
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 03 February 2021
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 12 - 2021 | https://doi.org/10.3389/fmicb.2021.592291
This article is part of the Research TopicGlobally or Regionally Spread of Epidemic Plasmids Carrying Clinically Important Resistance Genes: Epidemiology, Molecular Mechanism, and DriversView all 12 articles
 Paula Kurittu1*
Paula Kurittu1* Banafsheh Khakipoor1Maria Aarnio2
Banafsheh Khakipoor1Maria Aarnio2 Suvi Nykäsenoja2
Suvi Nykäsenoja2 Michael Brouwer3
Michael Brouwer3 Anna-Liisa Myllyniemi2
Anna-Liisa Myllyniemi2 Elina Vatunen4
Elina Vatunen4 Annamari Heikinheimo1,2
Annamari Heikinheimo1,2Plasmid-mediated extended-spectrum beta-lactamase (ESBL), AmpC, and carbapenemase producing Enterobacteriaceae, in particular Escherichia coli and Klebsiella pneumoniae, with potential zoonotic transmission routes, are one of the greatest threats to global health. The aim of this study was to investigate global food products as potential vehicles for ESBL/AmpC-producing bacteria and identify plasmids harboring resistance genes. We sampled 200 food products purchased from Finland capital region during fall 2018. Products originated from 35 countries from six continents and represented four food categories: vegetables (n = 60), fruits and berries (n = 50), meat (n = 60), and seafood (n = 30). Additionally, subsamples (n = 40) were taken from broiler meat. Samples were screened for ESBL/AmpC-producing Enterobacteriaceae and whole genome sequenced to identify resistance and virulence genes and sequence types (STs). To accurately identify plasmids harboring resistance and virulence genes, a hybrid sequence analysis combining long- and short-read sequencing was employed. Sequences were compared to previously published plasmids to identify potential epidemic plasmid types. Altogether, 14 out of 200 samples were positive for ESBL/AmpC-producing E. coli and/or K. pneumoniae. Positive samples were recovered from meat (18%; 11/60) and vegetables (5%; 3/60) but were not found from seafood or fruit. ESBL/AmpC-producing E. coli and/or K. pneumoniae was found in 90% (36/40) of broiler meat subsamples. Whole genome sequencing of selected isolates (n = 21) revealed a wide collection of STs, plasmid replicons, and genes conferring multidrug resistance. blaCTX–M–15-producing K. pneumoniae ST307 was identified in vegetable (n = 1) and meat (n = 1) samples. Successful IncFII plasmid type was recovered from vegetable and both IncFII and IncI1-Iγ types from meat samples. Hybrid sequence analysis also revealed chromosomally located beta-lactamase genes in two of the isolates and indicated similarity of food-derived plasmids to other livestock-associated sources and also to plasmids obtained from human clinical samples from various countries, such as IncI type plasmid harboring blaTEM–52C from a human urine sample obtained in the Netherlands which was highly similar to a plasmid obtained from broiler meat in this study. Results indicate certain foods contain bacteria with multidrug resistance and pose a possible risk to public health, emphasizing the importance of surveillance and the need for further studies on epidemiology of epidemic plasmids.
The increasing prevalence of bacteria producing extended-spectrum beta-lactamases (ESBL) and plasmid-encoded AmpC (pAmpC) enzymes mediating resistance to many commonly used antibiotics has led to global health problems. ESBL/pAmpC are commonly found in Gram-negative enterobacteria, such as Escherichia coli and Klebsiella pneumoniae, which belong to the normal human and animal intestinal microbiota. These bacteria may spread during food production, for example during the slaughter process, and thus contaminate food products, imposing a threat to consumers.
In addition to bacterial clonal spread, certain plasmids have been shown to be successful in transferring resistance genes between bacterial populations through horizontal gene transfer, thus increasing the spread of antimicrobial resistance (AMR) (Accogli et al., 2013; Carattoli, 2013).
Typically, ESBL/AmpC-producing bacteria are not more virulent than susceptible bacteria but problems may arise when bacteria cause infections requiring treatment with antimicrobials, such as urinary tract, bloodstream, intra-abdominal, and respiratory tract infections (Pitout and Laupland, 2008). In addition to healthcare associated infections, ESBL-producing E. coli have been recognized as a major cause of community-onset disease (Laupland et al., 2008). The World Health Organization (WHO) has issued guidelines on the use of medically important antimicrobials in food-producing animals to preserve the effectiveness of antimicrobials important for human medicine (WHO, 2017a). Cephalosporins (third, fourth, and fifth generation) and carbapenems have been classified as critically important antimicrobials for human medicine by the WHO (WHO, 2019). The WHO has additionally published a global priority list of antibiotic-resistant bacteria to guide research, which includes specific carbapenem-resistant and third-generation cephalosporin-resistant species of the Enterobacteriaceae family, including E. coli and K. pneumoniae (WHO, 2017b).
Antimicrobial resistance can disseminate in many environments, i.e., communities, hospitals, food production plants, and farms. International travel as well as import and export of goods work as mediators of the spread of AMR across country borders. The use of antimicrobials, both quantitatively and qualitatively, varies between countries, leading to differences in AMR levels (WHO et al., 2018; EFSA and ECDC, 2020). In Europe, antimicrobial sales for food-producing animals range from 3.1 to 423.1 mg/PCU (population correction unit) (European Medicines Agency [EMA], 2019). The median for all countries participating in the report was 61.9 mg/PCU. Northern European countries use less antimicrobials in animals in general compared with Southern European countries (European Medicines Agency [EMA], 2019). A recent study by Van Boeckel et al. (2019) identified the largest hot spots of AMR in animals in China and India, with predicted hot spots emerging in Kenya and Brazil. The use of antimicrobials affects the resistance levels in food-producing animals (Hoelzer et al., 2017) and the use of certain antimicrobials in livestock has been shown to correlate with the level of AMR in E. coli in pigs, poultry, and cattle (Chantziaras et al., 2014). Antimicrobial use in animals can even be seen in the levels of AMR in human populations (European Centre for Disease Prevention and Control [ECDC] et al., 2017; Tang et al., 2017). As the demand for animal-source nutrition is rising with the human population increase, more antimicrobials will be used in food-producing animals, especially in low- and middle-income countries (Van Boeckel et al., 2015; Schar et al., 2018). Regarding meat products, broiler meat has been recognized as having a high prevalence of ESBL/AmpC-producing E. coli (Ewers et al., 2012; EFSA and ECDC, 2020).
In addition to antimicrobial use, other factors such as lack of sanitation may also affect global AMR gene diversity and abundance (Hendriksen et al., 2019). In particular, fresh food products may be susceptible to bacterial contamination from environmental sources such as poor-quality irrigation water or by cross contamination in food-producing facilities (FAO and WHO, 2019). Application of manure of animal origin is another source of AMR dissemination in agriculture and food production (Hartmann et al., 2012). In addition, seafood is often grown in developing countries in unsanitary conditions, with an increased risk of AMR (Boss et al., 2016). Subsequently, fruits and vegetables may also acquire resistant bacteria through irrigation water contaminated by aquaculture production (Done et al., 2015). Animal crops fertilized with manure and contaminated by soil bacteria may be a source of resistant bacteria for food-producing animals and lead to amplification of AMR in animal gut microbiota with concurrent antimicrobial administration (Witte, 2000; Marshall and Levy, 2011; FAO and WHO, 2019). Also, human-derived pathogenic bacteria together with AMR may enter the food chain via wastewater or sewage sludge used for irrigation or fertilization in agriculture (Reinthaler et al., 2010).
Our objective was to study the occurrence of ESBL/AmpC-producing Enterobacteriaceae in food products from a wide selection of countries and multiple food categories to assess the risk of AMR. Furthermore, to study the epidemiology of bacterial isolates and their plasmids, a subset of ESBL/AmpC-producing Enterobacteriaceae isolates were subjected to whole genome sequencing (WGS) to determine sequence types (STs), resistance and virulence genes, plasmid incompatibility (Inc) groups and subtypes, and to verify bacterial species identification. In addition, short-read sequences were combined with long-read sequences to verify beta-lactamase harboring plasmids to gain insight to plasmid epidemiology and to compare plasmid sequences to previously published plasmids to identify similarities to plasmids identified from different sources. To the best of our knowledge, this study is the first of its kind using WGS for studying AMR and beta-lactamase-harboring plasmids from a large set of import countries, with 35 countries included.
Food products were collected from nine grocery stores in the Helsinki region during November 2018.
Altogether 200 individual products were collected from four different food categories, including vegetables (n = 60), fruits and berries (n = 50), meat (n = 60), and seafood products (n = 30). A detailed list of sampled products including information on country of origin, store of purchase and whether the product was fresh or frozen, and whether it originated from a same batch number with another sampled product is provided in Supplementary Table 1. Briefly, 11 batch numbers were identical for 32 sampled products. These 32 products included 10 raw broiler meat samples originating from the same batch and purchased from the same store. The other same-batch products consisted each of two or three products. Food products were divided into different categories according to how they are traditionally perceived; for example, herbs were categorized into vegetables. Products varied in size and consisted of raw, ready-to-eat, frozen, and cooked products. Country or region of origin for samples from different food categories in presented in Table 1.

Table 1. Country/region of origin for samples per food category.
The meat category included raw broiler meat products (n = 10/60) from the same batch number purchased from the same store. One package of broiler meat consisted of two kilograms of raw chicken wings, and 10 of these packages were included in the study. Additional subsamples were taken from each package for further characterization of bacterial isolates to study the diversity of Enterobacteriaceae, STs, plasmid replicons, and resistance genes in raw broiler meat, which has been identified as a rich reservoir of ESBL/AmpC-producing bacteria (EFSA Panel on Biological Hazards, 2011). Four subsamples were taken from each broiler meat package, totaling 40 subsamples. The aim of subsampling was to further study the diversity of ESBL/AmpC-producing Enterobacteriaceae found in a single product.
The country and region of origin of the samples per food category is shown in Table 1.
Samples were transported to the laboratory and stored at 4°C, and analysis was started within 24 h. Using sterile scissors and forceps, 25 g of each sample was dissected at various sites of the product and placed into a sterile Stomacher strainer bag (Seward Stomacher 400 Classic Strainer bag, Worthing, United Kingdom) with 225 ml of sterile buffered peptone water (Oxoid, Basingstoke, Hampshire, United Kingdom) and homogenized for 60 s (Stomacher 400 laboratory blender, Seward, United Kingdom). Samples were incubated at 37°C for 18–22 h. After incubation, a loopful (10 μl) of each enrichment was streaked onto two parallel MacConkey agar plates (Lab M, Lancashire, United Kingdom; Scharlau Chemie s.a, Sentmenat, Spain) with 1 mg/l cefotaxime. To improve the isolation of Enterobacteriaceae listed as the highest priority by the WHO (2017b), one of the plates was incubated at 44°C and the other at 37°C for 18–22 h. One colony from each morphologically different bacterial growth from each plate was re-streaked onto individual MacConkey agar plates with 1 mg/l cefotaxime and incubated overnight at 37°C. Bacterial colonies were re-streaked onto individual MacConkey agar plates with cefotaxime supplement until a pure culture was achieved.
Isolates were streaked onto a bovine blood agar plate and incubated at 37°C overnight for bacterial species determination with a matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI-TOF MS) based Bruker Biotyper (Bruker Daltonics). A score value of 2.0–3.0 was considered high and thus a confident match and was set as the criteria. All isolates identified as E. coli and K. pneumoniae were stored at −70°C for further characterization.
Antimicrobial susceptibility testing (AST) was performed on E. coli and K. pneumoniae isolates to confirm production of ESBL, AmpC, and/or carbapenemase. AST was performed with a disk diffusion method; susceptibility to third-generation cephalosporins was tested with ceftazidime (10 μg) (Neo-Sensitabs, Rosco Diagnostica, Taastrup, Denmark) and cefotaxime (5 μg) (Oxoid, Basingstoke, Hampshire, United Kingdom), to fourth-generation cephalosporins with cefepime (30 μg), to cephamycins with cefoxitin (30 μg), and to carbapenems with meropenem (10 μg) (Neo-Sensitabs, Rosco Diagnostica, Taastrup, Denmark). Epidemiological cut-off values were used as a reference (EUCAST, 2017). Synergism between third-generation cephalosporins and clavulanic acid was tested with a combination disk diffusion test with cefotaxime + clavulanic acid (30 μg + 10 μg) and ceftazidime + clavulanic acid (30 μg + 10 μg) (Neo-Sensitabs, Rosco Diagnostica, Taastrup, Denmark). E. coli ATCC 25922 was included as a quality control. In addition to resistance to third-generation cephalosporins, resistance to cephamycin and <5 mm difference in inhibition zones in the combination disc diffusion test were used as criteria for AmpC production, whereas ESBL production was evidenced by resistance to third-generation cephalosporins and ≥5 mm difference in the combination disk diffusion test.
