
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 09 July 2021
Sec. Fungi and Their Interactions
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.689374
This article is part of the Research TopicFungal Systematics and BiogeographyView all 27 articles
 Gislaine C. S. Melanda1†
Gislaine C. S. Melanda1† Alexandre G. S. Silva-Filho2†
Alexandre G. S. Silva-Filho2† Alexandre Rafael Lenz3
Alexandre Rafael Lenz3 Nelson Menolli Jr.4,5*
Nelson Menolli Jr.4,5* Alexandro de Andrade de Lima1
Alexandro de Andrade de Lima1 Renato Juciano Ferreira6
Renato Juciano Ferreira6 Nathalia Mendonça de Assis2
Nathalia Mendonça de Assis2 Tiara S. Cabral7
Tiara S. Cabral7 María P. Martín8
María P. Martín8 Iuri Goulart Baseia1,2
Iuri Goulart Baseia1,2The order Phallales (Basidiomycota) is represented by gasteroid fungi with expanded and sequestrate basidiomata, known as stinkhorns and false truffles. In phalloids, the first DNA sequence was published in 1997, and after that, some studies aimed to resolve phylogenetic conflicts and propose new species based on DNA markers; however, the number of families and genera in the order still generates controversies among researchers. Thus, this work aims to provide an overview of Phallales diversity represented by selected DNA markers available in public databases. We retrieved Phallales sequences from DNA databases (GenBank and UNITE) of seven markers: ITS (internal transcribed spacer), nuc-LSU (nuclear large subunit rDNA), nuc-SSU (nuclear small subunit rDNA), mt-SSU (mitochondrial small subunit rDNA), ATP6 (ATPase subunit 6), RPB2 (nuclear protein-coding second largest subunit of RNA polymerase), and TEF1-α (translation elongation factor subunit 1α). To compose our final dataset, all ITS sequences retrieved were subjected to BLASTn searches to identify additional ITS sequences not classified as Phallales. Phylogenetic analyses based on Bayesian and maximum likelihood approaches using single and combined markers were conducted. All ITS sequences were clustered with a cutoff of 98% in order to maximize the number of species hypotheses. The geographic origin of sequences was retrieved, as well as additional information on species lifestyle and edibility. We obtained a total of 1,149 sequences, representing 664 individuals. Sequences of 41 individuals were unidentified at genus level and were assigned to five distinct families. We recognize seven families and 22 genera in Phallales, although the delimitation of some genera must be further revisited in order to recognize only monophyletic groups. Many inconsistencies in species identification are discussed, and the positioning of genera in each family is shown. The clustering revealed 118 species hypotheses, meaning that approximately 20% of all described species in Phallales have DNA sequences available. Information related to geographic distribution represents 462 individuals distributed in 46 countries on all continents, except Antarctica. Most genera are saprotrophic with only one putative ectomycorrhizal genus, and 2.1% of the legitimate specific names recognized in Phallales are confirmed edible species. Great progress in the molecular analyses of phalloids has already been made over these years, but it is still necessary to solve some taxonomic inconsistencies, mainly at genus level, and generate new data to expand knowledge of the group.
The first molecular analyses including gasteroid fungi (Hibbett et al., 1997) showed that they represent a polyphyletic and artificial grouping of taxa that share a common ancestor with gilled and nongilled mushrooms that have active dispersion of the basidiospores. Hibbett et al. (1997) showed the gasteroid–phalloid fungi to group in a clade with other forms of gasteroid fungi, the Geastraceae Corda (earthstars) and Sphaerobolaceae J. Schröt. (cannonballs), all sharing a common ancestor with coralloid fungi. After that, Pine et al. (1999) confirmed the phylogenetic relationship between some cantharelloid, clavarioid, and phalloid fungi, naming all of them the gomphoid–phalloid clade, which was later confirmed as monophyletic (Hibbett and Thorn, 2001; Krüger et al., 2001; Binder and Hibbett, 2002; Hibbett and Binder, 2002) and designated as a subclass of Agaricomycetes: Phallomycetidae K. Hosaka, Castellano and Spatafora (Hosaka et al., 2006).
Phallales E. Fisch (Agaricomycetes, Phallomycetidae) includes representatives of gasteroid fungi with basidiospores that are passively dispersed mainly by insects and commonly named phalloid fungi, alien fungi, stinkhorns, and lattice or cage stinkhorns (Fischer, 1898; Cunningham, 1931; Miller and Miller, 1988; Pegler and Gomez, 1994). The phalloid fungi are mostly saprobic and characterized by hypogeous or epigeous immature basidiomata that are divided into chambers; thick white rhizomorphs are usually present; peridium with two or three layers, one of which is gelatinous; mature basidiomata that are usually epigeous or partially hypogeous, expanded, or sequestrate; pseudostipitate or sessile receptacle; receptacle carrying the green, olive to brown gleba; the usual presence of gelatinous to mucilaginous gleba that may be powdery at maturity, as in Gastrosporium Mattir.; and basidiospores mostly ellipsoid and smooth, with only a few genera with ornamentation on the basidiospore wall, as in Gastrosporium, Kjeldsenia Colgan, Castellano and Bougher, and Phlebogaster Fogel (Hosaka et al., 2006; Trierveiler-Pereira et al., 2014a).
Fischer (1898) grouped in Phallales the families Clathraceae Chevall. and Phallaceae Corda, which include specimens with expanded branched and unbranched basidiomata, respectively. Lysuraceae Corda was established in the same article as the establishment of Phallaceae (Corda, 1842); both families are characterized by basal pseudostipitate basidiomata, but unlike the morphology in Phallaceae, the apical part of the basidiomata in Lysuraceae is branched. Lysuraceae was not accepted by Fischer (1898) as an independent family in Phallales, and their species were considered by this author within Clathraceae, a view also adopted by a number of subsequent authors such as Cunningham (1944), Dring (1980), Jülich (1981), Miller and Miller (1988), Pegler and Gomez (1994), Hibbett and Thorn (2001), and Kirk et al. (2008). On the other hand, some authors (He et al., 2019; Wijayawardene et al., 2020) and well-known databases (Nilsson et al., 2019; Index Fungorum, 2020; MycoBank., 2020; Schoch et al., 2020) have placed in Phallaceae all the species traditionally classified as Lysuraceae. However, Hosaka et al. (2006), Degreef et al. (2013), Trierveiler-Pereira et al. (2014a), and Sulzbacher et al. (2016) consider these three families to be independent (Table 1).
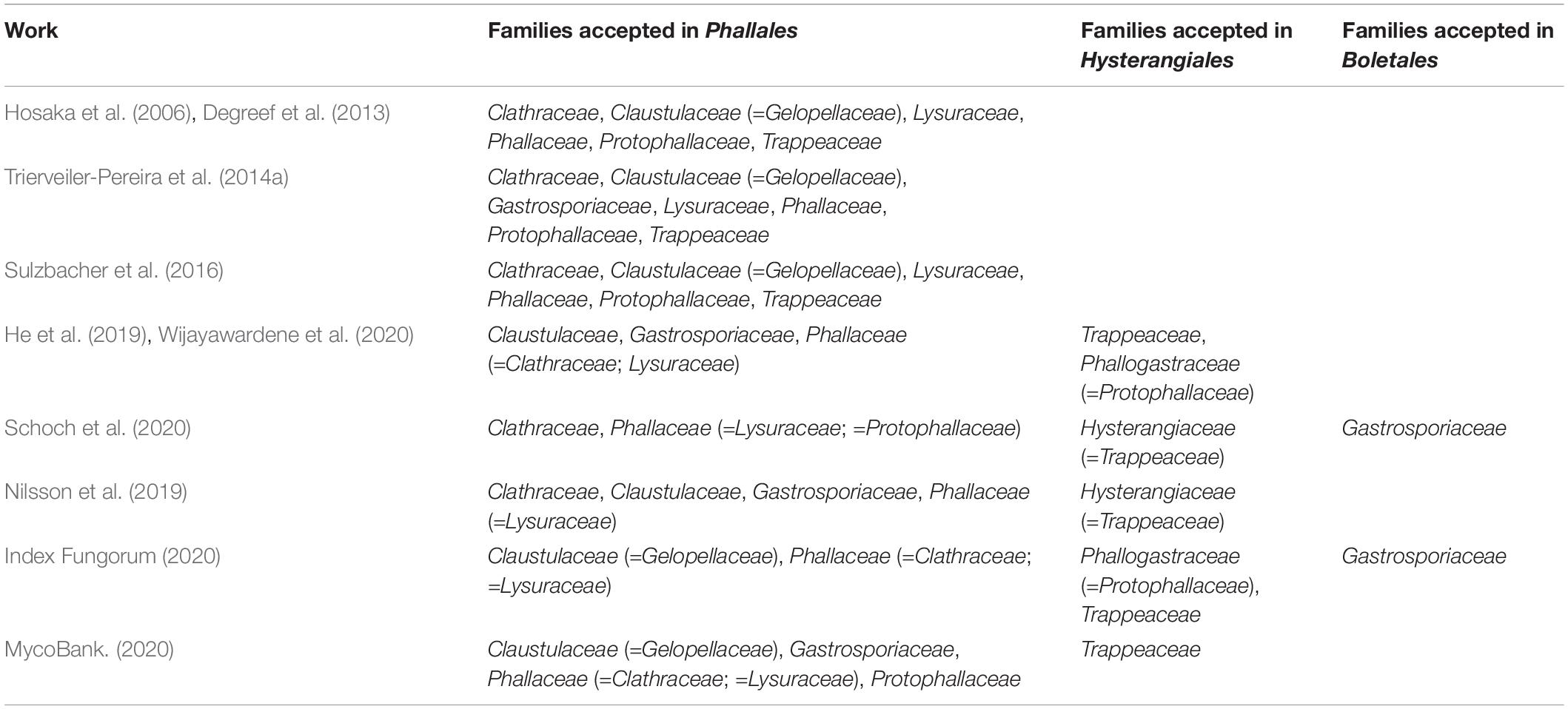
Table 1. Family classification of Phallales and related orders according to published molecular studies and taxonomic and molecular databases.
Cunningham (1931) accepted in Phallales sequestrate (truffle-like) basidiomata classified in the genus Claustula K.M. Curtis and established this genus as a type of the new monogeneric family Claustulaceae G. Cunn. As mentioned by Cunningham (1931), this family shares with other phalloid fungi the gelatinous peridium, immature basidiomata divided into chambers, and elliptical smooth basidiospores. Zeller (1939) also studied sequestrate fungi and proposed two new families: Gelopellaceae Zeller and Protophallaceae Zeller. However, these two families, as well as Hysterangiaceae E. Fisch., have been consolidated within the order Hysterangiales K. Hosaka and Castellano, according to Zeller (1939). Hysterangiales was first proposed by Zeller (1939), but it was considered a nomen nudum because it was published without a description or diagnosis; it was later formally established in Hosaka et al. (2006).
Moreover, Hosaka et al. (2006) established the subclass Phallomycetidae, including the new order Hysterangiales mentioned previously and the new Geastrales K. Hosaka and Castellano. Thus, in the study by Hosaka et al. (2006), Phallomycetidae comprises Geastrales, Gomphalles Jülich., Hysterangiales, and Phallales, a classification that is also accepted in He et al. (2019). In this way, Hosaka et al. (2006) agreed with Cunningham (1931) in the definition of Phallales including families with expanded basidiomata (Clathraceae, Lysuraceae, Phallaceae), as well as with sequestrate ones (Claustulaceae, Trappeaceae P.M. Kirk, and Protophallaceae). Although Trappeaceae was provisionally proposed in Phallales by Hosaka et al. (2006) with the genera Phallobata G. Cunn. and Trappea Castellano, the family was formally proposed two years later by Kirk et al. (2008), who classified it as part of Hysterangiales. Later, Sulzbacher et al. (2016) proposed a new genus in Trappeaceae (Restingomyces Sulzbacher, T. Grebenc and Baseia) and also considered the family in Phallales as previously pointed out by Hosaka et al. (2006).
After these 24 years of molecular studies, the sampling of some genera and families within molecular phylogenies of Phallales is incomplete. For example, Gastrosporium, which is part of the monogeneric family Gastrosporiaceae Pilát, was first incorporated in a molecular phylogenetic work by Hibbett and Binder (2002), who considered it as a sequestrate member of Phallales. This was later confirmed by Trierveiler-Pereira et al. (2014a), He et al. (2019), Kasuya et al. (2020), and Wijayawardene et al. (2020). However, Hosaka et al. (2006) did not include Gastrosporium in their phylogeny because of the lack of a protein code gene in their dataset. In the study by Trierveiler-Pereira et al. (2014a), Trappeaceae representatives were not included, and they recognized seven families in Phallales, which include Gastrosporiaceae plus the six families recognized by Hosaka et al. (2006).
On the other hand, the acceptance in Phallales of families composed of species characterized by sequestrate basidiomata (Claustulaceae, Gastrosporiaceae, Gelopellaceae, Trappeaceae, and Protophallaceae) has also generated controversies. For instance, morphological studies conducted by Zeller (1939, 1948) and Jülich (1981) considered Gelopellaceae and Protophallaceae in Hysterangiales, whereas Miller and Miller (1988) considered these families and Claustulaceae in Phallales. The last edition of Dictionary of the Fungi (Kirk et al., 2008) recognized three families in Phallales: Claustulaceae (=Gelopellaceae), Gastrosporiaceae, and Phallaceae (=Clathraceae; = Lysuraceae; = Protophallaceae), with Trappeaceae placed in Hysterangiales. Additionally, recent works based on Basidiomycota and general fungal classification (He et al., 2019; Wijayawardene et al., 2020) also considered Trappeaceae in Hysterangiales and only three families in Phallales: Claustulaceae, Gastrosporiaceae, and Phallaceae (Table 1). Representatives of some families of Phallales are shown in Figure 1.

