 Rafael Ayerbe-Algaba1,2
Rafael Ayerbe-Algaba1,2 Nuria Bayó3Ester Verdú3Raquel Parra-Millán1,2Jesús Seco3
Nuria Bayó3Ester Verdú3Raquel Parra-Millán1,2Jesús Seco3 Meritxell Teixidó3
Meritxell Teixidó3 Jerónimo Pachón2,4*
Jerónimo Pachón2,4* Ernest Giralt3,5
Ernest Giralt3,5 Younes Smani1,2*
Younes Smani1,2*- 1Clinical Unit of Infectious Diseases, Microbiology and Preventive Medicine, University Hospital Virgen del Rocío, Seville, Spain
- 2Institute of Biomedicine of Seville (IBiS), University Hospital Virgen del Rocío/Spanish National Research Council (CSIC)/University of Seville, Seville, Spain
- 3Institute for Research in Biomedicine (IRB Barcelona), Barcelona Institute for Science and Technology (BIST), Barcelona, Spain
- 4Department of Medicine, University of Seville, Seville, Spain
- 5Department of Inorganic and Organic Chemistry, University of Barcelona, Barcelona, Spain
Previously, we identified that a cyclic hexapeptide AOA-2 inhibited the interaction of Gram-negative bacilli (GNB) like Acinetobacter baumannii, Pseudomonas aeruginosa, and Escherichia coli to host cells thereby preventing the development of infection in vitro and in a murine sepsis peritoneal model. In this work, we aimed to evaluate in vitro a library of AOA-2 derivatives in order to improve the effect of AOA-2 against GNB infections. Ten AOA-2 derivatives were synthetized for the in vitro assays. Their toxicities to human lung epithelial cells (A549 cells) for 24 h were evaluated by determining the A549 cells viability using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. The effect of these peptide derivatives and AOA-2 at 250, 125, 62.5, and 31.25 μg/mL on the attachment of A. baumannii ATCC 17978, P. aeruginosa PAO1 and E. coli ATCC 25922 strains to A549 cells was characterized by adherence and viability assays. None of the 10 derivatives showed toxicity to A549 cells. RW01 and RW06 have reduced more the adherence of ATCC 17978, PAO1 and ATCC 2599 strains to A549 cells when compared with the original compound AOA-2. Moreover, both peptides have increased slightly the viability of infected A549 cells by PAO1 and ATCC 25922 than those observed with AOA-2. Finally, RW01 and RW06 have potentiated the activity of colistin against ATCC 17978 strain in the same level with AOA-2. The optimization program of AOA-2 has generated two derivatives (RW01 and RW06) with best effect against interaction of GNB with host cells, specifically against P. aeruginosa and E. coli.
Introduction
Multidrug resistance (MDR) in Gram-negative bacilli (GNB) is a current threat in global health that requires effective and urgent solutions, being a global priority for WHO (Tacconelli et al., 2018). The success of GNB as a community and nosocomial pathogen is attributed to its resistant to several antibiotic families. Whole-genome sequencing studies involving the GNB have demonstrated not only a vast array of antibiotic drug resistance determinants but also many pathogenicity islands (Kaper et al., 2004; Smith M. G. et al., 2007).
Studies of specific virulence mechanisms have demonstrated the contribution of outer membrane protein A (OmpA), as well as pili, fimbriae, outer membrane proteins and various secretion systems in the pathogenesis of GNB in vitro and in animal experimental models (Weiser and Gotschlich, 1991; Prasadarao et al., 1996; Wu et al., 2003; Teng et al., 2006; Mittal et al., 2011; Sánchez-Encinales et al., 2017; Nicholson et al., 2019).
OmpA is a β-barrel porin highly conserved among bacterial species, especially among Gram-negative organisms (Smith S. G. J. et al., 2007). This protein has a variety of interesting biological properties. It has been suggested to have involvement in the adherence to epithelia (Gaddy et al., 2009), translocation into the epithelial cell nucleus (Choi et al., 2008), induction of epithelial cell death (Choi et al., 2005), biofilm formation (Gaddy et al., 2009), and binding to factor H, which may allow A. baumannii to develop serum resistance (Kim et al., 2009).
Previous studies showed that A. baumannii and E. coli deficient in ompA reduced their pathogenecities in vitro and in vivo (Mittal et al., 2011; Sánchez-Encinales et al., 2017). Interestingly, we showed that the inhibition of OmpA by a cyclic hexapetide AOA-2 reduces bacterial dissemination between organs, development of pneumonia, and bacteremia and death in murine peritoneal sepsis model by A. baumannii, P. aeruginosa, and E. coli (Vila-Farrés et al., 2017). Moreover, AOA-2 has showed in vitro synergy with colistin which has presented a good therapeutic efficacy against colistin-susceptible and resistant A. baumannii through the overexpression of Omp25 (Parra-Millán et al., 2018). In the present study, we aimed to improve the efficacy of AOA-2 against GNB infections by optimizing the derivatives of AOA-2.
Materials and Methods
Materials for the Synthesis of Peptides
The protected amino acids and resins were supplied by Luxembourg Industries (Tel-Aviv, Israel), Neosystem (Strasbourg, France), Calbiochem-Novabiochem AG (Läufelfingen, Switzerland), PolyPeptides Labs (Torrance, CA, United States), Bachem AG (Bubendorf, Switzerland), and Iris Biotech (Marktredwitz, Germany). PyBOP was provided by Calbiochem-Novabiochem AG. Piperidine is obtained from SDS (Peypin, France); N, N-diisopropylethylamine (DIEA) was obtained from Merck (Darmstadt, Germany) and triisopropylsilane (TIS) and ninhydrin were from Fluka Chemika (Buchs, Switzerland). HOAt was acquired from GL Biochem Shanghai Ltd. (Shanghai, China). Solvents for the synthesis of peptides and RP-HPLC [dimethylformamide (DMF), dichloromethane (DCM) and acetonitrile (MeCN)] were from Scharlau or SDS (Barcelona, Spain). Trifluoroacetic acid (TFA) was purchased from Kali Chemie (Bad Wimpfen, Germany). The other chemicals used were from Aldrich (Milwaukee, WI, United States) and were of the highest commercially available purity.
