
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 19 July 2021
Sec. Microbial Physiology and Metabolism
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.707086
 Jose Carrasco-Pujante1†
Jose Carrasco-Pujante1† Carlos Bringas2†Iker Malaina3
Carlos Bringas2†Iker Malaina3 Maria Fedetz4
Maria Fedetz4 Luis Martínez3,5
Luis Martínez3,5 Gorka Pérez-Yarza2María Dolores Boyano2
Gorka Pérez-Yarza2María Dolores Boyano2 Mariia Berdieva6Andrew Goodkov6José I. López7
Mariia Berdieva6Andrew Goodkov6José I. López7 Shira Knafo1,8*
Shira Knafo1,8* Ildefonso M. De la Fuente3,9*
Ildefonso M. De la Fuente3,9*The capacity to learn new efficient systemic behavior is a fundamental issue of contemporary biology. We have recently observed, in a preliminary analysis, the emergence of conditioned behavior in some individual amoebae cells. In these experiments, cells were able to acquire new migratory patterns and remember them for long periods of their cellular cycle, forgetting them later on. Here, following a similar conceptual framework of Pavlov’s experiments, we have exhaustively studied the migration trajectories of more than 2000 individual cells belonging to three different species: Amoeba proteus, Metamoeba leningradensis, and Amoeba borokensis. Fundamentally, we have analyzed several relevant properties of conditioned cells, such as the intensity of the responses, the directionality persistence, the total distance traveled, the directionality ratio, the average speed, and the persistence times. We have observed that cells belonging to these three species can modify the systemic response to a specific stimulus by associative conditioning. Our main analysis shows that such new behavior is very robust and presents a similar structure of migration patterns in the three species, which was characterized by the presence of conditioning for long periods, remarkable straightness in their trajectories and strong directional persistence. Our experimental and quantitative results, compared with other studies on complex cellular responses in bacteria, protozoa, fungus-like organisms and metazoans that we discus here, allow us to conclude that cellular associative conditioning might be a widespread characteristic of unicellular organisms. This new systemic behavior could be essential to understand some key principles involved in increasing the cellular adaptive fitness to microenvironments.
An essential question of cellular behavior is to know if individual cells are capable of learning, thereby acquiring new adaptive systemic patterns to respond to changes in the external medium.
Associative learning (the emergence of behavioral modifications associated with new specific external stimuli) and memory are fundamental cognitive properties of multicellular organisms endowed with nervous systems to obtain critical information for survival, developing new complex behavior by the association of different stimuli and/or responses. This property is ubiquitous in many species, from cephalopods to humans (Hawkins and Byrne, 2015). Recently, we observed in a preliminary study the emergence of cellular conditioning in Amoeba proteus sp., which modified their migratory behavior by an association of external stimuli acquiring new systemic responses (De la Fuente et al., 2019a).
In continuation of this research and following a similar conceptual framework of Pavlov’s experiments, we have studied here the migration trajectories of more than 2000 individual cells belonging to three different species (Amoeba proteus, sp., Metamoeba leningradensis, sp. and Amoeba borokensis, sp.). First, we have analyzed the locomotion movements under three external conditions: in absence of stimuli, in an electric field (galvanotaxis), and under chemotactic gradient (chemotaxis). Next, we have conditioned a great number of cells, applying simultaneously galvanotactic and chemotactic stimuli (placing a specific peptide in the anode), and we verified that most of them (78%) can acquire a new systemic behavior (movement to the anode when the habitual behavior under galvanotactic conditions is to go to the cathode). It should be noted that cells were able to keep this new conditioned response for long periods of their cellular cycle, forgetting it later. Finally, we have quantitatively analyzed the behavior of these conditioned cells by studying the intensity of the locomotion responses, the directionality persistence, the persistence times, and several kinematic properties of the cell migration trajectories under conditioning such as the total distance traveled, the directionality ratio and the average speed. This exhaustive quantitative analysis unequivocally showed that the cellular conditioned behavior is robust, and according to the parameters here analyzed, these three species presented a similar motion structure characterized by the presence of conditioning for long periods, strong straightness in their trajectories, and high directional persistence. So, after the induction process (simultaneous galvanotaxis and chemotaxis), the three species exhibited the capacity to generate a new type of systemic motility pattern that prevailed for 40 min on average.
The organisms analyzed in our study are free-living naked lobose amoebae, the freshwater predators, belonging to the large and diverse supergroup Amoebozoa. Despite the seeming “simplicity” of these unicellular eukaryotes, the studies performed in Amoeba became a necessary basis for understanding various processes and phenomena, which are in the focus of attention of modern cellular and molecular biology, even if they are performed in organisms completely unrelated to protists phylogenetically distant to Amoebozoa. Thus, data on their locomotion mode, feeding behavior, and cell cycle features contributed to the understanding of the structure and function of metazoan (including human) cells, also those of cancer (Yudin, 1990; Jeon, 1995; Goodkov et al., 2014; Berdieva et al., 2019).
The amoebae’s life is a life in motion; they crawl over the substrates forming large or small protrusions searching for food and moving in response to external stimuli. Amoebae possess photo-, geo-, galvano-, and chemotaxis (Grebecki, 1980; Korohoda et al., 1997, 2000). Their locomotion’s type, known as “amoeboid movement,” is the most common locomotion mode among eukaryotic cells. Cells of multicellular organisms, including vertebrates, demonstrate a similar pattern of locomotion. Since the beginning of the 20th century, several hypotheses have been proposed in an attempt to explain it (reviewed in Smirnov, 2008); nevertheless, the essential mechanisms of systemic amoeboid movement remain to be elucidated.
Initial studies of the phagocytic ability of eukaryotic cells were preceded by investigations in Amoeba. Later, this activity was characterized in cells of the immune system of multicellular organisms, which are capable to engulf hostile cells. Interestingly, cancer cells also showed a clear feeding behavior primarily oriented against neighboring cells, live or dead (Lugini et al., 2006; Fais and Fauvarque, 2012).
Amoeba is an obligatory agamic organism that demonstrates a very special cell cycle, during which a strategy of the so-called cyclic polyploidy (alternation of polyploidization and depolyploidization stages preceding reproduction) is implemented (Demin et al., 2019). Such cycles of ploidy were described in cell cycles of some other unicellular eukaryotes. An analogous strategy is observed in mammalian cancer cells. Their similarity to unicellular organisms, especially amoebae, is considered a part of the atavistic theory of carcinogenesis, which suggests that cancer cells switch to a “selfish” lifestyle like unicellular organisms (Huna et al., 2015; Berdieva et al., 2019).
The experimental single-celled organisms for our study were not chosen randomly, but for a variety of reasons, which are briefly listed below. First, free-living amoebas - representatives of the Amoebidae family (Amoeba proteus and related species) - are in fact the only unicellular eukaryotes in which such concepts as “morphology” (body shape, appearance), “way of movement in space” (locomotion) and “behavior” is largely the same due to the specific type of cellular organization of these protists. But at the same time, they remain different organisms - species and genera, with their own biology - which is fundamentally important in a comparative analysis of the physiological characteristics of any biological objects. Secondly, Amoeba proteus (and related species), being a historically traditional model object of cell biology, are well characterized in many respects, including the mechanisms of cell motility, taxis, behavior, etc. Thirdly, the relatively large size of the cells and the nature of movement - movement over the substrate using the formation of directed discrete pseudopodia - provide an opportunity for subtle visual observation of all changes in the behavior of these unicellular organisms. Finally, fourthly, these amoebas are free-living freshwater unicellular organisms with a worldwide distribution. For them, there is a long-established standard laboratory cultivation technique that allows you to easily and quickly obtain the amount of cellular material required for experimental research.
The different types of unicellular organisms, even those that are very close, cannot fail to have, in addition to morphological, some differences and physiological characteristics, for example, in food or environmental preferences. Biochemical analysis showed, in particular, that isozyme spectra of acid phosphatase, glucose-6-phosphate dehydrogenase and esterases in all three of our species differ significantly, while strains of the same A. proteus species, but of different origin, do not have these differences (Sopina, 2000). And here it is just important that, although they are different species, they demonstrate basic similarities in conditioned behavioral responses.
In the present work, the results of our quantitative study were compared to other experimental findings in cellular behavior with complex responses in bacteria, protozoa, fungus-like organisms and metazoans, which allow us to conclude that associative conditioning might be a widespread characteristic of unicellular organisms.
The cellular capacity of learning and therefore acquiring new adaptive systemic patterns of response to changes in the external medium could represent a fundamental process for cellular adaptation as an evolutionary mechanism through which unicellular organisms increase their fitness in their respective habitats.
How cells efficiently regulate their locomotion movements is still unclear. A novel knowledge of the control of cellular systemic behavior such as cellular conditioning can define a new framework in the understanding of the mechanisms underlying the complex processes involved in the adaptive capacity of cells to the external medium.
Amoeba proteus (Carolina Biological Supply Company, Burlington, NC.Item # 131306) were cultured alongside Chilomonas as food organisms at around 21°C on Chalkley’s simplified medium (NaCl, 1.4 mM; KCl, 0.026 mM; CaCl2, 0.01 mM) (Carolina Biological Supply Company Item #131734), and previously baked wheat corns. Metamoeba leningradensis (Culture Collection of Algae and Protozoa, Oban, Scotland, United Kingdom, CCAP catalog number 1503/6) were cultured in the same conditions as Amoeba proteus. A. borokensis (Amoebae Cultures Collection of Institute of Cytology, ACCIC, Saint Petersburg, Russia) were cultured in Prescott & Carrier’s media, and supplied by 0.5 ml of Chilomonas sp., (Carolina Biological Supply Company Item #131734) and Colpydium sp. (ACCIC, Saint Petersburg, Russia, Goodkov et al., 2014) as food organisms twice a week; these two organisms were also cultured in Prescott & Carrier’s media (Prescott and Carrier, 1964) with flamed rice grains.
The experiments in this study were performed in a device shown in Supplementary Figures 1, 2 that was composed of two blocks of electrophoresis, 17.5 cm long each (Biorad Mini-Sub cell GT), a power supply (Biorad powerbank s2000), two bridges of agar (2% agar in 0.5 N KCl, 10-12 cm long), and a chamber made of glass covers and slides.
The first block was plugged to the power supply and connected to the second one by the two agar bridges this way preventing the direct contact of the second electrophoretic block with the power supply.
The agar bridges were necessarily new in every experiment to prevent contamination. Also, for this purpose there were different electrophoresis blocks, one was used only for chemotaxis with peptide, the other was used only for galvanotaxis without peptide, which in any case were thoroughly cleaned before the next experiment, just like the experimental chambers were reassembled each time.
The blocks were composed of three parts: two wells filled with the conductive medium (Chalkley’s simplified medium) and an elevated platform. The experimental glass structure was placed on the top of the elevated platform of the second block. This way, the implementation of a laminar flux was possible when the chamber was closed. Opening the chamber allowed to get it and extract the cells.
Standard glass slides and covers (total 4 pieces) composed the experimental chamber, that is, a 75 × 25 mm slide and three small pieces resulting from a careful cut of three standard long 40 × 24 × 0.1 mm cover glasses (Supplementary Figures 1, 2).
Three one-use-only cover glasses were trimmed to get one central piece of 3 × 24 × 0.1 mm and two lateral sliding pieces of 40 × 24 × 0.1 mm each.
This glass supports the central and sliding parts of the structure (experimental chamber) (Supplementary Figures 1, 2). We used silicone to glue two covers to a glass slide. This structure was left to dry for 24 h. Then, the protruding portions of the two cover slides (60 × 20 × 0.1 mm) were trimmed, leaving two 60 × 4 × 0.1 mm glass strips, which act as the lateral walls of the experimental chamber.
The resulting modified glass slide was placed on the top of the elevated central platform of the second electrophoresis block. Under it, an oil drop was placed to prevent that the Chalkley’s simplified medium can pass under the glass structure. The oil must fill the entire surface of the glass structure so that no liquid can pass through. On the top of the modified slide the central piece and the two sliding lateral glasses were placed.
Amoebae were placed under the central piece of the experimental glass chamber in 30 μl of clean Chalkley’s simplified medium. The process must be performed rapidly; otherwise the amoebae will adhere to the tip of the pipette and damage their cell membrane.
Cells were left to rest for around 2 min on the experimental chamber prior to every experiment. This way, they were allowed to adapt to the new environment, firmly attach to the glass surface and begin moving around the experimental chamber. The wells of both electrophoretic blocks were carefully filled with Chalkley’s simplified medium until reaching the base of the modified slide but not the sliding glasses. Then, the sliding glasses were pushed down with a pipette to put them in contact with the Chalkley’s simplified medium allowing the medium to sprawl by surface tension. Next, the two sliding glasses were gently moved alongside the longitudinal dimension of the experimental glass structure until touching the central glass piece to create a laminar flux.
When required, always before the establishment of the electric field, 750 μl of 2 × 10–4 M N-Formylmethionyl-leucyl-phenylalanine (nFMLP) peptide solution was added to the Chalkley simplified medium (75 ml) in the well corresponding to the positive pole of the second electrophoretic block. The same volume (750 μl) of Chalkley’s medium was removed from this well before the peptide was added. Keeping the same volume of medium in both wells is necessary to avoid harmful medium flows through the experimental chamber, which would affect the amoebae’s behavior.
Cells were rescued from the chamber by sliding the top lateral pieces of the glass structure using the tip of a micropipette.
Amoeba proteus, M. leningradensis, and A. borokensis may show different physiological variations due to slightly differing culture conditions. Prior any experiments, cells to be studied were starved for 24 h in clean axenic Chalkley simplified medium in the absence of any external stimuli. Only cells that were actively moving, did not display many thin or upward pseudopodia and showed a spindled or oval shape were selected for the experiments.
