
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 24 June 2020
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 11 - 2020 | https://doi.org/10.3389/fmicb.2020.01172
This article is part of the Research TopicExploiting Novel Combined Host- and Pathogen-Directed Therapies for Combating Bacterial Multidrug ResistanceView all 12 articles
 Sirijan Santajit1
Sirijan Santajit1 Watee Seesuay2Kodchakorn Mahasongkram2
Watee Seesuay2Kodchakorn Mahasongkram2 Nitat Sookrung2,3Pornpan Pumirat1
Nitat Sookrung2,3Pornpan Pumirat1 Sumate Ampawong4Onrapak Reamtong5Manas Chongsa-Nguan6
Sumate Ampawong4Onrapak Reamtong5Manas Chongsa-Nguan6 Wanpen Chaicumpa2
Wanpen Chaicumpa2 Nitaya Indrawattana1*
Nitaya Indrawattana1*The quorum sensing (QS) signaling molecule, N-(3-oxododecanoyl)-L-homoserine lactone (3O-C12-HSL), contributes to the pathogenesis of Pseudomonas aeruginosa by regulating expression of the bacterial virulence factors that cause intense inflammation and toxicity in the infected host. As such, the QS molecule is an attractive therapeutic target for direct-acting inhibitors. Several substances, both synthetic and naturally derived products, have shown effectiveness against detrimental 3O-C12-HSL activity. Unfortunately, these compounds are relatively toxic to mammalian cells, which limits their clinical application. In this study, fully human single-chain variable fragments (HuscFvs) that bind to P. aeruginosa haptenic 3O-C12-HSL were generated based on the principle of antibody polyspecificity and molecular mimicry of antigenic molecules. The HuscFvs neutralized 3O-C12-HSL activity and prevented mammalian cells from the HSL-mediated apoptosis, as observed by Annexin V/PI staining assay, sub-G1 arrest population investigation, transmission electron microscopy for ultrastructural morphology of mitochondria, and confocal microscopy for nuclear condensation and DNA fragmentation. Computerized homology modeling and intermolecular docking predicted that the effective HuscFvs interacted with several regions of the bacterially derived ligand that possibly conferred neutralizing activity. The effective HuscFvs should be tested further in vitro on P. aeruginosa phenotypes as well as in vivo as a sole or adjunctive therapeutic agent against P. aeruginosa infections, especially in antibiotic-resistant cases.
Pseudomonas aeruginosa, a versatile and ubiquitous Gram-negative bacterium, is an opportunistic microorganism that frequently causes severe nosocomial infections, particularly among immunocompromised patients and those suffering from cystic fibrosis, burns, HIV infection, and cancer (Tang et al., 1996; Sadikot et al., 2005; Silva Filho et al., 2013; Malhotra et al., 2019; Waters and Goldberg, 2019). The pathogenicity of P. aeruginosa is attributable mainly, if not solely, to the regulons of two complete N-acyl homoserine lactone (AHL)-dependent quorum sensing (QS) systems, called LasI/R and RhlI/R (Preston et al., 1997; Venturi, 2006). The two QS systems act in a hierarchical manner, i.e., the lasI/R system controls the activity of the rhlI/R circuit (Pearson et al., 1994, 1995). During bacterial infection, the LasI and RhlI synthases produce N-(3-oxododecanoyl)-L-homoserine lactone (3O-C12-HSL) and N-butanoyl-L-homoserine lactone (C4-HSL), respectively. The QS molecules then interact with their cognate LasR and RhlR, causing transcription of hundreds of target genes, including those coding for virulence factors such as lectins, elastases, proteases, exotoxin A, pyocyanin, and surface-active rhamnolipids important in the late stages of biofilm development, as well as genes involved in antibiotic resistance (Wagner et al., 2003; Venturi, 2006; Rutherford and Bassler, 2012; Moradali et al., 2017).
