 Viviane C. Oliveira1,2
Viviane C. Oliveira1,2 Felipe L. Bim1
Felipe L. Bim1 Rachel M. Monteiro1Ana Paula Macedo2Emerson S. Santos3Cláudia H. Silva-Lovato2Helena F. O. Paranhos2
Rachel M. Monteiro1Ana Paula Macedo2Emerson S. Santos3Cláudia H. Silva-Lovato2Helena F. O. Paranhos2 Luís D. R. Melo4
Luís D. R. Melo4 Sílvio B. Santos4
Sílvio B. Santos4 Evandro Watanabe1,5*
Evandro Watanabe1,5*- 1Human Exposome and Infectious Diseases Network, School of Nursing of Ribeirão Preto, University of São Paulo, Ribeirão Preto, Brazil
- 2Department of Dental Materials and Prostheses, School of Dentistry of Ribeirão Preto, University of São Paulo, Ribeirão Preto, Brazil
- 3Department of Clinical Toxicological and Bromatologic Analysis, School of Pharmaceutical Sciences of Ribeirão Preto, University of São Paulo, Ribeirão Preto, Brazil
- 4Centre of Biological Engineering, University of Minho, Braga, Portugal
- 5Department of Restorative Dentistry, School of Dentistry of Ribeirão Preto, University of São Paulo, Ribeirão Preto, Brazil
Studies involving antimicrobial-coated endotracheal tubes are scarce, and new approaches to control multidrug-resistant Pseudomonas aeruginosa biofilm on these devices should be investigated. In this study, five new P. aeruginosa bacteriophages from domestic sewage were isolated. All of them belong to the order Caudovirales, Myoviridae family. They are pH and heat stable and produce 27 to 46 particles after a latent period of 30 min at 37°C. Their dsDNA genome (ranging from ∼62 to ∼65 kb) encodes 65 to 89 different putative proteins. They exhibit a broad lytic spectrum and infect 69.7% of the P. aeruginosa strains tested. All the bacteriophages were able to reduce the growth of P. aeruginosa strains in planktonic form. The bacteriophages were also able to reduce the biofilm viability rates and the metabolic activity of P. aeruginosa strains in a model of biofilms associated with endotracheal tubes. In addition, scanning electron microscopy micrographs showed disrupted biofilms and cell debris after treatment of bacteriophages, revealing remarkable biofilm reduction. The lytic activity on multidrug-resistant P. aeruginosa biofilm indicates that the isolated bacteriophages might be considered as good candidates for therapeutic studies and for the application of bacteriophage-encoded products.
Introduction
Mechanical ventilation through an endotracheal or tracheostomy tube is a supportive intervention to critically ill patients and represents a risk factor for the occurrence of ventilator-associated pneumonia (VAP; Haas et al., 2014). VAP is the most common healthcare-associated infection in critically ill patients, involves high financial costs, and has negative prognosis (Barbier et al., 2013; Zimlichman et al., 2013; Russotto et al., 2015). VAP etiologies have been widely discussed, and the factors associated with its development include endotracheal tube biofilm formation which plays an important role as a reservoir for microorganisms (Nseir et al., 2011).
Knowing the potential risk factor of multidrug-resistant Pseudomonas aeruginosa biofilms on endotracheal tubes, interest in the development of antimicrobial-coated endotracheal tubes has emerged. The presence of antimicrobial agents, such as silver particles and chlorhexidine, on the surface of endotracheal tubes demonstrated a decrease in biofilm formation on the device surface’s (Berra et al., 2004; Tokmaji et al., 2015). However, chemical compounds have been implicated in bacterial resistance (Cieplik et al., 2019) and frequently linked to potential cytotoxic hazards (Lessa et al., 2010; Mao et al., 2018). Therefore, new approaches to control multidrug-resistant P. aeruginosa biofilm on endotracheal tubes should be investigated.
The applicability of bacteriophages in medical devices has aroused interest in several areas, including infections related to the urinary tract (Nzakizwanayo et al., 2015; Melo et al., 2016), venous catheters (Lungren et al., 2014), and orthopedic prostheses (Kaur et al., 2014; Barros et al., 2020). Recently, new lytic bacteriophages able to control multidrug-resistant P. aeruginosa planktonic cells and associated-biofilm forms were described (Yuan et al., 2019; Adnan et al., 2020). Although promising, there are few reports on bacteriophage applicability to control biofilms associated with endotracheal tubes (Aleshkin et al., 2016). Thus, we consider it essential to clarify whether newly isolated bacteriophages can be applied in order to control P. aeruginosa biofilms on the surface of endotracheal tubes. The null hypothesis of this study is that there would not be a difference in P. aeruginosa growth when challenged with bacteriophages.
Materials and Methods
Bacteriophage Isolation and Propagation
Bacteriophages were isolated from domestic sewage, and P. aeruginosa (ATCC 27853) was used as the host strain. Isolation, sample enrichment, and purification methods followed the protocol described previously (Sillankorva et al., 2008). Briefly, 20 ml of domestic effluents, collected from March to November 2018 (Domestic Sewage Treatment Station, Ribeirão Preto, Brazil and Araxá, Brazil), was mixed to 20 ml of double-strength tryptic soy broth (TSB; BD Difco, Sparks, MN, United States) and 100 μl of exponentially grown P. aeruginosa. The presence of bacteriophage was confirmed by inhibition haloes on double-layer agar plating [Tryptic Soy Agar Soft (0.8% agar)—TSAS (BD Difco)] (Sambrook and Russell, 2001). Repeated double-layer agar plating was done until single-plaque morphology was observed. For propagation, one isolated plaque was collected through a sterile toothpick and diluted in 25 μl of elution buffer [1 M Tris–HCl, pH 7.5 (Sigma Aldrich, Saint Louis, MO, United States), 10 mM MgSO4 (Sigma Aldrich), 10 mM NaCl (Dinâmica, Indaiatuba, SP, Brazil), 0.002% (w/v) gelatin (Dinâmica)]. The suspension was dropped onto a TSAS medium, with P. aeruginosa lawns, and incubated at 37°C for 24 h. Afterward, 10 ml of elution buffer was added to each plate to elute the bacteriophages, which were purified with chloroform (Sigma Aldrich), titrated, and stored at 4°C.
Transmission Electron Microscopy
One milliliter of bacteriophage suspension was centrifuged at 21,800 g for 60 min at 4°C. The sediment was washed twice in sterile ultrapure water, and 25 μl of each bacteriophage was applied on an electron microscopy grid. The grids were stained with 2% uranyl acetate for 5 min and then analyzed by transmission electron microscopy (JEOL JEM-100 CXII Akishima, Tokyo, Japan; Melo et al., 2014).
Acid–Base and Thermal Stability
In order to assess acid–base stability, 100 μl of the bacteriophage suspension (108 plaque forming units per milliliter—PFU/ml) were added to 900 μl of elution buffer, with pH adjusted to 3.0, 5.0, 7.0, 9.0, and 11.0. The samples were incubated for 2 h at 37°C. To evaluate thermal stability, 100 μl of the bacteriophage suspension (108 PFU/ml) was added to 900 μl of elution buffer and incubated at different temperatures (37, 42, 50, 60, 70, and 80°C) for 2 h (20). After the incubation period, the phage titer was determined through the double-layer agar method (Cui et al., 2017). The assays were carried out at three independent times in triplicate.
One-Step Growth Curve
An exponential-phase P. aeruginosa culture was centrifuged at 4,200 g for 5 min, and OD600 was adjusted to achieve ∼1011 colony-forming units per milliliter (CFU/ml). Subsequently, 5 ml of bacteria suspension was added to 5 ml of bacteriophage suspension at 5 × 108 PFU/ml (multiplicity of infection—MOI: 0.005). The mixture was incubated (120 rpm, 37°C, 5 min) and centrifuged (4,200 g, 5 min), and 10 ml of sterile TSB was added to the pellet. Samples were incubated (120 rpm, 37°C), and aliquots were taken every 5 min until 60 min post-infection. Using the double-layer agar plating, the number of bacteriophage particles was determined at each time. Latent period was determined as the time required for the phage to increase in number and burst size as the ratio between the number of bacteriophages after reaching the plateau and the number of infected bacteria (Sillankorva et al., 2008). The assay was carried out at three independent times.
Genome Sequencing Analysis
One milliliter of bacteriophage stock was incubated with 10 U DNAse I (Sigma Aldrich) and 10 μl RNAse A (20 mg/ml; Sigma Aldrich) at 37°C for 1 h. Enzymes were inactivated at 70°C for 15 min. Then, the suspensions were incubated with extraction buffer [40 μl EDTA 0.5 M (Sigma Aldrich), 5 μl proteinase K 10 mg/ml (Sigma Aldrich), and 50 μl SDS 10% (w/v; Sigma Aldrich)]. Extraction was performed with phenol/chloroform/isoamyl alcohol (25:24:1; Sigma Aldrich). Absence of bacterial contaminant DNA was confirmed by amplification of the 16S rRNA P. aeruginosa gene (Spilker et al., 2004).