Synthesis of Peptides
The peptides were synthesized on a 2-Chlorotrythyl chloride resin by solid-phase peptide synthesis using the 9-fluorenylmethoxycarbonyl/tert-butyl (Fmoc/tBu) strategy. The amino acids protected with Nα -Fmoc (3 eq)/HOAt (3 eq), PyBOP (4 eq), and DIEA (6 eq) were used for the couplings. The Fmoc protecting group was removed by treatment with a 20% piperidine solution in DMF. The peptides were cleaved using 2% TFA in DCM. For the cyclization step, the solvent used was DCM/DMF (98:2), PyAOP (2 eq) was dissolved in DMF and the peptide (5 mM) in DCM and once mixed in the appropriate proportions, 6 eq of DIEA was added, the reaction was completed in 2 or 3 h. After the cyclization carried out, deprotection of the side chains was carried out by TFA/TIS/H2O (95:2.5:2.5). Peptides were analyzed at λ = 220 nm by HPLC [Waters Alliance 2695 separation module equipped with a 2998 photodiode array detector, Sunfire C18 column (100 mm × 4.6 mm × 3.5 mm, 100 A, Waters)], and Empower software; flow rate = 1 ml/min. Peptides were then purified by semi-preparative HPLC [Waters 2700 Sample Manager equipped with a double absorbance detector λ Waters 2487, a Waters 600 controller, a Waters fraction collection II, a Symmetry C18 column (100 mm × 30 mm, 5 mm, 100 Å, Waters) and the Millenium chromatography manager software]. Flow rate = 15 mL/min; solvents: A = 0.1% trifluoroacetic acid in water, and B = 0.05% trifluoroacetic acid in acetonitrile. Peptides are characterized by MALDI-TOF mass spectrometry (Voyager-DE RP MALDI-TOF, PE Biosystems with a N2-laser of 337 nm) and a high-resolution ESI-MS model (LTQ-FT Ultra, Thermo Fisher Scientific).
Docking and Molecular Modeling
All molecular docking calculations, and preparation of protein and ligand systems were performed using Molecular Operating Environment (MOE; Chemical Computing Group, Quebec, Canada) software. Protein structure of the transmembrane domain of A. baumannii OmpA was obtained by homology modeling using Comparative Modeling with Rosetta (RosettaCM) (Song et al., 2013) using as template PDBs obtained by NMR (2GE4, 1G90) and crystallography (1BXW, 1QJP). Best-ranked homology models were minimized and further relaxed including short (2 ns) explicit solvent molecular dynamics (MD) simulations using Amber99 force field. To identify druggable pockets of interest and obtain a pharmacophoric description of the binding site, a modification of the original MDMix method (Seco et al., 2009) was applied. Briefly, the original isopropanol probe was replaced by alternative probe molecules compatible with the sidechain peptide chemistry, including as probes metylammonium, benzene and propane. The pharmacophore coordinates were profiled in terms of binding energetics and clustered to delimit the docking grid box extension and simultaneously, guide docking exploration toward compatible solutions. The process of pharmacophore identification and clustering was conducted with an in-house Python script. A library of cyclic hexapeptides was used for virtual screening. All initial cyclic peptide structures were subjected to conformational exploration (Chen and Foloppe, 2008) by using MOE, clustering the low-energy conformations with a backbone RMSD threshold of 0.25 units to yield a pool of conformations ranging between 10 and 25 per cyclic peptide structure. All generated conformations were docked using a hierarchical docking protocol using rDock (Ruiz-Carmona et al., 2014) and results were re-scored with AutoDock Vina.
Bacterial Strains
Three reference bacterial strains acquired from American Type Culture Collection (LGC, United Kingdom) were used: (a) A. baumannii ATCC 17978 (Baumann et al., 1968), (b) P. aeruginosa PAO1 (Holloway, 1955), and (c) E. coli ATCC 25922 (Reading and Cole, 1977). An OmpA-deficient mutant A. baumannii, JPAB01, was used (Parra-Millán et al., 2018).
Cell Culture and Infection
The human pneumocyte cell line type II A549, derived from a human lung carcinoma, obtained from the American Type Culture Collection was used and cultured in DMEM medium (Invitrogen, Spain) supplemented with 10% of heat-inactivated fetal bovine serum, 50 μg/mL vancomycin (Laboratorios Normon, Spain), 20 μg/mL gentamicin (Normon Laboratories, Spain), 0.25 μg/mL amphotericin B mL (Invitrogen, Spain) and 1% HEPES (Invitrogen, Spain) in a humidified incubator, 5% CO2 at 37°C. Routine passage of A549 cells was performed every 3–4 days. Cells were seeded 24 h in 96-well plates (Sarstedt, Germany) for the cell viability assay, and in 24-well plates (Sarstedt, Germany) for the bacterial adhesion assays.
In vitro Toxicity of Peptides
A549 cells were incubated with the peptide derivatives RW01, RW02, RW03, RW06, RW07, RW08, RW09, RW10, RW11, and RW13 (0, 3.5, 7, 15, 35, 62, 125, 250, and 500 μg/mL) for 24 h with 5% CO2 at 37°C. Prior the evaluation of the peptides cytotoxicity, A549 cells were washed three times with prewarmed PBS 1X. Subsequently, quantitative cytotoxicity was evaluated by measuring the mitochondrial reduction activity using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as described previously (Smani et al., 2013). The percentage of cytotoxicity was calculated from the absorbance at 570 nm as follow: [(Absorbance 570 nm of treated cells/Absorbance 570 nm mean of untreated cells) × 100].
Bacterial Adhesion Assay
A. baumannii ATCC 17978, P. aeruginosa PAO1 and E. coli ATCC 25922 strains were incubated with AOA-2 (31.25, 62.5, 125, and 250 μg/mL, 30 min), AOA-2 derivatives RW01, RW02, RW03, RW06, RW07, RW08, RW09, RW10, RW11, and RW13 (250 μg/mL, 30 min), or AOA-2 derivatives RW01, RW02, RW03, RW06 (31.25, 62.5, and 125 μg/mL, 30 min), and added at 108 CFU/mL, corresponding to a multiplicity of infection (MOI) of 500, to the A549 cells for 2 h at 5% CO2 and 37°C. Subsequently, infected A549 cells were washed five times with prewarmed PBS 1X and lysed with 0.5% Triton X-100. Serial dilutions of these lysates were made and plated on blood agar plates (Blood-Agar Columbia, Becton Dickinson Microbiology Systems, United States) and incubated at 37°C for 24 h for colony counting and then determination of CFU/mL that attached to A549 cells.