N-(3-Oxododecanoyl)-L-homoserine lactone (3O-C12-HSL) is the prominent autoinducer of the P. aeruginosa QS system (Duan and Surette, 2007; Rasamiravaka and El Jaziri, 2016). 3O-C12-HSL is a small, fatty acid-like, membrane-permeant signaling molecule that comprises a hydrophilic homoserine lactone ring linked to the hydrophobic 12-carbon-atom-long acyl side chain via an amide bond (Eberhard et al., 1981; Pearson et al., 1995; Ritchie et al., 2007; O’Connor et al., 2015). The roles of 3O-C12-HSL in pathogenesis and modulation of the host immune responses have been reviewed (Liu et al., 2015). Owing to its lipophilicity, the 3O-C12-HSL can traverse the mammalian cell membrane (Ritchie et al., 2007), causing mitochondrial damage and dysfunction, which subsequently activates the caspase pathway leading to apoptosis of several cell types, including macrophages, neutrophils, T lymphocytes, human vascular endothelial cells, murine fibroblasts, airway epithelial cells, goblet cells, and breast carcinoma cells (Tateda et al., 2003; Li et al., 2004; Shiner et al., 2006; Jacobi et al., 2009; Schwarzer et al., 2012; Tao et al., 2016, 2018). P. aeruginosa QS signaling molecules also modulate host immune responses by down-regulating the expression of co-stimulatory molecules on dendritic cells (DCs), leading to inhibition of DC maturation and their ability to activate effector T-cell responses (Boontham et al., 2008). Because the 3O-C12-HSL plays an important role in the virulence and pathogenesis of P. aeruginosa and host immunity suppression, it is an attractive target for novel therapeutics for P. aeruginosa infection. Substances that interfere with P. aeruginosa 3O-C12-HSL activity should mitigate bacterial-associated disease severity, although blocking the QS system alone does not necessarily abrogate all P. aeruginosa virulence factors, such as T3SS (Bleves et al., 2005; López-Jácome et al., 2019; Soto-Aceves et al., 2019). A therapeutic approach based on QS interference and/or attenuation of QS signals should result in greater sensitivity of the P. aeruginosa to stresses, such as antimicrobial drugs (Rasmussen and Givskov, 2006; Defoirdt et al., 2010; Maeda et al., 2012; Kalia et al., 2014; Krzyżek, 2019).
Recently, a murine monoclonal antibody (mAb), RS2-1G9, against a lactam mimetic of 3O-C12-HSL has been shown to prevent apoptosis through p38 mitogen-activated protein kinase activation and protected murine bone marrow-derived macrophages from the cytotoxic effects of the QS molecule (Kaufmann et al., 2006, 2008). The RS2-1G9 paratope was shown to enclose the polar lactam moiety of the 3O-C12-HSL molecule in the co-crystal structure of the Fab fragment of the RS2-1G9 mAb and the target 3O-C12-HSL completely (Debler et al., 2007). Active immunization of mice with 3O-C12-HSL-protein conjugate protected immunized mice from lethal P. aeruginosa infection (Miyairi et al., 2006). Antibody-based therapy directed to the QS molecule should not only block bacterial virulence, but also rescue the host immunity that had been modulated/suppressed by the QS system (Kaufmann et al., 2008; Palliyil and Broadbent, 2009). The present study generated engineered, fully human, single-chain antibody variable fragments (HuscFvs) that neutralize 3O-C12-HSL bioactivity. The HuscFvs should be tested, step-by-step, toward clinical application as a sole or adjunct therapy for the currently failing antibiotic treatment of patients with P. aeruginosa infection.
The QS molecule was synthesized commercially (Cayman Chemical, Ann Arbor, MI, United States) under the IUPAC name: 3-oxo-N-[(3S)-2-oxooxolan-3-yl]-dodecanamide. 3O-C12-HSL was stored in 100% dimethyl-sulfoxide (DMSO) and diluted with phosphate-buffered saline, pH 7.4 (PBS), to the desired concentration for use.