Libraries were prepared using the NexteraXT kit (Illumina, San Diego, CA, United States), and genomes were sequenced using Illumina MiSeq paired-end sequencing (2 × 300 bp/coverage 100×), according to the manufacturer’s instructions. Reads were trimmed and assembled into a single contig using Velvet de novo tool (Illumina). Potential coding sequences (CDSs) were initially annotated using myRAST (Aziz et al., 2008), and alternative start codons or non-annotated CDSs were further analyzed using Geneious version 2019.2.1 (Biomatters Ltd., Auckland, New Zealand). Encoded proteins were queried against the National Center for Biotechnology Information (NCBI) non-redundant database with BLASTP (coverage > 80%; E-value ≤ 10–5; Altschul et al., 1990). Protein families, signatures, and structure prediction were searched with PFAM (Finn et al., 2014), InterPro (Finn et al., 2017), and HHpred (Söding, 2005), while the presence of transmembrane domains was checked using TMHMM (Käll and Sonnhammer, 2002), and Phobius (Käll et al., 2004). To predict signal peptide cleavage sites, SignalP (Petersen et al., 2011), and Spoctopus (Viklund et al., 2008) were used. Transfer RNAs (tRNAs) were scanned using tRNAscan-SE (Schattner et al., 2005).
Host Range Investigation and EOP Analysis
Host range was evaluated by the presence of clear or turbid inhibition haloes in a spot test on double-layer agar against 26 clinical isolates and seven multidrug-resistant P. aeruginosa strains (Panel ATCC® MP-23TM; Table 1). The susceptibility profile of clinical isolates was determined by the disk diffusion method. Susceptibility test, antimicrobial classes, antibiotic concentration, and zone diameter breakpoint for P. aeruginosa strains were defined according to the Clinical and Laboratory Standards Institute (Clinical and Laboratory Standards Institute [CLSI], 2019).
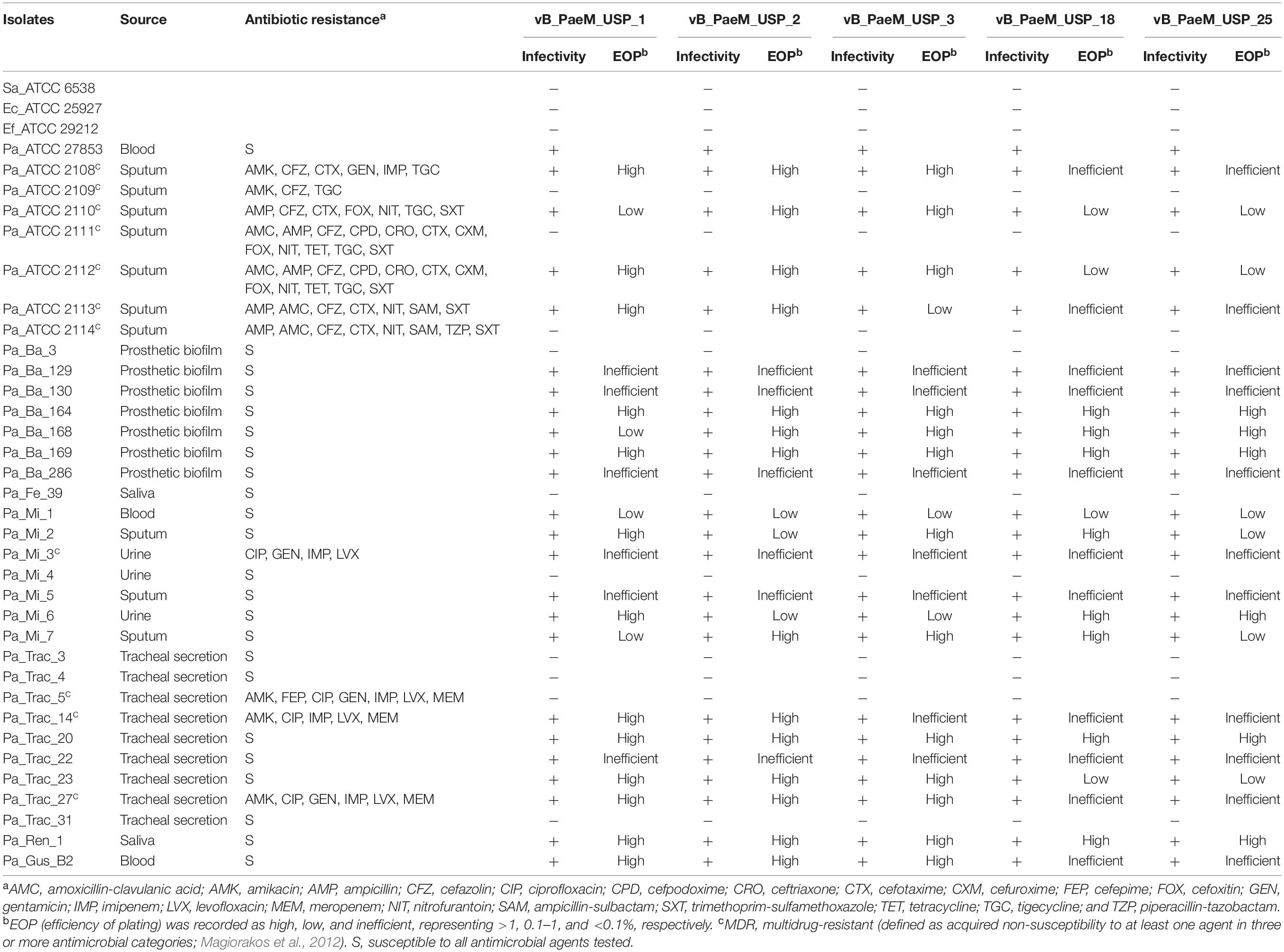
Table 1. Lytic spectrum and efficiency of plating of the bacteriophages against sensible and multidrug-resistant Pseudomonas aeruginosa isolated from different sources.
Efficiency of plating (EOP) was determined for all P. aeruginosa strains positive for bacteriophage lytic activity. Tenfold dilutions of the bacteriophages were spotted on double-layer agar inoculated with P. aeruginosa strains. EOP values were calculated as the ratio between the lysis plaques produced in each susceptible strain and the number of plaques produced in P. aeruginosa ATCC 27853. EOP was recorded as high, low, and inefficient, representing >1, 0.1–1, and <0.1%, respectively (Melo et al., 2014).
Infection of Planktonic Cells
Ten clinical isolates with the highest EOP values, P. aeruginosa ATCC 27853, and multidrug-resistant P. aeruginosa ATCC 2108, ATCC 2110, ATCC 2112, and ATCC 2113 were selected for this assay. Then, 100 μl of P. aeruginosa strains, OD600 adjusted to achieve ∼109 CFU/ml, was inoculated with 100 μl of each bacteriophage at a concentration of 5 × 107 PFU/ml (MOI: 0.05). The positive and negative control experiments were performed using a dilution buffer instead of bacteriophages and sterile TSB instead of bacterial suspension, respectively. The plates were incubated at 37°C, 80 rpm. The OD600 was measured after 2, 4, 6, 8, and 24 h (Melo et al., 2014). The assays were carried out at three independent times in triplicate.
Antibiofilm Activity Measurement
The same 15 P. aeruginosa strains applied in the infection of planktonic cells test were selected for this assay. The anti-biofilm activity of the bacteriophages was determined by means of biofilm viability rates (CFU/cm2), metabolic activity of the bacteria (XTT), and scanning electron microscopy (SEM). The assays were carried out at three independent times in triplicate.
For biofilm infection, strains were grown in TSB medium (37°C, 24 h), and OD600 was adjusted to ∼109 CFU/ml. Circular endotracheal tube specimens (Ø 5 mm; Rüsh, Meridian, MS, United States) were sterilized by hydrogen peroxide plasma (Sterrad 100S, Irvine, CA, United States) and randomly distributed in 48-well tissue culture plates. Each well received 400 μl of TSB medium containing the standardized cell suspension (∼107 CFU/ml). Negative control was performed using sterile TSB. The plates were incubated for 48 h at 37°C and 80 rpm. Afterward, the specimens were removed carefully from the plate, rinsed in 10 ml of phosphate-buffered saline (PBS), and transferred to a new plate with 1 ml of sterile culture medium and 108 PFU/ml of bacteriophages. The control samples received culture medium without bacteriophages. The plates were incubated for 24 h at 37°C and 80 rpm. After the incubation time, the specimens were removed from the wells through sterilized tweezers and rinsed in 10 ml of PBS.
