 Xi Yuan1,2
Xi Yuan1,2 Fusheng Chen1,2*
Fusheng Chen1,2*- 1Hubei International Scientific and Technological Cooperation Base of Traditional Fermented Foods, Huazhong Agricultural University, Wuhan, China
- 2College of Food Science and Technology, Huazhong Agricultural University, Wuhan, China
Cocultivation is an emerging and potential way to investigate microbial interaction in the laboratory. Extensive researches has been carried out over the years, but some microorganism cocultivation are not easy to implement in the laboratory, especially the fungus-fungus (FF) cocultivation, owing to the obstacles such as fungal different growth rate, limited growing space, hyphae intertwining, and difficulty of sample separation, etc. In this research, a double-sided petri dish (DSPD) was designed and carried out as a tool to study FF cocultivation in the laboratory. A natural FF cocultivation of Monascus spp. and Aspergillus niger inspired from black-skin-red-koji (BSRK), were studied. By using DSPD, the aforementioned obstacles in the FF cocultivation study were overcome through co-culturing Monascus spp. and A. niger on each side of DSPD. The characteristics of monocultured and co-cultured Monascus spp. and A. niger were compared and analyzed, including colonial and microscopic morphologies, and main secondary metabolites (SMs) of Monascus spp. analyzed by high performance liquid chromatography. And a novel SM was found to be produced by Monascus ruber M7 when co-cultured with A. niger CBS 513.88. Since the above mentioned obstacles, were overcome, we obtained good quality of transcriptome data for further analysis. These results indicate that DSPD might be an efficient tool for investigation of microbial interaction, in particular, for FF interaction.
Introduction
Black-skin-red-koji (BSRK), also known as Wuyihongqu (乌衣红曲) in Chinese, is a special traditional Chinese mixed starter which has been utilized for brewing rice wine and cereal vinegar for more than 1,000 years mainly in Fujian and Zhejiang provinces (Fang et al., 2011; Huang et al., 2018). As a representative model of fungus-fungus (FF) cocultivation, the main filamentous fungi in BSRK are Monascus spp. and Aspergillus niger (Huang et al., 2019). Usually, A. niger strains grow on the surface of the steamed rice, and Monascus spp. strains grow in the center of the steamed rice, so BSRK has a “red heart” and black “coat” (Figure 1).

Figure 1. Appearance and cross-sections of black-skin-red-koji (BSRK).
Monascus spp. belong to the family Monascaceae and were first screened from red yeast rice and characterized by van Tieghem (1884). Monascus spp. have been used to produce the food and health products due to their functional secondary metabolites (SMs) such as Monascus pigments (MPs), monacolin K (MK), dimerumic acid, and γ-aminobutyric acid (Chen et al., 2015; Feng et al., 2016). However, nephrotoxin citrinin (CIT) can also be produced by a few of Monascus spp. strains, which can lead to the food safety concerns of Monascus related products (He and Cox, 2016). A. niger is most widely used industrially for citric acid production and is also a tool for the production of enzymes such as α-amylase, cellulose, and pectinase (Nielsen et al., 2009; Andersen et al., 2011; Show et al., 2015). However, to date, as a natural FF cocultivation model, the reason why both of Monascus spp. and A. niger can harmoniously grow in BSRK and the mechanisms relating to how they affect each other are still undiscovered.
Cocultivation is regarded as an experimental imitation of the competition within natural microbe communities in laboratory conditions, microbial cocultivation has been studied for various uses, such as improving SMs production (Wakefield et al., 2017), accelerating the fermentation process (Yan et al., 2008), increasing bioenergy production (Camacho-Zaragoza et al., 2016), speeding up organic pollutants degradation (Kumari and Naraian, 2016), and especially mining novel SMs (Marmann et al., 2014; Yu M. et al., 2019). To date, microbial cocultivation research can be generally classified into three models including bacteria-bacteria (Zhang et al., 2015; Gao et al., 2021), bacteria-fungus (Moussa et al., 2019), and FF (Jimenez-Barrera et al., 2018). Since fungi are one of the biggest sources of natural products and enzymes (Meyer et al., 2016), research on the FF cocultivation has contributed to enhancing SMs production and discovering novel SMs (Shang et al., 2017; Vinale et al., 2017; Yu G. et al., 2019). However, compared to the other models, FF cocultivation is lesser studied, probably due to technical difficulties under laboratory conditions such as fungal different growth rates, limited growth spaces, hyphae intertwining, and difficulty in sample separation, etc.
Genome sequencing technology has revealed that there are a large number of gene clusters regulating SMs synthesis in fungi. However, when fungi are under laboratory culture conditions, most of fungal SMs gene clusters remaines silent (Pettit, 2009). Cocultivation is one of the most efficient methods to activate the silent fungal SMs gene clusters through interspecific communications by co-culture of two or more kinds of fungi in a specific culture environment (Nett et al., 2009; Chiang et al., 2011; Xu et al., 2015).
Since A. niger and Monascus spp. can be well symbiotic in BSRK, both of them may be an ideal research model of cocultivation. However, usually A. niger grows much faster than Monascus spp., which has become an obstacle to observing and analyzing the variation between monoculture and co-culture (Favela-Torres et al., 1998). The preliminary results also showed (data not shown) that Monascus spp. and A. niger could grow together which led to a difficulty in sample separations for further analysis. Furthermore, BSRK and its major fermentation products are all based on rice (Chen et al., 2009; Li et al., 2009), sugars and proteins from rice can cause serious interference to transcriptome sequencing.
In current study, a double-sided petri dish (DSPD, Figure 2) was designed to study the cocultivation of M. ruber M7 which can produce high yields of MPs and CIT but no MK, M. pilosus MS-1which can produce high yields of MK but no CIT (Feng et al., 2016; Wang et al., 2016), A. niger CBS 513.88 which is often used for enzyme production, and A. niger CBS 113.46 which is usually applied for citric acid production (Andersen et al., 2011), respectively. The colonial and microscopic morphologies were observed and compared, main SMs were analyzed, novel SMs were mined, and transcriptomes were elucidated, when the aforementioned fungal strains were monocultured and co-cultured on DSPD. The results are proven that DSPD is an efficient tool for FF cocultivation research.

Figure 2. Structure drawing, vertical view, and front view of the double-sided petri dish (DSPD).
Materials and Methods
Strains and Chemicals
The strains M. ruber M7 (CCAM 070120, Culture Collection of State Key Laboratory of Agricultural Microbiology, which is part of China Center for Type Culture Collection, Wuhan, China), M. pilosus MS-1 (CCTCC M 2013295, China Center for Type Culture Collection, Wuhan, China) and A. niger CBS 513.88, A. niger ATCC 1015 (Both were purchased from The Westerdijk Fungal Biodiversity Institute) were used in this study. The potato dextrose agar (PDA), potato dextrose broth (PDB), Czapek yeast extract agar (CYA), malt extract agar (MEA), and 25% glycerol nitrate agar (G25N) were taken as culture media. PDB: Potato (200 g/L), sugar (20 g/L), agar power (2 g/L); PDA: PDB with agar power (2 g/L); CYA: NaNO3 (3 g/L), K2HPO4 (1 g/L), KCl (0.5 g/L), MgSO4⋅7H2O (0.5 g/L), FeSO4 (0.01 g/L), sucrose (30 g/L), yeast extracts power (1 g/L), agar power (2 g/L); MEA: Malt extract power (30 g/L); G25N: CYA with 25% glycerol. Rice powders agar (RA): rice power (50 g/L), agar power (2 g/L). MK and CIT standard substances were purchased from Sigma Co. Other chemicals were bought from China National Medicines Corporation Ltd.
Design of a Double-Sided Petri Dish
A double-sided petri dish (DSPD, Figure 2) was produced by two resin petri dishes (ϕ = 9.0 cm). Firstly, the bottoms of two petri dishes were sticked together by glue, then a hole (ϕ = 2.0 cm) in the middle of the petri dishes was drilled. Each DSPD consists of one body part and two covers. When DSPD is used, the metabolites, signal factors, ect. that are produced by the tested microbial strains grown on each side of DSPD can pass and interact through the hole (Xu et al., 2018; Yan et al., 2008). Prior to pouring the media in to DSPD, a round piece of sterile cellophane (ϕ≈ 9.0 cm) is pasted on any side of the DSPD to cover the hole (communicating area) and prevent the media from leaking. After the media solidification on one side, the media are poured on another side.
Cocultivation Experiments
Two Monascus spp. strains were co-cultured with two A. niger strains by the following combinations: M. ruber M7 and A. niger CBS 513.88, M. ruber M7 and A. niger CBS 113.46, M. pilosus MS-1 and. niger CBS 513.88, and M. pilosus MS-1 and A. niger CBS 113.46. All the combinations were investigated by the conventional cocultivation and the cocultivation using DSPD.
Conventional Cocultivation
The conventional cocultivation, means co-culturing two fungal strains in the same petri dish directly (Zuck et al., 2011; Bertrand et al., 2013; Nonaka et al., 2015). Firstly, the strains were cultured on PDA slants at 30 ± 1°C for 7 days and then sterile water was added into the slants and spores were scraped using an inoculating loop. After the mycelia were filtered by sterile filter paper, the spore solution of Monascus spp. and A. niger were obtained, respectively (Feng et al., 2014). After that, two inoculation methods were used for cocultivation. Coating inoculation: 1 mL Monascu spp. and A. niger spore solutions were evenly spread on half of the same petri dish, respectively. Point inoculation: 1 μL Monascus spp. and A. niger spore solutions were inoculated on two points of the same petri dish, respectively, and the distance of the two points was about 2 cm (Shang et al., 2017; Figures 3, 4). After inoculation, all the petri dishes were incubated at 30 ± 1°C for 9 days.