From all food samples positive for ESBL/AmpC-producing E. coli or K. pneumoniae, a collection of isolates was chosen for WGS analysis in order to study the presence of AMR, virulence genes, and plasmid replicons, as well as to assess the multilocus sequence type (MLST). If applicable, a representative from each ESBL/AmpC enzyme type category (ESBL, AmpC, or ESBL together with AmpC) and bacterial species (E. coli or K. pneumoniae) was chosen from each positive food sample, excluding subsamples. Consequently, from one to three isolates were chosen for whole genome sequencing from each positive sample.
Bacterial DNA was extracted and purified with a PureLink Genomic DNA Mini Kit (Invitrogen by Thermo Fischer Scientific, Carlsbad, CA, United States) according to the manufacturer’s instructions. The assessment of DNA quality was carried out using a NanoDrop ND-1000 spectrophotometer (Thermo Fischer Scientific, Wilmington, DE, United States) and DNA quantity was measured using a Qubit 2.0 fluorometer (Invitrogen, Life Technologies, Carlsbad, CA, United States). Library preparation was performed with an Illumina Nextera XT and sequencing with an Illumina Novaseq 6000 (Center for Genomics and Transcriptomics, Tübingen, Germany) with paired-end reads. Samples were sequenced with 100× coverage and 2× 100 bp read length.
Subsequently, seven short-read sequenced E. coli isolates were chosen for long-read sequencing in order to study beta-lactamase harboring plasmids in more depth. Isolates were chosen from short-read sequenced isolates to represent a wide selection of different beta-lactamases, MLST types, and plasmid replicons. DNA extraction and purification were performed as described in Section “Short-Read Sequencing.”
DNA extracts from three or two isolates at a time were multiplexed using a SQK-LSK109 ligation sequence kit (Oxford Nanopore Technologies, United Kingdom) according to the manufacturer’s protocol. Libraries were loaded onto FLO-FLG001 R9.4.1 Flongle flow cells (Oxford Nanopore Technologies, United Kingdom) used with the MinION Mk1B sequencing device and sequenced with MinKNOW software v19.06.8 for 20–24 h.
All raw sequences have been deposited at European Nucleotide Archive (ENA) at EMBL-EBI under accession number PRJEB377791. Accession numbers for each sequenced isolate are provided in Supplementary Table 2.
Bioinformatic analyses of bacterial DNA sequences were run on a web-based service (Center for Genomic Epidemiology, DTU, Denmark). Raw reads were assembled with SPAdes 3.9 (Nurk et al., 2013) and the rest of the analyses were carried out with assembled contigs according to the service’s recommendations with default values for identity and coverage. Acquired AMR genes were determined using ResFinder 3.1 (Zankari et al., 2012), bacterial species identification was confirmed with KmerFinder 3.1 (Hasman et al., 2014; Larsen et al., 2014; Clausen et al., 2018), MLST was determined with MLST 2.0 (Larsen et al., 2012) using E. coli scheme 1 (Wirth et al., 2006), virulence genes for E. coli isolates were determined with VirulenceFinder 2.0 (Joensen et al., 2014), plasmid replicons were determined with PlasmidFinder 2.1 (Carattoli et al., 2014), and pMLST 2.0 (Carattoli et al., 2014) was used for typing plasmid replicons, where applicable.
Nanopore FAST5 read files were basecalled using Guppy v3.4.1 (Oxford Nanopore Technologies, United Kingdom) with FASTQ output and demultiplexed with Qcat v1.1.0 (Oxford Nanopore Technologies, United Kingdom). Quality trimming was performed with BBDuk (BBTools v38.71, Joint Genome Institute, United States) using a QTRIM value of seven. Hybrid assembly of Illumina and nanopore sequences was performed with Unicycler v0.4.8 (Wick et al., 2017) set at default values. Hybrid assembled FASTA files were uploaded to ResFinder 3.2 (Zankari et al., 2012) to determine acquired beta-lactamase resistance genes with default values for identity and coverage. PlasmidFinder 2.1 (Carattoli et al., 2014) was used to determine plasmid replicons located in the same contigs as beta-lactamase genes. Plasmid STs were determined for beta-lactamase harboring plasmid replicons with pMLST 2.0 (Carattoli et al., 2014). VirulenceFinder 2.0 (Joensen et al., 2014) was used to confirm virulence genes present on plasmid contigs. The plasmid sequences were annotated with Prokka v1.13 (Seemann, 2014) and manually curated with BLASTn/BLASTp. The newly developed tool MobileElementFinder v1.0.3 (Johansson et al., 2021) was utilized to search for mobile elements together with BLASTn/BLASTp. Plasmid structures were visualized with Easyfig v2.2.2 (Sullivan et al., 2011) for each different plasmid Inc group identified in the study, and comparisons to previously published plasmids with available metadata were visualized with BRIG v0.95 (Alikhan et al., 2011).
Altogether, 14 out of 200 food samples were positive for ESBL/AmpC-producing E. coli and/or K. pneumoniae (Table 2). ESBL/AmpC-producing E. coli was found in 3% (2/60) of vegetable samples. The positive samples were obtained from two coriander samples originating from Malaysia and were from the same batch and purchased from the same store. ESBL/AmpC-producing K. pneumoniae was found in 1% (1/60) of vegetable samples. The positive sample was obtained from chili pepper originating from Malaysia purchased from the same store as the coriander samples.
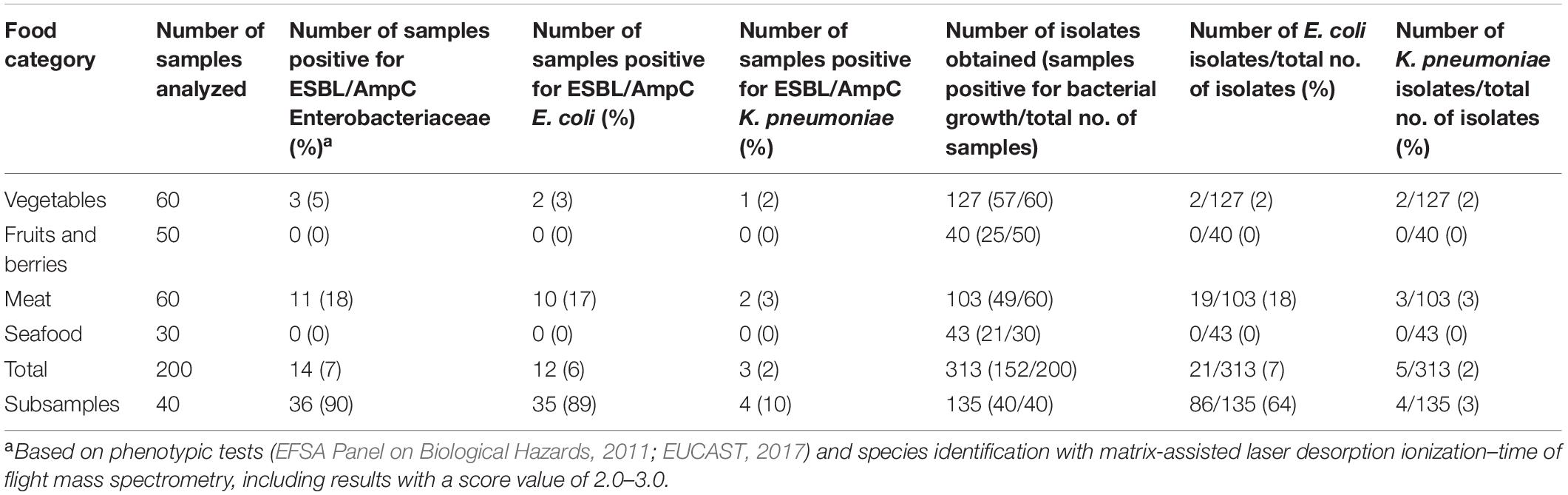
Table 2. Occurrence of ESBL/AmpC-producing Enterobacteriaceae in food samples.
ESBL/AmpC-producing E. coli was found in 17% (10/60) of meat samples, all originating from raw broiler meat from the same batch originating from Lithuania and purchased from the same store. In addition, ESBL/AmpC-producing K. pneumoniae was recovered from 3% (2/60) of meat samples, originating from the aforementioned raw broiler meat (n = 1) and frozen turkey meat (n = 1) originating from Poland. Positive samples originating from food products are presented in Figure 1.
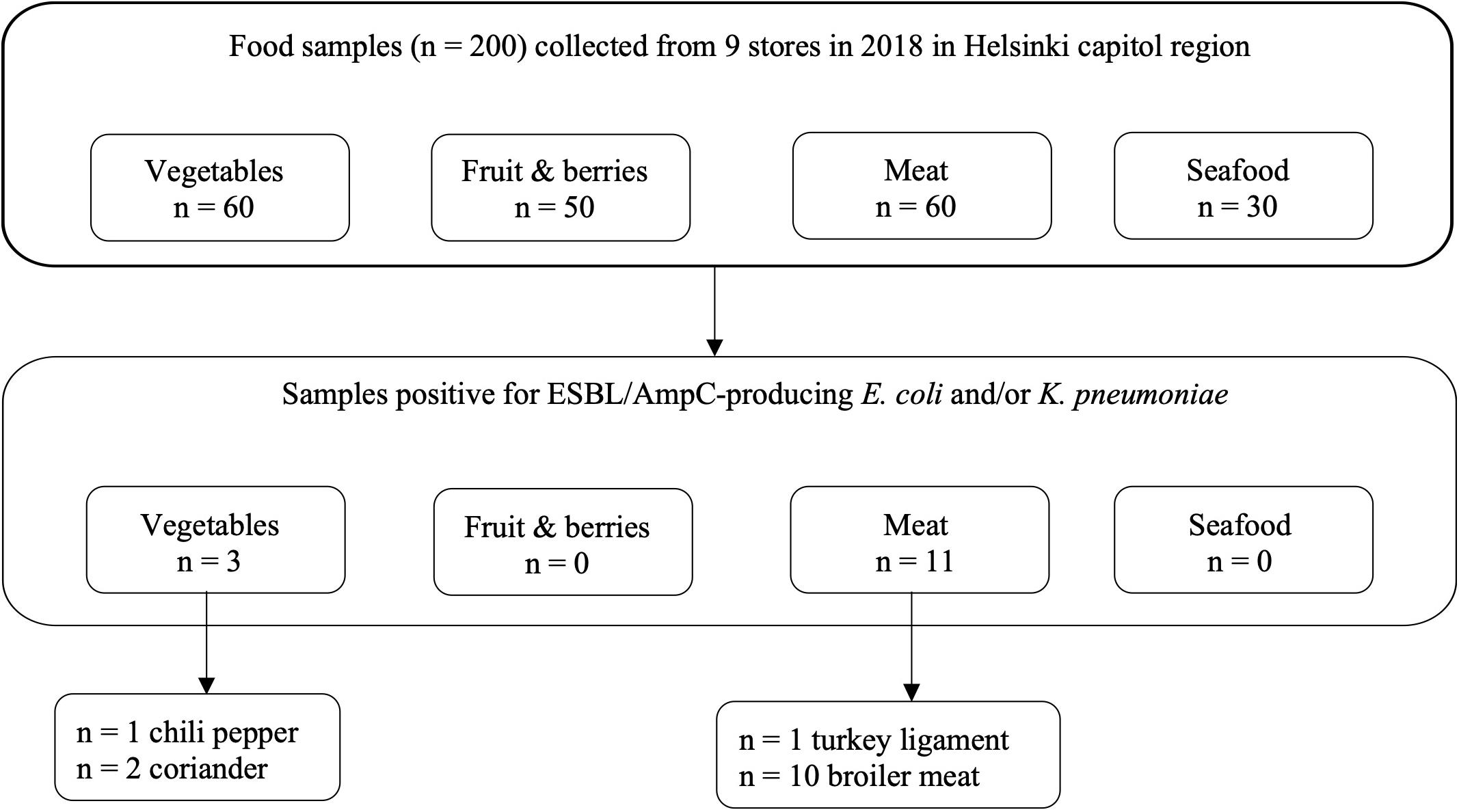
Figure 1. Food samples positive for ESBL/AmpC-producing Escherichia coli and/or Klebsiella pneumoniae based on phenotypic tests (EFSA Panel on Biological Hazards, 2011; EUCAST, 2017) and species identification with matrix- assisted laser desorption ionization–time of flight mass spectrometry, including results with a score value of 2.0–3.0.