Figure 1. Basidiomata of Phallales. Clathraceae: (A) Aseroë rubra; (B) Clathrus archeri MT13072101B (personal herbarium); (C) Laternea dringii (coll. AAL94); (D) Blumenavia baturitensis UFRN-Fungos 2868, holotype (coll. GCSM15); (E) Ileodictyon gracile MT141003001B (personal herbarium); (F) Clathrus ruber MT15100406B (personal herbarium); (G) Clathrus columnatus UFRN-Fungos 2912 (coll. AAL42); (H) Clathrus natalensis (coll. GSCM30); (I) Abrachium floriforme UFRN-Fungos 3271. Lysuraceae: (J) Lysurus arachnoideus INPA-Fungos 256537 (coll. TSC41). Phallaceae: (K) Phallus indusiatus INPA-Fungos 264931, neotype (coll. TSC148); (L) Phallus impudicus; (M) Itajahya sp. UFRN-Fungos 3342 (coll. AAL106); (N) Mutinus bambusinus UFRN-Fungos 3222 (coll. AAL100); (O) Mutinus caninus; (P) Xylophallus clavatus INPA-Fungos 271637 (coll. TSC237). Protophallaceae: (Q,R) Protubera maracuja (coll. GCSM18). Trappeaceae: (S) Restingomyces reticulatus UFRN-Fungos 1890, holotype (coll. MAS335). Photographs: (A) Clark L. Ovrebo; (B,E,F,L,O) Manuel Tabarés; (C,G,M,N) Alexandro de A. de Lima; (D,H,Q,R) Gislaine C. S. Melanda; (I) Alexandre G. S. Silva-Filho; (J,K,P) Tiara S. Cabral; (S) Marcelo A. Sulzbacher.
There is still no consensus in family level systematics of Phallales, based on a compilation of sources (Table 1), such as works of fungal classification (He et al., 2019; Wijayawardene et al., 2020), taxonomic (Index Fungorum, 2020; MycoBank., 2020) and molecular databases (Nilsson et al., 2019; Schoch et al., 2020), and phylogenetic studies focused on Phallales (Hosaka et al., 2006; Degreef et al., 2013; Trierveiler-Pereira et al., 2014a; Sulzbacher et al., 2016). Thus, based on the importance of molecular data for systematics and phylogenetic studies and the fact that DNA databases can be a good tool to assess the history behind the sequences generated over the years, as well as the geographic distribution of certain taxa, we have undertaken this study using Phallales as a target group to retrieve sequences in molecular databases and to provide an overview of Phallales diversity represented by selected DNA markers available in public databases. Moreover, we aim to test the phylogenetic positioning of named and unnamed sequences; to assess phylogenetic hypotheses using combined markers to recognize families and genera and to compare these data with the extant classification; to recognize the total number of species hypothesis (SH) represented in Phallales based on internal transcribed spacer (ITS) sequence clustering; and, finally, to record the global geographic distribution of their representative genera, their lifestyle, and edibility.
Our work used two databases to obtain Phallales sequences: NCBI GenBank1 and UNITE2. All sequences were downloaded on August 7 to 9, 2020, from both databases. GenBank is part of the International Nucleotide Sequence Database Collaboration and contains the vast majority of phalloid sequences from published articles. The UNITE database automatically clusters ITS sequences of eukaryotic organisms to approximately the species level (called SHs), and to facilitate unambiguous scientific communication, a DOI is given to each SH.
According to the revised bibliography already mentioned (Hibbett et al., 1997; Hosaka et al., 2006; Trierveiler-Pereira et al., 2014a; Sulzbacher et al., 2016; Kasuya et al., 2020), seven markers were selected to retrieve sequences in GenBank using query strings (Table 2): nuclear ribosomal ITS, nuclear large subunit rDNA (nuc-LSU), nuclear small subunit rDNA (nuc-SSU); mitochondrial small subunit rDNA (mt-SSU), mitochondrial protein-coding ATPase subunit 6 (ATP6), and nuclear protein-coding second largest subunit of RNA polymerase (RPB2); and nuclear protein-coding translation elongation factor subunit 1α (TEF1-α). GenBank query results were downloaded in TinySeq_XML format, and single datasets of nucleotide sequences were created for each marker. One single dataset of all ribosomal markers from GenBank was downloaded, and in order to identify each marker, this single dataset was separated manually, based on the marker name in each sequence title. From the UNITE database, the sequences under Phallales (DOI: TH005985) not placed in GenBank were retrieved manually. Furthermore, manual checking was done for some genera, as they are not classified in Phallales in GenBank (Gastrosporium, Kjeldsenia, Phallobata, Phlebogaster, and Trappea) and UNITE (Phlebogaster and Trappea).

Table 2. Query strings used to search for DNA sequences from phalloid fungi in GenBank.
Sequence metadata were retrieved from GenBank qualifiers and UNITE annotations. GenBank qualifiers include3 : country, collection_date, culture_collection, environmental_sample, clone, isolate, isolation_source, lat_lon, organism, specimen_voucher, strain, tissue_type, type_material, and PCR_primers, as well as information about authors, reference, title, and journal. UNITE annotations were obtained directly from the online database for each sequence; UNITE metadata include sampling_area (country), sample_type [Linked to (source)], and collection_date.
All ITS sequences obtained from GenBank and UNITE were used for additional Nucleotide BLAST searches in their respective database (NCBI BLAST, 2020; UNITE BLAST, 2020). These searches aimed to find ITS sequences of Phallales members that were deposited without being classified in this order. We retrieved unclassified sequences using the following cutoffs: query cover > 80%, identity > 70%, and e-value < e-1,000. The 80% sequence similarity represents the criterion to recognize the identity of sequences approximately at the order level (Tedersoo et al., 2014).
All information from the sequences retrieved from both databases and the BLAST searches was merged manually and organized in Supplementary Table 1 to better identify all markers of each individual based on their herbarium/culture accession number and/or other code given by whoever generated the sequences. An individual was considered repeated when it had more than one sequence of the same marker under different accession numbers or when it was indicated as a clone in the databases (individuals with “R” in the columns “R = repeated or duplicated voucher” of Supplementary Table 1). In these cases, two lines were created in Supplementary Table 1 for the same individual. The names of genera and species present in our Supplementary Table 1 are based on the qualifier organism in GenBank for each individual. In UNITE, the names adopted were according to the UNITE taxon name.
Phylogenetic analyses were performed using seven datasets, the combined one and the other six with each individual marker: ITS, nuc-LSU, mt-SSU, ATP6, RPB2, and TEF1-α. The marker nuc-SSU was not considered in the analyses because of the few (1%) sequences available (Supplementary Table 1). Sequences of Hysterangiales species were used as outgroup for individual marker analyses, as seen in Supplementary Table 2. The combined dataset (ITS + nuc-LSU + mt-SSU + ATP6 + RPB2 + TEF1-α) was constructed to confirm the organization in families, and this also used members of Hysterangiales as outgroup (Supplementary Table 3). Specimens from almost all genera retrieved in Phallales and that had most markers sequenced were chosen for the combined dataset (Supplementary Table 3), which includes 139 sequences: 69 ITS, 114 nuc-LSU, 26 mt-SSU, 76 ATP6, 65 RPB2, and 41 TEF1-α.
All datasets were aligned using MAFFT v.7 (Katoh and Standley, 2013) under the E-INS-i criteria. Seaview v.4 (Gouy et al., 2010) was used to visualize and adjust the alignments. The RPB2 alignment was partitioned into intron and exon, and TEF1-α alignment into intron 1/2/3 and exon 1/2 according to GenBank coding sequence notation. The best nucleotide substitution model was selected with BIC (Bayesian information criterion) using jModelTest 2v.1.6 (Darriba et al., 2012) for each individual dataset. Two strategies were used for phylogenetic reconstructions of each alignment: maximum likelihood and Bayesian inference. Maximum likelihood analyses were performed in RAxML v8.2.X (Stamatakis, 2006), combined with the rapid bootstrapping algorithm with 1,000 replicates under the GTRGAMMA option to obtain the maximum likelihood bootstrap (MLbs). Bayesian inferences were performed using MrBayes 3.2.6 (Ronquist et al., 2012) with two independent runs, each one beginning from random trees with four simultaneous independent chains, performing 2 × 107 Markov chain Monte Carlo (MCMC) generations, sampling one tree every 1 × 103 generation. The first 5 × 103 sampled trees were discarded as burn-in, whereas the remaining ones (all sampled after the average standard deviation of split frequencies reached < 0.01) were used to reconstruct a 50% majority-rule consensus tree and to calculate Bayesian posterior probabilities (PP) of the clades. The jModelTest 2v.1.6, RAxML v8.2.X, and MrBayes 3.2.6 were run from the CIPRES Science Gateway 176 3.1 (Miller et al., 2010). All final alignments and the resulting topologies were deposited in TreeBASE under the number 28016.
The ITS sequences of Phallales retrieved were clustered using CD-HIT-EST (Huang et al., 2010) at 98.0% sequence similarity threshold (Tedersoo et al., 2014) to assess species hypotheses in Phallales.
The qualifiers country and lat_lon from GenBank and sampling_area from UNITE were used to organize the global geographic distribution map of Phallales and each genus. To construct the maps based on this global distribution, the locality of each individual identified at genus level was retrieved from the databases (Supplementary Table 4). When possible, the missing localities in DNA databases were double searched in their original article or in secondary articles that give this information (Supplementary Table 4). When the geographic coordinate information was missing, we made an effort to establish it through Google Maps4 to complete the table. In the case of individuals for which the only origin information was the country, the geographic coordinates suggested by Google Maps were used based on the name of the respective country as a search keyword. One general map with all Phallales distribution was made, as well as separate maps according to genera.
To record the information related to lifestyle of each genus, we followed the FungalTraits database (Põlme et al., 2021) and additional data obtained from the qualifiers environmental_sample (GenBank), isolation_source (GenBank), and sample_type (UNITE). To complement and better explore the use of the phalloid species sampled with molecular data, the edibility status of the species was considered based on Li et al. (2021) and complementarily searched in the main text of the articles in which the sequences were generated.
Our final dataset led to a total of 1,149 DNA sequences of Phallales divided into 492 ITS, 303 nuc-LSU, 11 nuc-SSU, 75 mt-SSU, 129 ATP6, 88 RPB2, and 51 TEF1-α (Figure 2).
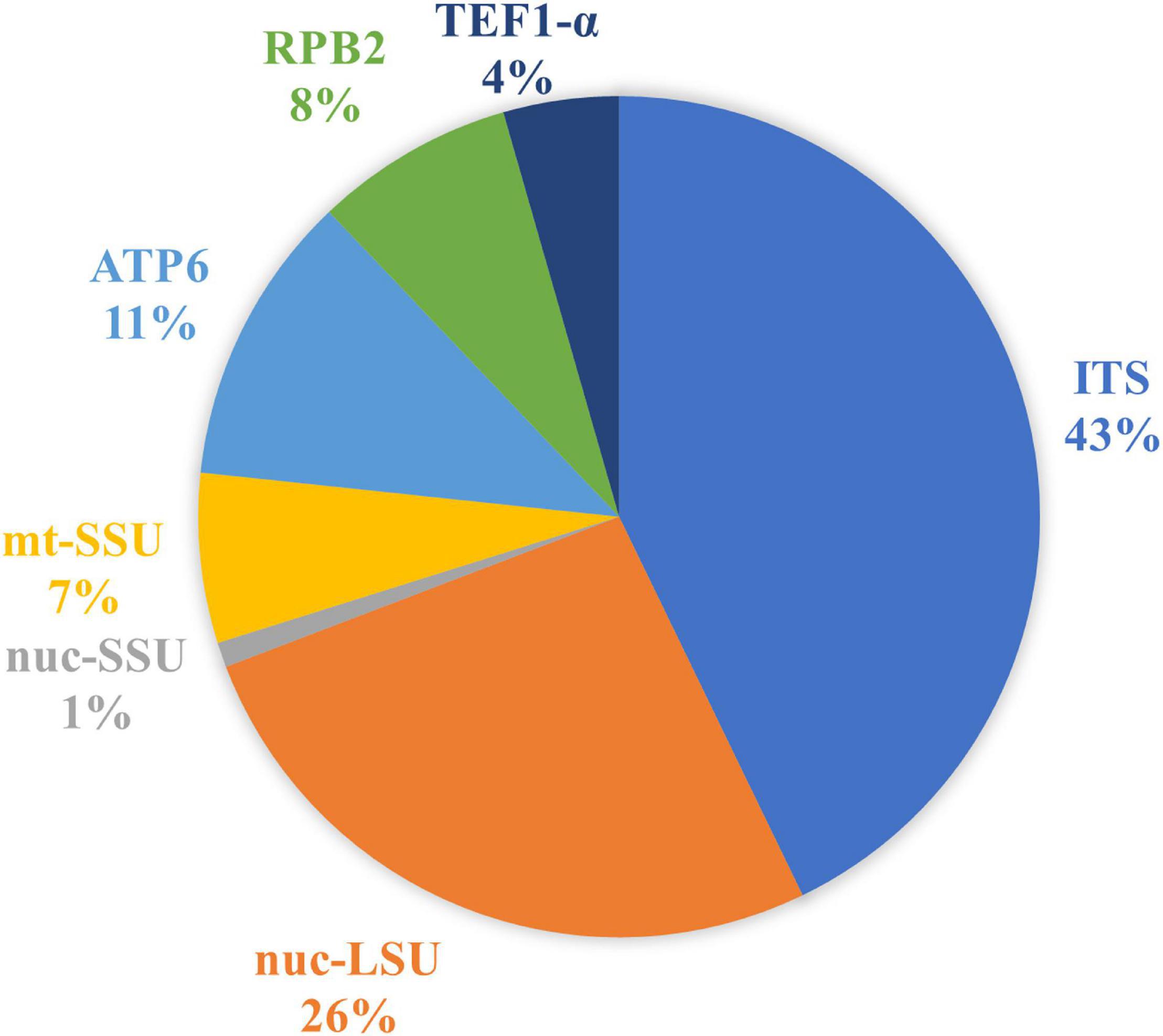
Figure 2. Relative proportion of phalloid DNA sequences deposited in DNA databases (GenBank and UNITE) according to the molecular marker.