The human single-chain variable fragments (HuscFvs) to the 3O-C12-HSL were generated based on the principles of the polyspecific property of an antibody, i.e., one antibody can bind different antigens by paratope adaptation to accommodate distinct antigens, such as through differential engagements of the complementarity determining regions (CDRs), and the molecular mimicry of the antigens (different antigens can share surface topologies in terms of shape or chemical nature) (Tapryal et al., 2013). In this study, HB2151 Escherichia coli clones carrying phagemids with inserted HuscFv genes (huscfvs) were previously selected from a HuscFv phage display library (Kulkeaw et al., 2009) using Pseudomonas exotoxin A (ETA) as antigen in the phage-biopanning process (Santajit et al., 2019). Genes coding for HuscFvs of individual E. coli clones were sequenced and deduced, and the canonical CDRs and framework regions (FRs) of both VH and VL domains were determined based on the numbering scheme of Chotia and Kobat (Abhinandan and Martin, 2008).
Three dimensional (3D) models of the selected HuscFvs were generated by subjecting their deduced amino acid sequences to the I-TASSER online server (Yang et al., 2015). The HuscFvs-3D models from the I-TASSER were further refined to improve local geometric and physical quality using ModRefiner (Xu and Zhang, 2011). The quality of the generated homology models of HuscFvs was then evaluated using the PROCHECK server to provide Ramachandran plots (Laskowski et al., 1993). Thereafter, the 3D structures of the individual HuscFvs were superimposed with the 3D structure of the mAb RS2-1G9 F(ab′)2 (PDB ID: 2NTF) (previously shown to neutralize 3O-C12-HSL pathogenic activity; hence, the mAb has been designated as a “quorum quenching antibody”) (Kaufmann et al., 2006, 2008) using the CLICK server, i.e., the topology-independent tool comparing 3D structures without a scoring function measuring structural similarity (Nguyen et al., 2011). The HuscFvs showing top-scored topological similarity with the RS2-1G9 antigen-binding site were selected. The 3D structure of 3O-C12-HSL was retrieved from the PubChem database, a resource of chemical molecules and their bioactivities (PubChem CID: 3246941) (Kim et al., 2015). The modeled-3O-C12-HSL F(ab′)2 was docked with the 3D model of each HuscFv receptor binding pocket using Autodock Vina software (Trott and Olson, 2010; Forli et al., 2016). The conformation of each HuscFv-ligand complex with the lowest binding free energy (ΔG) at the best docking position was selected for interaction analysis and visualization through the Discovery studio visualizer 3.5 program.
The E. coli clones carrying phagemids containing the DNA coding for the selected HuscFvs were subjected to sub-cloning for large scale HuscFv production. The huscfvs were PCR-amplified from the huscfv-pCANTAB5E phagemids of HB2151 E. coli clones using a Phusion High-Fidelity DNA polymerase (Thermo Fisher Scientific, Carlsbad, CA, United States). The PCR specific primers were forward-huscfv-LIC: 5′-GGTTGGGAATTGCAAGCGGC CCAGCCGGCC-3′ and reverse-E-tag-LIC: 5′-GGAGATGGGA AGTCATTAACGCGGTTCCAGCGGATCC-3′. The huscfv inserts were designed to consist of a HuscFv-coding sequence linked to specific sequences for ligation independent cloning (LIC) protocol (Thermo Fisher Scientific). The amplified huscfv-E-tag DNAs were cloned separately into the pLATE52 vector (Thermo Fisher Scientific). Recombinant pLATE52-huscfv plasmids were transformed into JM109 E. coli by the heat-shock method. After PCR screening and DNA sequencing, the recombinant plasmids were introduced into an expression host, NiCo21(DE3) E. coli (New England Biolabs, St. Albans, Herts, United Kingdom), and the transformed bacteria were grown at 37°C for 16 h on LB agar containing 100 μg/ml of ampicillin. A single colony of each transformed clone was cultured in LB broth containing 100 μg/ml ampicillin with shaking (250 rpm) at 37°C for 16 h. The overnight cultures (12.5 ml) were separately inoculated into the fresh LB medium (250 ml) containing ampicillin and grown at 37°C until an OD600 nm reached ∼0.6–0.8. Recombinant HuscFv expression was induced by adding isopropyl-β-D-1-thiogalactopyranoside (IPTG) to a final concentration of 1 mM and incubated at 30°C for 6 h. The bacterial pellets were collected by centrifugation at 5,000 × g at 4°C for 20 min.
