 Wenjun Deng
Wenjun Deng Dana K. Dittoe
Dana K. Dittoe Hilary O. Pavilidis
Hilary O. Pavilidis William E. Chaney
William E. Chaney Yichao Yang
Yichao Yang Steven C. Ricke
Steven C. Ricke
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 22 December 2020
Sec. Food Microbiology
Volume 11 - 2020 | https://doi.org/10.3389/fmicb.2020.583429
This article is part of the Research Topic Campylobacter-associated Food Safety View all 32 articles
Poultry has been one of the major contributors of Campylobacter related human foodborne illness. Numerous interventions have been applied to limit Campylobacter colonization in poultry at the farm level, but other strategies are under investigation to achieve more efficient control. Probiotics are viable microbial cultures that can establish in the gastrointestinal tract (GIT) of the host animal and elicit health and nutrition benefits. In addition, the early establishment of probiotics in the GIT can serve as a barrier to foodborne pathogen colonization. Thus, probiotics are a potential feed additive for reducing and eliminating the colonization of Campylobacter in the GIT of poultry. Screening probiotic candidates is laborious and time-consuming, requiring several tests and validations both in vitro and in vivo. The selected probiotic candidate should possess the desired physiological characteristics and anti-Campylobacter effects. Probiotics that limit Campylobacter colonization in the GIT rely on different mechanistic strategies such as competitive exclusion, antagonism, and immunomodulation. Although numerous research efforts have been made, the application of Campylobacter limiting probiotics used in poultry remains somewhat elusive. This review summarizes current research progress on identifying and developing probiotics against Campylobacter and presenting possible directions for future research efforts.
With the introduction of selective media that could be routinely employed for isolation, Campylobacter was identified as a critical clinical pathogen associated with the gastrointestinal tract (GIT; On, 2001; Butzler, 2004). By the mid to late 1980s, Campylobacter had been recognized as one of the most common bacterial agents causing gastroenteritis worldwide (Allos, 2001; Domingues et al., 2012; Geissler et al., 2017). Currently, Campylobacter is considered one of the leading causative agents of bacterial foodborne GIT disease globally (Silva et al., 2011; Mughini-Gras et al., 2014; Kaakoush et al., 2015; Marder et al., 2018), with poultry products being one of the main vehicles of Campylobacter exposure (Skarp et al., 2016; Taylor et al., 2013).
In 2010, foodborne transmission accounted for approximately 80 and 76% of campylobacteriosis cases in the United States and in the European Union (EU), respectively (Hald et al., 2016). According to the United States Centers for Disease Control and Prevention (2018), poultry contributed to 33 cases of the 209 foodborne Campylobacter outbreaks from 2010 to 2015. Similarly, in the EU, broiler meat and products contributed 24.2% of total foodborne campylobacteriosis outbreaks in 2017 (European Food Safety Authority, 2018). Certainly, preventative measures must be taken in order to reduce the incidence of Campylobacter among poultry and poultry products.
Previously the control measures of Campylobacter in poultry included antibiotic treatment, phage therapy, competitive exclusion, and vaccination (Lin, 2009; Meunier et al., 2017; Umaraw et al., 2017). However, the use of antibiotics in livestock can cause a selection of antibiotic-resistant pathogens which further transmit to humans during food consumption, leading to more severe illnesses because of the difficulties in treatment (Gupta et al., 2004; Iovine and Blaser, 2004; Yang et al., 2019). Due to the threat to public health, the use of antibiotics in poultry production has become more restricted (Tang et al., 2017; CDC, 2019). Consequently, in recent years alternative strategies and feed additives to effectively control the colonization of Campylobacter in poultry GIT have become of increasing interest (Park et al., 2016; Hossain et al., 2017). This review provides a historical perspective and recent updates on the development of anti-Campylobacter probiotics, the effect of host-microbiota on probiotics, and possible directions for future probiotic development research efforts (Figure 1).
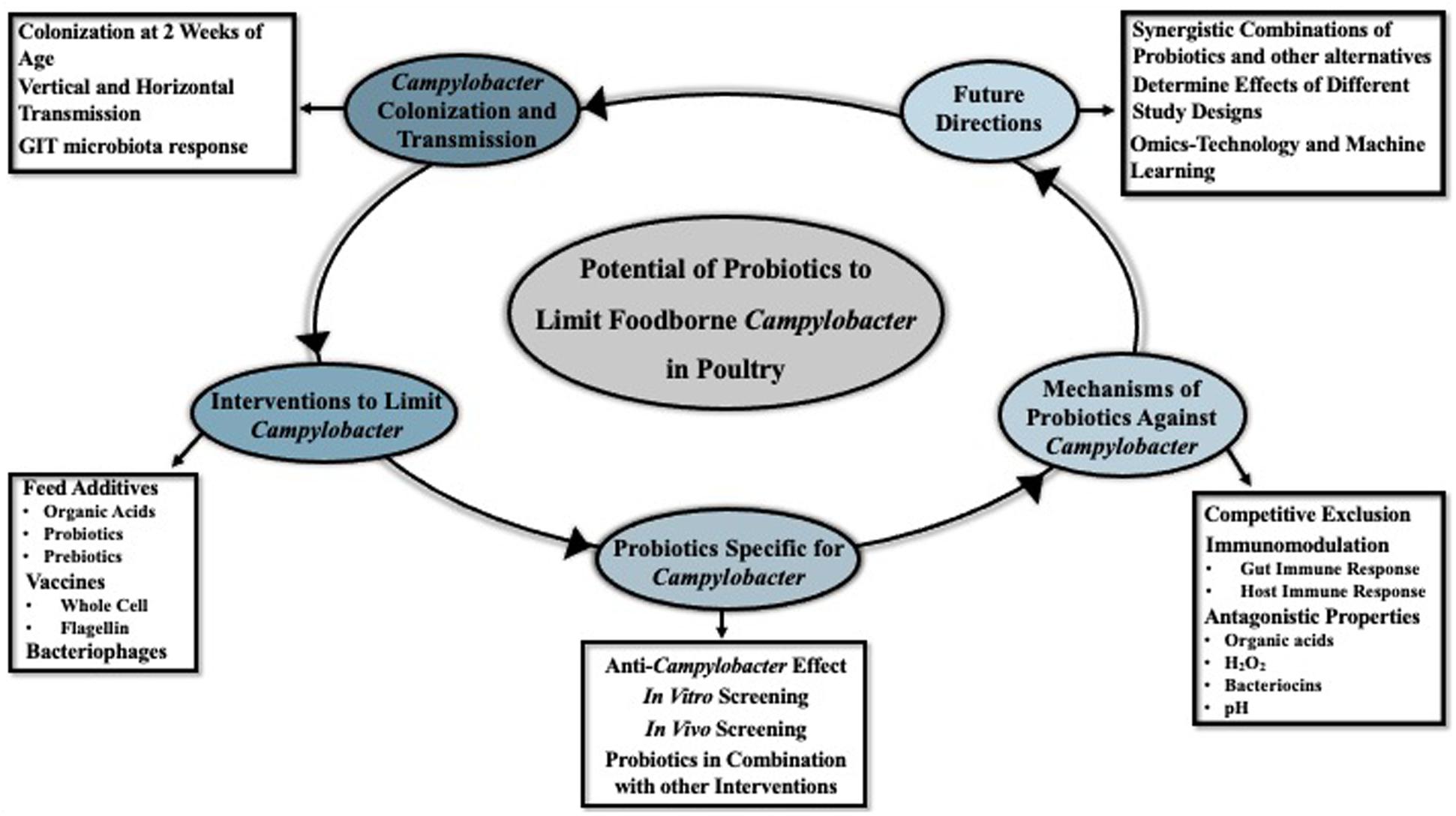
Figure 1. Brief overview of the topics and future directions that are discussed in the current review.
The two most common species, Campylobacter jejuni, and C. coli are thermophilic and obligate microaerophilic bacteria that colonize the intestinal mucosa of most warm-blood animals, including humans (Newell and Fearnley, 2003). Campylobacter spp. prefer to colonize avian species such as wild birds, broilers, turkeys, and ducks (Newell and Fearnley, 2003). The microaerophilic environment and internal body temperature of 41°C in the avian GIT provide optimal environmental conditions for Campylobacter (Robyn et al., 2015). The colonization of Campylobacter in chickens primarily occurs in the ceca and small intestine. It can still become invasive, appearing in the liver, spleen, deep muscle, thymus, bursa of Fabricius, and blood (Awad et al., 2018). It was reported that once ingested, C. jejuni colonization begins in the ileum, followed by dissemination to the jejunum and cecum (Lacharme-Lora et al., 2017). Campylobacter rapidly establishes in the chicken ceca and multiplies, eventually reaching very high cecal concentrations (109 CFU/g cecal content; Newell and Fearnley, 2003). Beery et al. (1988) determined the concentration of C. jejuni in different GIT locations of 8-day-old chickens, which were orally inoculated with 5 × 108 CFU bacteria cells. The concentrations in the proximal and distal small intestines and the large intestines reached 105 CFU/g and became undetectable at 5 days post-inoculation (minimum level of detection 102 CFU/g). The majority of the colonization occurred in the cecum, which attained a peak of 107 CFU/g 1 day after inoculation and remained detectable even at 7 days. Smith and Berrang (2006) compared the prevalence and concentration of food pathogens between crop and gizzard content in broiler carcasses. The Campylobacter prevalence in the crop contents (29 of 29 chickens) was higher than in gizzard contents (12 of 30 chickens). Moreover, it was found that the crops (4.6 log10 CFU/mL) contained significantly higher concentrations of Campylobacter than the gizzard (2.2 log10 CFU/mL).
The colonization of Campylobacter in chickens varies due to host age, bacterial strain type, and infective dose (Sahin et al., 2003). Newly hatched chickens are generally free of Campylobacter until 1–2 weeks of age; this delay of colonization is referred to as the lag phase (van Gerwe et al., 2009; Pielsticker et al., 2012; Kalupahana et al., 2013). The primary explanation for the lag phase is the protective effect of maternal antibodies (MAB), but the mechanisms behind it have not clearly been described yet (Sahin et al., 2003; Ringoir et al., 2007). The MAB levels are highest in newly hatched chickens but decrease gradually to the background level at approximately 3 weeks of age (Sahin et al., 2003; Shoaf-Sweeney et al., 2008). One study compared the colonization of C. jejuni S3B in 3-day-old chickens with and without anti-Campylobacter MAB, which were hatched from bacteria-infected and uninfected hens, respectively. When challenged with C. jejuni at the concentration of 5 × 105 CFU/bird, the MAB+ chickens exhibited a significantly lower percentage of shedding than MAB- birds at 2 and 4 post-inoculation days (PIDs). However, at 12 PIDs, both groups reached a 100% shedding rate (Sahin et al., 2003). These results indicated the partial protective effect of Campylobacter MAB against colonization in young chickens.
Furthermore, Cawthraw and Newell (2010) reported that there was no simple linear relationship between the level of MAB in chicken and their resistance to a C. jejuni challenge. It was noted that the resistance of 8-day-old chickens was greater than that for the day-of-hatch birds, although the maternally derived anti-C. jejuni IgY serum antibodies were at the peak levels (approximate ELISA OD450 2.1) in the latter one. From 8 to 21 days, the C. jejuni resistance of chickens and the antibody levels both decreased. Overall, the 1 to 2-week old birds were more resistant to C. jejuni than the 3 week old birds (Cawthraw and Newell, 2010). Another study reported that at least 5 × 104 and 5 × 103 of cells of a C. jejuni laboratory-maintained strain were required to colonize 2 and 14-day old chickens, respectively (Ringoir et al., 2007). Ringoir et al. (2007) demonstrated that 2-day old chickens were less susceptible to C. jejuni than 14-day chickens. Sahin et al. (2003) reported no interference between high levels of maternal antibodies and the development of the humoral immune system in young chickens. Furthermore, the 21-day-old chickens showed a much higher and more rapid humoral response than the 3-day-old chickens.
Previously Campylobacter was considered a non-pathogenic commensal in the poultry intestine, but this concept has been questioned more recently (Humphrey et al., 2014; Awad et al., 2018; Connerton et al., 2018). Humphrey et al. (2014) exposed four commercial broiler chicken breeds to C. jejuni M1, and initially, all four breeds produced inflammatory signals in the innate immune response to bacterial colonization. At 12 DPI, three breeds exhibited reduced inflammation responses. They remained asymptomatic after the expression of interleukin-10 (IL-10), while the other faster-growing breed failed to produce IL-10, which led to prolonged inflammation and diarrhea. In addition, Awad et al. (2015) reported changes in intestinal permeability and histomorphology, including decreased crypt depth, villi height, and surface area in Campylobacter colonized chickens.
Campylobacter can rapidly transmit from a colonized individual chicken to the entire flock in a matter of days (Awad et al., 2018). A study conducted with an Australian broiler flock quantified the transmission rate for each C. jejuni-infected bird, resulting in estimates of 2.37 ± 0.295 new bird infections per day. Based on this transmission rate, a flock consisting of 20,000 broilers exhibits a 95% prevalence of Campylobacter within 4.4 to 7.2 days after the appearance of the first infected bird (van Gerwe et al., 2009). The prevalence of Campylobacter in broiler chickens varies depending on the location and can range anywhere between 3 and 90% (Marotta et al., 2015). The transmission routes at the farm are considered to originate mainly from the surrounding environment and are transmitted horizontally between flock mates (Cox et al., 2012; Sibanda et al., 2018).
The vertical transmission (i.e., hen to the egg then to the chick) of Campylobacter remains debatable since the observations of vertical transmission have not been consistent across different studies (Cox et al., 2002; Callicott et al., 2006). In fact, Battersby et al. (2016) investigated the transmission of Campylobacter on three broiler farms (two flocks per farm) where they took fecal and environmental swabs and used polymerase chain reaction (PCR) assays to detect Campylobacter. Battersby et al. (2016) concluded that vertical transmission did not occur but rather the surrounding environment was one of the primary sources of Campylobacter and that biosecurity should be considered as a control measure. They concluded that once Campylobacter is spread among the flock, it is not long after that Campylobacter is detected outside the broiler house. In contrast, Rossi et al. (2012) determined that Campylobacter transmission can be due to vertical transmission from the breeder hen. Rossi et al. (2012) inoculated breeder hens intraesophageally and specific pathogenic free (SPF) eggs with Campylobacter coli. Rossi et al. (2012) demonstrated the transmission from inoculated hen to egg (offspring) and the potential dangers of C. coli reaching the amnion of SPF eggs. However, Rossi et al. (2012) contended that C. coli did not appear viable in the infected eggs produced by the inoculated hen, a potential limitation of C. coli transmission in the field.
A recent survey on United States broiler production revealed that none of the investigated farm managers (n = 18) reported Campylobacter tests in their farms, while 11% of poultry veterinarians (n = 2) and 90% of processing plants (n = 18) included Campylobacter in their microbiological tests (Hwang and Singer, 2020). Moreover, only 33% (n = 6) of the farms implemented the validated measurements for limiting Campylobacter contamination. In the survey, biosecurity was ranked as the most effective strategy against Campylobacter on the farm. Still, over half of the farmers and veterinarians reported that biosecurity is not adequate at limiting Campylobacter transmission and subsequent infection. These responses collectively indicated that while biosecurity might be the best available choice, it alone may not be sufficiently effective. Thus, further interventions must be utilized to mitigate Campylobacter prevalence among poultry, such as dietary supplements that alter Campylobacter colonization within the GIT.
The microbiota of the poultry GIT plays several important roles that benefit host health through the competitive exclusion of pathogens and other non-indigenous microorganisms, stimulation and development of the host immune system, and absorption of nutrients (Shang et al., 2018). It has been determined that most of the bacteria in ileum and ceca of broiler chickens are Gram-positive with low G + C content, which mainly includes Lactobacillus, clostridia, Bacillus, and Streptococci (Lu et al., 2003).
