 Carina Danchik
Carina Danchik Siqing Wang
Siqing Wang Petros C. Karakousis
Petros C. Karakousis
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 07 October 2021
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.744167
This article is part of the Research Topic Approaches to address Resistance, Drug Discovery, and Vaccine development in Mycobacterium Tuberculosis: Challenges and opportunities View all 9 articles
The stringent response is well conserved across bacterial species and is a key pathway involved both in bacterial survival and virulence and in the induction of antibiotic tolerance in Mycobacteria. It is mediated by the alarmone (p)ppGpp and the regulatory molecule inorganic polyphosphate in response to stress conditions such as nutrient starvation. Efforts to pharmacologically target various components of the stringent response have shown promise in modulating mycobacterial virulence and antibiotic tolerance. In this review, we summarize the current understanding of the stringent response and its role in virulence and tolerance in Mycobacteria, including evidence that targeting this pathway could have therapeutic benefit.
Current treatments for tuberculosis (TB) are lengthy and burdensome. Medical non-adherence and the continuing emergence of multi drug-resistant Mycobacterium tuberculosis strains contribute to the frequent failure of current antibiotic therapies to clear M. tuberculosis infection. The long duration required for curative TB treatment reflects the presence of a small population of bacteria characterized by antibiotic tolerance. Multiple strategies have been proposed to enhance the antibacterial activity of currently available antibiotics, thereby shortening the time required to achieve a stable cure, including boosting host defense pathways or inhibiting factors required for M. tuberculosis persistence and antibiotic tolerance, such as the stringent response (Kana et al., 2014; Frank et al., 2019).
Bacteria have evolved numerous strategies to evade killing by antibiotics. Reduced antibiotic susceptibility may result from genetic or phenotypic alterations and can be classified as resistance, persistence, or tolerance. Although each is associated with reduced antibiotic activity, it is important to distinguish among these three separate phenomena. A clear understanding of the diverse molecular pathways through which bacteria can become unresponsive to antibiotics is critical for the development of novel and more effective therapeutic approaches.
Antibiotic resistance is the most well-understood mechanism of bacterial insensitivity. The term applies to an entire bacterial population which exhibits an increase in the minimum inhibitory concentration of a particular antibiotic under optimal bacterial growth conditions. Antibiotic resistance may arise from mutations in genes encoding antibiotic targets or antibiotic-activating enzymes, mutations leading to increased enzymatic degradation of or reduced cell permeability to the antibiotic, or from increased activity of antibiotic efflux pumps (Hoffman, 2001; Karakousis, 2009; Westblade et al., 2020).
Antibiotic tolerance and persistence are both used to describe reversible states of broad insusceptibility to antibiotics. Although their usage in the literature remains ambiguous at times, antibiotic tolerance generally is used to describe an entire population of bacteria, while persistence refers to a subpopulation of phenotypically distinct bacteria within a clonal population. Both phenomena are frequently associated with a slowly growing or dormant state (Balaban et al., 2013; Maisonneuve and Gerdes, 2014).
Persistence is a non-heritable state in which a small proportion of bacteria in a clonal population can survive transient antibiotic exposure. This is characterized by a biphasic killing curve in which most of the bacterial population is killed rapidly, leaving behind a small number of persister bacteria which are insensitive to antibiotics and are killed at a much slower rate (Maisonneuve and Gerdes, 2014; Brauner et al., 2016). Persister cells may arise in the absence of any specific stimuli, perhaps due to stochastic differences in expression of specific genes, or may result from exposure to stress conditions. Persister cells are genetically identical to the rest of the bacterial population, and the antibiotic susceptibility patterns of daughter cells mirror those of the original parental population. Persistence has been attributed to reduced growth rate, metabolism, and protein synthesis as well as to the activity of toxin–antitoxin systems (Dahl et al., 2003; Sala et al., 2014; Harms et al., 2016; Talwar et al., 2020).
A wide array of stresses can induce antibiotic tolerance, including hypoxia, nitric oxide, and nutrient starvation (Gibson et al., 2018). Antibiotic tolerance is mediated by a variety of mechanisms, including the stringent response, discussed in more detail below, and drug efflux pumps (Goossens et al., 2021). Antibiotic tolerance may have a phenotypic or genetic basis and is often associated with an altered transcriptomic and metabolic state (Dahl et al., 2003; Dutta et al., 2019; Goossens et al., 2021). Additionally, actively dividing M. tuberculosis is able to develop antibiotic tolerance within macrophages through the activity of its efflux pumps (Adams et al., 2011, 2014; Szumowski et al., 2013; Black et al., 2014; Jang et al., 2017). These pumps can export antibiotics, reducing their intracellular concentrations and antibacterial effects, and pharmacological inhibition of the efflux pumps prevents the development of this macrophage-induced tolerance (Adams et al., 2011, 2014; Jang et al., 2017).
Although antibiotic resistance, tolerance, and persistence may reflect distinct phenomena, they may also be interrelated. Previous work has shown that bacterial mutations accumulate more rapidly under persistence- and tolerance-inducing stress conditions (Balaban et al., 2013). Such mutations can confer resistance to antibiotics, thus promoting the survival of tolerant bacteria (Levin-Reisman et al., 2017).
The stringent response is a conserved bacterial adaptation to nutrient starvation and other stress conditions. It is mediated by tetra- or penta-phosphorylated guanosine [(p)ppGpp] and inorganic polyphosphate [poly(P)]. Accumulation of these two regulatory molecules in E. coli leads to downregulation of growth-related pathways through altered transcription and metabolism (Traxler et al., 2008). In M. tuberculosis, the stringent response enables bacteria to survive host defenses by inducing metabolic quiescence, thereby contributing to long-term mycobacterial survival and virulence (Primm et al., 2000). By reducing mycobacterial growth and metabolism, the stringent response also contributes to antibiotic tolerance by suppressing the activity of numerous antibiotic targets.