The recombinant HuscFvs were purified from the bacterial inclusion bodies (IBs) as described previously (Jittavisutthikul et al., 2016). Two grams of E. coli wet cell pellets were resuspended in 10 ml of BugBusterTM protein extraction reagent (Novagen, Schwalbach, Germany) and 20 μl of LysonaseTM bioprocessing reagent (Novagen) were added to each preparation. The preparations were kept at 25°C on a rotator for 20 min and cell pellets were collected after centrifugation at 8,000 × g at 4°C for 30 min. The IBs were washed with Wash-100 reagent [50 mM sodium phosphate buffer, pH 8.0; 500 mM NaCl; 5 mM EDTA; 8% (w/v) glycerol; and 1% (v/v) Triton X-100] twice and once with Wash-114 buffer [50 mM Tris–HCl, pH 8.0; 300 mM NaCl; and 1% (v/v) Triton X-114] with shaking at high speed for 40 min, and the IB pellets were then collected. The IBs were then washed with Wash-Solvent solution [50 mM Tris–HCl, pH 8.0; and 60% (v/v) isopropanol] and sterile ultrapure distilled water on ice, also with vigorous shaking, and centrifuged. Thereafter, 2.5 mg of purified IB pellets were solubilized in 5 ml of solubilizing buffer [50 mM CAPS, pH 11.0; 0.3% (w/v) N-lauryl sarcosine; and 1 mM dithiothreitol (DTT)] and kept at 4°C for 16 h. After dissolving completely, the protein was loaded into Snakeskin dialysis tubing with a molecular weight cut-off of 10 kDa (Thermo Fisher Scientific), and dialyzed against 750 ml of refolding buffer (20 mM imidazole, pH 8.5, supplemented with 0.1 mM DTT) at 4°C with slow stirring. After 3 h, the buffer was changed to a fresh refolding buffer, and dialysis was continued for 16 h. The refolded protein was subsequently dialyzed against a dialysis buffer without DTT with slow stirring at 4°C for 16 h. Each preparation was filtered through a 0.2-μm low protein binding Acrodisc® Syringe Filter (Pall, Port Washington, NY, United States) and kept at 30°C in a water bath for 3 h before adding 60 mM trehalose. The protein concentration of the refolded HuscFvs was determined using Pierce® BCA Protein Assay, while the quality and purity of the recombinant proteins were analyzed by SDS-PAGE and stained with Coomassie Brilliant Blue G-250 (Bio-rad, Hercules, CA, United States). Refolded HuscFv preparations were concentrated using Amicon® Ultra 4 ml 3K centrifugal filter devices (Merck Millipore, Darmstadt, Germany) and stored at −20°C until use.
The buffer of the HuscFv preparations was changed to 20 mM sodium phosphate buffer, pH 8.5, at a protein concentration of 0.1 mg/ml, and the antibodies were subjected to CD measurement. The data were recorded using a JASCO spectrometer (model J-815) equipped with a Peltier temperature controller system (Jasco, Tokyo, Japan) in a 1 mm path-length quartz cuvette. The proteins were scanned at 50 nm/min at 25°C. The CD spectra were collected over a wavelength range of 190–260 nm.