However, the bacterial taxonomic composition of the ceca is significantly different from the jejunum and ileum, which may be attributable to the different functions of these two GIT regions (Gong et al., 2002; Stanley et al., 2014; Awad et al., 2016; Feye et al., 2020). The primary role of the jejunum and ileum is nutrient absorption. However, the ceca serve as the primary site for bacterial fermentation, further nutrient absorption, detoxification, and prevention of pathogen colonization (Gong et al., 2007). In addition, Oakley and Kogut (2016) determined that the fecal and cecal microbiota compositions also differ from one another. At 1-week of age, Gallibacterium and Lactobacillus were dominant in the feces, while Bacteroides was more abundant in the ceca. Both Clostridium and Caloramator increased significantly in the ceca, whereas Lactobacillus remained dominant in the feces of broilers at 6 weeks of age.
In addition to the location of GIT, the composition of microbiota varies depending on the age of broiler chickens. Awad et al. (2016) identified 24 phyla from the GIT contents of 1 to 28-day old broiler chickens. It was revealed that the Firmicutes and Proteobacteria were the most abundant phyla in all birds. In particular, the number of Proteobacteria were found to be significantly higher in newly hatched chickens and subsequently decreased with age, while the Firmicutes were predominant in older birds. This microbial community transition appeared to be related to oxygen availability. The facultative anaerobes of the Proteobacteria phylum initially colonized the GIT, but as oxygen gradually became depleted, the obligate anaerobes from Firmicutes emerged as the dominant microorganisms (Wise and Siragusa, 2007). Oakley et al. (2014) noted that age was more of a driving factor in the diversity and population of the cecal microbiota than dietary treatments. When the birds were 7-day-old, the cecal microbiota were primarily comprised of Flavonifractor, Pseudoflavonifractor, and Lachnospiracea. However, by 21-day of age, Faecalibacterium dominated the microbiota of broilers regardless of dietary treatment (Oakley et al., 2014).
Older birds (14–28 days old) exhibited significantly more microbial richness and diversity than young birds (1–7 days old) based on several diversity indices, which included the number of operational taxonomic units (OTUs), Chao1, abundance-based coverage estimator (ACE), Shannon’s index and Simpson index (p < 0.01; Awad et al., 2016). Oakley et al. (2014) and Oakley and Kogut (2016) also noted the increase in microbiota richness and diversity as birds matured.
Although some recent research has indicated that Campylobacter colonization does not influence the subsequent microbiota composition of commercial broiler flocks (Oakley et al., 2018), significant efforts have been made to determine the influence that Campylobacter has on the GIT microbiota. Discussion on the potential interactions between the microbiota and Campylobacter in the host GIT has occurred in more recent years (Indikova et al., 2015). Oakley et al. (2013) demonstrated that all samples along the “farm to fork” continuum had a common core microbiota consisting of recognized pathogens such as Clostridium, Campylobacter, and Shigella. Also, Oakley et al. (2013) revealed that Campylobacter did not appear to have a significant association with other taxa, concluding that this may be a reason as to why competitive exclusion is not effective against Campylobacter (2013). However, C. jejuni did have a significant association with Megamonas hypermegale (Oakley et al., 2013). Other significant research has revealed that the C. jejuni concentrations are higher in the cecal contents of antibiotic-treated or germ-free raised chickens than birds with a conventional microbiota, indicating a host protective role of microbiota against C. jejuni (Han et al., 2017).
Sakaridis et al. (2018) randomly sampled 100 birds from four different commercial farms to determine the association of Campylobacter loads and the subsequent microbiota composition using 16S rDNA. They determined that the inter-farm variation had a more pronounced effect on the microbiota than the intra-farm variation (Sakaridis et al., 2018). There was no correlation between Campylobacter load and the levels of Firmicutes, Bacteroidetes, or Ternicutes; however, there were increased levels of Enterobacteriaceae and decreased levels of Lactobacillus in the cecal microbiota of birds that had high levels of Campylobacter (Sakaridis et al., 2018). Therefore, Sakaridis et al. (2018) suggests that Lactobacillus and Enterobacteriaceae may be populations to modulate in order to decrease Campylobacter colonization.
In addition, Connerton et al. (2018) demonstrated that Campylobacter colonized chickens possess a different cecal microbiota composition than those not infected at 2 days post-inoculation. However, as time progressed, age was a more significant driver of the microbiota composition. Upon inoculation, Campylobacter colonization of the ceca was followed by a reduction of Lactobacillaceae and Clostridium cluster XIVa (Connerton et al., 2018). In contrast, Sofka et al. (2015), who compared the microbiota composition between C. jejuni positive and negative chickens in the fecal samples, saw no differences in the fecal microbiota composition. However, Sofka et al. (2015) noted that higher proportions of common GIT bacteria such as Firmicutes (62 versus 36.6%), Proteobacteria (44.6 versus 21.3%), and Bacteroidetes (15.4 versus 6.5%) were found among the non-inoculated birds (Campylobacter free).
Understanding the colonization of Campylobacter in the GIT of poultry and how the residential microbiota may impact or enhance Campylobacter infection in poultry enables stakeholders and the industry to combat this rather arduous foodborne pathogen. With this knowledge, researchers have investigated the use of multiple interventions, whether at the preharvest or postharvest level.
Interventions against Campylobacter can be employed at each step of the poultry production chain (i.e., farm, transport, slaughter, processing, and retail; Sahin et al., 2015; Umaraw et al., 2017; Upadhyay et al., 2019). The choice of control measures at each primary poultry production step plays a vital role against Campylobacter and could affect the following steps along the food chain (Meunier et al., 2016). Interventions applied at the farm level aim to reduce or eliminate the colonization of Campylobacter in broilers, while the remaining steps at postharvest primarily focus on decontamination during processing at the plant (FSAI Ireland, 2002). Due to the focus of the current review, only interventions at the farm level will be discussed. The interventions used at the poultry farm can be broadly categorized as biosecurity interventions, feed additives, vaccines, and bacteriophage (Hermans et al., 2011).
There is a wide range of feed additives, including organic acids such as SCFA, medium-chain fatty acids (MCFA) and their monoglycerides, plant-derived compounds, probiotics, prebiotics, and bacteriocins that are available as potential interventions (Guyard-Nicodème et al., 2016; Meunier et al., 2016; Dittoe et al., 2018; Kim et al., 2019; Micciche et al., 2019). The reported organic and fatty acids include but are not limited to caprylic acid, butyrate, formic acid, and sorbate (Dittoe et al., 2018). The SCFA exerts antibacterial effects by diffusing across the bacterial membrane as non-ionized acids, and dissociating in bacteria cells, which further leads to the decrease of intracellular pH and the dissipation of the proton motive force, and impacting cellular physiology (Ricke, 2003; Sun and O’Riordan, 2013). As reviewed by Meunier et al. (2016), the effectiveness of organic and fatty acids against Campylobacter varied widely among studies, and the reproducibility across experiments was low. A study of Guyard-Nicodème et al. (2016) compared the anti-Campylobacter effect of 12 feed additives (i.e., monoglyceride, SCFA, plant extracts, probiotics, and a prebiotic-like compound) in broiler chickens over the 42-day sampling period. At 14 days of age, eight additives elicited a significant reduction of C. jejuni, while at 35 days, only three additives remained effective, namely, a monoglyceride, a SCFA, and a multi-species probiotic. At 42 days of age, the SCFA still led to more than 2 log10 CFU/g reduction of C. jejuni, as well as a probiotic and a prebiotic-like compound, which only showed effectiveness at 42 days.
As a result, out of the array of feed amendments available to poultry producers to mitigate Campylobacter among their flocks, probiotics are promising. Probiotics, as will be discussed, are capable of directly competing for nutrients against pathogens, excluding pathogens from binding sites, expressing antagonistic mechanisms, and stimulating immunomodulation. A more detailed discussion on probiotics and bacteriocin formation is included in a later section of the current review.
Probiotics are considered non-pathogenic and non-toxic viable microorganisms that incur favorable impacts on host health when administrated via an oral route (Lutful Kabir, 2009). Probiotics can be bacteria or yeast and consist of either individual strains or a mixture of several organisms. The more common probiotics are generally comprised of species and strains of Lactobacillus, Streptococcus, Bacillus, Escherichia coli, Bifidobacterium, and Saccharomyces, among others (Dobson et al., 2012; Kergourlay et al., 2012; Helmy et al., 2017; Thibodeau et al., 2017; Massacci et al., 2019). The beneficial effects of probiotics in poultry production include maintaining an optimal balance of GIT microbiota, inhibition of pathogens (Bhatia et al., 1989; Santini et al., 2010; Ritzi et al., 2014), immunomodulation (Cox and Dalloul, 2015), positive histomorphological changes of the ileum (Olnood et al., 2015; Forte et al., 2018), and improving broiler growth performance among others (Kabir et al., 2004). The screening and selection process are critical to ensure that probiotic strains survive the GIT and play their beneficial roles in the animal host. The probiotic strain(s) should be able to survive under GIT conditions (e.g., low pH and bile salt), feed processing and storage conditions (e.g., heat, dry, and starvation), retain high viability and exhibit beneficial effects once reaching the target region of the GIT (Santini et al., 2010; Song et al., 2014; Park et al., 2016).
When used as a feed additive, probiotics help to mediate poultry GIT health and reduce the colonization of food and poultry pathogens in the chicken GIT (Cean et al., 2015; Eeckhaut et al., 2016). The addition of a butyrate-producing probiotic strain of Butyricicoccus pullicaecorum into a commercial chicken diet significantly reduced the percentage of necrotic lesions caused by Clostridium perfringens in all tested trials. It also reduced Campylobacter populations in cecal contents by 1.5 log10 gene copies/g at day 40 (Eeckhaut et al., 2016). More recently, several studies have screened specific probiotic strains targeted against Campylobacter in chickens (Kobierecka et al., 2016a, b; Wang et al., 2019). However, there were variations in reported results among different studies due to the experimental conditions, detection methods, intra- and inter-flock differences of the chicken host, and the complexity of probiotics and host interactions. In turn, these complex interactions led to difficulties in validating the subsequent effects of administering specific probiotic strains (Saint-Cyr et al., 2016; Johnson et al., 2017).
Taha-Abdelaziz et al. (2019) reported the inhibitory effects of six Lactobacilli spp. against C. jejuni in vitro. Both the neutralized cell-free supernatant and the Lactobacilli spp. cell culture inhibited C. jejuni growth with clear inhibition zones on Mueller-Hinton (MH) agar. Further investigation found that C. jejuni exposed to all Lactobacilli spp. except Lactobacillus reuteri exhibited a downregulation of genes responsible for motility and invasion, as well as reduced quorum sensing molecule AI-2 production. Another study by Mortada et al. (2020) tested a commercial probiotic for the anti-Campylobacter effect in vitro and in vivo. The overnight cultured gentamicin-resistant C. coli was co-cultured at different ratios (1:0, 1:1, 1:5 or 1:10) with the cell-free supernatant of four probiotic strains namely Lactobacillus reuteri, Pediococcus acidilactici, Bifidobacterium animalis, and Enterococcus faecium from the commercial product. It was found that the four strains were able to reduce C. coli in vitro at a 1:1 ratio or higher. However, when the probiotic product was added to the feed of Cobb-500 broilers, the probiotic supplementation was unable to reduce the colonization of the C. coli group at 42 days of age.
The commonly used probiotic strains belong to lactic acid bacteria (LAB), bifidobacteria, Bacillus spp., Bacteroides spp., and Streptococcus spp. (Applegate et al., 2010). Based on the regulation of the United States Food and Drug Administration (US Food and Drug Administration, 2018), the probiotic candidate strains should be generally regarded as safe (GRAS). The probiotic candidate strains usually originate from poultry feces and GIT contents, human feces, cheese, and plant silages (Santini et al., 2010; Ghareeb et al., 2012; Nishiyama et al., 2014; Kobierecka et al., 2017; Smialek et al., 2018). The selection of one or several optimally performing probiotic strains from a broader set of probiotic bacterial candidates is an intensive process. Therefore, tests need to be simple, rapid, and comprehensive (Taheri et al., 2009). The screening process can be roughly divided into two steps: pre-selection in vitro and evaluation in vivo (Lutful Kabir, 2009). Many screening studies are only carried out in vitro, while others are conducted as both in vitro and in vivo studies. The latter of the screening methods is more informative than the former in determining a probiotic’s candidacy.
In vitro screening methods can vary among studies. However, they generally include aggregation and co-aggregation, antibacterial activity, enzymatic activity, cell surface hydrophobicity, survival (acid and bile salt), strain identification and antibiotic sensitivity tests (Tables 1, 2; Taheri et al., 2009; Blajman et al., 2015; García-Hernández et al., 2016). These tests help determine if the probiotic strains would survive when exposed to the extreme conditions of the host GIT environment while still exerting their beneficial functions. The antibacterial test is essential for accurately screening anti-Campylobacter probiotics. The cell-free supernatants of probiotics are usually tested using a well-diffusion assay on agar plates to observe the range of anti-Campylobacter clearing zones in a Campylobacter lawn background. Live cultures of probiotic strains can be tested by methods such as agar spot, agar slab, and as co-cultures in suspension or with GIT cell monolayers (Robyn et al., 2012; Saint-Cyr et al., 2016; Kobierecka et al., 2017; Dec et al., 2018). The in vitro co-culture with intestinal cells mimics the interactions between probiotics and pathogens in the host GIT. Šikiæ Pogaèar et al. (2020) tested the Lactobacillus spp. strains (Lactobacillus plantarum PCS20, PCS22, PCS25, PCK9, and Lactobacillus rhamnosus LGG) for their competitive adhesion and infection prevention ability against C. jejuni in chicken B1OXI and pig PSI cl.1 epithelial polarized cells. Specifically, in the PSI cl.1 cell line, all tested bacterial strains significantly reduced the adhesion and infection of C. jejuni at 3, 17, and 24 h post-infection. In the B1OXI cell line, the bacterial strains PCS22, LGG, and PCK9 significantly reduced the adhesion of C. jejuni at 24 h. However, the invasion of C. jejuni in the B1OXI cell line was only observed at 3 h post-infection. At that time, the addition of PCS20, PCS22, and PCK9 significantly reduced the invasion of C. jejuni, whereas bacterial strains PCS25 and LGG prevented C. jejuni invasion.
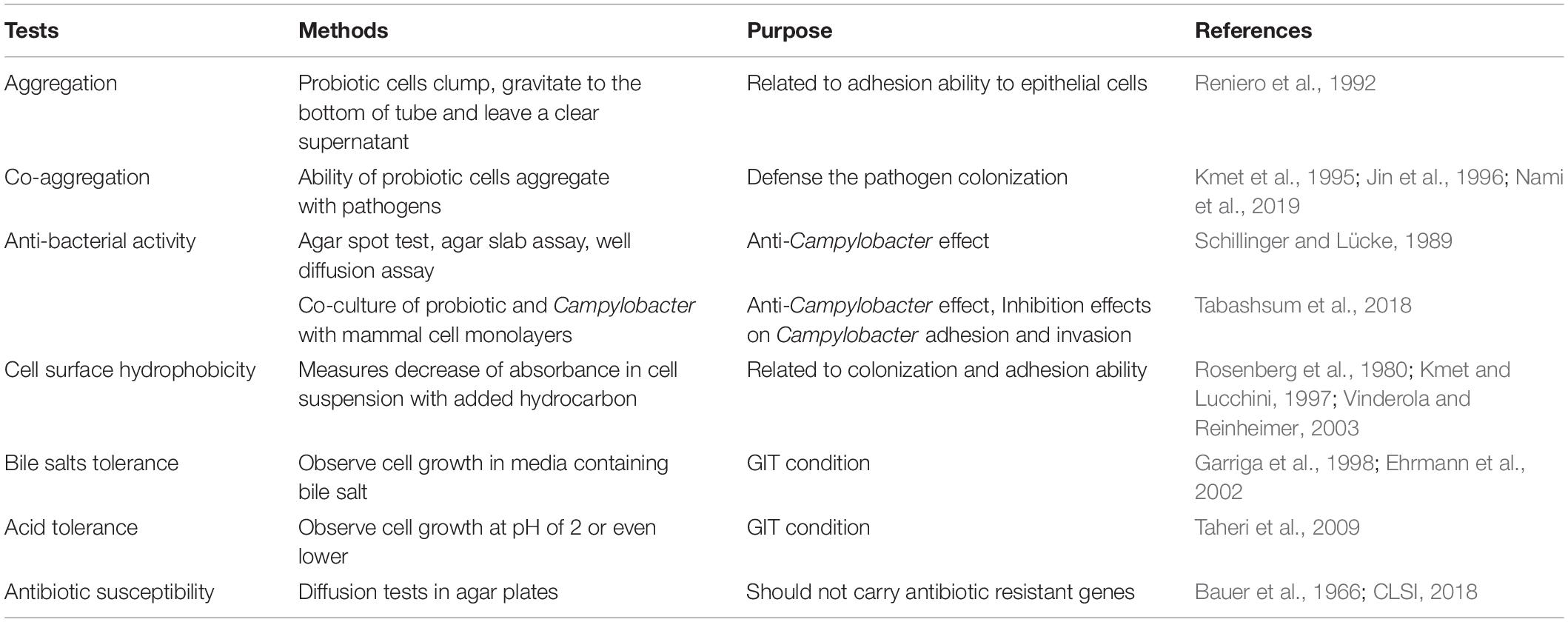
Table 1. Summary of tests in screening probiotics in vitro.