All cells analyzed were washed in Chalkley simplified medium prior to experimentation and then placed under the central piece of the glass chamber. The glass chamber was never closed before all amoebae appeared to be firmly attached to substrate, what happened in about a minute on average. The experiments were always performed using no more than 10 cells. Consecutive experiments tended to use always a smaller number of cells, since only those cells showing a potentially conditioned behavior were tested further. M. leningradensis were more difficult to handle than A. proteus, they showed less adherence to the glass and more tendency to stick to the pipette tip.
All experiments implying the use of galvanotaxis were traumatic for cells, and could alter their response to following experimentation. To minimize harmful effects, the amoebae must always be handled with extreme caution, only healthy and fit amoebae (as previously described in this section) shall be used and the electricity parameters’ value kept within the provided optimal ranges during experimentation.
During the investigation, some experimental replicates had to be discarded because of the following reasons: (i) the flow was of such intensity that it directly and by itself prevented cell adhesion, (ii) mechanical problems such as a displacement of the image recording system were identified, (iii) prior to any electrical or chemical stimulus, cells presented lack of adhesion, abnormal motility behaviors (non-existent or circular), or even spontaneous lysis. The deviations from standard culture conditions can affect cell motility, adhesion etc. However, after 24-48 h of standard culture, these cells recover their natural behavior. Only such experiments that met one or more of these conditions were discarded. All other experimental replicates have been reported. In our analysis, the percentage of discarded experiments was about 9.8%.
The electric field applied to the first electrophoresis block was conducted to the second by the two agar bridges. Direct measurements in the second block close to the experimental chamber made it possible to control and maintain the current between 58.5 and 60 V (334-342 mV/mm).
To ensure that the intensity required to successfully perform the experiments was supplied, it is important that during construction of the experimental chamber that the longitudinal strips were ≤4 mm in width and ≥0.1 mm longer. Height of the longitudinal strips was adapted by modifying the amount of silicone used to glue the cover slides pieces to the glass slide while constructing the chamber. Additionally, in order to have an immediate control of the intensity of the current, a variable resistance of 1 megohm was installed in series, and after it a milliampere meter (Supplementary Figure 3). During the experiments, by modifying the variable resistance in real-time we always kept the intensity of the electric field between the following values: 70-74 μA for Amoeba proteus; 70-80 μA for Metamoeba leningradensis; and 68-75 μA for Amoeba borokensis.
The power supply was turned-off after 30 min of cell exposure. In the experiments in which a chemotactic gradient was not used the electrophoresis block utilized had never been in contact with any chemotactic substance.
A 5 min galvanotaxis test was performed prior to any experiment that required the use of galvanotaxis. It is highly advisable to perform this check, as during experimentation we identified some cell populations that would not respond to galvanotaxis or even responded in an inverted manner, following to the anode.
Only when all the cells in every experiment were attached to the glass surface, the laminar flux was established and the peptide nFMLP introduced in the left well of the second electrophoresis block. 1 min later, the electric power supply was turned on. The process took 30 min and, after that, the power supply was turned off and only the cells that had moved toward the anode were rescued and placed in a Petri dish with clean Chalkley simplified medium for 10 min prior to future experiments.
Using the ranges of electricity other than those specified in the previous section will give abnormal results as, for example, the ones depicted in Supplementary Figure 4. The electric field’s intensity values in that experiment ranged between 83 and 90 μA.
After establishing the laminar flux, 750 μl of 2 × 10–4 M nFMLP (#F3506, Sigma-Aldrich) was added. The final concentration of the peptide solution was 2 × 10–6M. Immediately after introducing the peptide it was carefully mixed in the medium contained in the left well using a pipette until the amoebae started to move. Cell movements were recorded for 30 min.
The nFMLP peptide gradient was confirmed by measuring its concentration in the middle of the experimental chamber. 4 mM fluorescein-tagged peptide (#F1314, Invitrogen) was loaded in the left side of the set-up. The set-up was prepared as usual but leaving a small opening (the size of the tip of a 50-200 μL micropipette) between the sliding cover glasses and the central glass piece. This opening allowed to get 60 μL samples from the central part of the laminar flux in the chamber at 0, 2, 5, 10, 15, 20, and 30 min following the establishment of the laminar flux. The peptide concentration was calculated extrapolating the values from a standard curve in which the concentration of the fluorescein-tagged peptide was known (Supplementary Figure 5). All the measurements were performed twice and the experiment was repeated three times. The fluorescence was measured in 96 well glass bottom black plates (P96-1.5H-N, in vitro Scientific) employing a SynergyHTX plate reader (Biotek) at Excitation/Emission wavelengths of 460/528 following standard laboratory techniques as described by Green and Sambrook (Green and Sambrook, 2012).
Cell trajectories were recorded using a digital camera linked to a SM-2T stereomicroscope. Images were taken every 10 s for a minimum of 30 min (180 frames). Only the first 30 min were quantified. As suggested by Hilsenbeck et al. (2016), we did a manual tracking using the TrackMate software in ImageJ (1Tinevez et al., 2017), given the inaccuracies of automated tracking softwares (Hilsenbeck et al., 2016). Each track corresponds to the movements of an individual amoeba.
To estimate the significance of our results, we studied first if the distribution of cosines of angles came from a normal distribution, by applying the Kolmogorov-Smirnov test for single samples. Since the normality was rejected, the groups of cosines were compared by non-parametric tests, for groups by Kruskal-Wallis test, and pairs by the Wilcoxon rank-sum test, and therefore, the results were depicted as median/IQ instead of as mean ± SD. Besides the p-Values, we have reported the χ2 statistics and the Z statistics.
All the experiments were performed in a specific set-up (Supplementary Figures 1, 2) consisting of two standard electrophoresis blocks, a power supply, two agar bridges and a structure made from a standard glass slide and covers (the experimental glass chamber) commonly used in Pathology-Cytology Laboratories (“Method” Details).
1. Cellular displacement in the absence of stimuli.
First, we observed that the migration of 169 individual cells belonging to the three species, Amoeba proteus, Metamoeba leningradensis and Amoeba borokensis, under the absence of stimuli, exhibited random locomotion patterns through which the amoebae explored practically all the directions of the experimentation chamber without a clear preference (Figure 1A). To quantitatively analyze the directionality of the cells, we calculated the displacement cosine of each trajectory, where values close to −1 indicate a preference toward the left, and values close to 1 suggest preference toward the right. Since values did not follow a Gaussian distribution, values have been reported as median/interquartile-range, and non-parametric tests were applied to compare the data. In this experiment, the values ranged between −1 and 1 (A. proteus: 0.337/1.348, median/IQ; M. leningradensis: −0.431/1.194, median/IQ; A. borokensis: 0.173/1.422, median/IQ) confirming that in the absence of stimulus all cells moved randomly without any defined guidance. Experimental data: A. proteus, total cells 57, experimental replicates 8, number of cells per replicate 7-8; M. leningradensis, total cells 62, experimental replicates 8, number of cells per replicate 7-8; A. borokensis, total cells 50, experimental replicates 6, number of cells per replicate 7-10.
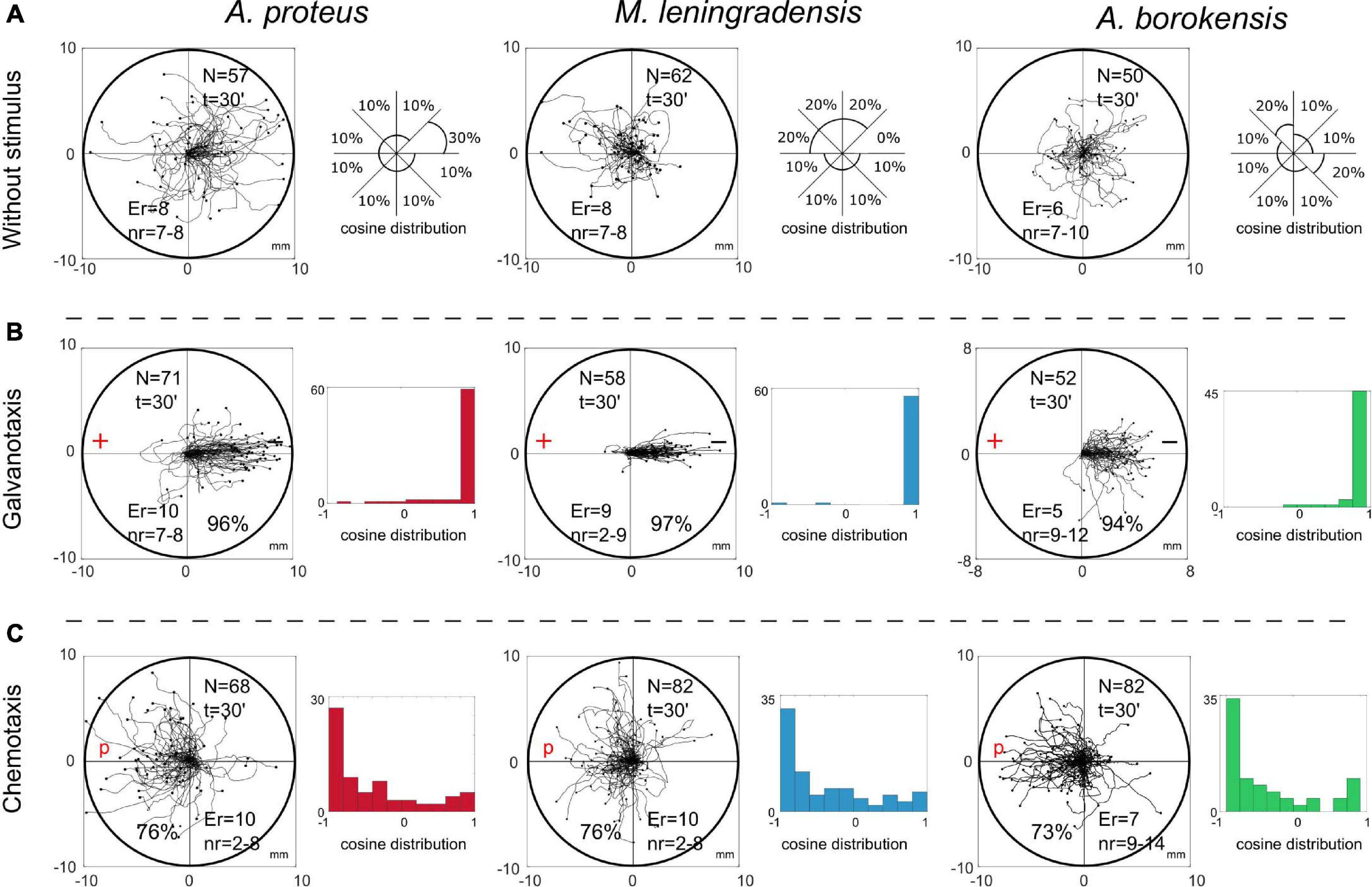
Figure 1. Migration trajectories of the three species under external experimental conditions. Panels (A) show the migration without any stimuli by each of the three species (A. proteus, M. leningradensis, and A. borokensis, respectively). Practically, the amoebae explored all the directions of the experimentation chamber. In addition to the trajectories, the approximated percentage of cells that displaced in certain portion of the exploration chamber is represented. The space has been divided in 8 sections (thus π/4 angle amplitude was used for determining the regions). In panels (B), the locomotion trajectories during galvanotaxis are depicted (96% of A. proteus, 97% of M. leningradensis, and 94% of A. borokensis migrated toward the cathode). Next to the trajectories, the respective histograms of the displacement cosines are illustrated. Finally, panels (C) represent the migration during chemotaxis. 76%, 76% and 73% of the respective cells migrated toward the chemotactic gradient. Alongside the trajectories, the histograms for the angles of each trajectory have been illustrated. “N” is the total number of cells, “Er” is the experimental replications, “nr” is the number of cells per replication, “t” time of galvanotaxis or chemotaxis, “p” chemotactic peptide (nFMLP), “+” anode, “−” cathode. Both the x and y-axis show the distance in mm, and the initial location of each cell has been placed at the center of the diagram.
2. Cell locomotion in an electric field (galvanotaxis).
To study the cellular migration under galvanotaxis conditions, a controlled external direct-current electric field of about 300-600 mV/mm was applied to 181 amoebae belonging to the three species and analyzed in small groups. The results showed that practically all the cells (96% of them) moved toward the cathode, indicating a strong directionality when the galvanotactic stimulus was applied (Figure 1B). Specifically, 96% of A. proteus, 97% of M. leningradensis and 94% of A. borokensis migrated to the cathode. The values of the displacement cosines ranged between −1 and 1 (A. proteus: 0.984/0.068, median/IQ; M. leningradensis: 0.994/0.02, median/IQ; A. borokensis: 0.97/0.097, median/IQ) which confirmed that a fundamental behavior characterized by an unequivocal directionality toward the cathode had emerged under these galvanotactic conditions (for more details, see the Supplementary Table 1). Next, the distributions of the values for the displacement cosines under galvanotaxis were compared to the values obtained in the experiment without stimulus, using Wilcoxon ran-sum test, which indicated that both behaviors were significantly different for the three species, and that this galvanotactic cellular behavior is extremely unlikely to be obtained by chance (p-Values: 10–9, 10–18 and 10–10; Z:−6.046, −8.78 and −6.27 for A. proteus, M. leningradensis, and A. borokensis, respectively). Experimental data: A. proteus, total cells 71, experimental replicates 10, number of cells per replicate 7-8; M. leningradensis, total cells 58, experimental replicates 9, number of cells per replicate 2-9; A. borokensis, total cells 52, experimental replicates 5, number of cells per replicate 9-12.