The production and metabolism of (p)ppGpp are mediated by members of the RelA/SpoT homology (RSH)-type protein family, which are conserved across bacterial species. In E. coli and some other gram-negative bacteria, (p)ppGpp synthesis and hydrolysis are regulated by two separate enzymes, RelA and SpoT, respectively, where RelA is a (p)ppGpp synthase and SpoT is a bifunctional protein with both (p)ppGpp synthesis and hydrolysis activities (Ronneau and Hallez, 2019). M. tuberculosis contains a single bifunctional RSH enzyme, designated RelMtb, with dual (p)ppGpp synthesis and hydrolysis activities (Avarbock et al., 1999). The constitutive activity of RelMtb maintains (p)ppGpp at a basal level and is required for bacterial growth and biofilm formation in vitro and in vivo (Weiss and Stallings, 2013).
The opposing activities of RelMtb are contained in distinct regions of the protein. RelMtb is a 738-amino acid enzyme, with catalytic domains in the N-terminal region and regulatory domains in the C-terminal region (Avarbock et al., 1999, 2005). In the N-terminal region (AA 1–394), residues 1–203 exhibit (p)ppGpp hydrolysis activity, and residues 87–394 exhibit (p)ppGpp synthesis activity, while the C-terminal regulatory domains are confined to residues 395–738 (Avarbock et al., 2005; Singal et al., 2017). The synthase domain catalyzes the transfer of the 5′-beta, gamma-pyrophosphate group from ATP to the 3’ OH group of GDP or GTP, producing ppGpp or pppGpp, respectively. The hydrolysis domain catalyzes the opposite reaction, i.e., hydrolysis of the pyrophosphate group on (p)ppGpp, producing GDP or GTP (Avarbock et al., 2005).
During amino acid deprivation, uncharged tRNA accumulates and competes with charged tRNA for the ribosome aminoacyl (A) site, giving rise to the RelMtb activating complex, which is comprised of uncharged tRNA, ribosomes, and cognate mRNA (Avarbock et al., 2000). This complex stimulates RelMtb synthase activity to produce (p)ppGpp. Conversely, under conditions of amino acid abundance, charged tRNA outcompetes uncharged tRNA at the ribosome A-site, suppressing (p)ppGpp synthesis (Avarbock et al., 2000).
(p)ppGpp is a global regulator of the stress and starvation response in bacteria. In E. coli, a basal level of (p)ppGpp is required to maintain normal low levels of the alternative sigma factor RpoS (Battesti et al., 2011). Under stress conditions, elevated (p)ppGpp levels induce production and reduce degradation of RpoS, which, in turn, regulates expression of 10% of the E. coli genome through direct or indirect interaction with RNA polymerase. This altered transcription yields a stationary phase-like phenotype marked by slowed growth and stress resistance (Battesti et al., 2011). In M. tuberculosis, a total of 159 genes, including many encoding virulence factors, important antigens, and proteins involved in transcription and translation, are regulated by (p)ppGpp expression (Dahl et al., 2003).
Inorganic polyphosphate [poly(P)], a linear polymer consisting of up to hundreds of phosphate residues, is ubiquitous across bacterial species and plays diverse roles in growth mediation, stress responses, and virulence (Rao et al., 2009). Poly(P) metabolism is controlled by polyphosphate kinases (PPKs) and exopolyphosphatases (PPXs), which are responsible for synthesis and hydrolysis, respectively. PPK1 (Rv2984), the primary polyphosphate kinase in M. tuberculosis, catalyzes the formation and elongation of a poly(P) chain using the gamma-phosphate from ATP. Ppk1 transcription is upregulated by the two-component system SenX3-RegX3 under phosphate-limited conditions, leading to increased poly(P) synthesis (Rifat et al., 2009; Sanyal et al., 2013; Namugenyi et al., 2017). PPK2 (Rv3232c), although historically annotated as a kinase due to its first described activity, is primarily responsible for poly(P) hydrolysis and may be more accurately characterized as a PPX. PPX1 (Rv0496) hydrolyzes short-chain poly(P), while PPX2 (Rv1026) hydrolyzes long-chain poly(P) (Sureka et al., 2007, 2009; Thayil et al., 2011; Chuang et al., 2015). Poly(P) levels increase during the stationary phase of growth and under various stress conditions, including upon exposure to antibiotics, and promote bacterial survival and virulence (Singh et al., 2013; Chuang et al., 2015). In E. coli, poly(P) binds and activates the ATP-dependent protease Lon, stimulating protein degradation to provide an alternative source of molecular building blocks during amino acid deprivation (Kuroda, 2006; Singh et al., 2013).
The two stringent response molecules are also able to positively regulate each other (Figure 1). (p)ppGpp also binds to and inhibits PPX1 and PPX2, decreasing poly(P) degradation and leading to poly(P) accumulation (Choi et al., 2012; Chuang et al., 2015). Conversely, poly(P) induces (p)ppGpp synthesis through the mprAB-sigE-relMtb regulatory cascade (Sureka et al., 2007). In this cascade, poly(P) serves as the phosphate donor for autophosphorylation of the histidine kinase MprB and its subsequent phosphorylation of MprA. MprA then regulates the expression of its own MprAB operon as well as that of sigE, a sigma factor which promotes relMtb transcription (Sureka et al., 2007). Thus, both (p)ppGpp and poly(P) regulate each other through a positive feedback mechanism. The intracellular levels of these regulatory molecules are controlled primarily by the bifunctional RSH enzyme RelMtb and by the various kinases and phosphatases involved in poly(P) metabolism and signaling pathways, which are critical for eliciting and maintaining the stringent response under stress conditions.