The aggregation and adhesion ability of probiotic bacterial strains facilitates their establishment in the GIT and the exclusion of pathogens (Lebeer et al., 2008). Tareb et al. (2013) investigated the auto-aggregation and co-aggregation ability of viable and heat-inactivated cultures of L. rhamnosus CNCM-I-3698 and L. farciminis CNCM-I-3699. Both living and dead cells of the two strains showed strong co-aggregation ability with C. jejuni CIP 70.2T, which was through the carbohydrate-lectin interaction and proteinaceous components. When probiotic strains were added to mucin at the same time or after C. jejuni, the inactivated probiotic cells were more effective than the living cells at preventing C. jejuni colonization. The authors indicated that the enhanced adhesion might be because of the production of EPS during the heat inactivation. One advantage of heat-inactivated cells over living cells is enhanced storage stability (Ostad et al., 2009; Ishikawa et al., 2010).
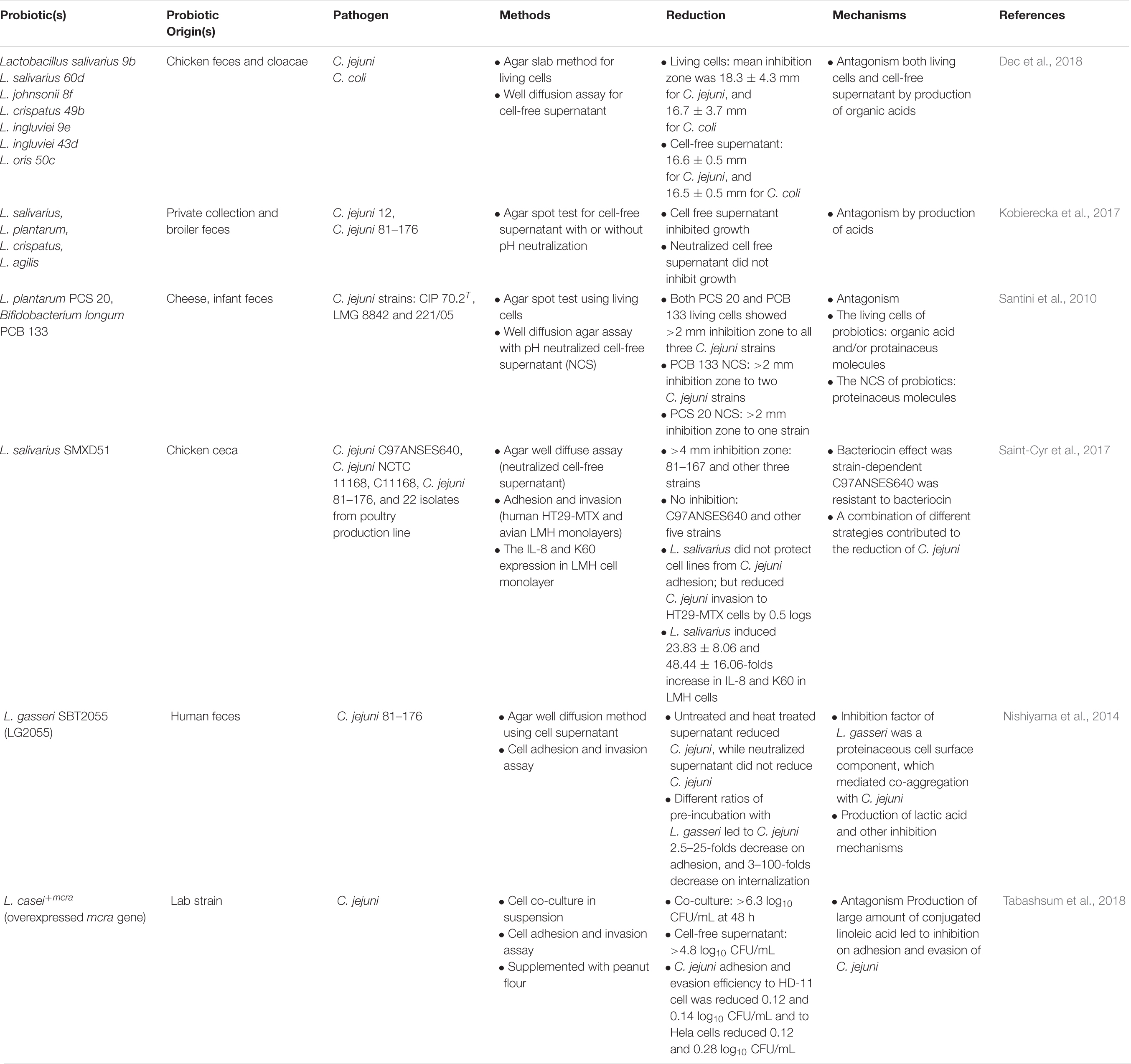
Table 2. Summary of in vitro screenings and methodologies of various probiotic strains against Campylobacter in Poultry.
The primary purpose of an antibiotic sensitivity test is to prevent the transposition of antibiotic-resistant genes to nearby resident GIT microbiota (Danielsen and Wind, 2003). It was believed that the probiotics should adhere to the poultry GIT mucosa and maintain viability under harsh conditions, which meant that probiotic strains could potentially have direct contact with intestinal microbiota and transfer antibiotic-resistance genes through horizontal transfer (Luangtongkum et al., 2009; Lutful Kabir, 2009; Verraes et al., 2013). If antibiotic-resistant genes are transferred to pathogens such as Campylobacter, it could represent a human health hazard (Imperial and Ibana, 2016). Lactobacillus spp. have been identified as candidates for anti-Campylobacter probiotics in several studies, but many strains of Lactobacillus spp. are resistant to certain antibiotics. Ocaña et al. (2006) reported that six Lactobacillus species were able to grow under high concentrations of streptomycin, kanamycin, quinolones (norfloxacin and ciprofloxacin), chloramphenicol, cephalosporins (ceftriaxone and ceftazidime), and aztreonam. Similarly, Taheri et al. (2009) reported a probiotic candidate for poultry, Lactobacillus crispatus, exhibited resistance to nalidixic acid and neomycin. Overall, the in vitro selection process helps exclude unqualified candidate strains and narrows the range of required screening. Ultimately, only a few strains that perform outstandingly well in the in vitro tests should be selected for in vivo tests.
Although the in vitro tests characterized probiotic strains under conditions mimic the intestinal environment, they cannot reproduce the exact interactions that occur among probiotics, the host GIT microbiota, and the possible GIT immune response (Table 3; Saint-Cyr et al., 2016, 2017; Mortada et al., 2020). The selected candidate strains that were inhibitory in vitro may not elicit a reduction of Campylobacter in vivo (Robyn et al., 2012, 2013). Thus, the selected probiotic strains should be further evaluated in vivo to determine their colonization ability, anti-pathogen effects, and persistence in chicken GIT (Lutful Kabir, 2009). Blajman et al. (2015) conducted a study to select chicken-originated probiotic strains for feed supplementation. In that study, 360 bacterial strains from broiler chicken GIT contents were screened through a series of in vitro tests including aggregation test, antagonistic activity, bacterial identification, cell surface hydrophobicity, acid resistance, bile tolerance, and H2O2 production tests, and the three best performing probiotic strains were selected for follow up in vivo testing. These three strains were constructed to be rifampicin-resistant to track their colonization within the chicken GIT. The administered strain concentrations in the liver, crop, and cecum were determined by direct plating on de Man, Rogasa, and Sharpe agar plates supplemented with rifampicin (MRSrif). In the end, Lactobacillus salivarius DSPV001P was selected as the candidate probiotic strain since it successfully colonized and maintained significantly higher population levels in the broiler GIT.
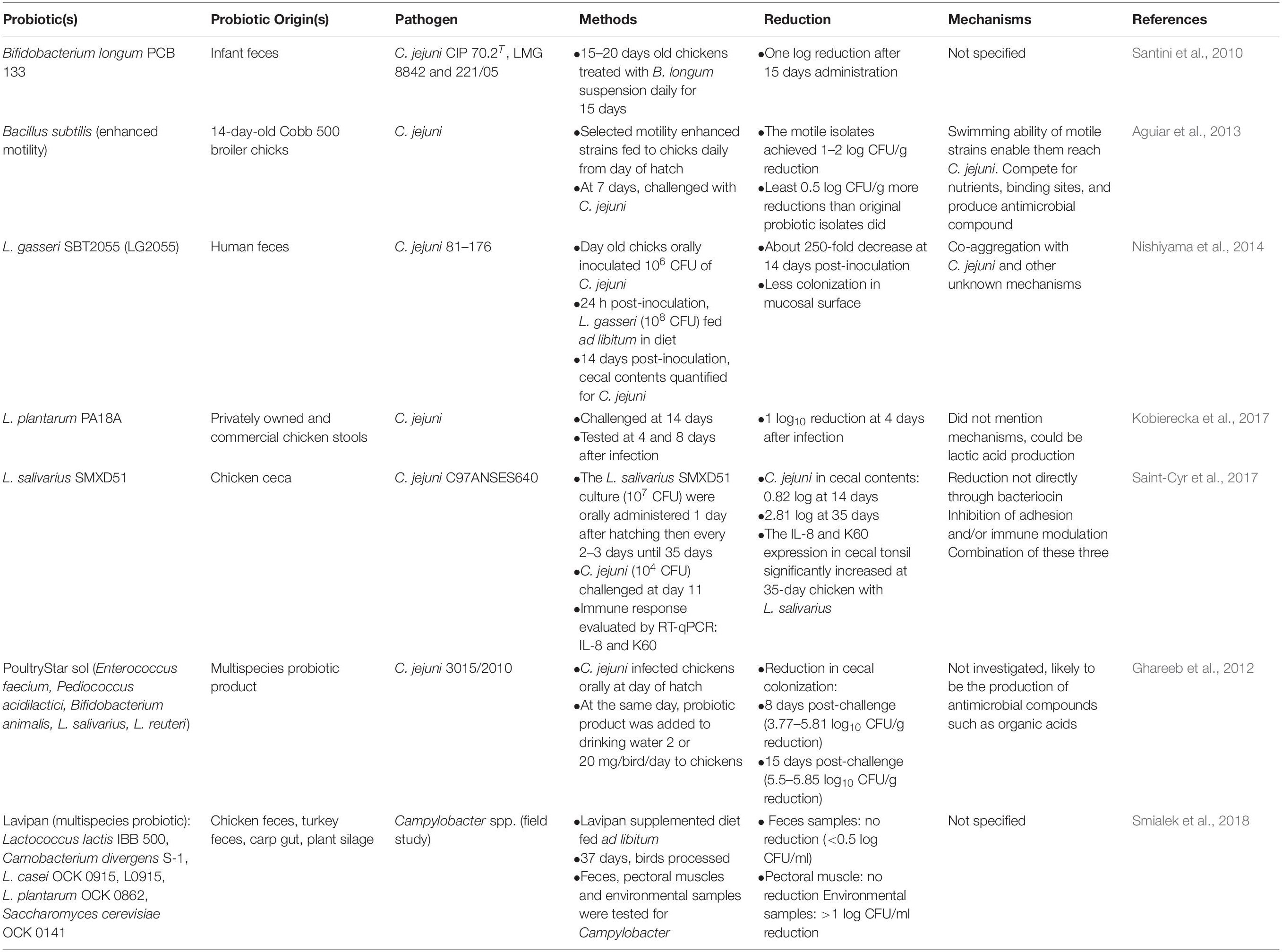
Table 3. Summary of the in vivo screening of probiotic strains against Campylobacter in poultry.
One way to ensure the colonization of the lower GIT is through encapsulation or microencapsulation of the probiotic. In fact, some probiotic strains with anti-Campylobacter properties cannot survive the acidity of the host stomach, so encapsulation is necessary. Arsi et al. (2015a) used intracloacal inoculation of probiotics to introduce them more directly to the lower GIT of birds for in vivo screening without the extra cost for encapsulation. They compared intracloacal administration to oral gavage on the anti-Campylobacter effects of ten pre-selected probiotic strains. The birds were challenged with C. jejuni on day 7, and on day 14, C. jejuni cecal concentrations were quantified. Only one strain of oral administered probiotics achieved a 1-log reduction in the ceca, whereas the six strains introduced via intracloacal administration resulted in 1–3 log reductions of C. jejuni.
During the in vivo screening procedures, additional characteristics enable the probiotic strains to achieve better anti-Campylobacter performance in the poultry host. Motility enhancement of probiotic strains was identified as a critical characteristic for the reduction of Campylobacter colonization in the GIT. Aguiar et al. (2013) developed a screening technique for selecting probiotic strains with enhanced motility. The in vivo experiments indicated that motile strains reduced at least 0.5 log10 CFU/g more C. jejuni in the cecum than the original strains. The enhanced ability allowed the motile strains to reach the deep mucosal surface of cecal crypts, and overcome C. jejuni by occupying the binding sites, competing for nutrients, and/or by the production of antibacterial compounds.
In order to meet the needs of industrial production, the viability and persistence of probiotics under storage conditions such as lyophilization need to be evaluated as a practical consideration following the in vivo studies (Blajman et al., 2015). The in-feed stability and viability of probiotics will ensure that a sufficient level is administered to the host to deliver the expected anti-Campylobacter effects under production conditions (Ren et al., 2019).
Probiotics can also be applied with other Campylobacter control measures to improve the effectiveness of the overall intervention. Such combinations that exist are probiotics coupled with vaccines, phytochemicals, and prebiotics. Nothaft et al. (2017) reported that the co-administration of probiotic strains Anaerosporobacter mobilis or L. reuteri increased the efficacy of a C. jejuni vaccine in both broiler and Leghorn layer chickens. This research group developed an N-glycan-expressing Escherichia coli live vaccine that induced a specific immune response and achieved a several log reduction of C. jejuni in the host. However, the vaccine was not protective in some birds where lower numbers of A. mobilis were present in the GIT. When co-administered with either A. mobilis or L. reuteri, a higher proportion of chickens were protected against C. jejuni by the vaccine, accompanied by increased body weight and production of antibodies against the C. jejuni N-glycan antigen.
