacterial viruses, with over 65% of viral sequences from surface waters being unrecognized when compared to existing databases (Palermo et al., 2019).
Flood and Ashbolt (2000) observed that wetland biofilms could entrap viral-sized particles and concentrate them over 100-fold, compared to suspended numbers in the surrounding water column. Further, in the different stages of WWTPs, phage have been observed at concentrations ranging from 108 to 1010 ml–1, which is 10–1,000 times higher than in natural (unimpacted) aquatic environments, indicating WWTPs as important reservoirs of bacteriophage (Wu and Liu, 2009). While bacteriophage do not represent a direct threat to human or domestic animals, they shape the diversity and genetic architecture of their hosts (Chopyk et al., 2020). Since there are records of phage invading biofilms by disrupting the EPS matrix or by killing embedded cells (Sutherland et al., 2004), it was originally believed that biofilm formation and phage transmission were mutually exclusive (Abe et al., 2020). However, more recent work has described phage as playing supporting beneficial roles in high density microbial colonies such as biofilms, as cell lysis mediated by phage infection often leads to production of e-DNA. The e-DNA released by phage may then be integrated by the biofilm’s matrix, thus increasing the opportunity for acquisition of resistance genes through HGT (Fernández et al., 2018).
Most studies of bacteriophage composition have focused on WWTP microbial communities, where the order Caudovirales comprises most of the viral sequences, with dominant viral families including Myoviridae, Siphoviridae, and Podoviridae (Parsley et al., 2010; Allen et al., 2011; Cantalupo et al., 2011; Tamaki et al., 2012; Chopyk et al., 2020; Petrovich et al., 2020). A similar virome composition was observed in benthic biofilms of three different freshwater streams, where Siphoviridae, Podoviridae and Myoviridae were the dominant viral taxa (Bekliz et al., 2019). Moreover, it was observed that ARGs such as sul1, blaTEM and bla CTX–M, were more persistent after WWTP treatments (chlorination, thermal treatment and natural inactivation) when more of these ARGs were found within the protein capsid of phage particles than found within bacteria (Calero-Cáceres and Muniesa, 2016). It is thought that the protein capsid could provide DNA fragments with enough protection to survive various environmental stressors, thereby serving as a significant reservoir. Furthermore, the possibility of ARGs being transferred to a suitable host would also increase under such a scenario (Calero-Cáceres and Muniesa, 2016).
One study that described both the bacterial and viral composition of the swine gut identified ARGs such as MDR efflux pumps [bcrA, macB, mef(A)] within the virome and that the abundance of phage integrase-encoding genes were significantly increased in the viromes of medicated swine relative to non-medicated swine, indicating induction of bacteriophage in the presence of antibiotics (Allen et al., 2011). However, the authors did not observe significant changes in bacteriophage abundance, suggesting that while they were induced in the swine gut, this did not result in the transfer of ARGs via prophage to bacteria in the swine gut. In contrast, a study by Zhang and LeJeune (2008) demonstrated that blaCMY-2, tetA and tetB were transferred via phage between Salmonella serovars of bovine origin, providing evidence of ARG propagation by phage-mediated transduction events.
Bacteriophage are becoming increasingly recognized for their potential role in the dissemination of antimicrobial resistance, as well as other genes that shape bacterial community composition (i.e., virulence factors, auxiliary metabolic genes) (Colavecchio et al., 2017; Breitbart et al., 2018; Chopyk et al., 2020). Evidence suggests that dissemination of ARGs by bacteriophage may be possible between different, and even remote, environments not contaminated by antibiotics as a result of anthropogenic activities (Muniesa et al., 2013). Just as exposure to antibiotics leads to selective pressure on susceptible bacteria, the constant presence of phage and/or competing viruses may exert a selective pressure on microbial communities, influencing the prevalence and abundance of antibiotic resistant genes (Madsen et al., 2012; Beceiro et al., 2013). In studies that compared viral and bacterial taxonomic composition, dsDNA phage were the most abundant of the viral members observed, though it was noted that virome studies targeting bacteriophage, and the availability of sequenced viromes, is currently limited (Calero-Cáceres et al., 2019).
There is some evidence to suggest viral taxa may not have a great impact on ARG transfer via transduction (Colavecchio et al., 2017; Enault et al., 2017). However, transduction tends to be more common in sites with high viral and bacterial density (e.g., WWTPs) (Petrovich et al., 2020), suggesting a different pattern for biofilms where their high microbial diversity and density could possibly enable them to be more susceptible to viral-ARG associations. Nonetheless, there is an ongoing debate on whether viruses are associated with antimicrobial resistance spread and if they contribute significantly to HGT (Parsley et al., 2010; Enault et al., 2017; Petrovich et al., 2020). Notably, very few studies have compared the dynamics of bacterial and viral communities in biofilms. Moreover, few studies have attempted to correlate the effects of antiviral drugs in combination with antibiotics on microbial communities, and monitoring of antivirals in wastewaters, surface waters, groundwater, and drinking water is not currently conducted. However, some antivirals are highly bioactive and are proposed to negatively affect non-target organisms and persist in aquatic environments (Jain et al., 2013; Sims and Kasprzyk-Hordern, 2020). For this reason, it is important to investigate the simultaneous occurrence of antiviral drugs with antibiotics.
Further research is clearly needed to understand associations between bacterial and viral taxa, particularly in critical environmental niches (e.g., biofilms) to better address whether viruses contribute to the spread or transfer of ARG. More insight into the relations of bacteriophage and the prevalence of antimicrobial resistance would provide data for improved methods for effluent water treatment, better comprehension of the interactions between viromes and sub-MIC-exposed environmental resistomes, and offer a new framework for monitoring strategies to mitigate the spread of pathogenic viruses and antimicrobial resistance.