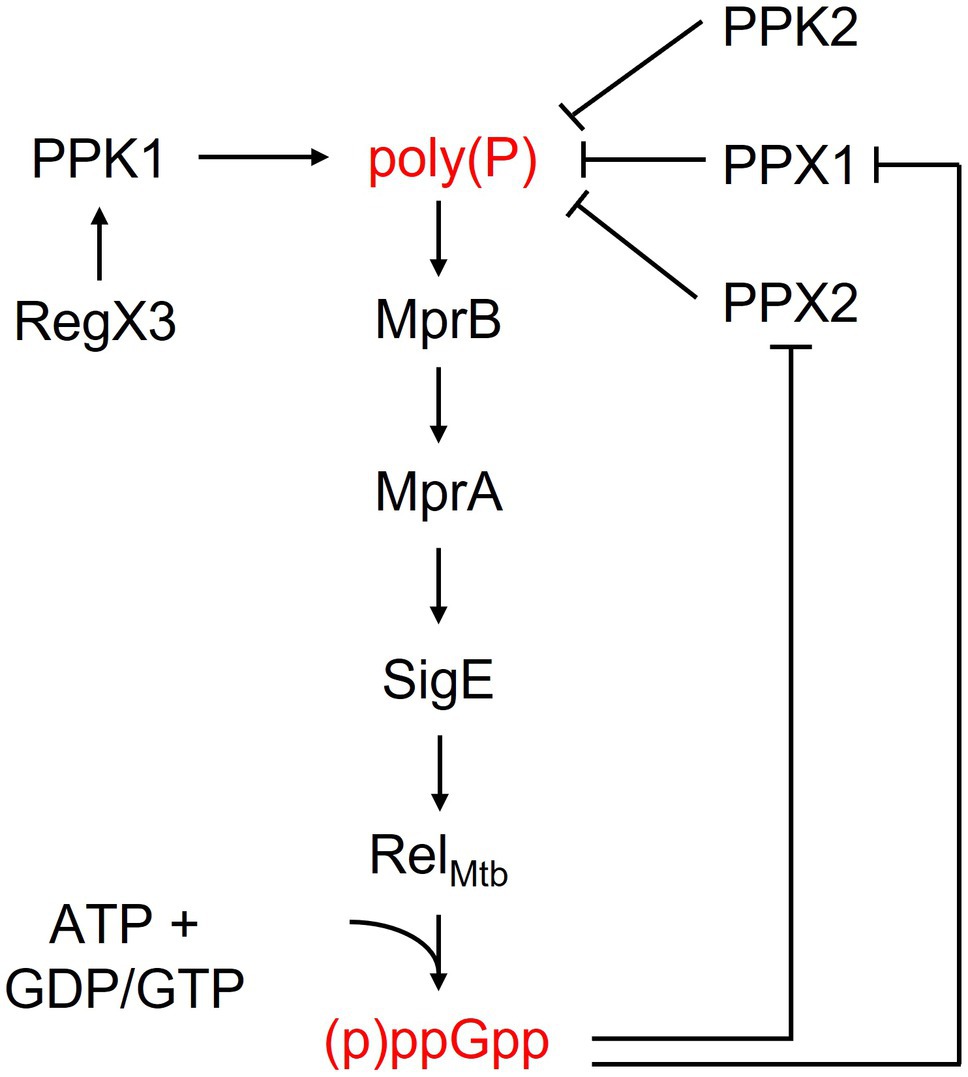
Figure 1. Activity and regulation of key bacterial stringent response factors.
The stringent response contributes to M. tuberculosis survival under physiologically relevant stress conditions and to virulence within the infected mammalian host. Disruption of RelMtb, PPK, or PPX leads to abnormal levels of (p)ppGpp, poly(P), and other cell metabolites, as well as to reduced viability and virulence (Primm et al., 2000; Dutta et al., 2019).
Deletion of relMtb is associated with defective M. tuberculosis survival under nutrient starvation and hypoxia conditions (Primm et al., 2000). Recently, a relMtb-deficient mutant was found to have an impaired ability to slow its replication rate or downregulate lipid metabolism during nutrient starvation, resulting in reduced viability (Dutta et al., 2019). Although aerosol infection of C57BL/6 mice with ΔrelMtb resulted in normal initial bacterial growth and containment, the long-term survival of this strain in mouse lungs and spleens was severely impaired, and the histopathology of these organs was markedly reduced in mice infected with ΔrelMtb relative to those infected with the isogenic wild-type strain (Dahl et al., 2003). Similarly, a relMtb-deficient mutant was found to have reduced extracellular survival in a murine hypoxic granuloma model of latent TB infection (Karakousis et al., 2004; Klinkenberg et al., 2008). Deletion of relMtb resulted in impaired initial M. tuberculosis growth and survival, as well as a striking absence of gross tubercle lesions and histological evidence of caseous granulomas in the lungs of infected guinea pigs (Klinkenberg et al., 2010). Finally, C3HeB/FeJ mice, which develop necrotic TB lung granulomas characterized by tissue hypoxia, had significantly prolonged survival after aerosol infection with ∆relMtb than after infection with the isogenic wild-type strain (Harper et al., 2012; Dutta et al., 2019).
Poly(P) accumulates in response to stress conditions, and disruption of the enzymes involved in poly(P) synthesis and hydrolysis affects the bacteria’s ability to adapt to these conditions. M. tuberculosis mutants deficient in PPX1, PPX2, and PPK2 have impaired hydrolysis activity and exhibit constitutively higher levels of intracellular poly(P) relative to wild type (Thayil et al., 2011; Chuang et al., 2013, 2016). Conversely, a PPK1-deficient mutant showed impaired synthesis activity and exhibited lower levels of poly(P) (Prusa et al., 2018). Poly(P)-accumulating strains deficient in PPX1, PPX2, and PPK2 were found to have defective biofilm formation (Chuang et al., 2015, 2016).
Poly(P) accumulation in M. tuberculosis is also associated with increased lipid oxidation and citrate cycle activity, as well as altered expression of genes involved in glycerol-3-phosphate (G3P) homeostasis, contributing to reduced intracellular G3P content in poly(P)-accumulating strains (Chuang et al., 2016). G3P can be used as a scaffold for phospholipid biosynthesis, and overexpression of G3P dehydrogenase in E. coli leads to reduced intracellular G3P levels and increased formation of persister cells following exposure to antibiotics (Spoering et al., 2006; Yao and Rock, 2013). Poly(P) accumulation also appears to alter peptidoglycan synthesis in M. tuberculosis (Chuang et al., 2016). A ppx2 knockdown strain displayed altered cell wall thickness and permeability, and a ppk2-deficient mutant showed reduced Nile red staining relative to control strains (Chuang et al., 2015, 2016). Furthermore, a PPK2-deficient mutant was found to be more sensitive to the toxic hydroxy-1,4-naphthoquinone, plumbagin, relative to isogenic wild-type and complemented strains (Chuang et al., 2016).