In an in vitro co-culture conducted by Tabashsum et al. (2019), berry pomace phenolic extracts (BPPE) stimulated the growth and enhanced the anti-Campylobacter effects of a conjugated linoleic acid overproducing Lactobacillus casei (LC-CLA). The co-culture of the cell-free supernatant from LC-CLA with the presence of BPPE reduced C. jejuni over 3.2 log10 CFU/ml while BPPE alone or LC-CLA+ BPPE only decreased C. jejuni by approximately 1.8 log10 CFU/ml (Tabashsum et al., 2019). Also, in the presence of BPPE, the LC-CLA living cell and cell-free supernatant both exhibited a much stronger inhibition against the C. jejuni adhesion and invasion of the DF-1, HD-11, and HeLa cell monolayers. For instance, LC-CLA + BPPE reduced both adhesion and invasion of C. jejuni to HD-11 by 1 log10 CFU/ml. However, in the same research group’s previous study, the reduction by LC-CLA alone reduced adhesion and invasion to HD-11 by approximately only 0.07 and 0.14 log10 CFU/ml (Tabashsum et al., 2018, 2019).
Fooks and Gibson (2002) investigated the anti-pathogen effect of probiotic strains L. plantarum 0407 and Bifidobacterium. bifidum Bb12 that utilized various prebiotics as carbohydrate sources. The probiotics were co-cultured with pathogens in a basal media that was supplemented with different prebiotics [fructooligosaccharide (FOS), inulin, and xylooligosaccharides (XOS)], and their paired mixtures. Regardless of experimental conditions, the probiotic and prebiotic combinations of L. plantarum + FOS, B. bifidum + FOS, B. bifidum + Inulin: FOS (80:20 w/w), and B. bifidum + FOS: XOS (50:50 w/w) significantly reduced C. jejuni growth in the basal medium compared to the probiotic strains-only groups. The anti-Campylobacter effects of probiotic and prebiotic combinations varied depending on the type of prebiotics used. This variation might be because fermentation was affected by the different prebiotic structures, which impacted pathogen inhibition and the end products produced by the probiotics. Arsi et al. (2015b) also attempted to improve the anti-Campylobacter efficacy of three probiotic strains (Bacillus spp., L. salivarius subsp. salivarius and subsp. salicinius) by the supplementation with prebiotics in broiler chickens. Two prebiotics, FOS (0.125, 0.25, or 0.5% concentration) and mannan oligosaccharide (MOS, 0.04%, 0.08 or 0.16% concentration) were combined with each probiotic strain and fed to the day-of-hatch chickens. The chickens were challenged with Campylobacter on day 7, and cecal concentrations were quantified on day 14. The combination of 0.04% MOS and L. salivarius subsp. salicinius led to a 3-log reduction of Campylobacter, whereas the probiotic alone only resulted in a 1 to 2 log reduction. In addition, Baffoni et al. (2017) found that the life-long administration of a synbiotic (probiotic Bifidobacterium longum subsp. longum PCB133 and prebiotic XOS) effectively protected the chicken host against Campylobacter more than a short-term supplementation (starting at 14-day old). Compared to 10-day-old chickens, the plate counts of Campylobacter in 39-day-old chickens were reduced by approximately 4.8 and 3.8 log CFU/g with the prolonged and discontinued supplementation of the synbiotic treatment. However, qPCR-based quantification showed no significant reduction of Campylobacter between 10 and 39-day old chickens in both treatment groups. Thus, the choice of detection method can contribute to a different conclusion in a study.
Ultimately, the screening procedures, in vitro and in vivo, allow for the implementation and use of probiotics able to survive and modulate the GIT. Without the ability to survive the GIT, the probiotic would never be able to limit or reduce Campylobacter in the hindgut. Further, these methodologies help us better understand how to combine these screened probiotics with other feed amendments. These combinations are the future of feed amendments in poultry as they allow for sustained affects in the GIT. However, it is important to understand the exact mechanisms behind the probiotic in order to combine with other amendments or supplements.
Probiotics play multiple roles in the poultry host, delivering beneficial effects, such as increasing nutrient uptake, and body weight gains. The mechanisms behind the host beneficial effects are complex and not always well defined, so only the mechanisms of antibacterial effects are discussed in this review (Lutful Kabir, 2009; Park et al., 2016; Popova, 2017; Wang et al., 2017; Peralta-Sánchez et al., 2019). The current known modes of action for probiotics as an antimicrobial is demonstrated in three ways including, but not limited to, competitive exclusion, antagonism, and stimulation of the host immune system (Zhang et al., 2007; Bratz et al., 2015; Cox and Dalloul, 2015; Schneitz and Hakkinen, 2016; Dec et al., 2018).
The competitive exclusion (CE) concept was first developed by Nurmi and Rantala (1973) when they attempted to limit the Salmonella proliferation in broiler flocks. Conceptionally, this approach introduced the intestinal or fecal bacteria from healthy Salmonella-free adult chickens to newly hatched chicks to reduce the Salmonella colonization in these chicks (Lutful Kabir, 2009; Boulianne et al., 2019). Later the CE concept was applied for controlling other enteropathogens, including Campylobacter, Clostridium, and E. coli in poultry production (Stern and Meinersmann, 1989; Stern et al., 2001; La Ragione et al., 2004; Schneitz, 2005). The traditional CE cultures utilize a mixture of undefined bacterial species and populations from the chicken GIT instead of known bacterial species, leading to conflicting observations among studies. Furthermore, the specific mechanisms of the undefined CE cultures were difficult to ascertain (Schoeni and Doyle, 1992; Schoeni and Wong, 1994). Numerous studies have been carried out to derive candidate anti-Campylobacter strains from chicken GIT contents and elaborate on the potential mechanisms of CE specifically directed toward Campylobacter. In general, the CE characteristic of probiotics acts through the nutrient competition and occupation of mucosal niches to reduce the Campylobacter colonization in the poultry host (Mead, 2000; Chaveerach et al., 2004; Pan and Yu, 2014). As reported by Nishiyama et al. (2014), the probiotic strain Lactobacillus gasseri SBT2055 (LG2055) reduced up to 25- and 100- fold of C. jejuni 81-176 adhesion and internalization to the human epithelial cell monolayer (Int407) in vitro. When daily orally administered to chickens, L. gasseri reduced C jejuni colonization by 250- fold in the cecum of 14-day-old birds post-challenge compared to the levels in the control group. It was found that a proteinaceous component on the LG2055 cell surface resulted in its co-aggregation with C. jejuni or competitive adhesion to Int407 cells, indicating that this surface component might play a vital role in the CE activity against C. jejuni (Nishiyama et al., 2014). To gain insight into the inhibition mechanism, the same group of researchers carried out another study focusing on the role of cell surface aggregation-promoting factors (APFs) of LG2055 (Nishiyama et al., 2015). The APFs are associated with the self-aggregation, maintenance of cell shape, and adhesion of L. gasseri. The study revealed that the primary inhibition mechanism of the AFP mediated competition was through adhesion to epithelial cells, instead of co-aggregation with C. jejuni. The LG2055 apf1 gene deletion mutant lost its inhibition effect to C. jejuni on Int407 cell monolayer and in the chicken GIT, while the wild type LG2055 reduced C. jejuni invasion in vitro by 177- fold and colonization in vivo by 230- fold.
Ganan et al. (2013) reported the protective effect of commercial human probiotic strains, L. rhamnosus GG, Propionibacterium freudenreichii ssp. shermanii JS and Lactococcus lactis ssp. lactis, on the poultry GIT mucus layer against Campylobacter infection. The intestinal mucus from broiler and turkeys was isolated and coated on microtiter plate wells for in vitro exclusion and competitive inhibition assays. When applied before Campylobacter infection, the probiotics reduced Campylobacter colonization by occupying the binding sites on intestinal mucus isolated from jejunum, colon, and cecum. However, when the probiotics and Campylobacter were simultaneously exposed to the mucus, the probiotics increased Campylobacter adhesion to the mucus. In vitro, L. casei outcompeted the attachment of C. jejuni to human epithelial cells INT407 when they were introduced at 1:1 ratio in a co-infection assay (Salaheen et al., 2014). When the ratio of L. casei and C. jejuni was lowered to 1:10, L. casei still significantly reduced C. jejuni colonization. However, further decreases in the number of L. casei to a 1:100 ratio failed to reduce C. jejuni attachment. The authors suggested that the initial number of L. casei should be high enough to exclude the C. jejuni. Therefore, the inhibition mechanism of L. casei may have been through the occupation of the host cell surface receptors that C. jejuni uses to recognize and subsequently bind to.
The antagonistic effects of probiotics include the production of antibacterial metabolites such as organic acids, H2O2, and bacteriocins. As many probiotic candidates are Lactic Acid Producing Bacteria (LAB), it is common for probiotics to produce a sufficient amount of organic acids to alter the pH of the surrounding environment and reduce pathogens. As such, Chaveerach et al. (2004) reported a probiotic candidate (Lactobacillus P93) isolated from the chicken GIT inhibited the growth of Campylobacter by producing organic acids and anti-Campylobacter proteins. In an in vitro co-culture, Neal-McKinney et al. (2012) reported that lactic acids produced by Lactobacillus were able to disrupt the cell membranes of C. jejuni leading to cell death. Similarly, the in vitro screening of the cell-free culture supernatants of seven Lactobacillus strains in a well-diffusion agar assay revealed the inhibition of C. jejuni and C. coli (Bratz et al., 2015). This inhibitory effect was achieved by the production of organic acids, which decreased the pH.
When the supernatants were pH neutralized before testing, they lost the inhibitory effect against Campylobacter. Another in vitro study also reported the inhibition of Campylobacter. Seven Lactobacillus isolates from the chicken GIT inhibited the growth of Campylobacter by producing organic acids that decreased the surrounding pH (Dec et al., 2018). However, the anti-Campylobacter effect was no longer observed after the pH of the cell-free supernatant was neutralized to 6.5 to 7.0. In some cases, the anti-Campylobacter ability of probiotics is due to the combined effects of the antagonistic and competitive exclusion properties. For example, in an in vitro study, the Lactobacillus fermentum 3872 was reported to bind to the same C. jejuni attachment receptor in host GIT as well as releasing lactic acid that inhibited C. jejuni (Lehri et al., 2017).
To support their invasion and establishment in the host, all major lineages of bacteria and archaea produce bacteriocins, antimicrobial peptides (Gillor et al., 2009; Svetoch and Stern, 2010; Hoang et al., 2011a; Lagha et al., 2017). The antibacterial function of bacteriocins is through the formation of pores in the cell wall. Bacteriocins bind to the cell wall of the target microorganism and interact with the outer cell membrane, leading to the formation of these pores and leakage of ions which, in turn, causes the death of the target cell (Svetoch and Stern, 2010; Prudêncio et al., 2015).
Based on in vitro studies, L. salivarius has been identified as a promising probiotic candidate due to its well-characterized ability to produce bacteriocins and anti-Salmonella and anti-C. jejuni effects (Messaoudi et al., 2013). More recently, 44 strains of LAB were screened as potential probiotics by Ayala et al. (2019). Of the 44 screened strains, L. salivarius L28 was the “top-ranking” strain and possessed the most antagonistic features such as no antimicrobial resistance (AMR)-encoding genes in mobile elements, ability to produce bacteriocins and adhere to the epithelial cells, and low cytotoxicity percentages. In other studies, L. salivarius SMXD51, MMS122, and MMS151 were reported to be ideal bacteriocin producers against Campylobacter (Messaoudi et al., 2011). Saint-Cyr et al. (2017) tested the bacteriocin of L. salivarius SMXD51 in vitro against 23 C. jejuni strains isolated from poultry farms and retail operations. The bacteriocin exhibited inhibition (<4 mm inhibition zone) of 15 strains and strong inhibition (>4 mm inhibition zone) of two strains of C. jejuni (AC4700 and C94). The remaining four C. jejuni strains were not inhibited by the bacteriocin, indicating the difference in sensitivity of the C. jejuni strains to the L. salivarius SMXD51 bacteriocin.
Thus, the variation of bacteriocin sensitivity among Campylobacter strains presents difficulties in applying bacteriocin as an on-farm control measure. Hoang et al. (2011b) tested the prevalence of bacteriocin-resistance in C. jejuni and C. coli isolates from various sources including humans (15), bovines (5), chickens (121), turkeys (1), pigs (4) and environment [i.e., trapped mice (5), bird droppings (5) and lagoons (1)]. Except for one C. coli strain, all the strains were susceptible to the tested bacteriocins, OR-7 and E-760, produced by chicken derived probiotic strains L. salivarius and E. faecium, respectively. The MIC of C. coli was 64 mg/mL, compared to other strains with MIC ranges between 0.25 and 4 mg/mL. To identify the genes involved with bacteriocin resistance, the authors compared the OR-7 resistant C. jejuni mutant strain with the parent strain using microarray analysis. It was concluded that the multidrug efflux pump CmeABC played a role in the bacterial resistance to the OR-7 bacteriocin. Previous research had revealed the contribution of CmeABC to the resistance against antibiotics and natural antimicrobials in Campylobacter.
Although the multidrug efflux pump CmeABC is responsible for the resistance of Campylobacter against bacteriocins, bacteriocins are still effective against Campylobacter. Stern et al. (2008) demonstrated that two cell-free bacteriocins (250 mg/kg feed) produced by Paenibacillus polymyxa and L. salivarius reduced C. jejuni by at least a 6 log10 CFU in chickens. In contrast, the ingestion of living cells (107–108 CFU/chick) of the producer strains did not elicit any inhibitory effects toward C. jejuni. These two strains failed to exclude the C. jejuni and occupy the colonization sites in chicken intestine. The bacteriocin was potentially produced in the host GIT in limited amounts. However, there was no reason for the strain to overproduce bacteriocins, given the high energy costs for carrying plasmid and toxin production (Dobson et al., 2012). Another potential reason for the ineffectiveness of the bacteriocin producers was that their populations were too low compared to the surrounding microorganisms. In this case, the impact of the bacteriocin was limited. The advantage of killing other competitors could not compensate for the metabolic cost for the production of bacteriocin (Riley and Wertz, 2002). Also, the GIT might not present the optimal environmental conditions for triggering the maximum in vivo production of bacteriocin, unlike laboratory growth medium and conditions (Telke et al., 2019).
Thus far, few studies have demonstrated a probiotic-triggered immune response against Campylobacter in poultry. However, the interaction between Campylobacter and the chicken immune system is not well characterized, unlike other food pathogens such as E. coli and Salmonella (Shaughnessy et al., 2009; Chintoan-Uta, 2016). In chickens, the gut-associated lymphoid tissues (GALTs) play a crucial role in the poultry intestinal immune system. The GALT is comprised of lymphoid structures such as the bursa of Fabricius, cecal tonsils, Peyer’s patch, Meckel’s diverticulum, and lymphocyte aggregates distributed in the epithelial lining and the lamina propria (Lillehoj and Trout, 1996; Kim and Lillehoj, 2019). The poultry immune system of poultry includes the innate and adaptive immune responses, with the latter system further divided into the humoral and cell-mediated immune response. The initiation of the innate immune response begins with the recognition of pathogen-associated molecular patterns (PAMPs) by the pathogen recognition receptors (PRRs; Kim and Lillehoj, 2019). There are several important receptors in chicken GIT responsible for bacteria recognition, including the Toll-like receptor (TLR)-2 for peptidoglycan, TLR-4 for lipopolysaccharides, TLR-5 for flagellin, and TLR-21 for unmethylated CpG DNA of bacteria (Wigley, 2013). The corresponding recognition by these receptors leads to the production of antimicrobial peptides and cytokines (e.g., IL-10) by the epithelial cells, further activating the lymphocytes. In addition, the signal triggers the B cells to produce secretory IgA (sIgA) (Kim and Lillehoj, 2019).
The adaptive immune response is an antigen-specific response by B cells and T cells. The triggered B cell differentiates and produces antigen-specific immunoglobulin (Ig) antibodies that interact and destroy the extracellular antigens (Lillehoj and Trout, 1996; Erf, 2004). However, when antigens have already entered host cells, the cell-mediated response plays a role in eliminating the intracellular antigens (Erf, 2004). The cell-mediated immune response includes the activation of different cells such as T lymphocytes, NK cells, and macrophages, whereas T cells are further divided into cytotoxic T lymphocytes (CD8+) and helper T cells (CD4+) (Lillehoj and Trout, 1996).