Bacterial Cytotoxicity
A549 cells were infected with 108 CFU/mL, corresponding to a MOI of 500, of A. baumannii ATCC 17978, P. aeruginosa PAO1 and E. coli ATCC 25922 strains pretreated with AOA-2 or AOA-2 derivatives RW01, RW02, RW03, and RW06 (31.25, 62.5, and 125 μg/mL, 30 min) for 24 h with 5% CO2 at 37°C. Prior to the evaluation of bacterial cytotoxicity, viable bacteria are first removed from A549 cell culture by washing these A549 cells five times with PBS 1X. Subsequently, cell viability was evaluated as described above.
In vitro Susceptibility Testing
The minimal inhibitory concentration (MIC) of colistin in absence and presence of RW01 or RW06 at 125μμg/mL for A. baumannii ATCC 17978 strain was determined by microdilution assay in two independent experiments as previously reported (Parra-Millán et al., 2018).
Statistical Analysis
Group data are presented as mean ± SEM. Student t-test was used to determine differences between means. Differences were considered significant at P < 0.05. The SPSS (version 23.0) statistical package was used (SPSS Inc.).
Results
AOA-2 Derivatives Synthesis
The optimization of the AOA-2 peptide was carried out through an iterative computational design process. This computational design was based on the use of docking techniques with flexible side chains and molecular dynamics, using the data on the interaction of AOA-2 and OmpA in detailed (Vila-Farrés et al., 2017). Previously, an exhaustive exploration of its accessible conformational space was made for each molecule. All the interaction calculations were carried out thanks to the access of the “Mare Nostrum” supercomputer at the Barcelona Supercomputer Center (BSC). Thus, a total of 10 peptides derived from the lead compound, AOA-2 were synthesized (Table 1).
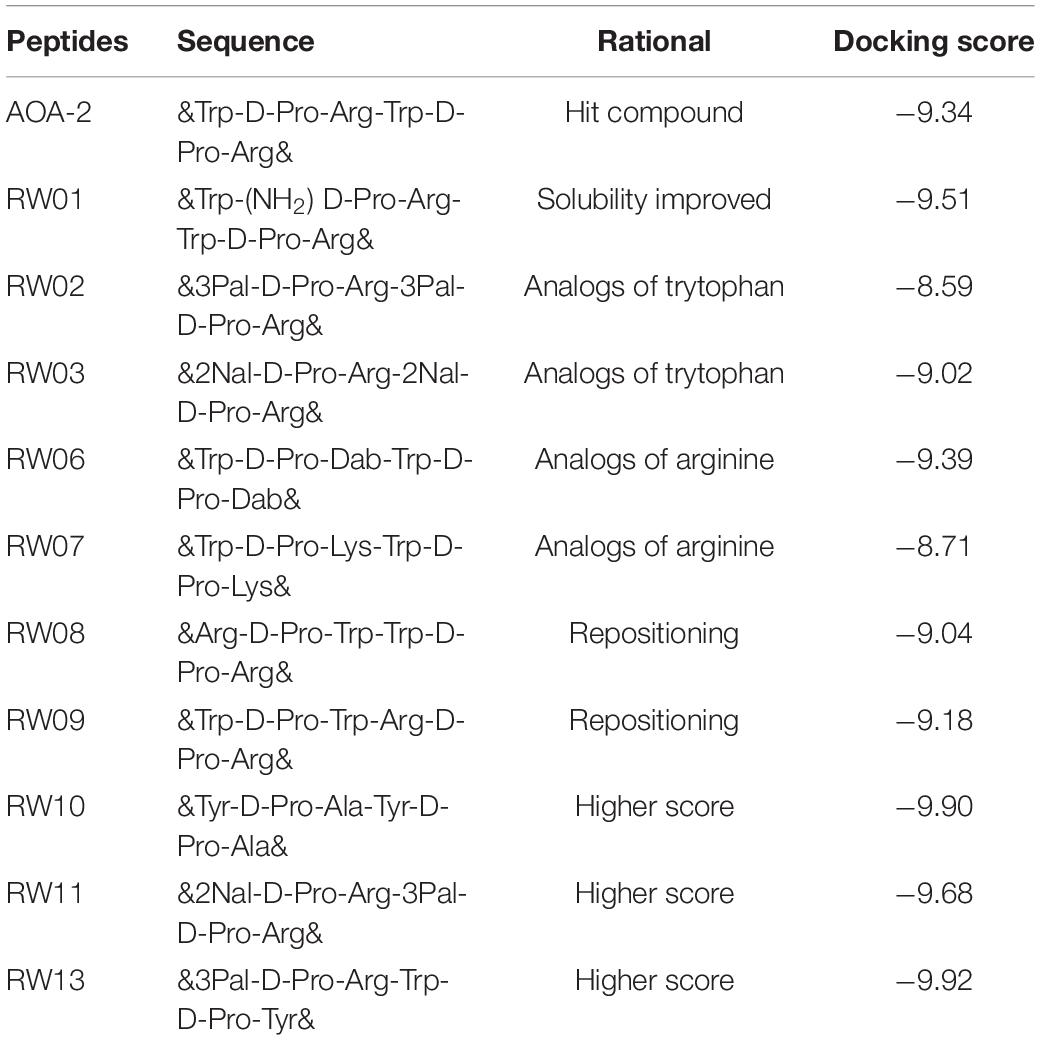
Table 1. Peptide sequences of the AOA-2 and its derivatives, and their docking score.
Different changes were carried out to optimize the lead compound. In RW01 solubility was enhanced by the addition of NH2 group. In RW02 and RW03 tryptophan analogs [3Pal: beta-(3-pyridyl)-alanine, and 2Nal: beta-(2-naphtyl)-alanine] were added. In RW06 and RW07 arginine analogs (Dab: 2,4-Diaminobutanoic acid, and lysine) were added. In RW08 and RW09 a repositioning of the hit sequence were made. The remaining peptides RW10, RW11, and RW13 showed a higher docking score than the AOA-2 peptide (Table 1 and Figure 1).