Altogether, 152 out of 200 food samples yielded a total of 313 isolates, which were subjected to bacterial species identification with MALDI-TOF MS. Isolates originated from samples incubated at both 44 and 37°C. Samples with bacterial growth (n = 152) yielded from one to four isolates per parallel agar plate. Information on samples and isolates positive for bacterial growth and ESBL/AmpC-producing E. coli and K. pneumoniae is provided in Figure 2. Altogether, 21 isolates were identified as E. coli and five as K. pneumoniae. Isolates positive for ESBL/AmpC-producing E. coli and/or K. pneumoniae and the selection process for WGS are presented in Figure 2.
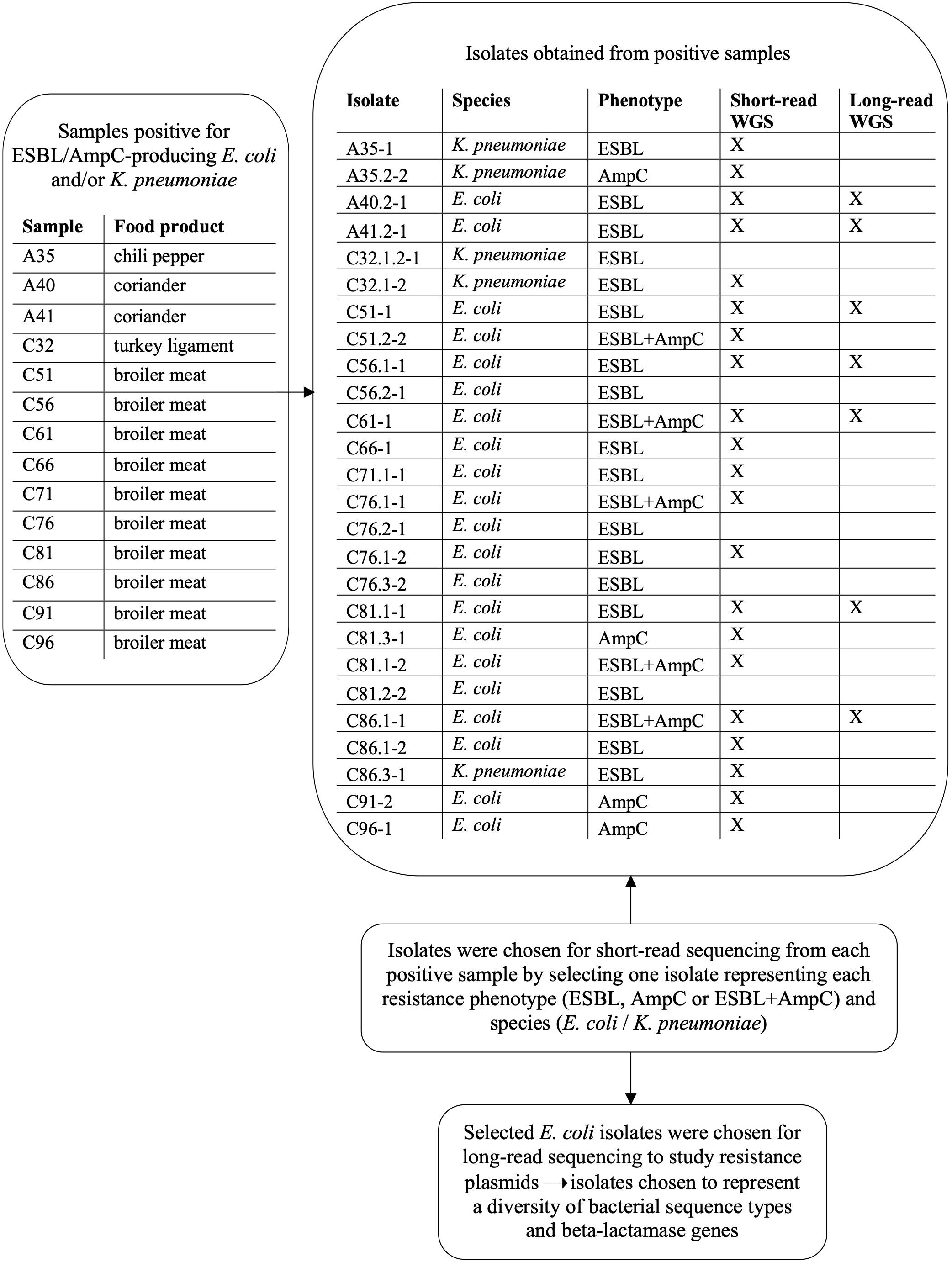
Figure 2. Isolates obtained from food samples positive for ESBL/AmpC-producing Escherichia coli and/or Klebsiella pneumoniae and subsequent isolate selection for short-read and long-read whole genome sequencing (WGS).
Altogether, ESBL/AmpC-producing E. coli and/or K. pneumoniae were found in 90% (36/40) of subsamples. The subsamples were taken from 10 raw broiler meat samples that were included in the main samples. These raw broiler meat samples originated all from the same batch from the same store. Altogether, 135 isolates were recovered from 40 raw broiler meat subsamples. From these subsample isolates, 109 were identified with MALDI-TOF MS with a score value of 2.0–3.0. From these isolates, 86 were identified as E. coli and four as K. pneumoniae. For 23 isolates, species identification was not possible.
Altogether, 21 isolates from the main samples were identified as E. coli. From these, 13 (62%) were phenotypically ESBL producers, three (14%) AmpC producers, and five (24%) produced both AmpC and ESBL. From the five isolates from the main samples identified as K. pneumoniae, four were phenotypically ESBL producers and one an AmpC producer. All isolates were resistant to third-generation cephalosporin (cefotaxime and ceftazidime), except one E. coli isolate that was susceptible to ceftazidime but resistant to cefotaxime. None of the isolates were resistant to carbapenem (meropenem).
According to AST with disk diffusion, from the 86 isolates identified as E. coli, 43 (50%) were ESBL producers, 20 (23%) AmpC producers, and 23 (27%) produced both AmpC and ESBL. All four K. pneumoniae isolates were ESBL producers. All isolates from the subsamples were resistant to third-generation cephalosporin (cefotaxime and ceftazidime), except one K. pneumoniae isolate that was susceptible to ceftazidime but resistant to cefotaxime. None of the subsample isolates were resistant to carbapenem (meropenem).
Altogether, 21 isolates were subjected to short-read WGS with Illumina, consisting of 17 E. coli and four K. pneumoniae isolates, originating from one chili pepper, two coriander, one turkey, and 10 broiler meat samples. Genotypic results are presented in Table 3.
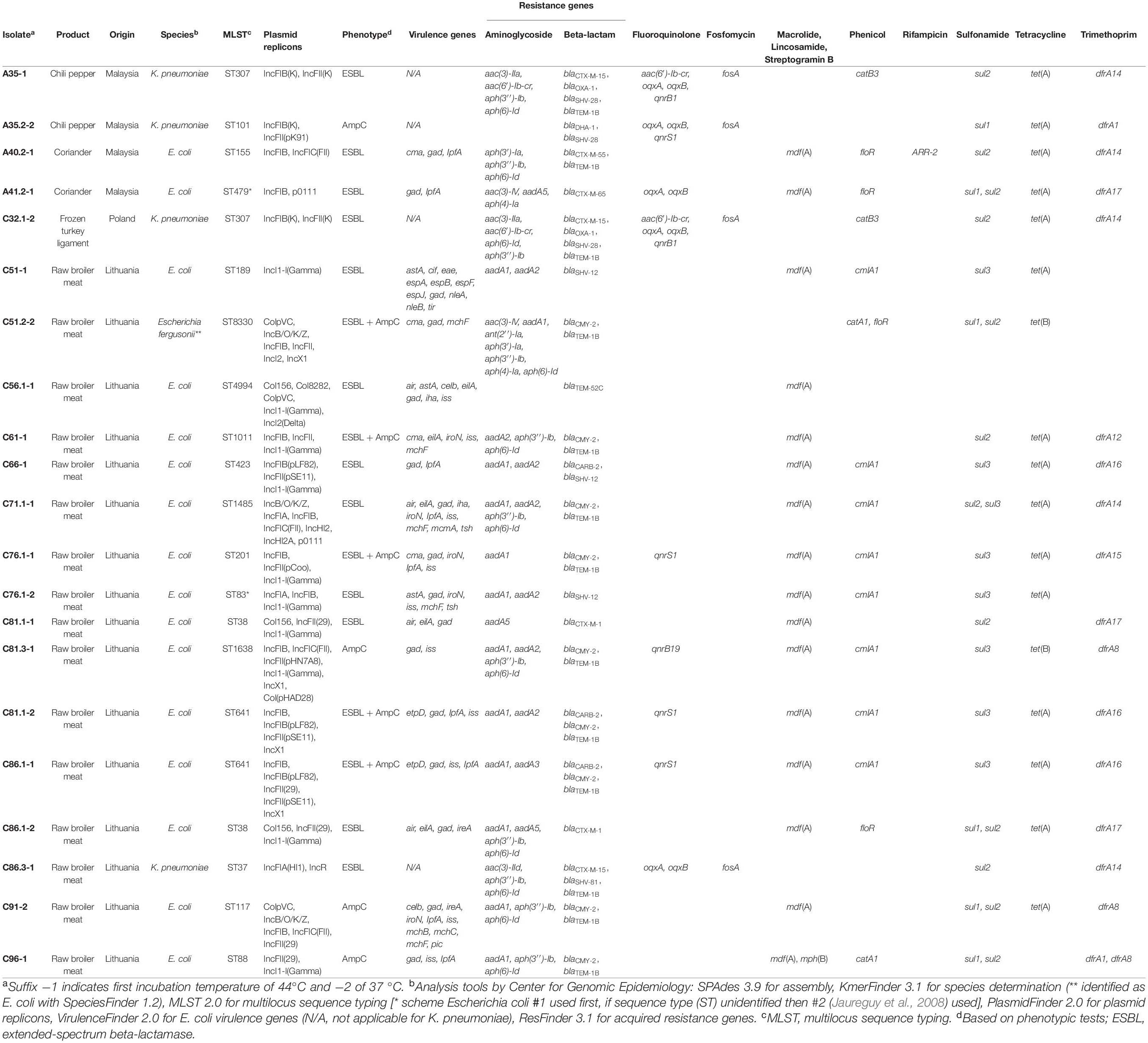
Table 3. Genomic characteristics of Escherichia coli and Klebsiella pneumoniae strains isolated from globally produced food.
The sequenced isolates were confirmed to be either E. coli or K. pneumoniae. Altogether 18 MLST types were identified, 15 STs from the 17 sequenced E. coli isolates and three STs from the four sequenced K. pneumoniae isolates. Four different STs were identified from the four vegetable samples: ST155 and ST479 E. coli isolates from coriander, and ST307 and ST101 K. pneumoniae isolates from chili. The K. pneumoniae isolate from turkey meat was identified as ST307. The 16 isolates originating from broiler meat consisted of 14 different STs, including ST189, ST8330, ST4994, ST1011, ST423, ST1485, ST201, ST83, ST38, ST1638, ST641, ST117, and ST88 E. coli isolates and ST37 K. pneumoniae isolate.
Multiple AMR genes were observed in all of the sequenced isolates. All isolates harbored beta-lactamase gene(s) (blaCTX–M–1, blaCTX–M–15, blaCTX–M–55, blaCTX–M–65, blaSHV–12, blaSHV–81, blaSHV–28, blaTEM–1B, blaTEM–52C, blaCARB–2, blaOXA–1, blaDHA–1, and/or blaCMY–2) and almost all isolates harbored genes conferring resistance to aminoglycoside, macrolide, lincosamide, streptogramin B, phenicol, sulfonamide, tetracycline, and trimethoprim. Additionally, a few isolates harbored plasmid-mediated quinolone resistance (PMQR), fosfomycin, and/or rifampicin resistance genes. Nine out of the 16 sequenced broiler meat samples harbored the AmpC beta-lactamase blaCMY–2.
All isolates were carrying AMR genes conferring resistance toward critically important antimicrobials (WHO, 2019), including aminoglycosides, third- and fourth-generation cephalosporins, macrolides, and quinolones. All of the E. coli isolates harbored the macrolide resistance gene mdf(A). All of the K. pneumoniae isolates harbored PMQR and fosfomycin resistance genes, whereas only five E. coli isolates harbored PMQR genes and none harbored fosfomycin resistance genes. Aminoglycoside resistance genes were found in all but one E. coli and one K. pneumoniae isolate. Two K. pneumoniae isolates harbored gene aac(6′)-Ib-cr, which confers resistance toward both aminoglycosides and fluoroquinolones (Frasson et al., 2011). Phenicol resistance was found in 16 isolates, rifampicin resistance in one isolate, sulfonamide resistance in all but one isolate, and tetracycline and trimethoprim resistance in 17 isolates.