For CFU quantification, the specimens were transferred to a tube containing 2 ml of PBS. In order to detach the remaining biofilm, the tubes were vortexed for 30 s, sonicated (200 W, 40 kHz; Altsonic, Clean 9CA, Ribeirão Preto, SP, Brazil) for 20 min, and vortexed again for 2 min. Afterward, 10-fold-dilution aliquots (100–10–6) were seeded in Petri dishes containing TSA medium. The Petri dishes were incubated at 37°C for 24 h, and the number of colonies was registered and expressed as log CFU/cm2.
For the evaluation of metabolic activity, the specimens were transferred to another 48-well plate containing 316 μl PBS supplemented with 100 mM glucose (Sigma Aldrich), 80 μl XTT 1 mg/ml (Sigma Aldrich), and 4 μl of 0.4 mM menadione (Sigma Aldrich). The plates were statically incubated, protected from light, at 37°C for 2 h, and the absorbance of the resulting solution was measured (492 nm).
For SEM, P. aeruginosa ATCC 27853 was analyzed. The biofilms were removed from the plates and washed twice in distilled water. Samples were fixed with 2.5% glutaraldehyde for 60 min and then dehydrated in a graded ethanol series (30, 50, 70, 90, and 100%). After chemical drying (hexamethyldisilazane; Sigma Aldrich), the specimens were mounted on a metal stub and gold-coated. The surface morphology of the biofilms was examined under high vacuum with a JEOL JSM-35CF microscope (Tokyo, Japan).
Statistical Analysis
Statistical tests were performed through the IBM SPSS Statistics 25.0 software (IBM Corp Armonk, NY, United States). The significance level was set to 0.05. All data showed normal distribution and homogeneous variance and were analyzed using two-way ANOVA, with independent levels and Bonferroni post hoc test.
Results
Bacteriophage Isolation and Propagation
Five new lytic bacteriophages against P. aeruginosa were isolated from domestic sewage and designated as vB_PaeM_USP_1, vB_PaeM_USP_2, vB_PaeM_USP_3, vB_PaeM_USP_18, and vB_PaeM_USP_25. All bacteriophages were able to form clear plaques, having well-defined boundaries. It was observed that none of the bacteriophages produced double haloes, and it was not possible to differentiate lysis plaques among the different isolated bacteriophages (Figures 1A–E).
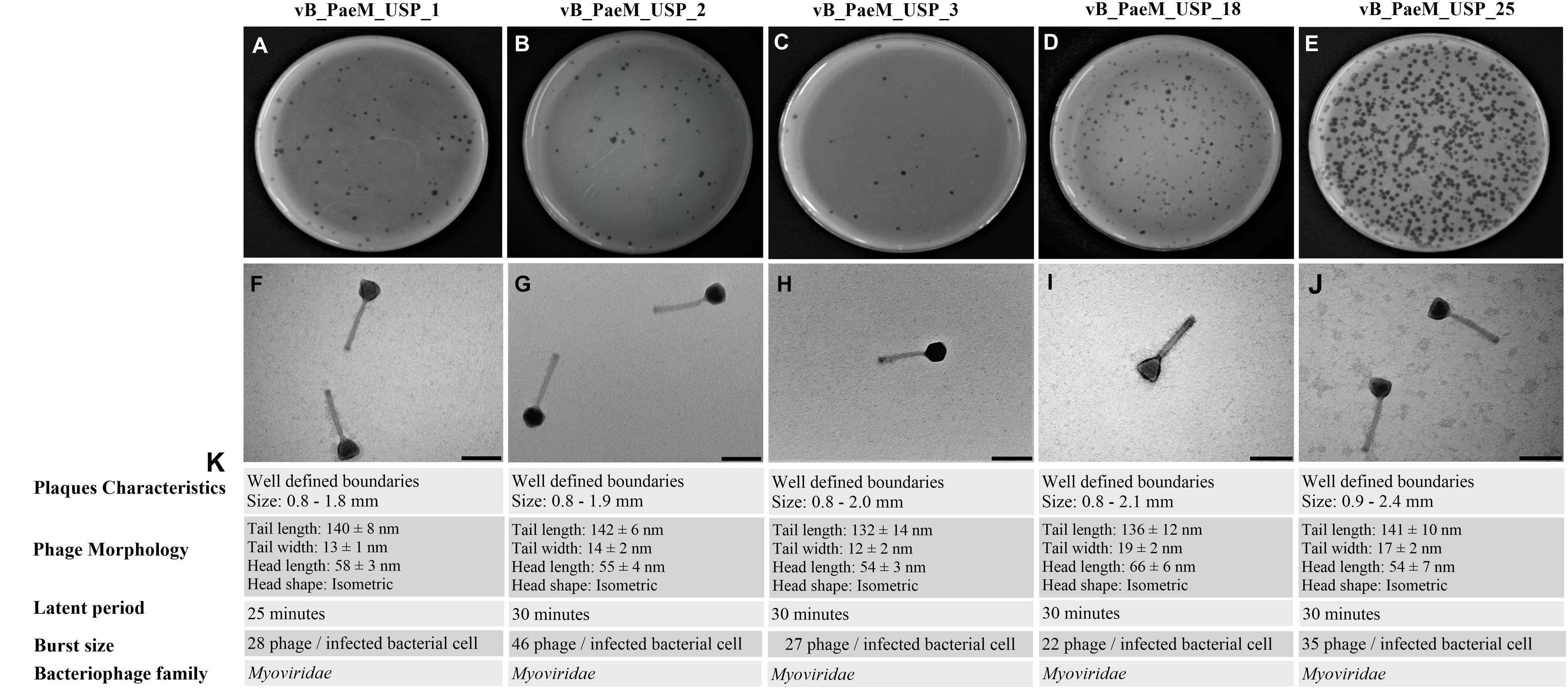
Figure 1. (A–E) Representative image of phage isolates depicting different plaque morphologies of vB_PaeM_USP_1, vB_PaeM_USP_2, vB_PaeM_USP_3, vB_PaeM_USP_18, and vB_PaeM_USP_25. (F–J) Transmission electron micrographs of individual bacteriophages. (K) Panel of plaque characteristics, phage morphology, latent period, and burst size.
Electron Microscopy
The morphological aspects were slightly different among the different isolated viruses (Figures 1F–J). According to the International Committee on Taxonomy of Viruses, all bacteriophages belong to the order Caudovirales and are members of the Myoviridae family.
Acid–Base and Thermal Stability
The values of pH 3.0 and pH 11.0 contributed to reduced bacteriophage viability (p < 0.05). Neutral pH (7.0) and small variations in hydrogenionic potential (pH 5.0 and pH 9.0) did not cause significant changes in viability. vB_PaeM_USP_2 and vB_PaeM_USP_3 were the most resistant to pH variation (Figure 2A). Reduced bacteriophage viability after exposure to 70 and 80°C was noticed. Bacteriophage vB_PaeM_USP_25 was the most resistant to temperature variation (Figure 2B).
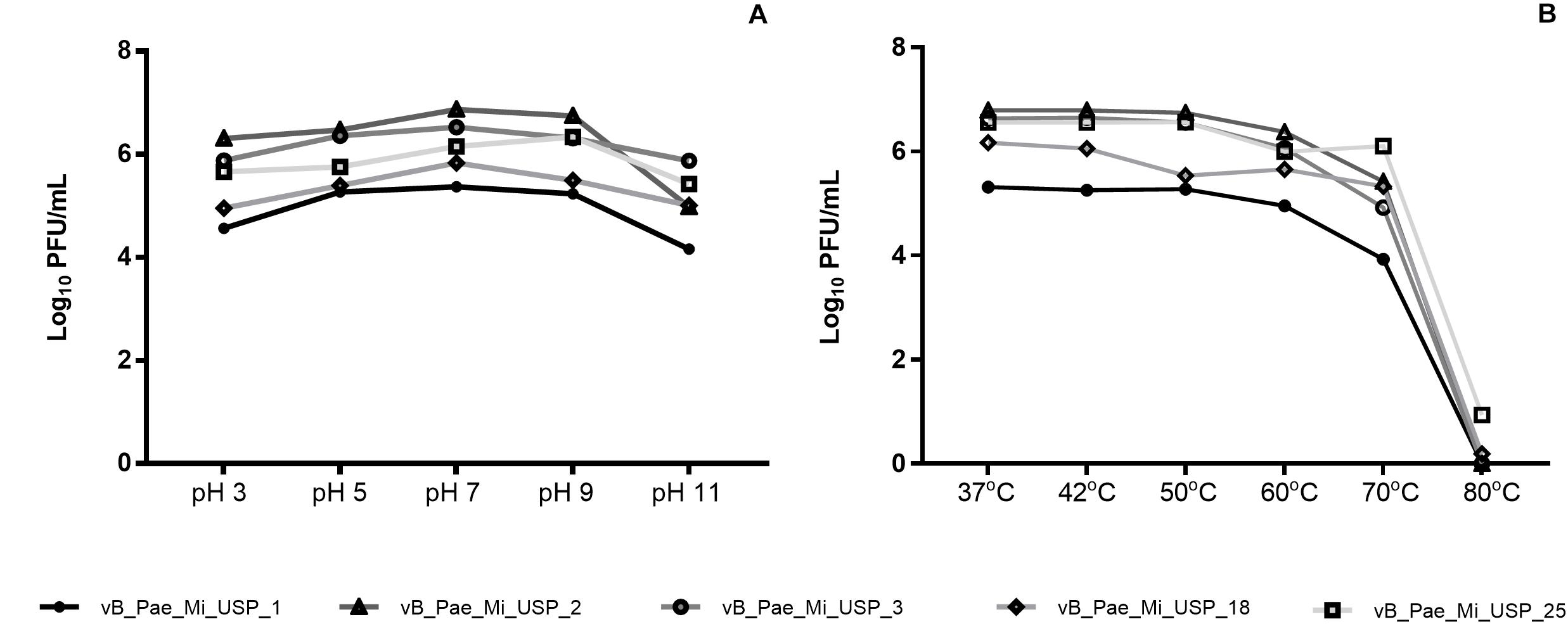
Figure 2. Effects of different pH and temperature values (log10UFP/ml) on vB_PaeM_USP_1, vB_PaeM_USP_2, vB_PaeM_USP_3, vB_PaeM_USP_18, and vB_PaeM_USP_25.