Based on the available records of collection date (Figure 3), the first uploaded sequences of Phallales individuals were the nuc-SSU sequences of Pseudocolus fusiformis (E. Fisch.) Lloyd, dated October 31, 1997 (AF026623), and November 5 of the same year (AF026666), both part of the work published by Hibbett et al. (1997). In 2006, there was a peak of deposited sequences, including new markers such as ATP6, RPB2, and TEF1-α (Figure 3), and in 2012, there was a higher constancy of deposited sequences, in which ITS and nuc-LSU are the most represented markers (Figure 3). Interestingly, the sequence obtained from the oldest phalloid individual belongs to Colus hirudinosus Cavalier and Séchier (voucher UC 955042, dated February 1, 1952) and was uploaded in GenBank on April 4, 2020 (nuc-LSU accession code MK607412, author of sequence: Kuo, M.).
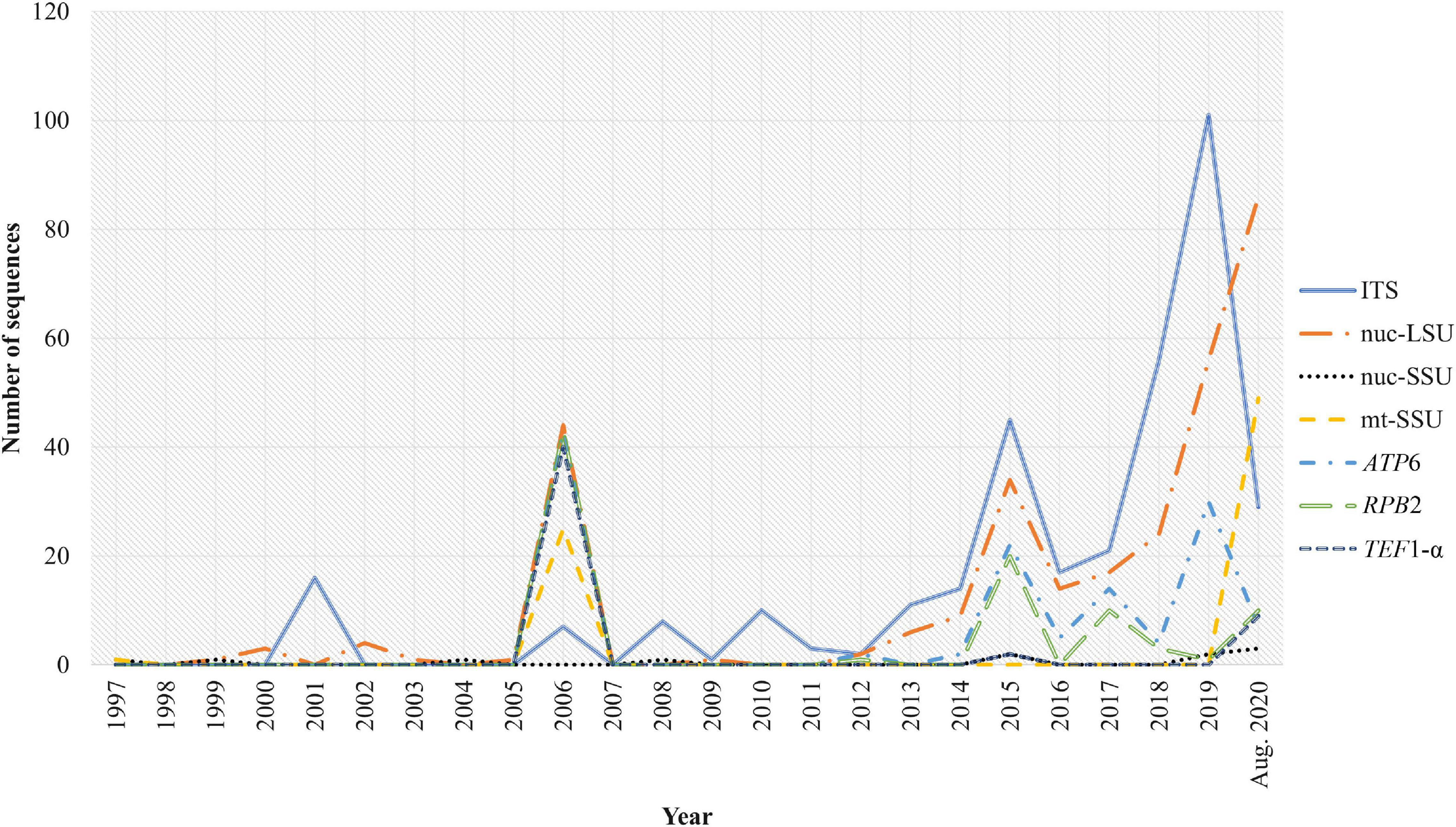
Figure 3. Number of Phallales DNA sequences uploaded in DNA databases (GenBank and UNITE) along the years.
The 1,149 sequences comprise 664 individuals, with 19 of them comprising repeated sequences for the same maker. A total of 58.7% of all individuals have only one sequenced marker, 23.9% have two sequenced markers, and 17.4% have three to five sequenced markers. There are 122 sequences representing 41 type collections (Supplementary Table 5).
Sequences of 22 recognized genera were retrieved: Abrachium Baseia and T.S. Cabral, Aseroë Labill., Blumenavia Möller, Clathrus P. Micheli ex L., Claustula, Colus Cavalier and Séchier, Gastrosporium, Gelopellis Zeller, Ileodictyon Tul. and C. Tul., Itajahya Möller, Kjeldsenia, Laternea Turpin, Lysurus Fr., Mutinus Fr. (=Jansia Penz.), Phallobata, Phallus Junius ex L. (=Dictyophora Desv.), Phlebogaster, Protubera Möller, Pseudocolus Lloyd, Restingomyces (as Phallales sp.), Trappea, and Xylophallus (Schltdl.) E. Fisch. (Table 3). Sequences named Gymnotelium Syd. were retrieved as Phallales, but they are excluded from the present study because of their doubtful quality and because it is a genus classified in Pucciniaceae Chevall., Pucciniales Clem. and Shear (He et al., 2019). The genus Calvarula Zeller was not included in the combined analyses, because only one TEF1-α sequence is available and its placement at family level is questionable (see Phylogenetic analyses).
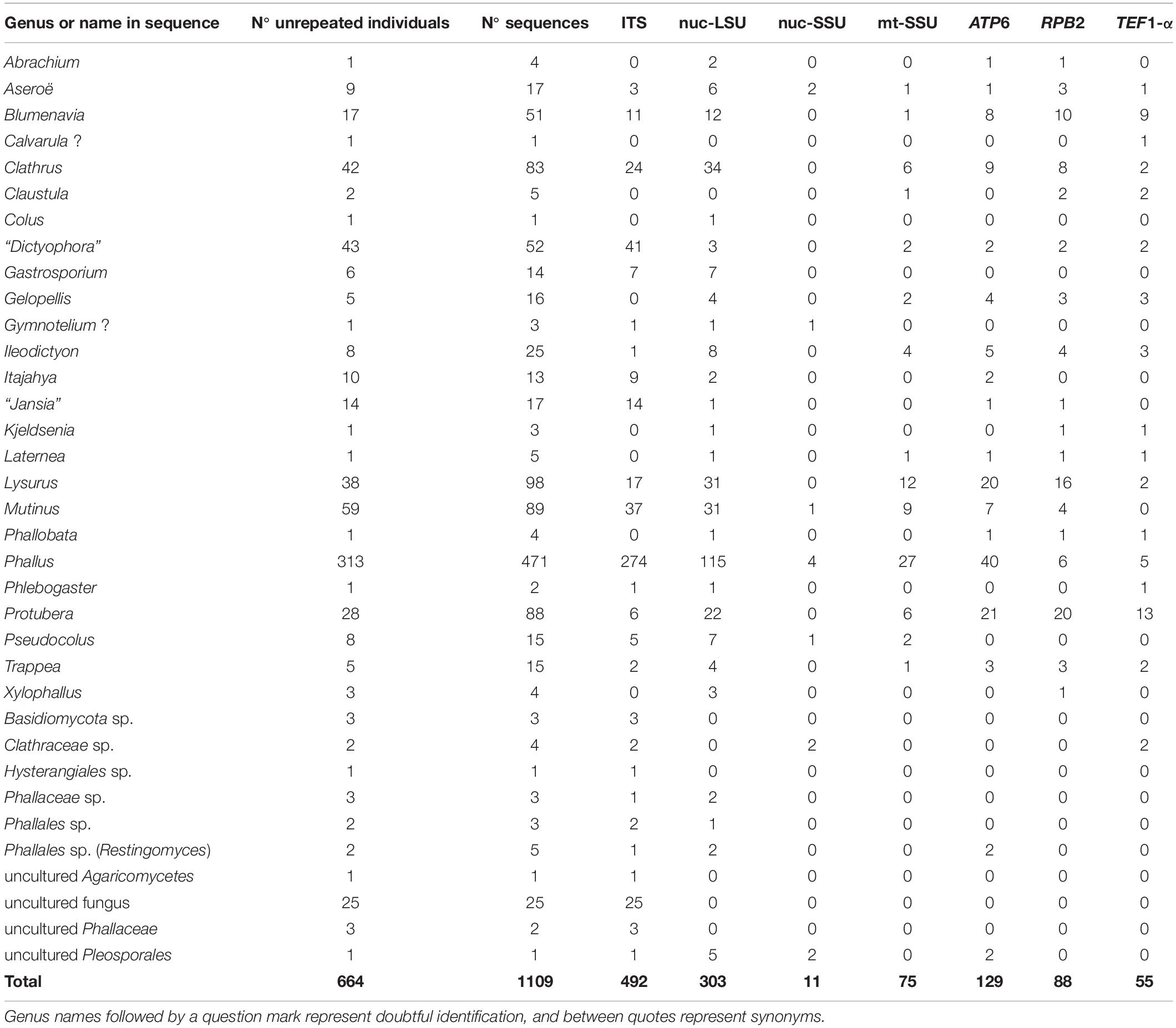
Table 3. Total of individuals and number of sequences of each molecular marker for each taxon retrieved from GenBank and UNITE databases from phalloid fungi searches.
Phallus is the most highly represented genus, with 313 individuals and 471 sequences (Table 3). Forty-three unidentified Phallales individuals were not classified at genus level (see individuals with “NO” in genus columns in Supplementary Table 1); they are specified in Table 3. The possible classification of these individuals is discussed in Phylogenetic analyses.
The final aligned matrices for the analyses of each independent marker contain 481 sequences of ITS (1,210 positions), 306 nuc-LSU (1,245 positions), 77 mt-SSU (600 positions), 131 ATP6 (743 positions), 90 RPB2 (822 positions), and 53 TEF1-α (849 positions), and 139 of the combined matrix (4637 positions). The evolutionary models selected for the final dataset were as follows: ITS: TIM1 + I + G; nuc-LSU: TrN + I + G; mt-SSU: TPM3uf + G; ATP6: TVM + I + G; RPB2 Intron: TIM3 + I + G, RPB2 exon: TIM3 + I + G; TEF1-α intron 1: TrNef + G, TEF1-α exon 1: TrN + I + G, TEF1-α intron 2: TPM1 + G; TEF1-α exon 2: TPM1 + G, and TEF1-α intron 3: K80 + G.
The concatenated tree is shown in Figure 4, in which some names were changed according to the current name recognized by their phylogenetic positioning. The trees that resulted from the independent analyses of each marker are available in Supplementary Figures 1–3.