Figure 1. Structure of AOA-2 and its optimized derivatives.
In vitro Toxicity of AOA-2 Derivatives
The study of the toxicity of the synthesized compounds was carried out. The percentage of cell viability on the A549 epithelial cells incubated for 24 h with the different peptides at decreasing concentrations was determined from 500 to 3.5 μg/mL. None of the studied compounds showed cytotoxicity at the different concentrations used, being the cellular viability is between 94 and 100% (Table 2).
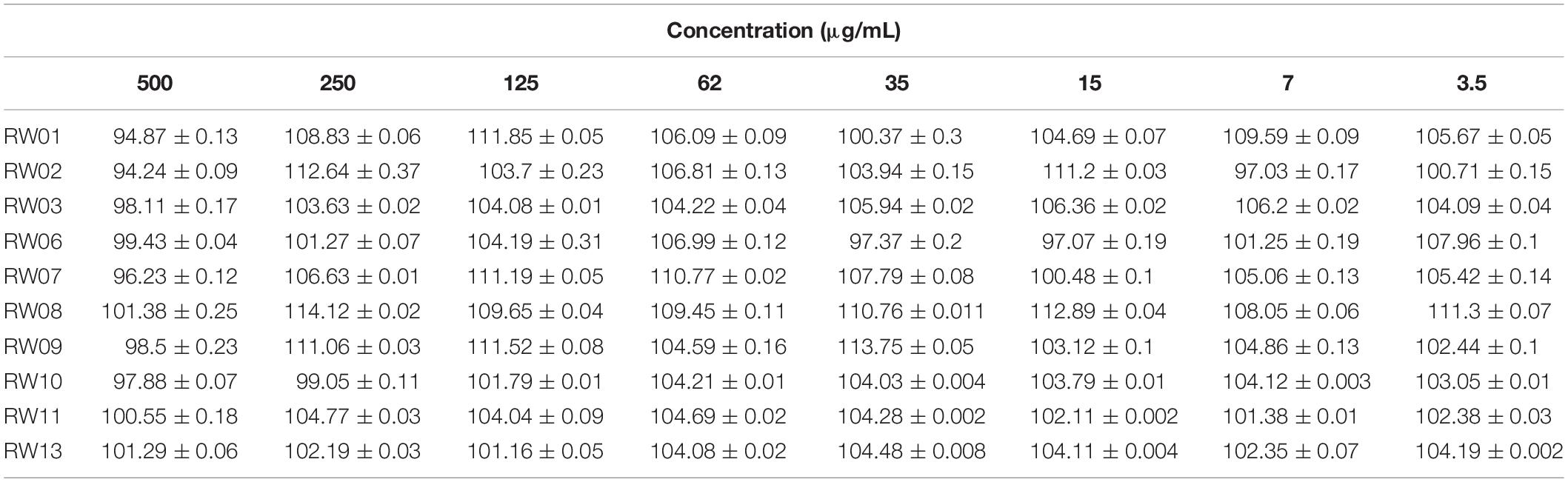
Table 2. Percentages of A549 cellular toxicity assessed through mitochondrial reduction activity (MTT assay) in presence AOA-2 derivatives. Data expressed as a percentage of cell viability.
In vitro Effect of AOA-2 Derivatives on A. baumannii, P. aeruginosa, E. coli Interaction With Host Cells
Bacterial Adhesion Assay
An initial stage of screening of the effect of all peptide derivatives and the original compound AOA-2 on adherence of A. baumannii ATCC 17978, P. aeruginosa PAO1 and E. coli ATCC 25922 strains to host cells was carried out. First we showed that AOA-2, RW01, RW02, RW03, and RW06 at 250 μg/mL reduced the adherence of A. baumannii to 36.62, 26.18, 43.58, 55.47, and 32.83%, respectively, the adherence of P. aeruginosa to 84.47, 40.08, 42.51, 20.71, and 31.14%, respectively and the adherence of E. coli to 65.05, 66.66, 61.11, 29.62, and 44.44%, respectively. In addition, we found that RW07 and RW13 reduced the adherence of PAO1 and ATCC 25922 strains to A549 cells; whereas, RW08 reduced the adherence of ATCC 17978 and PAO1 strains to A549 cells. In the case of RW09, RW10 and RW11, only the adherence of PAO1 strain to A549 cells was reduced (Figure 2A). From these data, we suggest that RW01, RW02, RW03 and RW06 present an inhibitory effect on the adherence of the three pathogens to A549 cells.