One-Step Growth Curve
A one-step growth curve revealed a similar behavior among the bacteriophages (Supplementary Figure 1). Latent period and yields at burst size are presented in Figure 1K.
Genome Sequencing Analysis
The complete genome sequence of each bacteriophage was deposited in the NCBI database under the accession numbers MT491204–MT491208. Genome analysis revealed that the five bacteriophages are potentially virulent since they do not encode for any known protein associated to lysogeny. Moreover, no known virulence-associated or toxic proteins were identified, revealing that the isolated bacteriophages are potentially safe for therapeutic purposes. All bacteriophage genomes are composed of double-stranded DNA molecules, and no putative genes coding for tRNAs were found. The general genomic properties of the five bacteriophages are presented in Table 2. Two different subgroups of bacteriophages could be observed (Figure 3). The first one, composed of bacteriophages vB_PaeM_USP_1, vB_PaeM_USP_2, and vB_PaeM_USP_3, presents similar genome sizes (approximately 65 kb), G + C content (approximately 55.5%), a high nucleotide pairwise identity (94.6%), and proteome homology (sharing 83 protein homologs between them), with roughly 62% of the proteins with unknown function (Figure 3). Despite their similarity, they present some differences at the beginning (0–5 kb) and at the end of their genome (53 kb end; Figure 4A). Their genomes share high nucleotide identity with other P. aeruginosa bacteriophage members of the Pbunavirus genus, namely, Pseudomonas virus Pa193, Epa22, vB_PaeM_PAO1_Ab27, and BrSP1, with identities above 96% and coverages of up to 98%.
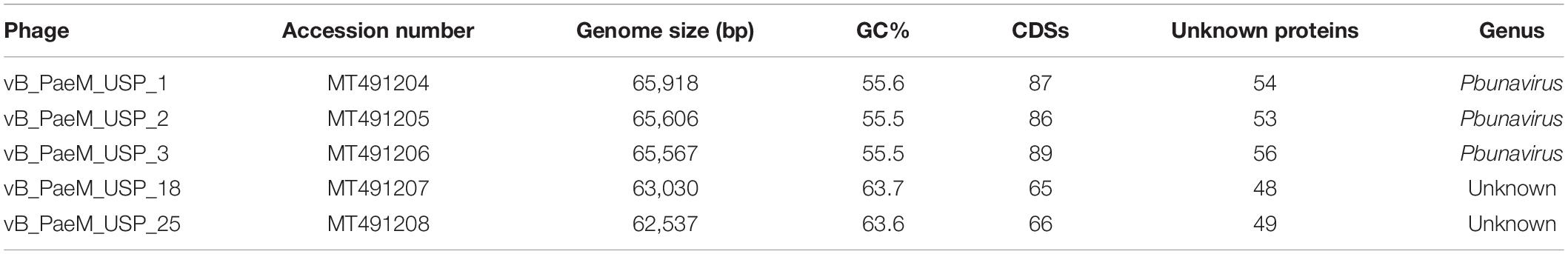
Table 2. Genomic properties of Pseudomonas aeruginosa bacteriophages.
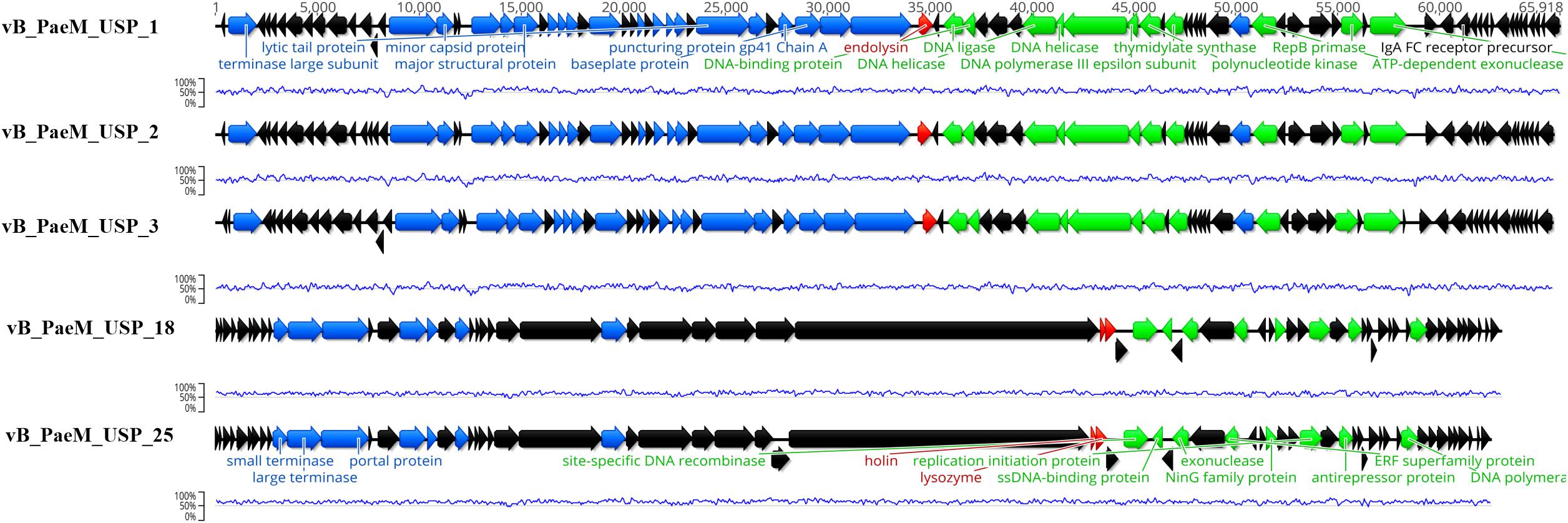
Figure 3. Genome map of the five sequenced bacteriophages depicting the predicted open reading frames colored according to their predicted function (black for hypothetical proteins, green for genes involved in DNA replication and transcription, blue for DNA packaging and morphogenesis genes, and red for cell lysis genes). The blue line represents the guanine-cytosine content (%).