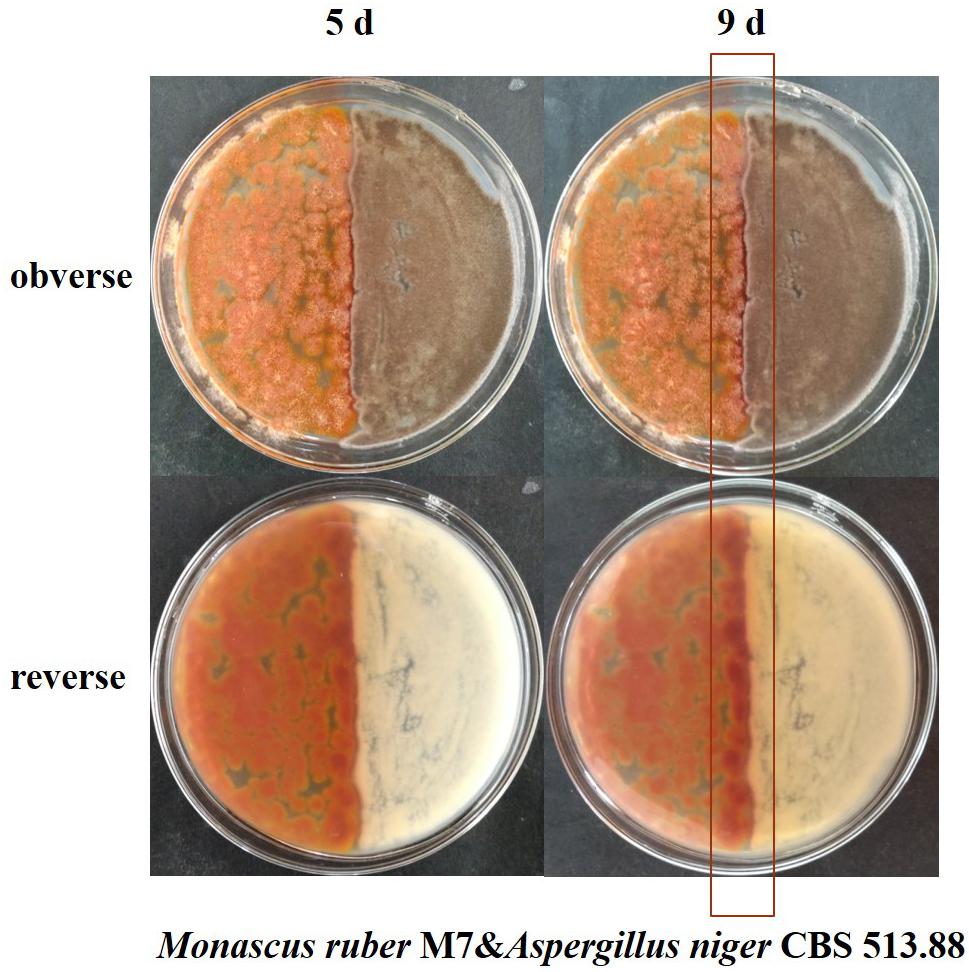
Figure 3. Cocultivation of Monascus ruber M7 and Aspergillus niger CBS 513.88 by coating inoculation method at 5 and 9 days &-co-cultured with.
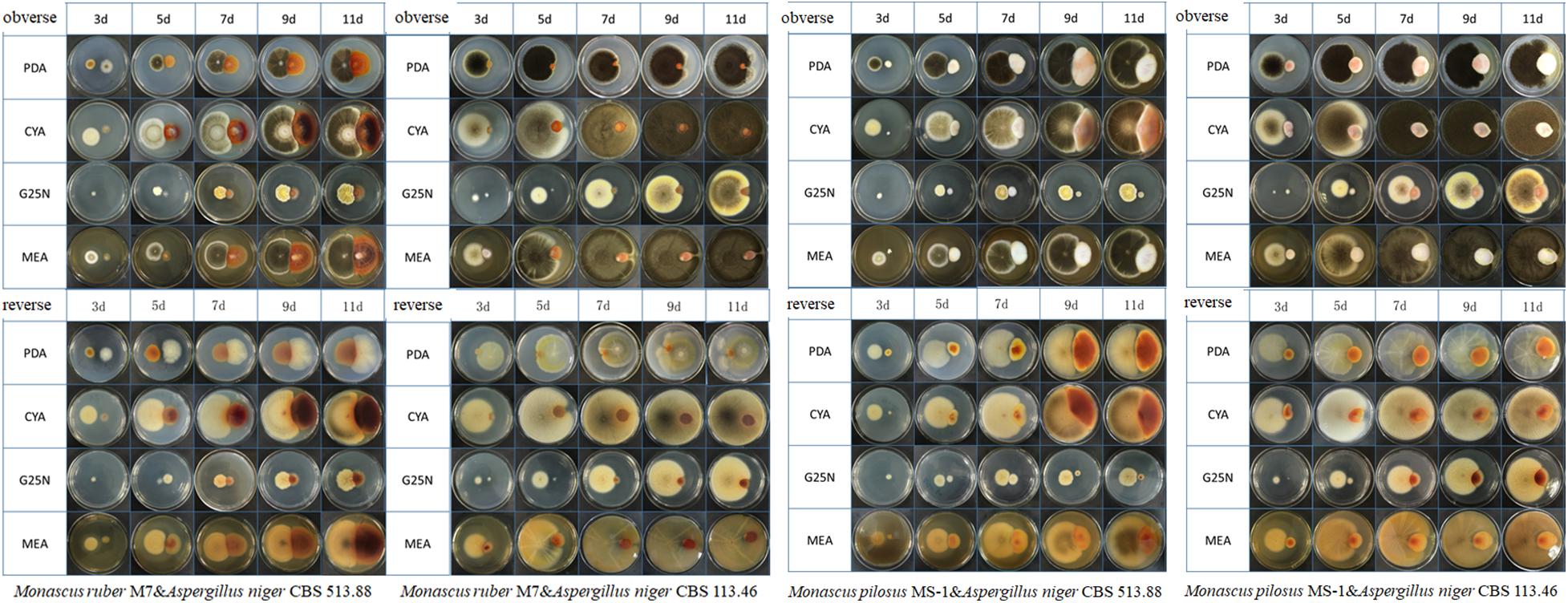
Figure 4. Colonial morphologies of Monascus ruber M7, Monascus pilosus MS-1 when co-cultured with Aspergillus niger CBS 513.88 and A. niger CBS 113.46 by conventional (direct) cocultivation at 3, 5,7,9, and 11 days &-co-cultured with.
Cocultivation in DSPD
After the media solidification in two sides, the tested Monascus spp. and A. niger strains were inoculated on each side of DSPD, respectively. The cocultivation conditions were the same as the conventional cocultivation.
Observation of Colonial and Microscopic Morphologies
The colonial morphologies of M. ruber M7, M. pilosus MS-1, A. niger CBS 513.88, and A. niger CBS 113.46 were observed by the naked eye, and photos were taken by camera at 3, 5, 7, and 9 days (Figures 3–5). The micromorphologies were observed and photos were taken by the microscope (Leica, DM300, Wetzlar, Germany), focusing on mycelia, conidia, and cleistothecia (Figure 6).
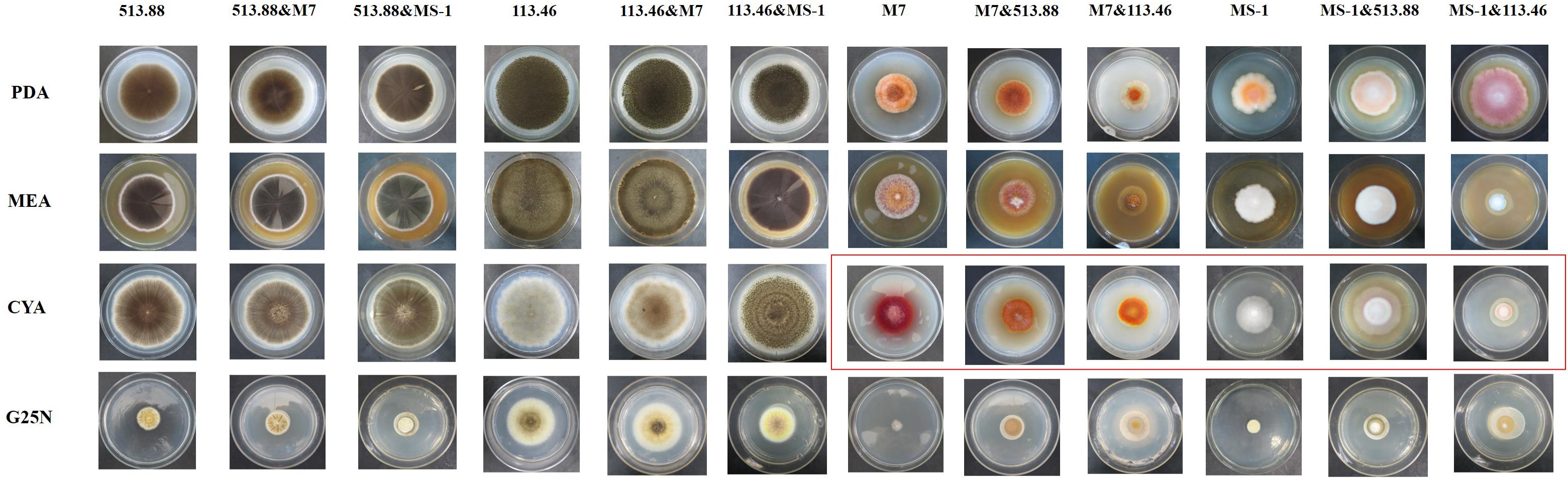
Figure 5. Colonial morphologies of Monascus ruber M7, Monascus pilosus MS-1 when co-cultured with Aspergillus niger CBS 513.88 and A. niger CBS 113.46 by using DSPD at 9 days. M7-M. ruber M7; MS-1-M. pilosus MS-1; 513.88-A. niger CBS 513.88; 113.46-A. niger CBS 113.46; &-co-cultured with.