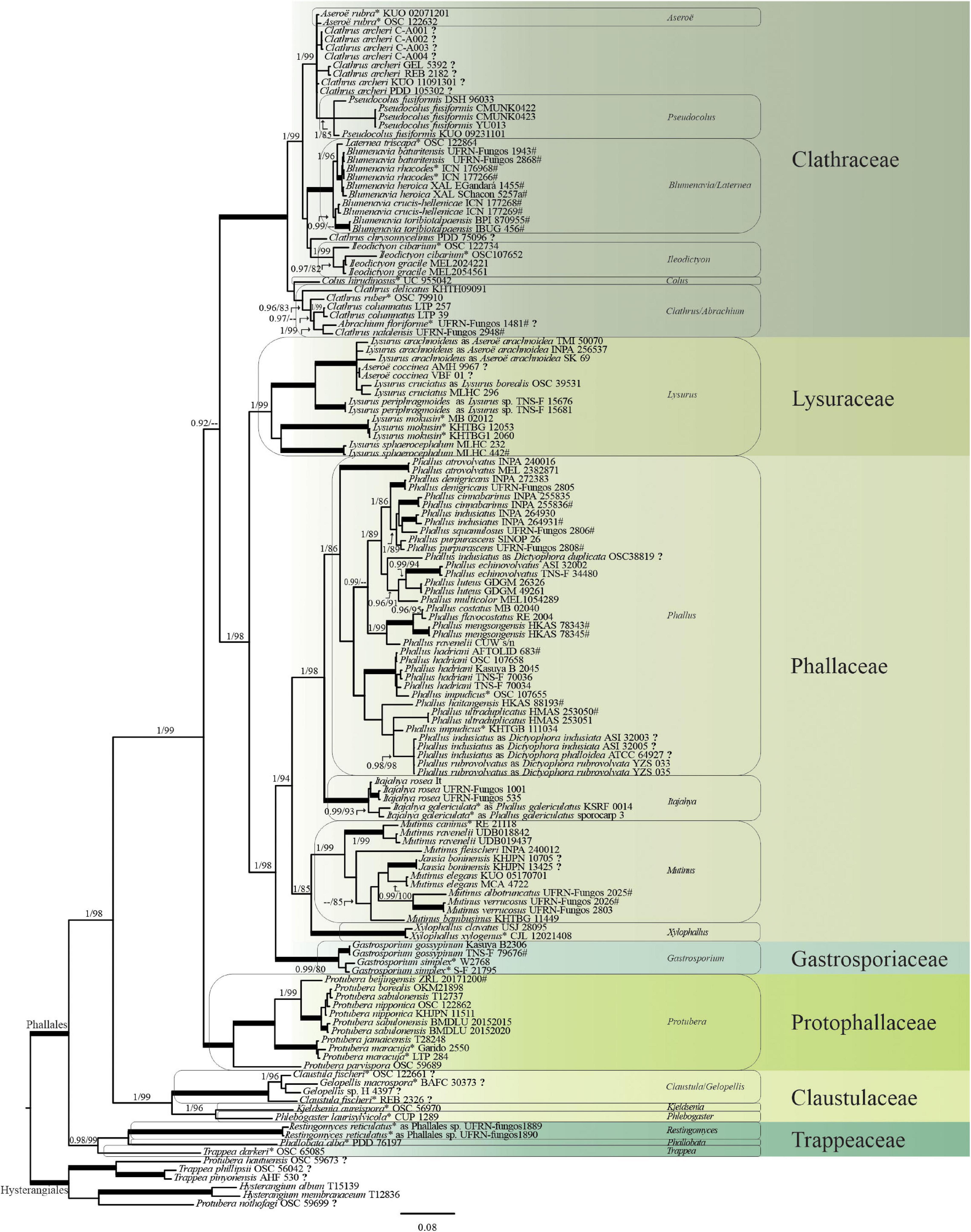
Figure 4. Phylogenetic tree of the Phallales based on a combined dataset (ITS/nuc-LSU/mt-SSU/ATP6/RPB2/TEF1-α). Family clades are colored in green shades and named on the right. Names of recognized genera are highlighted in the boxes. (*) Type species. (?) Sequences that need review of their identity or generic status. (#) Type specimens. Tree topology is based on the Bayesian analyses. Numbers on branches are posterior probabilities (PP, before slash) and maximum likelihood bootstrap values (MLbs, after slash). Thickened branches in boldface indicate fully supported nodes (PP = 1, MLbs = 100). Scale bar indicates expected changes per site.
Two ITS sequences of environmental samples retrieved from GenBank (EF218792 and MF487330) did not match any phalloid sequence in BLAST, and thus, they were eliminated from our data and not included in Supplementary Tables 1, 3. Thirteen ITS sequences (MK518965, UDB015101 (JQ657782), MT644888, UDB018620, UDB0673787, UDB0317538, UDB0321542, UDB0215586, UDB0196057, UDB089976, MT512648, UDB0180761, and MH930315) and five nuc-LSU (MK518662, MH532563, MH532564, MH532565, and MH532566) were excluded from the dataset because they had many ambiguous bases and long gaps, possibly the result of poorly edited sequences. These sequences were also checked on NCBI BLAST, and they do not correspond to any species of Phallales. Six sequences were divided into ITS and nuc-LSU and incorporated in both ITS and nuc-LSU alignments (see individuals with “YES” in the column “nuc-LSU sequence incorporated in phylogenetic analyses…” in Supplementary Table 1).
Among the sequences that need revision because of possible misidentification or doubtful positioning are those of the monospecific genus Calvarula, as Calvarula excavata Zeller (TEF1-α, DQ219293), which is positioned in Lysuraceae (Supplementary Figure 1A), although its classification was in Protophallaceae (Zeller, 1939). Sequences named Protubera sp. (T20068), Protubera hautuensis Castellano and Beever (OSC59673), Protubera nothofagi Castellano and Beever (OSC59699), Trappea phillipsii (Harkn.) Castellano (OSC56042), and Trappea pinyonensis States (AHF530) are grouped outside the Phallales core, in the outgroup of Hysterangiales. Thus, these sequences, as well as Hysterangium sequences, were used to root the trees as representative of Hysterangiales, and their identification must be further investigated. Sequences named Protubera sp. (vouchers FLAS-F60616 and FLAS-F 61859), Protubera canescens G.W. Beaton and Malajczuk, and Protubera clathroidea Dring also need to be investigated because they clustered in Clathraceae or Lysuraceae (Supplementary Figures 1–3), despite the classification of the genus in Protophallaceae (Hosaka et al., 2006; Trierveiler-Pereira et al., 2014a,b). Finally, some sequences of Gelopellis need further studies because they clustered out of the expected Claustulaceae (Hosaka et al., 2006; Trierveiler-Pereira et al., 2014a); Gelopellis purpurascens G.W. Beaton and Malajczuk (voucher H292) is grouped in Phallaceae (Supplementary Figure 3B); Gelopellis sp. (voucher MEL 2063389) is in Clathraceae (Supplementary Figures 2B, 3A) or external to Phallaceae (Supplementary Figure 3B).
For the total of 43 unidentified individuals not classified at genus level (Table 3), 39 were placed in five distinct families, two did not group with any family in Phallales (Figure 5 and Supplementary Table 6), and the last two (Hysterangiales sp. UDB015101, and Phallales sp. UDB018620), as mentioned previously, were excluded from analyses. A total of 13 individuals were assigned to a specific genus: Ileodictyon, Gastrosporium, Mutinus, and Phallus (Supplementary Table 6). Fifteen individuals were identified to species level: Blumenavia crucis-hellenicae G. Coelho, Sulzbacher, Grebenc and Cortez, Phallus impudicus L., Phallus hadriani Vent., Restingomyces reticulatus Sulzbacher, B.T. Goto and Baseia (Supplementary Table 6). The identification of the sequences of R. reticulatus was possible by consulting the original article that proposed the new taxon (Sulzbacher et al., 2016). Supplementary Figures 1A, 2A,B, 3B show the phylogenetic positioning and the possible identification of some of the 41 unidentified individuals retrieved according to TEF1-α, ITS, nuc-LSU, and ATP6, respectively.
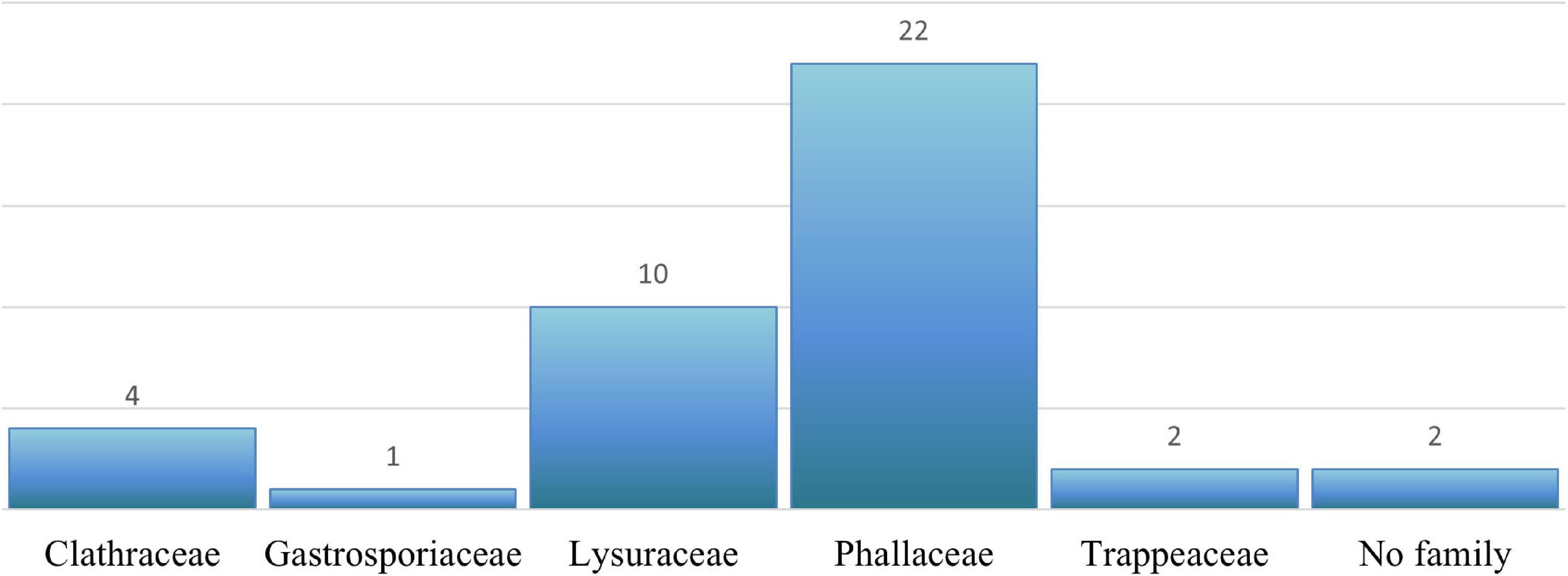
Figure 5. Number of undetermined Phallales individuals and their putative family level classification according to the phylogenetic position of the sequences recovered.
Based on the phylogenetic inferences using the concatenated data matrix (ITS + nuc-LSU + mt-SSU + ATP6 + RPB2 + TEF1-α), our results show Phallales as a strongly supported monophyletic order (PP = 1, MLbs = 100) and composed of seven families (Figure 4). We recognized 22 genera in Phallales, with Dictyophora and Jansia confirmed as synonyms of Phallus and Mutinus, respectively, although the recognition of some genera and the placement of their representatives deserve attention: Abrachium, Aseroë, Blumenavia, Clathrus, Claustula, Gelopellis, Laternea, Protubera, Pseudocolus, and Trappea.
The composition of each family based on our analyses is presented below.
This family grouped the genera Abrachium, Aseroë, Blumenavia, Clathrus, Ileodictyon, Laternea, and Pseudocolus (Figure 4). In the combined analyses, Aseroë in the Clathraceae clade is represented by the type species Aseroë rubra Labill. but is not recognized as monophyletic and forms a paraphyletic group with other sequences named Clathrus archeri (Berk.) Dring. These sequences warrant further examination. Pseudocolus, represented by P. fusiformis, is within the well-supported clade formed by A. rubra and C. archeri.
Laternea, based on the type species Laternea triscapa Turpin, forms a monophyletic clade together with sequences of Blumenavia, which includes the epitype specimen Blumenavia rhacodes Möller (voucher ICN 177266).
Ileodictyon is recognized as monophyletic (PP = 1, MLbs = 99), including seven sequences of two individuals of the type species Ileodictyon cibarium Tul. and C. Tul and another eight sequences of two individuals of I. gracile Berk. The specific identity of OSC107652, named as I. cibarium, must be investigated because it is positioned closer to sequences identified as I. gracile. Although with no support, Clathrus chrysomycelinus Möller is external to Ileodictyon, and the identity or generic status of the individual PDD75096 must be investigated.
Colus is represented by one nuc-LSU sequence of the type species C. hirudinosus Cavalier and Séchier, and it is closer to Clathrus, although this relationship is not supported.
Clathrus is recovered as polyphyletic with representatives clustered in at least three clades (Figure 4). Its core is represented by the type species Clathrus ruber P. Micheli ex Pers. plus Clathrus delicatus Berk. and Broome, Clathrus columnatus Bosc, Clathrus natalensis G.S. Medeiros, Melanda, T.S. Cabral, B.D.B Silva and Baseia, and Abrachium floriforme (Baseia and Calonge) Baseia and T.S. Cabral. Abrachium is represented here by only one collection (holotype of the type species). Sequences of C. archeri and C. chrysomycelinus are related to species of Aseroë and Ileodictyon, respectively; as mentioned previously, these should be further investigated for a possible reannotation or recombination.
In the family Lysuraceae, sequences named under Lysurus and Aseroë are included, although the latter has been formally classified in Clathraceae. Sequences are distributed in four well-supported clades (Figure 4). The most well-sampled clade includes sequences of Lysurus arachnoideus (E. Fischer) Trierv.-Per. and Hosaka (=Aseroë aracnoidea E. Fisch.), Aseroë coccinea Imazeki and Yoshimi, and Lysurus borealis (Burt) Henn. (=Lysurus cruciatus Henn.). Our results confirm the synonymizing of A. aracnoidea in Lysurus (Trierveiler-Pereira et al., 2014a), and based on the individuals retrieved, L. borealis is the confirmed synonym of L. cruciatus (Dring, 1980). The other three clades represent the following three taxa: Lysurus periphragmoides (Klotzsch) Dring from Japan (Caffot et al., 2018), the type species Lysurus mokusin (L.) Fr., and Lysurus sphaerocephalum (Schltdl.) Hern. Caff., Urcelay, Hosaka and L.S. Domínguez., with the latter considered an invalid name (Index Fungorum, 2020; MycoBank., 2020) according to Art. F.5.1 (Shenzhen), due to the absence of an identifier-issued citation in a recognized repository.