ESBL/AmpC phenotypes correlated with the detected genotype in all but four (isolates A35.2-2, C51.2-2, C61-1, and C71.1-1) of the sequenced isolates. Isolate A35.2-2 was phenotypically an AmpC producer but harbored both blaDHA–1 and blaSHV–28. Isolates C51.2-2 and C61-1 were phenotypically both ESBL and AmpC producers, although they possessed only blaTEM–1B in addition to blaCMY–2. Isolate C71.1-1 was phenotypically an ESBL producer but harbored blaCMY–2 in addition to blaTEM–1B. blaTEM–1B confers resistance toward penicillin and ampicillin but not significantly toward extended-spectrum cephalosporins (Paterson and Bonomo, 2005). Complete resistance gene profiles are shown in Table 3.
A total of 24 different plasmid replicons were identified from the sequenced isolates with PlasmidFinder 2.1 (Carattoli et al., 2014) with IncF subtype IncFIB and IncI1-Iγ being the most common and appearing in 11 and 10 isolates, respectively. Plasmid replicons belonging to IncF groups were overall detected in 19 isolates. Each isolate harbored from one to seven different plasmid replicons, with the average amount of replicons being three per isolate. Typing of plasmids with plasmid multilocus sequence typing (pMLST) revealed 14 different STs, with the most common ones being ST12 and ST95-CC9 (clonal complex) appearing each in three isolates. Three ST/clonal complex clusters were linked to the IncI1-Iγ plasmid group. IncI1-Iγ ST3-CC3 was found in two ST38 E. coli isolates, both harboring resistance gene blaCTX–M–1. IncI1-Iγ ST95-CC9 was associated with resistance gene blaSHV–12, which was found in three E. coli isolates, all with different STs: ST189, ST423, and ST83. IncI1-Iγ ST12 was identified in three E. coli isolates harboring blaCMY–2. All of the IncI1-Iγ plasmid replicons originated from broiler meat samples originating from the same batch and collected from the same store. pMLST results are presented in Supplementary Table 3.
The isolates harbored multiple different virulence genes. Virulence gene results are presented in Table 3. One of the isolates, originating from broiler meat, was positive for adhesin intimin coding eae gene, which is associated with enteropathogenic E. coli (Frankel et al., 1998; Müller et al., 2016).
Altogether, seven E. coli isolates were subjected to long-read sequencing with Oxford Nanopore Technologies, consisting of two coriander and five broiler meat isolates. Results of the hybrid assembled plasmid sequences are presented in Table 4. All long-read sequenced and hybrid assembled isolates were found to harbor resistance genes and plasmid replicons matching with short-read sequence analysis. In the two coriander isolates, beta-lactamase genes were located in plasmid replicons belonging to the IncF group, whereas in four out of five broiler isolates beta-lactamase genes were located on IncI1-Iγ plasmids. Isolate C86.1-1 included blaCARB–2-carrying IncF type plasmid replicon and blaTEM–1-carrying IncX1 plasmid replicon in addition to a chromosomal blaCMY–2. A chromosomal blaTEM–1B gene was detected also in isolate C61-1. All detected plasmid replicons represented different plasmid STs. Additionally, all IncF plasmid replicons, the IncX1 replicon and one IncI1-Iγ replicon were found to harbor multiple resistance genes.
Table 4. Hybrid assembled Escherichia coli food isolates and corresponding beta-lactamase harboring plasmids for comparative analysis.
Three of the hybrid assembled food isolates were found to carry plasmids belonging to the IncF group. Two of the isolates were of coriander origin (A40.2-1 and A41.2-1) and one from raw broiler meat (C86.1-1). RST analysis with pMLST tool (Carattoli et al., 2014) indicated new FIB alleles for pZPK-A41.2-1 and pZPK-C86.1-1 which were submitted to PubMLST database2 and assigned FIB76 and FIB77, respectively. IncFII/IncFIB replicon carrying narrow-spectrum beta-lactamase blaCARB–2 with a FAB formula of F68:A-:B77 from isolate C86.1-1 additionally carried blaCMY–2 in the chromosome and an IncX1 plasmid harboring blaTEM–1. From this isolate, the IncX1 plasmid (pZPK-C86.1-1_X1) was selected for further comparison analysis to achieve diverse ESBL-plasmid comparisons (Table 4).
pZPK40.2-1 from isolate A40.2-1 carried a multireplicon IncFIB/IncFIC with a size of 120.7 kb and C + G content of 51.45% and 136 predicted coding sequences (CDSs). pZPK41.2-1 from isolate A41.2-1 carried IncFIB replicon together with p0111 and was 153.3 kb in size with G + C content of 49.93% and 165 CDSs. Comparison to a previously published and annotated IncF plasmid with FAB formula F18:A-:B1:C4 (GenBank accession: MK878890.1) indicated high variability between the plasmids with multiple insertions of mobile elements in both pZPK-A40.2-1 and pZPK-A41.2-1 (Figure 3A).
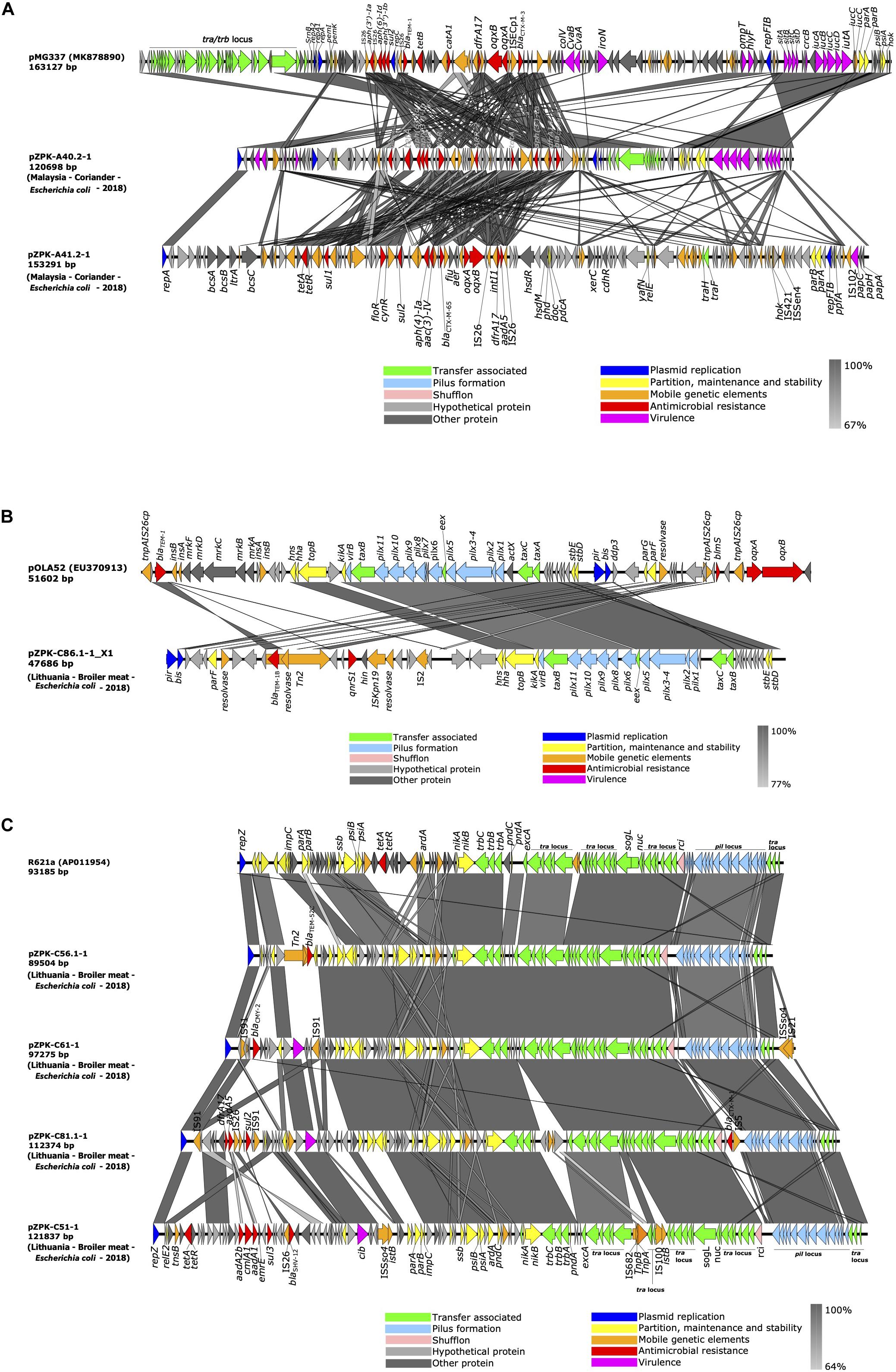
Figure 3. Linear comparison of (A) IncFIB/IncFIC(FII) and IncFIB/p0111, (B) IncX1 and (C) IncI1-Iγ plasmids identified in this study with previously published plasmids (GenBank accession numbers in parentheses for reference plasmids; for plasmids identified in this study the country of origin, source, bacterial species and year of isolation is provided in parentheses). Gray areas between plasmid sequences indicate the percentage of nucleotide sequence identity. The arrows represent coding sequences and their orientation and are colored based on their predicted function.
pZPK-A40.2-1 harbored multiple resistance genes in addition to its beta-lactamases blaCTX–M–55 and blaTEM–1B, including ARR-2, floR, dfrA14, aph(3′′)-Ib, aph(3′)-Ia, aph(6)-Id, tet(A) and sul2. In addition, multiple virulence genes (cma, cvaC, hlyF, iucC, iutA, ompT, sitA) were detected. Top BLAST hits with available metadata in NCBI included an IncFII/IncFIC type plasmid with a FAB profile F18:A-B1 and blaCTX–M–55, blaTEM–1B and mcr-1 from a human urine sample from United States (GenBank accession: KX276657.1) with 89% coverage and 99.73% identity. Another plasmid with the FAB profile F18:A-B1 with blaCTX–M–55 obtained from a turkey in Canada (GenBank accession: CP059932.1) was found to be similar with 88% coverage and 99.78% identity (Figure 4A).
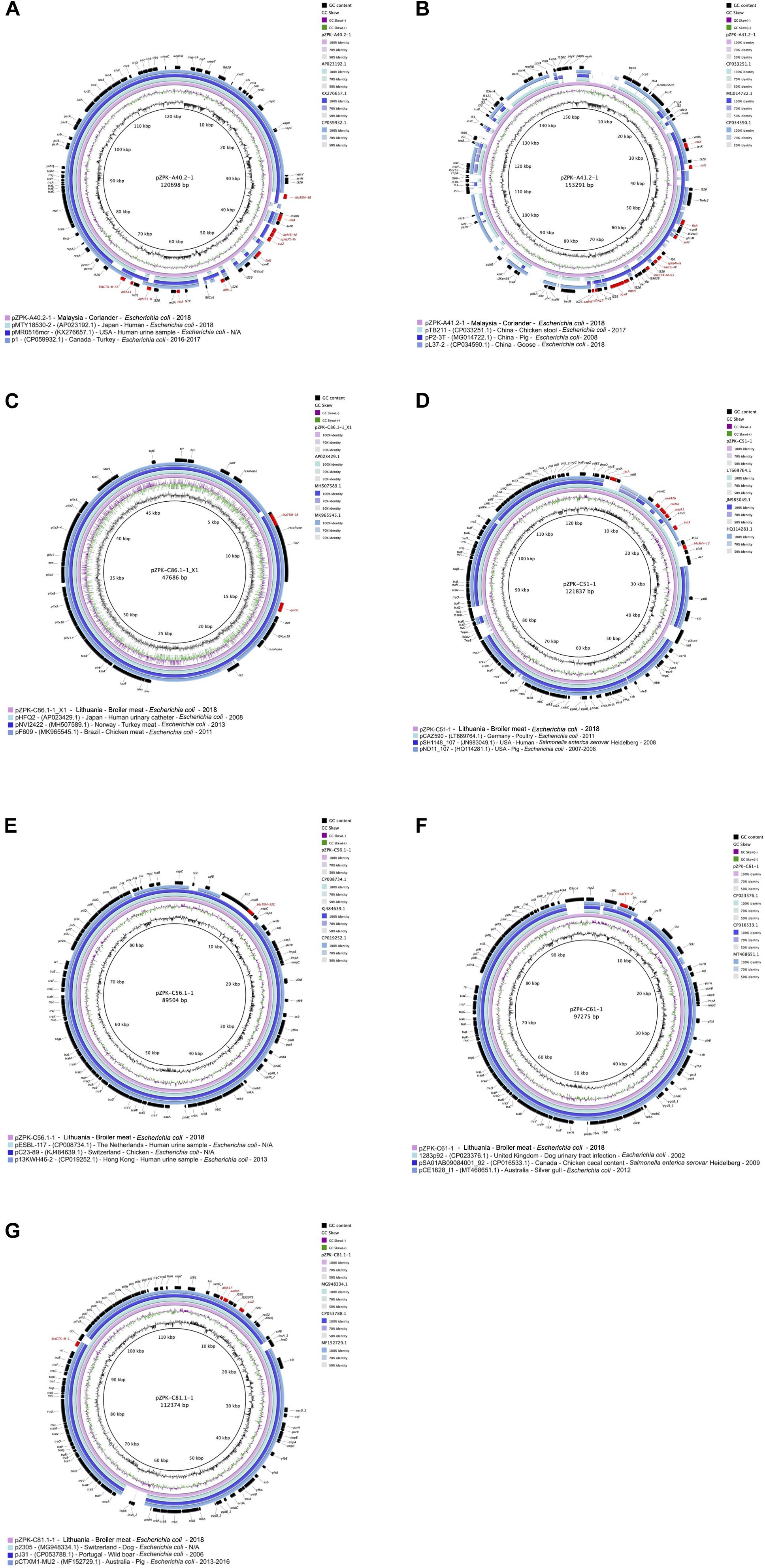
Figure 4. Circular comparisons of the studied plasmids to previously published similar plasmids (GenBank accession numbers are provided in parentheses in the figure legends after plasmid name, followed by country, source, bacterial species and year of isolation; N/A, not available). GC content and GC skew of the studied plasmids are depicted in the inner map with distance scale and the outer ring represents predicted coding sequences with antimicrobial resistance genes highlighted in red. The plasmids in this study included (A) pZPK-A40.2-1, (B) pZPK-A41.2-1, (C) pZPK-C86.1-1_X1, (D) pZPK-C51-1, (E) pZPK-C56.1-1, (F) pZPK-C61-1 and (G) pZPK-C81.1-1.