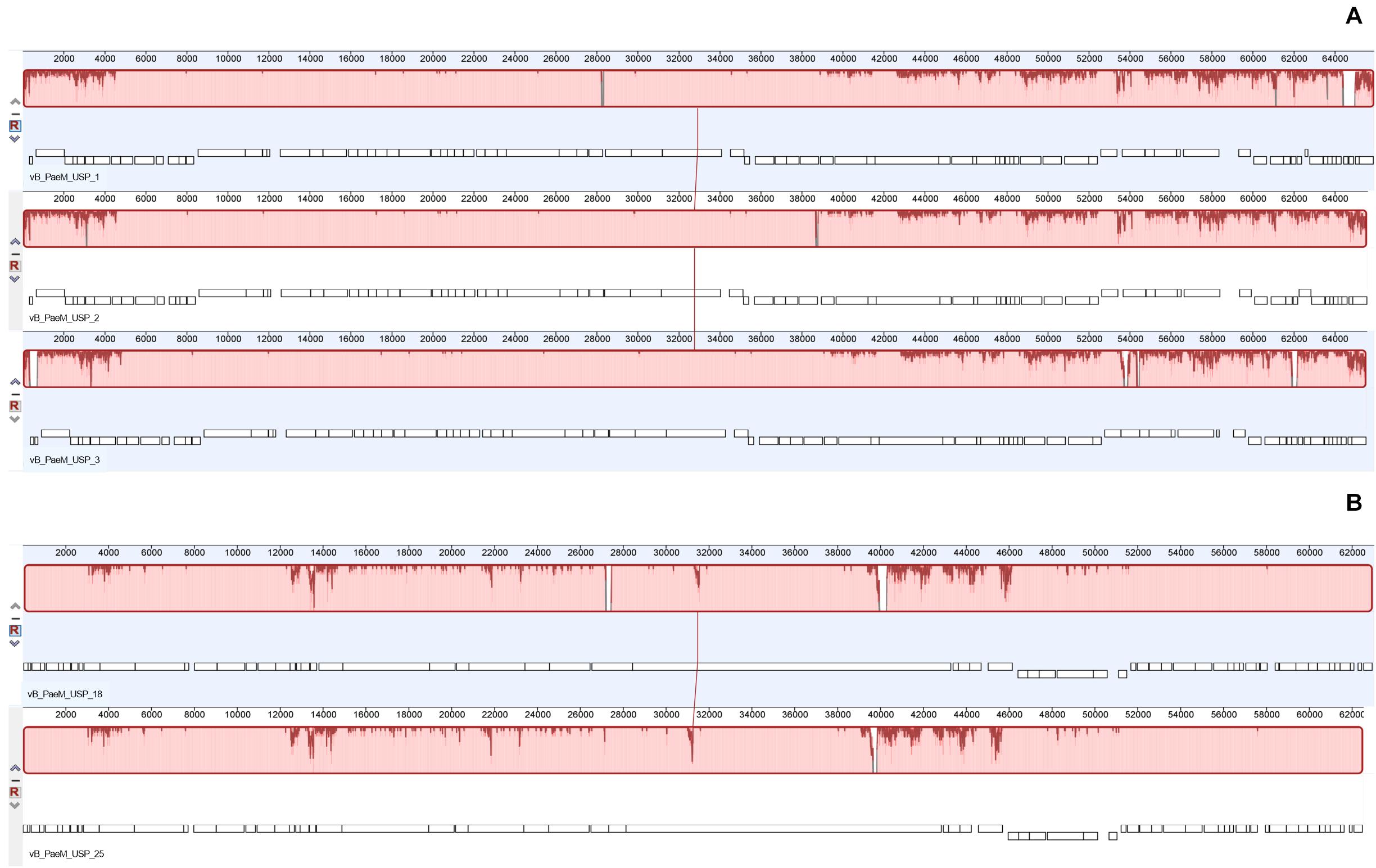
Figure 4. Whole genome alignment (using progressive Mauve algorithm) of the bacteriophage genomes demonstrating the high identity between the two groups. (A) Mauve alignment of the group containing bacteriophages vB_PaeM_USP_1, vB_PaeM_USP_2, and vB_PaeM_USP_3. (B) Mauve alignment of the group containing bacteriophages vB_PaeM_USP_18 and vB_PaeM_USP_25.
The second subgroup, composed of vB_PaeM_USP_18 and vB_PaeM_USP_25, has nucleotide pairwise identity of 97.1% and shares 65 homologs, ca. 74% of them without an assigned function (Figure 3). A slightly lower homology is observed in the genomic regions 3–4.5, 12.5–14, and mainly from 39–46.5 kb (Figure 4B). The low homology with genomes from other phages deposited with the NCBI does not allow grouping these phage genomes with any known genus or subfamily.
Host Range Investigation and EOP Analysis
All five bacteriophages exhibited a broad lytic spectrum and were able to infect 69.7% (n = 23) of the P. aeruginosa strains, of which seven were multidrug resistant. Furthermore, the bacteriophages showed lytic activity on clinical strains isolated from different infection sites (Table 1).
The EOP values of vB_PaeM_USP_1 (high: 13; insufficient: 6; low: 4), vB_PaeM_USP_2 (high: 14; insufficient: 6; low: 3), and vB_PaeM_USP_3 (high: 13; insufficient: 7; low: 3) were higher than vB_PaeM_USP_18 (high: 8; insufficient: 11; low: 4) and vB_PaeM_USP_25 (high: 6; insufficient: 11; low: 6). The last two viruses had low or inefficient EOP for all multidrug-resistant strains (Table 1). Although during host range investigation the bacteriophages had lysed the same strains, inefficient EOP results indicate that the haloes observed previously refer to “lysis from without events.”
Infection of Planktonic Cells
All five bacteriophages reduced bacterial growth up to 8 h of incubation in 13 strains (p < 0.05; Figure 5A). The reduction was above 60% after 6 and 8 h of incubation. After 24 h of incubation, all bacteriophages were able to maintain lower OD values in three strains, but with reductions in bacterial growth of 45% (p < 0.05). In five other strains, the growth rates were reduced by different bacteriophages, and the growth of five strains was not affected by the bacteriophages. Concerning multidrug-resistant P. aeruginosa, the bacteriophages promoted alteration in growth rates in one of the two strains. vB_PaeM_USP_1 and vB_PaeM_USP_3 promoted the greatest growth reductions (Supplementary Tables 1–5).
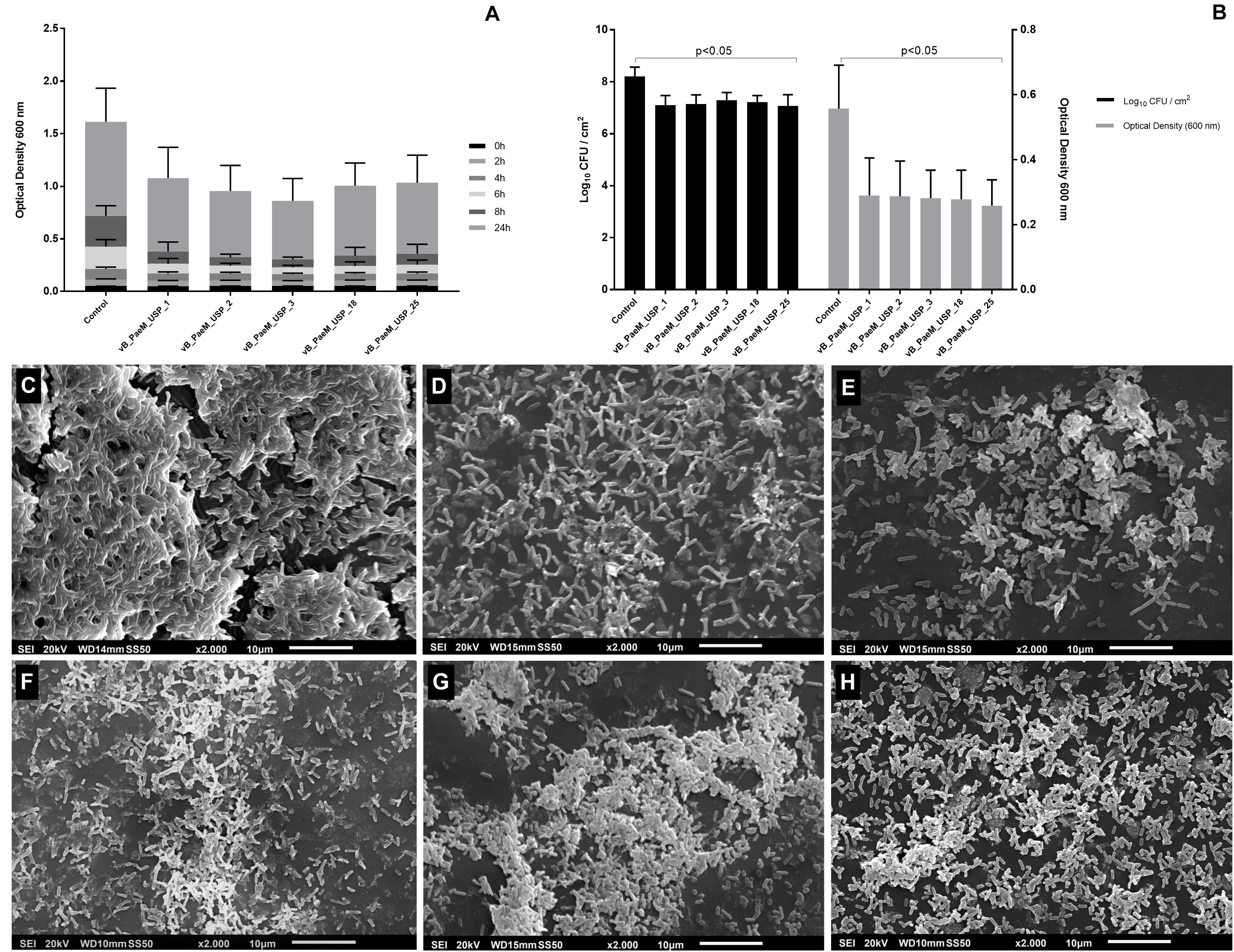
Figure 5. (A) Optical density of planktonic Pseudomonas aeruginosa strains after 2, 4, 6, 8, and 24 h of co-incubation with different bacteriophages. (B) Anti-biofilm activity of different bacteriophages on biofilm viability rates expressed in log10 CFU/cm2 (left y axis) and metabolic activity expressed in absorbance at 492 nm (right y axis). Inhibition of planktonic growth and anti-biofilm activity assays was carried out at three independent times in triplicate against 15 strains of P. aeruginosa. (C–H) Scanning electron micrographs after phage treatment: (C) control, (D) vB_PaeM_USP_1, (E) vB_PaeM_USP_2, (F) vB_PaeM_USP_3, (G) vB_PaeM_USP_18, and (H) vB_PaeM_USP_25. Scale bar = 100 nm. After the treatment of bacteriophages, it is possible to observe lower planktonic growth, biofilm viability rates, and metabolic activity. Disrupted biofilms and cell debris are also noticed in SEM images.