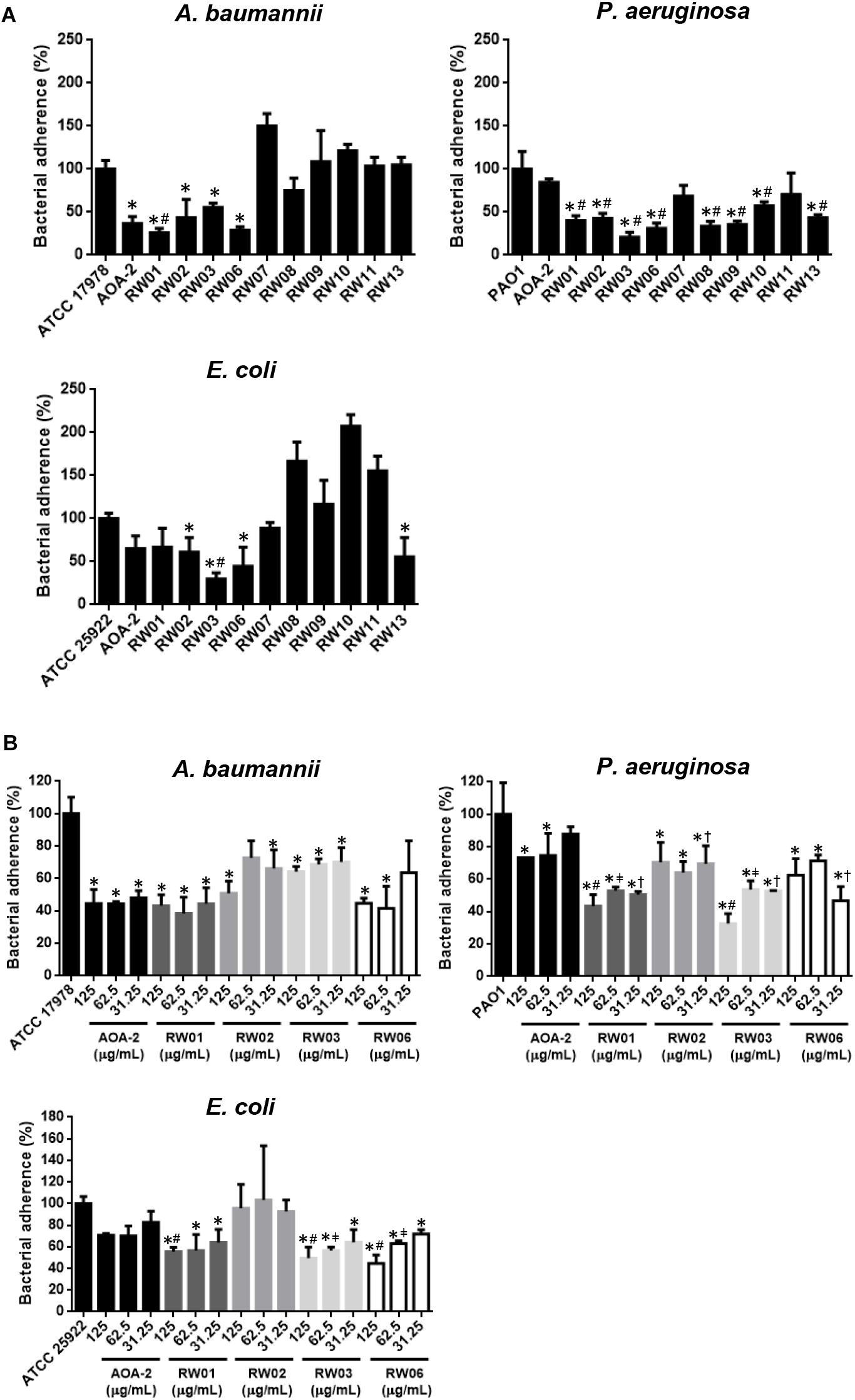
Figure 2. Bacterial adhesion assay of A. baumannii ATCC 17978, P. aeruginosa PAO1 and E. coli ATCC 25922 in presence of AOA-2 and its derivatives at concentration of 250 μg/mL (A), and 31.25, 62.5, and 125 μg/mL (B). ∗compared to the untreated bacteria, #compared to AOA-2 at 125 or 250 μg/mL, μcompared to AOA-2 at 62.5 μg/mL, †compared to AOA-2 at 31.25 μg/mL, Student’s t-test with P < 0.05.
Next, RW01, RW02, RW03, and RW06 were selected for the following studies, in which the reduction of bacterial adherence was studied again but at decreasing concentrations of these peptides (125, 62.5, and 31.25 μg/mL) and compared with those of the original peptide AOA-2 (Figure 2B). We found that the four peptides reduced better the adherence of PAO1 strain with some statistical differences when compared with AOA-2. While, RW01, RW03, and RW06 have reduced with some statistical differences in comparison with AOA-2 the adherence of ATCC 25922 to host cells. Against ATCC 17978 strain, only RW01 and RW06 have reduced non-statistically the adherence of ATCC 17978 to A549 cells when compared with AOA-2 (except for treatment with RW06 at 31.25 μg/mL). From these data, we suggest that RW01 and RW06 improve the effect of AOA-2 on the adherence of P. aeruginosa and E. coli to host cells, and with slight level against A. baumannii.
Cell Viability Assay
In order to confirm that AOA-2 derivatives have improved the effect of the original compound AOA-2 against A. baumannii, P. aeruginosa, and E. coli, only the effects of RW01 and RW06 vs. AOA-2 at 125, 62.5, and 31.25 μg/mL on cell viability in infected A549 cells were explored (Figure 3). Treatment of ATCC 17978, PAO1 and ATCC 25922 strains with AOA-2 at these concentrations did not increase significantly the cell viability with respect to untreated strains. Whereas, only the treatment of ATCC 17978 strain with RW01 or RW06 at 125 μg/mL increased significantly the cell viability with respect to untreated ATCC 17978 strain.
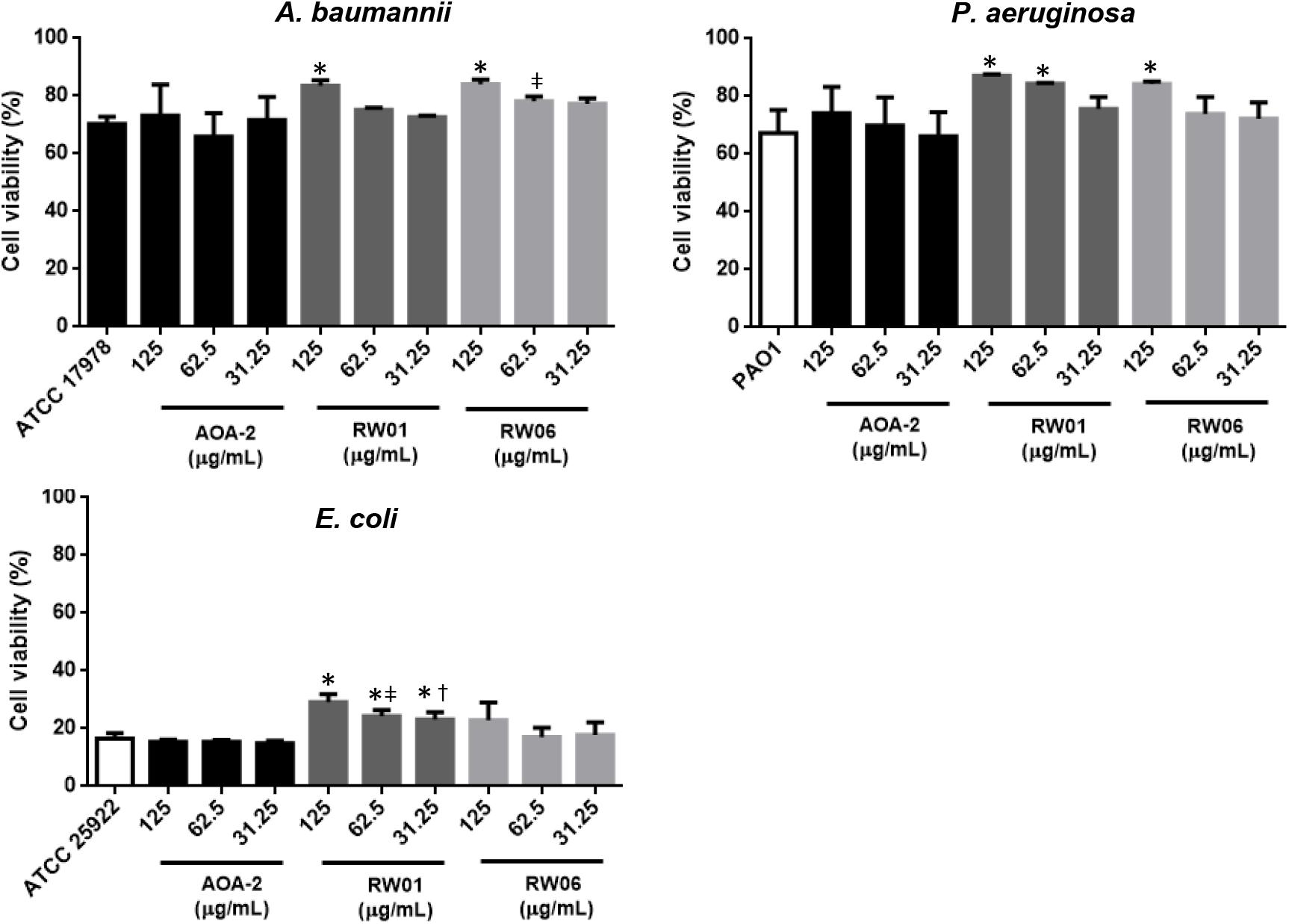
Figure 3. Cell viability assay of A. baumannii ATCC 17978, P. aeruginosa PAO1, and E. coli ATCC 25922 at different concentrations of AOA-2 and its derivatives RW01 and RW06. ∗compared to the untreated bacteria, μcompared to AOA-2 at 62.5 μg/mL, †compared to AOA-2 at 31.25 μg/mL, Student’s t-test with P < 0.05.
Similar results were observed with PAO1 and ATCC 25922 strains. Treatment of both strains with RW01 at the three concentrations increased significantly the cell viability with respect to untreated strains (except for PAO1 strain treatment with RW01 at 31.25 μg/mL). In contrast, treatment of both strains with RW06 (except for PAO1 treatment with RW06 at 125 at μg/mL) did not increase significantly the cell viability with respect to untreated strains.
Comparing the results of RW01 and RW06 with those of AOA-2, we found that only RW06 at 62.5 μg/mL has protected more significantly the cell death caused by ATCC 17978, whereas RW01 at 62.5 and 31.25 μg/mL has been more protective against ATCC 25922.
Synergistic Effect of AOA-2 Derivatives Against A. baumannii