pZPK-A41.2-1 also carried multiple resistance genes in addition to the beta-lactamase blaCTX–M–65, including dfrA17, oqxB, oqxA, sul2, sul1, aac(3)-IV, aph(4)-Ia, aadA5, tet(A) and floR. The only virulence gene detected was the P fimbriae encoding papC. BLASTn search against NCBI database identified only partly similar plasmid sequences with lower coverage values than for the other plasmids in the study. pZPK-A41.2-1 aligned with 68% coverage and 99.97% identity with another IncFIB/p0111 type plasmid obtained from chicken stool in China (GenBank accession: CP033251.1). This plasmid carried a blaCTX–M–14 instead of blaCTX–M–65. Another IncFIB/p0111 plasmid with blaTEM–1B from China recovered from a goose sample (GenBank accession: CP034590.1) aligned with 64% coverage and 99.5% identity (Figure 4B).
IncX1 type plasmid (pZPK-C86.1-1_X1) was recovered from isolate C86.1-1 from raw broiler meat. The plasmid carried blaTEM–1B and PMQR gene qnrS1 and no virulence genes were identified. The size of the plasmid was 47.7 kb with G + C content 43.13% and 56 CDSs predicted. Comparison of the plasmid structure to a IncX1 reference plasmid pOLA52 (GenBank accession: EU370913.1) identified the IncX plasmid backbone in the sequenced plasmid, including replication genes pir and bis, pilus associated genes pilX and genes involved in partition (par), stability (stb) and conjugation (tax) (Johnson et al., 2012) (Figure 3B).
BLASTn search against NCBI database identified a highly similar plasmid obtained from E. coli from a human urinary catheter sample in Japan (GenBank accession: AP023429.1), aligning with 100% coverage and 99.98% identity and also carrying resistance genes blaTEM–1B and qnrS1. Another highly homologous plasmid with the same resistance profile was a plasmid obtained from turkey meat in Norway (GenBank accession: MH507589.1), aligning with 99% coverage and 99.99% identity. An IncX1 plasmid carrying blaCTX–M–15, blaTEM–1B and qnrS1 recovered from chicken meat in Brazil (GenBank accession: MK965545.1) was also found to be similar, with 97% coverage and 100% identity to pZPK-C86.1-1_X1 (Figure 4C).
IncI1-Iγ type plasmid replicons were identified from four hybrid sequenced food isolates, all originating from broiler meat from the same batch and ranging in size from 89.5 to 121.8 kb. Overall G + C content ranged in size from 50.24 to 51.65% and between 97 and 136 CDSs were predicted per plasmid. Only one of the four IncI1-Iγ plasmids harbored additional resistance genes other than a beta-lactamase; this pZPK-C51-1 blaSHV–12-harboring ST95(CC-9) plasmid with aadA1, aadA2b, tet(A), sul3, cmlA1 was the largest of the four plasmids. The only virulence genes detected in IncI1-Iγ plasmids were the channel-forming colicin gene cia or cib. Plasmid pZPK-C56.1-1 (ST36 and CC-3) carried blaTEM–52C and no virulence genes. The pndCA plasmid addiction system was detected in all the IncI1-Iγ plasmid sequences.
Aligning the plasmid sequences with BLASTn with both IncI1 type reference plasmid R64 (GenBank accession: AP005147.1) and IncIγ type reference plasmid R621a (GenBank accession: AP011954.1) indicated three of the four of the plasmids were slightly more similar to the IncIγ type R621a, which was chosen for comparative genomic visualization (Figure 3C). The plasmids in our study aligned with a query coverage of 61–85% and with >98% identity with R621a. All studied plasmids had typical IncI type backbones including the conjugational, pilus formation and maintenance and stability regions, except for blaCTX–M–1 and IS5 located near the shufflon region in pZPK-C81.1-1. The accessory module was variable between plasmids with different inserted elements and resistance genes (Figure 3C). Alignment of excA and traY regions with both references (R64 and R621a) indicated more similarity with IncI1 type R64. The parAB region was also more similar to corresponding regions of R64 in all sequenced IncI1-Iγ plasmids except for pZPK-C51-1 which shared more similarity with parAB region of R621 plasmid. The results indicate the plasmids in our study share similarities with both I1 and Iγ replicon types for which reason plasmid results are here referred to as IncI1-Iγ.
BLASTn search against NCBI database indicated highly similar plasmids have been isolated from different locations and sources, mostly from different livestock but also human clinical samples and wild animal sources (Figures 4D–G). pZPK-C56.1-1 was found to be highly similar with a plasmid of the same ST type (ST36) also carrying blaTEM–52C isolated from a human urine sample in the Netherlands (GenBank accession: CP008734.1) with a coverage of 100% and identity of 99.99%. pZPK-C56.1-1 was found to align also with another plasmid from a human urine sample from Hong Kong (GenBank accession: CP019252.1) with 91% coverage and 98.98% identity. Also, pZPK-C51-1 carrying blaSHV–12 matched with 99.76% identity and 80% coverage to a plasmid isolated from Salmonella enterica serovar Heidelberg from a human source in United States (GenBank accession: JN983049.1). blaCMY–2-harboring pZPK-C61-1 was similar with a plasmid with the same sequence type ST2 CC-2 obtained from a dog urinary tract infection sample in the United Kingdom (GenBank accession: CP023376.1), aligning with 96% coverage and 99.99% identity. pZPK-C61-1 was also similar to a plasmid isolated from a silver gull sample from Australia (GenBank accession: MT468651.1) with 93% coverage and 98.65% identity. Also, pZPK-C81.1-1 with blaCTX–M–1 matched with a plasmid obtained from a dog source in Switzerland (GenBank accession: MG948334.1) with 96% coverage and 99.92% identity and with a wild animal derived plasmid from a wild boar in Portugal (GenBank accession: CP053788.1) with 94% coverage and 99.98% identity. All IncI1-Iγ type plasmids in this study were found to match with similar plasmids from livestock-associated sources, such as a ST95 plasmid with blaSHV–12 from poultry in Germany (GenBank accession: LT669764.1) with 94% coverage and 99.97% identity with pZPK-C51-1 and ST36 plasmid with blaTEM–52C from chicken in Switzerland (GenBank accession: KJ484639.1) with 100% coverage and 99.96% identity with pZPK-C56. 1-1.
Our results demonstrate that imported food products from various origins, acquired in Finland, possess wide genetic variety of ESBL/AmpC-producing Enterobacteriaceae. Hybrid sequence analysis combining long- and short-read sequencing proved beneficial in determining resistance gene locations on specific plasmid replicons. Hybrid assembly of plasmid sequences also resolved the chromosomal location of beta-lactamase genes in two studied isolates. The spread of ESBL/pAmpC genes is highly attributed to epidemic and highly transmissible plasmids, which emphasizes the importance of plasmid replicon determination in epidemiological studies. Although the sample size was limited, our study provides a global indicative insight into AMR prevalence in global food products from 35 countries. The diversity of AMR genes was high in the isolates from food samples positive for ESBL/AmpC-producing Enterobacteriaceae. In addition, resistance toward critically important antimicrobials was found in all sequenced isolates. Raw meat, in particular broiler meat, was recognized as a common source of ESBL/AmpC-producing Enterobacteriaceae. Broiler meat samples originated from the same batch, which emphasizes the finding that one source can contain a wide variety of different resistance genes, STs, and plasmid replicons.
Compared with an earlier study investigating ESBL/AmpC-producing E. coli in Finnish poultry production, the isolates recovered from broiler meat in the current study showed broader resistance to multiple antimicrobial groups (Oikarainen et al., 2019). The isolates in the present study carried AMR genes conferring resistance against critically important antimicrobials (WHO, 2019), including aminoglycosides, third- and fourth-generation cephalosporins, macrolides, and quinolones, whereas no macrolide or PMQR was found in the isolates recovered from Finnish poultry production in the earlier study (Oikarainen et al., 2019). Another study of broilers at slaughterhouses found only limited sulfonamide resistance in addition to ESBL/AmpC with no other resistance genes in broiler meat and cecum samples (Päivärinta et al., 2020).
Our results show that vegetables, coriander and chili from Malaysia contained ESBL/AmpC-producing bacteria. Vegetables are often eaten without heating processes, leading to a greater risk of acquiring resistant bacteria. The three vegetable samples positive for ESBL/AmpC-producing Enterobacteriaceae in our study yielded a total of four isolates with varying STs, plasmid replicons, and a wide variety of beta-lactamase encoding genes, including blaCTX–M–15, blaSHV–28, blaCTX–M–55, and blaCTX–M–65. Samples from all food categories yielded many isolates, although most isolates were recovered from the vegetable samples. Citrobacter spp. and Hafnia alvei were also identified in the isolates, but these were left out of further analysis as our study focused on E. coli and K. pneumoniae, which are categorized as critical for research by the WHO (WHO, 2017b).
The finding of resistance genes blaCTX–M–15, blaSHV–12, and blaOXA–1 commonly linked to human sources (Cantón et al., 2008; Livermore et al., 2019) highlights the potential transmission route of AMR via food products and emphasizes the importance of hygiene measures. In the current study, two of the three isolates carrying blaCTX–M–15 were harboring the plasmid replicon IncFII(K), which has been previously identified from human infections with K. pneumoniae harboring blaCTX–M–15 (Dolejska et al., 2012; Bi et al., 2018). Plasmids of the IncFII type have been identified as being epidemic (Carattoli, 2009; Mathers et al., 2015). The AmpC-type resistance gene blaCMY–2 and ESBL types blaCTX–M–1, blaCTX–M–55, and blaCTX–M–65 are associated with food-producing animals, especially poultry (EFSA Panel on Biological Hazards, 2011; Cormier et al., 2019; Day et al., 2019). Interestingly, blaCMY–2 and blaCTX–M–1 were recovered from broiler meat samples and blaCTX–M–55, and blaCTX–M–65 from vegetable samples in our study. One explanation for the finding of AMR genes associated with food-producing animals in food products could be the possible use of contaminated irrigation water or animal manure in the farming process. As low-income countries have higher AMR abundance in their wastewater (Hendriksen et al., 2019), it can be speculated that the risk of ESBL/AmpC dissemination is greater, especially when water is contaminated by animal manure or wastewater from human sources is used for crop watering. However, no conclusions can be drawn from the current study regarding the possible differences in ESBL/AmpC-producing Enterobacteriaceae prevalence between different countries, because the sample size per country was limited. More attention should be paid to the comparison of different food categories from the results of the current study.
Food samples were found to harbor a variety of bacterial STs, some of which have been described from human infections, i.e., ST37 from Chinese strains (Zhu et al., 2015; Zhang et al., 2016; Xiao et al., 2017) and ST307 from Italian, Colombian, and United Kingdom strains (Villa et al., 2017) of K. pneumoniae and ST38 from Bangladeshi and United Kingdom strains of E. coli (Hasan et al., 2015; Day et al., 2019). Also, same ST types (ST88 and ST117) found in the Lithuanian broiler meat in the present study have been previously found also in Finnish broiler production (Heikinheimo et al., 2016; Oikarainen et al., 2019) and ST117 more widely in Nordic broiler production (Ronco et al., 2017). In a Canadian study (Vincent et al., 2010), E. coli ST117 isolates were identified from human infections and poultry sources with related pulsed-field gel electrophoresis profiles, pointing to a possible poultry source of human infections. With the large variety of different bacterial STs, it is difficult to draw any further conclusions about possible bacterial transmission routes from our study, but it seems that certain E. coli STs and their resistance genes are common in international food production and are found in food products produced in various countries. Noteworthy is the finding of clinically relevant bacterial STs.