While many studies of ARG and ARB spread have focused on hotspots where high antibiotic concentrations are released (e.g., hospitals, WWTPs and large animal production facilities), the majority of downstream environments are routinely exposed to much lower concentrations of these metabolically-active agents. The extent to which these more prevalent antibiotic contaminants drive proliferation of antibiotic resistance in the environmental resistome remains to be fully elucidated. Interactions of antiviral and antibiotic drugs between the microbial community of pristine (unimpacted) environmental reservoirs are complex and also poorly understood, although seemingly offer myriad non-mutually exclusive events with potential to impact the persistence and spread of antibiotic resistance.
Most environmental resistomes are becoming increasingly influenced by the constant selective pressure from human activities, and in particular, from industrialized agriculture practices. In this review, we have focused on the effects of low antibiotic concentration exposure of biofilms colonizing freshwater environments and as a result of agricultural practices given the growing evidence suggesting sub-MIC selectively influences the prevalence of ARGs and ARB. Furthermore, the rich microbial diversity of biofilms commonly includes high concentrations of viruses, and in particular, bacteriophage, which are a potentially important and under-appreciated mechanism of antibiotic resistance propagation. The environmental virome as an integrated part of the environmental resistome, and specifically the bacteriophage’s role in the environmental transfer of antimicrobial resistance, requires more study to better understand the role of the environment in antibiotic resistance transmission.
GF-V: writing—original draft. JB: supervision and writing—review and editing. JRL: writing—review and editing. DRK: supervision, writing—review and editing, and funding acquisition. All authors contributed to the article and approved the submitted version.
This study was supported by NSERC discovery grant funding to DRK.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abe, K., Nomura, N., and Suzuki, S. (2020). Biofilms: hot spots of horizontal gene transfer (HGT) in aquatic environments, with a focus on a new HGT mechanism. FEMS Microbiol. Ecol. 96:fiaa031. doi: 10.1093/femsec/fiaa031
Ahmad, J. I., Dignum, M., Liu, G., Medema, G., and van der Hoek, J. P. (2021). Changes in biofilm composition and microbial water quality in drinking water distribution systems by temperature increase induced through thermal energy recovery. Environ. Res. 194:110648. doi: 10.1016/j.envres.2020.110648
Allen, H. K., Donato, J., Wang, H. H., Cloud-Hansen, K. A., Davies, J., and Handelsman, J. (2010). Call of the wild: antibiotic resistance genes in natural environments. Nat. Rev. Microbiol. 8, 251–259. doi: 10.1038/nrmicro2312
Allen, H. K., Looft, T., Bayles, D. O., Humphrey, S., Levine, U. Y., Alt, D., et al. (2011). Antibiotics in feed induce prophages in swine fecal microbiomes. mBio 2:e00260-11. doi: 10.1128/mBio.00260-11
Altindis, E., Fu, Y., and Mekalanos, J. J. (2014). Proteomic analysis of Vibrio cholerae outer membrane vesicles. Proc. Natl. Acad. Sci. U.S.A. 111, E1548—E1556. doi: 10.1073/pnas.1403683111
Andersson, D. I., and Hughes, D. (2012). Evolution of antibiotic resistance at non-lethal drug concentrations. Drug Resist. Updat. 15, 162–172. doi: 10.1016/j.drup.2012.03.005
Asante, J., and Osei Sekyere, J. (2019). Understanding antimicrobial discovery and resistance from a metagenomic and metatranscriptomic perspective: advances and applications. Environ. Microbiol. Rep. 11, 62–86. doi: 10.1111/1758-2229.12735
Aubertheau, E., Stalder, T., Mondamert, L., Ploy, M. C., Dagot, C., and Labanowski, J. (2017). Impact of wastewater treatment plant discharge on the contamination of river biofilms by pharmaceuticals and antibiotic resistance. Sci. Total Environ. 579, 1387–1398. doi: 10.1016/j.scitotenv.2016.11.136
Auguet, O., Pijuan, M., Borrego, C. M., Rodriguez-Mozaz, S., Triadó-Margarit, X., della Giustina, S. V., et al. (2017). Sewers as potential reservoirs of antibiotic resistance. Sci. Total Environ. 605–606, 1047–1054. doi: 10.1016/j.scitotenv.2017.06.153
Baker-Austin, C., Wright, M. S., Stepanauskas, R., and McArthur, J. V. (2006). Co-selection of antibiotic and metal resistance. Trends Microbiol. 14, 176–182. doi: 10.1016/j.tim.2006.02.006
Balcázar, J. L., Subirats, J., and Borrego, C. M. (2015). The role of biofilms as environmental reservoirs of antibiotic resistance. Front. Microbiol. 6:1216. doi: 10.3389/fmicb.2015.01216
Battin, T. J., Besemer, K., Bengtsson, M. M., Romani, A. M., and Packmann, A. I. (2016). The ecology and biogeochemistry of stream biofilms. Nat. Rev. Microbiol. 14, 251–263. doi: 10.1038/nrmicro.2016.15
Beceiro, A., Tomás, M., and Bou, G. (2013). Antimicrobial resistance and virulence: a successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 26, 185–230. doi: 10.1128/cmr.00059-12
Bekliz, M., Brandani, J., Bourquin, M., Battin, T. J., and Peter, H. (2019). Benchmarking protocols for the metagenomic analysis of stream biofilm viromes. PeerJ 7:e8187. doi: 10.7717/peerj.8187
Bengtsson-Palme, J., Kristiansson, E., and Larsson, D. G. J. (2018). Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 42, 68–80. doi: 10.1093/femsre/fux053
Bott, T. L., Montgomery, D. S., Arscott, D. B., and Dow, C. L. (2006). Primary productivity in receiving reservoirs: links to influent streams. J. North Am. Benthol. Soc. 25, 1045–1061.