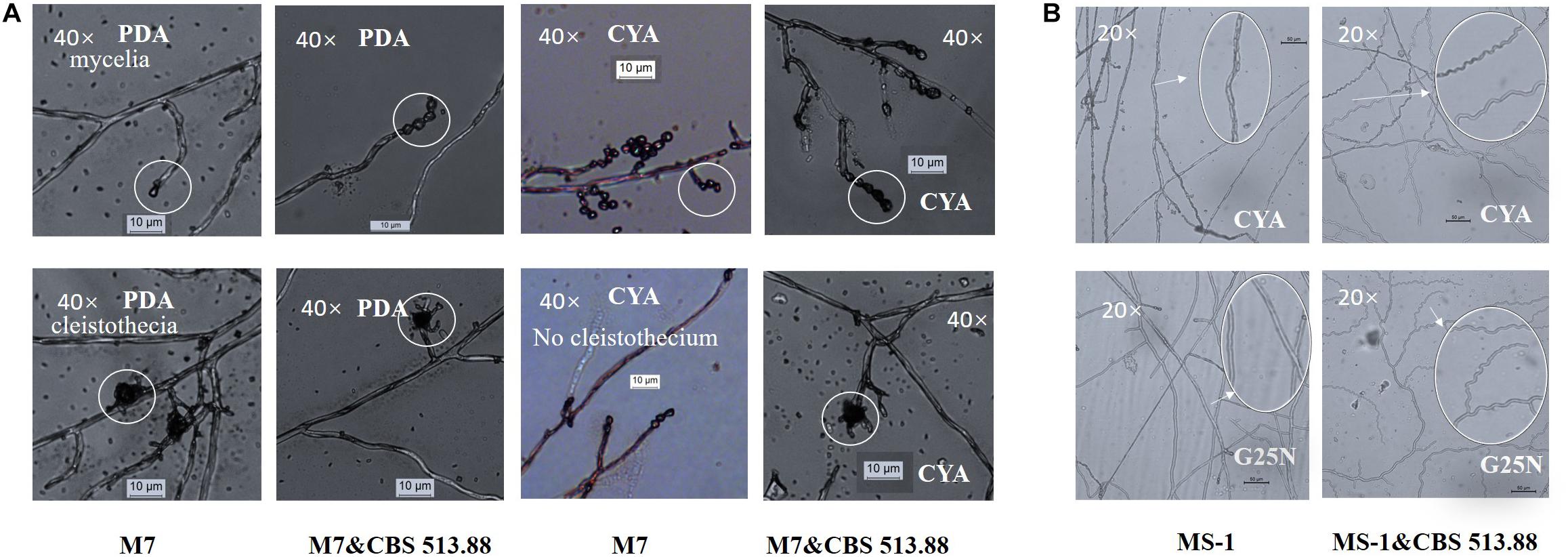
Figure 6. Cocultivation micromorphologies: (A) comparison of monocultured and co-cultured (with Aspergillus niger CBS 513.88) M. ruber M7 on PDA and CYA media; (B) mycelia shape variation of M. pilosus MS-1 when co-cultured with A. niger CBS 513.88. Abbreviations: M7-M. ruber M7; M7&CBS 513.88-M. ruber M7 co-cultured with A. niger CBS 513.88; MS-1-M. pilosus MS-1; MS-1&CBS 513.88-M. pilosus MS-1 co-cultured with A. niger CBS 513.88.
Determination of Color Value of MPs
MPs can be divided into Monascus yellow pigment (MYP, 330–450 nm), Monascus orange pigment (MOP, 460–480 nm), and Monascus red pigment (MRP, 490–530 nm) according to MPs maximum absorption wavelength (Feng et al., 2012; Chen et al., 2017). The color values, which can represent the MPs contents, are defined as optical density values at the given wavelength per milliliter or gram (Feng et al., 2012). Samples were dried by vacuum freeze drying and then weighed (about 1 mg), dissolved by 70% ethanol, and subject to ultrasonic extraction. The extracted solutions were filtrated by filter paper and diluted by 70% ethanol. Diluted solutions were detected by UV–vis UV-1700 Spectrophotometer (Shimadzu, Tokyo, Japan). One unit of optical density at a given wavelength corresponded to one unite (U) of color value (Li et al., 2019):
where A is the absorbance of the pigment extract at a given wavelength, V is the total volume of extracted solutions, and m is the dry weight of the samples used for pigment extraction. The given wavelengths used to detect MYP, MOP, and MRP were 380, 470, and 520 nm, respectively. In addition, 505 nm was taken as the given wavelength to measure total MPs (Ding et al., 2008; Wang et al., 2016).
Determination of SMs
MK, CIT and MPs varieties were detected by high performance liquid chromatography (HPLC; Feng et al., 2014, 2016; Li et al., 2019). Samples for MK and MPs detection were ultrasonically extracted for 1 h, then centrifugated at 12000 rpm (Heal Force Neofuge 15R, Shanghai, China) for 10 min. Samples for CIT detection were dried and ultrasonically extracted with 80% methanol solvent, then centrifugated at 12000 rpm for 40 min. The supernatants were filtered through a 0.22 μm membrane before HPLC detection. Then MK, CIT and MPs were assessed by HPLC, which were performed on a chromatographic system (Shimadzu, model LC-20A Prominence, Tokyo, Japan), equipped with a diode-array detector (Shimadzu, Tokyo, Japan). The column of inertsil ODS-3 (4.6 mm × 250 mm, id, 5 μm) was employed. For MK detection, the mobile phases were acetonitrile (ACN) and water (contains 0.1% phosphoric acid) (60%:40%, v/v) by isocratic elution, the column temperature was set at 25°C, the flow rate was maintained at 1.0 mL/min and the injection volume was 20 μL. Both acid and lactone forms of MK were calculated as MK yield (Feng et al., 2014). For MPs detection, the mobile phases were ACN and water (contains 0.1% formic acid). Gradient elution was performed as follows: step gradient for ACN was 55% (v/v) to 65% (v/v) in 3 min; 65% (v/v) to 90% (v/v) in 22 min; 90% (v/v) for 5 min; 90% (v/v) to 55% in 1 min; 55% (v/v) for 9 min. The column temperature was kept at 30°C. The detection wavelength was 210 nm to 600 nm. For CIT detection, the mobile phases were ACN and water (contains 0.1% formic acid) (70%:30%, v/v) by isocratic elution, and the flow rate was 1 mL/min. The observation wavelength was 331 nm (Liu et al., 2016). The column temperature was set at 30°C, and the injection volume was 10 μL. When co-cultured with A. niger, MK producing strain M. pilosus MS-1 was selected to study MK variation while M. ruber M7 was selected to study CIT and MPs variation.
Mining Novel SMs
Our preliminary results (data not shown) found two new SMs from the cocultivation of M. ruber M7 and A. niger CBS 513.88 in the steamed rice. In this study, DSPD was applied for further research and rice agar was employed to simulated steamed rice. By using DSPD, the mycelia of M. ruber M7 were harvested and extracted by ethyl acetate, rotary evaporated, redissolved by 70% ethyl alcohol, then detected by HPLC. The mobile phase condition was ACN and water (contains 0.1% phosphoric acid) (70%:30%, v/v). The observation wavelength was 420 nm according to the maximum absorption wavelength.
Mycelia Preparation for Transcriptome
DSPD was utilized to separate the tested strains, and cellophane was covered on the surfaces of rice agar. After 5 days of cocultivation, both fungi grew well and the mycelia of co-cultured M. ruber M7 and A. niger CBS 513.88 were scraped down and collected from the cellophane, respectively. After liquid nitrogen freezing, the mycelia were stored at −80°C and sent to Shanghai Majorbio Bio-pharm Technology Co., Ltd. for transcriptome sequencing.
RNA Isolation, Library Preparation, and Sequencing
Total RNA was extracted from mycelia using the TRIzol method (Invitrogen, Carlsbad, CA, United States), and the concentration and purity of the extracted RNA were detected by Nanodrop2000. RNA integrity was detected by agarose gel electrophoresis, and the RNA integrity number (RIN) value was determined by Agilent2100. The libraries were constructed after the RNA samples were qualified by using magnetic beads with Oligo(dT) and polyA to pair A-T bases to enrich mRNA. The fragmentation buffer solution was added to randomly break the mRNA into small fragments of about 300 bp. Double-stranded cDNA was synthesized using a SuperScript double-stranded cDNA synthesis kit (Invitrogen, Carlsbad, CA, United States) with random hexamer primers (Illumina). Then the synthesized cDNA was subjected to end-repair, phosphorylation, and “A” base addition according to Illumina’s library construction protocol. The cDNA library was constructed with an Illumina Paired End Sample Prep kit (Illumina, San Diego, CA, United States), quantified by TBS380 (Picogreen, Invitrogen, Carlsbad, CA, United States), and was then sequenced on the Illumina HiSeqTM2500 (2 × 150 bp read length) platform. The transcriptomeic data were analyzed online platform of Majorbio Cloud Platform (www.majorbio.com).