Plasmid replicons linked to the spread of AMR, particularly of the IncI type (Carattoli, 2013), were detected in our study. IncI1-Iγ ST12 was recovered from three broiler isolates, and it has been previously linked to blaSHV–12 from avian E. coli (Accogli et al., 2013). In addition, IncI plasmids with the blaCTX–M–1 gene have been found in E. coli from poultry and humans in the Netherlands (Leverstein-van Hall et al., 2011). Also, plasmids of the IncF group were detected in 90% (19/21) of the sequenced isolates. Plasmids of the IncF type are one of the most common Inc types identified in humans and animals, especially in Asia, and most often carry AMR genes of the bla family (Rozwandowicz et al., 2018).
Long-read sequencing and hybrid assembly with Illumina short reads allowed more in-depth analysis of plasmids harboring beta-lactamases in our study. Hybrid assembly of the sequences resolved the correct plasmid replicons carrying bla genes, which would not have been possible with confidence from the fragmented short read data. As new assembly tools, such as Trycycler3 have been developed to resolve genomes from WGS data it is important to bear in mind the requirements for high-quality assemblies with these new tools, such as high enough coverage of the long reads. With the advancement and improved availability of long-read sequencing technologies plasmid analyses regarding AMR studies should prove to be even more affordable and accessible in the future.
Our analysis comparing plasmids carrying bla genes in this study with plasmid sequences deposited in open databases showed the plasmids in our study were similar with mostly livestock-derived plasmids obtained previously from various countries, but also with human clinical samples, especially from urinary tract infections. pZPK-A41.2-1 carried the P fimbriae encoding virulence gene papC which has been associated with uropathogenic E. coli infections (Yazdanpour et al., 2020). pZPK-A40.2-1 was also similar to a mcr-1-carrying plasmid isolated from a human urine sample from the United States (GenBank accession: KX276657.1), indicating a potential for IncF type plasmids being able to obtain new resistance genes. pZPK-C86.1-1_X1 was found the be a typical IncX1 plasmid in regards of carrying the PMQR gene qnrS1, which has been linked to Salmonella and E. coli from animal and human sources (Dobiasova and Dolejska, 2016). This plasmid was also found to be highly similar to a plasmid isolated from a human urinary catheter sample in Japan (GenBank accession: AP023429.1).
Five of the hybrid assembled plasmid sequences were obtained from raw broiler meat samples originating from the same batch number. Four of these carried the IncI-Iγ type plasmid, but all harbored a different bla gene, indicating diversity even among plasmids obtained from a homogenous origin. The similarity of all of the seven hybrid assembled plasmids, even from vegetable origin, to poultry and other livestock sources published previously also indicates certain plasmid-resistance gene combinations flourish in specific host environments.
Similarity to plasmids from small animal sources was also observed. pZPK-C61-1 of the IncI1-Iγ type with blaCMY–2 was highly similar to a plasmid obtained from a canine urinary tract infection sample in the UK (GenBank accession: CP023376.1). pZPK-C81.1-1 with blaCTX–M–1 also of IncI1-Iγ type was also similar to a blaCTX–M–1-carrying plasmid obtained from E. coli from a dog in Switzerland (GenBank accession: MG948334.1). The finding of similar transmissible plasmids from food products and small animals indicates a possible transmission route between food, animals and potentially humans who are in close contact with pets. This highlights the need for plasmid studies implementing a One Health approach, since horizontal gene transmission via plasmids does not know country or species borders.
Interestingly, long-read sequencing and hybrid assembly revealed chromosomally located beta-lactamases in two of the sequenced isolates. Long-read sequencing provides reliable information on the true location of resistance genes, as plasmid replicons and genes can be matched together. As plasmids have been recognized as successful drivers of AMR, more studies combining short- and long-read sequencing are needed in order to sequence plasmids as a whole (Valcek et al., 2019) and to gain a deeper knowledge of the multifactorial epidemiology behind the spread of AMR.
Interestingly, no ESBL/AmpC-producing Enterobacteriaceae were obtained from fruit or seafood samples in our study. It is noteworthy, however, that sample sizes were limited. In another study conducted in Spain, raw fish products and sushi were found to have a prevalence of ESBL-producing Enterobacteriaceae of 10.6 and 19.4%, respectively (Vitas et al., 2018). Local fish products in Vietnam have been found to have an ESBL-producing E. coli prevalence of 62.5% with a high level of multidrug resistance (Le et al., 2015). Other AMR bacteria have also been recovered from seafood products in previous studies (Boss et al., 2016; Ellis-Iversen et al., 2019). In our study, seafood samples consisted largely of frozen or precooked products, which may, together with limited sample size, explain the absence of ESBL producers. Fortunately, no cross contamination with ESBL/AmpC-producing Enterobacteriaceae was found in cooked meat products, as these products are often eaten without additional heating, posing a greater risk of transmission of bacteria. The finding is in line with an earlier study conducted in Spain (Vitas et al., 2018) but not in line with a study by Jiang et al. (2014), which found a 6.7% prevalence of ESBL producers in E. coli isolates recovered from cooked meat products in China. blaCTX–M–1, blaCTX–M–9, and blaTEM–1 were identified in the latter study, which may point to inadequate hygiene measures after heat processing or cross contamination, in which bacteria are introduced to products after heat processing. We did not recover any carbapenemase producing Enterobacteriaceae (CPE) in our study, although CPE have been recovered previously from meat, vegetable, and seafood products of Southeast Asian origin (Zurfluh et al., 2015a; Janecko et al., 2016; Sugawara et al., 2019). However, the selection with cefotaxime in the pre-enrichment of samples in our study might cause carbapenemase producing strains to be missed.
Earlier studies investigating the presence of ESBL/AmpC-producing bacteria in food products have partly differing results. A German study found only one out of 399 vegetable samples to be positive for cefotaxime-resistant E. coli (Kaesbohrer et al., 2019) but found cefotaxime-resistant E. coli in 74.9% of chicken meat and 40.1% of turkey meat. This is in line with our results, pointing to a higher prevalence of resistant Enterobacteriaceae in broiler meat and limited prevalence in vegetables. However, a Swiss study found 25.4% of vegetables imported from the Dominican Republic, India, Thailand, and Vietnam positive for ESBL-producing Enterobacteriaceae (Zurfluh et al., 2015b). This is remarkably higher than in our study with 5% of vegetable samples being positive for ESBL/AmpC-producing Enterobacteriaceae, although our study included samples from around the world, not only Asia and South America. In these continents, however, wastewater of human origin is commonly used for agriculture (Jiménez, 2006; Raschid-Sally and Jayakody, 2008). Using contaminated irrigation water may serve as a direct link to AMR contamination, especially if the water is applied to edible parts of fruits and vegetables (FAO and WHO, 2019). Antimicrobial-resistant bacteria have also been recovered from frozen products, vegetables, and ready-to-eat products from Chinese retail food (Ye et al., 2018). Ready-to-eat vegetables, in particular sprouts, have been previously recognized as a possible source of ESBL/AmpC-producing E. coli and K. pneumoniae (Kim et al., 2015).
Although food is not considered as the main source of human ESBL infections (Belmar Campos et al., 2014; Carmo et al., 2014; Börjesson et al., 2016; Day et al., 2019), similar ESBL strains have been occasionally found from human infections and foods (Irrgang et al., 2017; Yamaji et al., 2018; Day et al., 2019; Roer et al., 2019; Valcek et al., 2019). In a recent population-based modeling study, 18.9% of human ESBL/AmpC-E. coli carriage was attributed to a food source (Mughini-Gras et al., 2019), highlighting the importance of AMR surveillance and proper hygiene measures for food products. Our results also show highly similar plasmids have been previously identified from human samples, indicating a possible transmission route of ESBL-harboring plasmids via food sources.
Antimicrobial resistance is a global problem with an uneven distribution around the globe and an accumulated burden especially in low-income countries (Alsan et al., 2015; Seale et al., 2017; Hendriksen et al., 2019). Although the burden of AMR is not equal in all countries, globalization, traffic of people, animals, and food enables resistant bacteria to disseminate around the world. To track the spread of AMR, it is of vital importance to execute AMR surveillance programs and to study different food products from various food categories in order to detect changes in the distribution of resistant bacteria, genes, and plasmids. As plasmids have been recognized as important drivers of AMR, efforts should be put into developing rapid WGS pipelines able to accurately detect plasmids carrying antimicrobial resistance genes of high importance.
Food products from different food categories around the world contain a wide variety of bacterial species resistant to third-generation cephalosporins among other critically important antimicrobials, with poultry meat serving as a rich reservoir of ESBL/AmpC-producing E. coli and K. pneumoniae. Although raw poultry meat was identified as the most common source of ESBL/AmpC-producing Enterobacteriaceae, the sporadic finding of resistant bacteria from fresh vegetable samples highlights the need for diverse One Health surveillance of AMR from multiple sources. Further studies should focus on identifying plasmids, resistance and virulence genes, and bacterial STs in food products relevant for human infections and food safety. The finding of high similarity between food-derived plasmids carrying bla genes with plasmids recovered previously from various sources globally highlights the risks posed by international trade, such as food products.
The datasets generated for this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
PK, AH, MA, SN, EV, and A-LM contributed to the concept and design of the study. PK collected samples. PK and SN contributed to bacterial laboratory analysis. BK, MB, PK, and AH analyzed whole genome sequence data. MB and PK performed long-read sequencing. BK, PK, and MB performed subsequent analysis. BK and PK made visualizations for plasmid comparisons. PK and AH drafted the manuscript. MB, MA, and SN revised the manuscript. All authors have read and approved the final draft of the manuscript.
Financial support was obtained from the Walter Ehrström Foundation, the Finnish Food Research Foundation, and the European Union’s Horizon 2020 Research and Innovation Program under grant agreement no. 773830 OneHealth EJP: FULL-FORCE.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank senior laboratory technicians Kirsi Ristkari and Arie Kant and laboratory analyst students Veronika Konstantinova, Noora Nenonen, and Santeri Soininen for assistance with laboratory work. We are also grateful for the valuable and helpful discussions with Hanna Castro.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2021.592291/full#supplementary-material
Accogli, M., Fortini, D., Giufrè, M., Graziani, C., Dolejska, M., Carattoli, A., et al. (2013). IncI1 plasmids associated with the spread of CMY-2, CTX-M-1 and SHV-12 in Escherichia coli of animal and human origin. Clin. Microbiol. Infect. 19, E238–E240. doi: 10.1111/1469-0691.12128
Alikhan, N. F., Petty, N. K., Ben Zakour, N. L., and Beatson, S. A. (2011). BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12:402. doi: 10.1186/1471-2164-12-402
Alsan, M., Schoemaker, L., Eggleston, K., Kammili, N., Kolli, P., and Bhattacharya, J. (2015). Out-of-pocket health expenditures and antimicrobial resistance in low-income and middle-income countries: an economic analysis. Lancet Infect. Dis. 15, 1203–1210. doi: 10.1016/S1473-3099(15)00149-8
Belmar Campos, C., Fenner, I., Wiese, N., Lensing, C., Christner, M., Rohde, H., et al. (2014). Prevalence and genotypes of extended spectrum beta-lactamases in Enterobacteriaceae isolated from human stool and chicken meat in Hamburg, Germany. Int. J. Med. Microbiol. 304, 678–684.