We previously demonstrated that AOA-2 synergize with colistin against A. baumannii (Parra-Millán et al., 2018). Therefore, we thought it would be interesting to examine the capacity of RW01 and RW06 to potentiate the activity of colistin against A. baumannii ATCC 17978 strain. The addition of RW01 or RW06 at 125 μg/mL to ATCC 17978 strain increases the susceptibility of this strain to colistin from 0.5 to = 0.015 μg/mL. In contrast, the addition of RW01 or RW06 at 125 μg/mL to JPAB01 did not change the activity (MIC 0.0625 μg/mL) of colistin against this mutant strain. This last data suggest that RW01 and RW06 might act on OmpA of A. baumannii.
Discussion
Nowadays the threat of antibiotic resistance requires a multiple approach that includes better administration of known antibiotics and the development of new alternative strategies to treat or prevent bacterial infections.
This study focuses on the paradigm of interference in pathogenesis, that is, disarming pathogenic bacteria by neutralizing their virulence factors (Dickey et al., 2017). It was already demonstrated that a cyclic hexapeptide, AOA-2, was able to inhibit the interaction of GNB with host cells and consequently increased the viability of these host cells (Vila-Farrés et al., 2017). At the same time, in vivo administration of this compound showed a large decrease in bacterial load in tissues as well as a reduction in bacteremia, together with a significant decrease in mice mortality, especially in A. baumannii. In this scenario, the drug discovery, such as AOA-2, should be considered as an initial stage for the development of a new class of agents with anti-virulence capacity, like other studies that are currently in preclinical stages of development, such as the compound M64 developed against the virulence factor MvfR of P. aeruginosa (Starkey et al., 2014) or the compound 22, a byarilmannoside, against the FimK of uropathogenic E. coli (Cusumano et al., 2011; Jarvis et al., 2016).
The therapeutic efficacy of these virulence factors should also be further evaluated in combination with antimicrobial agents used in the clinical setting. For example, AOA-2 has presented good therapeutic efficacy when combined with colistin in murine peritoneal sepsis model by colistin-resistant A. baumannii (Parra-Millán et al., 2018). Taking into account this approach as an adjuvant to antibiotic treatment, it is also necessary to optimize this peptide in order to develop a new library of compounds that improve the efficacy of the lead compound AOA-2.
A number of peptides with biological activity, particularly in the field of antibacterial research, have led to new classes of antibacterial drugs (Gordon et al., 2005). On the basis of the fact that AOA-2 has previously proved its utility as an effective source of anti-virulence compound with specific mechanism of action (Vila-Farrés et al., 2017), we have maintained this backbone as the fundamental core in our new derivatives. Three main points of variation have been considered: (1) increasing of peptide solubility by NH2 addition, (2) substituting tryptophan and arginine by their analogs, (3) and repositioning the general structure of AOA-2.
All these derivatives did not show toxicities in human lungs epithelial cells. Similarly, the lead compound AOA-2 did not show toxicity in these host cells (Vila-Farrés et al., 2017). This data confirm that the variation made in the AOA-2 structure did not affect the viability of host cells.