3. Cell migration under chemotactic gradient (chemotaxis).
Here, we analyzed the trajectories of 232 cells belonging to the three species, which were exposed for 30 min to an nFMLP peptide gradient placed on the left side of the set-up (Figure 1C). In this case, 75% of all cells migrated toward the attractant stimulus, the peptide (see the Supplementary Table 1). Specifically, 76% of A. proteus, 76% of M. leningradensis and 73% of A. borokensis migrated to the peptide. The values of the displacement cosines were −0.657/0.827, median/IQ for A. proteus, −0.656/0.919, median/IQ for M. leningradensis and −0.632/0.9, median/IQ for A. borokensis. Since the medians for the three cosines of displacement were negatives, it was corroborated that most of the cells tended to migrate toward the left side, where the peptide was placed. The comparison between the displacement cosines under chemotaxis and galvanotaxis (p-Values: 10–18, 10–21 and 10–17; Z: 8.74, 9.37, and 8.4 for A. proteus, M. leningradensis, and A. borokensis, respectively) corroborated that the systemic locomotion behavior of cells under the chemotactic gradient was totally different with respect to the observed under an electric field. Experimental data: A. proteus, total cells 68, experimental replicates 10, number of cells per replicate 2-8; M. leningradensis, total cells 82, experimental replicates 10, number of cells per replicate 2-8; A. borokensis, total cells 82, experimental replicates 7, number of cells per replicate 9-14.
4. Migratory behavior under simultaneous galvanotactic and chemotactic stimuli (induction process).
After studying the behavior of the three species without stimulus, under galvanotaxis and chemotaxis conditions, we exposed 458 cells in total to simultaneous galvanotactic and chemotactic stimuli for 30 min (induction process). The nFMLP peptide was placed on the left, in the anode (Figures 2A–C). This experiment showed that 55% of all the amoebae moved toward the peptide-anode, while the remaining 45% migrated toward the cathode (Supplementary Table 1). Specifically, 49% of A. proteus, 55% of M. leningradensis and 60% of A. borokensis migrated to the peptide-anode. The displacement cosines ranged from −1 to 1 (0.045/1.543, median/IQ) for A. proteus, (−0.2/1.706, median/IQ), for M. leningradensis, and (−0.332/1.685, median/IQ) for A. borokensis. This analysis quantitatively verified that two main cellular migratory behaviors had emerged in the experiment, one toward the anode and another toward the cathode (Figures 2D–F). The statistical analysis (Wilcoxon rank-sum test) confirmed the presence of these two different behaviors for A. proteus (p-Value = 10–27; Z = 10.74), M. leningrandensis (p-Value = 10–23; Z = 9.93) and A. borokensis (p-Value = 10–28; Z = 11.008). Experimental data: A. proteus, total cells 155, experimental replicates 21, number of cells per replicate 7-8; M. leningradensis, total cells 134, experimental replicates 18, number of cells per replicate 7-8; A. borokensis, total cells 169, experimental replicates 22, number of cells per replicate 5-11.
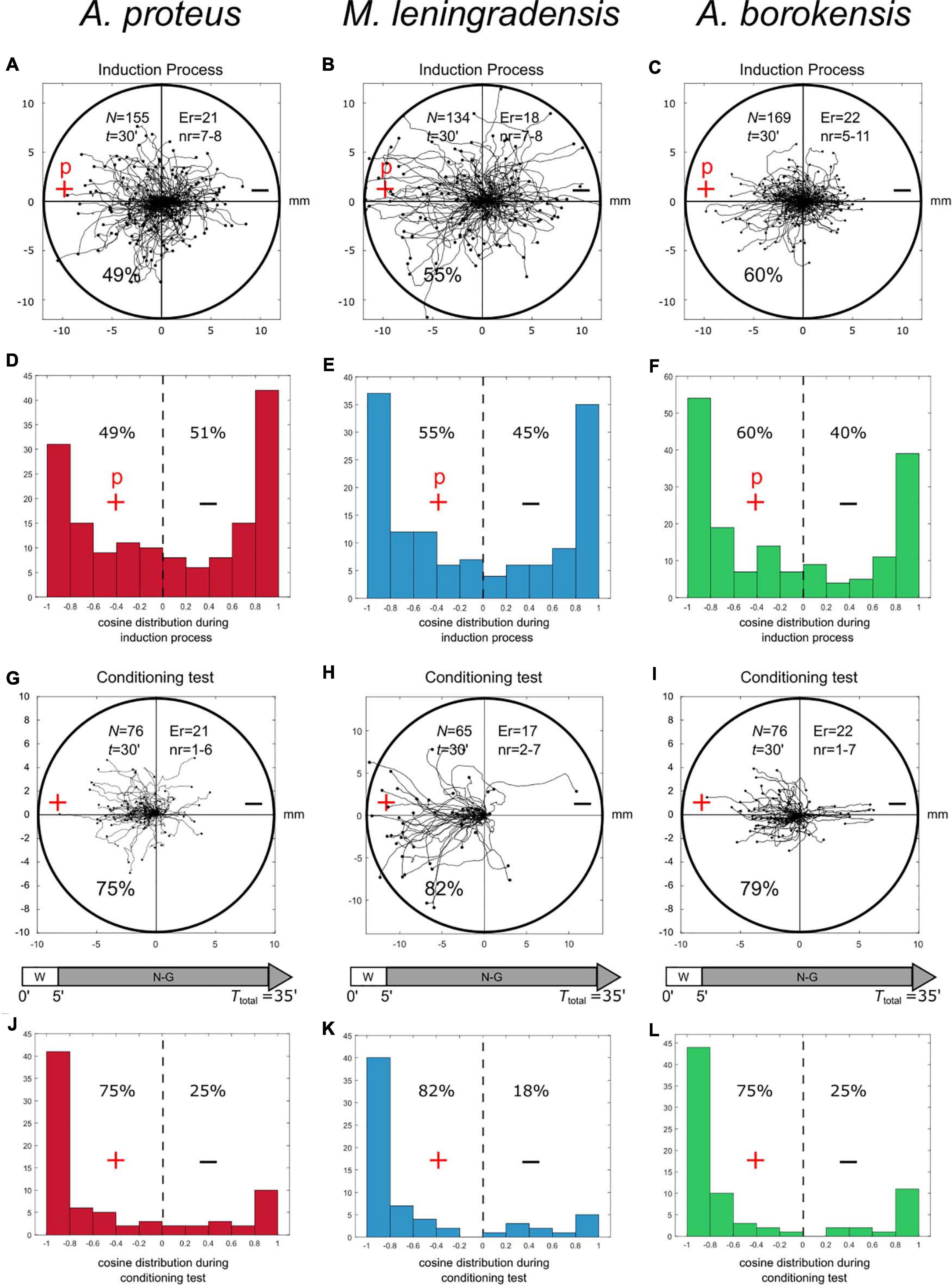
Figure 2. Induction processes and conditioning test. Panels (A–C) show the cellular migration under simultaneous galvanotactic and chemotactic stimuli (induction process) for A. proteus, M. leningradensis, and A. borokensis, respectively. Panels (D–F) illustrate the histograms of the displacement cosines from panels (A–C), respectively. In panels (G–I), the trajectories under galvanotactic conditions of those cells that had previously migrated toward the anode-peptide is represented. It can be observed that the 75%, 82%, and 79% of A. proteus, M. leningradensis and A. borokensis cells migrated toward the left positive pole, where the peptide was absent. Panels (J–L)-illustrate the histograms of the displacement cosines from panels (G–I), respectively. “N” is the total number of cells, “Er” is the experimental replications, “nr” is the number of cells per replication, “t” time of galvanotaxis or chemotaxis, “p” chemotactic peptide (nFMLP), “+” anode, “−” cathode. Both the x and y-axis show the distance in mm, and the initial location of each cell has been placed at the center of the diagram.
5. Emergence of cellular conditioned behavior.
To test if the cells that moved toward the anode during the induction process, presented some kind of associative conditioning in their migratory trajectories, we performed a conditioned behavior test (Figures 2G–I). For such a purpose, those cells that had previously migrated toward the anode-peptide during the exposition to the two simultaneous stimuli, were manually extracted and placed in a Petri dish with a normal culture medium (Chalkley’s medium) for 5 min, without any external stimuli, and then, they were re-exposed, for the second time, to the same single electric field, but without peptide. Note that when amoebae were placed in an electric field for long periods (30 min during the induction process), the probability of dying or, at least, detaching from the substrate and adopting a spherical shape increases sharply; therefore, after the induction process the cells were physically extracted and replated for 5 min to minimize cell damage. Next, we placed the cells in a new identical setup that had never been in contact with the peptide nFMLP, and filled it with clean Chalkley’s medium. In this context, the cells were again exposed to galvanotaxis for another 30 min.
Under these conditions, the analysis of the individual trajectories of 217 amoebae showed that most of the cells (78%) moved to the anode where the peptide was absent (Figures 2G–I, 3). Specifically, 75% of A. proteus, 82% of M. leningradensis and 79% of A. borokensis migrated to the positive pole. The fact that the majority of cells moved toward the anode in the absence of peptide corroborated that a new locomotion pattern had appeared in the cells (note that without the induction process, practically all the cells migrated toward the cathode under galvanotactic conditions).
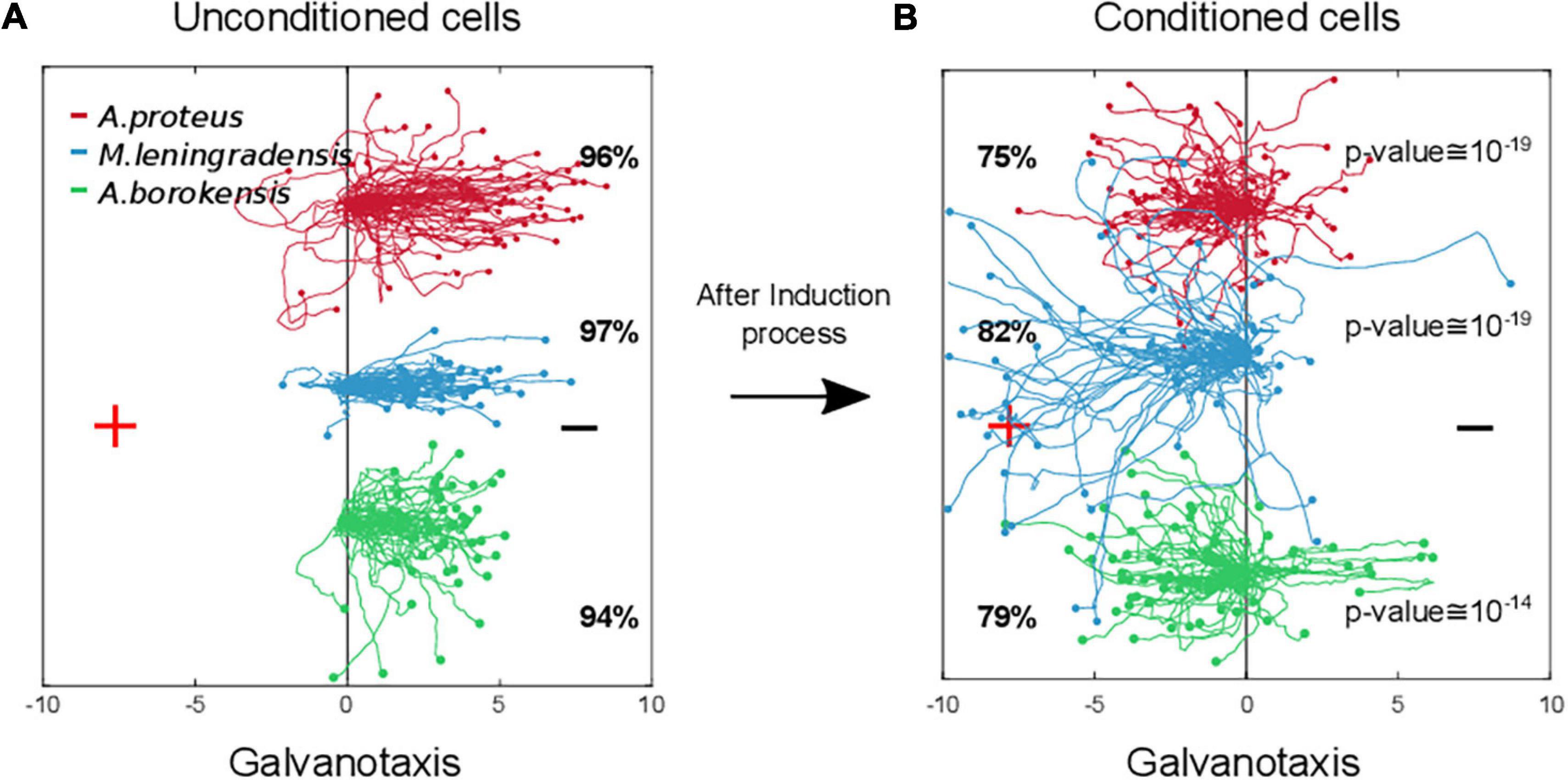
Figure 3. Comparison of amoebae migrations under two galvanotaxis conditions, before and after being conditioned. In red Amoebae proteus, in blue Metamoeba leningradensis, and in green Amoebae borokensis. (A) When galvanotactic stimulus was applied, practically all the cells of the three species migrated toward the cathode. (B) After induction process (simultaneous galvanotactic and chemotactic stimuli for 30 min, see Figure 2) a new locomotion pattern had appeared in the cells, and the majority of amoebae moved toward the anode in the absence of peptide (75, 82, and 79% of the A. proteus, M. leningradensis and A. borokensis, respectively). The p-Values obtained when the displacement cosines of galvanotaxis and conditioning tests for each species were depicted, these values indicated that this newly acquired cellular behavior is extremely unlikely to be obtained by chance. The three species responded similarly, suggesting the presence of associative conditions in most of them (78% on average). “+” indicates anode, while “−” cathode. The x-axis shows the distance in mm.