This family is composed of Itajahya, Phallus (=Dictyophora), Mutinus (=Jansia), and Xylophallus (Figure 4). The type species P. impudicus is represented by two individuals that are positioned in two different clades, although the collection OSC107655 is closer to other sequences of P. hadriani Vent. and most likely is misidentified as P. impudicus. Anyway, the identity and positioning of P. impudicus deserve further investigations. Sequences of Phallus indusiatus Vent. closer to Phallus rubrovolvatus (M. Zang, D.G. Ji and X.X. Liu) Kreisel also most likely represent misidentification.
Itajahya, recognized as monophyletic and sister to Phallus, is represented by sequences of Itajahya rosea (Delile) E. Fisch. and Itajahya galericulata Möller. Sister to Itajahya and Phallus is the clade formed by Mutinus and Xylophallus. Mutinus is represented by eight species, including the type species Mutinus caninus (Huds.) Fr. and Jansia boninensis Lloyd. Xylophallus is represented by Xylophallus clavatus T.S. Cabral, M.P. Martín, C.R. Clement, K. Hosaka and Baseia and the type species Xylophallus xylogenus (Mont.) E. Fisch.
Gastrosporiaceae is delimited in our analysis with high support (PP = 0.98, MLbs = 80) and sharing a common ancestor with Phallaceae, with both families (Gastrosporiaceae and Phallaceae) as sister of Lysuraceae (Figure 4). This monogeneric family is here represented by sequences of Gastrosporium gossypinum T. Kasuya, S. Hanawa and K. Hosaka, and the type species Gastrosporium simplex Mattir.
Protophallaceae is represented by the genus Protubera with the type species Protubera maracuja Möller plus Protubera beijingensis G.J. Li and R.L. Zhao, Protubera borealis S. Imai, Protubera jamaicensis (Murrill) Zeller, Protubera nipponica Kobayasi, Protubera sabulonensis Malloch, and Protubera parvispora Castellano and Beever (Figure 4). Protubera parvispora (OSC59689) was not placed in the Protophallaceae clade in the ATP6 tree (Supplementary Figure 3B), but it was in Protophallaceae in the other analyses (combined, nuc-LSU, RPB2, and TEF1-α).
In the Claustulaceae, clustered sequences were named Claustula, Gelopellis, Kjeldsenia, and Phlebogaster (Figure 4). Claustula and Gelopellis formed a monophyletic clade represented by four individuals including their type species: Claustula fischeri K.M. Curtis (type country New Zealand) and Gelopellis macrospora Zeller (type country Chile). Gelopellis macrospora is represented by a sample from Argentina, whereas C. fischeri is represented by two individuals from New Zealand, but these did not cluster together in our analyses, which puts into doubt the identification of one or more of these individuals as C. fischeri. The other clade in Claustulaceae is formed by Kjeldsenia and Phlebogaster represented by their type species: Kjeldsenia aureispora W. Colgan, Castellano and Bougher and Phlebogaster laurisylvicola Fogel, respectively. We retrieved two sequences (nuc-LSU and TEF1-α) of one individual of P. laurisylvicola (CUP 1289), which appears close to Hysterangium species in the nuc-LSU analyses (Supplementary Figure 2B) but is in Claustulaceae in the TEF1-α (Supplementary Figure 1A) and the concatenated analyses (Figure 4), which support our classification of Phlebogaster in Claustulaceae (Phallales).
This family is represented as monophyletic (PP = 0.98, MLbs = 99) in the combined analyses by Restingomyces, Phallobata, and Trappea with their respective type species R. reticulatus, Phallobata alba G. Cunn., and Trappea darkeri (Zeller) Castellano, respectively (Figure 4). However, the sequences of these genera were not recovered as a single-family clade in some of the unique marker analyses: in TEF1-α (Supplementary Figure 1A) and RPB2 (Supplementary Figure 3A), P. alba and T. darkeri are recovered separately; in ITS (Supplementary Figure 2A), R. reticulatus and T. darkeri do not form a clade (although the individuals of T. darkeri in the ITS analyses are not the same as all the other analyses); in nuc-LSU (Supplementary Figure 2B) T. darkeri is external (PP = 0.96) to all Phallales, whereas R. reticulatus and P. alba clustered together but with no support.
The clustering of the 479 sequences of ITS revealed 118 species hypotheses in Phallales, based on a sequence similarity threshold of 98.0%. In MycoBank. (2020), 576 legitimate specific names have already been deposited in Phallales. Our clustering shows that almost 20% of the total recognized species of Phallales have DNA sequences available, revealing that a lot of work remains to be done in this area.
Information on the country of origin included in DNA databases is available for 462 individuals (69.8%), from which 191 contain detailed information of the location but only 36 with the exact geographic coordinates. These individuals were distributed in 46 countries, which are concentrated in tropical and subtropical areas, with lower occurrence closer to the polar circles. Estonia is the source of the highest number of sequences (131) and individuals (131) deposited, all ITS. The United States is the second most sampled country, with 121 sequences of 79 individuals, and China is the third, with 108 sequences of 72 individuals. A map of global distribution of all phalloid individuals segregated by genera can be observed in Figure 6. For all 22 recognized genera, only Phallobata is not represented on the map, because there is no location information for the voucher. Individuals named under Jansia are represented as Mutinus, and those under Dictyophora as Phallus (Supplementary Table 4).
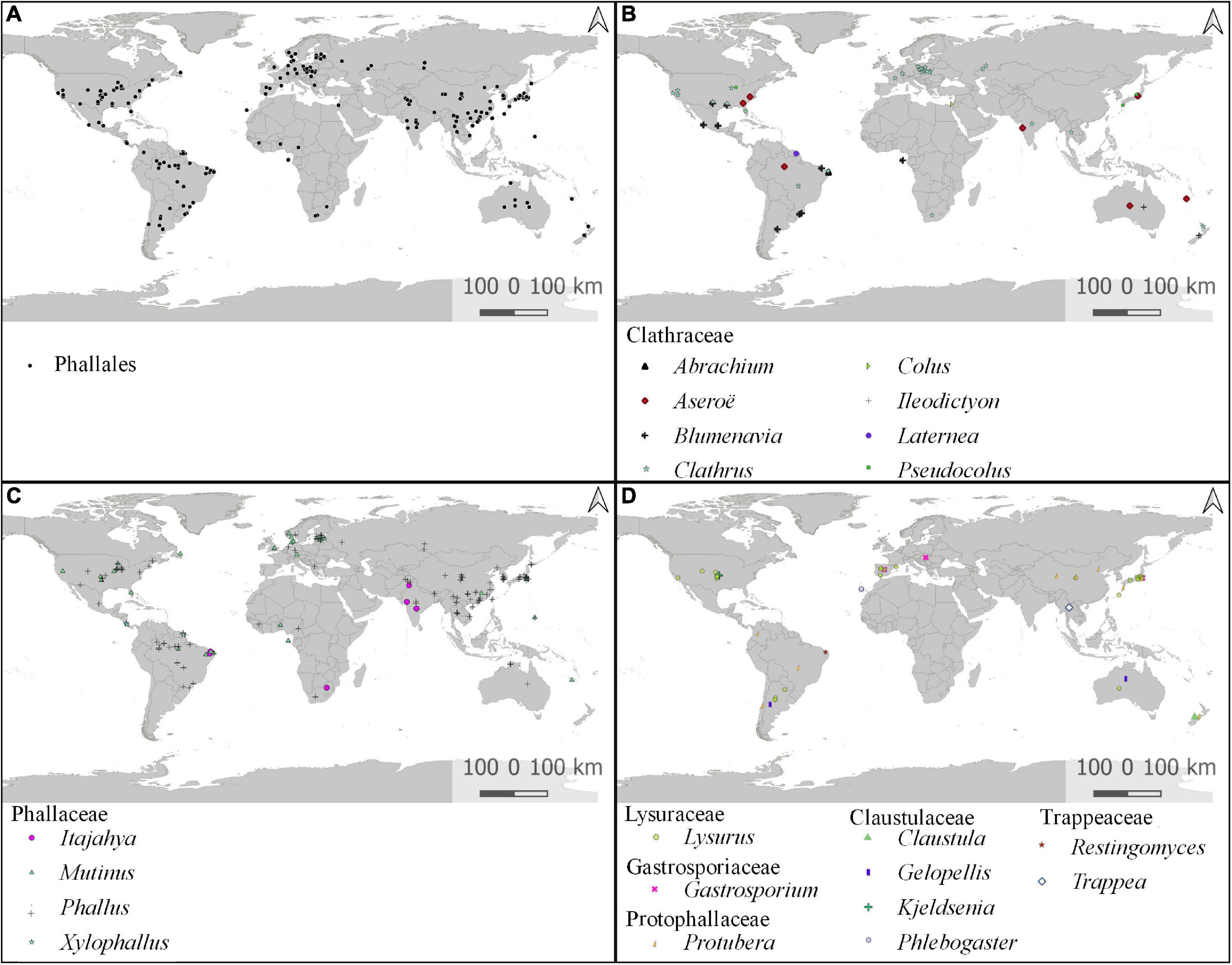
Figure 6. Global geographic distribution of Phallales based on samples with molecular sequences available and locality information. (A) Global distribution of all Phallales samples. Global distribution of (B) Clathraceae genera; (C) Phallaceae genera; (D) Lysuraceae, Gastrosporiaceae, Protophallaceae, Claustulaceae, and Trappeaceae genera.
A total of 168 individuals (25.3%) are from environmental samples, including 148 individuals from soil, eight from roots of Bouteloua gracilis (Willd. ex Kunth) Lag. ex Griffiths (blue grama), two from air filters, three from seedling stem [two from Hevea nitida Mart. ex Müll. Arg and one from Micrandra spruceana (Baill.) R. E. Schult.], one from heavy metal–contaminated soil, one from marine subsurface sediments, and one from house dust. According to the FungalTraits database (Põlme et al., 2021), a total of 21 phalloid genera recognized in our work are soil saprotrophic, and only Phlebogaster is mentioned as ectomycorrhizal.
Among the Phallales sequences sampled in our work and their respective published references, only Phallus dongsun T.H. Li, T. Li, Chun Y. Deng, W.Q. Deng and Zhu L. Yang is reported by Li et al. (2020) as an edible species commercially cultivated in China. To complement the list of edible species of Phallales with molecular data available, according to edibility categories proposed Li et al. (2021), our study includes 11 confirmed edible species (E1): Ileodictyon cibarium, Phallus echinovolvatus (M. Zang, D.R. Zheng and Z.X. Hu) Kreisel, Phallus fuscoechinovolvatus T.H. Li, B. Song and T. Li, P. hadriani, P. indusiatus, Phallus luteus (Liou and L. Hwang) T. Kasuya, Phallus merulinus (Berk.) Cooke (as Dictyophora merulina Berk.), P. rubrovolvatus, Phallus ultraduplicatus X.D. Yu, W. Lv, S.X. Lv, Xu H. Chen and Qin Wang, Protubera nipponica [as Kobayasia nipponica (Kobayasi) S. Imai and A. Kawam.], and Pseudocolus fusiformis; three confirmed edible species, but with conditions (E2): Clathrus archeri, Phallus impudicus, and Phallus multicolor (Berk. and Broome) Cooke (as Dictyophora multicolor Berk. and Broome); six unconfirmed edible species (E3): Clathrus columnatus, Ileodictyon gracile, Jansia boninensis, Lysurus mokusin, L. periphragmoides, Phallus rugulosus (E. Fisch.) Lloyd; and three poisonous species (P): Lysurus arachnoideus, Mutinus bambusinus (Zoll.) E. Fisch., and M. caninus.
Molecular data are available for 81.5% of the Phallales species mentioned by Li et al. (2021) as having a known edibility status. Only Mutinus borneensis Ces., Phallus armeniacus Pat., Phallus fragrans M. Zang, and Phallus tenuis (E. Fisch.) Kuntze lack public sequence data. Mutinus borneensis and P. armeniacus are categorized by the authors as E3 (unconfirmed edible species), P. fragrans as E1 (confirmed edible species), and P. tenuis as P (poisonous species).
The year 2006 can be marked as a huge advance in Phallales taxonomy with the work of Hosaka et al. (2006), a study aided by the Deep Hypha initiative (Blackwell et al., 2006). Hosaka et al. (2006) were responsible for 68 phalloid sequences deposited in GenBank (Figure 3), as well as the introduction of new markers (ATP6, RPB2, and TEF1-α). Another important turning point observed in Figure 3 is the increased use of molecular data in many works focused on Phallales diversity since 2013: nuc-LSU sequences – Degreef et al. (2013); ITS sequences – Moreno et al. (2013), Lu et al. (2014), Cabral et al. (2015), Kim et al. (2015), Elwess and Latourelle (2016), Patel et al. (2018), and Bobade and Dahanukar (2020); nuc-LSU and ATP6 sequences – Marincowitz et al. (2015); ITS and nuc-LSU sequences – Pietras et al. (2016); ITS, nuc-LSU, RPB1 and ATP6 sequences – Garnica et al. (2016). Trierveiler-Pereira et al. (2014a) also merit a spotlight because of their phylogenetic review of Phallales using nuc-LSU, ATP6, and RPB2 markers and by generating 53 sequences.