Human cervical carcinoma, HeLa, cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, Carlsbad, CA, United States) supplemented with 10% (v/v) fetal bovine serum (Hyclone, Novato, CA, United States) and 1% (v/v) penicillin-streptomycin (complete DMEM) at 37°C in a 5% CO2 atmosphere.
The monolayer of HeLa cells established in individual wells of a 24-well tissue culture plate (∼2 × 105 cells/well) were washed with sterile PBS, added with 2 μM of individual HuscFv preparations in complete DMEM, and incubated at 37°C in 5% CO2 atmosphere for 24 h. Cells in the medium alone served as a background control. After 24 h, the percent viability of cells of each treatment was analyzed using an FITC-Annexin V Apoptosis Detection Kit (BD Biosciences, San Jose, CA, United States) according to the manufacturer’s protocols. The cells were washed with Dulbecco’s phosphate-buffered saline (DPBS) and resuspended in binding buffer. Five microliters of Annexin V-FITC conjugate and 5 μl of propidium iodide (PI) were added. After 15-min incubation at room temperature (25°C) in darkness, apoptotic cells were enumerated by flow cytometric analysis (BD LSRFortessaTM, San Jose, CA, United States) using BD FACSDivaTM software (BD Biosciences). At least 20,000 events of single cells per sample were collected.
HeLa cells (∼2 × 105 cells/well) were treated with various concentrations of 3O-C12-HSL, i.e., 10, 25, 50, 75, and 100 μM. The background control comprised of cells incubated with medium alone. After incubation at 37°C in a CO2 incubator for 18 h, the cells were harvested and subjected to Annexin V/PI binding assay, as described above.
Fifty micromolars of 3O-C12-HSL in 0.25% DMSO were mixed with various concentrations of individual HuscFv preparations (0.25, 0.5, 1.0, and 1.2 μM) for 1 h before adding to HeLa cells (∼2 × 105 cells/well) and incubated at 37°C in a CO2 incubator for 18 h. After incubation, the cells were collected, washed, double-stained with Annexin V-FITC and PI, and analyzed by flow cytometry, as described above. Three independent experiments were performed.
HeLa cells (∼2 × 105 cells/well) were treated with a mixture of 50 μM 3O-C12-HSL and 1 μM of individual HuscFvs for 18 h. HeLa cells exposed to medium alone served as a control. After incubation, cells were washed with ice-cold PBS, fixed in 70% ethanol, and kept at −20°C overnight. Cells were then washed 3 times with ice-cold PBS and incubated in 500 μl of stain solution [10 μg/ml PI, 100 μg/ml RNase, and 0.1% (v/v) Triton X-100 in DPBS, pH 7.4] at room temperature in darkness for 30 min. The DNA contents of the cells were measured, and cell cycle histograms/distributions were generated. Then, the percentage of cells in the sub-G1 phase was determined by flow cytometry (BD LSRFortessaTM) using BD FACSDivaTM software (BD Biosciences), with at least 10,000 recorded events per sample.
Nuclear damage was studied using 4’,6-diamidino-2-phenylindole (DAPI) staining. Briefly, HeLa cells (1 × 106 cells) were seeded on a 22 × 22 mm square coverslip (Menzel-Glaser, Braunschweig, Germany) in a 6-well plate (Costar, New York, NY, United States) and kept at 37°C in a 5% CO2 incubator for 24 h. The culture medium was removed and the cells were replenished with complete DMEM containing a mixture of 3O-C12-HSL (50 μM) and HuscFvs (2 μM). After 18 h, the cells were washed and fixed with 4% (v/v) paraformaldehyde in PBS, permeabilized with 1% (v/v) Triton X-100 in PBS, blocked with 3% (w/v) bovine serum albumin (BSA) in PBS at room temperature for 30 min, then washed. The permeabilized cells were stained and mounted with DAPI (1:5,000) (Molecular Probes, Carlsbad, CA, United States) in the anti-fade mounting medium. DNA fragmentation and chromatin condensation were observed under a confocal microscope (Carl Zeiss Laser Scanning System LSM 700, Jena, Germany). Images were processed using the Zeiss LSM Image Browser (version 6.0.0.309).