The colonization of Campylobacter has been reported to trigger both innate and adaptive immune responses of the chicken host. Vaezirad et al. (2017) reported that the immunosuppressed chickens exhibited more rapid C. jejuni colonization and dissemination to the liver. At 17 days of age, the chickens were treated with glucocorticoid (GC), which dampened the innate immune response. In that study, higher concentrations of bacteria were present in the cecal contents and liver compared to the non-GC treated group 2–4 days post the C. jejuni challenge. These results indicated that the chicken immune system might play a role in limiting the invasion and dissemination of C. jejuni. The role of B lymphocytes on the colonization of C. jejuni in broilers has also been studied. Lacharme-Lora et al. (2017) compared healthy chickens to B lymphocyte depleted chickens to determine the exact role of B lymphocytes on C. jejuni colonization (Lacharme-Lora et al., 2017). At day-of-hatch, chickens were bursectomized using cyclophosphamide. The bursectomy depleted over 90% of bursal B cells and disabled the anti-C. jejuni IgY and IgM production under C. jejuni exposure. At 14 and 28 days post-inoculation (DPI), the cecal C. jejuni levels were high regardless of the bursectomized or control groups. However, C. jejuni in the jejunum and ileum cleared by 28 DPI in the control groups and not in the bursectomized group. By 63 DPI, the shedding levels of C. jejuni in the control birds decreased while the bursectomized group remained unchanged. Therefore, the B lymphocytes played an essential role in the small intestine but not in the ceca during C. jejuni colonization.
The GIT microbiota and probiotics are known to stimulate the immune response against pathogens (Mahfuz et al., 2017; Willson et al., 2018). Several studies have reported the enhancing effects of probiotics on the chicken immune response (Brisbin et al., 2015; Shojadoost et al., 2019; Šefcová et al., 2020). The presence of probiotics trigger the host immune response, depending on the bacterial strains and experimental conditions.
Haghighi et al. (2006) reported elevated production of several antibodies in day-of-hatch chickens fed a probiotic cocktail containing Lactobacillus acidophilus, B. bifidum, and Streptococcus faecalis. Specifically, there were increases of immunoglobulin G (IgG) in the GIT against tetanus toxoid (TT) and IgG and IgM in serum against TT and the Clostridium perfringens alpha-toxin, compared to non-probiotic treated chickens. At 6 weeks of age, the supplemented probiotic Bacillus subtilis enhanced the serum IgM levels and the cell-mediated immune response in chickens grown under high ambient temperatures (ave. 29–32°C) compared to the negative control group (Fathi et al., 2017). The probiotics showed no significant effect on IgA and IgY levels (Fathi et al., 2017). In contrast, Bai et al. (2017) reported increased levels of IgA and IgG levels in the serum of chickens fed B. subtilis fmbJ supplemented diets at 42 days of age.
Furthermore, Sadeghi et al. (2015) reported an enhanced response to the Newcastle and infectious bursal viruses when Salmonella challenged broilers were fed diets supplemented with a commercial probiotic (Gallipro, Chr Hansen, Milwalkee, WI, United States). The challenge of Salmonella enterica decreased the antibody titer, lymphocyte count, and the weight of spleen and bursa in chickens fed a no-probiotic supplemented diet. However, broilers fed the B. subtilis supplemented diets showed increased levels of antibodies against the Newcastle virus (18%) at 21-days of age and against the Newcastle (21%) and infectious bursal viruses (14%) at day 42. Also, the virus antibodies were not affected by the B. subtilis probiotic under Salmonella-free conditions. Though the antibody increase due to the supplementation of the probiotic diet was not further investigated, the authors offered some potential explanations. First, B. subtilis could reduce Salmonella colonization through inhibitory mechanisms such as CE and antagonism, thus reduce the negative effect of Salmonella on antibody levels. Also, Salmonella is known to stimulate the production of interleukin-1β (IL-1β), and B. subtilis were reported to suppress the pro-inflammatory cytokines, including IL-1β and increase the anti-inflammatory cytokines. Another more recent in vitro study investigated the immune-modulation caused by L. salivarius, L. johnsonii, L. reuteri, L. crispatus, and L. gasseri. The treatment of chicken macrophages with every species of the heat-killed Lactobacilli at a multiplicity of infection (MOI) of 100 increased nitric oxide (NO) production (an indicator of macrophage activation) except for L. reuteri. Moreover, the treatment of the heat-killed single or mixed cultures significantly increased the phagocytosis by macrophages on the fluorescein isothiocyanate labeled C. jejuni (Taha-Abdelaziz et al., 2019).
The feed supplementation of probiotics has been reported as an effective pathogen control strategy for poultry over several decades; however, a thorough mechanistic understanding is still missing for it to be utilized as a routine measure to limit Campylobacter colonization. In the present review, the characteristics of Campylobacter colonization and transmission in poultry were discussed. Also, the current preharvest intervention measures were briefly addressed, followed by a discussion on anti-Campylobacter probiotics. The screening process and functioning mechanisms of probiotics were reviewed. The number of studies on probiotics screening is considerable, but the variations among studies led to difficulties for direct comparisons to reach general conclusions. Several studies may observe the reduction of C. jejuni by the same probiotic strain, but their reported number of log reductions may differ. This variation might be contributed by the difference of experimental conditions and design, such as the broiler age, the time of administration of the probiotic treatment, and subsequent Campylobacter challenge. In addition, the potential functional mechanisms of probiotics were discussed, which may help understand the kinetics of Campylobacter reduction and provide directions for future screening work.
Overall, the addition of probiotics to feed is effective for reducing Campylobacter in poultry, but it cannot achieve complete elimination. Therefore, more studies on the effects of combining probiotics with other control preventions such as prebiotics and postbiotics represent an opportunity to reduce or eliminate Campylobacter. The inconsistency of Campylobacter reduction by probiotic treatment is another barrier hindering the widespread application of probiotics at the farm level. The effect on probiotic performance by factors including Campylobacter strain, variations among farms, different genetic lines, feed composition, and environmental procedures should be further elaborated. In addition, the laborious and time-consuming drawbacks of probiotic screening work have hindered the progress of the application of probiotics in poultry production. In more recent years, the availability of omics technology (i.e., metagenomic, transcriptomic, and metabolomic methods) has provided researchers a means to estimate the functions of GIT bacteria and their host and their responses to pathogens based on the characterization of the bacterial and host genetic information (Ricke et al., 2019). Thus, the utilization of omics can be employed to efficiently facilitate the identification of probiotic candidates (Papadimitriou et al., 2015; Rebollar et al., 2016). These approaches, combined with machine-learning techniques, have been widely studied for applications in numerous fields, including microbiology. The ability to predict the interactions among microorganisms can be applied in probiotic screening and therefore enable a more rapid screening process (Qu et al., 2019; van den Bogert et al., 2019; Zhou and Gallins, 2019).
All authors significantly contributed to the work of the current review. WD wrote the current review with the assistance from DD who formatted, edited, and submitted the review with direction from SR. HP, WC, YY, and SR edited the review before submission.
The authors declare that this review received funding from Diamond V to cover publishing fees. The funder was not involved in the design, interpretation of data, the writing of this article, or the decision to submit it for publication.
HP and WC are employees of Diamond V.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
DD would like to knowledge the Graduate College at the University of Arkansas for its support through the Distinguished Academy Fellowship and the continued support from the Cell and Molecular Biology Program and Department of Food Science at the University of Arkansas.
Aguiar, V. F., Donoghue, A. M., Arsi, K., Reyes-Herrera, I., Metcalf, J. H., de los Santos, F. S., et al. (2013). Targeting motility properties of bacteria in the development of probiotic cultures against Campylobacter jejuni in broiler chickens. Foodborne Pathog. Dis. 10, 435–441. doi: 10.1089/fpd.2012.1302
Allos, B. M. (2001). Campylobacter jejuni infections: update on emerging issues and trends. Clin. Infect. Dis. 32, 1201–1206. doi: 10.1086/319760
Applegate, T. J., Klose, V., Steiner, T., Ganner, A., and Schatzmayr, G. (2010). Probiotics and phytogenics for poultry: myth or reality? J. Appl. Poultry Res. 19, 194–210. doi: 10.3382/japr.2010-00168
Arsi, K., Donoghue, A. M., Woo-Ming, A., Blore, P. J., and Donoghue, D. J. (2015a). Intracloacal inoculation, an effective screening method for determining the efficacy of probiotic bacterial isolates against Campylobacter colonization in broiler chickens. J. Food Prot. 78, 209–213. doi: 10.4315/0362-028X.JFP-14-326
Arsi, K., Donoghue, A. M., Woo-Ming, A., Blore, P. J., and Donoghue, D. J. (2015b). The efficacy of selected probiotic and prebiotic combinations in reducing Campylobacter colonization in broiler chickens. J. Appl. Poult. Res. 24, 327–334. doi: 10.3382/japr/pfv032
Awad, W. A., Hess, C., and Hess, M. (2018). Re-thinking the chicken–Campylobacter jejuni interaction: a review. Avian. Pathol. 47, 352–363. doi: 10.1080/03079457.2018.1475724
Awad, W. A., Mann, E., Dzieciol, M., Hess, C., Schmitz-Esser, S., Wagner, M., et al. (2016). Age-related differences in the luminal and mucosa-associated gut microbiome of broiler chickens and shifts associated with Campylobacter jejuni infection. Front. Cell Infect. Microbiol. 6:154. doi10.3389/fcimb.2016.00154
Awad, W. A., Molnár, A., Aschenbach, J. R., Ghareeb, K., Khayal, B., Hess, C., et al. (2015). Campylobacter infection in chickens modulates the intestinal epithelial barrier function. Innate Immun. 21, 151–160. doi: 10.1177/1753425914521648
Ayala, D. I, Cook, P. W., Franco, J. G., Bugarel, M., Loneragan, G. H., Brashears, M. M., et al. (2019). A systematic approach to identify and characterize the effectiveness and safety of novel probiotic strains to control foodborne pathogens. Front. Microbiol. 10:1108. doi: 10.3389/fmicb.2019.01108
Baffoni, L., Gaggìa, F., Garofolo, G., Di Serafino, G., Buglione, E., Di Giannatale, E., et al. (2017). Evidence of Campylobacter jejuni reduction in broilers with early synbiotic administration. Int. J. Food Microbiol. 251, 41–47. doi: 10.1016/j.ijfoodmicro.2017.04.001
Bai, K., Huang, Q., Zhang, J., He, J., Zhang, L., and Wang, T. (2017). Supplemental effects of probiotic Bacillus subtilis fmbJ on growth performance, antioxidant capacity, and meat quality of broiler chickens. Poultry Sci. 96, 74–82. doi: 10.3382/ps/pew246
Battersby, T., Whyte, P., and Bolton, D. J. (2016). The pattern of Campylobacter contamination on broiler farms; external and internal sources. J. Appl. Microbiol. 120, 1108–1118. doi: 10.1111/jam.13066
Bauer, A. W., Kirby, W. M. M., Sherris, J. C., and Turck, M. (1966). Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 45, 493–496. doi: 10.1093/ajcp/45.4_ts.493
Beery, J. T., Hugdahl, M. B., and Doyle, M. P. (1988). Colonization of gastrointestinal tracts of chicks by Campylobacter jejuni. Appl. Environ. Microbiol. 54, 2365–2370. doi: 10.1128/aem.54.10.2365-2370.1988
Bhatia, S. J., Kochar, N., Abraham, P., Nair, N. G., and Mehta, A. P. (1989). Lactobacillus acidophilus inhibits growth of Campylobacter pylori in vitro. J. Clin. Microbiol. 27, 2328–2330. doi: 10.1128/jcm.27.10.2328-2330.1989
Blajman, J., Gaziano, C., Zbrun, M. V., Soto, L., Astesana, D., Berisvil, A., et al. (2015). In vitro and in vivo screening of native lactic acid bacteria toward their selection as a probiotic in broiler chickens. Res. Vet. Sci. 101, 50–56. doi: 10.1016/j.rvsc.2015.05.017
Boulianne, M., Logue, C. M., McDougald, L. R., Nair, V., and Suarez, D. L. (2019). Diseases of Poultry. Hoboken, NJ: John Wiley & Sons.
Bratz, K., Gölz, G., Janczyk, P., Nöckler, K., and Alter, T. (2015). Analysis of in vitro and in vivo effects of probiotics against Campylobacter spp. Berliner und Münchener tierärztliche Wochenschrift 128, 155–162. doi: 10.2376/0005-9366-128-155
Brisbin, J. T., Davidge, L., Roshdieh, A., and Sharif, S. (2015). Characterization of the effects of three Lactobacillus species on the function of chicken macrophages. Res. Vet. Sci. 100, 39–44. doi: 10.1016/j.rvsc.2015.03.003
Butzler, J.-P. (2004). Campylobacter, from obscurity to celebrity. Clin. Microbiol. Infect. 10, 868–876. doi: 10.1111/j.1469-0691.2004.00983.x
Callicott, K. A., Friðriksdóttir, V., Reiersen, J., Lowman, R., Bisaillon, J.-R., Gunnarsson, E., et al. (2006). Lack of evidence for vertical transmission of Campylobacter spp. in chickens. Appl. Environ. Microbiol. 72, 5794–5798. doi: 10.1128/AEM.02991-05
Cawthraw, S. A., and Newell, D. G. (2010). Investigation of the presence and protective effects of maternal antibodies against Campylobacter jejuni in chickens. AvianDis. 54, 86–93. doi: 10.1637/9004-072709-Reg.1
CDC (2019). Antibiotic Resistance | Campylobacter | CDC. Available online at: https://www.cdc.gov/campylobacter/campy-antibiotic-resistance.html (accessed May 16, 2020).
Cean, A., Stef, L., Simiz, E., Julean, C., Dumitrescu, G., Vasile, A., et al. (2015). Effect of human isolated probiotic bacteria on preventing Campylobacter jejuni colonization of poultry. Foodborne Pathog. Dis. 12, 122–130. doi: 10.1089/fpd.2014.1849
Centers for Disease Control and Prevention (2018). Reports of Selected Campylobacter Outbreak Investigations. Available online at: https://www.cdc.gov/campylobacter/outbreaks/outbreaks.html (accessed May 12, 2019).
Chaveerach, P., Lipman, L. J. A., and van Knapen, F. (2004). Antagonistic activities of several bacteria on in vitro growth of 10 strains of Campylobacter jejuni/coli. Int. J. Food Microbiol. 90, 43–50. doi: 10.1016/S0168-1605(03)00170-3
Chintoan-Uta, C. (2016). The host-pathogen interaction in Campylobacter jejuni infection of chickens: an understudied aspect that is crucial for effective control. Virulence 8, 241–243. doi: 10.1080/21505594.2016.1240860
CLSI (2018). M100Ed29 | Performance Standards for Antimicrobial Susceptibility Testing, 29th Edn. Available online at: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed May 12, 2019).