Brandt, K. K., Amézquita, A., Backhaus, T., Boxall, A., Coors, A., Heberer, T., et al. (2015). Ecotoxicological assessment of antibiotics: a call for improved consideration of microorganisms. Environ. Int. 85, 189–205. doi: 10.1016/j.envint.2015.09.013
Breitbart, M., Bonnain, C., Malki, K., and Sawaya, N. A. (2018). Phage puppet masters of the marine microbial realm. Nat. Microbiol. 3, 754–766. doi: 10.1038/s41564-018-0166-y
Brooun, A., Liu, S., and Lewis, K. (2000). A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 44, 640–646. doi: 10.1128/AAC.44.3.640-646.2000
Bruchmann, J., Kirchen, S., and Schwartz, T. (2013). Sub-inhibitory concentrations of antibiotics and wastewater influencing biofilm formation and gene expression of multi-resistant Pseudomonas aeruginosa wastewater isolates. Environ. Sci. Pollut. Res. 20, 3539–3549. doi: 10.1007/s11356-013-1521-4
Burcham, Z. M., Schmidt, C. J., Pechal, J. L., Brooks, C. P., Rosch, J. W., Benbow, M. E., et al. (2019). Detection of critical antibiotic resistance genes through routine microbiome surveillance. PLoS One 14:e0213280. doi: 10.1371/journal.pone.0213280
Bürgmann, H., Frigon, D., Gaze, W. H., Manaia, C. M., Pruden, A., Singer, A. C., et al. (2018). Water and sanitation: an essential battlefront in the war on antimicrobial resistance. FEMS Microbiol. Ecol. 94:fiy101. doi: 10.1093/femsec/fiy101
Caldwell, D. E., Korber, D. R., and Lawrence, J. R. (1992). Imaging of bacterial cells by fluorescence exclusion using scanning confocal laser microscopy. J. Microbiol. Methods 15, 249–261. doi: 10.1016/0167-7012(92)90045-6
Caldwell, D. E., Korber, D. R., and Lawrence, J. R. (1993). Analysis of biofilm formation using 2D vs 3D digital imaging. J. Appl. Bacteriol. 74, 52S–66S. doi: 10.1111/j.1365-2672.1993.tb04342.x
Calero-Cáceres, W., and Muniesa, M. (2016). Persistence of naturally occurring antibiotic resistance genes in the bacteria and bacteriophage fractions of wastewater. Water Res. 95, 11–18. doi: 10.1016/j.watres.2016.03.006
Calero-Cáceres, W., Ye, M., and Balcázar, J. L. (2019). Bacteriophages as environmental reservoirs of antibiotic resistance. Trends Microbiol. 27, 570–577. doi: 10.1016/j.tim.2019.02.008
Cantalupo, P. G., Calgua, B., Zhao, G., Hundesa, A., Wier, A. D., Katz, J. P., et al. (2011). Raw sewage harbors diverse viral populations. mBio 2:e00180-11. doi: 10.1128/mBio.00180-11
Celli, J., and Trieu-Cuot, P. (1998). Circularization of Tn916 is required for expression of the transposon-encoded transfer functions: characterization of long tetracycline-inducible transcripts reading through the attachment site. Mol. Microbiol. 28, 103–117. doi: 10.1046/j.1365-2958.1998.00778.x
Chekabab, S. M., Lawrence, J. R., Alvarado, A., Predicala, B., and Korber, D. R. (2020). A health metadata-based management approach for comparative analysis of high-throughput genetic sequences for quantifying antimicrobial resistance reduction in Canadian hog barns. Comput. Struct. Biotechnol. J. 18, 2629–2638. doi: 10.1016/j.csbj.2020.09.012
Chekabab, S. M., Lawrence, J. R., Alvarado, A. C., Predicala, B. Z., and Korber, D. R. (2021). Piglet gut and in-barn manure from farms on a raised without antibiotics program display reduced antimicrobial resistance but an increased prevalence of pathogens. Antibiotics 10:1152. doi: 10.3390/antibiotics10101152
Chopyk, J., Nasko, D. J., Allard, S., Callahan, M. T., Bui, A., Ferelli, A. M. C., et al. (2020). Metagenomic analysis of bacterial and viral assemblages from a freshwater creek and irrigated field reveals temporal and spatial dynamics. Sci. Total Environ. 706:135395. doi: 10.1016/j.scitotenv.2019.135395
Chow, L., Waldron, L., and Gillings, M. R. (2015). Potential impacts of aquatic pollutants: sub-clinical antibiotic concentrations induce genome changes and promote antibiotic resistance. Front. Microbiol. 6:803. doi: 10.3389/fmicb.2015.00803
Chow, L. K. M., Ghaly, T. M., and Gillings, M. R. (2021). A survey of sub-inhibitory concentrations of antibiotics in the environment. J. Environ. Sci. 99, 21–27. doi: 10.1016/j.jes.2020.05.030
Colavecchio, A., Cadieux, B., Lo, A., and Goodridge, L. D. (2017). Bacteriophages contribute to the spread of antibiotic resistance genes among foodborne pathogens of the Enterobacteriaceae family – a review. Front. Microbiol. 8:1108. doi: 10.3389/fmicb.2017.01108
Danner, M. C., Robertson, A., Behrends, V., and Reiss, J. (2019). Antibiotic pollution in surface fresh waters: occurrence and effects. Sci. Total Environ. 664, 793–804. doi: 10.1016/j.scitotenv.2019.01.406
Donlan, R., Murga, R., Carpenter, J., Brown, E., Besser, R., and Fields, B. (2002). “Monochloramine disinfection of biofilm-associated Legionella pneumophila in a potable water model system,” in Legionella, eds R. Marre, Y. A. Kwaik, and C. Bartlett (Washington, DC: American Society of Microbiology), doi: 10.1128/9781555817985.ch82
Ebmeyer, S., Kristiansson, E., and Larsson, D. G. J. (2021). A framework for identifying the recent origins of mobile antibiotic resistance genes. Commun. Biol. 4:8. doi: 10.1038/s42003-020-01545-5
Enault, F., Briet, A., Bouteille, L., Roux, S., Sullivan, M. B., and Petit, M.-A. (2017). Phages rarely encode antibiotic resistance genes: a cautionary tale for virome analyses. ISME J. 11, 237–247. doi: 10.1038/ismej.2016.90
EUCAST (2019). The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 9.0. Available online at: http://www.eucast.org (accessed November 09, 2021).