Bacterial adherence assays were employed to screen the efficacity of these derivatives. Through a various screening steps, two derivatives RW01 (solubility enhancer: NH2) and RW06 (analog of arginine: 2,4-Diaminobutanoic acid) have showed extended spectrum effect on the bacterial adherence to human lung epithelial cells and have been selected for the rest of the experiments. Whereas other tryptophan (3Pal: beta-(3-pyridyl)-alanine, and 2Nal: beta-(2-naphtyl)-alanine) and arginine (lysine) analogs or amino acids repositioning in the lead compound have showed short spectrum effect. This could be explained by the fact that these derivatives did not bind equally to OmpA and its homologous of the all studied GNB, due to difference in the homology of OmpA sequence. Of note, OmpA of A. baumannii present a 58% of homology with OmpA of E. coli and its homologous in P. aeruginosa (Gribun et al., 2003). Further structural analyses are required to decipher why RW01 and RW06 showed greater activity than AOA-2.
Interestingly, RW01 and RW06 have prevented significantly the death of human lung epithelial cells caused by A. baumannii and P. aeruginosa, and with lesser effect against E. coli. These results are consistent with previous observations that the lead compound AOA-2 prevents more the cell death caused by A. baumannii and P. aeruginosa than by E. coli (Vila-Farrés et al., 2017). The failure of both derivatives against E. coli may be attributable to the presence of other highly virulent factors circumventing the loss of OmpA (Nielubowicz and Mobley, 2010).
Conclusion
The optimization program of AOA-2 has generated two derivatives with in vitro promising results against P. aeruginosa and E. coli. In vivo studies are necessary to support these in vitro results and to evaluate the therapeutic efficacy of both derivatives in monotherapy and in combination with clinically relevant antibiotics in severe infections by these GNB.
Data Availability Statement
The datasets generated for this study can be found in the online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
MT, JP, EG, and YS conceived the study and designed the experiments. RA-A, NB, EV, RP-M, and JS carried out the experiments. RA-A and YS analyzed the data and wrote the manuscript. MT, EG, and JP have reviewed the manuscript and the experiments. All authors read and approved the final manuscript.
Funding
This study was supported by the Instituto de Salud Carlos III, Proyectos de Investigación en Salud (Grants PI15/01358 and PI16/01378) and by Plan Nacional de I + D + i 2013–2016 and Instituto de Salud Carlos III, Subdirección General de Redes y Centros de Investigación Cooperativa, Ministerio de Ciencia, Inno-vación y Universidades, Spanish Network for Research in Infectious Diseases (REIPI RD16/0016/0009)—co-financed by the European Development Regional Fund “A way to achieve Europe,” Operative program Intelligent Growth 2014–2020. YS was supported by the Subprograma Miguel Servet Tipo I from the Ministerio de Economía y Competitividad of Spain (CP15/00132).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.634323/full#supplementary-material
References
Baumann, P., Doudoroff, M., and Stanier, R. Y. (1968). A study of the Moraxella group. II. Oxidative-negative species (genus Acinetobacter). J. Bacteriol. 95, 1520–1541.
Chen, I. J., and Foloppe, N. (2008). Conformational sampling of druglike molecules with MOE and catalyst: implications for pharmacophore modeling and virtual screening. J. Chem. Inf. Model. 48, 1773–1791. doi: 10.1021/ci800130k
Choi, C. H., Hyun, S. H., Lee, J. Y., Lee, J. S., Lee, Y. S., Kim, S. A., et al. (2008). Acinetobacter baumannii outer membrane protein A targets the nucleus and induces cytotoxicity. Cell. Microbiol. 10, 309–319. doi: 10.1111/j.1462-5822.2007.01041.x
Choi, C. H., Lee, E. Y., Lee, Y. C., Park, T. I., Kim, H. J., Hyun, S. H., et al. (2005). Outer membrane protein 38 of Acinetobacter baumannii localizes to the mitochondria and induces apoptosis of epithelial cells. Cell. Microbiol. 7, 1127–1138. doi: 10.1111/j.1462-5822.2005.00538.x
Cusumano, C. K., Pinkner, J. S., Han, Z., Greene, S. E., Ford, B. A., Crowley, J. R., et al. (2011). Treatment and prevention of urinary tract infection with orally active FimH inhibitors. Sci. Transl. Med. 3:109ra115. doi: 10.1126/scitranslmed.3003021
Dickey, S. W., Cheung, G. Y. C., and Otto, M. (2017). Different drugs for bad bugs: antivirulence strategies in the age of antibiotic resistance. Nat. Rev. Drug Discov. 16, 457–471. doi: 10.1038/nrd.2017.23
Gaddy, J. A., Tomaras, A. P., and Actis, L. A. (2009). The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic surfaces and in the interaction of this pathogen with eukaryotic cells. Infect. Immun. 77, 3150–3160. doi: 10.1128/IAI.00096-09
Gordon, Y. J., Romanowski, E. G., and McDermott, A. M. (2005). A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Curr. Eye Res. 30, 505–515. doi: 10.1080/02713680590968637
Gribun, A., Nitzan, Y., Pechatnikov, I., Hershkovits, G., and Katcoff, D. J. (2003). Molecular and structural characterization of the HMP-AB gene encoding a pore-forming protein from a clinical isolate of Acinetobacter baumannii. Curr. Microbiol. 47, 434–443. doi: 10.1007/s00284-003-4050-4
Holloway, I. W. (1955). Genetic recombination in Pseudomonas aeruginosa. J. Gen. Microbial. 13, 572–581.