In this experiment the displacement cosines ranged between −1 and 1, with −0.84/1.02 (median/IQ) for A. proteus, −0.908/0.447 (median/IQ) for M. leningradensis, and −0.905/0.588 (median/IQ) for A. borokensis, thus verifying that the majority of the amoebae displayed a new locomotion pattern characterized by movement to the anode during a galvanotactic stimulus without peptide (Figures 2J–L). See experimental data and quantitative analysis in the Supplementary Information. The results recorded in this experiment were compared to the ones obtained in the galvanotaxis without previous induction, and the test indicated that this newly acquired cellular behavior is extremely unlikely to be obtained by chance (p-Values: 10–19, 10–19 and 10–14; Z: 8.99, 8.91, and 7.52 for A. proteus, M. leningradensis, and A. borokensis, respectively). Additionally, the comparison of all the cells from the conditioning tests against the galvanotactic responses was made, highlighting that this behavior was extremely unlikely to be obtained by chance (p-Value: 10–49, Z = 14.75). Experimental data: A. proteus, total cells 76, experimental replicates 21, number of cells per replicate 1-6; M. leningradensis, total cells 65, experimental replicates 17, number of cells per replicate 2-7; A. borokensis, total cells 76, experimental replicates 22, number of cells per replicate 1-7.
6. Control tests.
Different types of controls were established to complete the analysis of the previously mentioned experimental observations (Figure 4). First, to test the possible capacity of nFMLP peptide to change the migration of cells in an electric field, we have simultaneously exposed 55 Amoeba proteus to galvanotactic and chemotactic stimuli for 30 min, but in this case, placing the nFMLP peptide in the cathode (now in the left part, Figure 4A). Afterward, the same 55 amoebae were subjected to a single electric field, without peptide, for 30 min. Under the simultaneous exposition to the peptide and the electric field (placing the nFMLP peptide in the cathode), 93% of the cells migrated to the negative pole, while the remaining 7% exhibited a stochastic behavior without a clear directionality toward any pole. Under the galvanotactic control, we observed that 98% of the cells ran toward the cathode. The displacement angles were distributed between −1 and 0.98 (−0.998/0.03 median/IQ). This experiment was compared to the Induction Process previously described, and the test indicated that the response of the amoebas was significantly different (p = 10–15; Z = 7.76). Therefore, the previous exposition of cells to the peptide nFMLP in an electric field (placing the nFMLP peptide in the cathode) did not induce any cellular behavior characterized by movement toward the anode under galvanotactic conditions (total cells: 55, experimental replicates: 14, number of cells per replicate: 7-9). During this galvanotactic control of these cells (Figure 4A, right panel), we also observed that 98% of the cells moved toward the cathode. No cell showed a clear migratory movement toward the anode. The cosines of displacements ranged between −1 and 0.17 (−0.998/0.03 median/IQ), supporting mathematically that practically all cells moved toward the cathode. The comparison between the cosines of displacements obtained during the conditioned behavior test and this second galvanotactic control shows that the cellular migration was significantly different between both experiments (p = 10–20; Z = 9.13), indicating that the exposition of cells to the peptide nFMLP in an electric field never developed “per se” a cellular behavior characterized by cells moving toward the anode. Moreover, our results indicated that a cell showing a strong directionality toward the anode was extremely improbable. Therefore, the conditioned behavior can only be obtained from a cellular associative process that links the positive pole with the peptide nFMLP (Figure 2).
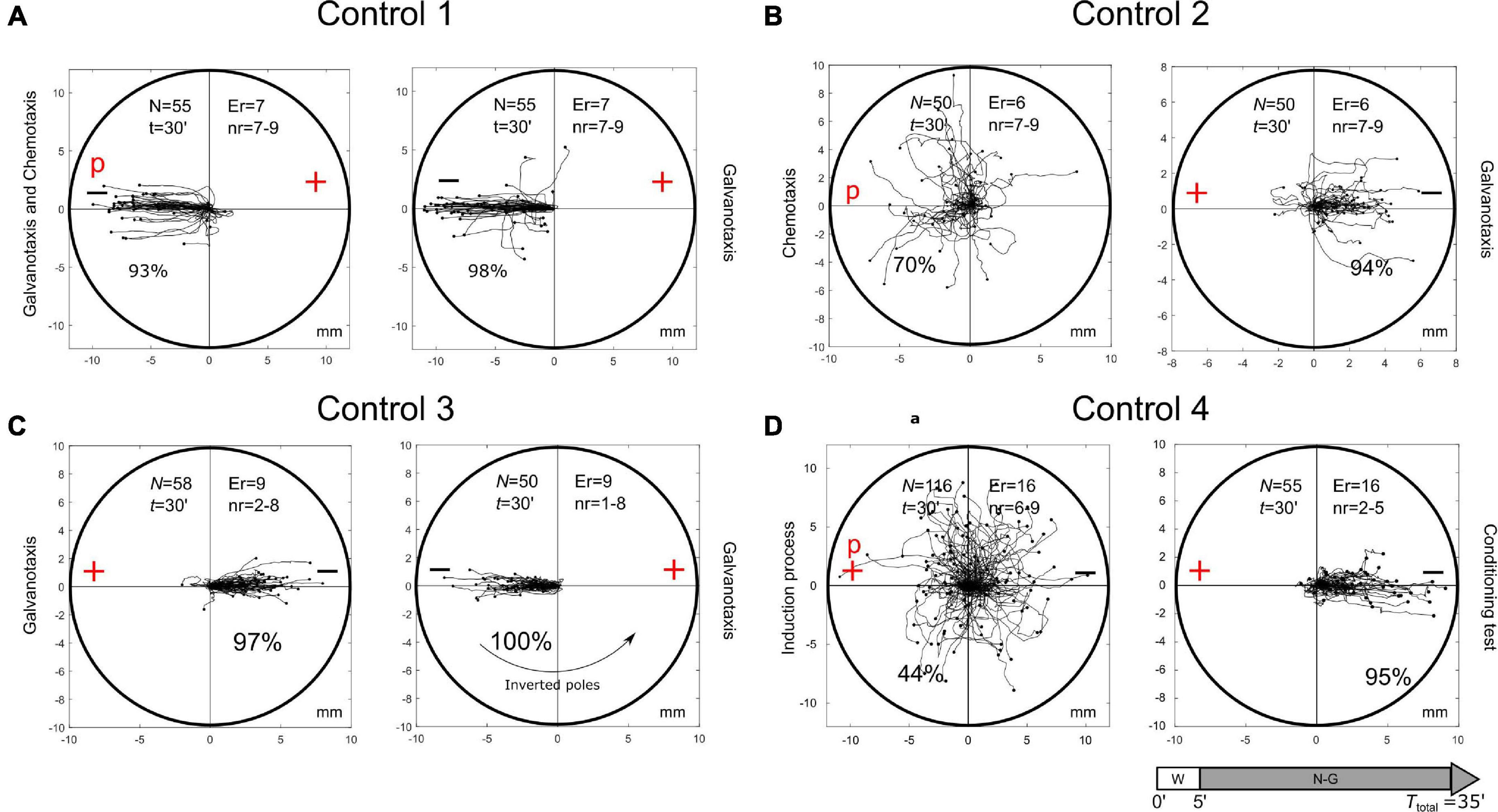
Figure 4. Control tests. (A) Control test on the capacity of nFMLP to change the migration of Amoeba proteus in an electric field. A total of 55 Amoeba proteus were exposed simultaneously to galvanotactic and chemotactic stimuli for 30 min, placing the nFMLP peptide in the cathode. The same 55 amoebae previously exposed simultaneously to galvanotactic and chemotactic stimuli were subjected to a single electric field without peptide for 30 min. In both experiments, no cell showed clear directionality toward the anode. (B) Chemotactic and subsequent galvanotactic control. A total of 50 M. leningradensis were exposed to nFMLP peptide for 30 min, and 72% of them migrated toward the anode. Then, those 50 cells were subjected to a galvanotactic stimulus during 30 min, and 90% of the cells migrated toward the cathode, therefore exhibiting normal galvanotaxis confirming that the exposure to the peptide did not alter the normal galvanotactic response. (C) Galvanotactic control with inverted polarity. A total of 58 M. leningradensis were exposed to a galvanotactic stimulus for 30 min; next, the cells were exposed to another identical electric field with inverted polarity (experimental replicates: 9, number of cells per replicate: 1-8). As it can be observed, practically all the cells showed a normal galvanotactic behavior in both occasions. (D) Induction process and subsequent conditioning test of non-induced cells. A total of 55 M. leningradensis were exposed simultaneously to the electric field and to nFMLP peptide for 30 min, and 44% of them migrated toward the anode. Then, those cells that migrated to cathode (56%) were subjected to a galvanotactic stimulus during 30 min, and 95% of such cells migrated toward the cathode, exhibiting normal galvanotaxis, and confirming that the previous exposure to the peptide did not alter the normal galvanotactic response. “N” is the total number of cells, “Er” is the experimental replicates, “nr” is the number of cells per replicate, “t” galvanotaxis time, “+” anode, “−” cathode, “P” nFMLP peptide. Both the x and y-axis show the distance in mm, and the initial location of each cell has been placed at the center of the diagram.
To test the cells that move toward the cathode during chemotaxis, 50 M. leningradensis were exposed to nFMLP peptide for 30 min and 72% of them migrated toward the peptide. Then, these 50 cells were subjected to a galvanotactic stimulus for 30 min (Figure 4B). The results showed that 90% of the cells migrated toward the cathode, therefore exhibiting normal galvanotaxis. This experiment confirmed that the exposure to the peptide did not alter the normal galvanotactic response of cells (total cells: 50, experimental replicates: 12, number of cells per replicate: 7-9).
Next, a galvanotactic control of cells that moved toward the cathode was also performed. For such a purpose, 58 M. leningradensis were exposed to a galvanotactic stimulus for 30 min, next they were placed in a Petri dish filled with Chalkley’s medium for 5 min, and finally exposed to another identical electric field with inverted polarity (Figure 4C). The results of this experiment showed that all the amoebae exhibited normal galvanotactic behavior on both occasions. By inverting the position of the electric field, we demonstrated that the amoebae did not migrate to a specific point in the space (total cells: 58, experimental replicates: 18, number of cells per replicate: 1-8).
Finally, we tested the cells that moved toward the cathode during the induction process (simultaneous galvanotactic and chemotactic stimuli for 30 min) Figure 4D. For such a purpose, 55 M. leningradensis that migrated toward the cathode during the induction process were again subjected to a controlled electric field (galvanotaxis for 30 min). The results showed that practically all the amoebae (95%) migrated toward the cathode, confirming that these cells were unconditioned, and that their habitual behavior, that is, to go to the cathode, was not altered by the induction process (total cells: 116, experimental replicates: 32, number of cells per replicate: 6-9).
The total number of independent experimental replicates in these control tests was 76 and the total number of individual cells studied in such control tests were 279.
7. Intensity of response in the conditioned cells.
Here, we have analyzed the displacement module (measured in mm) of the conditioned cells which showed the intensity of the response in those cells that modified their migratory behavior by the association of stimuli. Such a module corresponds to the vector, starting in the origin of the migratory trajectory and finishing at the end of it.
The results showed that the displacement module for A. proteus, values ranged from 0.103 to 8.017 (2.445/2.457, median/IQ), for M. leningrandensis was comprehended between 0.368 and 14.96 (7.5/8.26, median/IQ), and for A. borokensis, from 0.3701 to 9.229 (3.232/2.706, median/IQ). See quantitative analysis in the Supplementary Information. In Figures 5A,B, the distribution of these statistics by boxplots and frequency polygons, respectively, is depicted. Clearly, M. leningradensis responded much more intensively than the other two species, which exhibited a similar intensity in the conditioned responses. However, the values of metamoebae cells were more heterogeneous than those obtained from the other species, indicating more variability in its migratory conditioned behavior; A. proteus presented the most stable intensity of the conditioned responses, with the lesser variability. Additionally, the response intensity pattern followed by M. leningradensis was very different from the observed in the other two species (Figure 5B). The total number of conditioned cells analyzed was 167, and the experimental time 30 min. Kruskal-Wallis test indicated that there were significant differences regarding the intensity of response (p-Value = 10–6, χ2 statistic = 23.83), and therefore, pair-wise comparisons were made by using Wilcoxon rank-sum test. The obtained results indicated that significant differences were found between the values of A. proteus and M. leningradensis (p-Value = 10–6, Z = −4.468), as well as for the values of A. borokensis and M. leningradensis (p-Value = 0.00013, Z = 3.81) but not between A. borokensis and A. proteus (p-Value = 0.2084, Z = −1.258).