Anti-biofilm Activity Measurement
All five bacteriophages reduced the biofilm viability rates in 12 strains (p < 0.05) in about 10-fold CFU/cm2 (Figure 5B). vB_PaeM_USP_2 and vB_PaeM_USP_18 presented increased anti-biofilm activity against two multidrug-resistant strains.
Concerning the metabolic activity (XTT), after treatment of the bacteriophages, the absorbance values reduced in about 50% in 11 strains (p < 0.05; Figure 5B). Bacteriophage vB_PaeM_USP_2 was able to significantly reduce the metabolic activity. The mean and the standard deviation values (CFU/cm2 and XTT) after exposure to different bacteriophages are shown in the Supplementary Data (Supplementary Tables 6, 7).
Results from SEM micrographs showed densely associated bacteria in the control biofilm (Figure 5C). In contrast, after exposure to bacteriophages, it is possible to observe disrupted biofilms and cell debris, demonstrating remarkable biofilm reduction (Figures 5D–H).
Discussion
This study comprises the isolation and the full characterization of five new P. aeruginosa bacteriophages isolated from domestic sewage. The bacteriophages showed a broad lytic spectrum, able to disrupt P. aeruginosa biofilms on endotracheal tube surfaces. Based on the results, the null hypothesis was rejected since there was a statistical difference on planktonic bacteria growth, viability rates, and metabolic activity of the biofilm.
The vast majority of the already characterized bacteriophages are tailed (Hatfull and Hendrix, 2011). Through morphological and genomics analysis, here there is evidence that all the five bacteriophages have similar features, belonging to the order Caudovirales and Myoviridae family. Moreover, genome sequencing revealed that vB_PaeM_USP_1, vB_PaeM_USP_2, and vB_PaeM_USP_3 were most closely related to Epa22, sharing about 96% of identity. Their genomes are typically organized in clusters. Both guanine-cytosine content and genome length were similar to other Pbunaviruses as previously reported (de Melo et al., 2019; Yuan et al., 2019). Regarding vB_PaeM_USP_18 and vB_PaeM_USP_25, only ca. 20% of their genomes could be aligned with other phage genomes deposited at the NCBI. This low identity with other known genomes does not allow grouping these two phages with any known genus or subfamily. The inability to group these two genomes suggests that they belong to a new bacteriophage genus. Genomic bioinformatics analysis allowed identifying the endolysin gene on all bacteriophages. Commonly, this enzyme is responsible for the lytic ability of tailed bacteriophages (Wu et al., 2019). In addition, several open reading frames could only be annotated as coding for hypothetical proteins and thus with an unknown function, something that is common in bacteriophage studies (Guo et al., 2019).
The one-step growth curve analysis revealed that the latent period was similar in the five bacteriophages (about 30 min; Figure 1H), with a progeny per infected cell varying from 22 to 46 bacteriophage particles. Regarding the latent period, the same result was reported previously. Nonetheless, comparing burst sizes with bacteriophages with a similar genome, we noticed that all bacteriophages produced lower burst sizes (Guo et al., 2019; Yuan et al., 2019). The reduction observed here could be explained by differences in bacterial growth rates since the burst size increases linearly with bacterial growth rate (Nabergoj et al., 2018). Adsorption and latent period also vary with bacterial growth rate, decreasing at higher growth rates.
The infectivity of the bacteriophages remained stable for a pH range from 5.0 to 9.0 and temperature range from 37 to 60°C (Figure 2). Although pH and temperature do not undergo abrupt variation in physiological systems, comprehending this aspect is important in order to maintain bacteriophage viability and infectivity. Even though new antimicrobial agents should be investigated, the large pH and temperature tolerance indicate a broad workability range to search and apply bacteriophage-based products to setups of different infection models. For instance, skin becomes more alkaline by bacterial colonization, and wounds show pH variations in relation to healthy skin (Schneider et al., 2007).
The lytic activity of bacteriophages was tested against 37 strains (33 P. aeruginosa and four different bacterial species). Our results reveal that all bacteriophages could efficiently lyse 69.7% of the P. aeruginosa strains, of which seven were multidrug resistant. Infection of multidrug-resistant P. aeruginosa indicates that bacteriophages or bacteriophage-based products could potentially be applied for therapeutic purposes. The lytic rates presented here are higher than in other Pbunavirus (de Melo et al., 2019), which can be related to the origin and the number of tested strains. In fact, host broad range and species specificity are desirable for bacteriophage application in phage therapy. Moreover, bacteriophages should be strictly lytic in order to avoid the risk of potential transmission of virulent bacterial genes (Canchaya et al., 2003). Here five lytic bacteriophages had promising results in disrupting mature P. aeruginosa biofilms and can be further explored in studies involving phage therapy.
Efficiency of plating assays indicated a considerable variation from low to high between the different bacteriophages on the different strains. Moreover, the bacteriophages showed inefficient EOP for six strains. Differences in EOP and absence of lysis can be explained by the recognition process, which can diverge in different strains according to the expression of bacteriophage receptors on the bacterial outer membrane (Uchiyama et al., 2016). Furthermore, the localization, volume, and density of surface molecules can affect bacteriophage adsorption (Rakhuba et al., 2010). Considering the mentioned variation in molecules for bacteriophage adsorption, the application of a bacteriophage cocktail, which retains distinct adsorption mechanisms, is important to maintain bacteriophage infectivity (Forti et al., 2018). Apart from the recognition process, well-known bacterial resistance systems against bacteriophages could have led to low EOP values. Literature has pointed out that many antiviral mechanisms may be employed by the host cell to evade infection from bacteriophages, such as restriction nucleases and CRISPR (Seed, 2015).
Assessing the efficacy of bacteriophages in reducing P. aeruginosa growth in planktonic form, we observed significant reductions for up to 8 h of co-incubation for almost all investigated strains. However, following 24 h of co-incubation, only three strains showed reduced growth rates. Two multidrug-resistant P. aeruginosa did not show reduced growth rates at the evaluated times. Guo et al. (2019) demonstrated that two new phages at low MOI (0.01) had good performance both on preventing biofilm formation and eradicating preformed biofilms. Moreover, Melo et al. (2014), using a MOI of 0.05, demonstrated reduction in the optical density of planktonic cells in about 83% of clinical isolates after 24 h of infection. We considered that antiviral mechanisms might result in the emergence of bacteriophage-insensitive bacterial strains through blocking of bacteriophage receptors, production of extracellular matrix, production of competitive inhibitors, prevention of bacteriophage DNA entry, slicing of bacteriophage nucleic acids, and abortive infection mechanisms (Labrie et al., 2010).
We observed a great reduction in planktonic growth and anti-biofilm activity after treatment of bacteriophages, and it was widely coincident in both assays. Slight differences were detected among the strains, which can be related to distinct analysis methods. Planktonic assays were performed by OD, while anti-biofilm activity was evaluated by CFU counts and activity metabolic measurement. Comparing OD and CFU results, only one strain showed a discordant outcome. Considering the three analysis methods (OD, CFU, and XTT), the results of six strains did not seem to be concordant. The lytic activity of the bacteriophages was relevant to biofilm control on the endotracheal tube surface. Although no depolymerase was identified in these bacteriophage genomes, the anti-biofilm action could be related to the penetration of viruses at the deeper layers of the biofilm by means of the degradation of the polysaccharide matrix (Lin et al., 2017).
Anti-biofilm results support the hypothesis that bacteriophages might be applied as a biocontrol tool against multidrug-resistant P. aeruginosa on the endotracheal tube surface in the near future. Systemic treatment with antibiotics did not fully inhibit biofilm development on endotracheal tube surfaces (Fernández-Barat et al., 2012). Therefore, direct administration of non-antibiotic approaches, such as antimicrobial agents (Guillon et al., 2018) and bacteriophages, seem to be a suitable option for the fight against persistent biofilms. Indeed bacteriophages could be potentially applied on endotracheal tube biofilm as prophylactic and therapeutic agent. Bacteriophage-coated tubes could be attended to inhibit bacterial adhesion, whereas direct application may be useful in reducing microbial bioburden. Aleshkin et al. (2016) reported the intragastric administration of a phage cocktail in patients on mechanical ventilation, aiming for the treatment of acute infections caused by nosocomial pathogens. The authors demonstrated an important reduction in bacterial counts after phage therapy, indicating it as an alternative purpose to antibiotics to treat healthcare-associated infections.