Poly(P) accumulation also affects M. tuberculosis virulence in vivo. A ppk2-deficient mutant showed reduced survival at day 7 in activated and naive J774 macrophages relative to the wild-type strain, and naive macrophages showed increased expression of interleukin 2 (IL-2), IL-9, IL-10, IL-12p70, and gamma interferon (IFN-γ) following infection with the mutant relative to those infected with the wild type (Chuang et al., 2013). Consistent with a requirement of ppk2 for full M. tuberculosis virulence in vivo, a ppk2-deficient mutant exhibited a significantly lower lung bacillary burden during acute murine infection compared to the control groups (Chuang et al., 2013). Similarly, a ppx1-deficient mutant showed a significant survival defect in activated human macrophages and reduced survival in the lungs of guinea pigs (Thayil et al., 2011). Furthermore, M. tuberculosis deficiency of the phosphate-specific transport PhoU orthologs, PhoY1 and PhoY2, led to increased transcription of ppk1 in a RegX3-dependent manner, accumulation of poly(P) during log phase growth, and reduced survival relative to wild type in the lungs of mice 12weeks after infection (Namugenyi et al., 2017).
As in the case of poly(P) accumulation, poly(P) deficiency is also associated with reduced M. tuberculosis stress adaptation and virulence. Thus, compared to the wild-type strain, a ppk1-deficient mutant displayed a survival defect in response to nitrosative stress in vitro and in THP-1 macrophages, as well as a lower mycobacterial load and fewer necrotic granulomas in the lungs of guinea pigs at 10weeks post-infection (R. Singh et al., 2013). Taken together, these data suggest that poly(P) levels are tightly regulated in M. tuberculosis, and disturbance of poly(P) homeostasis is associated with a reduced stress resistance and virulence.
Perturbation of the M. tuberculosis stringent response leads to altered antibiotic susceptibility, suggesting that, in addition to stress adaptation and virulence, this pathway also plays a role in antibiotic tolerance.
Although deletion of the relMtb gene did not alter M. tuberculosis susceptibility to multiple antibiotics during logarithmic growth in nutrient-rich broth, the MBC of isoniazid was reduced 512-fold against ΔrelMtb relative to the wild-type and complemented strains during nutrient starvation (Primm et al., 2000; Dutta et al., 2019). Furthermore, 2weeks of oral therapy with human-equivalent doses of isoniazid significantly reduced the lung bacillary burden of mice chronically infected with ΔrelMtb (2.03 log10 CFU reduction) relative to those infected with control strains (0.11 log10 CFU reduction; p<0.0001; Dutta et al., 2019). These findings suggest that the stringent response is required for M. tuberculosis tolerance to bactericidal antibiotics during stress exposure in vitro and in animal tissues.
Dysregulation of poly(P) homeostasis also alters the antibiotic susceptibility profile of M. tuberculosis. Thus, a poly(P)-deficient ppk1 deletion mutant showed increased susceptibility to several antibiotics, including isoniazid and levofloxacin (Singh et al., 2013). Alternatively, a poly(P)-accumulating ΔphoY1 ΔphoY2 double mutant was found to have increased susceptibility to rifampin in vitro and in mouse lungs during the chronic phase of injection, and this phenotype was not associated with altered cell wall permeability (Namugenyi et al., 2017). Two other poly(P)-accumulating strains, namely a ppx2 knockdown mutant (Chuang et al., 2015) and a ppk2 transposon mutant (Chuang et al., 2013), displayed reduced susceptibility to isoniazid, although the latter strain showed increased susceptibility to meropenem (Chuang et al., 2016). Another poly(P)-accumulating strain deficient in PPX1 showed increased susceptibility to clofazimine (Chuang et al., 2016).
Taken together, these data suggest that proper regulation of the stringent response is critical for antibiotic tolerance in M. tuberculosis.
Since the stringent response is critical for M. tuberculosis stress adaptation, virulence, and antibiotic tolerance, targeting this pathway represents an attractive approach for improving TB treatment outcomes. The two main strategies proposed so far are inhibition via small molecules (summarized in Table 1) and priming of the host immune response via DNA vaccination.

Table 1. Small molecule inhibitors of bacterial stringent response factors.
Disruption of the stringent response by perturbing poly(P) homeostasis has been proposed as a novel TB treatment strategy. To this end, inhibitors of PPK1 and PPK2 have been developed. A small in silico screen of 18 predicted inhibitors was performed for PPK1. The three lead compounds from this screen showed robust activity with inhibition constants ranging from 255 to 866nM (Shahbaaz et al., 2019). A high-throughput screen of 2,300 compounds for PPK2 inhibitors identified two lead compounds with >80% inhibition of enzyme activity at 100μM (M. Singh et al., 2016). A DNA aptamer with an IC50 of 40nM in a biochemical assay has also been designed for PPK2 (Shum et al., 2011). The finding that a ppk2 transposon mutant displayed a 4-fold increase in the MIC of isoniazid relative to the wild-type and complemented strains is concerning, however, as this effect could counteract any potential benefits of incorporating a PPK2 inhibitor into current TB treatment regimens (Chuang et al., 2013; Singh et al., 2016).
Since regulation of (p)ppGpp levels is critical for the activation of the stringent response and for bacterial adaption to stress conditions, inhibition of the bifunctional RelMtb protein has also been proposed as a treatment strategy (Wexselblatt et al., 2012; Dutta et al., 2019).
Multiple groups have sought to identify RelA inhibitors. The earliest of these inhibitors, Relacin, was identified using modeling based on the crystal structure of Rel/Spo from Streptococcus equisimilis (Wexselblatt et al., 2012). This small molecule was able to decrease (p)ppGpp levels and disrupt biofilm formation in Bacillus subtilis. It also inhibited sporulation in a dose-dependent manner (Wexselblatt et al., 2012). Deoxyguanosine-based analogs were subsequently designed to optimize the compound, and one of these had increased potency against RelA from E. coli (Wexselblatt et al., 2013). Two additional analogs (a benzoylated and an acetylated benzoylated version) were also synthesized and tested for their ability to decrease levels of (p)ppGpp in M. smegmatis (Syal et al., 2017b). The acetylated benzoylated analog had an IC50 of ~40μM and caused elongation of the cells (Syal et al., 2017b), which is in line with previous morphological characterization of mycobacteria lacking rel (Gupta et al., 2016). This compound also decreased long-term survival of M. smegmatis and inhibited biofilm formation in both M. smegmatis and M. tuberculosis (Syal et al., 2017b). The activity of Relacin and its analogs against multiple bacterial species suggest that these Rel inhibitors could have broad applications as antibiotics.