Connerton, P. L., Richards, P. J., Lafontaine, G. M., O’Kane, P. M., Ghaffar, N., Cummings, N. J., et al. (2018). The effect of the timing of exposure to Campylobacter jejuni on the gut microbiome and inflammatory responses of broiler chickens. Microbiome 6:88. doi: 10.1186/s40168-018-0477-5
Cox, C. M., and Dalloul, R. A. (2015). Immunomodulatory role of probiotics in poultry and potential in ovo application. Benef Microbes 6, 45–52. doi: 10.3920/BM2014.0062
Cox, N. A., Richardson, L. J., Maurer, J. J., Berrang, M. E., Fedorka-Cray, P. J., Buhr, R. J., et al. (2012). Evidence for horizontal and vertical transmission in Campylobacter passage from hen to her progeny. J. Food Prot. 75, 1896–1902. doi: 10.4315/0362-028.JFP-11-322
Cox, N. A., Stern, N. J., Hiett, K. L., and Berrang, M. E. (2002). Identification of a new source of Campylobacter contamination in poultry: transmission from breeder hens to broiler chickens. Avian Dis. 46, 535–541. doi: 10.1637/0005-2086(2002)046[0535:ioanso]2.0.co;2
Danielsen, M., and Wind, A. (2003). Susceptibility of Lactobacillus spp. to antimicrobial agents. Int. J. Food Microbiol. 82, 1–11. doi: 10.1016/S0168-1605(02)00254-4
Dec, M., Nowaczek, A., Urban-Chimiel, R., Stêpień-pyśniak, D., and Wernicki, A. (2018). Probiotic potential of Lactobacillus isolates of chicken origin with anti-Campylobacter activity. J. Vet. Med. Sci. 80, 1195–1203. doi: 10.1292/jvms.18-0092
Dittoe, D. K., Ricke, S. C., and Kiess, A. S. (2018). Organic acids and potential for modifying the avian gastrointestinal tract and reducing pathogens and disease. Front. Vet. Sci. 5:216. doi: 10.3389/fvets.2018.00216
Dobson, A., Cotter, P. D., Ross, R. P., and Hill, C. (2012). Bacteriocin production: a probiotic trait? Appl. Environ. Microbiol. 78, 1–6. doi: 10.1128/AEM.05576-11
Domingues, A. R., Pires, S. M., Halasa, T., and Hald, T. (2012). Source attribution of human campylobacteriosis using a meta-analysis of case-control studies of sporadic infections. Epidem. Infect. 140, 970–981. doi: 10.1017/S0950268811002676
Eeckhaut, V., Wang, J., Van Parys, A., Haesebrouck, F., Joossens, M., Falony, G., et al. (2016). The probiotic Butyricicoccus pullicaecorum reduces feed conversion and protects from potentially harmful intestinal microorganisms and necrotic enteritis in broilers. Front. Microbiol. 7:1416. doi: 10.3389/fmicb.2016.01416
Ehrmann, M. A., Kurzak, P., Bauer, J., and Vogel, R. F. (2002). Characterization of lactobacilli towards their use as probiotic adjuncts in poultry. J. Appl. Microbiol. 92, 966–975. doi: 10.1046/j.1365-2672.2002.01608.x
Erf, G. F. (2004). Cell-mediated immunity in poultry. Poultry Sci. 83, 580–590. doi: 10.1093/ps/83.4.580
European Food Safety Authority (2018). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 16:e05500. doi: 10.2903/j.efsa.2018.5500
Fathi, M. M., Ebeid, T. A., Al-Homidan, I., Soliman, N. K., and Abou-Emera, O. K. (2017). Influence of probiotic supplementation on immune response in broilers raised under hot climate. Br. Poultry Sci. 58, 512–516. doi: 10.1080/00071668.2017.1332405
Feye, K. M., Baxter, M. F. A., Tellez-Isaias, G., Kogut, M. H., and Ricke, S. C. (2020). Influential factors on the composition of the conventionally raised broiler gastrointestinal microbiomes. Poultry Sci. 99, 653–659. doi: 10.1016/j.psj.2019.12.013
Fooks, L. J., and Gibson, G. R. (2002). In vitro investigations of the effect of probiotics and prebiotics on selected human intestinal pathogens. FEMS Microbiol. Ecol. 39, 67–75. doi: 10.1111/j.1574-6941.2002.tb00907.x
Forte, C., Manuali, E., Abbate, Y., Papa, P., Vieceli, L., Tentellini, M., et al. (2018). Dietary Lactobacillus acidophilus positively influences growth performance, gut morphology, and gut microbiology in rurally reared chickens. Poultry Sci. 97, 930–936. doi: 10.3382/ps/pex396
FSAI Ireland (2002). Control of Campylobacter species in the food chain. Available online at: https://www.lenus.ie/handle/10147/44795 (accessed April 5, 2019).
Ganan, M., Martinez-Rodriguez, A. J., Carrascosa, A. V., Vesterlund, S., Salminen, S., and Satokari, R. (2013). Interaction of Campylobacter spp. and human probiotics in chicken intestinal mucus. Zoonoses Pub. Health 60, 141–148. doi: 10.1111/j.1863-2378.2012.01510.x
García-Hernández, Y., Pérez-Sánchez, T., Boucourt, R., Balcázar, J. L., Nicoli, J. R., Moreira-Silva, J., et al. (2016). Isolation, characterization and evaluation of probiotic lactic acid bacteria for potential use in animal production. Res. Vet. Sci. 108, 125–132. doi: 10.1016/j.rvsc.2016.08.009
Garriga, M., Pascual, M., Monfort, J. M., and Hugas, M. (1998). Selection of lactobacilli for chicken probiotic adjuncts. J. Appl. Microbiol. 84, 125–132. doi: 10.1046/j.1365-2672.1997.00329.x
Geissler, A. L., Bustos Carrillo, F., Swanson, K., Patrick, M. E., Fullerton, K. E., Bennett, C., et al. (2017). Increasing Campylobacter infections, outbreaks, and antimicrobial resistance in the United States, 2004-2012. Clin. Infect. Dis. 65, 1624–1631. doi: 10.1093/cid/cix624
Ghareeb, K., Awad, W. A., Mohnl, M., Porta, R., Biarnés, M., Böhm, J., et al. (2012). Evaluating the efficacy of an avian-specific probiotic to reduce the colonization of Campylobacter jejuni in broiler chickens. Poultry Sci. 91, 1825–1832. doi: 10.3382/ps.2012-02168
Gillor, O., Giladi, I., and Riley, M. A. (2009). Persistence of colicinogenic Escherichia coli in the mouse gastrointestinal tract. BMC Microbiol. 9:165. doi: 10.1186/1471-2180-9-165
Gong, J., Forster, R. J., Yu, H., Chambers, J. R., Wheatcroft, R., Sabour, P. M., et al. (2002). Molecular analysis of bacterial populations in the ileum of broiler chickens and comparison with bacteria in the cecum. FEMS Microbiol. Ecol. 41, 171–179. doi: 10.1111/j.1574-6941.2002.tb00978.x
Gong, J., Si, W., Forster, R. J., Huang, R., Yu, H., Yin, Y., et al. (2007). 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: from crops to ceca. FEMS Microbiol. Ecol. 59, 147–157. doi: 10.1111/j.1574-6941.2006.00193.x
Gupta, A., Nelson, J. M., Barrett, T. J., Tauxe, R. V., Rossiter, S. P., Friedman, C. R., et al. (2004). Antimicrobial resistance among Campylobacter strains, United States, 1997–2001. Emerg. Infect. Dis. 10, 1102–1109. doi: 10.3201/eid1006.030635
Guyard-Nicodème, M., Keita, A., Quesne, S., Amelot, M., Poezevara, T., Le Berre, B., et al. (2016). Efficacy of feed additives against Campylobacter in live broilers during the entire rearing period. Poultry Sci. 95, 298–305. doi: 10.3382/ps/pev303
Haghighi, H. R., Gong, J., Gyles, C. L., Hayes, M. A., Zhou, H., Sanei, B., et al. (2006). Probiotics stimulate production of natural antibodies in chickens. Clin. Vaccine Immunol. 13, 975–980. doi: 10.1128/CVI.00161-06
Hald, T., Aspinall, W., Devleesschauwer, B., Cooke, R., Corrigan, T., Havelaar, A. H., et al. (2016). World Health Organization estimates of the relative contributions of food to the burden of disease due to selected foodborne hazards: a structured expert elicitation. PLoS One 11:e0145839. doi: 10.1371/journal.pone.0145839
Han, Z., Willer, T., Li, L., Pielsticker, C., Rychlik, I., Velge, P., et al. (2017). Influence of the gut microbiota composition on Campylobacter jejuni colonization in chickens. Infect. Immun. 85:e00380-17. doi: 10.1128/IAI.00380-17
Helmy, Y. A., Kassem, I. I., Kumar, A., and Rajashekara, G. (2017). In vitro evaluation of the impact of the probiotic E. coli Nissle 1917 on Campylobacter jejuni’s invasion and intracellular survival in human colonic cells. Front. Microbiol. 8:1588. doi: 10.3389/fmicb.2017.01588
Hermans, D., Van Deun, K., Messens, W., Martel, A., Van Immerseel, F., Haesebrouck, F., et al. (2011). Campylobacter control in poultry by current intervention measures ineffective: urgent need for intensified fundamental research. Vet. Microbiol. 152, 219–228. doi: 10.1016/j.vetmic.2011.03.010
Hoang, K. V., Stern, N. J., and Lin, J. (2011a). Development and stability of bacteriocin resistance in Campylobacter spp. J. Appl. Microbiol. 111, 1544–1550. doi: 10.1111/j.1365-2672.2011.05163.x
Hoang, K. V., Stern, N. J., Saxton, A. M., Xu, F., Zeng, X., and Lin, J. (2011b). Prevalence, development, and molecular mechanisms of bacteriocin resistance in Campylobacter. Appl. Environ. Microbiol. 77, 2309–2316. doi: 10.1128/AEM.02094-10
Hossain, M. I., Sadekuzzaman, M., and Ha, S.-D. (2017). Probiotics as potential alternative biocontrol agents in the agriculture and food industries: a review. Food Res. Inter. 100, 63–73. doi: 10.1016/j.foodres.2017.07.077
Humphrey, S., Chaloner, G., Kemmett, K., Davidson, N., Williams, N., Kipar, A., et al. (2014). Campylobacter jejuni is not merely a commensal in commercial broiler chickens and affects bird welfare. mBio 5:e01364-14. doi: 10.1128/mBio.01364-14
Hwang, H., and Singer, R. S. (2020). Survey of the U.S. broiler industry regarding pre- and post-harvest interventions targeted to mitigate Campylobacter contamination on broiler chicken products. J. Food Prot. 83, 1137–1148. doi: 10.4315/JFP-19-527
Imperial, I. C. V. J., and Ibana, J. A. (2016). Addressing the antibiotic resistance problem with probiotics: reducing the risk of its double-edged sword effect. Front. Microbiol. 7:1983. doi: 10.3389/fmicb.2016.01983
Indikova, I., Humphrey, T. J., and Hilbert, F. (2015). Survival with a helping hand: Campylobacter and microbiota. Front. Microbiol. 6:1266. doi: 10.3389/fmicb.2015.01266
Iovine, N. M., and Blaser, M. J. (2004). Antibiotics in animal feed and spread of resistant Campylobacter from poultry to humans. Emerg. Infect. Dis. 10, 1158–1189. doi: 10.3201/eid1006.040403
Ishikawa, H., Kutsukake, E., Fukui, T., Sato, I., Shirai, T., Kurihara, T., et al. (2010). Oral administration of heat-killed Lactobacillus plantarum strain b240 protected mice against Salmonella enterica serovar Typhimurium. Biosci. Biotechnol. Biochem. 74, 1338–1342. doi: 10.1271/bbb.90871
Jin, L. Z., Ho, Y. W., Abdullah, N., Ali, M. A., and Jalaludin, S. (1996). Antagonistic effects of intestinal Lactobacillus isolates on pathogens of chicken. Lett. Appl. Microbiol. 23, 67–71. doi: 10.1111/j.1472-765x.1996.tb00032.x
Johnson, T. J., Shank, J. M., and Johnson, J. G. (2017). Current and potential treatments for reducing Campylobacter colonization in animal hosts and disease in humans. Front. Microbiol. 8:487. doi: 10.3389/fmicb.2017.00487
Kaakoush, N. O., Castaño-Rodríguez, N., Mitchell, H. M., and Man, S. M. (2015). Global epidemiology of Campylobacter infection. Clin. Microbiol. Rev. 28, 687–720. doi: 10.1128/CMR.00006-15
Kabir, S. M. L., Rahman, M. M., Rahman, M. B., Rahman, M. M., and Ahmed, S. U. (2004). The dynamics of probiotics on growth performance and immune response in broilers. Int. J. Poultry Sci. 3, 361–364. doi: 10.3923/ijps.2004.361.364
Kalupahana, R. S., Kottawatta, K. S. A., Kanankege, K. S. T., van Bergen, M. A. P., Abeynayake, P., and Wagenaar, J. A. (2013). Colonization of Campylobacter spp. in broiler chickens and laying hens reared in tropical climates with low-biosecurity housing. Appl. Environ. Microbiol. 79, 393–395. doi: 10.1128/AEM.02269-12
Kergourlay, G., Messaoudi, S., Dousset, X., and Prévost, H. (2012). Genome sequence of Lactobacillus salivarius SMXD51, a potential probiotic strain isolated from chicken cecum, showing anti-Campylobacter activity. J. Bacteriol. 194, 3008–3009. doi: 10.1128/JB.00344-12
Kim, J.-A., Bayo, J., Cha, J., Choi, Y. J., Jung, M. Y., Kim, D.-H., et al. (2019). Investigating the probiotic characteristics of four microbial strains with potential application in feed industry. PLoS One 14:e0218922. doi: 10.1371/journal.pone.0218922
Kim, W. H., and Lillehoj, H. S. (2019). Immunity, Immunomodulation, and Antibiotic Alternatives to Maximize the Genetic Potential of Poultry for Growth and Disease Response. Available online at: https://pubag.nal.usda.gov/catalog/6161486 (accessed June 23, 2019).