Fernández, L., and Hancock, R. E. W. (2012). Adaptive and mutational resistance: role of porins and efflux pumps in drug resistance. Clin. Microbiol. Rev. 25, 661–681. doi: 10.1128/CMR.00043-12
Fernández, L., Rodríguez, A., and García, P. (2018). Phage or foe: an insight into the impact of viral predation on microbial communities. ISME J. 12, 1171–1179. doi: 10.1038/s41396-018-0049-5
Finley, R. L., Collignon, P., Larsson, D. G. J., Mcewen, S. A., Li, X. Z., Gaze, W. H., et al. (2013). The scourge of antibiotic resistance: the important role of the environment. Clin. Infect. Dis. 57, 704–710. doi: 10.1093/cid/cit355
Flemming, H. C., and Wingender, J. (2010). The biofilm matrix. Nat. Rev. Microbiol. 8, 623–633. doi: 10.1038/nrmicro2415
Flood, J. A., and Ashbolt, N. J. (2000). Virus-sized particles can be entrapped and concentrated one hundred fold within wetland biofilms. Adv. Environ. Res. 3, 403–411.
Friman, V.-P., Guzman, L. M., Reuman, D. C., and Bell, T. (2015). Bacterial adaptation to sublethal antibiotic gradients can change the ecological properties of multitrophic microbial communities. Proc. R. Soc. B Biol. Sci. 282:20142920. doi: 10.1098/rspb.2014.2920
Fulaz, S., Vitale, S., Quinn, L., and Casey, E. (2019). Nanoparticle–biofilm interactions: the role of the EPS matrix. Trends Microbiol. 27, 915–926. doi: 10.1016/j.tim.2019.07.004
Gebreyohannes, G., Nyerere, A., Bii, C., and Sbhatu, D. B. (2019). Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 5:e02192. doi: 10.1016/j.heliyon.2019.e02192
Gillings, M. R., and Stokes, H. W. (2012). Are humans increasing bacterial evolvability? Trends Ecol. Evol. 27, 346–352. doi: 10.1016/j.tree.2012.02.006
Goulas, A., Belhadi, D., Descamps, A., Andremont, A., Benoit, P., Courtois, S., et al. (2020). How effective are strategies to control the dissemination of antibiotic resistance in the environment? A systematic review. Environ. Evid. 9, 1–32. doi: 10.1186/s13750-020-0187-x
Grenni, P., Ancona, V., and Barra Caracciolo, A. (2018). Ecological effects of antibiotics on natural ecosystems: a review. Microchem. J. 136, 25–39. doi: 10.1016/j.microc.2017.02.006
Gullberg, E., Cao, S., Berg, O. G., Ilbäck, C., Sandegren, L., Hughes, D., et al. (2011). Selection of resistant bacteria at very low antibiotic concentrations. PLoS Pathog. 7:e1002158. doi: 10.1371/journal.ppat.1002158
Guo, X., Yang, Y., Lu, D., Niu, Z., Feng, J., Chen, Y., et al. (2018). Biofilms as a sink for antibiotic resistance genes (ARGs) in the Yangtze Estuary. Water Res. 129, 277–286. doi: 10.1016/j.watres.2017.11.029
Hall, C. W., and Mah, T.-F. (2017). Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 41, 276–301. doi: 10.1093/femsre/fux010
Han, X.-M., Hu, H.-W., Chen, Q.-L., Yang, L.-Y., Li, H.-L., Zhu, Y.-G., et al. (2018). Antibiotic resistance genes and associated bacterial communities in agricultural soils amended with different sources of animal manures. Soil Biol. Biochem. 126, 91–102. doi: 10.1016/j.soilbio.2018.08.018
Henriot, C. P., Martak, D., Dagot, C., Petit, F., Topp, E., Bertrand, X., et al. (2021). “The fate of antibiotic-resistant bacteria in the environment,” in Emerging Contaminants: Occurrence and Impact, Vol. 1, eds N. Morin-Crini, E. Lichtfouse, and G. Crini (Cham: Springer International Publishing), 207–260. doi: 10.1007/978-3-030-69079-3_4
Hoffman, L. R., D’Argenio, D. A., MacCoss, M. J., Zhang, Z., Jones, R. A., and Miller, S. I. (2005). Aminoglycoside antibiotics induce bacterial biofilm formation. Nature 436, 1171–1175. doi: 10.1038/nature03912
Høiby, N., Bjarnsholt, T., Givskov, M., Molin, S., and Ciofu, O. (2010). Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 35, 322–332. doi: 10.1016/j.ijantimicag.2009.12.011
Hooban, B., Joyce, A., Fitzhenry, K., Chique, C., and Morris, D. (2020). The role of the natural aquatic environment in the dissemination of extended spectrum beta-lactamase and carbapenemase encoding genes: a scoping review. Water Res. 180, 1–12. doi: 10.1016/j.watres.2020.115880
Jackson, L. M. D., Kroukamp, O., Yeung, W. C., Ronan, E., Liss, S. N., and Wolfaardt, G. M. (2019). Species interaction and selective carbon addition during antibiotic exposure enhances bacterial survival. Front. Microbiol. 10:2730. doi: 10.3389/fmicb.2019.02730
Jain, S., Kumar, P., Vyas, R. K., Pandit, P., and Dalai, A. K. (2013). Occurrence and removal of antiviral drugs in environment: a review. Water Air Soil Pollut. 224:1410. doi: 10.1007/s11270-012-1410-3
Kaplan, J. B., Izano, E. A., Gopal, P., Karwacki, M. T., Kim, S., Bose, J. L., et al. (2012). Low levels of β-lactam antibiotics induce extracellular DNA release and biofilm formation in Staphylococcus aureus. mBio 3:e00198-12. doi: 10.1128/mBio.00198-12