Clathraceae is characterized by fungi with clathrate (branched) or pseudostipitate and clathrate basidiomata that are named cage or lattice stinkhorns (Pegler and Gomez, 1994; Melanda et al., 2020). Individuals with clathrate format belong to Blumenavia, Clathrus, Ileodictyon, and Laternea, whereas those with prominent pseudostipe and a clathrate part composed of arms, armless, or lattice belong to Abrachium, Aseroë, Colus, and Pseudocolus. Cabral et al. (2012) accepted in Clathraceae the genera Abrachium, Aseroë, Blumenavia, Clathrus, Pseudocolus, Lysurus, and Ileodictyon. Our phylogenetic analyses, in agreement with Hosaka et al. (2006) and Trierveiler-Pereira et al. (2014a), show Lysurus as part of Lysuraceae. Based on morphological taxonomy using as the main diagnostic feature the disposition of the gleba in the receptacle, Pegler and Gomez (1994) classified the following genera in the Clathraceae series Lysuroid: Aseroë, Colus, Lysurus, Kalchbrennera Berk., Neolysurus O.K. Mill., Pseudocolus, and Simblum Klotzsch ex Hook. According to Pegler and Gomez (1994), these genera share the receptacle composed of a tubular and sterile pseudostipe, with the gleba attached to the upper portion of the receptacle. For Dring (1980), Kalchbrennera and Simblum were considered synonyms of Lysurus. However, our phylogeny (Figure 4) does not confirm this morphological approach as a natural character, with representatives of Clathraceae series Lysuroid sensu Pegler and Gomez (1994) in both Clathraceae (Aseroë and Colus) and Lysuraceae (Lysurus).
The relationship of the type species of Aseroë, A. rubra from Australia (type country), and C. archeri (Figure 4) has been previously shown by Hosaka et al. (2006), Degreef et al. (2013), and Trierveiler-Pereira et al. (2014a). The name C. archeri is a result of the combination of Lysurus archeri Berk. (Dring, 1980), which also has Anthurus archeri (Berk.) E. Fisch. as a synonym. The morphological research into Clathraceae performed by Dring (1980) considered Anthurus as a synonym of Clathrus. No other sequences of any other name previously treated as Anthurus are available. Considering that the type species of Clathrus is found in another clade, another generic name must be investigated for C. archeri, with Anthurus and Aseroë being two putative names, considering this taxonomic history and the relationship shown in our analyses.
The positions of Laternea and Blumenavia indicate that they may belong to the same genus, in which Laternea would have nomenclatural priority. Blumenavia and Laternea are also grouped in a monophyletic clade, as previously observed by Degreef et al. (2013) and Trierveiler-Pereira et al. (2014a), although both of these works performed their phylogenetic analyses with only one individual of each genus. Melanda et al. (2020) reviewed Blumenavia but did not present any Laternea species in the phylogeny. Considering that the molecular global sampling of Laternea is represented by only one representative, new molecular studies involving Laternea are encouraged, so as to better understand the relationship between Laternea and Blumenavia.
The genus Colus was considered as a member of the family Clathraceae, based on a morphological approach by Cunningham (1944), Dring (1980), and Pegler and Gomez (1994); here, we confirm this classification and also encourage the generation of more sequences of this genus.
Clathrus was shown to be polyphyletic by Hosaka et al. (2006) and Trierveiler-Pereira et al. (2014a). The relationship between Abrachium and Clathrus was observed by Trierveiler-Pereira et al. (2014a), who showed A. floriforme in an unsupported clade with C. ruber (type species) and C. columnatus. Abrachium is a monospecific genus proposed by Cabral et al. (2012), with the type species A. floriforme being a combination from Aseroë floriformis Baseia and Calonge. Cabral et al. (2012) emended the family Clathraceae to include the armless sunflower-shaped receptacle characteristic of Abrachium. Based on the high morphological variation between Abrachium and Clathrus, we have to wait for more sequences of abrachinoid individuals to infer any synonymy among the two genera, with Clathrus having nomenclatural priority. Abrachium appears to be widely distributed in the Atlantic Forest and Caatinga Biomes5, and additional studies may show unknown diversity.
Linderia G. Cunn, Linderiella G. Cunn., Ligiella J.A. Sáenz, and other possible representatives of Clathraceae do not have any sequences available and need to be included in further molecular phylogenetic studies. MycoBank. (2020) and Index Fungorum (2020) consider Linderiella and Linderia as synonyms of Clathrus, whereas He et al. (2019) consider only Linderiella as a synonym of Clathrus, but the inclusion of sequences of these genera in phylogenetic studies can confirm their relationships.
The positioning of the type species of Lysurus (L. mokusin, type country: China) from the United States and Japan has been previously shown by Trierveiler-Pereira et al. (2014a). Simblum, Kalchbrennera, and Neolysurus, other possible representatives of Lysuraceae, do not have any sequences available and need to be included in further studies. Simblum and Kalchbrennera are considered synonyms of Lysurus by Dring (1980), He et al. (2019), and in Index Fungorum (2020), whereas MycoBank. (2020) considers only Kalchbrennera to be a synonym of Lysurus, but the inclusion of sequences of these genera in phylogenetic studies can clarify their relationships. Lysurus periphragmoides and L. sphaerocephalum (nom. inval.) are names that have been previously placed in Simblum, and the phylogenetic placement of sequences under these names in our analyses could confirm the synonymizing between Lysurus and Simblum. Simblum periphragmoides Klotzsch and S. sphaerocephalum Schltdl. were considered as heterotypic synonyms by Dring (1980), who proposed the combination L. periphragmoides, but this synonymizing was not accepted by Caffot et al. (2018), and based on our analyses (Figure 4) it is also not confirmed. Considering the positioning of sequences named A. coccinea in the Lysuraceae, we encourage further studies to investigate the generic status of this taxon and the identity of the individuals under this name.
Li et al. (2014, 2016) provided phylogenetic data to show that Dictyophora and Phallus should be treated as a single genus, Phallus. Cabral et al. (2019) described a high diversity in Phallus indusiatus, a species complex well-known for the presence of an indusium; however, P. indusiatus actually represents at least four phylogenetic species. Li et al. (2020) published the new species P. dongsun, which is not included in our concatenated tree, and mentioned that samples named P. impudicus from China represent P. dongsun, as P. impudicus is a species described from Europe (the type locality of P. impudicus). As previously mentioned, the identity of the sequences named P. impudicus recovered in our analyses needs further examination in order to obtain a robust view of the phylogenetic position of the species.
Kreisel (1996) accepted the genus Itajahya as Phallus subgenus Itajahya. In the same work, the type species of Itajahya, I. galericulata, originally described from South Brazil (Möller, 1895), was combined in Phallus galericulatus (Möller) Kreisel (Kreisel, 1996). However, based on our analyses (Figure 4), I. galericulata is clustered in a clade separate from Phallus and together with other Itajahya species, which does not justify the classification and the synonymizing proposed by Kreisel (1996). For Mutinus and Jansia, however, they are considered synonymous as previously pointed out by Crous et al. (2017). The positioning of Xylophallus as sister to Mutinus has been previously shown by Trierveiler-Pereira et al. (2014a) and Crous et al. (2018).
There are no molecular data for Aporophallus Möller, Floccomutinus Henn., and Staheliomyces E. Fisch., all genera also accepted in Phallaceae (Index Fungorum, 2020; MycoBank., 2020), although Floccomutinus is considered a synonym of Mutinus by He et al. (2019) and in MycoBank. (2020). Aporophallus and Staheliomyces are monospecific genera, and their revision will be highly relevant. Sequences of taxa dealt under Floccomutinus could be useful to better assess their relationships with Mutinus.
The monogeneric family Gastrosporiaceae was confirmed as part of Phallales in our study and also by previous authors: Kirk et al. (2008), Trierveiler-Pereira et al. (2014a), He et al. (2019), Kasuya et al. (2020), and Wijayawardene et al. (2020). The genus Gastrosporium contains three described species: G. simplex (type species) from Italy, G. asiaticum Dörfelt and Bumžaa from Mongolia, and G. gossypinum from Japan (Honshu). The positioning of the family Gastrosporiaceae external to Phallaceae in our phylogeny (Figure 4) rejects the suggestion by Hosaka et al. (2006), who did not include Gastrosporium in their analyses and thus suggested that the ancestor of Clathraceae, Phallaceae, and Lysuraceae could be the point of transition from sequestrated to expanded basidiomata in Phallales.
Protophallaceae is represented in our analyses only by Protubera species, as also shown by Trierveiler-Pereira et al. (2014a, b). Protophallaceae was proposed by Zeller (1939) with species of Calvarula, Protophallus Murril, and Protubera. Calvarula is not included in our combined analyses (Figure 4), and the identity of the individual EF5-1 as C. excavata needs to be investigated. Additional individuals and sequences of Calvarula are needed to clarify its family level positioning. Protophallus is considered a synonym of Protubera (Malloch, 1989; He et al., 2019), and the positioning of Protubera jamaicensis (=Protophallus jamaicensis Murrill) in our analyses (Figure 4) confirms this synonymization.
Sanshi and Kawamura (1958) analyzed specimens of Protubera from Japan and proposed a new genus named Kobayasia S. Imai and A. Kawam. to accommodate Protubera nipponica. However, Hosaka et al. (2006) and Trierveiler-Pereira et al. (2014a, b) have shown sequences of P. nipponica within the clade formed by other Protubera, as also shown in our analyses (Figure 4), which does not justify Kobayasia as a separate genus and confirms its synonymization under Protubera. Kobayasia kunmingica M. Zang, K. Tao and X.X. Liu is the only other species described in Kobayasia, and its phylogenetic positioning should be investigated to better understand the relationship between Protubera and Kobayasia.
The relationship of Protubera borealis (voucher OKM21898) and P. sabulonensis has been shown by Hosaka et al. (2006) and Cabral et al. (2012). Sanshi and Kawamura (1958) considered Protubera borealis under the genus Protuberella S. Imai and A. Kawam. as Protuberella borealis (S. Imai) S. Imai and A. Kawam. However, the recognition of Protuberella as a separate genus was not accepted by Hosaka et al. (2006), Cabral et al. (2012), and Li et al. (2018), and it is also not confirmed here (Figure 4), although it is still considered as a distinct genus by He et al. (2019) and in Index Fungorum (2020).
Besides the sequences of Protubera clustered in Phallales, some individuals (T20068, OSC59673, and OSC59699) named under Protubera (including P. nothofagi and P. hautuensis) clustered in Hysterangiales (our outgroup), in agreement with Hosaka et al. (2006) and Trierveiler-Pereira et al. (2014b), who classified P. nothofagi in Gallaceae and P. hautuensis in Phallogastraceae. However, based on the positioning of the type species P. maracuja in Phallales, this must be the best classification for Protubera and Protophallaceae. The positioning of the individuals named Protubera canescens and P. clathroidea in Clathraceae and Lysuraceae, respectively (Supplementary Figures 1–3), has been shown also by Hosaka et al. (2006), Degreef et al. (2013), and Trierveiler-Pereira et al. (2014b). According to May et al. (2010), P. canescens is an egg (immature) form of Ileodictyon, and for Trierveiler-Pereira et al. (2014b), these Protubera species out of Protophallaceae may be egg forms of expanded phalloid taxa or misidentification.
In this family clade, the four genera Claustula, Gelopellis, Kjeldsenia, and Phlebogaster clustered together as in Hosaka et al. (2006). In Trierveiler-Pereira et al. (2014a), the family clade was represented only by Gelopellis and Claustula. Although Claustula and Gelopellis clustered together, it would be important to have sequences of nondoubtful identification and more representatives of both genera to confirm whether they could be considered synonyms, of which Claustula has nomenclatural priority. Individuals of Gelopellis clustered out of Claustulaceae, in Clathraceae (Supplementary Figures 2B, 3A), or external to Phallaceae (Supplementary Figure 3B), and they also need revision to confirm their identity or generic status, mainly because immature basidiomata can lead to misidentification of some expanded phalloids, as seen in individuals of Protubera.
The composition of Trappeaceae in our study by the three genera Restingomyces, Phallobata, and Trappea is in agreement with Sulzbacher et al. (2016). Trappea is the type genus of the family as established by Castellano (1990), in a study that mentioned that the genus represents a transition between Clathraceae and Hysterangium, and proposed the type species of Trappea based on Hysterangium darkeri Zeller. The positioning of Trappea in Phallales as a member of Trappeaceae is based on an individual of T. darkeri without information on the location, as also previously shown by Hosaka et al. (2006) and Sulzbacher et al. (2016). As in Protubera, some species of Trappea are also placed in Hysterangiales, but considering the positioning of its type species in Phallales, we confirm the classification of Trappea in Phallales. The positioning of some Trappea in Hysterangiales has also been observed by Hosaka et al. (2006) and Trierveiler-Pereira et al. (2014b), and both authors showed T. phillipsii and T. pinyonensis positioned in the Phallogastraceae (Hysterangiales) family. The identity or the generic status of these individuals of Trappea in Hysterangiales needs further revision.
The DNA databases contain many sequences of Phallales representatives without precise information on the collection location or even without any metadata. In addition, the sequences have a low representation across continents, which makes it impossible to draw any robust inferences about the biogeographical distribution of families, genera, species, and clades. Thus, we encourage the inclusion of DNA sequences from representatives of Phallales from underexplored locations, such as Africa and maritime Southeast Asia, as well as the sequencing of collections already deposited in herbaria around the world and of taxa not yet sequenced. These further data will contribute to a better understanding of the evolutionary processes and distribution patterns in Phallales.
The explanation for the high number of sequences from Estonia is the increase of studies of ITS using environmental samples and the high number of these sequences deposited in UNITE (see Supplementary Table 1). Other locations with a large number of Phallales records and DNA sequences are partly due to the proximity of the research centers and specialists in the group.
Studies of environmental biodiversity or other ecological approaches including phalloid species started in 2003 (Bidartondo et al., 2003) and has continued in a few works (Sato et al., 2012; Kellner et al., 2014; Skaltsas et al., 2019; Vu et al., 2019).