Bi, D., Zheng, J., Li, J. J., Sheng, Z. K., Zhu, X., Ou, H. Y., et al. (2018). In silico typing and comparative genomic analysis of incfiik plasmids and insights into the evolution of replicons, plasmid backbones, and resistance determinant profiles. Antimicrob. Agents Chemother. 62:e00764-18. doi: 10.1128/AAC.00764-18
Börjesson, S., Ny, S., Egervärn, M., Bergström, J., Rosengren, Å., Englund, S., et al. (2016). Limited Dissemination of Extended-Spectrum β-Lactamase- and Plasmid-Encoded AmpC-Producing Escherichia coli from Food and Farm Animals, Sweden. Emerg. Infect. Dis. 22, 634–640. doi: 10.3201/eid2204.151142
Boss, R., Overesch, G., and Baumgartner, A. (2016). Antimicrobial Resistance of Escherichia coli, Enterococci, Pseudomonas aeruginosa, and Staphylococcus aureus from Raw Fish and Seafood Imported into Switzerland. J. Food Prot. 79, 1240–1246. doi: 10.4315/0362-028X.JFP-15-463
Cantón, R., Novais, A., Valverde, A., Machado, E., Peixe, L., Baquero, F., et al. (2008). Prevalence and spread of extended-spectrum β-lactamase-producing Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 14, 144–153. doi: 10.1111/J.1469-0691.2007.01850.X
Carattoli, A. (2009). Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 53, 2227–2238. doi: 10.1128/AAC.01707-08
Carattoli, A. (2013). Plasmids and the spread of resistance. Int. J. Med. Microbiol. 303, 298–304. doi: 10.1016/J.IJMM.2013.02.001
Carattoli, A., Zankari, E., García-Fernández, A., Voldby Larsen, M., Lund, O., Villa, L., et al. (2014). In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58, 3895–3903. doi: 10.1128/AAC.02412-14
Carmo, L. P., Nielsen, L. R., da Costa, P. M., and Alban, L. (2014). Exposure assessment of extended-spectrum beta-lactamases/AmpC beta-lactamases-producing Escherichia coli in meat in Denmark. Infect. Ecol. Epidemiol. 4:10.3402/iee.v4.22924. doi: 10.3402/iee.v4.22924
Chantziaras, I., Boyen, F., Callens, B., and Dewulf, J. (2014). Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: a report on seven countries. J. Antimicrob. Chemother. 69, 827–834. doi: 10.1093/jac/dkt443
Clausen, P. T. L. C., Aarestrup, F. M., and Lund, O. (2018). Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinformatics 19:307. doi: 10.1186/s12859-018-2336-6
Cormier, A., Zhang, P. L. C., Chalmers, G., Weese, J. S., Deckert, A., Mulvey, M., et al. (2019). Diversity of CTX-M-positive Escherichia coli recovered from animals in Canada. Vet. Microbiol. 231, 71–75. doi: 10.1016/J.VETMIC.2019.02.031
Day, M. J., Hopkins, K. L., Wareham, D. W., Toleman, M. A., Elviss, N., Randall, L., et al. (2019). Extended-spectrum β-lactamase-producing Escherichia coli in human-derived and foodchain-derived samples from England, Wales, and Scotland: an epidemiological surveillance and typing study. Lancet Infect. Dis. 19, 1325–1335. doi: 10.1016/S1473-3099(19)30273-7
Dobiasova, H., and Dolejska, M. (2016). Prevalence and diversity of IncX plasmids carrying fluoroquinolone and β-lactam resistance genes in Escherichia coli originating from diverse sources and geographical areas. J. Antimicrob. Chemother. 71, 2118–2124. doi: 10.1093/jac/dkw144
Dolejska, M., Brhelova, E., Dobiasova, H., Krivdova, J., Jurankova, J., Sevcikova, A., et al. (2012). Dissemination of IncFIIK-type plasmids in multiresistant CTX-M-15-producing Enterobacteriaceae isolates from children in hospital paediatric oncology wards. Int. J. Antimicrob. Agents 40, 510–515. doi: 10.1016/j.ijantimicag.2012.07.016
Done, H. Y., Venkatesan, A. K., and Halden, R. U. (2015). Does the recent growth of aquaculture create antibiotic resistance threats different from those associated with land animal production in agriculture? AAPS J. 17, 513–524. doi: 10.1208/s12248-015-9722-z
EFSA and ECDC (2020). The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017/2018. EFSA J. 18:6007. doi: 10.2903/j.efsa.2020.6007
EFSA Panel on Biological Hazards (2011). Scientific Opinion on the public health risks of bacterial strains producing extended-spectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 9:2322. doi: 10.2903/j.efsa.2011.2322
Ellis-Iversen, J., Seyfarth, A. M., Korsgaard, H., Bortolaia, V., Munck, N., and Dalsgaard, A. (2019). Antimicrobial resistant E. coli and enterococci in pangasius fillets and prawns in Danish retail imported from Asia. Food Control 114:106958. doi: 10.1016/J.FOODCONT.2019.106958
EUCAST (2017). European Committee on Antimicrobial Susceptibility Testing. EUCAST Disk Diffusion Method Manual 6.0. Available online at: https://www.eucast.org/ (accessed December 19, 2018).
European Centre for Disease Prevention and Control [ECDC], European Food Safety Authority [EFSA], and European Medicines Agency [EMA] (2017). ECDC/EFSA/EMA second joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals – Joint Interagency Antimicrobial Consumption and Resistance Analysis (JIACRA) Report. EFSA J. 15:e4872. doi: 10.2903/j.efsa.2017.4872
European Medicines Agency [EMA] (2019). European Surveillance of Veterinary Antimicrobial Consumption (ESVAC), 2019. Sales of Veterinary Antimicrobial agents in 31 European Countries in 2017. Trends from 2010 to 2017. Ninth ESVAC report. Amsterdam: EMA.
Ewers, C., Bethe, A., Semmler, T., Guenther, S., and Wieler, L. H. (2012). Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin. Microbiol. Infect. 18, 646–655. doi: 10.1111/j.1469-0691.2012.03850.x
FAO and WHO (2019). Joint FAO/WHO Expert Meeting in Collaboration with OIE on Foodborne Antimicrobial Resistance: Role of the Environment, Crops and Biocides – Meeting report. Microbiological Risk Assessment Series no. 34. Rome: FAO.
Frankel, G., Phillips, A. D., Rosenshine, I., Dougan, G., Kaper, J. B., and Knutton, S. (1998). Enteropathogenic and enterohaemorrhagic Escherichia coli: more subversive elements. Mol. Microbiol. 30, 911–921. doi: 10.1046/j.1365-2958.1998.01144.x
Frasson, I., Cavallaro, A., Bergo, C., Richter, S. N., and Palù, G. (2011). Prevalence of aac(6′)-Ib-cr plasmid-mediated and chromosome-encoded fluoroquinolone resistance in Enterobacteriaceae in Italy. Gut Pathog. 3:12. doi: 10.1186/1757-4749-3-12
Hartmann, A., Locatelli, A., Amoureux, L., Depret, G., Jolivet, C., Gueneau, E., et al. (2012). Occurrence of CTX-M Producing Escherichia coli in Soils, Cattle, and Farm Environment in France (Burgundy Region). Front. Microbiol. 3:83. doi: 10.3389/fmicb.2012.00083
Hasan, B., Olsen, B., Alam, A., Akter, L., and Melhus, Å (2015). Dissemination of the multidrug-resistant extended-spectrum β-lactamase-producing Escherichia coli O25b-ST131 clone and the role of house crow (Corvus splendens) foraging on hospital waste in Bangladesh. Clin. Microbiol. Infect. 21, 1000.e1–1000.e4. doi: 10.1016/J.CMI.2015.06.016
Hasman, H., Saputra, D., Sicheritz-Ponten, T., Lund, O., Svendsen, C. A., Frimodt-Møller, N., et al. (2014). Rapid whole-genome sequencing for detection and characterization of microorganisms directly from clinical samples. J. Clin. Microbiol. 52, 139–146. doi: 10.1128/JCM.02452-13
Heikinheimo, A., Latvio, S.-M., Päivärinta, M., and Fredriksson-Ahomaa, M. (2016). “Identification of blaCMY-2 and blaCTX-M-1 carrying Escherichia coli in slaughterhouse broilers and broiler meat in Finland – whole genome sequencing reveals high genetic diversity with multiple MLST types,” in Proceedings of the 4th International Conference on Responsible Use of Antibiotics in Animals, Hague.
Hendriksen, R. S., Munk, P., Njage, P., van Bunnik, B., McNally, L., Lukjancenko, O., et al. (2019). Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 10:1124. doi: 10.1038/s41467-019-08853-3
Hoelzer, K., Wong, N., Thomas, J., Talkington, K., Jungman, E., and Coukell, A. (2017). Antimicrobial drug use in food-producing animals and associated human health risks: What, and how strong, is the evidence? BMC Vet. Res. 13:211. doi: 10.1186/s12917-017-1131-3
Irrgang, A., Falgenhauer, L., Fischer, J., Ghosh, H., Guiral, E., Guerra, B., et al. (2017). CTX-M-15-Producing E. coli isolates from food products in Germany are mainly associated with an IncF-Type plasmid and belong to two predominant clonal E. coli lineages. Front. Microbiol. 8:2318. doi: 10.3389/fmicb.2017.02318
Janecko, N., Martz, S.-L., Avery, B. P., Daignault, D., Desruisseau, A., Boyd, D., et al. (2016). Carbapenem-Resistant Enterobacter spp. in Retail Seafood Imported from Southeast Asia to Canada. Emerg. Infect. Dis. 22, 1675–1677. doi: 10.3201/eid2209.160305
Jaureguy, F., Landraud, L., Passet, V., Diancourt, L., Frapy, E., Guigon, G., et al. (2008). Phylogenetic and genomic diversity of human bacteremic Escherichia coli strains. BMC Genomics 9:560. doi: 10.1186/1471-2164-9-560
Jiang, X., Yu, T., Wu, N., Meng, H., and Shi, L. (2014). Detection of qnr, aac(6’)-Ib-cr and qepA genes in Escherichia coli isolated from cooked meat products in Henan, China. Int. J. Food Microbiol. 187, 22–25. doi: 10.1016/J.IJFOODMICRO.2014.06.026
Jiménez, B. (2006). Irrigation in Developing Countries Using Wastewater. Available online at: https://www.semanticscholar.org/paper/Irrigation-in- Developing-Countries-Using-Wastewater-Jiménez/cfdb21d4680b59e12f057ba 2757315d72e7820b7 (accessed December 4, 2019).
Joensen, K. G., Scheutz, F., Lund, O., Hasman, H., Kaas, R. S., Nielsen, E. M., et al. (2014). Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Microbiol. 52, 1501–1510. doi: 10.1128/JCM.03617-13
Johansson, M. H. K., Bortolaia, V., Tansirichaiya, S., Aarestrup, F. M., Roberts, A. P., and Petersen, T. N. (2021). Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 76, 101–109. doi: 10.1093/jac/dkaa390
Johnson, T. J., Bielak, E. M., Fortini, D., Hansen, L. H., Hasman, H., Debroy, C., et al. (2012). Expansion of the IncX plasmid family for improved identification and typing of novel plasmids in drug-resistant Enterobacteriaceae. Plasmid 68, 43–50. doi: 10.1016/j.plasmid.2012.03.001
Kaesbohrer, A., Bakran-Lebl, K., Irrgang, A., Fischer, J., Kämpf, P., Schiffmann, A., et al. (2019). Diversity in prevalence and characteristics of ESBL/pAmpC producing E. coli in food in Germany. Vet. Microbiol. 233, 52–60. doi: 10.1016/J.VETMIC.2019.03.025
Kim, H.-S., Chon, J.-W., Kim, Y.-J., Kim, D.-H., Kim, M., and Seo, K.-H. (2015). Prevalence and characterization of extended-spectrum-β-lactamase-producing Escherichia coli and Klebsiella pneumoniae in ready-to-eat vegetables. Int. J. Food Microbiol. 207, 83–86. doi: 10.1016/J.IJFOODMICRO.2015.04.049
Larsen, M. V., Cosentino, S., Lukjancenko, O., Saputra, D., Rasmussen, S., Hasman, H., et al. (2014). Benchmarking of methods for genomic taxonomy. J. Clin. Microbiol. 52, 1529–1539. doi: 10.1128/JCM.02981-13
Larsen, M. V., Cosentino, S., Rasmussen, S., Friis, C., Hasman, H., Marvig, R. L., et al. (2012). Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 50, 1355–1361. doi: 10.1128/jcm.06094-11
Laupland, K. B., Church, D. L., Vidakovich, J., Mucenski, M., and Pitout, J. D. D. (2008). Community-onset extended-spectrum β-lactamase (ESBL) producing Escherichia coli: importance of international travel. J. Infect. 57, 441–448. doi: 10.1016/J.JINF.2008.09.034
Le, H. V., Kawahara, R., Khong, D. T., Tran, H. T., Nguyen, T. N., Pham, K. N., et al. (2015). Widespread dissemination of extended-spectrum β-lactamase-producing, multidrug-resistant Escherichia coli in livestock and fishery products in Vietnam. Int. J. Food Contam. 2:17. doi: 10.1186/s40550-015-0023-1
Leverstein-van Hall, M. A., Dierikx, C. M., Stuart, J. C., Voets, G. M., van den Munckhof, M. P., van Essen-Zandbergen, A., et al. (2011). Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17, 873–880. doi: 10.1111/J.1469-0691.2011.03497.X
Livermore, D. M., Day, M., Cleary, P., Hopkins, K. L., Toleman, M. A., Wareham, D. W., et al. (2019). OXA-1 β-lactamase and non-susceptibility to penicillin/β-lactamase inhibitor combinations among ESBL-producing Escherichia coli. J. Antimicrob. Chemother. 74, 326–333. doi: 10.1093/jac/dky453
Marshall, B. M., and Levy, S. B. (2011). Food animals and antimicrobials: impacts on human health. Clin. Microbiol. Rev. 24, 718–733. doi: 10.1128/CMR.00002-11
Mathers, A. J., Peirano, G., and Pitout, J. D. D. (2015). The role of epidemic resistance plasmids and international high- risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin. Microbiol. Rev. 28, 565–591. doi: 10.1128/CMR.00116-14
Mughini-Gras, L., Dorado-García, A., van Duijkeren, E., van den Bunt, G., Dierikx, C. M., Bonten, M. J. M., et al. (2019). Attributable sources of community-acquired carriage of Escherichia coli containing β-lactam antibiotic resistance genes: a population-based modelling study. Lancet Planet. Health 3, e357–e369. doi: 10.1016/S2542-5196(19)30130-5
Müller, A., Stephan, R., and Nüesch-Inderbinen, M. (2016). Distribution of virulence factors in ESBL-producing Escherichia coli isolated from the environment, livestock, food and humans. Sci. Total Environ. 541, 667–672. doi: 10.1016/j.scitotenv.2015.09.135
Nurk, S., Bankevich, A., Antipov, D., Gurevich, A., Korobeynikov, A., Lapidus, A., et al. (2013). “Assembling Genomes and Mini-metagenomes from Highly Chimeric Reads,” in Research in Computational Molecular Biology. RECOMB 2013. Lecture Notes in Computer Science, Vol. 7821, eds M. Deng, R. Jiang, F. Sun, and X. Zhang (Heidelberg: Springer), 158–170. doi: 10.1007/978-3-642-37195-0_13
Oikarainen, P. E., Pohjola, L. K., Pietola, E. S., and Heikinheimo, A. (2019). Direct vertical transmission of ESBL/pAmpC-producing Escherichia coli limited in poultry production pyramid. Vet. Microbiol. 231, 100–106. doi: 10.1016/j.vetmic.2019.03.001
Päivärinta, M., Latvio, S., Fredriksson-Ahomaa, M., and Heikinheimo, A. (2020). Whole genome sequence analysis of antimicrobial resistance genes, multilocus sequence types and plasmid sequences in ESBL/AmpC Escherichia coli isolated from broiler caecum and meat. Int. J. Food Microbiol. 315:108361. doi: 10.1016/J.IJFOODMICRO.2019.108361
Paterson, D. L., and Bonomo, R. A. (2005). Extended-spectrum β-lactamases: a clinical update. Clin. Microbiol. Rev. 18, 657–686. doi: 10.1128/CMR.18.4.657-686.2005
Pitout, J. D., and Laupland, K. B. (2008). Extended-spectrum β-lactamase-producing Enterobacteriaceae: an emerging public-health concern. Lancet Infect. Dis. 8, 159–166. doi: 10.1016/S1473-3099(08)70041-0
Raschid-Sally, L., and Jayakody, P. (2008). Drivers and Characteristics of Wastewater Agriculture in Developing Countries: Results from a Global Assessment. IWMI Res. Reports. Available online at: https://ideas.repec.org/p/iwt/rerpts/h041686.html (Accessed December 4, 2019).