Karkman, A., Do, T. T., Walsh, F., and Virta, M. P. J. (2018). Antibiotic-resistance genes in waste water. Trends Microbiol. 26, 220–228. doi: 10.1016/j.tim.2017.09.005
Khan, G. A., Berglund, B., Khan, K. M., Lindgren, P.-E., and Fick, J. (2013). Occurrence and abundance of antibiotics and resistance genes in rivers, canal and near drug formulation facilities – a study in Pakistan. PLoS One 8:e62712. doi: 10.1371/journal.pone.0062712
Kraemer, S. A., Ramachandran, A., and Perron, G. G. (2019). Antibiotic pollution in the environment: from microbial ecology to public policy. Microorganisms 7:180. doi: 10.3390/microorganisms7060180
Lawrence, J. R., Chenier, M. R., Roy, R., Beaumier, D., Fortin, N., Swerhone, G. D. W., et al. (2004). Microscale and molecular assessment of impacts of nickel, nutrients, and oxygen level on structure and function of river biofilm communities. Appl. Environ. Microbiol. 70, 4326–4339. doi: 10.1128/aem.70.7.4326-4339.2004
Lawrence, J. R., Korber, D. R., Hoyle, B. D., Costerton, J. W., and Caldwell, D. E. (1991). Optical sectioning of microbial biofilms. J. Bacteriol. 173, 6558–6567. doi: 10.1128/jb.173.20.6558-6567.1991
Lawrence, J. R., Neu, T. R., Paule, A., Korber, D. R., and Wolfaardt, G. M. (2015). “Aquatic biofilms: development, cultivation, analyses, and applications,” in Manual of Environmental Microbiology, eds M. Yates, C. Nakatsu, R. Miller, and S. Pillai (Washington, DC: ASM), 4.2.3-1–4.2.3-33. doi: 10.1128/9781555818821.ch4.2.3
Lawrence, J. R., Zhu, B., Swerhone, G. D. W., Roy, J., Tumber, V., Waiser, M. J., et al. (2012). Molecular and microscopic assessment of the effects of caffeine, acetaminophen, diclofenac, and their mixtures on river biofilm communities. Environ. Toxicol. Chem. 31, 508–517. doi: 10.1002/etc.1723
Lawrence, J. R., Zhu, B., Swerhone, G. D. W., Roy, J., Wassenaar, L. I., Topp, E., et al. (2009). Comparative microscale analysis of the effects of triclosan and triclocarban on the structure and function of river biofilm communities. Sci. Total Environ. 407, 3307–3316. doi: 10.1016/j.scitotenv.2009.01.060
Lehmann, K., Bell, T., Bowes, M. J., Amos, G. C. A., Gaze, W. H., Wellington, E. M. H., et al. (2016). Trace levels of sewage effluent are sufficient to increase class 1 integron prevalence in freshwater biofilms without changing the core community. Water Res. 106, 163–170. doi: 10.1016/j.watres.2016.09.035
Lépesová, K., Kraková, L., Pangallo, D., Medved’ová, A., Olejníková, P., Mackul’ak, T., et al. (2018). Prevalence of antibiotic-resistant coliform bacteria, Enterococcus spp. and Staphylococcus spp. in wastewater sewerage biofilm. J. Glob. Antimicrob. Resist. 14, 145–151. doi: 10.1016/j.jgar.2018.03.008
Lerminiaux, N. A., and Cameron, A. D. S. (2019). Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 65, 34–44. doi: 10.1139/cjm-2018-0275
Lewandowski, Z. (2000). Notes on biofilm porosity. Water Res. 34, 2620–2624. doi: 10.1016/S0043-1354(00)00186-X
Li, Y.-H., Lau, P. C. Y., Lee, J. H., Ellen, R. P., and Cvitkovitch, D. G. (2001). Natural genetic transformation of Streptococcus mutans growing in biofilms. J. Bacteriol. 183, 897–908. doi: 10.1128/jb.183.3.897-908.2001
Lundström, S. V., Östman, M., Bengtsson-Palme, J., Rutgersson, C., Thoudal, M., Sircar, T., et al. (2016). Minimal selective concentrations of tetracycline in complex aquatic bacterial biofilms. Sci. Total Environ. 553, 587–595. doi: 10.1016/j.scitotenv.2016.02.103
Madsen, J. S., Burmølle, M., Hansen, L. H., and Sørensen, S. J. (2012). The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 65, 183–195. doi: 10.1111/j.1574-695X.2012.00960.x
Maestre-Carballa, L., Lluesma Gomez, M., Angla Navarro, A., Garcia-Heredia, I., Martinez-Hernandez, F., and Martinez-Garcia, M. (2019). Insights into the antibiotic resistance dissemination in a wastewater effluent microbiome: bacteria, viruses and vesicles matter. Environ. Microbiol. 21, 4582–4596. doi: 10.1111/1462-2920.14758
Manaia, C. M., Graham, D., Topp, E., Martinez, J. L., Collignon, P., and Gaze, W. H. (2020). “Antibiotic resistance in the environment: expert perspectives,” in Antibiotic Resistance in the Environment: A Worldwide Overview, eds C. M. Manaia, E. Donner, I. Vaz-Moreira, and P. Hong (Cham: Springer International Publishing), 1–18. doi: 10.1007/698_2020_472
Mangalappalli-Illathu, A. K., Lawrence, J. R., Swerhone, G. D. W., and Korber, D. R. (2008a). Architectural adaptation and protein expression patterns of Salmonella enterica serovar enteritidis biofilms under laminar flow conditions. Int. J. Food Microbiol. 123, 109–120. doi: 10.1016/j.ijfoodmicro.2007.12.021