In the FungalTraits database (Põlme et al., 2021), all their genera in Phallales are classified as saprotrophic. The genera Phallobata, Phlebogaster, Restingomyces, and Trappea are classified as part of Hysterangiales, but we recognize them in Phallales. Phlebogaster is considered an ectomycorrhizal genus in FungalTraits. Thus, considering Phlebogaster in Phallales, as supported by our analyses, the Phallales lifestyle must be expanded to include both saprotrophic and ectomycorrhizal genera. The ectomycorrhizal habit of Phlebogaster is most likely based on Fogel (1980), who described P. laurisylvicola as a hypogeous taxon under the plant species Laurus azorica (Seub.) Franco (Lauraceae Lindley). Kreisel (2001), in a checklist of gasteroid fungi, also mentioned P. laurisylvicola in association with L. azorica. However, we did not find any confirmation and description of these associations as an ectomycorrhizal symbiosis. We regard this species as putatively ectomycorrhizal, but further investigation is needed.
Hosaka et al. (2006) cited Protubera canescens as the only Phallales ectomycorrhizal, but according to May et al. (2010), this species is an immature form of Ileodictyon, as mentioned previously. In addition, Trierveiler-Pereira et al. (2014b) affirmed that all Protubera species are saprotrophic as reported in FungalTraits (Põlme et al., 2021). Therefore, we suggest further investigation to clarify the ectomycorrhizal association in this phalloid species.
Wild edible fungi are an important renewable natural resource in some regions, constituting important sources of income and nutrition. Despite their unpleasant odor, many representatives of Phallales are edible but are considered mushrooms with little culinary value. Stinkhorns are the most popular edible Phallales (Boa, 1988). The stinkhorn Phallus is the genus with the largest number of species with edible status (14: nine E1, two E2, two E3, and one P), followed by Lysurus (three E3 and one P) and Mutinus (one E3 and two P) that include only poisonous and unconfirmed edible species (Li et al., 2021). Phallus was reported by Li et al. (2021) as a genus with 43 described species, whereas He et al. (2019) reported 34 known species. Based on these numbers, the percentage of confirmed edible species in the genus represents 20.9 to 26.5%; however, the number of known species is probably underestimated because MycoBank. (2020) lists approximately 125 legitimate specific names for the genus. For Phallales, considering the 576 legitimate specific names recognized in MycoBank. (2020), the number of confirmed edible species in the order represents only 2.1%, whereas the number of poisonous species represents only 0.7%. The proportion of the number of confirmed edible and poisonous species in Phallales in relation to the 118 SHs that we found is 10.2 and 3.4%, respectively. This confirms that the edibility of Phallales species is poorly explored.
Many stinkhorns are consumed in the egg stage because of their tasty flavor (Phillips et al., 2018). In Germany and North America, the egg stage of Phallus is sold canned or fresh (Læssøe and Spooner, 1994). In China, L. mokusin and P. rubrovolvatus (as Dictyophora rubrovolvata M. Zang, D.G. Ji and X.X. Liu) are considered edible (Læssøe and Spooner, 1994). In addition, P. indusiatus and P. dongsun stand out in China for their flavor, where they are commercially cultivated (Boa, 1988; Li et al., 2020) and represent an important economic product. Despite the nutritional and commercial importance, the consumption of wild stinkhorns is not recommended unless their taxonomic affiliation is known with certainty because some of them are poisonous, such as species of Lysurus and Mutinus. Additionally, some species of the same genus are confirmed edible, whereas others are controversial, such as P. tenuis, which is considered poisonous (Li et al., 2021).
This work presents a summary of studies using molecular tools in Phallales. In general, as in other groups of fungi, these tools clarify results of previous studies based on morphology; also, the use of combined markers has allowed a clearer delimitation and positioning of the families and genera. Although we recognized seven families and 22 genera in Phallales, an extra effort is needed in taxonomic studies of the genera Abrachium, Aseroë, Blumenavia, Clathrus, Claustula, Gelopellis, Laternea, Protubera, Pseudocolus, and Trappea, because some inconsistencies in species identification and positioning of their representatives should be clarified. It is also necessary to include sequences of the genera Aporophallus (Phallaceae), Floccomutinus (Phallaceae), Kalchbrennera (Lysuraceae), Ligiella (Clathraceae), Linderia (Clathraceae), Linderiella (Clathraceae), Neolysurus (Lysuraceae), and Staheliomyces (Phallaceae). The DNA databases are an excellent source of molecular data, both when searching for sequences and in helping to understand the evolution of traits shown in previous studies, but missing metadata can be a stalemate for that. Therefore, it is vital to fill in the records in DNA databases correctly and accurately.
Since the inclusion of molecular tools for systematics studies, considerable progress has been achieved in the taxonomic and evolutionary study of fungi. Sequences of different markers deposited in databases represent a valuable repository that is still poorly exploited. The data we utilized have been shown to be a good, no-cost tool to clarify taxonomic and systematics problems, test phylogenetic position of misidentified sequences, examine the geographic distribution of groups, explore the ecology and use of phalloid species, and to visualize where our knowledge gaps are. We therefore encourage all mycologists to conduct extensive reviews of molecular data available for their particular fungal taxa of expertise.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
GM and NM designed the project. ALe, GM, and NM extracted the data. GM organized the metadata. ALi, AS-F, ALe, GM, NA, and RF analyzed the data. ALi and GM performed quantitative analyses. AS-F performed BLAST and phylogenetic analyses. ALe performed species hypothesis recognition. GM, MM, AS-F, ALi, ALe, TC, NM, and RF wrote the draft. IB, MM, and NM supervised the project. All authors reviewed and approved the manuscript.
This work was supported by ‘Coordenação de Aperfeiçoamento de Pessoal de Nível Superior’ (CAPES) – Finance code 001 (Process 23077.056677/2021-47), and for AS-F and NA’s scholarships; ‘Conselho Nacional de Desenvolvimento Científico e Tecnológico’ (CNPq) for GM’s scholarship (Process 140541/2018-7); ‘Fundação de Amparo a Ciência e Tecnologia do Estado de Pernambuco’ (FACEPE) for ALi’s scholarship; Special Visiting Researcher – PVE/CNPq (Process 407474/2013–7) given to IB and MM; and ‘Fundação de Amparo à Pesquisa do Estado de São Paulo’ (FAPESP) Grant #2018/15677-0 given to NM.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All authors thank the funding agency CAPES for paying the fees of publication. AS-F and NA thank the funding agency CAPES; GM thanks CNPq; and ALi thanks FACEPE, for the scholarships. IB and MM thank to PVE/CNPq; and NM thanks to FAPESP, for the researchers grants. The authors are grateful to Paul Kirk for giving data from Index Fungorum of Phallales in Excel format; and to Clark L. Ovrebo, Manuel Tabarés, and Marcelo A. Sulzbacher for some photographs of basidiomata included in Figure 1. Thanks to Susan Catherine Casement Moreira and Marian Glenn for English revision.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.689374/full#supplementary-material
Supplementary Figure 1 | Phylogenetic trees of the Phallales order obtained with TEF1-α (A) and mt-SSU (B). Names in blue and question marks indicate individuals with uncertain position or that represent a misidentification. Individuals retrieved without a genus name are represented in red with the possible species name based on our analyses given first. Family clades are colored in green shades and named on the right. Tree topology is based on the Bayesian analyses. Numbers on branches are posterior probabilities (PP, before slash) and maximum likelihood bootstrap values (MLbs, after slash). Thickened branches in boldface indicate fully supported nodes (PP = 1, MLbs = 100). Scale bar indicates expected changes per site.
Supplementary Figure 2 | Phylogenetic trees of the Phallales order obtained with ITS (A) and nuc-LSU (B). Names in blue and question marks indicate individuals with uncertain position or that represent a misidentification. Individuals retrieved without a genus name are represented in red with the possible species name based on our analyses given first. Family clades are colored in green shades and named on the right. Tree topology is based on the Bayesian analyses. Numbers on branches are posterior probabilities (PP, before slash) and maximum likelihood bootstrap values (MLbs, after slash). Thickened branches in boldface indicate fully supported nodes (PP = 1, MLbs = 100). Scale bar indicates expected changes per site.
Supplementary Figure 3 | Phylogenetic tree of the Phallales order obtained with RPB2 (A) and ATP6 (B). Names in blue and question marks indicate individuals with uncertain position or that represent a misidentification. Individuals retrieved without a genus name are represented in red with the possible species name based on our analyses given first. Family clades are colored in green shades and named on the right. Tree topology is based on the Bayesian analyses. Numbers on branches are posterior probabilities (PP, before slash) and maximum likelihood bootstrap values (MLbs, after slash). Thickened branches in boldface indicate fully supported nodes (PP = 1, MLbs = 100). Scale bar indicates expected changes per site.
Supplementary Table 1 | General data compilation of all Phallales sequences recovered.
Supplementary Table 2 | GenBank accession numbers for the outgroup used in phylogenetic analyses for individual markers.
Supplementary Table 3 | Taxa, individuals, and accession numbers of sequences used in phylogenetic analyses from the combined dataset.
Supplementary Table 4 | Taxa, individuals, and their respective geographic location used for the global distribution map.
Supplementary Table 5 | Type collections and collections of reference (REF.) in Phallales with the respective GenBank accession numbers. When the status of the type is unknown, only “type” is indicated.
Supplementary Table 6 | Putative identification for the undetermined Phallales individuals recovered.
Bidartondo, M. I., Bruns, T. D., Weiß, M., Sérgio, C., and Read, D. J. (2003). Specialized cheating of the ectomycorrhizal symbiosis by an epiparasitic liverwort. Proc. R. Soc. B Biol. Sci. 270, 835–842. doi: 10.1098/rspb.2002.2299
Binder, M., and Hibbett, D. S. (2002). Higher-level phylogenetic relationships of homobasidiomycetes (mushroom-forming fungi) inferred from four rDNA regions. Mol. Phylogenet. Evol. 22, 76–90. doi: 10.1006/mpev.2001.1043
Blackwell, M., Hibbett, D. S., Taylor, J. W., and Spatafora, J. W. (2006). Research coordination networks: a phylogeny for kingdom Fungi (Deep Hypha). Mycologia 98, 829–837. doi: 10.1080/15572536.2006.11832613
Bobade, V., and Dahanukar, N. (2020). Molecular characterization of stinkhorn fungus Aseroë coccinea Imazeki et Yoshimi ex Kasuya 2007 (Basidiomycota: Agaricomycetes: Phallales) from India. J. Threat. Taxa 12, 15530–15534. doi: 10.11609/jott.5091.12.4.15530-15534
Cabral, T. S., Clement, C. R., and Baseia, I. G. (2015). Amazonian phalloids: New records for Brazil and South America. Mycotaxon 130, 315–320. doi: 10.5248/130.315
Cabral, T. S., Marinho, P., Goto, T. B., and Baseia, I. G. (2012). Abrachium, a new genus in the Clathraceae, and Itajahya reassessed. Mycotaxon 119, 419–429. doi: 10.5248/119.419
Cabral, T. S., Silva, B. D. B., Martín, M. P., Clement, C. R., Hosaka, K., and Baseia, I. G. (2019). Behind the veil - exploring the diversity in Phallus indusiatus s.l. (Phallomycetidae, Basidiomycota). MycoKeys 58, 103–127. doi: 10.3897/mycokeys.58.35324
Caffot, M. L. H., Hosaka, K., Domínguez, L. S., and Urcelay, C. (2018). Molecular and morphological data validate the new combination of Lysurus sphaerocephalum from Argentina, with some additional records on Phallales (Agaricomycetes). Mycologia 110, 419–433. doi: 10.1080/00275514.2018.1456834
Castellano, M. A. (1990). The new genus Trappea (Basidiomycotina, Hysterangiaceae), a segregate from Hysterangium. Mycotaxon 38, 1–9.
Crous, P. W., Wingfield, M. J., Burgess, T. I., Carnegie, A. J., Hardy, G. E. S. J., Smith, D., et al. (2017). Fungal Planet description sheets: 625–715. Persoonia 39, 270–467. doi: 10.3767/persoonia.2017.39.11
Crous, P. W., Wingfield, M. J., Burgess, T. I., Hardy, G. E. S. J., Gené, J., Guarro, J., et al. (2018). Fungal Planet description sheets: 716–784. Persoonia 40, 240–393. doi: 10.3767/persoonia.2018.40.10
Cunningham, G. H. (1931). The Gasteromycetes of Australasia. XI. The Phallales. Part II. Proc. Linn. Soc. New South Wales 56, 182–200.
Cunningham, G. H. (1944). The Gasteromycetes of Australia and New Zealand. New Zealand: Dunedin, J. Mclndoe, 236.
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and high-performance computing Europe PMC Funders Group. Nat. Methods 9, 772. doi: 10.1038/nmeth.2109
Degreef, J., Amalfi, M., Decock, C., and Demoulin, V. (2013). Two rare Phallales recorded from São Tomé. Cryptogam. Mycol. 34, 3–13. doi: 10.7872/crym.v34.iss1.2013.3
Dring, D. M. (1980). Contributions towards a rational arrangement of the Clathraceae. Kew Bull. 35, 1–96. doi: 10.2307/4117008
Elwess, N. L., and Latourelle, S. M. (2016). DNA Barcoding and phylogenetic characterization of mushrooms with in the adirondack park. J. Agric. Res. 2, 28–43.
Fischer, E. (1898). “Phallineae,” in Die natürlichen Pflanzenfamilien, Teil. Abteilung. Leipzig, eds A. Engler and K. Prantl 276–296.