Transmission electron microscopy (TEM) was used to examine the ultrastructural changes of the HeLa cell mitochondria after various treatments. The cells from each treatment group were fixed with 2.5% (v/v) glutaraldehyde in 0.1 M sucrose phosphate buffer (SPB) at room temperature for 1 h, washed three times with 0.1 M SPB, post-fixed with 1.0% (w/v) osmium tetroxide in the same buffer for 1 h, and dehydrated with a graded series of ethanol. The dehydrated cells were infiltrated with pure LR white embedded medium (EMS®, Hatfield, PA, United States) in 70% (v/v) ethanol, embedded in a capsule beam, and incubated at 65°C for 48 h. The ultrathin (100 nm) sections of the cells were prepared; the sections were positioned on a 200 square-mesh copper grid and stained with ethanolic uranyl acetate and lead citrate. The morphological and structural characteristics of mitochondria were observed under a transmission electron microscope (model HT7700, Hitachi, Tokyo, Japan).
Statistical analyses of all experiments were performed using GraphPad Prism 5 software (La Jolla, CA, United States). One-way ANOVA followed by Tukey’s post hoc multiple comparison tests were used to analyze the differences between groups. All data are shown as mean ± SD. Statistically significant difference was set at p < 0.05.
The refined models of HuscFvs for the selected HB2151 E. coli clones derived from phage biopanning with P. aeruginosa exotoxin A revealed that HuscFvs of three E. coli clones, i.e., E44 (HuscFv-E44), F15 (HuscFv-F15), and F19 (HuscFv-F19), showed reliable Ramachandran plots. The percent residues in the most favored regions, the additional allowed regions, the generously allowed regions, and the disallowed regions of the Ramachandran diagrams of the HuscFv-E44, HuscFv-F15, and HuscFv-F19 were 90.1, 6.8, 1.0, and 2.1 %; 91.3, 7.7, 0.5, and 0.5%; and 88.2, 9.4, 1.0, and 1.5%, respectively (Supplementary Figure S1).