Literature has presented that bacteriophages or the synergistic action of bacteriophage and antibiotics constitute promising strategies against multidrug-resistant P. aeruginosa biofilm (Akturk et al., 2019; Adnan et al., 2020). Given the ability of new lytic bacteriophages to disrupt and reduce the microbial load of endotracheal-associated biofilms in vitro, several aspects must be taken into consideration before the transfer of phage therapy to clinical practice. As the device is not in contact with vascularized tissues, potential bacteriophage administration routes should be discussed. Moreover, in clinical settings, it is highly unlikely that the endotracheal tubes will continuously be exposed to the phages, as demonstrated in this in vitro study. Thus, we suggest that phage therapy could be applied during endotracheal suctioning, as recommended for ventilated patients. The instillation of bacteriophages during the aspiration of pulmonary secretions could favor its direct application (Guillon et al., 2018). Inhalation of aerosol-containing antibiotics and phage formulations is another potential administration route.
In addition, it should be considered that in vivo biofilms are different from the in vitro ones studied here, probably due to the presence of body fluids, host cells, and different interactions among microorganisms. In light of the aforementioned assumptions, future investigations considering mixed biofilm models and the synergistic action of phage and antibiotic therapy should be conducted. Although treatment of bacteriophage reduced the concentration of P. aeruginosa both on planktonic and biofilm forms, further research is required to determine the correct concentration and the possible combinations of different bacteriophages to establish a cocktail. Based on EOP and anti-biofilm activity and considering the similarities among the five bacteriophages, we suggest that, in further studies, a cocktail composed of vB_PaeM_USP_2 and vB_PaeM_USP_18 should be used. Moreover, only one administration with a single bacteriophage concentration to treat biofilms was evaluated. Assessing different times of infection, a strong lytic activity could be evidenced. In addition, special attention should be paid to the potential development of bacteriophage resistance. In view of this, we also consider it necessary to evaluate the phenotype of bacteriophage-insensitive mutants after a long exposure to bacteriophages.
Conclusion
Biofilms associated with endotracheal tubes are critical concerns in the development of VAP. Since antibiotic therapy is not profitable to eliminate biofilms on the surface of medical devices, bacteriophage therapy offers a possible alternative to reduce bacterial colonization on the surface of endotracheal tubes. The studied bacteriophages were efficient in controlling the growth of P. aeruginosa and promoted biofilm disruption on the surface of endotracheal tubes, including multidrug-resistant strains. Thus, they might be considered as good candidates for therapeutic studies and the application of bacteriophage-encoded proteins.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/ Supplementary Material.
Author Contributions
EW contributed to conceptualization and funding acquisition. VO carried out the main body of research and wrote the original manuscript. FB and RM contributed to the phage growth experiments. AM contributed to data analysis. CS-L and HP supervised the work’s progress and edited the manuscript. LM and SS contributed to bioinformatics analysis and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the São Paulo Research Foundation (FAPESP) under grant numbers 2018/09757-0 and 2019/13271-9 and the National Council for Scientific and Technological Development (CNPq) under grant number 405622/2018-0.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.580779/full#supplementary-material
References
Adnan, M., Shah, M. R. A., Jamal, M., Jalil, F., Andleeb, S., Nawaz, M. A., et al. (2020). Isolation and characterization of bacteriophage to control multidrug-resistant Pseudomonas aeruginosa planktonic cells and biofilm. Biologicals 63, 89–96. doi: 10.1016/j.biologicals.2019.10.003
Akturk, E., Oliveira, H., Santos, S. B., Costa, S., Kuyumcu, S., Melo, L. D. R., et al. (2019). Synergistic action of phage and antibiotics: parameters to enhance the killing efficacy against mono and dual-species biofilms. Antibiotics 8:103. doi: 10.3390/antibiotics8030103
Aleshkin, A. V., Ershova, O. N., Volozhantsev, N. V., Svetoch, E. A., Popova, A. V., Rubalskii, E. O., et al. (2016). Phagebiotics in treatment and prophylaxis of healthcare-associated infections. Bacteriophage 6:e1251379. doi: 10.1080/21597081.2016.1251379
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Aziz, R. K., Bartels, D., Best, A. A., DeJongh, M., Disz, T., Edwards, R. A., et al. (2008). The RAST Server: rapid annotations using subsystems technology. BMC Genom. 9:75. doi: 10.1186/1471-2164-9-75
Barbier, F., Andremont, A., Wolff, M., and Bouadma, L. (2013). Hospital-acquired pneumonia and ventilator-associated pneumonia: recent advances in epidemiology and management. Curr. Opin. Pulm. Med. 19, 216–228. doi: 10.1097/MCP.0b013e32835f27be
Barros, J. A. R., Melo, L. D. R., Silva, R. A. R., Ferraz, M. P., Azeredo, J. C. V. R., Pinheiro, V. M. C., et al. (2020). Encapsulated bacteriophages in alginate-nanohydroxyapatite hydrogel as a novel delivery system to prevent orthopedic implant-associated infections. Nanomedicine 24:102145. doi: 10.1016/j.nano.2019.102145
Berra, L., De Marchi, L., Yu, Z. X., Laquerriere, P., Baccarelli, A., and Kolobow, T. (2004). Endotracheal tubes coated with antiseptics decrease bacterial colonization of the ventilator circuits, lungs, and endotracheal tube. Anesthesiology 100, 1446–1456. doi: 10.1097/00000542-200406000-00017
Canchaya, C., Fournous, G., Chibani-Chennoufi, S., Dillmann, M. L., and Brussow, H. (2003). Phage as agents of lateral gene transfer. Curr. Opin. Microbiol. 6, 417–424. doi: 10.1016/s1369-5274(03)00086-9
Cieplik, F., Jakubovics, N. S., Buchalla, W., Maisch, T., Hellwig, E., and Al-Ahmad, A. (2019). Resistance toward Chlorhexidine in oral bacteria - is there cause for concern? Front. Microbiol. 10:587. doi: 10.3389/fmicb.2019.00587
Clinical and Laboratory Standards Institute [CLSI] (2019). Performance Standards for Antimicrobial Susceptibility Testing, Twenty-Nine Informational Supplement. Wayne: CLSI.
Cui, Z. L., Feng, T. T., Gu, F. F., Li, Q. T., Dong, K., Zhang, Y., et al. (2017). Characterization and complete genome of the virulent Myoviridae phage JD007 active against a variety of Staphylococcus aureus isolates from different hospitals in Shanghai, China. Virol. J. 14:26. doi: 10.1186/s12985-017-0701-0
de Melo, A. C. C., Gomes, A. D., Melo, F. L., Ardisson-Araujo, D. M. P., de Vargas, A. P. C., Ely, V. L., et al. (2019). Characterization of a bacteriophage with broad host range against strains of Pseudomonas aeruginosa isolated from domestic animals. BMC Microbiol. 19:134. doi: 10.1186/s12866-019-1481-z
Fernández-Barat, L., Ferrer, M., Sierra, J. M., Soy, D., Guerrero, L., Vila, J., et al. (2012). Linezolid limits burden of methicillin-resistant Staphylococcus aureus in biofilm of tracheal tubes. Crit. Care Med. 40, 2385–2389. doi: 10.1097/CCM.0b013e31825332fc
Finn, R. D., Attwood, T. K., Babbitt, P. C., Bateman, A., Bork, P., Bridge, A. J., et al. (2017). InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 45, 190–199. doi: 10.1093/nar/gkw1107
Finn, R. D., Bateman, A., Clements, J., Coggill, P., Eberhardt, R. Y., Eddy, S. R., et al. (2014). Pfam: the protein families database. Nucleic Acids Res. 42, 222–230. doi: 10.1093/nar/gkt1223
Forti, F., Roach, D. R., Cafora, M., Pasini, M. E., Horner, D. S., Fiscarelli, E. V., et al. (2018). Design of a broad-range bacteriophage cocktail that reduces Pseudomonas aeruginosa biofilms and treats acute infections in two animal models. Antimicrob. Agents Chemother. 62:e02573-17. doi: 10.1128/AAC.02573-17
Guillon, A., Fouquenet, D., Morello, E., Henry, C., Georgeault, S., Si-Tahar, M., et al. (2018). Treatment of Pseudomonas aeruginosa biofilm present in endotracheal tubes by Poly-l-Lysine. Antimicrob. Agents Chemother. 62:e0564-18.