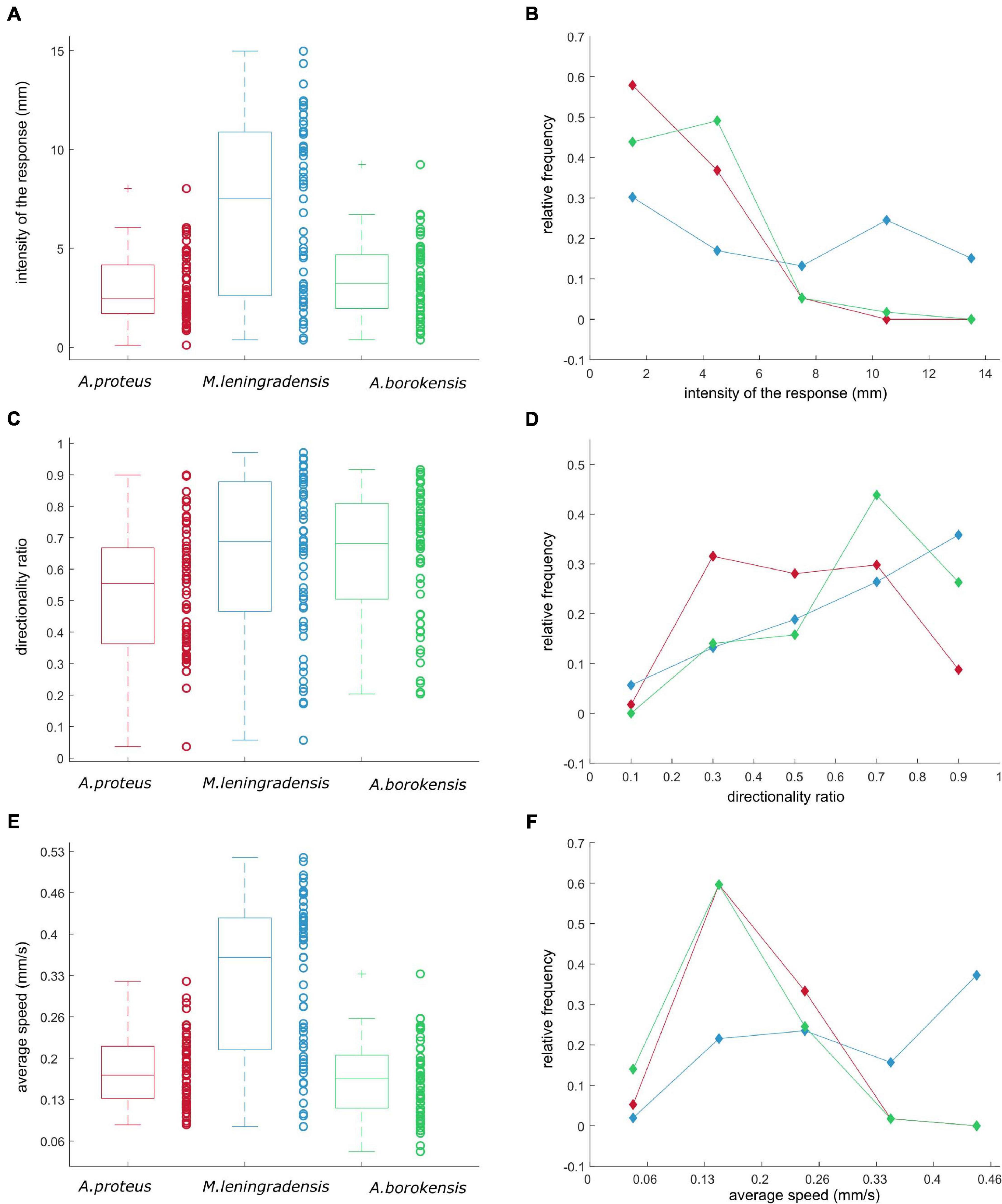
Figure 5. The intensity of response, directionality ratio, and average speed of the cells that acquired conditioned behavior. (A,C,E) panels illustrate boxplots for the intensity of response (measured in mm), the directionality ratio, and the average speed (measured in mm/s), respectively (where A. proteus is represented in red, M. leningradensis in blue, and A. borokensis in green). On the side of each box, the values of the statistic for each cell were depicted by circles. Panels (B,D,F) illustrate the frequency polygons corresponding to the statistics in panels (A,C,E), respectively. Here, the x-axis indicates the value of the statistic, while the y-axis represents the relative frequency (i.e., the number of cases divided by the total number of cells of that species) for the three cell types.
8. Straightness of the conditioned cell trajectories.
To analyze the straightness in the direction followed by the conditioned cell we have calculated the directionality ratio (Figures 5C,D). This statistic test quantifies the trajectory straightness, which ranged between 0 (for fully curved trajectories) and 1 (for fully straight tracks), and is calculated by the quotient between the displacement module and the total distance traveled. First, we computed the total trajectory length, next, we obtained the Euclidean distance between the start point and the endpoint, and finally, we defined the directionality ratio as the quotient between these two values. The results showed that for A. proteus, values ranged between 0.0362 and 0.899 (0.555/0.304 median/IQ), for M. leningradensis from 0.056 to 0.970(0.688/0.412, median/IQ), and for A. borokensis, from 0.2032 to 0.916 (0.681/0.304 median/IQ). This data suggested that A. borokensis and M. leningradensis followed similar and strong straightness in their conditioned migratory direction, going to the migratory target much more directly than A. proteus cells, which exhibited a more wandering trajectory in their movements. Interestingly, again, M. leningradensis showed more heterogeneity in their directionality values than the obtained from the other species, indicating more variability in their conditioned responses. Moreover, A. borokensis presented the largest number of cells with high straightness in their migratory movements (Figure 5D). The total number of conditioned cells analyzed was 170, and the experimental time 30 min. Kruskal-Wallis test indicated that at least the directionality ratio of one of the species was significantly different (p-Value = 0.002, χ2 statistic = 12.66), and therefore, pair-wise comparisons were made. The obtained results showed that no significant differences were found between the directionality ratio values of A. borokensis and M. leningradensis (p-Value = 0.697, Z = 0.3888), however the values of A. proteus were significantly different to the values of M. leningradensis (p-Value = 0.0055, Z = −2.775), and A. borokensis (p-Value = 0.0008, Z = −3.34).
9. Average speed of the conditioned cells.
Another kinematic property analyzed was the average speed (measured in mm/s) of the conditioned cells. The results showed that the values for A. proteus ranged from 0.093 to 0.324 (0.173/0.084, median/IQ), for M. leningrandensis between 0.09 and 0.523 (0.362/0.212, median/IQ), and for A. borokensis, from 0.05 to 0.336 (0.167/0.086, median/IQ), see Figures 5E,F. Conditioned M. leningradensis cells moved significantly faster than the other two species. However, these metamoebae cells migrated more dispersedly that A. proteus and A. borokensis cells, which in turn, showed a similar behavior for the average speed (Figure 5E). In the case of A. proteus and A. borokensis cells, the majority of speed values were concentrated around 0.16 mm/s (Figure 5F). In addition, the distance traveled (measured in mm) was previously estimated to obtain the average speed. These results showed that the values for A. proteus ranged from 2.777 to 9.714 (5.18/2.523, median/IQ), for M. leningradensis varied between 2.692 and 15.7 (10.871/6.373, median/IQ), and for A. borokensis, from 1.485 to 10.073 (5.0022/2.5647, median/IQ). The total number of conditioned cells analyzed was 170, and the experimental time 30 min. The quantitative control test comparing the average speed of the three species indicated that the distributions were significantly different (p-Value = 10–11, χ2 statistic = 48.43). The obtained results indicated that no significant differences were found between the speed values of A. proteus and A. borokensis (p-Value = 0.2918, Z = 1.054), but the average speed was significantly different between A. proteus and M. leningradensis (p-Value = 10–9, Z = −5.814), and between A. borokensis and M. leningradensis (p-Value = 10–10, Z = 6.197).
10. Directionality persistence level in the conditioned cells.
Next, we studied the persistence of the movement by analyzing Spearman’s correlation coefficient relating the x coordinate of the displacement of the cells, and the respective time step (Figure 6). We used this coefficient instead of Pearson’s correlation coefficient because the data was not normally distributed. Values close to −1 indicated that cells migrated persistently toward the left pole (anode), while values close to 1 corresponded to a strong tendency toward the right pole (cathode), and intermediate values suggested little persistence of migration to any pole. Results of our analysis indicated that values of A. proteus ranged between −1 and 0.171 (−0.934/0.184 median/IQ), of M. leningrandensis from −1 to 0.017 (−0.976/0.137, median/IQ), and of A. borokensis, from −1 to 0.663 (−0.99/0.058 median/IQ). The results showed that the persistence values of A. borokensis were higher than those of A. proteus, but no differences were found when these values were compared to those obtained from M. leningradensis. All the amoebae analyzed exhibited a very strong preference toward the anode. To illustrate this behavior, in Figures 6A–C we have represented three prototypical series and their correlation coefficient calculation. Also, in Figures 6D–F we have represented through histograms all the correlation values obtained for the trajectories of the three species. The total number of conditioned cells analyzed was 170, and the experimental time 30 min. The quantitative control test showed the presence of significant differences between the statistic values (p-Value = 0.0008, χ2 statistic = 14.18). The results indicated that no significant differences were found between the correlation values of A. borokensis and M. leningradensis (p-Value = 0.459, Z = 0.739), however, the values of A. proteus were statistically different compared to those of A. borokensis (p-Value = 0.00016, Z = 3.769) and M. leningradensis (p-Value = 0.0127, Z = 2.492).
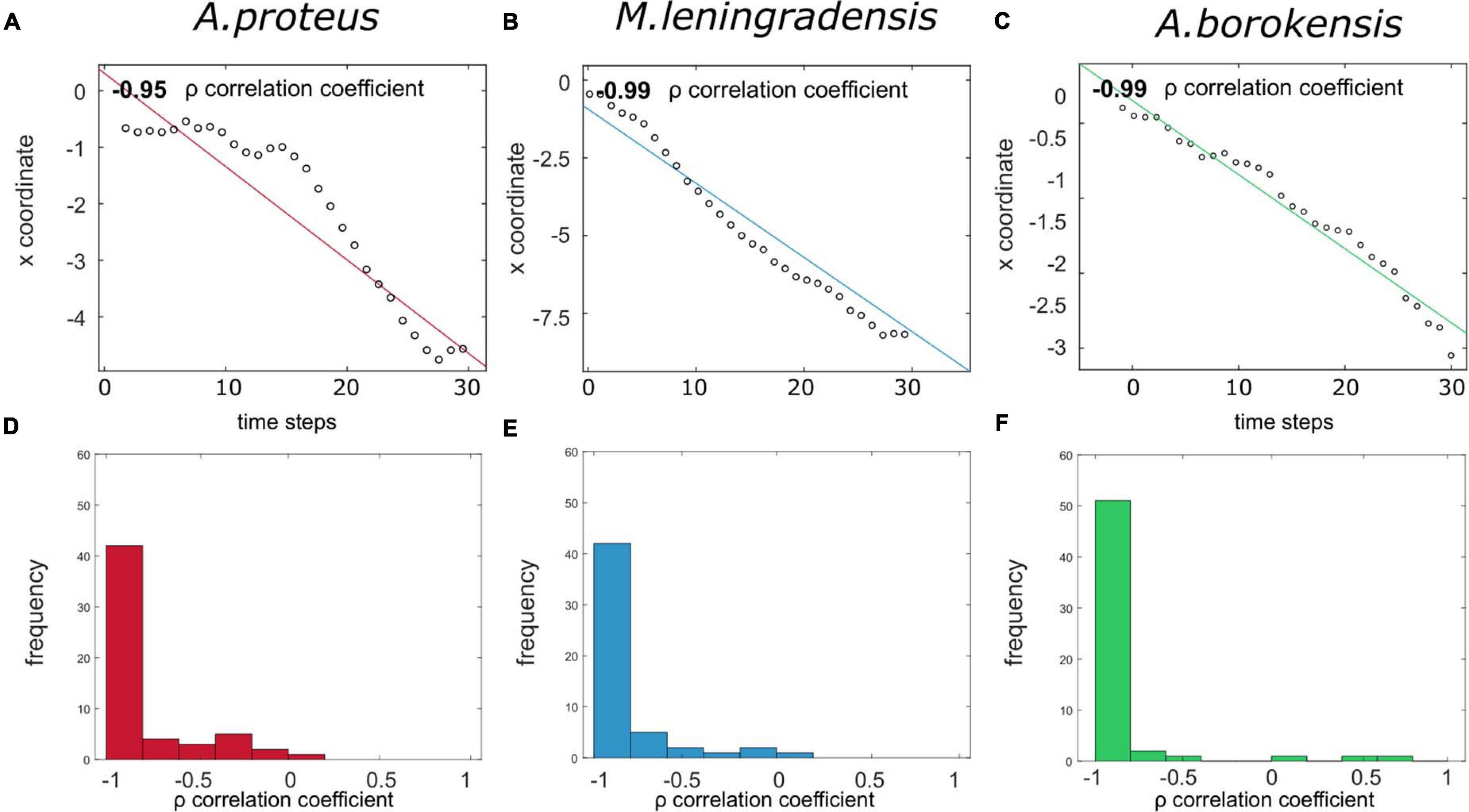
Figure 6. Persistence toward the anode for the cells that acquired conditioned behavior. (A–C) panels illustrate Spearman’s correlation coefficient ρ relating the x coordinate and the time steps for three prototypical cells, each one of them corresponding to one of the species (A. proteus, M. leningradensis, and A. borokensis). As it can be observed, there is a strong negative correlation, denoting that as time increases, cells tend to maintain their migratory direction toward the anode, which indicates high persistence during the displacement. (D–F) panels represent histograms for Spearman’s correlation coefficients for the three species, respectively. It can be noticed that the majority of the cells present a strong negative correlation, corresponding to strong persistence toward the anode.
11. Persistence time in the conditioned cells.
The motility patterns acquired by amoebae are also characterized by a limited period of conditioned behavior. Figures 7C-E display how the trajectories of 15 amoebae, that previously had acquired such conduct after the induction process, gradually lost the persistence toward the anode and turned back to the cathode, under galvanotaxis, thus losing their acquired behavior.