Kmet, V., Callegari, M. L., Bottazzi, V., and Morelli, L. (1995). Aggregation-promoting factor in pig intestinal Lactobacillus strains. Lett. Appl. Microbiol. 21, 351–353. doi: 10.1111/j.1472-765X.1995.tb01079.x
Kmet, V., and Lucchini, F. (1997). Aggregation-promoting factor in human vaginal Lactobacillus strains. FEMS Immunol. Med. Microbiol. 19, 111–114. doi: 10.1111/j.1574-695X.1997.tb01079.x
Kobierecka, P. A., Olech, B., Ksiâżek, M., Derlatka, K., Adamska, I., Majewski, P. M., et al. (2016a). Cell wall anchoring of the Campylobacter antigens to Lactococcus lactis. Front. Microbiol. 7:165. doi: 10.3389/fmicb.2016.00165
Kobierecka, P. A., Wyszyńska, A. K., Aleksandrzak-Piekarczyk, T., Kuczkowski, M., Tuzimek, A., Piotrowska, W., et al. (2017). In vitro characteristics of Lactobacillus spp. strains isolated from the chicken digestive tract and their role in the inhibition of Campylobacter colonization. MicrobiologyOpen 6:e00512. doi: 10.1002/mbo3.512
Kobierecka, P. A., Wyszyńska, A. K., Gubernator, J., Kuczkowski, M., Wiśniewski, O., Maruszewska, M., et al. (2016b). Chicken anti-Campylobacter vaccine – comparison of various carriers and routes of immunization. Front. Microbiol. 7:740. doi: 10.3389/fmicb.2016.00740
La Ragione, R. M., Narbad, A., Gasson, M. J., and Woodward, M. J. (2004). In vivo characterization of Lactobacillus johnsonii FI9785 for use as a defined competitive exclusion agent against bacterial pathogens in poultry. Lett. Appl. Microbiol. 38, 197–205. doi: 10.1111/j.1472-765x.2004.01474.x
Lacharme-Lora, L., Chaloner, G., Gilroy, R., Humphrey, S., Gibbs, K., Jopson, S., et al. (2017). B lymphocytes play a limited role in clearance of Campylobacter jejuni from the chicken intestinal tract. Sci. Rep. 7:45090. doi: 10.1038/srep45090
Lagha, A. B., Haas, B., Gottschalk, M., and Grenier, D. (2017). Antimicrobial potential of bacteriocins in poultry and swine production. Vet. Res. 48:22. doi: 10.1186/s13567-017-0425-6
Lebeer, S., Vanderleyden, J., and De Keersmaecker, S. C. J. (2008). Genes and molecules of Lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 72, 728–764. doi: 10.1128/MMBR.00017-08
Lehri, B., Seddon, A. M., and Karlyshev, A. V. (2017). Lactobacillus fermentum 3872 as a potential tool for combatting Campylobacter jejuni infections. Virulence 8, 1753–1760. doi: 10.1080/21505594.2017.1362533
Lillehoj, H. S., and Trout, J. M. (1996). Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 9, 349–360. doi: 10.1128/cmr.9.3.349-360.1996
Lin, J. (2009). Novel approaches for Campylobacter control in poultry. Foodborne Pathog. Dis. 6, 755–765. doi: 10.1089/fpd.2008.0247
Lu, J., Idris, U., Harmon, B., Hofacre, C., Maurer, J. J., and Lee, M. D. (2003). Diversity and succession of the intestinal bacterial community of the maturing broiler chicken. Appl. Environ. Microbiol. 69, 6816–6824. doi: 10.1128/AEM.69.11.6816-6824.2003
Luangtongkum, T., Jeon, B., Han, J., Plummer, P., Logue, C. M., and Zhang, Q. (2009). Antibiotic resistance in Campylobacter: emergence, transmission and persistence. Future Microbiol. 4, 189–200. doi: 10.2217/17460913.4.2.189
Lutful Kabir, S. M. (2009). The role of probiotics in the poultry industry. Int. J. Mol. Sci. 10, 3531–3546. doi: 10.3390/ijms10083531
Mahfuz, S., Nahar, M. J., Mo, C., Ganfu, Z., Zhongjun, L., and Hui, S. (2017). Inclusion of probiotic on chicken performance and immunity: a review. Int. J. Poultry Sci. 16, 328–335. doi: 10.3923/ijps.2017.328.335
Marder, E. P., Griffin, P. M., Cieslak, P. R., Dunn, J., Hurd, S., Jervis, R., et al. (2018). Preliminary incidence and trends of infections with pathogens transmitted commonly through food — foodborne diseases active surveillance network, 10 U.S. Sites, 2006–2017. MMWR Morb. Mortal. Wkly. Rep. 67, 324–328. doi: 10.15585/mmwr.mm6711a3
Marotta, F., Garofolo, G., Di Donato, G., Aprea, G., Platone, I., Cianciavicchia, S., et al. (2015). Population diversity of Campylobacter jejuni in poultry and its dynamic of contamination in chicken meat. BioMed Res. Int. 2015:859845. doi: 10.1155/2015/859845
Massacci, F. R., Lovito, C., Tofani, S., Tentellini, M., Genovese, D. A., De Leo, A. A. P., et al. (2019). Dietary Saccharomyces cerevisiae boulardii CNCM I-1079 Positively affects performance and intestinal ecosystem in broilers during a Campylobacter jejuni infection. Microorganisms 7:596. doi: 10.3390/microorganisms7120596
Mead, G. C. (2000). Prospects for “competitive exclusion” treatment to control Salmonellas and other foodborne pathogens in poultry. Vet. J. 159, 111–123. doi: 10.1053/tvjl.1999.0423
Messaoudi, S., Kergourlay, G., Rossero, A., Ferchichi, M., Prévost, H., Drider, D., et al. (2011). Identification of lactobacilli residing in chicken ceca with antagonism against Campylobacter. Int. Microbiol. 14, 103–110. doi: 10.2436/20.1501.01.140
Messaoudi, S., Manai, M., Kergourlay, G., Prévost, H., Connil, N., Chobert, J.-M., et al. (2013). Lactobacillus salivarius: bacteriocin and probiotic activity. Food Microbiol. 36, 296–304. doi: 10.1016/j.fm.2013.05.010
Meunier, M., Guyard-Nicodème, M., Dory, D., and Chemaly, M. (2016). Control strategies against Campylobacter at the poultry production level: biosecurity measures, feed additives and vaccination. J. Appl. Microbiol. 120, 1139–1173. doi: 10.1111/jam.12986
Meunier, M., Guyard-Nicodème, M., Vigouroux, E., Poezevara, T., Beven, V., Quesne, S., et al. (2017). Promising new vaccine candidates against Campylobacter in broilers. PLoS One 12:e0188472. doi: 10.1371/journal.pone.0188472
Micciche, A., Rothrock, M. J. J., Yang, Y., and Ricke, S. C. (2019). Essential oils as an intervention strategy to reduce Campylobacter in poultry production: a review. Front. Microbiol. 10:1051. doi: 10.3389/fmicb.2019.01058
Mortada, M., Cosby, D. E., Shanmugasundaram, R., and Selvaraj, R. K. (2020). In vivo and in vitro assessment of commercial probiotic and organic acid feed additives in broilers challenged with Campylobacter coli. J. Appl. Poultry Res. 29, 435–446. doi: 10.1016/j.japr.2020.02.001
Mughini-Gras, L., Smid, J. H., Wagenaar, J. A., Boer, A. D., Havelaar, A. H., Friesema, I. H. M., et al. (2014). Campylobacteriosis in returning travelers and potential secondary transmission of exotic strains. Epid. Infect. 142, 1277–1288. doi: 10.1017/S0950268813002069
Nami, Y., Vaseghi Bakhshayesh, R., Mohammadzadeh Jalaly, H., Lotfi, H., Eslami, S., and Hejazi, M. A. (2019). Probiotic properties of Enterococcus isolated from artisanal dairy products. Front. Microbiol. 10:300. doi: 10.3389/fmicb.2019.00300
Neal-McKinney, J. M., Lu, X., Duong, T., Larson, C. L., Call, D. R., Shah, D. H., et al. (2012). Production of organic acids by probiotic Lactobacilli can be used to reduce pathogen load in poultry. PLoS One 7:e0043928. doi: 10.1371/journal.pone.0043928
Newell, D. G., and Fearnley, C. (2003). Sources of Campylobacter colonization in broiler chickens. Appl. Environ. Microbiol. 69, 4343–4351. doi: 10.1128/AEM.69.8.4343-4351.2003
Nishiyama, K., Nakazato, A., Ueno, S., Seto, Y., Kakuda, T., Takai, S., et al. (2015). Cell surface-associated aggregation-promoting factor from Lactobacillus gasseri SBT2055 facilitates host colonization and competitive exclusion of Campylobacter jejuni. Mol. Microbiol. 98, 712–726. doi: 10.1111/mmi.13153
Nishiyama, K., Seto, Y., Yoshioka, K., Kakuda, T., Takai, S., Yamamoto, Y., et al. (2014). Lactobacillus gasseri SBT2055 reduces infection by and colonization of Campylobacter jejuni. PLoS One 9:e108827. doi: 10.1371/journal.pone.0108827
Nothaft, H., Perez-Muñoz, M. E., Gouveia, G. J., Duar, R. M., Wanford, J. J., Lango-Scholey, L., et al. (2017). Co-administration of the Campylobacter jejuni N-glycan based vaccine with probiotics improves vaccine performance in broiler chickens. Appl. Environ. Microbiol. 83:e01523-17. doi: 10.1128/AEM.01523-17
Nurmi, E., and Rantala, M. (1973). New aspects of Salmonella infection in broiler production. Nature 241:210. doi: 10.1038/241210a0
Oakley, B. B., Buhr, R. J., Ritz, C. W., Kiepper, B. H., Berrang, M. E., Seal, B. S., et al. (2014). Successional changes in the chicken cecal microbiome during 42 days of growth are independent of organic acid feed additives. BMC Vet. Res. 10:282. doi: 10.1186/s12917-014-0282-8
Oakley, B. B., and Kogut, M. H. (2016). Spatial and temporal changes in the broiler chicken cecal and fecal microbiomes and correlations of bacterial taxa with cytokine gene expression. Front. Vet. Sci. 3:11. doi: 10.3389/fvets.2016.00011
Oakley, B. B., Morales, C. A., Line, J., Berrang, M. E., Meinersmann, R. J., Tillman, G. E., et al. (2013). The poultry-associated microbiome: network analysis and farm-to-fork characterization. PLoS One 8:e57190. doi: 10.1371/journal.pone.0057190
Oakley, B. B., Vasconcelos, E. J. R., Diniz, P. P. V. P., Calloway, K. N., Richardson, E., Meinersmann, R. J., et al. (2018). The cecal microbiome of commercial broiler chickens varies significantly by season. Poult. Sci. 97, 3635–3644. doi: 10.3382/ps/pey214
Ocaña, V., Silva, C., and Nader-Macías, M. E. (2006). Antibiotic susceptibility of potentially probiotic vaginal Lactobacilli. Infect. Dis. Obstet. Gynecol. 2006:18182. doi: 10.1155/IDOG/2006/18182
Olnood, C. G., Beski, S. S. M., Choct, M., and Iji, P. A. (2015). Novel probiotics: their effects on growth performance, gut development, microbial community and activity of broiler chickens. Anim. Nutr. 1, 184–191. doi: 10.1016/j.aninu.2015.07.003
On, S. L. W. (2001). Taxonomy of Campylobacter, Arcobacter, Helicobacter and related bacteria: current status, future prospects and immediate concerns. J. Appl. Microbiol. 90, 1S–15S. doi: 10.1046/j.1365-2672.2001.01349.x
Ostad, S. N., Salarian, A. A., Ghahramani, M. H., Fazeli, M. R., Samadi, N., and Jamalifar, H. (2009). Live and heat-inactivated lactobacilli from feces inhibit Salmonella typhi and Escherichia coli adherence to Caco-2 cells. Folia Microbiol. 54, 157–160. doi: 10.1007/s12223-009-0024-7
Pan, D., and Yu, Z. (2014). Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 5, 108–119. doi: 10.4161/gmic.26945
Papadimitriou, K., Zoumpopoulou, G., Foligné, B., Alexandraki, V., Kazou, M., Pot, B., et al. (2015). Discovering probiotic microorganisms: in vitro, in vivo, genetic and omics approaches. Front. Microbiol. 6:58. doi: 10.3389/fmicb.2015.00058
Park, Y. H., Hamidon, F., Rajangan, C., Soh, K. P., Gan, C. Y., Lim, T. S., et al. (2016). Application of probiotics for the production of safe and high-quality poultry meat. Korean J. Food Sci. Anim. Resour. 36, 567–576. doi: 10.5851/kosfa.2016.36.5.567
Peralta-Sánchez, J. M., Martín-Platero, A. M., Ariza-Romero, J. J., Rabelo-Ruiz, M., Zurita-González, M. J., Baños, A., et al. (2019). Egg Production in poultry farming is improved by probiotic bacteria. Front. Microbiol. 10:1042. doi: 10.3389/fmicb.2019.01042
Pielsticker, C., Glünder, G., and Rautenschlein, S. (2012). Colonization properties of Campylobacter jejuni in chickens. Eur. J. Microbiol. Immunol. 2, 61–65. doi: 10.1556/EuJMI.2.2012.1.9
Popova, T. (2017). Effect of probiotics in poultry for improving meat quality. Curr. Opin. Food Sci. 14, 72–77. doi: 10.1016/j.cofs.2017.01.008
Prudêncio, C. V., dos Santos, M. T., and Vanetti, M. C. D. (2015). Strategies for the use of bacteriocins in Gram-negative bacteria: relevance in food microbiology. J. Food Sci. Technol. 52, 5408–5417. doi: 10.1007/s13197-014-1666-2
Qu, K., Guo, F., Liu, X., Lin, Y., and Zou, Q. (2019). Application of machine learning in microbiology. Front. Microbiol. 10:827. doi: 10.3389/fmicb.2019.00827
Rebollar, E. A., Antwis, R. E., Becker, M. H., Belden, L. K., Bletz, M. C., Brucker, R. M., et al. (2016). Using “omics” and integrated multi-omics approaches to guide probiotic selection to mitigate chytridiomycosis and other emerging infectious diseases. Front. Microbiol. 7:68. doi: 10.3389/fmicb.2016.00068
Ren, H., Zentek, J., and Vahjen, W. (2019). Optimization of production parameters for probiotic Lactobacillus strains as feed additive. Molecules 24:3286. doi: 10.3390/molecules24183286
Reniero, R., Cocconcelli, P., Bottazzi, V., and Morelli, L. (1992). High frequency of conjugation in Lactobacillus mediated by an aggregation-promoting factor. Microbiology 138, 763–768. doi: 10.1099/00221287-138-4-763
Ricke, S. C. (2003). Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 82, 632–639. doi: 10.1093/ps/82.4.632
Ricke, S. C., Feye, K. M., Chaney, W. E., Shi, Z., Pavlidis, H., and Yang, Y. (2019). Developments in rapid detection methods for the detection of foodborne Campylobacter in the United States. Front. Microbiol. 9:3280. doi: 10.3389/fmicb.2018.03280
Riley, M. A., and Wertz, J. E. (2002). Bacteriocins: evolution, ecology, and application. Annu. Rev. Microbiol. 56, 117–137. doi: 10.1146/annurev.micro.56.012302.161024
Ringoir, D. D., Szylo, D., and Korolik, V. (2007). Comparison of 2-day-old and 14-day-old chicken colonization models for Campylobacter jejuni. FEMS Immunol. Med. Microbiol. 49, 155–158. doi: 10.1111/j.1574-695X.2006.00181.x
Ritzi, M. M., Abdelrahman, W., Mohnl, M., and Dalloul, R. A. (2014). Effects of probiotics and application methods on performance and response of broiler chickens to an Eimeria challenge. Poultry Sci. 93, 2772–2778. doi: 10.3382/ps.2014-04207
Robyn, J., Rasschaert, G., Hermans, D., Pasmans, F., and Heyndrickx, M. (2013). In vivo broiler experiments to assess anti-Campylobacter jejuni activity of a live Enterococcus faecalis strain. Poult. Sci. 92, 265–271. doi: 10.3382/ps.2012-02712
Robyn, J., Rasschaert, G., Messens, W., Pasmans, F., and Heyndrickx, M. (2012). Screening for lactic acid bacteria capable of inhibiting Campylobacter jejuni in in vitro simulations of the broiler chicken caecal environment. Benef. Microbes 3, 299–308. doi: 10.3920/BM2012.0021
Robyn, J., Rasschaert, G., Pasmans, F., and Heyndrickx, M. (2015). Thermotolerant Campylobacter during broiler rearing: risk factors and intervention. Compr. Rev. Food Sci. Food Saf. 14, 81–105. doi: 10.1111/1541-4337.12124
Rosenberg, M., Gutnick, D., and Rosenberg, E. (1980). Adherence of bacteria to hydrocarbons: a simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 9, 29–33. doi: 10.1111/j.1574-6968.1980.tb05599.x
Rossi, D. A., Fonsec, B. B., de Melo, R. T., Felipe, G. S., da Silva, P. L., Mendonça, E. P., et al. (2012). Transmission of Campylobacter coli in chicken embryos. Braz. J. Microbiol. 43, 535–543. doi: 10.1590/S1517-8382201200002000014