Fogel, R. (1980). Additions to the hypogeous mycoflora of the Canary Islands and Madeira. Contrib. Univ. Michigan Herb. 14, 75–82.
Garnica, S., Riess, K., Schön, M. E., Oberwinkler, F., and Setaro, S. D. (2016). Divergence times and phylogenetic patterns of Sebacinales, a highly diverse and widespread fungal lineage. PLoS One 11:0149531. doi: 10.1371/journal.pone.0149531
Gouy, M., Guindon, S., and Gascuel, O. (2010). SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 27, 221–224. doi: 10.1093/molbev/msp259
He, M. Q., Zhao, R. L., Hyde, K. D., Begerow, D., Kemler, M., Yurkov, A., et al. (2019). Notes, outline and divergence times of Basidiomycota. Fungal Divers. 99, 105–367. doi: 10.1007/s13225-019-00435-4
Hibbett, D. S., and Binder, M. (2002). Evolution of complex fruiting-body morphologies in homobasidiomycetes. Proc. R. Soc. B 269, 1963–1969. doi: 10.1098/rspb.2002.2123
Hibbett, D. S., Pine, E. M., Langer, E., Langer, G., and Donoghue, M. J. (1997). Evolution of gilled mushrooms and puffballs inferrred from ribosomal DNA sequences. Proc. Natl. Acad. Sci. 94, 12002–12006. doi: 10.1073/pnas.94.22.12002
Hibbett, D. S., and Thorn, R. G. (2001). “Basidiomycota: Homobasidiomycetes,” in The Mycota VII Part B: Systematics and Evolution, eds D. J. McLaughlin, E. G. McLaughlin, and P. A. Lemke (Berlin Heidelberg: Springer-Verlag), 121–168. doi: 10.1007/978-3-662-10189-6_5
Hosaka, K., Bates, S. T., Beever, R. E., Castellano, M. A., Colgan, W., Domínguez, L. S., et al. (2006). Molecular phylogenetics of the gomphoid-phalloid fungi with an establishment of the new subclass Phallomycetidae and two new orders. Mycologia 98, 949–959. doi: 10.3852/mycologia.98.6.949
Huang, Y., Niu, B., Gao, Y., Fu, L., and Li, W. (2010). CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 26, 680–682. doi: 10.1093/bioinformatics/btq003
Index Fungorum (2020). Excel version of the list of Phallales present in Index Fungorum send by e-mail by Paul Kirk. Available Online at: http://www.indexfungorum.org/ (accessed August 14, 2020).
Kasuya, T., Hanawa, S., and Hosaka, K. (2020). A new species of Gastrosporium (Phallales) from coastal sand dunes of Ibaraki Prefecture, central Japan. Truffology 3, 9–16.
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kellner, H., Luis, P., Pecyna, M. J., Barbi, F., Kapturska, D., Krug̈er, D., et al. (2014). Widespread occurrence of expressed fungal secretory peroxidases in forest soils. PLoS One 9:e95557. doi: 10.1371/journal.pone.0095557
Kim, C. S., Jo, J. W., Kwag, Y. N., Sung, G. H., gee Lee, S., Kim, S. Y., et al. (2015). Mushroom flora of ulleung-gun and a newly recorded Bovista species in the Republic of Korea. Mycobiology 43, 239–257. doi: 10.5941/MYCO.2015.43.3.239
Kirk, P. M., Cannon, D. W., Minter, D. W., and Stalpers, J. A. (2008). Dictionary of the Fungi, 10th Edn. Switzerland: CABI Europe.
Kreisel, H. (1996). A preliminary survey of the genus Phallus sensu lato. Czech Mycol. 48, 273–281. doi: 10.33585/cmy.48407
Kreisel, H. (2001). Checklist of the gasteral and secotioid Basidiomycetes of Europe, Africa, and the Middle East. Osterr. Z. Pilzkd. 10, 213–313.
Krüger, D., Binder, M., Fischer, M., and Kreisel, H. (2001). The Lycoperdales. A molecular approach to the systematics of some gasteroid mushrooms. Mycologia 93, 947–957. doi: 10.2307/3761759
Li, G., Deng, D., Wei, J., Zhang, C., Zhao, R., and Lin, F. (2018). Protubera beijingensis sp. nov. (Protophallaceae, Phallales) from China. Phytotaxa 348, 133–140. doi: 10.11646/phytotaxa.348.2.8
Li, H., Ma, X., Mortimer, P. E., Karunarathna, S. C., Xu, J., and Hyde, K. D. (2016). Phallus haitangensis, a new species of stinkhorn from Yunnan province, China. Phytotaxa 280, 116–128. doi: 10.11646/phytotaxa.280.2.2
Li, H., Mortimer, P. E., Karunarathna, S. C., Xu, J., and Hyde, K. D. (2014). New species of Phallus from a subtropical forest in Xishuangbanna, China. Phytotaxa 163, 91–103. doi: 10.11646/phytotaxa.163.2.3
Li, H., Tian, Y., Menolli, N., Ye, L., Karunarathna, S. C., Perez-Moreno, J., et al. (2021). Reviewing the world’s edible mushroom species: A new evidence-based classification system. Compr. Rev. Food Sci. Food Saf. 20, 1982–2014. doi: 10.1111/1541-4337.12708
Li, T., Li, T., Deng, W., Song, B., Deng, C., and Yang, Z. L. (2020). Phallus dongsun and P. lutescens, two new species of Phallaceae (Basidiomycota) from China. Phytotaxa 443, 19–37. doi: 10.11646/phytotaxa.443.1.3
Lu, Y., Gui, Y., Gong, G., Wei, S., and Zhu, G. (2014). Genetic diversity of 18 Dictyophora rubrovolvata germplasm resources from Guizhou. Guizhou Agric. Sci. 42, 17–20.
Læssøe, T., and Spooner, B. (1994). The uses of ‘Gasteromycetes’. Mycologist 8, 154–159. doi: 10.1016/S0269-915X(09)80179-1
Marincowitz, S., Coetzee, M. P. A., Wilken, P. M., Wingfield, B. D., and Wingfield, M. J. (2015). Phylogenetic placement of Itajahya: An unusual Jacaranda fungal associate. IMA Fungus 6, 257–262. doi: 10.5598/imafungus.2015.06.02.01
May, T. W., Sinnott, N., and Sinnott, A. (2010). The truffle-like “Protubera canescens” is an early developmental stage of the cage fungus “Ileodictyon.”. Vic. Nat. 127, 49–54.
Melanda, G. C. S., Accioly, T., Ferreira, R. J., Rodrigues, A. C. M., Cabral, T. S., Coelho, G., et al. (2020). Diversity trapped in cages: Revision of Blumenavia Möller (Clathraceae, Basidiomycota) reveals three hidden species. PLoS One 15:e0232467. doi: 10.1371/journal.pone.0232467
Miller, M. A., Pfeiffer, W., and Schwartz, T. (2010). Creating the CIPRES Science Gateway for inference of large phylogenetic trees. 2010 Gatew. Comput. Environ. Work. GCE 2010, 1–8. doi: 10.1109/GCE.2010.5676129
Miller, O. K., and Miller, H. H. (1988). Gasteromycetes: Morphological and Development Features. Eureka, CA: Mad Rivers Press.
Möller, A. (1895). Brasilische Pilzblumen. Botanische Mitteilungen aus den Tropen 7. Germen: Jena, G. Fischer.
Moreno, G., Khalid, A. N., Alvarado, P., and Kreisel, H. (2013). Phallus hadriani and P. roseus from Pakistan. Mycotaxon 125, 45–51. doi: 10.5248/125.45
MycoBank (2020). Excel version of the list of taxa present in MycoBank. Available Online at: https://www.mycobank.org/ (accessed August 11, 2020).
NCBI BLAST (2020). NCBI Nucleotide BLAST. Available Online at: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed September 1, 2020).
Nilsson, R. H., Larsson, K. H., Taylor, A. F. S., Bengtsson-Palme, J., Jeppesen, T. S., Schigel, D., et al. (2019). The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 47, D259–D264. doi: 10.1093/nar/gky1022
Patel, R. S., Vasava, A. M., and Rajput, K. S. (2018). Morphological and molecular evidence for the occurrence of Itajahya galericulata (Basidiomycota, Phallales) in India. Plant Fungal Syst. 63, 39–44. doi: 10.2478/pfs-2018-0006
Pegler, D. N., and Gomez, L. D. (1994). An unusual member of the cage fungus family. Mycologist 8, 54–59. doi: 10.1016/s0269-915x(09)80124-9
Phillips, E., Gillett-Kaufman, J. L., and Smith, M. E. (2018). Stinkhorn mushrooms (Agaricomycetes: Phallales: Phallaceae). EDIS 2018, 1–5. doi: 10.32473/edis-pp345-2018
Pietras, M., Rudawska, M., Iszkuło, G., Kujawa, A., and Leski, T. (2016). Distribution and molecular characterization of an alien fungus, Clathrus archeri, in Poland. Polish J. Environ. Stud. 25, 1197–1204. doi: 10.15244/pjoes/61230
Pine, E. M., Hibbett, D. S., and Donoghue, M. J. (1999). Phylogenetic relationships of cantharelloid and clavarioid Homobasidiomycetes based on mitochondrial and nuclear rDNA sequences. Mycologia 91, 944–963. doi: 10.2307/3761626
Põlme, S., Abarenkov, K., Henrik Nilsson, R., Lindahl, B. D., Clemmensen, K. E., Kauserud, H., et al. (2021). FungalTraits: a user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 105, 1–16. doi: 10.1007/s13225-020-00466-2
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Sanshi, I., and Kawamura, A. (1958). On the japanese species of Protubera. Sci. reports Yokohama Natl. Univ. Sect. II Biol. Geol. Sci. 7, 1–6.
Sato, H., Tsujino, R., Kurita, K., Yokoyama, K., and Agata, K. (2012). Modelling the global distribution of fungal species: New insights into microbial cosmopolitanism. Mol. Ecol. 21, 5599–5612. doi: 10.1111/mec.12053
Schoch, C. L., Ciufo, S., Domrachev, M., Hotton, C. L., Kannan, S., Khovanskaya, R., et al. (2020). NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020:baaa062. doi: 10.1093/database/baaa062
Skaltsas, D. N., Badotti, F., Vaz, A. B. M., da Silva, F. F., Gazis, R., Wurdack, K., et al. (2019). Exploration of stem endophytic communities revealed developmental stage as one of the drivers of fungal endophytic community assemblages in two Amazonian hardwood genera. Sci. Rep. 9:12685. doi: 10.1038/s41598-019-48943-2
Stamatakis, A. (2006). RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690. doi: 10.1093/bioinformatics/btl446
Sulzbacher, M. A., Grebenc, T., Cabral, T. S., Giachini, A. J., Goto, B. T., Smith, M. E., et al. (2016). Restingomyces, a new sequestrate genus from the Brazilian Atlantic rainforest that is phylogenetically related to early-diverging taxa in Trappeaceae (Phallales). Mycologia 108, 954–966. doi: 10.3852/15-265
Tedersoo, L., Bahram, M., Põlme, S., Kõljalg, U., Yorou, N. S., Wijesundera, R., et al. (2014). Global diversity and geography of soil fungi. Science 346:1256688. doi: 10.1126/science.1256688
Trierveiler-Pereira, L., da Silveira, R. M. B., and Hosaka, K. (2014a). Multigene phylogeny of the Phallales (Phallomycetidae, Agaricomycetes) focusing on some previously unrepresented genera. Mycologia 106, 904–911. doi: 10.3852/13-188
Trierveiler-Pereira, L., Meijer, A. A. R., Hosaka, K., and Silveira, R. M. B. (2014b). Updates on Protubera (Protophallaceae, Phallales) and additional notes on P. maracuja. Mycoscience 55, 35–42. doi: 10.1016/j.myc.2013.05.001
UNITE BLAST (2020). UNITE BLAST. Available Online at: https://unite.ut.ee/analysis.php (accessed September 1, 2020).
Vu, D., Groenewald, M., de Vries, M., Gehrmann, T., Stielow, B., Eberhardt, U., et al. (2019). Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 92, 135–154. doi: 10.1016/j.simyco.2018.05.001
Wijayawardene, N. N., Hyde, K. D., Al-Ani, L. K. T., Tedersoo, L., Haelewaters, D., Rajeshkumar, K. C., et al. (2020). Outline of Fungi and fungus-like taxa. Mycosphere 11, 1060–1456. doi: 10.5943/mycosphere/11/1/8
Keywords: gasteroid fungi, GenBank, phylogeny, stinkhorns, UNITE
Citation: Melanda GCS, Silva-Filho AGS, Lenz AR, Menolli N Jr, Lima AA, Ferreira RJ, Assis NM, Cabral TS, Martín MP and Baseia IG (2021) An Overview of 24 Years of Molecular Phylogenetic Studies in Phallales (Basidiomycota) With Notes on Systematics, Geographic Distribution, Lifestyle, and Edibility. Front. Microbiol. 12:689374. doi: 10.3389/fmicb.2021.689374
Received: 31 March 2021; Accepted: 21 May 2021;
Published: 09 July 2021.
Edited by:
Dhanushka Nadeeshan Wanasinghe, Chinese Academy of Sciences, ChinaReviewed by:
Henrik R. Nilsson, University of Gothenburg, SwedenCopyright © 2021 Melanda, Silva-Filho, Lenz, Menolli, Lima, Ferreira, Assis, Cabral, Martín and Baseia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nelson Menolli Jr., bWVub2xsaWpyQHlhaG9vLmNvbS5icg==
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.