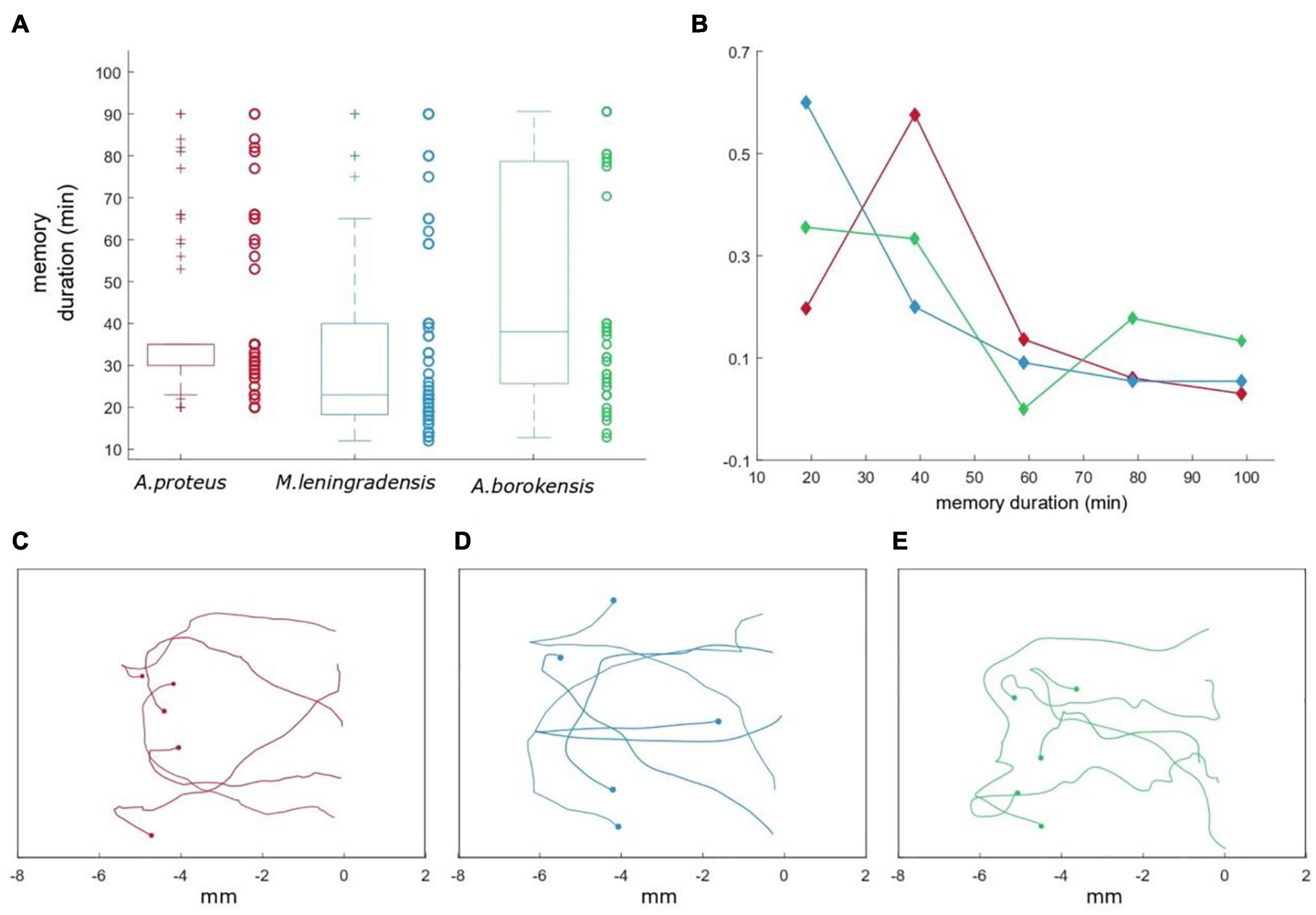
Figure 7. Persistence time for the cells that acquired conditioned behavior. (A) Boxplot representing the duration of the acquired associative behavior (up to 90 min) for the three species (A. proteus in red, M. leningradensis in blue, and A. borokensis in green). On the side of each box, we depict the persistence time values with circles. (B) Frequency polygons representing the duration for each species. The x-axis indicates the duration in minutes, while the y-axis represents the relative frequency for the three cell types. As it can be observed, the majority of the cells present persistence times ranging between 20 and 50 min. Panels (C–E) show how the trajectories of 15 prototypical amoebae (belonging to A. proteus, M. leningradensis, and A. borokensis) after the induction process lost gradually the persistence toward the anode and turned back to the cathode, under new galvanotaxis conditions. Here, the x-axis indicates the distance traveled in mm.
We estimated the temporal persistence of the conditioned behavior calculating the time (measured in minutes) during which these cells showed a directional response to anode during galvanotaxis after an induction process. For such a purpose, 166 amoebae (66 A. proteus, 55 M. lenigradensis, 45 A. borokensis) that had previously migrated toward the anode-peptide during the exposition to the two simultaneous stimuli (induction process) were manually extracted and placed for 5 min on a normal culture medium (Chalkley’s medium) in a small Petri dish in absence of stimuli. Next, the cells were deposited on a new identical set-up that had never been in contact with the chemotactic peptide nFMLP and exposed for the second time to a single electric field, without peptide, for 30 min. This process was repeated two more consecutive times. The time elapsed until the cells forgot the conditioned response, turned around, and returned to the cathode was estimated (Figure 7) and the results indicated that the temporal persistence for A. proteus, ranged between 20 and 90 (35/5, median/IQ), for M. leningradensis between 12 and 90 (23/21.75, median/IQ), and from 13 to 90 (38/52.5, median/IQ) for A. borokensis. Figure 7B shows that the patterns of loss of persistence toward the anode (loss of conditioning) were very similar in the three species. The whole analysis indicated that the total average time of the cells belonging to the three species that lost the acquired motility pattern was 37/36 (median/IQ) minutes. So, cells belonging to three species of unicellular organisms were able to acquire a new systemic behavior from the association of stimuli; they were also able to keep it for long periods compared to their cellular cycle, and they forgot them later. The quantitative analysis test indicated that at least the persistence times of one of the species was significantly different (p-Value = 0.0017, χ2 statistic = 12.7). Temporal values of M. leningradensis were found to be significantly different when compared to the values of A. proteus (p-Value = 0.0017, Z = 3.134) and A. borokensis (p-Value = 0.0032, Z = −2.945), but no differences were found between the values of A. proteus and A. borokensis (p-Value = 0.466, Z = −0.728).
Finally, a violin graph (Figure 8) depicts the most relevant results of our quantitative analysis. From this exhaustive analysis, we concluded that the cellular conditioning was robust, and these three species presented a similar motion pattern structure, characterized by persistence times larger than 20 min, directionality ratios close to 0.6, and persistence directionalities close to −1, indicating the presence of conditioning for long periods, remarkable straightness in their trajectories and strong directionality persistence toward the anode pole, respectively.
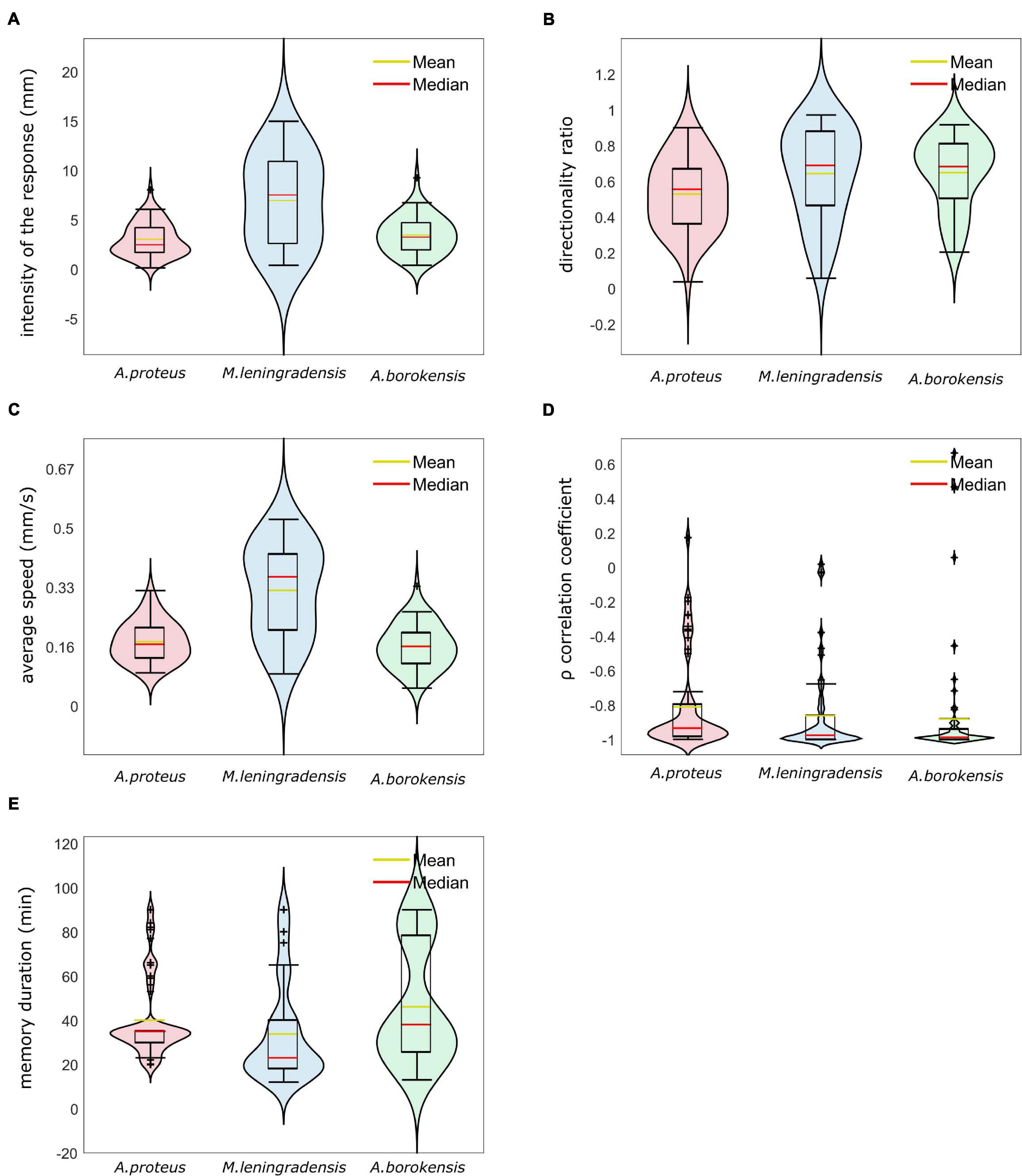
Figure 8. Violin plots for cells of the three species that acquired conditioned behavior. In this graph, we represent by Violin plots the probability distribution, the boxplots, and the mean value of the five main quantitative characteristics. Panel (A) depicts the intensity of response (in mm), panel (B) illustrates the directionality ratio, panel (C) represents average speed (in mm/s), panel (D) depicts Spearman’s correlation coefficient between time and the x-axis of the trajectory, and (E) illustrates the persistence time of the acquired new behavior (mins). A. proteus cells presented conditioned responses characterized by strong persistence, less speed and less distance traveled than the other species. M. leningradensis were the less stable cells, i.e., demonstrated less homogeneity and more variability than the other species; additionally, this species showed more intensive and fast migration than the other two. A. borokensis cells exhibited more directionality persistence than the other two species, with high straightness toward the anode. The three species analyzed show extremal values in their correlation coefficient (close to -1), and it can be concluded that the cellular conditioning for the three species is a very robust behavior with strong directional persistence.
Over centuries, the combined work of naturalists, philosophers, physiologists and scientists has shaped a very plentiful and venerable history of the essential principles that define the basic forms of associative memory and learning processes (Finger, 2001). Among all of them, it is worth noting the Nobel Prize Laureate Ivan Pavlov who was the first to establish the fundaments of the associative memory in his classic studies with dogs (Pavlov, 1927). Following a similar conceptual framework of Pavlov’s experiments, and using an appropriate direct-current electric field (galvanotaxis) and a specific peptide (nFMLP, typically secreted by bacteria) as a chemoattractant (chemotaxis), we have addressed here essential aspects of the cellular systemic behavior. The migration trajectories of more than 2000 individual cells belonging to three different species of unicellular organisms, A. proteus, M. leningradensis, and A. borokensis, have been exhaustively studied in this work, confirming that these three species were able to modify their systemic response to a specific stimulus by associative conditioning.
First, we have observed that cell locomotion in absence of stimuli exhibited a random directional distribution by which amoebae explored practically all the directions of the experimentation chamber. Second, cells showed an unequivocal systemic response consisting of the migration to the cathode when exposed to a direct electric field (galvanotaxis). Third, amoebae were studied during chemical guidance by exposing them to an nFMLP peptide gradient. In this experiment, 75% of cells on average migrated toward the chemotactic stimulus showing stochastic locomotion movements with robust directionality toward the peptide (chemotaxis). All these unicellular organisms belonging to the three species here studied responded in concordance with other similar experimental observations, carried out in absence of stimuli (De la Fuente et al., 2019b), in galvanotactic conditions (Prusch and Britton, 1987; Bray, 2000), and under a chemotactic stimulus (Dunigan et al., 2019).
An essential question of contemporary biology is to know if cells are capable of acquiring new behavioral systemic patterns to adapt themselves to changes in the external medium. For such a purpose, those cells that had previously migrated toward the anode-peptide during the dual simultaneous exposition to galvanotactic and chemotactic stimuli (induction process) were exposed to a single electric field, without peptide. Under these conditions, most of the cells (78% on average) ran to the anode where the peptide was absent. The fact that the majority of induced cells moved toward the anode in the absence of peptide corroborated that a new locomotion pattern had appeared in the cells. Note that without the induction process practically all the cells migrated toward the cathode under galvanotactic conditions and, conversely, after induction process most of the cells went to the anode.
When the exposition to a stimulus related to the amoebae nourishment (the peptide nFMLP) is accompanied by an electric field, and the peptide is placed in the anode (induction process), some amoebae seemed to associate the anode with the food (the peptide). After the induction process, testing under galvanotactic conditions, most of the conditioned amoebae ran toward the anode, where the peptide was absent, in contrast to their known tendency to move to the cathode, which demonstrates their modified systemic conduct.
To better understand this cellular conditioning, we analyzed a great number of individual conditioned amoebae by studying the intensity of the migratory movements, the directionality persistence, the total distance traveled, the average speed, the directionality ratio, and the persistence times.
This analysis has shown that A. proteus cells presented the most stable conditioned responses, with less variability, as well as exhibiting strong persistence, less distance traveled, and less speed of locomotion movements. The temporal persistence was high in these amoebae. However, they exhibited greater wandering in their migratory trajectories.