Mangalappalli-Illathu, A. K., Vidović, S., and Korber, D. R. (2008b). Differential adaptive response and survival of Salmonella enterica serovar enteritidis planktonic and biofilm cells exposed to benzalkonium chloride. Antimicrob. Agents Chemother. 52, 3669–3680. doi: 10.1128/AAC.00073-08
Manyi-Loh, C., Mamphweli, S., Meyer, E., and Okoh, A. (2018). Antibiotic use in agriculture and its consequential resistance in environmental sources: potential public health implications. Molecules 23:795. doi: 10.3390/molecules23040795
Mao, D., Yu, S., Rysz, M., Luo, Y., Yang, F., Li, F., et al. (2015). Prevalence and proliferation of antibiotic resistance genes in two municipal wastewater treatment plants. Water Res. 85, 458–466. doi: 10.1016/j.watres.2015.09.010
Marti, E., Variatza, E., and Balcazar, J. L. (2014). The role of aquatic ecosystems as reservoirs of antibiotic resistance. Trends Microbiol. 22, 36–41. doi: 10.1016/j.tim.2013.11.001
Martínez, J. L., Coque, T. M., and Baquero, F. (2015). What is a resistance gene? Ranking risk in resistomes. Nat. Rev. Microbiol. 13, 116–123. doi: 10.1038/nrmicro3399
Miller, M. B., and Bassler, B. L. (2001). Quorum sensing in bacteria. Annu. Rev. Microbiol. 55, 165–199. doi: 10.1146/annurev.micro.55.1.165
Miyaue, S., Suzuki, E., Komiyama, Y., Kondo, Y., Morikawa, M., and Maeda, S. (2018). Bacterial memory of persisters: bacterial persister cells can retain their phenotype for days or weeks after withdrawal from colony–biofilm culture. Front. Microbiol. 9:1396. doi: 10.3389/fmicb.2018.01396
Muniesa, M., Colomer-Lluch, M., and Jofre, J. (2013). Potential impact of environmental bacteriophages in spreading antibiotic resistance genes. Future Microbiol. 8, 739–751. doi: 10.2217/fmb.13.32
Murphy, K., Park, A. J., Hao, Y., Brewer, D., Lam, J. S., and Khursigara, C. M. (2014). Influence of O polysaccharides on biofilm development and outer membrane vesicle biogenesis in Pseudomonas aeruginosa PAO1. J. Bacteriol. 196, 1306–1317. doi: 10.1128/JB.01463-13
Nakao, R., Myint, S. L., Wai, S. N., and Uhlin, B. E. (2018). Enhanced biofilm formation and membrane vesicle release by Escherichia coli expressing a commonly occurring plasmid gene, kil. Front. Microbiol. 9:2605. doi: 10.3389/fmicb.2018.02605
Obst, U., Schwartz, T., and Volkmann, H. (2006). Antibiotic resistant pathogenic bacteria and their resistance genes in bacterial biofilms. Int. J. Artif. Organs 29, 387–394. doi: 10.1177/039139880602900408
O’Neill, J. (2016). Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Government of the United Kingdom. Available online at: https://apo.org.au/node/63983 (accessed May 19, 2016).
Orazi, G., and O’Toole, G. A. (2019). “It takes a village”: mechanisms underlying antimicrobial recalcitrance of polymicrobial biofilms. J. Bacteriol. 202:e00530-19. doi: 10.1128/jb.00530-19
Palermo, C. N., Fulthorpe, R. R., Saati, R., and Short, S. M. (2019). Metagenomic analysis of virus diversity and relative abundance in a eutrophic freshwater harbour. Viruses 11:792. doi: 10.3390/v11090792
Pärnänen, K. M. M., Narciso-da-Rocha, C., Kneis, D., Berendonk, T. U., Cacace, D., Do, T. T., et al. (2019). Antibiotic resistance in European wastewater treatment plants mirrors the pattern of clinical antibiotic resistance prevalence. Sci. Adv. 5:eaau9124. doi: 10.1126/sciadv.aau9124
Parsley, L. C., Consuegra, E. J., Thomas, S. J., Bhavsar, J., Land, A. M., Bhuiyan, N. N., et al. (2010). Census of the viral metagenome within an activated sludge microbial assemblage. Appl. Environ. Microbiol. 76, 2673–2677. doi: 10.1128/AEM.02520-09
Partridge, S. R., Kwong, S. M., Firth, N., and Jensen, S. O. (2018). Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 31:e00088-17. doi: 10.1128/CMR.00088-17
Petrovich, M. L., Zilberman, A., Kaplan, A., Eliraz, G. R., Wang, Y., Langenfeld, K., et al. (2020). Microbial and viral communities and their antibiotic resistance genes throughout a hospital wastewater treatment system. Front. Microbiol. 11:153. doi: 10.3389/fmicb.2020.00153
Poole, K. (2017). At the nexus of antibiotics and metals: the impact of Cu and Zn on antibiotic activity and resistance. Trends Microbiol. 25, 820–832. doi: 10.1016/j.tim.2017.04.010
Proia, L., Vilches, C., Boninneau, C., Kantiani, L., Farré, M., Romaní, A. M., et al. (2013). Drought episode modulates the response of river biofilms to triclosan. Aquat. Toxicol. 127, 36–45. doi: 10.1016/j.aquatox.2012.01.006
Quinlan, E. L., Nietch, C. T., Blocksom, K., Lazorchak, J. M., Batt, A. L., Griffiths, R., et al. (2011). Temporal dynamics of periphyton exposed to tetracycline in stream mesocosms. Environ. Sci. Technol. 45, 10684–10690. doi: 10.1021/es202004k