Statistical Analysis
All experiments were carried out in triplicate. Analysis of variance (ANOVA) was computed for testing the significance of the experiment. Data were analyzed in Microsoft Excel.
Results
Comparison of Colonial Morphologies of the Strains on DSPD
Conventional cocultivation of Monascus spp. and A. niger was carried out at first. Taking M. ruber M7 and A. niger CBS 513.88 as an example of coating inoculation method (Figure 3), no antagonistic zone can be observed between M. ruber M 7 and A. niger CBS 513.88. The area color of the Monascus spp. colony, which had contacted with A. niger CBS 513.88, became redder. By point inoculation method of a different combination of Monascus spp. and A. niger, there was still no antagonistic zone and the contacted area color was also redder (Figure 4). No significant change in the A. niger colony was observed but A. niger CBS 113.46 can inhibit the growth of Monascus spp. As it was also limited by the growth space, the shape of the colony was irregular. Figure 5 showed that by using DSPD, besides the results observed by conventional cocultivation, all the colonies kept the original shape and the color of the communicating area of A. niger, which seemed redder than other areas in the colonies of Monascus strains. On the PDA media, the color of co-cultured M. ruber M7 was redder than those of the monocultured examples. Especially on CYA media, the colony color of M. ruber M7 changed from red to orange when co-cultured with A. niger CBS 513.88.
Comparison of Micromorphologies of the Strains on DSPD
The micromorphologies of monocultured and co-cultured Monascus spp. and A. niger could hardly compared by conventional cocultivation due to the difficulty in separation and distinguishing of the tested strains. The DSPD solved the problem. The results showed that at the 3rd day, on CYA media, cleistothecia could not be observed in monocultured M. ruber M7 but could be observed in co-cultured M. ruber M7 (with A. niger CBS 513.88) (Figure 6A). At the 7th days, on CYA and G25N media, the mycelia of M. pilosus MS-1 changed significantly from straight to curve (Figure 6B). For the micromorphologies of tested A. niger strains, no significant change was observed.
Analysis of SMs From Monascus spp. Strains on DSPD
Color Value Variation by Cocultivation
As a MP producing strain, M. ruber M7 was selected to investigate the changes of color value and MPs (Chen et al., 2017). The results demonstrated a significant change in color value, which reflected the MPs variation of M. ruber M7 when co-cultured with A. niger strains, respectively. As illustrated in Figure 7, it was clear that both A. niger CBS 513.88 and A. niger CBS 113.46 could induce M. ruber M7 to produce more intracellular (mycelia) MPs (Figure 7A), among which A. niger CBS 513.88 acted better than A. niger CBS 113.46 in increasing MPs yields. The color value of orange pigments, red pigments, and total pigments increased nearly 6-fold, 4-fold, and 3-fold, respectively. Compared to A. niger CBS 513.88, A. niger CBS 113.46 was less efficient in the increase of orange and red pigments but was equivalent to an increase of yellow pigments. However, both selected A. niger strains had little effect on extracellular (media) MPs of M. ruber M7 (Figure 7B).
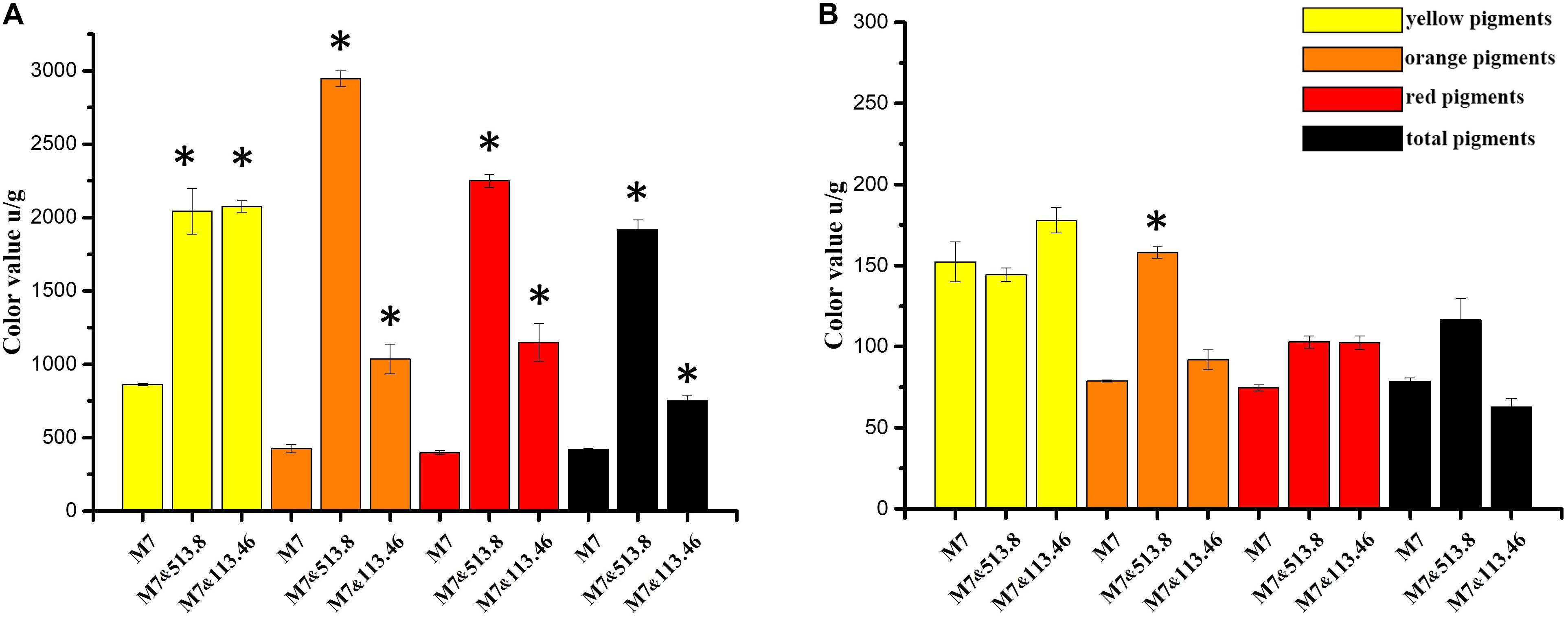
Figure 7. Color value variation of Monascus spp. and Aspergillus niger cocultivation: (A) comparison of Monascus ruber M7 intracellular (mycelia) color value among monoculture, co-cultured with A. niger CBS 513.88 and co-cultured with A. niger CBS 113.46; (B) comparison of M. ruber M7 extracellular (media) color value among monoculture, co-cultured with A. niger CBS 513.88 and co-cultured with A. niger CBS 113.46. Abbreviations: M7-monocultured M. ruber M7; M7&513.88-M. ruber M7 co-cultured with A. niger CBS 513.88; M7&113.46-M. ruber M7 co-cultured with A. niger CBS 113.46. *Represents significant difference compared with control (P < 0.05).
MK and CIT Variation by Cocultivation
MK and CIT are both crucial SMs of Monascus spp. As detailed in Table 1, MK was only produced by monocultured M. pilosus MS-1 on PDA media. When M. pilosus MS-1 was co-cultured with A. niger CBS 513.88 and A. niger CBS 113.46, respectively MK was undetectable. As a non-CIT producing strain, CIT was undetected in M. pilosus MS-1 samples as reported (Feng et al., 2016). According to the CIT content of monocultured and co-cultured M. ruber M7, both A. niger strains could decrease the intracellular CIT content while increasing the extracellular CIT content of M. ruber M7.
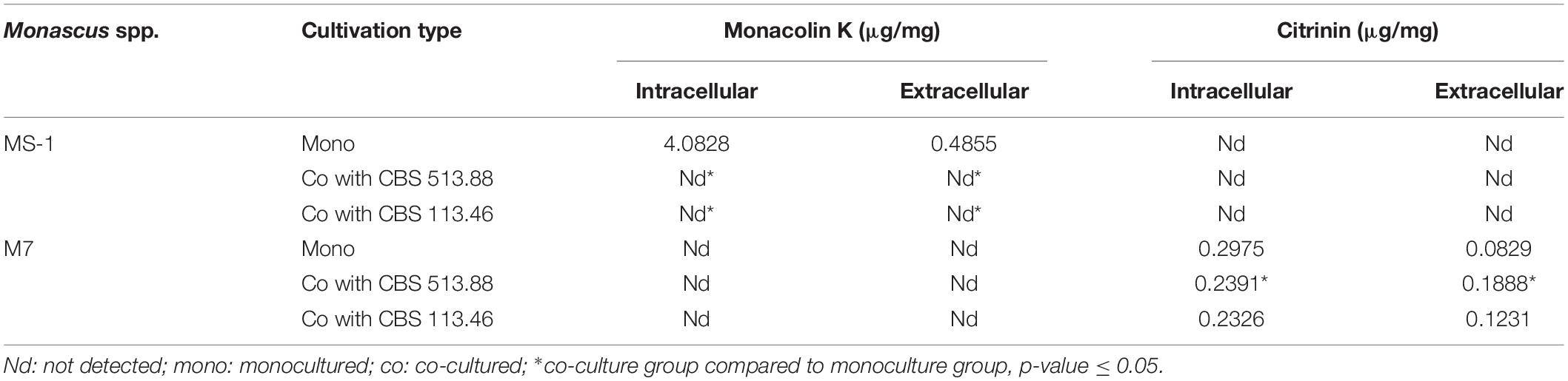
Table 1. Monacolin K and citrinin variation when Monascus spp. were co-cultured with Aspergillus niger on PDA media.