Sadeghi, A. A., Shawrang, P., and Shakorzadeh, S. (2015). Immune response of Salmonella challenged broiler chickens fed diets containing Gallipro®, a Bacillus subtilis probiotic. Probiot. Antimicro. Prot. 7, 24–30. doi: 10.1007/s12602-014-9175-1
Sahin, O., Kassem, I. I., Shen, Z., Lin, J., Rajashekara, G., and Zhang, Q. (2015). Campylobacter in poultry: ecology and potential interventions. Avian Dis. 59, 185–200. doi: 10.1637/11072-032315-Review
Sahin, O., Luo, N., Huang, S., and Zhang, Q. (2003). Effect of Campylobacter-specific maternal antibodies on Campylobacter jejuni colonization in young chickens. Appl. Environ. Microbiol. 69, 5372–5379. doi: 10.1128/AEM.69.9.5372-5379.2003
Saint-Cyr, M. J., Guyard-Nicodème, M., Messaoudi, S., Chemaly, M., Cappelier, J.-M., Dousset, X., et al. (2016). Recent advances in screening of anti-Campylobacter activity in probiotics for use in poultry. Front. Microbiol. 7:553. doi: 10.3389/fmicb.2016.00553
Saint-Cyr, M. J., Haddad, N., Taminiau, B., Poezevara, T., Quesne, S., Amelot, M., et al. (2017). Use of the potential probiotic strain Lactobacillus salivarius SMXD51 to control Campylobacter jejuni in broilers. Int. J. Food Microbiol. 247, 9–17. doi: 10.1016/j.ijfoodmicro.2016.07.003
Sakaridis, I., Ellis, R. J., Cawthraw, S. A., van Vliet, A. H. M., Stekel, D. J., Penell, J., et al. (2018). Investigating the association between the caecal microbiomes of broilers and Campylobacter burden. Front. Microbiol. 9:927. doi: 10.3389/fmicb.2018.00927
Salaheen, S., White, B., Bequette, B. J., and Biswas, D. (2014). Peanut fractions boost the growth of Lactobacillus casei that alters the interactions between Campylobacter jejuni and host epithelial cells. Food Res. Int. 62, 1141–1146. doi: 10.1016/j.foodres.2014.05.061
Santini, C., Baffoni, L., Gaggia, F., Granata, M., Gasbarri, R., Di Gioia, D., et al. (2010). Characterization of probiotic strains: an application as feed additives in poultry against Campylobacter jejuni. Int. J. Food Microbiol. 141, (Suppl. 1), S98–S108. doi: 10.1016/j.ijfoodmicro.2010.03.039
Schillinger, U., and Lücke, F. K. (1989). Antibacterial activity of Lactobacillus sake isolated from meat. Appl. Environ. Microbiol. 55, 1901–1906. doi: 10.1128/aem.55.8.1901-1906.1989
Schneitz, C. (2005). Competitive exclusion in poultry—-30 years of research. Food Control 16, 657–667. doi: 10.1016/j.foodcont.2004.06.002
Schneitz, C., and Hakkinen, M. (2016). The efficacy of a commercial competitive exclusion product on Campylobacter colonization in broiler chickens in a 5-week pilot-scale study. Poultry Sci. 95, 1125–1128. doi: 10.3382/ps/pew020
Schoeni, J. L., and Doyle, M. P. (1992). Reduction of Campylobacter jejuni colonization of chicks by cecum-colonizing bacteria producing anti-C. jejuni metabolites. Appl. Environ. Microbiol. 58, 664–670. doi: 10.1128/aem.58.2.664-670.1992
Schoeni, J. L., and Wong, A. C. (1994). Inhibition of Campylobacter jejuni colonization in chicks by defined competitive exclusion bacteria. Appl. Environ. Microbiol. 60, 1191–1197. doi: 10.1128/aem.60.4.1191-1197.1994
Šefcová, M., Larrea-Álvarez, M., Larrea-Álvarez, C., Karaffová, V., Revajová, V., Gancarèíková, S., et al. (2020). Lactobacillus fermentum administration modulates cytokine expression and lymphocyte subpopulation levels in broiler chickens challenged with Campylobacter coli. Foodborne Pathog. Dis. 17, 485–493. doi: 10.1089/fpd.2019.2739
Shang, Y., Kumar, S., Oakley, B., and Kim, W. K. (2018). Chicken gut microbiota: importance and detection technology. Front. Vet. Sci. 5:254. doi: 10.3389/fvets.2018.00254
Shaughnessy, R. G., Meade, K. G., Cahalane, S., Allan, B., Reiman, C., Callanan, J. J., et al. (2009). Innate immune gene expression differentiates the early avian intestinal response between Salmonella and Campylobacter. Vet. Immunol. Immunop. 132, 191–198. doi: 10.1016/j.vetimm.2009.06.007
Shoaf-Sweeney, K. D., Larson, C. L., Tang, X., and Konkel, M. E. (2008). Identification of Campylobacter jejuni proteins recognized by maternal antibodies of chickens. Appl. Environ. Microbiol. 74, 6867–6875. doi: 10.1128/AEM.01097-08
Shojadoost, B., Kulkarni, R. R., Brisbin, J. T., Quinteiro-Filho, W., Alkie, T. N., and Sharif, S. (2019). Interactions between lactobacilli and chicken macrophages induce antiviral responses against avian influenza virus. Res. Vet. Sci. 125, 441–450. doi: 10.1016/j.rvsc.2017.10.007
Sibanda, N., McKenna, A., Richmond, A., Ricke, S. C., Callaway, T., Stratakos, A. C., et al. (2018). A review of the effect of management practices on Campylobacter prevalence in poultry farms. Front. Microbiol. 9:2002. doi: 10.3389/fmicb.2018.02002
Šikiæ Pogaèar, M., Langerholc, T., Mièetiæ-Turk, D., Možina, S. S., and Klanènik, A. (2020). Effect of Lactobacillus spp. on adhesion, invasion, and translocation of Campylobacter jejuni in chicken and pig small-intestinal epithelial cell lines. BMC Vet. Res. 16:34. doi: 10.1186/s12917-020-2238-5
Silva, J., Leite, D., Fernandes, M., Mena, C., Gibbs, P. A., and Teixeira, P. (2011). Campylobacter spp. as a foodborne pathogen: a review. Front. Microbiol. 2:200. doi: 10.3389/fmicb.2011.00200
Skarp, C. P. A., Hänninen, M.-L., and Rautelin, H. I. K. (2016). Campylobacteriosis: the role of poultry meat. Clin. Microbiol. Infect. 22, 103–109. doi: 10.1016/j.cmi.2015.11.019
Smialek, M., Burchardt, S., and Koncicki, A. (2018). The influence of probiotic supplementation in broiler chickens on population and carcass contamination with Campylobacter spp. - Field study. Res. Vet. Sci. 118, 312–316. doi: 10.1016/j.rvsc.2018.03.009
Smith, D. P., and Berrang, M. E. (2006). Prevalence and numbers of bacteria in broiler crop and gizzard contents. Poultry Sci. 85, 144–147. doi: 10.1093/ps/85.1.144
Sofka, D., Pfeifer, A., Gleiß, B., Paulsen, P., and Hilbert, F. (2015). Changes within the intestinal flora of broilers by colonisation with Campylobacter jejuni. Berl. Munch. Tierarztl. Wochenschr. 128, 104–110. doi: 10.2376/0005-9366-128-104
Song, J., Xiao, K., Ke, Y. L., Jiao, L. F., Hu, C. H., Diao, Q. Y., et al. (2014). Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poultry Sci. 93, 581–588. doi: 10.3382/ps.2013-03455
Stanley, D., Hughes, R. J., and Moore, R. J. (2014). Microbiota of the chicken gastrointestinal tract: influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 98, 4301–4310. doi: 10.1007/s00253-014-5646-2
Stern, D. N. J., Eruslanov, B. V., Pokhilenko, V. D., Kovalev, Y. N., Volodina, L. L., Perelygin, V. V., et al. (2008). Bacteriocins reduce Campylobacter jejuni colonization while bacteria producing bacteriocins are ineffective. Microb. Ecol. Health. Dis. 20, 74–79. doi: 10.1080/08910600802030196
Stern, N. J., Cox, N. A., Bailey, J. S., Berrang, M. E., and Musgrove, M. T. (2001). Comparison of mucosal competitive exclusion and competitive exclusion treatment to reduce Salmonella and Campylobacter spp. colonization in broiler chickens. Poultry Sci. 80, 156–160. doi: 10.1093/ps/80.2.156
Stern, N. J., and Meinersmann, R. J. (1989). Potentials for colonization control of Campylobacter jejuni in the chicken. J. Food Prot. 52, 427–430. doi: 10.4315/0362-028X-52.6.427
Sun, Y., and O’Riordan, M. X. D. (2013). Regulation of bacterial pathogenesis by intestinal short-chain fatty acids. Adv. Appl. Microbiol. 85, 93–118. doi: 10.1016/B978-0-12-407672-3.00003-4
Svetoch, E. A., and Stern, N. J. (2010). Bacteriocins to control Campylobacter spp. in poultry-A review. Poultry Sci. 89, 1763–1768. doi: 10.3382/ps.2010-00659
Tabashsum, Z., Peng, M., Kahan, E., Rahaman, S. O., and Biswas, D. (2019). Effect of conjugated linoleic acid overproducing Lactobacillus with berry pomace phenolic extracts on Campylobacter jejuni pathogenesis. Food Funct. 10, 296–303. doi: 10.1039/c8fo01863d
Tabashsum, Z., Peng, M., Salaheen, S., Comis, C., and Biswas, D. (2018). Competitive elimination and virulence property alteration of Campylobacter jejuni by genetically engineered Lactobacillus casei. Food Control 85, 283–291. doi: 10.1016/j.foodcont.2017.10.010
Taha-Abdelaziz, K., Astill, J., Kulkarni, R. R., Read, L. R., Najarian, A., Farber, J. M., et al. (2019). In vitro assessment of immunomodulatory and anti- Campylobacter activities of probiotic lactobacilli. Sci. Rep. 9, 1–15. doi: 10.1038/s41598-019-54494-3
Taheri, H. R., Moravej, H., Tabandeh, F., Zaghari, M., and Shivazad, M. (2009). Screening of lactic acid bacteria toward their selection as a source of chicken probiotic. Poultry Sci. 88, 1586–1593. doi: 10.3382/ps.2009-00041
Tang, K. L., Caffrey, N. P., Nóbrega, D. B., Cork, S. C., Ronksley, P. E., Barkema, H. W., et al. (2017). Restriction in the Use of Antibiotics in Food Animals and Antibiotic Resistance in Food Animals and Humans – a Systematic Review and Meta-Analysis (University of Calgary, Canada). Geneva: World Health Organization.
Tareb, R., Bernardeau, M., Gueguen, M., and Vernoux, J.-P. (2013). In vitro characterization of aggregation and adhesion properties of viable and heat-killed forms of two probiotic Lactobacillus strains and interaction with foodborne zoonotic bacteria, especially Campylobacter jejuni. J. Med. Microbiol. 62, 637–649. doi: 10.1099/jmm.0.049965-0
Taylor, E. V., Herman, K. M., Ailes, E. C., Fitzgerald, C., Yoder, J. S., Mahon, B. E., et al. (2013). Common source outbreaks of Campylobacter infection in the USA, 1997-2008. Epidemiol. Infect. 141, 987–996. doi: 10.1017/S0950268812001744
Telke, A. A., Ovchinnikov, K. V., Vuoristo, K. S., Mathiesen, G., Thorstensen, T., and Diep, D. B. (2019). Over 2000-fold increased production of the leaderless bacteriocin garvicin KS by increasing gene dose and optimization of culture conditions. Front. Microbiol. 10:389. doi: 10.3389/fmicb.2019.00389
Thibodeau, A., Letellier, A., Yergeau, É, Larrivière-Gauthier, G., and Fravalo, P. (2017). Lack of evidence that selenium-yeast improves chicken health and modulates the caecal microbiota in the context of colonization by Campylobacter jejuni. Front. Microbiol. 8:451. doi: 10.3389/fmicb.2017.00451
Umaraw, P., Prajapati, A., Verma, A. K., Pathak, V., and Singh, V. P. (2017). Control of Campylobacter in poultry industry from farm to poultry processing unit: a review. Crit. Rev. Food Sci. Nutr. 57, 659–665. doi: 10.1080/10408398.2014.935847
Upadhyay, A., Arsi, K., Upadhyaya, I., Donoghue, A. M., and Donoghue, D. J. (2019). “Natural and environmentally friendly strategies for controlling Campylobacter jejuni colonization in poultry, survival in poultry products and infection in humans,” in Food Safety in Poultry Meat Production Food Microbiology and Food Safety, eds K. Venkitanarayanan, S. Thakur, and S. C. Ricke (Cham: Springer), 67–93. doi: 10.1007/978-3-030-05011-5_4
US Food and Drug Administration (2018). Generally Recognized as Safe (GRAS). Available online at: /food/food-ingredients-packaging/generally-recognized-safe-gras (accessed May 11, 2019).
Vaezirad, M. M., Keestra-Gounder, A. M., de Zoete, M. R., Koene, M. G., Wagenaar, J. A., and van Putten, J. P. M. (2017). Invasive behavior of Campylobacter jejuni in immunosuppressed chicken. Virulence 8, 248–260. doi: 10.1080/21505594.2016.1221559
van den Bogert, B., Boekhorst, J., Pirovano, W., and May, A. (2019). On the role of bioinformatics and data science in industrial microbiome applications. Front. Genet. 10:721. doi: 10.3389/fgene.2019.00721
van Gerwe, T., Miflin, J. K., Templeton, J. M., Bouma, A., Wagenaar, J. A., Jacobs-Reitsma, W. F., et al. (2009). Quantifying transmission of Campylobacter jejuni in commercial broiler flocks. Appl. Environ. Microbiol. 75, 625–628. doi: 10.1128/AEM.01912-08
Verraes, C., Van Boxstael, S., Van Meervenne, E., Van Coillie, E., Butaye, P., Catry, B., et al. (2013). Antimicrobial resistance in the food chain: a review. Int. J. Environ. Res. Public. Health 10, 2643–2669. doi: 10.3390/ijerph10072643
Vinderola, C. G., and Reinheimer, J. A. (2003). Lactic acid starter and probiotic bacteria: a comparative “in vitro” study of probiotic characteristics and biological barrier resistance. Food Res. Int. 36, 895–904. doi: 10.1016/S0963-9969(03)00098-X
Wang, C., Zhou, H., Guo, F., Yang, B. S., Su, X., Lin, J., et al. (2019). Oral immunization of chickens with Lactococcus lactis expressing cjaA temporarily reduces Campylobacter jejuni colonization. Foodborne Pathog. Dis. 17, 366–372. doi: 10.1089/fpd.2019.2727
Wang, Y., Sun, J., Zhong, H., Li, N., Xu, H., Zhu, Q., et al. (2017). Effect of probiotics on the meat flavour and gut microbiota of chicken. Sci. Rep. 7, 1–13. doi: 10.1038/s41598-017-06677-z
Wigley, P. (2013). Immunity to bacterial infection in the chicken. Dev. Comp. Immunol. 41, 413–417. doi: 10.1016/j.dci.2013.04.008
Willson, N.-L., Nattrass, G. S., Hughes, R. J., Moore, R. J., Stanley, D., Hynd, P. I., et al. (2018). Correlations between intestinal innate immune genes and cecal microbiota highlight potential for probiotic development for immune modulation in poultry. Appl. Microbiol. Biotechnol. 102, 9317–9329. doi: 10.1007/s00253-018-9281-1
Wise, M. G., and Siragusa, G. R. (2007). Quantitative analysis of the intestinal bacterial community in one- to three-week-old commercially reared broiler chickens fed conventional or antibiotic-free vegetable-based diets. J. Appl. Microbiol. 102, 1138–1149. doi: 10.1111/j.1365-2672.2006.03153.x
Yang, Y., Feye, K. M., Shi, Z., Pavlidis, H. O., Kogut, M., Ashworth, J., et al. (2019). A historical review on antibiotic resistance of foodborne Campylobacter. Front. Microbiol. 10:1509. doi: 10.3389/fmicb.2019.01509
Zhang, G., Ma, L., and Doyle, M. P. (2007). Potential competitive exclusion bacteria from poultry inhibitory to Campylobacter jejuni and Salmonella. J. Food Prot. 70, 867–873. doi: 10.4315/0362-028x-70.4.867
Keywords: probiotics, Campylobacter, poultry, gastrointestinal tract, foodborne pathogens
Citation: Deng W, Dittoe DK, Pavilidis HO, Chaney WE, Yang Y and Ricke SC (2020) Current Perspectives and Potential of Probiotics to Limit Foodborne Campylobacter in Poultry. Front. Microbiol. 11:583429. doi: 10.3389/fmicb.2020.583429
Received: 14 July 2020; Accepted: 03 November 2020;
Published: 22 December 2020.
Edited by:
Michael Konkel, Washington State University, United StatesReviewed by:
Alessandra De Cesare, University of Bologna, ItalyCopyright © 2020 Deng, Dittoe, Pavilidis, Chaney, Yang and Ricke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Steven C. Ricke, c3JpY2tlQHdpc2MuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.