Reinthaler, F. F., Feierl, G., Galler, H., Haas, D., Leitner, E., Mascher, F., et al. (2010). ESBL-producing E. coli in Austrian sewage sludge. Water Res. 44, 1981–1985. doi: 10.1016/j.watres.2009.11.052
Roer, L., Overballe-Petersen, S., Hansen, F., Johannesen, T. B., Stegger, M., Bortolaia, V., et al. (2019). ST131 fimH 22 Escherichia coli isolate with a bla CMY-2/IncI1/ST12 plasmid obtained from a patient with bloodstream infection: highly similar to E. coli isolates of broiler origin. J. Antimicrob. Chemother. 74, 557–560. doi: 10.1093/jac/dky484
Ronco, T., Stegger, M., Olsen, R. H., Sekse, C., Nordstoga, A. B., Pohjanvirta, T., et al. (2017). Spread of avian pathogenic Escherichia coli ST117 O78:H4 in Nordic broiler production. BMC Genomics 18:13. doi: 10.1186/s12864-016-3415-6
Rozwandowicz, M., Brouwer, M. S. M., Fischer, J., Wagenaar, J. A., Gonzalez-Zorn, B., Guerra, B., et al. (2018). Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 73, 1121–1137. doi: 10.1093/jac/dkx488
Schar, D., Sommanustweechai, A., Laxminarayan, R., and Tangcharoensathien, V. (2018). Surveillance of antimicrobial consumption in animal production sectors of low- and middle-income countries: optimizing use and addressing antimicrobial resistance. PLoS Med. 15:e1002521. doi: 10.1371/journal.pmed.1002521
Seale, A. C., Gordon, N. C., Islam, J., Peacock, S. J., and Scott, J. A. G. (2017). AMR surveillance in low and middle-income settings - A roadmap for participation in the Global Antimicrobial Surveillance System (GLASS). Wellcome Open Res. 2:92. doi: 10.12688/wellcomeopenres.12527.1
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Sugawara, Y., Hagiya, H., Akeda, Y., Aye, M. M., Myo Win, H. P., Sakamoto, N., et al. (2019). Dissemination of carbapenemase-producing Enterobacteriaceae harbouring blaNDM or blaIMI in local market foods of Yangon, Myanmar. Sci. Rep. 9:14455. doi: 10.1038/s41598-019-51002-5
Sullivan, M. J., Petty, N. K., and Beatson, S. A. (2011). Easyfig: a genome comparison visualizer. Bioinformatics 27, 1009–1010. doi: 10.1093/bioinformatics/btr039
Tang, K. L., Caffrey, N. P., Nóbrega, D. B., Cork, S. C., Ronksley, P. E., Barkema, H. W., et al. (2017). Restricting the use of antibiotics in food-producing animals and its associations with antibiotic resistance in food-producing animals and human beings: a systematic review and meta-analysis. Lancet Planet. Health 1, e316–e327. doi: 10.1016/S2542-5196(17)30141-9
Valcek, A., Roer, L., Overballe-Petersen, S., Hansen, F., Bortolaia, V., Leekitcharoenphon, P., et al. (2019). IncI1 ST3 and IncI1 ST7 plasmids from CTX-M-1-producing Escherichia coli obtained from patients with bloodstream infections are closely related to plasmids from E. coli of animal origin. J. Antimicrob. Chemother. 74, 2171–2175. doi: 10.1093/jac/dkz199
Van Boeckel, T. P., Brower, C., Gilbert, M., Grenfell, B. T., Levin, S. A., Robinson, T. P., et al. (2015). Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. U.S.A. 112, 5649–5654. doi: 10.1073/pnas.1503141112
Van Boeckel, T. P., Pires, J., Silvester, R., Zhao, C., Song, J., Criscuolo, N. G., et al. (2019). Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 365:eaaw1944. doi: 10.1126/science.aaw1944
Villa, L., Feudi, C., Fortini, D., Brisse, S., Passet, V., Bonura, C., et al. (2017). Diversity, virulence, and antimicrobial resistance of the KPC-producing Klebsiella pneumoniae ST307 clone. Microb. Genomics 3:e000110. doi: 10.1099/mgen.0.000110
Vincent, C., Boerlin, P., Daignault, D., Dozois, C. M., Dutil, L., Galanakis, C., et al. (2010). Food reservoir for Escherichia coli causing urinary tract infections. Emerg. Infect. Dis. 16, 88–95. doi: 10.3201/eid1601.091118
Vitas, A. I., Naik, D., Pérez-Etayo, L., and González, D. (2018). Increased exposure to extended-spectrum β-lactamase-producing multidrug-resistant Enterobacteriaceae through the consumption of chicken and sushi products. Int. J. Food Microbiol. 269, 80–86. doi: 10.1016/J.IJFOODMICRO.2018.01.026
WHO (2017a). Guidelines on Use of Medically Important Antimicrobials in Food-Producing Animals: Policy Brief. Geneva: World Health Organization.
WHO (2017b). Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug Resistant Bacterial Infections, Including Tuberculosis. Geneva: World Health Organization.
WHO (2019). Critically Important Antimicrobials for Human Medicine, 6th Revision. Geneva: World Health Organization.
WHO FAO and OIE (2018). Monitoring Global Progress on Addressing Antimicrobial Resistance: Analysis Report of the Second Round of Results of AMR Country Self-Assessment Survey 2018. Available online at: https://www.who.int/antimicrobial-resistance/publications/Analysis-report-of-AMR-country-se/en/ (accessed December 4, 2019).
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 13:e1005595. doi: 10.1371/journal.pcbi.1005595
Wirth, T., Falush, D., Lan, R., Colles, F., Mensa, P., Wieler, L. H., et al. (2006). Sex and virulence in Escherichia coli: an evolutionary perspective. Mol. Microbiol. 60, 1136–1151. doi: 10.1111/j.1365-2958.2006.05172.x
Witte, W. (2000). Ecological impact of antibiotic use in animals on different complex microflora: environment. Int. J. Antimicrob. Agents 14, 321–325. doi: 10.1016/S0924-8579(00)00144-8
Xiao, S.-Z., Wang, S., Wu, W.-M., Zhao, S.-Y., Gu, F.-F., Ni, Y.-X., et al. (2017). The resistance phenotype and molecular epidemiology of Klebsiella pneumoniae in Bloodstream Infections in Shanghai, China, 2012-2015. Front. Microbiol. 8:250. doi: 10.3389/fmicb.2017.00250
Yamaji, R., Friedman, C. R., Rubin, J., Suh, J., Thys, E., McDermott, P., et al. (2018). A population-based surveillance study of shared genotypes of Escherichia coli isolates from retail meat and suspected cases of urinary tract infections. mSphere 3:e00179-18. doi: 10.1128/mSphere.00179-18
Yazdanpour, Z., Tadjrobehkar, O., and Shahkhah, M. (2020). Significant association between genes encoding virulence factors with antibiotic resistance and phylogenetic groups in community acquired uropathogenic Escherichia coli isolates. BMC Microbiol. 20:241. doi: 10.1186/s12866-020-01933-1
Ye, Q., Wu, Q., Zhang, S., Zhang, J., Yang, G., Wang, J., et al. (2018). Characterization of extended-spectrum β-lactamase-producing Enterobacteriaceae from retail food in China. Front. Microbiol. 9:1709. doi: 10.3389/fmicb.2018.01709
Zankari, E., Hasman, H., Cosentino, S., Vestergaard, M., Rasmussen, S., Lund, O., et al. (2012). Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 67, 2640–2644. doi: 10.1093/jac/dks261
Zhang, J., Zhou, K., Zheng, B., Zhao, L., Shen, P., Ji, J., et al. (2016). High Prevalence of ESBL-Producing Klebsiella pneumoniae causing community-onset infections in China. Front. Microbiol. 7:1830. doi: 10.3389/fmicb.2016.01830
Zhu, J., Ding, B., Xu, X., Zhu, D., Yang, F., Zhang, H., et al. (2015). Klebsiella pneumoniae: development of Carbapenem resistance due to acquisition of bla NDM-1 during antimicrobial therapy in twin infants with pneumonia. Front. Microbiol. 6:1399. doi: 10.3389/fmicb.2015.01399
Zurfluh, K., Nüesch-Inderbinen, M., Morach, M., Zihler Berner, A., Hächler, H., and Stephan, R. (2015a). Extended-spectrum-β-lactamase-producing Enterobacteriaceae isolated from vegetables imported from the Dominican Republic, India, Thailand, and Vietnam. Appl. Environ. Microbiol. 81, 3115–3120. doi: 10.1128/AEM.00258-15
Keywords: antimicrobial resistance, whole genome sequencing, extended-spectrum beta-lactamases, multidrug resistance, imported food, hybrid sequencing, one health
Citation: Kurittu P, Khakipoor B, Aarnio M, Nykäsenoja S, Brouwer M, Myllyniemi A-L, Vatunen E and Heikinheimo A (2021) Plasmid-Borne and Chromosomal ESBL/AmpC Genes in Escherichia coli and Klebsiella pneumoniae in Global Food Products. Front. Microbiol. 12:592291. doi: 10.3389/fmicb.2021.592291
Received: 06 August 2020; Accepted: 14 January 2021;
Published: 03 February 2021.
Edited by:
Vincent Burrus, Université de Sherbrooke, CanadaReviewed by:
Jian-Hua Liu, South China Agricultural University, ChinaCopyright © 2021 Kurittu, Khakipoor, Aarnio, Nykäsenoja, Brouwer, Myllyniemi, Vatunen and Heikinheimo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paula Kurittu, cGF1bGEua3VyaXR0dUBoZWxzaW5raS5maQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.