A high-throughput strategy using a truncated recombinant version of RelMtb and a fluorescence polarization assay to screen a library of over 2million compounds for enzymatic inhibition of (p)ppGpp synthesis activity yielded 178 hits with favorable physicochemical properties (Dutta et al., 2019). In whole-cell assays, one of these hits (X9) dosed at 2μM induced significant killing of nutrient-starved, wild-type M. tuberculosis, phenocopying the survival defect of the untreated ΔrelMtb mutant during nutrient starvation, and hypersensitized the nutrient-starved wild type to cell wall-active agents, decreasing the MBC of isoniazid 16-fold. These findings suggest that pharmacological inhibition of the M. tuberculosis stringent response results in direct mycobacterial killing under growth-limiting conditions, as well as reversal of tolerance to cell wall-active agents.
Several natural products have also demonstrated anti-Rel activity. Vitamin C inhibits the (p)ppGpp synthesis activity of Rel and decreases cell viability and biofilm formation in M. smegmatis (Syal et al., 2017a). The compound, 4-(4,7-dimethyl-1,2,3,4-tetrahydronaphthalene-1-yl)pentanoic acid (DMNP), a derivative of erogorgiaene, was identified as an inhibitor of both RelMsm and RelZ in M. smegmatis (Tkachenko et al., 2021). DMNP showed antibacterial activity against M. smegmatis in a whole cell assay, and overexpression of either RelMsm or RelZ reduced the compound’s antibiotic effect and increased the number of persisters. DMNP further inhibited the (p)ppGpp synthase activity of purified RelMsm and showed similar predicted binding sites for RelMsm or RelZ in molecular docking simulations (Tkachenko et al., 2021).
One FDA-approved antitubercular drug appears to target the M. tuberculosis stringent response, among other putative mechanisms of action. Pyrazinamide is a critical component of current treatment regimens for drug-susceptible TB and is formulated as a prodrug which is converted by M. tuberculosis pyrazinamidase to pyrazinoic acid. Pyrazinoic acid, but not the prodrug, was found to inhibit Rv2783c, which is involved in the RNA degradosome, a multi-enzyme complex important in RNA processing and mRNA degradation, with a Kd1 of 1.05mM and a Kd2 of 3.17mM (Njire et al., 2017). This binding was completely abolished in an D67N mutant and in enzyme isolated from naturally pyrazinamide-resistant M. smegmatis. In addition to its role in mRNA processing and degradation, Rv2783c has multifunctional activities, including polymerization and phosphorolysis of single-stranded DNA and (p)ppGpp hydrolysis in vitro. Each of these activities can be inhibited by pyrazinoic acid. The point mutation which eliminated pyrazinoic acid binding to Rv2783c resulted in a 5-fold increase in the MIC of pyrazinamide against M. tuberculosis, further underscoring the importance of Rv2783c as a target for this antitubercular drug (Njire et al., 2017).
In addition to pharmacological modulation of the stringent response, recent work has focused on potentiating host immunity to M. tuberculosis stringent response factors as a treatment-shortening strategy since this pathway is important for long-term mycobacterial survival in host tissues, and is induced during chronic TB infection in the lungs of mice (Talaat et al., 2004).
Intramuscular immunization of mice with a DNA vaccine targeting four different M. tuberculosis stringent response genes (relMtb, sigE, ppk2, and ppx) induced significant antigen-specific IgG responses, antigen-specific IFN-γ-producing CD4+ T cells, and relMtb-specific TNF-α-producing CD4+ T cells (Chuang et al., 2016). Although DNA vaccine containing all four stringent response genes was not protective against aerosol challenge with virulent M. tuberculosis, it enhanced the activity of isoniazid in a mouse model of chronic infection, significantly reducing the M. tuberculosis burden in the lungs relative to mice receiving the control DNA vaccine (Chuang et al., 2016).
Of the four stringent response genes included in the above vaccine, isoniazid exposure of M. tuberculosis-infected macrophages increased expression of relMtb most robustly (Chuang et al., 2020), prompting generation of a DNA vaccine targeting this gene alone. The relMtb DNA vaccine conferred increased susceptibility of M. tuberculosis to isoniazid relative to the control DNA vaccine in a mouse model of chronic TB infection and reduced regrowth of bacteria after cessation of antibiotic treatment (Chuang et al., 2020). Thus, RelMtb appears to be an important persistence antigen, which can be targeted by cellular immune responses in conjunction with antitubercular drugs to assist with clearance of M. tuberculosis infection from host tissues.
The stringent response is involved in bacterial adaptation to stress, contributing to survival, virulence, and drug tolerance within the host. (p)ppGpp and poly(P) play essential roles in mediating the stringent response, and mutations in M. tuberculosis proteins involved in their metabolism lead to reduced mycobacterial virulence and enhanced sensitivity to antibiotics.
Inhibition of stringent response factors such as RelMtb shows promise as an anti-tuberculosis strategy. While there are numerous reports of small molecule inhibitors of stringent response factors, further medicinal chemistry campaigns are required to optimize the activity, bioavailability, and pharmacokinetics of these compounds. Additionally, relapse studies in clinically relevant animal models are needed to determine whether the addition of a stringent response inhibitor to current TB treatment regimens can shorten the time required to achieve a stable cure. Based on the established role of the stringent response in M. tuberculosis antibiotic tolerance, we predict that addition of a stringent response modulator, whether a small molecule inhibitor or a therapeutic vaccine enhancing antigen-specific T cell responses, may reduce the number of persisters, thereby promoting more rapid eradication of infection from host tissues.
Novel strategies targeting the stringent response pathway are predicted to have treatment-shortening potential against both drug-susceptible and drug-resistant M. tuberculosis. This is a particularly important consideration for multidrug-resistant (MDR) TB and extensively drug-resistant (XDR) TB, since standard regimens to treat these infections have markedly reduced efficacy and require more than 18months of continuous therapy. Thus, the stringent response represents a promising target for the development of novel antimycobacterial strategies.