MPs Variation by Cocultivation
As mentioned in the colonial morphologies results (Figure 5), on DSPD we observed that on PDA and CYA media, the colony color of M. ruber M7 changed significantly. The HPLC results in Figure 8 could support the above conclusion. The MPs peaks of M. ruber M7 can be identified by retention time and UV spectrum according to our previous work (Chen et al., 2017). Under the observing wavelength of 380 nm (Chen et al., 2017), one yellow pigment decreased while one orange pigment increased when M. ruber M7 was co-cultured with A. niger CBS 513.88 on PDA media. On the CYA media, the colony color changed from red to orange due to the production of more yellow pigments when M. ruber M7 was stimulated by A. niger CBS 513.88.
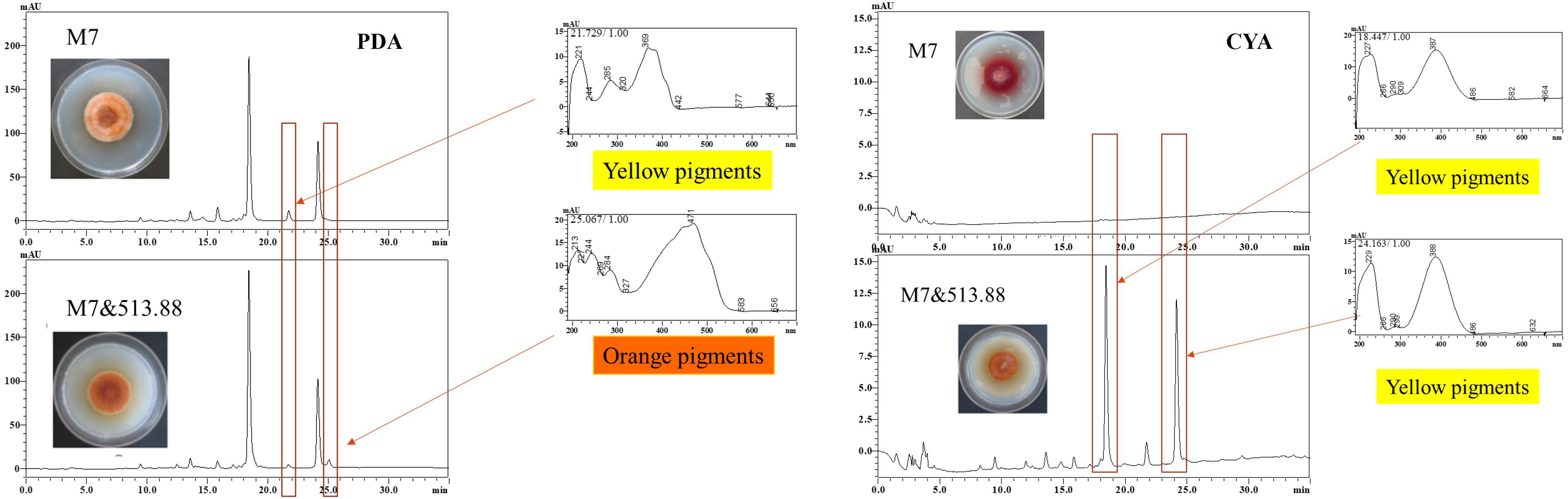
Figure 8. High performance liquid chromatography results of MPs of monocultured Monascus ruber M7 and co-cultured M. ruber M7 with Aspergillus niger CBS 513.88.
Mining and Primary Localization of Novel SMs by DSPD
Two novel SMs, namely substance A and B, were discovered by cocultivation of M. ruber M7 and A. niger CBS 513.88 on the steamed rice in our preliminary experiments (Data not shown). However, the rice-based mixture was not easily analyzed further on account of sample separation. As we saw from Figure 9, when DSPD was applied, substance A can only be detected in the mycelia of co-cultured M. ruber M7. Substance B with low yield was detected in culture media and A. niger CBS 513.88 mycelia. Compared to monocultured A. niger CBS 513.88, a new peak appeared in co-cultured A. niger CBS 513.88 mycelia. According to the retention time and UV spectrum, this new peak was similar to peak C that appeared in M. ruber M7 mycelia. According to the previous work of our lab (Chen et al., 2017), peak C was identified as a yellow Monascus pigment.
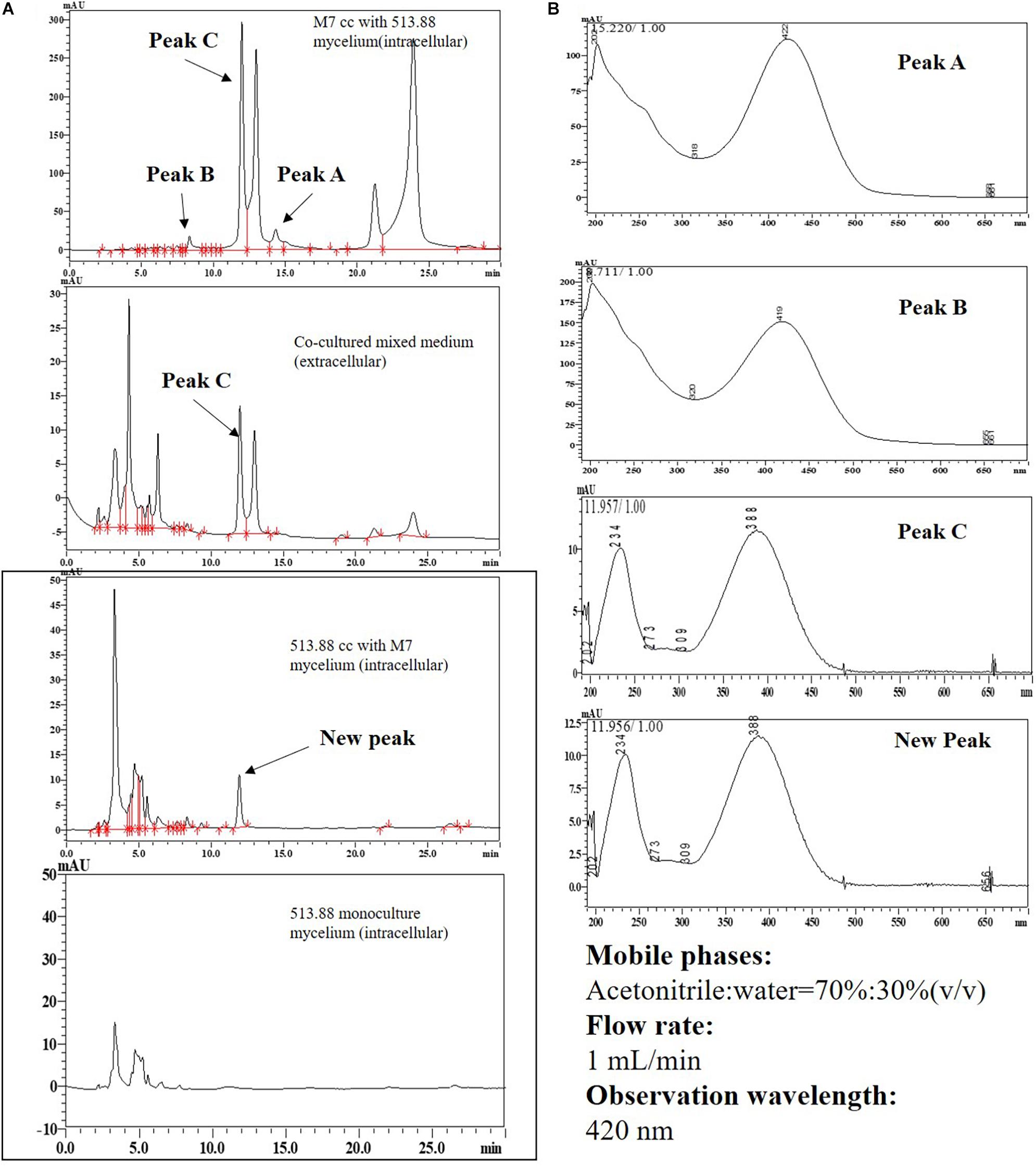
Figure 9. High performance liquid chromatography results of substance A and B in different cocultivation samples: (A) HPLC results; (B) UV-spectrum results. Abbreviations: CC-cocultured; M7-Monascus ruber M7; 513.88- A. niger CBS 513.88.
Transcriptomic Analysis of the Strains on DSPD
Considering that novel SMs can only be detected in the combination of M. ruber M7 and A. niger CBS 513.88, and that the genomes of these two fungi have been sequenced or published on NCBI (Andersen et al., 2011). the combination of M. ruber M7 and A. niger CBS 513.88 was selected for transcriptome analysis. The samples of co-cultured M. ruber M7. and co-cultured A. niger CBS 513.88 on DSPD were used for total RNA isolation, respectively. Taken samples on the fifth day as an example, in this case monocultured M. ruber M7, co-cultured M. ruber M7, monocultured A. niger CBS 513.88, and co-cultured A. niger CBS 513.88, total four samples were chosen as samples to study transcriptome variation during cocultivation. The results of agarose gel electrophoresis (Figure 10) showed that all samples had clear 28S and 18S bands, no significant contamination of proteins, sugars, and other impurities were observed and good RNA integrity was achieved. All samples were in line with sequencing requirements and met the requirements for library construction (Table 2).