Jarvis, C., Han, Z., Kalas, V., Klein, R., Pinkner, J. S., Ford, B., et al. (2016). Antivirulence isoquinolone mannosides: optimization of the biaryl aglycone for FimH lectin binding affinity and efficacy in the treatment of chronic UTI. Chem. Med. Chem. 11, 367–373. doi: 10.1002/cmdc.201600006
Kaper, J. B., Nataro, J. P., and Mobley, H. L. (2004). Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123–140. doi: 10.1038/nrmicro818
Kim, S. W., Choi, C. H., Moon, D. C., Jin, J. S., Lee, J. H., Shin, J. H., et al. (2009). Serum resistance of Acinetobacter baumannii through the binding of factor H to outer membrane proteins. FEMS Microbiol. Lett. 301, 224–231. doi: 10.1111/j.1574-6968.2009.01820.x
Mittal, R., Krishnan, S., González-Gómez, I., and Prasadarao, N. V. (2011). Deciphering the roles of outer membrane protein a extracellular loops in the pathogenesis of Escherichia coli K1 meningitis. J. Biol. Chem. 286, 2183–2193. doi: 10.1074/jbc.M110.178236
Nicholson, T. F., Watts, K. M., and Hunstad, D. A. (2019). OmpA of uropathogenic Escherichia coli promotes postinvasion pathogenesis of cystitis. Infect. Immun. 77, 5245–5251. doi: 10.1128/IAI.00670-09
Nielubowicz, G. R., and Mobley, H. L. (2010). Host-pathogen interactions in urinary tract infection. Nat. Rev. Urol. 7, 430–441. doi: 10.1038/nrurol.2010.101
Parra-Millán, R., Vila-Farrés, X., Ayerbe-Algaba, R., Varese, M., Sánchez-Encinales, V., Bayó, N., et al. (2018). Synergistic activity of an OmpA inhibitor and colistin against colistin-resistant Acinetobacter baumannii: mechanistic analysis and in vivo efficacy. J. Antimicrob. Chemother. 73, 3405–3412. doi: 10.1093/jac/dky343
Prasadarao, N. V., Wass, C. A., Weiser, J. N., Stins, M. F., Huang, S. H., and Kim, K. S. (1996). Outer membrane protein A of Escherichia coli contributes to invasion of brain microvascular endothelial cells. Infect. Immun. 64, 146–153.
Reading, C., and Cole, M. (1977). Clavulanic acid: a beta-lactamase-inhibiting beta-lactam from Streptomyces clavuligerus. Antimicrob. Agents Chemother. 11, 852–857.
Ruiz-Carmona, S., Alvarez-Garcia, D., Foloppe, N., Garmendia-Doval, A. B., Juhos, S., Schmidtke, P., et al. (2014). rDock: a fast, versatile and open source program for docking ligands to proteins and nucleic acids. PLoS Comput. Biol. 10:e1003571. doi: 10.1371/journal.pcbi.1003571
Sánchez-Encinales, V., Álvarez-Marín, R., Pachón-Ibáñez, M. E., Fernández-Cuenca, F., Pascual, A., Garnacho-Montero, J., et al. (2017). Overproduction of outer membrane protein a by Acinetobacter baumannii as a risk factor for nosocomial pneumonia, bacteremia, and mortality rate increase. J. Infect. Dis. 215, 966–974. doi: 10.1093/infdis/jix010
Seco, J., Luque, F. J., and Barril, X. (2009). Binding site detection and druggability index from first principles. J. Med. Chem. 52, 2363–2371. doi: 10.1021/jm801385d
Smani, Y., Dominguez-Herrera, J., and Pachón, J. (2013). Association of the outer membrane protein Omp33 with fitness and virulence of Acinetobacter baumannii. J. Infect. Dis. 208, 1561–1570. doi: 10.1093/infdis/jit386
Smith, M. G., Gianoulis, T. A., Pukatzki, S., Mekalanos, J. J., Ornston, L. N., and Gerstein, M. (2007). New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev. 21, 601–614. doi: 10.1101/gad.1510307
Smith, S. G. J., Mahon, J., Lambert, M. A., and Fagan, R. P. (2007). A molecular Swiss army knife: OmpA structure, function and expression. FEMS Microbiol. Lett. 273, 1–11. doi: 10.1111/j.1574-6968.2007.00778.x
Song, Y., DiMaio, F., Wang, R. Y. R., Kim, D., Miles, C., Brunette, T., et al. (2013). High-resolution comparative modeling with RosettaCM. Structure 21, 1735–1742. doi: 10.1016/j.str.2013.08.005
Starkey, M., Lepine, F., Maura, D., Bandyopadhaya, A., Lesic, B., He, J., et al. (2014). Identification of anti-virulence compounds that disrupt quorum-sensing regulated acute and persistent pathogenicity. PLoS Pathog. 10:e1004321. doi: 10.1371/journal.ppat.1004321
Tacconelli, E., Carrara, E., Savoldi, A., Harbarth, S., Mendelson, M., Monnet, D. L., et al. (2018). Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327. doi: 10.1016/S1473-3099(17)30753-3
Teng, C. H., Xie, Y., Shin, S., Di Cello, F., Paul-Satyaseela, M., Cai, M., et al. (2006). Effects of ompA deletion on expression of type 1 fimbriae in Escherichia coli K1 strain RS218 and on the association of E. coli with human brain microvascular endothelial cells. Infect. Immun. 74, 5609–5616. doi: 10.1128/IAI.00321-06
Vila-Farrés, X., Parra-Millán, R., Sánchez-Encinales, V., Varese, M., Ayerbe-Algaba, R., Bayó, N., et al. (2017). Combating virulence of gram-negative bacilli by OmpA inhibition. Sci. Rep. 7:14683. doi: 10.1038/s41598-017-14972-y
Weiser, J. N., and Gotschlich, E. C. (1991). Outer membrane protein A (OmpA) contributes to serum resistance and pathogenicity of Escherichia coli K-1. Infect. Immun. 59, 2252–2258.
Keywords: AOA-2 derivatives, outer membrane protein A, inhibitor, Gram-negative bacilli, peptides
Citation: Ayerbe-Algaba R, Bayó N, Verdú E, Parra-Millán R, Seco J, Teixidó M, Pachón J, Giralt E and Smani Y (2021) AOA-2 Derivatives as Outer Membrane Protein A Inhibitors for Treatment of Gram-Negative Bacilli Infections. Front. Microbiol. 12:634323. doi: 10.3389/fmicb.2021.634323
Received: 27 November 2020; Accepted: 21 January 2021;
Published: 12 February 2021.
Edited by:
Santi M. Mandal, Indian Institute of Technology Kharagpur, IndiaReviewed by:
Shakiba Darvish Alipour Astaneh, Semnan University, IranJuan Carlos Vázquez Ucha, Institute of Biomedical Research of A Coruña (INIBIC), Spain
Iraj Rasooli, Shahed University, Iran
William Frank Penwell, Jacksonville University, United States
Copyright © 2021 Ayerbe-Algaba, Bayó, Verdú, Parra-Millán, Seco, Teixidó, Pachón, Giralt and Smani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jerónimo Pachón, cGFjaG9uQHVzLmVz; Younes Smani, eV9zbWFuaUBob3RtYWlsLmNvbQ==; eXNtYW5pLWliaXNAdXMuZXM=