The conditioned M. leningradensis showed the maximum intensity of response during its locomotion movements but presented much less stable behavior than A. proteus, i.e., they exhibited greater variability and heterogeneity in all the tests performed. Also, these amoebae cells were those that traveled the most distance, exhibiting maximum speed in their movements, and sharp persistence with strong straightness. Their persistence times were the shortest of the three species.
Conditioned A. borokensis were the cells that showed greater persistence with high preference toward the anode as well as robust straightness. These amoebae exhibited similar distances traveled compared to the A. proteus with very close migration speeds and persistence times, but with greater dispersion and variability. Besides, their persistence times were the longest of the three species.
Cells belonging to the three species analyzed showed extremal values in their Spearman’s correlation coefficient (close to −1), i.e., there is very high directionality persistence during the migratory displacement of all amoebae. In fact, this is a key characteristic of conditioned cells. The strong negative correlation denoted that as time increases, cells sharply tended to maintain their migratory direction toward the anode during a large period (despite their usual behavior is to move toward the cathode). Additionally, this magnitude is scale-invariant for the two variables considered, in this case, the x coordinate indicating the position and the time step t. This analysis showed that cellular conditioning was robust for the three species with strong directional persistence, and such a result represents a fundamental aspect of the behavior of conditioned cells.
The kinematic variables studied (intensity of the migratory movements, the total distance traveled, the average speed, and the directionality ratio) highlighted the different responses that cellular life exhibited at individual level, even within the same species. However, we can conclude that the cellular conditioning for the three species was robust because these three species presented a similar motion migratory structure, characterized by persistence times larger than 20 min, directionality ratios close to 0.6, and persistence directionalities close to −1, indicating the presence of conditioning for long periods, remarkable straightness in their trajectories and strong directionality persistence toward the anode. Most conditioned cells moved toward the anode in the absence of peptide thus corroborating that a new locomotion pattern had appeared in the cells (specifically, 75% of A. proteus, 82% of M. leningradensis, and 79% of A. borokensis migrated to the anode). These cells were able to keep this new conditioned response for long periods of time with respect to their cellular cycle (39.58 ± 22.48 min), forgetting it later. The results were compared to those obtained in the galvanotaxis without previous induction, and all tests indicated that this newly acquired cellular behavior is extremely unlikely to be obtained by chance (p-Values: 10–19, 10–19 and 10–14 for A. proteus, M. leningradensis, and A. borokensis, respectively). As a matter of fact, the comparison of all the cells from the conditioning tests against the galvanotactic responses emphasized that it was extremely unlikely to obtain such conditioned behavior by chance (A. proteus, M. leningradensis, A. borokensis p-Value: 10–49). There is no scientific evidence (in our works and the literature) that neither the nFMLP peptide nor the electric field “per se” may originate such cellular behavior.
On the other hand, all controls indicated that cells that were exposed independently to galvanotaxis or chemotaxis or under simultaneous galvanotactic and chemotactic stimuli (but placing the nFMLP peptide in the cathode) did not present any observable conditioned pattern. Pavlov described four fundamental types of persistent behavior provoked by two stimuli. The experiments on cellular conditioning exposed here were based in one of them, the so-called “simultaneous conditioning,” in which both stimuli are applied at the same time. Nevertheless, strictly speaking we cannot classify our findings as the classical Pavlovian conditioning since not all controls for classical conditioning studies were performed yet (Rescorla, 1967).
In parallel to our Pavlovian-like experiments, for better understanding the dynamic characteristics of the locomotion movements and to quantitatively study the role of the nucleus in the migration of Amoeba proteus, we have previously analyzed the movement trajectories of enucleated and non-enucleated amoebae using advanced non-linear physical-mathematical tools and computational methods (De la Fuente et al., 2019b). Specifically, in this study, we calculated the relative move-step fluctuation along their migratory trajectories by applying the root mean square fluctuation, a classical method in Statistical Mechanics based on Gibbs and Einstein studies (Gibbs, 1902; Einstein, 1909) that has been developed and widely applied to quantify different time-series. The results showed that both cells and cytoplasts displayed migration trajectories characterized by non-trivial long-range positive correlations with periods of about 41.5 min on average in all the analyzed cells and cytoplasts, which corresponded to non-trivial dependencies of the past movements. Therefore, each cellular move-step at a given point is strongly influenced by its previous trajectory. This dynamic memory (non-trivial correlations) represents a key characteristic of the movements of Amoeba proteus during cell migration (De la Fuente et al., 2019b). It is worth noting that this temporal persistence (non-trivial correlations with 41.5 min on average) in both enucleated and nucleated cells matches with the Pavlovian-like dynamic memory (40 min on average) obtained here.
Our results unequivocally evidence that cellular conditioning is a robust and stable cellular property, and that the acquired systemic responses are very similar in the three species analyzed. The fact that unicellular organisms belonging to three different species such as A. proteus, M. leningradensis, and A. borokensis are able to associate external signals and consequently generate new robust migratory responses, which can be remembered for long periods, show that cellular conditioned behavior may be an essential property common to more cells in Nature.
Cellular behavior consistent with memory patterns and learning has been observed in different kingdoms including bacteria, protozoa, fungus-like organisms and metazoan. For instance, gene expression patterns in E. coli are almost identical when responding to either a drop in oxygen concentration levels or to a rise in the ambient temperature. This behavior is thought to be a consequence of the association of higher temperatures with an anaerobic environment, such as it is the prevailing context in mammalian intestinal tracts (Tagkopoulos et al., 2008). Besides, the ability to encode robust and persistent memory patterns has been demonstrated in collectives of bacteria (Yang et al., 2020). In other experiments it has been observed that Pseudomonas aeruginosa cells also use multigenerational memory via rhythmic pattern of cAMP (Lee et al., 2018). Armus et al. (2006) have identified in Paramecium caudatum that this protist is able to develop a preference for illumination level using a mild electric shock as a reinforce. Fungus-like organisms such as Physarum polycephalum has been shown to exhibit a primitive type of learning (Boisseau et al., 2016; Boussard et al., 2019; Kramar and Alim, 2021). Moreover, this unicellular multinucleate plasmodium was observed to acquire new thermotactic behaviors when exposed to a temperature gradient and a source of nutrients. While under normal circumstances, Physarum polycephalum tend to migrate toward higher temperatures up to 30°C, after conditioning, it shows a preference for cooler temperatures (Shirakawa et al., 2011). Dictyostelium cells exhibit cellular memory during chemotaxis in traveling waves (Skoge et al., 2014). Human pancreatic β cells have also demonstrated to exhibit associative conditioning behaviors. In short, temporal potentiation of sensitivity to glucose was achieved by replacing glucose concentrations with other secretagogues in combination with carbachol, giving rise to a newly formed behavior that could be explained by a short-term associative conditioning process (Sanchez-Andres et al., 2020). Electrophysiological studies with individual Purkinje cells have showed that these organisms can remember the interval between the onset of an artificial conditioned stimulus and the onset of an artificial unconditioned stimulus (Gallistel, 2017). Recently, the almost forgotten experiments on Pavlovian conditioning in the ciliate Stentor coeruleus and Paramecium aurelia has been reconsidered (Tang and Marshall, 2018; Gershman et al., 2021).
Our exhaustive experimental and quantitative analysis with three different species analyzed, compared with these studies on complex cellular responses in bacteria, protozoa, fungus-like organisms and metazoans, allow us to conclude that cellular associative conditioning might be a widespread characteristic of unicellular organisms.
It is still too early to hypothesize about the molecular mechanisms supporting the cellular associative conditioning. In this sense, we want to point out that the Pavlovian-like experiments presented here were originally conceived in 2013 when we studied self-organized enzymatic activities arranged in dynamic metabolic networks (De la Fuente, 2014). From these studies, we could verify using advanced tools of Statistic Mechanics and techniques of Artificial Intelligence the emergence of Hopfield-like dynamics in self-organized metabolic networks which were characterized by exhibiting associative memory, similar to neural networks (De la Fuente et al., 2013). In that study, the associative memory emerges as a consequence of the complex metabolic dynamics that take place within the cell at systemic level. Consequently, our work quantitatively showed, for the first time, that an associative memory was also possible in some unicellular organisms, and such type of memory would correspond to an epigenetic type of cellular memory (De la Fuente, 2015).
Cell migration, including the conditioned behavior by which cells are able to acquire new migratory patterns, is a systemic emergent property characterized by integrated physiological processes, highly coordinated and carefully regulated. This property is originated through complex non-linear interaction of the molecular-metabolic reactions of the cell, which allow self-organization of the biochemical system as a whole. The emergent systemic properties are therefore not found in any of their molecular-metabolic parts, or individual physiological process, and represent an increase in the information and complexity of the cell (De la Fuente et al., 2021).
Cellular conditioning could be an essential issue to understand some key evolutionary principles involved in increasing the cellular adaptive fitness to microenvironments. Throughout evolution, the organisms that adapt successfully their behavior to the environment do increase their possibilities to survive and reproduce compared to those that cannot. Associative learning in animals has long been considered essential for the efficient adaptation and processing information about the environment. In fact, learning processes have been shaped through evolution to enable animals to detect and store information about causal interactions with their surroundings. In free-living cells, unlike habituation and sensitization, associative conditioning results in responses that are reliably correlated with the state of the external medium, allowing unicellular organisms to recognize significant events and respond quickly and appropriately to specific signals. Associative conditioning could enable cellular organisms to detect contingency relations between different stimuli, or to establish different types of connections between a specific behavior and its consequences for survival. The association of these cues will benefit tracking current conditions and map them for optimal or near-optimal responses. These responses will confer adaptive advantages to organisms enabling them to anticipate to future events and discriminate between different types of stimuli. Cellular conditioning could lead also to very rapid, rather than gradual, behavioral changes, that enable fast detection of correlated features in complex environments. All these elements allow to consider cellular associative conditioning as a powerful optimizing mechanism to perform efficiently different behavioral sequences under variable ecological circumstances. Moreover, learning has long been regarded as one of the hallmarks of cognition, and this form of cellular elementary learning also could have essential implications in the origin of primitive forms of cognition and the role of convergent evolution in biological cognition. Cellular associative conditioning would be a good example of how evolution can find elegant solutions to an adequate adaptation to the microenvironments.
In short, we have analyzed several important characteristics of cellular migration, a systemic behavior essential in cellular development, and the functional maintenance of both free-living cells and cells of multicellular organisms. In humans, embryogenesis, organogenesis, immune responses, and tissue repair, for instance, require very precise and complex migratory movements of cells. Any error in the control of these migratory processes may result in important consequences such as mental retardation, cardiovascular diseases, and cancer (Stuelten et al., 2018). The metastatic process allowing tumor cells to abandon the primary tumor and migrate to distant organs is a leading cause of death in patients with cancer. A better knowledge of the processes that control such migration will contribute to reducing the cancer-associated mortality (De la Fuente and López, 2020; Mezu-Ndubuisi and Maheshwari, 2020).
Cells are continuously receiving different biochemical and bioelectric signals, and our experiments have evidenced that unicellular organisms are capable to associate these signals and generate new adaptive migratory behaviors that can be remembered for long periods. This finding might be essential to understand some main principles of cellular systemic behavior and its adaptive strategies involved in enhancing their fitness to external medium.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://figshare.com/s/fa621110f0063d333d7c.
JC-P, CB, and MF performed the experiments. CB, MF, and ID designed the setup. CB performed the methodology with experimental glass chamber. MB and AG did the cell cultures and gave advice on cellular behavior. JC-P and CB performed the digitalization of trajectories. IM performed the quantitative studies. SK performed the gradient analysis. SK, MDB, and LM performed the main funding. MDB contributed to the laboratory facilities. SK, LM, MDB, GP-Y, JIL, and IM carried out the analysis and design of the research mapping. ID conceived, designed, and directed the investigation. All authors wrote the manuscript and agreed with its submission.
This work was supported by a grant of the University of Basque Country (UPV/EHU), GIU17/066, the Basque Government grant IT974-16, the UPV/EHU and Basque Center of Applied Mathematics, grant US18/21, and the Israel Science Foundation (536/19).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank Florentino Onandía Yague for valuable advice related to the experimental setup, and Laura Pérez Gómez for assistance in designing Supplementary Figure 1. We also express our gratitude to Dr. Alexey Smirnov (Saint Petersburg University, Russia) for valued help with amoeba imaging.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.707086/full#supplementary-material
Supplementary Figure 1 | Experimental set-up. (A) illustrates the top and lateral views of the experimental system (two standard electrophoresis blocks), in red, the anode where the peptide is initially dissolved, in gray the cathode, a: agar bridges b: mixing pipette c: experimental glass structure; arrow illustrating the flow of the laminar flux. (B) shows the glass chamber where the cells are placed. 1: standard glass slide; 2: longitudinal cover glasses fixed to the glass slide; 3: sliding cover glasses; 4: central cover glass under which the cells are initially placed; 5: experimental chamber where a laminar flux is created and the cells can migrate.
Supplementary Figure 2 | Experimental chamber. The specific set-up consists of two standard electrophoresis blocks, two agar bridges and a structure made from a standard glass slide and covers (the experimental chamber) (D–F) The glass structure is composed of one slide and two covers (A). The two covers are fixed to the glass slide with silicone (B), and then they are trimmed with a methacrylate ruler (C). This modified slide is placed in the central platform of the second electrophoresis block (D). To avoid that the medium goes across the modified slide, we placed an oil drop under, it in the central platform of the block of electrophoresis. Finally, a central piece of cover glass about 3 × 24 × 0.1 mm (E) and two sliding lateral cover pieces are placed on the modified slide (F). When the sliding cover pieces are moved closing the central part (see Supplementary Figure 1) an inner laminar flux is generated in the chamber and when they are open, cell placement and rescue are possible under the central piece of coverslips.