CD and PK designed and directed the project. CD and SW performed the literature search. CD, SW, and PK wrote the article. All authors contributed to the article and approved the submitted version.
This work was supported by NIH grants R01AI148710, R21AI140860, and K24AI143447 to PK. The funding source had no role in the design and conduct of the study.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adams, K. N., Szumowski, J. D., and Ramakrishnan, L. (2014). Verapamil, and its metabolite norverapamil, inhibit macrophage-induced, bacterial efflux pump-mediated tolerance to multiple anti-tubercular drugs. J. Infect. Dis. 210, 456–466. doi: 10.1093/INFDIS/JIU095
Adams, K. N., Takaki, K., Connolly, L. E., Wiedenhoft, H., Winglee, K., Humbert, O., et al. . (2011). Drug tolerance in replicating mycobacteria mediated by a macrophage-induced efflux mechanism. Cell 145, 39–53. doi: 10.1016/J.CELL.2011.02.022
Avarbock, D., Avarbock, A., and Rubin, H. (2000). Differential regulation of opposing Rel(Mtb) activities by the aminoacylation state of a tRNA·ribosome·mRNA·Rel(Mtb) complex. Biochemistry 39, 11640–11648. doi: 10.1021/bi001256k
Avarbock, A., Avarbock, D., Teh, J.-S., Buckstein, M., Wang, Z.-M., and Rubin, H. (2005). Functional Regulation of the Opposing (p)ppGpp synthetase/hydrolase activities of Rel Mtb from Mycobacterium tuberculosi. Biochemistry 44, 9913–9923. doi: 10.1021/bi0505316
Avarbock, D., Salem, J., Li, L. S., Wang, Z. M., and Rubin, H. (1999). Cloning and characterization of a bifunctional RelA/SpoT homologue from Mycobacterium tuberculosis. Gene 233, 261–269. doi: 10.1016/S0378-1119(99)00114-6
Balaban, N. Q., Gerdes, K., Lewis, K., and McKinney, J. D. (2013). A problem of persistence: still more questions than answers? Nat. Rev. Microbiol. 11, 587–591. doi: 10.1038/nrmicro3076
Battesti, A., Majdalani, N., and Gottesman, S. (2011). The RpoS-mediated general stress response in Escherichia coli. Annu. Rev. Microbiol. 65, 189–213. doi: 10.1146/annurev-micro-090110-102946
Black, P. A., Warren, R. M., Louw, G. E., Helden, P. D.van , Victor, T. C., and Kana, B. D. (2014). Energy metabolism and drug efflux in Mycobacterium tuberculosis. Antimicrob. Agents Chemother., 58, 2491–2503. doi: 10.1128/AAC.02293-13
Brauner, A., Fridman, O., Gefen, O., and Balaban, N. Q. (2016). Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 14, 320–330. doi: 10.1038/nrmicro.2016.34
Choi, M. Y., Wang, Y., Wong, L. L. Y., Lu, B. T., Chen, W. Y., Huang, J. D., et al. . (2012). The two PPX-GppA homologues from Mycobacterium tuberculosis have distinct biochemical activities. PLoS One 7:e42561. doi: 10.1371/journal.pone.0042561
Chuang, Y. M., Bandyopadhyay, N., Rifat, D., Rubin, H., Bader, J. S., and Karakousis, P. C. (2015). Deficiency of the novel exopolyphosphatase Rv1026/PPX2 leads to metabolic downshift and altered cell wall permeability in Mycobacterium tuberculosis. MBio 6:e02428. doi: 10.1128/mBio.02428-14
Chuang, Y. M., Belchis, D. A., and Karakousis, P. C. (2013). The polyphosphate kinase gene ppk2 is required for Mycobacterium tuberculosis inorganic polyphosphate regulation and virulence. MBio 4, e00039–e00013. doi: 10.1128/mBio.00039-13
Chuang, Y. M., Dutta, N. K., Gordy, J. T., Campodónico, V. L., Pinn, M. L., Markham, R. B., et al. . (2020). Antibiotic treatment shapes the antigenic environment during chronic TB infection, offering novel targets for therapeutic vaccination. Front. Immunol. 11:680. doi: 10.3389/fimmu.2020.00680
Chuang, Y. M., Dutta, N. K., Hung, C. F., Wu, T. C., Rubin, H., and Karakousis, P. C. (2016). Stringent response factors PPX1 and PPK2 play an important role in Mycobacterium tuberculosis metabolism, biofilm formation, and sensitivity to isoniazid in vivo. Antimicrob. Agents Chemother. 60, 6460–6470. doi: 10.1128/AAC.01139-16
Dahl, J. L., Kraus, C. N., Boshoff, H. I. M., Doan, B., Foley, K., Avarbock, D., et al. . (2003). The role of RelMtb-mediated adaptation to stationary phase in long-term persistence of Mycobacterium tuberculosis in mice. Proc. Natl. Acad. Sci. U. S. A. 100, 10026–10031. doi: 10.1073/pnas.1631248100
Dutta, N. K., Klinkenberg, L. G., Vazquez, M. J., Segura-Carro, D., Colmenarejo, G., Ramon, F., et al. . (2019). Inhibiting the stringent response blocks Mycobacterium tuberculosis entry into quiescence and reduces persistence. Sci. Adv. 5:eaav2104. doi: 10.1126/sciadv.aav2104
Frank, D. J., Horne, D. J., Dutta, N. K., Shaku, M. T., Madensein, R., Hawn, T. R., et al. . (2019). Remembering the host in tuberculosis drug development. J. Infect. Dis. 219, 1518–1524. doi: 10.1093/infdis/jiy712
Gibson, S. E. R., Harrison, J., and Cox, J. A. G. (2018). Modelling a silent epidemic: a review of the in vitro models of latent tuberculosis. Pathogens 7:88. doi: 10.3390/PATHOGENS7040088
Goossens, S. N., Sampson, S. L., and Van Rie, A. (2021). Mechanisms of drug-induced tolerance in Mycobacterium tuberculosis. Clin. Microbiol. Rev. 34, 1–21. doi: 10.1128/CMR.00141-20