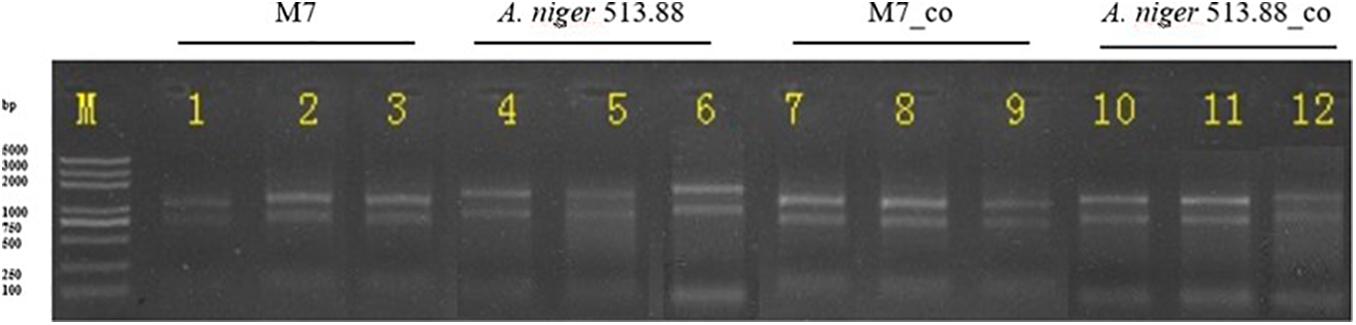
Figure 10. Total RNA of 12 samples for RNA-Seq in 1% agarose gel electrophoresis.
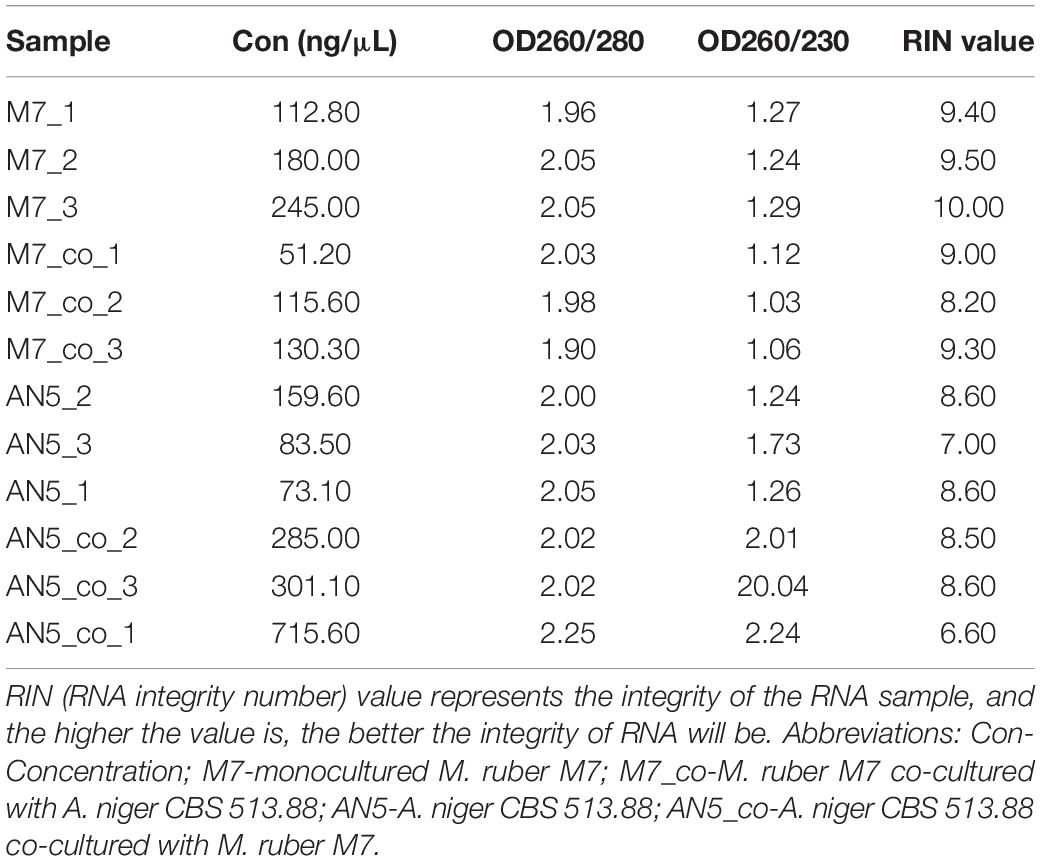
Table 2. The quality results of the total RNA of 12 samples.
For M. ruber M7 samples, after quality control processing, a total of 46.14 Gb of clean data were obtained. The clean data of each sample reached more than 6.76 Gb. A total of 9,165 genes were detected in this analysis, including 8,671 known genes and 494 new genes. There were 15,985 expressed transcripts, including 8,262 known transcripts and 7,723 new transcripts. The quality values of base Q20 and Q30 were higher than 98% and 95%, respectively, and GC content was higher than 52%, indicating that the data quantity was rich and reliable, which could be used for subsequent analysis in this study. The obtained clean reads were compared with the reported M. purpureus genome1, and there were six libraries (as listed in Table 3), and more than 95.13% of clean reads in each library could be compared with the reference genome.
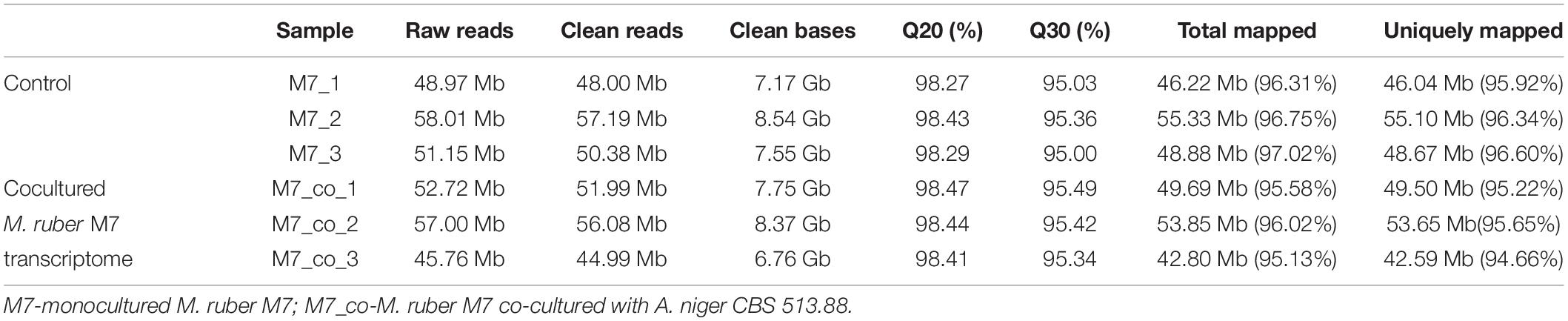
Table 3. RNA-Seq clean reads and the counts, percentage of reads mapping statistics to Monascus purpureus genome.
For A. niger CBS 513.88 samples, after quality control processing, a total of 46.33 Gb clean data were obtained, and the clean data of each sample reached more than 7.39 Gb. A total of 12,635 expressed genes were detected in this analysis, including 12,428 known genes and 207 new genes. There were 17,068 expressed transcripts, including 11,653 known transcripts and 5,415 new transcripts. The quality values of base Q20 and Q30 were higher than 98% and 95%, respectively, and GC content was higher than 54%, indicating that the data quantity was rich and reliable, which could be used for subsequent analysis in this study. The obtained clean reads were compared with the reported A. niger CBS 513.88 genome2, and there were six libraries (as listed in Table 4), and more than 93.37% of clean reads in each library that could be compared with the reference genome.

Table 4. RNA-Seq clean reads and the counts, percentage of reads mapping statistics to Aspergillus niger CBS 513.88 genome.
Discussion
Black-skin-red-koji, as a natural FF cocultivation model, has been applied for wine and vinegar production for over 1,000 years but has been seldom studied (Fang et al., 2011; Mao et al., 2011). In this work, a DSPD was developed and applied to explore BSRK, started from the cocultivation of Monascus spp. and A. niger. In conventional cultivation of Monascus spp. and A. niger, we found no antagonism but a symbiosis between Monascus spp. and A. niger (Figures 3, 4). Moreover, Monascus spp. may be stimulated to produce more MPs (Figure 7). Although Monascus spp. and A. niger can grow together, A. niger, especially A. niger CBS 113.46 can inhibit the growth of Monascus spp. strains. It became more meaningful to lucubrate the reason why Monascus spp. and A. niger can grow together and has been applied to fermented food for over 1,000 years. Further molecular mechanisms should be studied to reveal the meaning of this cocultivation mode in the biological and food industry. According to the micromorphologic results, at the third day, A. niger CBS 513.88 affected M. ruber M7 to produce cleistothecia on CYA media. Since cleistothecia are the sexual reproductive organ of M. ruber M7 (Ojeda-Lopez et al., 2018), it is speculated that A. niger CBS 513.88 might change the proliferation of M. ruber M7 under nutrient deficiency conditions. When using PDA as a culture media, there was no significant differences between monocultured M. ruber M7 and M. pilosus MS-1 on micromorphologic characterics. However, by using CYA and G25N as culture media, A. niger can affect the mycelia of M. pilosus MS-1 significantly, from straight to curved. The recent research shows that A. niger might be stimulated by carbon starving and produce a variety of pectinases which can break cell walls (van Munster et al., 2014). Therefore, considering CYA and G25N are all carbon-starved culture media, it is likely that A. niger can be forced to produce pectinase to achieve more carbon resources, meanwhile, the cell wall of M. pilosus MS-1 was broken by pectinase and caused a change of mycelia shape (Figure 6B). No significant change was observed by conventional cocultivation of A. niger, while on DSPD, the color of the communicating area of A. niger was darker than other areas in the colony. Research shows that the black color (melanin) of A. niger originated from spores (Kumaran et al., 2017). Monascus spp. strains may promote the production of the spores of A. niger. The molecular mechanisms about this phenomenon are still being studied.