Ramanan, R., Kim, B.-H., Cho, D.-H., Oh, H.-M., and Kim, H.-S. (2016). Algae–bacteria interactions: evolution, ecology and emerging applications. Biotechnol. Adv. 34, 14–29. doi: 10.1016/j.biotechadv.2015.12.003
Rodriguez-Mozaz, S., Chamorro, S., Marti, E., Huerta, B., Gros, M., Sànchez-Melsió, A., et al. (2015). Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 69, 234–242. doi: 10.1016/j.watres.2014.11.021
Romaní, A. M. (2009). “Freshwater biofilms,” in Biofouling, eds S. Dürr and J. C. Thomason (Oxford: John Wiley & Sons, Ltd), 137–153. doi: 10.1002/9781444315462.ch10
Romaní, A. M., Borrego, C. M., Díaz-Villanueva, V., Freixa, A., Gich, F., and Ylla, I. (2014). Shifts in microbial community structure and function in light- and dark-grown biofilms driven by warming. Environ. Microbiol. 16, 2550–2567. doi: 10.1111/1462-2920.12428
Romaní, A. M., Guasch, H., Muñoz, I., Ruana, J., Vilalta, E., Schwartz, T., et al. (2004). Biofilm structure and function and possible implications for riverine DOC dynamics. Microb. Ecol. 47, 316–328. doi: 10.1007/s00248-003-2019-2
Romero, F., Acuña, V., Font, C., Freixa, A., and Sabater, S. (2019). Effects of multiple stressors on river biofilms depend on the time scale. Sci. Rep. 9:15810. doi: 10.1038/s41598-019-52320-4
Sasikaladevi, R., Kiruthika Eswari, V., and Nambi, I. M. (2020). “Antibiotic resistance and sanitation in India: current situation and future perspectives,” in Antibiotic Resistance in the Environment: A Worldwide Overview, eds C. M. Manaia, E. Donner, I. Vaz-Moreira, and P. Hong (Cham: Springer International Publishing), 217–244. doi: 10.1007/698_2020_608
Schroeder, M., Brooks, B. D., and Brooks, A. E. (2017). The complex relationship between virulence and antibiotic resistance. Genes (Basel) 8:39. doi: 10.3390/genes8010039
Schwartz, T., Kohnen, W., Jansen, B., and Obst, U. (2003). Detection of antibiotic-resistant bacteria and their resistance genes in wastewater, surface water, and drinking water biofilms. FEMS Microbiol. Ecol. 43, 325–335. doi: 10.1111/j.1574-6941.2003.tb01073.x
Sims, N., and Kasprzyk-Hordern, B. (2020). Future perspectives of wastewater-based epidemiology: monitoring infectious disease spread and resistance to the community level. Environ. Int. 139:105689. doi: 10.1016/j.envint.2020.105689
Slizovskiy, I. B., Mukherjee, K., Dean, C. J., Boucher, C., and Noyes, N. R. (2020). Mobilization of antibiotic resistance: are current approaches for colocalizing resistomes and mobilomes useful? Front. Microbiol. 11:1376. doi: 10.3389/fmicb.2020.01376
Stewart, P. S., and Costerton, J. W. (2001). Antibiotic resistance of bacteria in biofilms. Lancet 358, 135–138. doi: 10.1016/S0140-6736(01)05321-1
Subirats, J., Timoner, X., Sànchez-Melsió, A., Balcázar, J. L., Acuña, V., Sabater, S., et al. (2018). Emerging contaminants and nutrients synergistically affect the spread of class 1 integron-integrase (intI1) and sul1 genes within stable streambed bacterial communities. Water Res. 138, 77–85. doi: 10.1016/j.watres.2018.03.025
Sutherland, I. W., Hughes, K. A., Skillman, L. C., and Tait, K. (2004). The interaction of phage and biofilms. FEMS Microbiol. Lett. 232, 1–6. doi: 10.1016/S0378-1097(04)00041-2
Szomolay, B., Klapper, I., Dockery, J., and Stewart, P. S. (2005). Adaptive responses to antimicrobial agents in biofilms. Environ. Microbiol. 7, 1186–1191. doi: 10.1111/j.1462-2920.2005.00797.x
Tamaki, H., Zhang, R., Angly, F. E., Nakamura, S., Hong, P.-Y., Yasunaga, T., et al. (2012). Metagenomic analysis of DNA viruses in a wastewater treatment plant in tropical climate. Environ. Microbiol. 14, 441–452. doi: 10.1111/j.1462-2920.2011.02630.x
Thakur, S., and Gray, G. C. (2019). The mandate for a global “One Health” approach to antimicrobial resistance surveillance. Am. J. Trop. Med. Hyg. 100, 227–228. doi: 10.4269/ajtmh.18-0973
Thieme, L., Hartung, A., Tramm, K., Klinger-Strobel, M., Jandt, K. D., Makarewicz, O., et al. (2019). MBEC versus MBIC: the lack of differentiation between biofilm reducing and inhibitory effects as a current problem in biofilm methodology. Biol. Proced. Online 21:18. doi: 10.1186/s12575-019-0106-0
Tiseo, K., Huber, L., Gilbert, M., Robinson, T. P., and Van Boeckel, T. P. (2020). Global trends in antimicrobial use in food animals from 2017 to 2030. Antibiotics 9:918. doi: 10.3390/antibiotics9120918
Topp, E., Larsson, D. G. J., Miller, D. N., Van den Eede, C., and Virta, M. P. J. (2018). Antimicrobial resistance and the environment: assessment of advances, gaps and recommendations for agriculture, aquaculture and pharmaceutical manufacturing. FEMS Microbiol. Ecol. 94:fix185. doi: 10.1093/femsec/fix185