Guo, Y. Y. J., Chen, P., Lin, Z. L., and Wang, T. T. (2019). Characterization of two Pseudomonas aeruginosa viruses vB_PaeM_SCUT-S1 and vB_PaeM_SCUT-S2. Viruses 11:e318. doi: 10.3390/v11040318
Haas, C. F., Eakin, R. M., Konkle, M. A., and Blank, R. (2014). Endotracheal tubes: old and new. Respir. Care 59, 933–952. doi: 10.4187/respcare.02868
Hatfull, G. F., and Hendrix, R. W. (2011). Bacteriophages and their genomes. Curr. Opin. Virol. 1, 298–303. doi: 10.1016/j.coviro.2011.06.009
Käll, L., Krogh, A., and Sonnhammer, E. L. L. (2004). A combined transmembrane topology and signal peptide prediction method. J. Mol. Biol. 338, 1027–1036. doi: 10.1016/j.jmb.2004.03.016
Käll, L., and Sonnhammer, E. L. L. (2002). Reliability of transmembrane predictions in whole-genome data. FEBS Lett. 532, 415–418. doi: 10.1016/S0014-5793(02)03730-4
Kaur, S., Harjai, K., and Chhibber, S. (2014). Bacteriophage mediated killing of Staphylococcus aureus in vitro on orthopaedic K wires in presence of linezolid prevents implant colonization. PLoS One 9:e90411. doi: 10.1371/journal.pone.0090411
Labrie, S. J., Samson, J. E., and Moineau, S. (2010). Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 8, 317–327. doi: 10.1038/nrmicro2315
Lessa, F. C. R., Aranha, A. M. F., Nogueira, I., Giro, E. M. A., Hebling, J., and Costa, C. A. S. (2010). Toxicity of Chlorhexidine on odontoblast-like cells. J. Appl. Oral Sci. 18, 50–58. doi: 10.1590/s1678-77572010000100010
Lin, D. M., Koskella, B., and Lin, H. C. (2017). Phage therapy: an alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 8:162. doi: 10.4292/wjgpt.v8.i3.162
Lungren, M. P., Donlan, R. M., Kankotia, R., Paxton, B. E., Falk, I., Christensen, D., et al. (2014). Bacteriophage K antimicrobial-lock technique for treatment of Staphylococcus aureus central venous catheter-related infection: a leporine model efficacy analysis. J. Vasc. Interv. Radiol. 25, 1627–1632. doi: 10.1016/j.jvir.2014.06.009
Magiorakos, A. P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., et al. (2012). Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. doi: 10.1111/j.1469-0691.2011.03570.x
Mao, B. H., Chen, Z. Y., Wang, Y. J., and Yan, S. J. (2018). Silver nanoparticles have lethal and sublethal adverse effects on development and longevity by inducing ROS-mediated stress responses. Sci. Rep. 8:2445. doi: 10.1038/s41598-018-20728-z
Melo, L. D. R., Sillankorva, S., Ackermann, H. W., Kropinski, A. M., Azeredo, J., and Cerca, N. (2014). Isolation and characterization of a new Staphylococcus epidermidis broad-spectrum bacteriophage. J. Gen. Virol. 95, 506–515. doi: 10.1099/vir.0.060590-0
Melo, L. D. R., Veiga, P., Cerca, N., Kropinski, A. M., Almeida, C., Azeredo, J., et al. (2016). Development of a phage cocktail to control Proteus mirabilis catheter-associated urinary tract infections. Front. Microbiol. 7:1024. doi: 10.3389/fmicb.2016.01024
Nabergoj, D., Modic, P., and Podgornik, A. (2018). Effect of bacterial growth rate on bacteriophage population growth rate. Microbiologyopen 7:e00558. doi: 10.1002/mbo3.558
Nseir, S., Ader, F., Lubret, R., and Marquette, C. H. (2011). Pathophysiology of airway colonization in critically ill COPD patient. Curr. Drug Targets 12, 514–520. doi: 10.2174/138945011794751537
Nzakizwanayo, J., Hanin, A., Alves, D. R., McCutcheon, B., Dedi, C., Salvage, J., et al. (2015). Bacteriophage can prevent encrustation and blockage of urinary catheters by Proteus mirabilis. Antimicrob. Agents Chemother. 60, 1530–1536. doi: 10.1128/AAC.02685-15
Petersen, T. N., Brunak, S., Heijne, G. V., and Nielsen, H. (2011). SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat. Methods 8, 785–786. doi: 10.1038/nmeth.1701
Rakhuba, D. V., Kolomiets, E. I., Dey, E. S., and Novik, G. I. (2010). Bacteriophage receptors, mechanisms of phage adsorption and penetration into host cell. Pol. J. Microbiol. 59, 145–155. doi: 10.33073/pjm-2010-023
Russotto, V., Cortegiani, A., Raineri, S. M., and Giarratano, A. (2015). Bacterial contamination of inanimate surfaces and equipment in the intensive care unit. J. Intensive Care 3:54. doi: 10.1186/s40560-015-0120-125
Sambrook, J., and Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual. New York, NY: Cold Spring Harbor Laboratory Press.
Schattner, P., Brooks, A. N., and Lowe, T. M. (2005). The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 33, W686–W689. doi: 10.1093/nar/gki366
Schneider, L. A., Korber, A., Grabbe, S., and Dissemond, J. (2007). Influence of pH on wound-healing: a new perspective for wound-therapy? Arch. Dermatol. Res. 298, 413–420. doi: 10.1007/s00403-006-0713-x
Seed, K. D. (2015). Battling phages: how bacteria defend against viral attack. PLoS Pathog. 11:e1004847. doi: 10.1371/journal.ppat.1004847
Sillankorva, S., Neubauer, P., and Azeredo, J. (2008). Isolation and characterization of a T7-like lytic phage for Pseudomonas fluorescens. BMC Biotechnol. 8:80. doi: 10.1186/1472-6750-8-80
Söding, J. (2005). Protein homology detection by HMM-HMM comparison. Bioinformatics 21, 951–960. doi: 10.1093/bioinformatics/bti125
Spilker, T., Coenye, T., Vandamme, P., and LiPuma, J. J. (2004). PCR-Based assay for differentiation of Pseudomonas aeruginosa from other Pseudomonas species recovered from cystic fibrosis patients. J. Clin. Microbiol. 42, 2074–2079. doi: 10.1128/jcm.42.5.2074-2079.2004
Tokmaji, G., Vermeulen, H., Muller, M. C. A., Kwakman, P. H. S., Schultz, M. J., and Zaat, S. A. J. (2015). Silver-coated endotracheal tubes for prevention of ventilator-associated pneumonia in critically ill patients. Cochrane Database Syst. Rev 8:CD009201. doi: 10.1002/14651858.CD009201
Uchiyama, J., Suzuki, M., Nishifuji, K., Kato, S. I., Miyata, R., Nasukawa, T., et al. (2016). Analyses of short-term antagonistic evolution of Pseudomonas aeruginosa strain PAO1 and phage KPP22 (Myoviridae Family, PB1-like virus Genus). Appl. Environ. Microbiol. 82, 4482–4491. doi: 10.1128/AEM.00090-16
Viklund, H., Bernsel, A., Skwark, M., and Elofsson, A. (2008). Spoctopus: a combined predictor of signal peptides and membrane protein topology. Bioinformatics 24, 2928–2929. doi: 10.1093/bioinformatics/btn550
Wu, M. L., Hu, K. Y., Xie, Y. H., Liu, Y. L., Mu, D., Guo, H. M., et al. (2019). A novel phage PD-6A3, and its endolysin Ply6A3, with extended lytic activity against Acinetobacter baumannii. Front. Microbiol. 9:3302. doi: 10.3389/fmicb.2018.03302
Yuan, Y. Y., Qu, K. L., Tan, D. M., Li, X. Y., Wan, L. L., Cong, C., et al. (2019). Isolation and characterization of a bacteriophage and its potential to disrupt multi-drug resistant Pseudomonas aeruginosa biofilms. Microb. Pathog. 128, 329–336. doi: 10.1016/j.micpath.2019.01.032
Keywords: bacteriophage, biofilm, Pseudomonas aeruginosa, multidrug resistance, phage therapy
Citation: Oliveira VC, Bim FL, Monteiro RM, Macedo AP, Santos ES, Silva-Lovato CH, Paranhos HFO, Melo LDR, Santos SB and Watanabe E (2020) Identification and Characterization of New Bacteriophages to Control Multidrug-Resistant Pseudomonas aeruginosa Biofilm on Endotracheal Tubes. Front. Microbiol. 11:580779. doi: 10.3389/fmicb.2020.580779
Received: 24 July 2020; Accepted: 08 September 2020;
Published: 06 October 2020.
Edited by:
Jian Li, Monash University, AustraliaReviewed by:
Oana Ciofu, University of Copenhagen, DenmarkYoon Kyung (Rachel) Chang, The University of Sydney, Australia
Copyright © 2020 Oliveira, Bim, Monteiro, Macedo, Santos, Silva-Lovato, Paranhos, Melo, Santos and Watanabe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Evandro Watanabe, ZXZhbmRyb3dhdGFuYWJlQGdtYWlsLmNvbQ==