Gupta, K. R., Baloni, P., Indi, S. S., and Chatterji, D. (2016). Regulation of growth, cell shape, cell division, and gene expression by second messengers (p)ppGpp and cyclic Di-GMP in Mycobacterium smegmatis. J. Bacteriol. 198, 1414–1422. doi: 10.1128/JB.00126-16
Harms, A., Maisonneuve, E., and Gerdes, K. (2016). Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 354, aaf4268. doi: 10.1126/SCIENCE.AAF4268
Harper, J., Skerry, C., Davis, S. L., Tasneen, R., Weir, M., Kramnik, I., et al. . (2012). Mouse model of necrotic tuberculosis granulomas develops hypoxic lesions. J. Infect. Dis. 205, 595–602. doi: 10.1093/INFDIS/JIR786
Hoffman, S. B. (2001). Mechanisms of antibiotic resistance. Compend. Contin. Educ. Pract. Vet. 4, 464–472. doi: 10.1128/microbiolspec.vmbf-0016-2015
Jang, J., Kim, R., Woo, M., Jeong, J., Park, D. E., Kim, G., et al. . (2017). Efflux attenuates the antibacterial activity of Q203 in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 61, e02637–e02616. doi: 10.1128/AAC.02637-16
Kana, B. D., Karakousis, P. C., Parish, T., and Dick, T. (2014). Future target-based drug discovery for tuberculosis? Tuberculosis 94, 551–556. doi: 10.1016/j.tube.2014.10.003
Karakousis, P. C. (2009). Mechanisms of action and resistance of antimycobacterial agents. Anti. Drug Res., 271–291. doi: 10.1007/978-1-59745-180-2_24
Karakousis, P. C., Yoshimatsu, T., Lamichhane, G., Woolwine, S. C., Nuermberger, E. L., Grosset, J., et al. . (2004). Dormancy phenotype displayed by extracellular Mycobacterium tuberculosis within artificial granulomas in mice. J. Exp. Med. 200, 647–657. doi: 10.1084/jem.20040646
Klinkenberg, L. G., Lee, J., Bishai, W. R., and Karakousis, P. C. (2010). The stringent response is required for full virulence of Mycobacterium tuberculosis in Guinea pigs. J. Infect. Dis. 202, 1397–1404. doi: 10.1086/656524
Klinkenberg, L. G., Sutherland, L. A., Bishai, W. R., and Karakousis, P. C. (2008). Metronidazole lacks activity against Mycobacterium tuberculosis in an in vivo hypoxic granuloma model of latency. J. Infect. Dis. 198, 275–283. doi: 10.1086/589515
Kuroda, A. (2006). A polyphosphate-lon protease complex in the adaptation of Escherichia coli to amino acid starvation. Biosci. Biotechnol. Biochem. 70, 325–331. doi: 10.1271/bbb.70.325
Levin-Reisman, I., Ronin, I., Gefen, O., Braniss, I., Shoresh, N., and Balaban, N. Q. (2017). Antibiotic tolerance facilitates the evolution of resistance. Science 355, 826–830. doi: 10.1126/science.aaj2191
Maisonneuve, E., and Gerdes, K. (2014). Molecular mechanisms underlying bacterial persisters. Cell 157, 539–548. doi: 10.1016/j.cell.2014.02.050
Namugenyi, S. B., Aagesen, A. M., Elliott, S. R., and Tischler, A. D. (2017). Mycobacterium tuberculosis PhoY proteins promote persister formation by mediating Pst/SenX3-RegX3 phosphate sensing. MBio 8, e00494–e00417. doi: 10.1128/MBIO.00494-17
Njire, M., Wang, N., Wang, B., Tan, Y., Cai, X., Liu, Y., et al. . (2017). Pyrazinoic acid inhibits a bifunctional enzyme in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 61, e00070–e00017. doi: 10.1128/AAC.00070-17
Primm, T. P., Andersen, S. J., Mizrahi, V., Avarbock, D., Rubin, H., and Barry, C. E. (2000). The stringent response of Mycobacterium tuberculosis is required for long-term survival. J. Bacteriol. 182, 4889–4898. doi: 10.1128/JB.182.17.4889-4898.2000
Prusa, J., Zhu, D. X., and Stallings, C. L. (2018). The stringent response and Mycobacterium tuberculosis pathogenesis. Pathog. Dis. 76:54. doi: 10.1093/femspd/fty054
Rao, N. N., Gómez-García, M. R., and Kornberg, A. (2009). Inorganic polyphosphate: essential for growth and survival. Annu. Rev. Biochem. 78, 605–647. doi: 10.1146/annurev.biochem.77.083007.093039
Rifat, D., Bishai, W. R., and Karakousis, P. C. (2009). Phosphate depletion: a novel trigger for mycobacterium tuberculosis persistence. J. Infect. Dis. 200, 1126–1135. doi: 10.1086/605700
Ronneau, S., and Hallez, R. (2019). Make and break the alarmone: regulation of (p)ppGpp synthetase/hydrolase enzymes in bacteria. FEMS Microbiol. Rev. 43, 389–400. doi: 10.1093/femsre/fuz009
Sala, A., Bordes, P., and Genevaux, P. (2014). Multiple toxin-antitoxin systems in Mycobacterium tuberculosis. Toxins 6, 1002–1020. doi: 10.3390/toxins6031002
Sanyal, S., Banerjee, S. K., Banerjee, R., Mukhopadhyay, J., and Kundu, M. (2013). Polyphosphate kinase 1, a central node in the stress response network of Mycobacterium tuberculosis, connects the two-component systems MprAB and SenX3-RegX3 and the extracytoplasmic function sigma factor, sigma E. Microbiology 159, 2074–2086. doi: 10.1099/mic.0.068452-0
Shahbaaz, M., Nkaule, A., and Christoffels, A. (2019). Designing novel possible kinase inhibitor derivatives as therapeutics against Mycobacterium tuberculosis: an in silico study. Sci. Rep. 9:4405. doi: 10.1038/s41598-019-40621-7
Shum, K. T., Lui, E. L. H., Wong, S. C. K., Yeung, P., Sam, L., Wang, Y., et al. . (2011). Aptamer-mediated inhibition of mycobacterium tuberculosis polyphosphate kinase 2. Biochemistry 50, 3261–3271. doi: 10.1021/bi2001455