Tu, C., Chen, T., Zhou, Q., Liu, Y., Wei, J., Waniek, J. J., et al. (2020). Biofilm formation and its influences on the properties of microplastics as affected by exposure time and depth in the seawater. Sci. Total Environ. 734:139237. doi: 10.1016/j.scitotenv.2020.139237
Van Boeckel, T. P., Pires, J., Silvester, R., Zhao, C., Song, J., Criscuolo, N. G., et al. (2019). Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 365:eaaw1944. doi: 10.1126/science.aaw1944
Vasudevan, S., Swamy, S. S., Kaur, G., Princy, S. A., and Balamurugan, P. (2018). “Synergism between quorum sensing inhibitors and antibiotics: combating the antibiotic resistance crisis,” in Biotechnological Applications of Quorum Sensing Inhibitors, ed. V. C. Kalia (Singapore: Springer Singapore), 209–225. doi: 10.1007/978-981-10-9026-4_10
Villanueva, V. D., Font, J., Schwartz, T., and Romaní, A. M. (2011). Biofilm formation at warming temperature: acceleration of microbial colonization and microbial interactive effects. Biofouling 27, 59–71. doi: 10.1080/08927014.2010.538841
Waiser, M. J., Swerhone, G. D. W., Roy, J., Tumber, V., and Lawrence, J. R. (2016). Effects of erythromycin, trimethoprim and clindamycin on attached microbial communities from an effluent dominated prairie stream. Ecotoxicol. Environ. Saf. 132, 31–39. doi: 10.1016/j.ecoenv.2016.05.026
Wang, F., Fu, Y.-H., Sheng, H.-J., Topp, E., Jiang, X., Zhu, Y.-G., et al. (2021). Antibiotic resistance in the soil ecosystem: a One Health perspective. Curr. Opin. Environ. Sci. Health 20:100230. doi: 10.1016/j.coesh.2021.100230
Wang, N., Guo, X., Yan, Z., Wang, W., Chen, B., Ge, F., et al. (2016). A comprehensive analysis on spread and distribution characteristic of antibiotic resistance genes in livestock farms of Southeastern China. PLoS One 11:e0156889. doi: 10.1371/journal.pone.0156889
Wellington, E. M. H., Boxall, A. B. A., Cross, P., Feil, E. J., Gaze, W. H., Hawkey, P. M., et al. (2013). The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect. Dis. 13, 155–165. doi: 10.1016/S1473-3099(12)70317-1
White, A., and Hughes, J. M. (2019). Critical importance of a One Health approach to antimicrobial resistance. Ecohealth 16, 404–409. doi: 10.1007/s10393-019-01415-5
WHO (2017). Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis. License: CC BY-NC-SA 3.0 IGO. Geneva: World Health Organization.
Wingender, J., and Flemming, H. C. (2011). Biofilms in drinking water and their role as reservoir for pathogens. Int. J. Hyg. Environ. Health 214, 417–423. doi: 10.1016/j.ijheh.2011.05.009
Wu, Q., and Liu, W.-T. (2009). Determination of virus abundance, diversity and distribution in a municipal wastewater treatment plant. Water Res. 43, 1101–1109. doi: 10.1016/j.watres.2008.11.039
Xiang, S., Wang, X., Ma, W., Liu, X., Zhang, B., Huang, F., et al. (2020). Response of microbial communities of karst river water to antibiotics and microbial source tracking for antibiotics. Sci. Total Environ. 706:135730. doi: 10.1016/j.scitotenv.2019.135730
Yang, K., Chen, Q.-L., Chen, M.-L., Li, H.-Z., Liao, H., Pu, Q., et al. (2020). Temporal dynamics of antibiotic resistome in the plastisphere during microbial colonization. Environ. Sci. Technol. 54, 11322–11332. doi: 10.1021/acs.est.0c04292
Zhang, Y., and LeJeune, J. T. (2008). Transduction of blaCMY-2, tet(A), and tet(B) from Salmonella enterica subspecies enterica serovar Heidelberg to S. typhimurium. Vet. Microbiol. 129, 418–425. doi: 10.1016/j.vetmic.2007.11.032
Zhao, Y., Yang, Q. E., Zhou, X., Wang, F. H., Muurinen, J., Virta, M. P., et al. (2020). Antibiotic resistome in the livestock and aquaculture industries: status and solutions. Crit. Rev. Environ. Sci. Technol. 51, 1–38. doi: 10.1080/10643389.2020.1777815
Zhou, L., Li, T., An, J., Liao, C., Li, N., and Wang, X. (2017). Subminimal inhibitory concentration (sub-MIC) of antibiotic induces electroactive biofilm formation in bioelectrochemical systems. Water Res. 125, 280–287. doi: 10.1016/j.watres.2017.08.059
Keywords: biofilms, antibiotic resistance, reservoirs, environmental resistome, bacteriophage, sub-minimum inhibitory concentration
Citation: Flores-Vargas G, Bergsveinson J, Lawrence JR and Korber DR (2021) Environmental Biofilms as Reservoirs for Antimicrobial Resistance. Front. Microbiol. 12:766242. doi: 10.3389/fmicb.2021.766242
Received: 28 August 2021; Accepted: 22 November 2021;
Published: 13 December 2021.
Edited by:
Sanket J. Joshi, Sultan Qaboos University, OmanReviewed by:
Lei Wang, Charité University Medicine Berlin, GermanyCopyright © 2021 Flores-Vargas, Bergsveinson, Lawrence and Korber. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Darren R. Korber, ZHJrMTM3QG1haWwudXNhc2suY2E=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.