By conventional cocultivation of fungi, the mycelia are very hard to separate and harvest if there is no antagonism phenomenon between the tested strains, which is an obstacle for SM analysis in this study. DSPD overcame this obstacle efficiently. From MK and CIT results (Table 1), we speculated that A. niger may affect the biosynthesis of MK and CIT produced by Monascus spp. These results can guide us on transcriptome analysis. With the help of DSPD, more intuitive and completed Monascus spp. colony pictures demonstrated the effects of A. niger on the MPs production. The above conclusions were further illustrated by the detection of color value and the HPLC analysis of MPs. Both tested A. niger can increase the MPs production of M. ruber M7, especially the orange pigments. Among them, on increasing MPs of M. ruber M7, A. niger CBS 513.88 was more efficient (Figure 7). The ultraviolet spectrograms and retention time of MPs peaks from HPLC results were compared to the published research about MPs (Chen et al., 2017), which have demonstrated that A. niger can cause variation of several known MPs contents. It was reported that the cocultivation of yeast and Monascus spp. and MPs production increased significantly (Shin et al., 1998). This may provide a reference to study the molecular mechanisms about MPs variation. Research has shown that CIT synthesis is related to MPs synthesis (He and Cox, 2016), which may explain the effects of A. niger CBS 513.88 on changing the intracellular and extracellular CIT content of M. ruber M7.
Research on SMs determination has shown that by using PDA, MEA, CYA, and G25N, no new peak can be observed by HPLC which represents a new compound. This study thus mined novel SMs by other attempts, changing culture temperatures, culture media, and extraction methods. When the rice was taken as a culture media, at 420 nm, two new compounds, namely substance A and B. The chemical structure of A and B are still being identified by LCMS-MS and NMR and will be brought out in the future. DSPD offered an effective method for sample preparation, which is necessary for the preparation and purification of substances A and B. In addition, with the help of DSPD, substance A was almost verified to be produced by M. ruber M7 when co-cultured with A. niger CBS 513.88. Surprisingly, a peak whose retention time and UV-spectrum were a similar yellow MP, identified by our previous work (Chen et al., 2017), was detected in A. niger CBS 513.88 mycelia, indicating that M. ruber M7 may stimulate A. niger CBS 513.88 to produce MP. These results offer important information for follow-up research. The related experiments on the identification of these new compounds and transcriptome analysis are still ongoing and will be presented in future studies.
To study the most natural co-cultured method of Monascus spp. and A. niger, which is growing on rice together in a way similar to BSRK, sample preparation represented a huge challenge for transcriptome analyses, due to the separation problems. Monascus spp. can grow together with A. niger on rice, the mycelia and spores of both sides are intertwined, and difficult to separate from each other. The tested strains were rooted in rice, which made it difficult to separate strains from rice. Rice contains proteins and saccharides which would interfere with the transcriptome sequencing results. The DSPD presented a breakthrough for overcoming this problem. Out of the tested strains, M. ruber M7 and A. niger CBS 513.88 were chosen as the advisable research subjects for further transcriptome analysis after comprehensive consideration according to the sequenced genome, based on similar growth rate and SMs richness. DSPD broke down the first barrier of transcriptome analysis. All the separation problems about strain-strains and strain-media were solved and good quality transcriptome data were obtained (Tables 4, 5).
This article reveals some promising superficial results relating to Monascus spp. and A. niger cocultivation that are worth further exploration. Although we choose M. ruber M7 and A. niger CBS 513.88 as the main research subjects for some reasonable causes, the results achieved by the DSPD showed that the different pairs of A. niger and Monascus spp. strains had different interactions, which indicates that it is important to choose appropriate strains for cocultivation to increase beneficial metabolites, decrease detrimental ones, or even find new substances. Beyond the two new substances found by the SMs profiles of M. ruber M7 when co-cultured with A. niger CBS 513.88, there still may be more undiscovered compounds that were in this case limited by the culture conditions, extraction methods, and instrumental methods. The MPs only changed on yields and not in terms of kind, and the reasons for this on a molecular level are also worth studying in the future. For transcriptome analyses, the present study overcomes the barrier of sample preparation, opening further avenues for the study of the molecular mechanisms of M. ruber M7 and A. niger CBS 513.88, and obtaining good quality transcriptome results. The DSPD developed in the present study presents an excellent stepping-stone to success. All the above conclusions obtained by DSPD provide the theoretical foundations for further study of the molecular mechanisms between Monascus spp. and A. niger.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: NCBI Sequence Read Archive (SRA) with BioProject number PRJNA721011.
Author Contributions
XY and FC conceived the study and designed the experiments. XY carried out the experiments, analyzed the data, wrote the manuscript, undertook investigation, methodology, writing, reviewing, editing, and supervision. FC reviewed the manuscript and the experiments, oversaw conceptualization, and project administration. Both authors contributed to the article and approved the submitted manuscript.
Funding
This work was supported by the Major Program of the National Natural Science Foundation of China (Nos. 31730068 and 31330059 to FC), the National Key Research and Development Program of China (No. 2018YFD0400404 to FC), the Shandong Province Taishan Industry Leading Talents High-Efficiency Agriculture Innovation Project (No. tscy20180120 to FC), and Project supported by the Fundamental Research Funds for the Central Universities of China (No. 2662018JC013).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
- ^ https://genome.jgi.doe.gov/portal/pages/dynamicOrganismDownload.jsf?organism=Monpu1
- ^ http://www.aspergillusgenome.org/cache/A_niger_CBS_513_88_genomeSnapshot.html
References
Andersen, M. R., Salazar, M. P., Schaap, P. J., Van De Vondervoort, P. J., Culley, D., Thykaer, J., et al. (2011). Comparative genomics of citric-acid-producing Aspergillus niger ATCC 1015 versus enzyme-producing CBS 513.88. Genome Res. 21, 885–897. doi: 10.1101/gr.112169.110
Bertrand, S., Schumpp, O., Bohni, N., Monod, M., Gindro, K., and Wolfender, J. L. (2013). De novo production of metabolites by fungal co-culture of Trichophyton rubrum and Bionectria ochroleuca. J. Nat. Prod. 76, 1157–1165. doi: 10.1021/np400258f
Camacho-Zaragoza, J. M., Hernandez-Chavez, G., Moreno-Avitia, F., Ramirez-Iniguez, R., Martinez, A., Bolivar, F., et al. (2016). Engineering of a microbial coculture of Escherichia coli strains for the biosynthesis of resveratrol. Microb. Cell Fact. 15:163. doi: 10.1186/s12934-016-0562-z
Chen, F., Li, L., Qu, J., and Chen, C. (2009). “Cereal vinegars made by solid-state fermentation in China,” in Vinegars of the world, eds L. Solieri and P. Giudici (Milano: Springer), 243–259.
Chen, W., Chen, R., Liu, Q., He, Y., He, K., Ding, X., et al. (2017). Orange, red, yellow: biosynthesis of azaphilone pigments in Monascus fungi. Chem. Sci. 8, 4917–4925. doi: 10.1039/c7sc00475c
Chen, W., He, Y., Zhou, Y., Shao, Y., Feng, Y., Li, M., et al. (2015). Edible filamentous fungi from the species Monascus: early traditional fermentations, modern molecular biology, and future genomics. Compr. Rev. Food Sci. Food Saf. 14, 555–567.
Chiang, Y., Chang, S., Oakley, B. R., and Wang, C. C. (2011). Recent advances in awakening silent biosynthetic gene clusters and linking orphan clusters to natural products in microorganisms. Curr. Opin. Chem. Biol. 15, 137–143. doi: 10.1016/j.cbpa.2010.10.011
Ding, G., Zhao, J., Zhang, W., Yao, J. C., Xu, H., Ding, Y. Z., et al. (2008). National Standard GB4926-2008: Food Additive-Red Kojic Rice (Powder) AQSIQ. China. 1–5.
Fang, S., Yang, Y., Zhen, D., and Chen, M. (2011). Production of koji by black-skin-red-koji. Adv. Mater. Res. 236-238, 2748–2752.
Favela-Torres, E., Cordova-López, J., Garda-Rivero, M., and Gutirrrez-Rojas, M. (1998). Kinetics of growth of Aspergillus niger during submerged, agar surface and solid state fermentations. Process Biochem. 33, 103–107.
Feng, Y., Chen, W., and Chen, F. (2016). A Monascus pilosus MS-1 strain with high-yield monacolin K but no citrinin. Food Sci. Biotechnol. 25, 1115–1122. doi: 10.1007/s10068-016-0179-3
Feng, Y., Shao, Y., and Chen, F. (2012). Monascus pigments. Appl. Microbiol. Biotechnol. 96, 1421–1440.
Feng, Y., Shao, Y., Zhou, Y., and Chen, F. (2014). Monacolin K production by citrinin-free Monascus pilosus MS-1 and fermentation process monitoring. Eng. Life Sci. 14, 538–545.
Gao, G., Fan, H., Zhang, Y., Cao, Y., Li, T., Qiao, W., et al. (2021). Production of nisin-containing bacterial cellulose nanomaterials with antimicrobial properties through co-culturing Enterobacter sp. FY-07 and Lactococcus lactis N8. Carbohydr. Polym. 251:117131. doi: 10.1016/j.carbpol.2020.117131
He, Y., and Cox, R. J. (2016). The molecular steps of citrinin biosynthesis in fungi. Chem. Sci. 7, 2119–2127.