Singal, B., Balakrishna, A. M., Nartey, W., Manimekalai, M. S. S., Jeyakanthan, J., and Grüber, G. (2017). Crystallographic and solution structure of the N-terminal domain of the Rel protein from Mycobacterium tuberculosis. FEBS Lett. 591, 2323–2337. doi: 10.1002/1873-3468.12739
Singh, R., Singh, M., Arora, G., Kumar, S., Tiwari, P., and Kidwai, S. (2013). Polyphosphate deficiency in Mycobacterium tuberculosis is associated with enhanced drug susceptibility and impaired growth in Guinea pigs. J. Bacteriol. 195, 2839–2851. doi: 10.1128/JB.00038-13
Singh, M., Tiwari, P., Arora, G., Agarwal, S., Kidwai, S., and Singh, R. (2016). Establishing virulence associated polyphosphate kinase 2 as a drug target for Mycobacterium tuberculosis. Sci. Rep. 6:26900. doi: 10.1038/srep26900
Spoering, A. L., Vulić, M., and Lewis, K. (2006). GlpD and PlsB participate in persister cell formation in Escherichia coli. J. Bacteriol. 188, 5136–5144. doi: 10.1128/JB.00369-06
Sureka, K., Dey, S., Datta, P., Singh, A. K., Dasgupta, A., Rodrigue, S., et al. . (2007). Polyphosphate kinase is involved in stress-induced mprAB-sigE-rel signalling in mycobacteria. Mol. Microbiol. 65, 261–276. doi: 10.1111/j.1365-2958.2007.05814.x
Sureka, K., Sanyal, S., Basu, J., and Kundu, M. (2009). Polyphosphate kinase 2: a modulator of nucleoside diphosphate kinase activity in mycobacteria. Mol. Microbiol. 74, 1187–1197. doi: 10.1111/j.1365-2958.2009.06925.x
Syal, K., Bhardwaj, N., and Chatterji, D. (2017a). Vitamin C targets (p)ppGpp synthesis leading to stalling of long-term survival and biofilm formation in Mycobacterium smegmatis. FEMS Microbiol. Lett. 364:fnw282. doi: 10.1093/femsle/fnw282
Syal, K., Flentie, K., Bhardwaj, N., Maiti, K., Jayaraman, N., Stallings, C. L., et al. . (2017b). Synthetic (p)ppGpp analogue is an inhibitor of stringent response in mycobacteria. Antimicrob. Agents Chemother. 61, e00443–e00417. doi: 10.1128/AAC.00443-17
Szumowski, J. D., Adams, K. N., Edelstein, P. H., and Ramakrishnan, L. (2013). Antimicrobial efflux pumps and Mycobacterium tuberculosis drug tolerance: evolutionary considerations. Curr. Top. Microbiol. Immunol. 374, 81–108. doi: 10.1007/82_2012_300
Talaat, A. M., Lyons, R., Howard, S. T., and Johnston, S. A. (2004). The temporal expression profile of Mycobacterium tuberculosis infection in mice. Proc. Natl. Acad. Sci. U. S. A. 101, 4602–4607. doi: 10.1073/PNAS.0306023101
Talwar, S., Pandey, M., Sharma, C., Kutum, R., Lum, J., Carbajo, D., et al. . (2020). Role of VapBC12 toxin-antitoxin locus in cholesterol-induced mycobacterial persistence. mSystems 5, e00855–e00820. doi: 10.1128/msystems.00855-20
Thayil, S. M., Morrison, N., Schechter, N., Rubin, H., and Karakousis, P. C. (2011). The role of the novel exopolyphosphatase MT0516 in Mycobacterium tuberculosis drug tolerance and persistence. PLoS One 6:e28076. doi: 10.1371/journal.pone.0028076
Tkachenko, A. G., Kashevarova, N. M., Sidorov, R. Y., Nesterova, L. Y., Akhova, A. V., Tsyganov, I. V., et al. . (2021). A synthetic diterpene analogue inhibits mycobacterial persistence and biofilm formation by targeting (p)ppGpp synthetases. Cell Chem. Biol. doi: 10.1016/j.chembiol.2021.01.018 [Epub ahead of print]
Traxler, M. F., Summers, S. M., Nguyen, H.-T., Zacharia, V. M., Smith, J. T., and Conway, T. (2008). The global, ppGpp-mediated stringent response to amino acid starvation in Escherichia coli. Mol. Microbiol. 68, 1128–1148. doi: 10.1111/J.1365-2958.2008.06229.X
Weiss, L. A., and Stallings, C. L. (2013). Essential roles for mycobacterium tuberculosis rel beyond the production of (p)ppGpp. J. Bacteriol. 195, 5629–5638. doi: 10.1128/JB.00759-13
Westblade, L. F., Errington, J., and Dörr, T. (2020). Antibiotic tolerance. PLoS Pathog. 16:e1008892. doi: 10.1371/journal.ppat.1008892
Wexselblatt, E., Kaspy, I., Glaser, G., Katzhendler, J., and Yavin, E. (2013). Design, synthesis and structure-activity relationship of novel Relacin analogs as inhibitors of Rel proteins. Eur. J. Med. Chem. 70, 497–504. doi: 10.1016/j.ejmech.2013.10.036
Wexselblatt, E., Oppenheimer-Shaanan, Y., Kaspy, I., London, N., Schueler-Furman, O., Yavin, E., et al. . (2012). Relacin, a novel antibacterial agent targeting the stringent response. PLoS Pathog. 8:e1002925. doi: 10.1371/journal.ppat.1002925
Keywords: Mycobacterium tuberculosis, stringent response, antibiotic tolerance, (p)ppGpp, hyperphosphorylated guanosine, inorganic polyphosphate, small molecule inhibitor, vaccination
Citation: Danchik C, Wang S and Karakousis PC (2021) Targeting the Mycobacterium tuberculosis Stringent Response as a Strategy for Shortening Tuberculosis Treatment. Front. Microbiol. 12:744167. doi: 10.3389/fmicb.2021.744167
Edited by:
Sandeep Sharma, Lovely Professional University, IndiaReviewed by:
Julien Briffotaux, Shenzhen Nanshan Center for Chronic Disease Control, ChinaCopyright © 2021 Danchik, Wang and Karakousis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Petros C. Karakousis, cGV0cm9zQGpobWkuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.