Supplementary Figure 3 | Supplementary electric device. The goal of these devices is to regulate and measure the electrical intensity that circulates through the system. 1, Standard Electrophoresis power supply, set at constant voltage. 2, 1 MΩ linear variable resistor that regulates the current of the system. 3, a micro ampere meter connected in series to the system, to measure the current intensity. 4, experimental setup described in Supplementary Figures 1, 2.
Supplementary Figure 4 | Conditioning process in Amoeba proteus using non-optimal electric field intensity. (A) Under simultaneous galvanotaxis and chemotaxis (induction process), 47% of the cells moved toward the anode-peptide (induced cells). These results agree with those obtained when using optimal intensity values for Amoeba proteus (70-74 μA, see Figure 2). (B) After the induction process, the cells were placed in Chalkley’s medium without any stimulus for 5 min, and then they were exposed to galvanotaxis for 30 min using non-optimal electric field intensity values (83-90 μA). 35% of the induced cells presented lasting directionality toward the anode (where the chemotactic peptide was absent) compared to 75% when using optimal intensity values for Amoeba proteus (see Figure 2). However, despite using non-optimal calibration, the cosine values obtained from the conditioned test were significantly different to those from the galvanotaxis (p−value = 10−5), indicating the emergence of a new behavior in the migration patterns. “N” total number of cells, “Er” experimental replicates, “nr” number of cells per replicate, “t” time of galvanotaxis or chemotaxis, “+” anode, “−” cathode. Both the x and y-axis show the distance in mm, and the initial location of each cell has been placed at the center of the diagram.
Supplementary Figure 5 | Fluorescein-tagged peptide concentration in the middle part of the laminar chamber flux. Fluorescein-tagged peptide concentration in the middle part of the laminar chamber flux as determined with a microplate reader. (A) heat map presenting individual fluorescence measurements at different times. (B) average levels of nFMLP-fluorescein concentration as time-function. The data represent the Mean ± SEM of 6 measurements (A–F, taken at 0, 2, 5, 10, 20, and 30 min). The peptide concentration in the middle of the glass experimental chamber (where the amoebae are placed) increases immediately following the flow establishment (within 2 min the concentration rises from zero to approximately 0.2 μM) and this concentration increases further (to 0.6 μM) for at least 30 min.
Armus, H. L., Montgomery, A. R., and Jellison, J. L. (2006). Discrimination learning in Paramecia (P. caudatum). Psychol. Record 56, 489–498. doi: 10.1007/BF03396029
Berdieva, M., Demin, S., and Goodkov, A. (2019). Amoeba proteus and ploidy cycles: from simple model to complex issues. Protistology 13, 166–173.
Boisseau, R. P., Vogel, D., and Dussutour, A. (2016). Habituation in non-neural organisms: evidence from slime moulds. Proc. R. Soc. Biol. Sci. Ser. B 283, 2–8. doi: 10.1098/rspb.2016.0446
Boussard, A., Delescluse, J., Pérez-Escudero, A., and Dussutour, A. (2019). Memory inception and preservation in slime moulds: the quest for a common mechanism. Philos. Trans. R. Soc. B 374:20180368. doi: 10.1098/rstb.2018.0368
De la Fuente, I. M. (2014). “Metabolic dissipative structures,” in Systems Biology of Metabolic and Signaling Networks: Energy, Mass and Information Transfer, eds M. A. Aon, V. Saks, and U. Schlattner (Berlin: Springer Berlin Heidelberg), 179–211. doi: 10.1007/978-3-642-38505-6_8
De la Fuente, I. M. (2015). Elements of the cellular metabolic structure. Front. Mol. Biosci. 2:16. doi: 10.3389/fmolb.2015.00016
De la Fuente, I. M., Bringas, C., Malaina, I., Fedetz, M., Carrasco-Pujante, J., Morales, M., et al. (2019a). Evidence of conditioned behavior in amoebae. Nat. Commun. 10, 3690–3690. doi: 10.1038/s41467-019-11677-w
De la Fuente, I. M., Bringas, C., Malaina, I., Regner, B., Pérez-Samartín, A., Boyano, M. D., et al. (2019b). The nucleus does not significantly affect the migratory trajectories of amoeba in two-dimensional environments. Sci. Rep. 9:16369. doi: 10.1038/s41598-019-52716-2
De la Fuente, I. M., Cortes, J. M., Pelta, D. A., and Veguillas, J. (2013). Attractor metabolic networks. PLoS One 8:e58284. doi: 10.1371/journal.pone.0058284
De la Fuente, I. M., Martínez, L., Carrasco-Pujante, J., Fedetz, M., López, J. I., and Malaina, I. (2021). Self-Organization and information processing: from basic enzymatic activities to complex adaptive cellular behavior. Front. Genet. 12:644615. doi: 10.3389/fgene.2021.644615
Demin, S., Berdieva, M., and Goodkov, A. (2019). Cyclic polyploidy in obligate agamic amoebae. Cell Tissue Biol. 13, 242–246. doi: 10.1134/s1990519x19030027
Dunigan, D. D., Al-Sammak, M., Al-Ameeli, Z., Agarkova, I. V., DeLong, J. P., and Van Etten, J. L. (2019). Chloroviruses lure hosts through long-distance chemical signaling. J. Virol. 93, e01688-18. doi: 10.1128/JVI.01688-18
Fais, S., and Fauvarque, M.-O. (2012). TM9 and cannibalism: how to learn more about cancer by studying amoebae and invertebrates. Trends Mol. Med. 18, 4–5. doi: 10.1016/j.molmed.2011.09.001
Finger, S. (2001). Origins of Neuroscience: A History of Explorations Into Brain Function. Oxford: Oxford University Press.
Gallistel, C. R. (2017). The coding question. Trends Cogn. Sci. 21, 498–508. doi: 10.1016/j.tics.2017.04.012
Gershman, J. S., Balbi, E. M. P., Gallistel, C. R., and Gunawardena, J. (2021). Reconsidering the evidence for learning in single cells. ELife 2021:e61907. doi: 10.7554/eLife.61907
Gibbs, J. W. (1902). Elementary Principles in Statistical Mechanics: Developed with Especial Reference to the Rational Foundations of Thermodynamics. New York, NY: Charles Scribner’s sons.
Goodkov, A., Yudin, A., and Podlipaeva, Y. (2014). Collection of the proteus-type amoebae at the Institute of Cytology, Russian Academy of Sciences. I. History, goals and research fields. Protistology 8, 71–75.
Grebecki, A. (1980). Behaviour of Amoeba proteus exposed to light-shade difference. Protistologica 16, 103–113.
Green, M. R., and Sambrook, J. (2012). Molecular Cloning: A Laboratory Manual. New York, NY: Cold Spring Harbor Laboratory Press.
Hawkins, R. D., and Byrne, J. H. (2015). Associative learning in invertebrates. Cold Spring Harb. Perspect. Biol. 7:a021709. doi: 10.1101/cshperspect.a021709
Hilsenbeck, O., Schwarzfischer, M., Skylaki, S., Schauberger, B., Hoppe, P. S., Loeffler, D., et al. (2016). Software tools for single-cell tracking and quantification of cellular and molecular properties. Nat. Biotechnol. 34, 703–706. doi: 10.1038/nbt.3626
Huna, A., Salmina, K., Erenpreisa, J., Vazquez-Martin, A., Krigerts, J., Inashkina, I., et al. (2015). Role of stress-activated OCT4A in the cell fate decisions of embryonal carcinoma cells treated with etoposide. Cell Cycle 14, 2969–2984. doi: 10.1080/15384101.2015.1056948
Jeon, K. W. (1995). The large, free-living amoebae: wonderful cells for biological studies. J. Eukaryot. Microbiol. 42, 1–7. doi: 10.1111/j.1550-7408.1995.tb01532.x
Korohoda, W., Golda, J., Sroka, J., Wojnarowicz, A., Jochym, P., and Madeja, Z. (1997). Chemotaxis of Amoeba proteus in the developing pH gradient within a pocket-like chamber studied with the computer assisted method. Cell Motil. Cytoskeleton 38, 38–53. doi: 10.1002/(sici)1097-0169(1997)38:1<38::aid-cm5>3.0.co;2-d
Korohoda, W., Mycielska, M., Janda, E., and Madeja, Z. (2000). Immediate and long-term galvanotactic responses of Amoeba proteus to dc electric fields. Cell Motil. Cytoskeleton 45, 10–26. doi: 10.1002/(sici)1097-0169(200001)45:1<10::aid-cm2>3.0.co;2-t
Kramar, M., and Alim, K. (2021). Encoding memory in tube diameter hierarchy of living flow network. Proc. Natl. Acad. Sci. U.S.A. 118:e2007815118. doi: 10.1073/pnas.2007815118
Lee, C. K., de Anda, J., Baker, A. E., Bennett, R. R., Luo, Y., Lee, E. Y., et al. (2018). Multigenerational memory and adaptive adhesion in early bacterial biofilm communities. Proc. Natl. Acad. Sci. U.S.A. 115, 4471–4476. doi: 10.1073/pnas.1720071115
Lugini, L., Matarrese, P., Tinari, A., Lozupone, F., Federici, C., Iessi, E., et al. (2006). Cannibalism of live lymphocytes by human metastatic but not primary melanoma cells. Cancer Res. 66, 3629–3638. doi: 10.1158/0008-5472.CAN-05-3204
Mezu-Ndubuisi, O. J., and Maheshwari, A. (2020). The role of integrins in inflammation and angiogenesis. Pediatr. Res. 89, 1619–1626. doi: 10.1038/s41390-020-01177-9
Pavlov, I. P. (1927). Conditioned Reflexes: An Investigation of the Physiological Activity of the Cerebral Cortex. Oxford: Oxford University Press.
Prescott, D. M., and Carrier, R. F. (1964). “experimental procedures and cultural methods for euplotes eurystomus and Amoeba proteus,” in Methods in Cell Biology, ed. D. M. Prescott (New York, NY: Academic Press), 85–95. doi: 10.1016/s0091-679x(08)62087-7
Prusch, R. D., and Britton, J. C. (1987). Peptide stimulation of phagocytosis in Amoeba proteus. Cell Tissue Res. 250, 589–593. doi: 10.1007/BF00218951
Rescorla, R. A. (1967). Pavlovian conditioning and its proper control procedures. Psychol. Rev. 74, 71–80. doi: 10.1037/h0024109
Sanchez-Andres, J. V., Pomares, R., and Malaisse, W. (2020). Adaptive short-term associative conditioning in the pancreatic β−cell. Physiol. Rep. 8:e14403. doi: 10.14814/phy2.14403
Shirakawa, T., Gunji, Y. P., and Miyake, Y. (2011). An associative learning experiment using the plasmodium of Physarum polycephalum. Nano Commun. Netw. 2, 99–105. doi: 10.1016/j.nancom.2011.05.002
Skoge, M., Yue, H., Erickstad, M., Bae, A., Levine, H., Groisman, A., et al. (2014). Cellular memory in eukaryotic chemotaxis. Proc. Natl. Acad. Sci. U.S.A. 111, 14448–14453. doi: 10.1073/pnas.1412197111
Smirnov, A. V. (2008). “Amoebas, lobose,” in Encyclopedia of Microbiology, ed. M. Schaechter (Oxford: Elsevier), 558–577. doi: 10.1016/b978-012373944-5.00359-x
Sopina, V. A. (2000). Electrophoretic forms of glucose-6-phosphate dehydrogenase, acid phosphatase and esterase in Amoeba species amoebas. Tsitologiia 42, 1134–1143.
Stuelten, C. H., Parent, C. A., and Montell, D. J. (2018). Cell motility in cancer invasion and metastasis: insights from simple model organisms. Nat. Rev. Cancer 18, 296–312. doi: 10.1038/nrc.2018.15
Tagkopoulos, I., Liu, Y. C., and Tavazoie, S. (2008). Predictive behavior within microbial genetic networks. Science 320, 1313–1317. doi: 10.1126/science.1154456
Tang, S. K. Y., and Marshall, W. F. (2018). Cell learning. Curr. Biol. 28, R1180–R1184. doi: 10.1016/j.cub.2018.09.015
Tinevez, J. Y., Perry, N., Schindelin, J., Hoopes, G. M., Reynolds, G. D., Laplantine, E., et al. (2017). TrackMate: an open and extensible platform for single-particle tracking. Methods 115, 80–90. doi: 10.1016/j.ymeth.2016.09.016
Yang, C. Y., Bialecka-Fornal, M., Weatherwax, C., Larkin, J. W., Prindle, A., Liu, J., et al. (2020). Encoding membrane-potential-based memory within a microbial community. Cell Syst. 10, 417-423.e3. doi: 10.1016/j.cels.2020.04.002
Keywords: cellular migration, systemic behavior, galvanotaxis, chemotaxis, associative conditioning, learning, Pavlov’s experiments
Citation: Carrasco-Pujante J, Bringas C, Malaina I, Fedetz M, Martínez L, Pérez-Yarza G, Dolores Boyano M, Berdieva M, Goodkov A, López JI, Knafo S and De la Fuente IM (2021) Associative Conditioning Is a Robust Systemic Behavior in Unicellular Organisms: An Interspecies Comparison. Front. Microbiol. 12:707086. doi: 10.3389/fmicb.2021.707086
Received: 18 May 2021; Accepted: 22 June 2021;
Published: 19 July 2021.
Edited by:
Haike Antelmann, Freie Universität Berlin, GermanyReviewed by:
Falk Hillmann, Leibniz Institute for Natural Product Research and Infection Biology, GermanyCopyright © 2021 Carrasco-Pujante, Bringas, Malaina, Fedetz, Martínez, Pérez-Yarza, Dolores Boyano, Berdieva, Goodkov, López, Knafo and De la Fuente. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shira Knafo, c2hpcmFrQHBvc3QuYmd1LmFjLmls; Ildefonso M. De la Fuente, bXRwbWFkZWlAZWh1LmV1cw==; aWxkZWZvbnNvQGNlYmFzLmNzaWMuZXM=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.