Huang, L., Chen, K., Zhang, M., Wang, J., Ren, X., Yang, H., et al. (2019). Comparative analysis of physicochemical components and volatile components in rice wines brewed by Aspergillus niger and Monascus purpureus in Chinese. J. Food Saf. Qual. 10, 4896–4904.
Huang, Z., Hong, J., Xu, J., Li, L., Guo, W., Pan, Y., et al. (2018). Exploring core functional microbiota responsible for the production of volatile flavour during the traditional brewing of Wuyi Hong Qu glutinous rice wine. Food Microbiol. 76, 487–496. doi: 10.1016/j.fm.2018.07.014
Jimenez-Barrera, D., Chan-Cupul, W., Fan, Z., and Osuna-Castro, J. A. (2018). Fungal co-culture increases ligninolytic enzyme activities: statistical optimization using response surface methodology. Prep. Biochem. Biotechnol. 48, 787–798. doi: 10.1080/10826068.2018.1509084
Kumaran, S., Abdelhamid, H. N., and Wu, H. (2017). Quantification analysis of protein and mycelium contents upon inhibition of melanin for Aspergillus niger: a study of matrix assisted laser desorption/ionization mass spectrometry (MALDI-MS). RSC Adv. 7, 30289–30294.
Kumari, S., and Naraian, R. (2016). Decolorization of synthetic brilliant green carpet industry dye through fungal co-culture technology. J. Environ. Manag. 180, 172–179. doi: 10.1016/j.jenvman.2016.04.060
Li, K., Chen, M., and Zhu, Z. (2009). Production of koji by black-skin-red-koji. China Brewing 4, 124–126.
Li, L., Chen, S., Gao, M., Ding, B., Zhang, J., Zhou, Y., et al. (2019). Acidic conditions induce the accumulation of orange Monascus pigments during liquid-state fermentation of Monascus ruber M7. Appl. Microbiol. Biotechnol. 103, 8393–8402. doi: 10.1007/s00253-019-10114-8
Liu, Q., Cai, L., Shao, Y., Zhou, Y., Li, M., Wang, X., et al. (2016). Inactivation of the global regulator LaeA in Monascus ruber results in a species-dependent response in sporulation and secondary metabolism. Fungal Biol. 120, 297–305. doi: 10.1016/j.funbio.2015.10.008
Mao, Q., Tu, X., Jia, X., and Zhou, L. (2011). The fermentation technology of black-skin-red-koji and the brewing of indica rice wine. Jiangsu Condiment Subsidiary Food 28, 28–31.
Marmann, A., Aly, A. H., Lin, W., Wang, B., and Proksch, P. (2014). Co-cultivation–a powerful emerging tool for enhancing the chemical diversity of microorganisms. Mar. Drugs 12, 1043–1065. doi: 10.3390/md12021043
Meyer, V., Andersen, M. R., Brakhage, A. A., Braus, G. H., Caddick, M. X., Cairns, T. C., et al. (2016). Current challenges of research on filamentous fungi in relation to human welfare and a sustainable bio-economy: a white paper. Fungal Biol. Biotechnol. 3:6. doi: 10.1186/s40694-016-0024-8
Moussa, M., Ebrahim, W., Bonus, M., Gohlke, H., M’Andi, A., Kurt’An, T., et al. (2019). Co-culture of the fungus Fusarium tricinctum with Streptomyces lividans induces production of cryptic naphthoquinone dimers. RSC Adv. 9, 1491–1500.
Nett, M., Ikeda, H., and Moore, B. S. (2009). Genomic basis for natural product biosynthetic diversity in the actinomycetes. Nat. Prod. Rep. 26, 1362–1384.
Nielsen, K. F., Mogensen, J. M., Johansen, M., Larsen, T. O., and Frisvad, J. C. (2009). Review of secondary metabolites and mycotoxins from the Aspergillus niger group. Anal. Bioanal. Chem. 395, 1225–1242. doi: 10.1007/s00216-009-3081-5
Nonaka, K., Iwatsuki, M., Horiuchi, S., Shiomi, K., Ômura, S., and Masuma, R. (2015). Induced production of BE-31405 by co-culturing of Talaromyces siamensis FKA-61 with a variety of fungal strains. J. Antibiot. 68, 573–578.
Ojeda-Lopez, M., Chen, W., Eagle, C. E., Gutierrez, G., Jia, W. L., Swilaiman, S. S., et al. (2018). Evolution of asexual and sexual reproduction in the aspergilli. Stud. Mycol. 91, 37–59.
Pettit, R. K. (2009). Mixed fermentation for natural product drug discovery. Appl. Microbiol. Biotechnol. 83, 19–25.
Shang, Z., Salim, A. A., and Capon, R. J. (2017). Chaunopyran A: co-Cultivation of marine mollusk-derived fungi activates a rare class of 2-alkenyl-tetrahydropyran. J. Natural Prod. 80, 1167–1172. doi: 10.1021/acs.jnatprod.7b00144
Shin, C. S., Kim, H. J., Kim, M. J., and Ju, J. Y. (1998). Morphological change and enhanced pigment production of Monascus when cocultured with Saccharomyces cerevisiae or Aspergillus oryzae. Biotechnol. Bioeng. 59, 576–581. doi: 10.1002/(sici)1097-0290(19980905)59:5<576::aid-bit7>3.0.co;2-7
Show, P. L., Oladele, K. O., Siew, Q. Y., Aziz Zakry, F. A., Lan, J. C.-W., and Ling, T. C. (2015). Overview of citric acid production from Aspergillus niger. Front. Life Sci. 8:271–283. doi: 10.1080/21553769.2015.1033653
van Munster, J. M., Daly, P., Delmas, S., Pullan, S. T., Blythe, M. J., Malla, S., et al. (2014). The role of carbon starvation in the induction of enzymes that degrade plant-derived carbohydrates in Aspergillus niger. Fungal Genet. Biol. 72, 34–47. doi: 10.1016/j.fgb.2014.04.006
Vinale, F., Nicoletti, R., Borrelli, F., Mangoni, A., Parisi, O. A., Marra, R., et al. (2017). Co-culture of plant beneficial microbes as source of bioactive metabolites. Sci. Rep. 7:14330. doi: 10.1038/s41598-017-14569-5
Wakefield, J., Hassan, H. M., Jaspars, M., Ebel, R., and Rateb, M. E. (2017). Dual induction of new microbial secondary metabolites by fungal bacterial co-cultivation. Front. Microbiol. 8:1284. doi: 10.3389/fmicb.2017.01284
Wang, L. W., Dai, Y., Chen, W., Shao, Y., and Chen, F. (2016). Effects of light intensity and color on the biomass, extracellular red pigment, and citrinin production of Monascus ruber. J. Agric. Food Chem. 64, 9506–9514. doi: 10.1021/acs.jafc.6b04056
Xu, D., Wang, L., and Du, C. (2015). Progress in microbial co-culture -A review. Acta Microbiol. Sin. 55, 1089–1096.
Xu, M., Bai, H. Y., Fu, W. Q., Sun, K., Wang, H. W., Xu, D. L., et al. (2021). Endophytic bacteria promote the quality of Lyophyllum decastes by improving non-volatile taste components of mycelia. Food Chem. 336, 127672. doi: 10.1016/j.foodchem.2020.127672
Xu, T., Li, J., Yu, B., Liu, L., Zhang, X., Liu, J., et al. (2018). Transcription factor SsSte12 was involved in mycelium growth and development in Sclerotinia sclerotiorum. Front. Microbiol. 9:2476. doi: 10.3389/fmicb.2018.02476
Yan, P., Xue, W., Tan, S., Zhang, H., and Chang, X. (2008). Effect of inoculating lactic acid bacteria starter cultures on the nitrite concentration of fermenting Chinese paocai. Food Control 19, 50–55.
Yu, G., Sun, Z., Peng, J., Zhu, M., Che, Q., Zhang, G., et al. (2019). Secondary metabolites produced by combined culture of Penicillium crustosum and a Xylaria sp. J. Natural Prod. 82, 2013–2017. doi: 10.1021/acs.jnatprod.9b00345
Yu, M., Li, Y., Banakar, S. P., Liu, L., Shao, C., Li, Z., et al. (2019). New metabolites from the co-culture of marine-derived actinomycete Streptomyces rochei MB037 and Fungus Rhinocladiella similis 35. Front. Microbiol. 10:915. doi: 10.3389/fmicb.2019.00915
Zhang, H., Li, Z., Pereira, B., and Stephanopoulos, G. (2015). Engineering E. coli-E. coli cocultures for production of muconic acid from glycerol. Microb. Cell Fact. 14:134. doi: 10.1186/s12934-015-0319-0
Keywords: Aspergillus niger, cocultivation, HPLC, transcriptome, Monascus spp., double-sided petri dish
Citation: Yuan X and Chen F (2021) Cocultivation Study of Monascus spp. and Aspergillus niger Inspired From Black-Skin-Red-Koji by a Double-Sided Petri Dish. Front. Microbiol. 12:670684. doi: 10.3389/fmicb.2021.670684
Received: 22 February 2021; Accepted: 12 April 2021;
Published: 09 June 2021.
Edited by:
Van-Tuan Tran, Vietnam National University, VietnamReviewed by:
Laurent Dufossé, Université de la Réunion, FranceZhilong Wang, Shanghai Jiao Tong University, China
Lourdes Morales-Oyervides, Universidad Autónoma de Coahuila, Mexico
Copyright © 2021 Yuan and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fusheng Chen, Y2hlbmZzQG1haWwuaHphdS5lZHUuY24=