From structural comparisons of individual HuscFvs with the antigen-binding site of the well-characterized quorum quenching mAb, i.e., RS2-1G9 (shown previously to bind to and neutralize the activities of P. aeruginosa 3O-C12-HSL), it was found that the binding pockets of the three HuscFvs were superimposed with the antigen-binding site of RS2-1G9. The coverage percentages of the overlapping structures between the modeled HuscFv-E44, HuscFv-F15, and HuscFv-F19 and the RS2-1G9 were 90.83, 89.29, and 88.58%, respectively (Figure 1 and Supplementary Table S1).
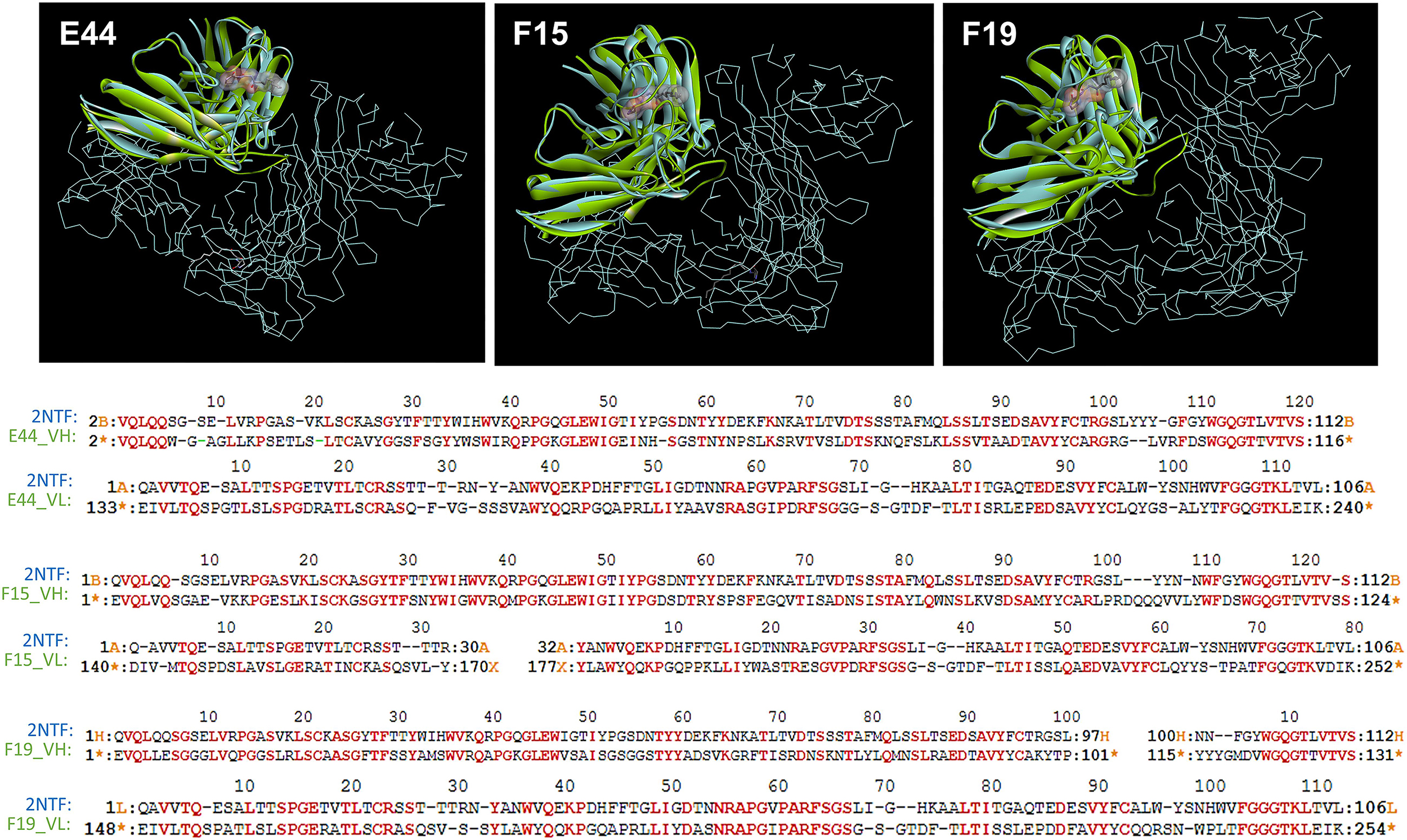
Figure 1. The 3D complex of RS2-1G9 F(ab′)2 fragment and N-Acyl-L-homoserine lactone analog (PDB:2NTF) was superimposed by the HuscFv-E44 (left), HuscFv-F15 (middle), and HuscFv-F19 (right) using CLICK: http://mspc.bii.a-star.edu.sg/click (Upper panel). One antigen-binding site of the mAb RS2-1G9 F(ab′)2 fragment (VH and VL domains, shown in blue) was superimposed by the HuscFvs (green). The trace illustration is the remaining portion of the RS2-1G9 F(ab′)2. Lower panel, the superimposed amino acids of the RS2-1G9 antigen-binding site (2NTF) and the VH and VL of HuscFv-E44, HuscFv-F15, and HuscFv-F19, are shown in red alphabets.
In silico intermolecular docking was performed to investigate the interaction of the HuscFvs with the 3O-C12-HSL. The residues of HuscFv-E44, HuscFv-F15, and HuscFv-F19 that tentatively formed interactive bonds with the haptenic 3O-C12-HSL target are shown in Figure 2 and Table 1. The Gibbs free energy (ΔG) of the representative complexes of respective HuscFvs with the ligand were −5.6, −5.8, and −5.4 kcal/mol, respectively.
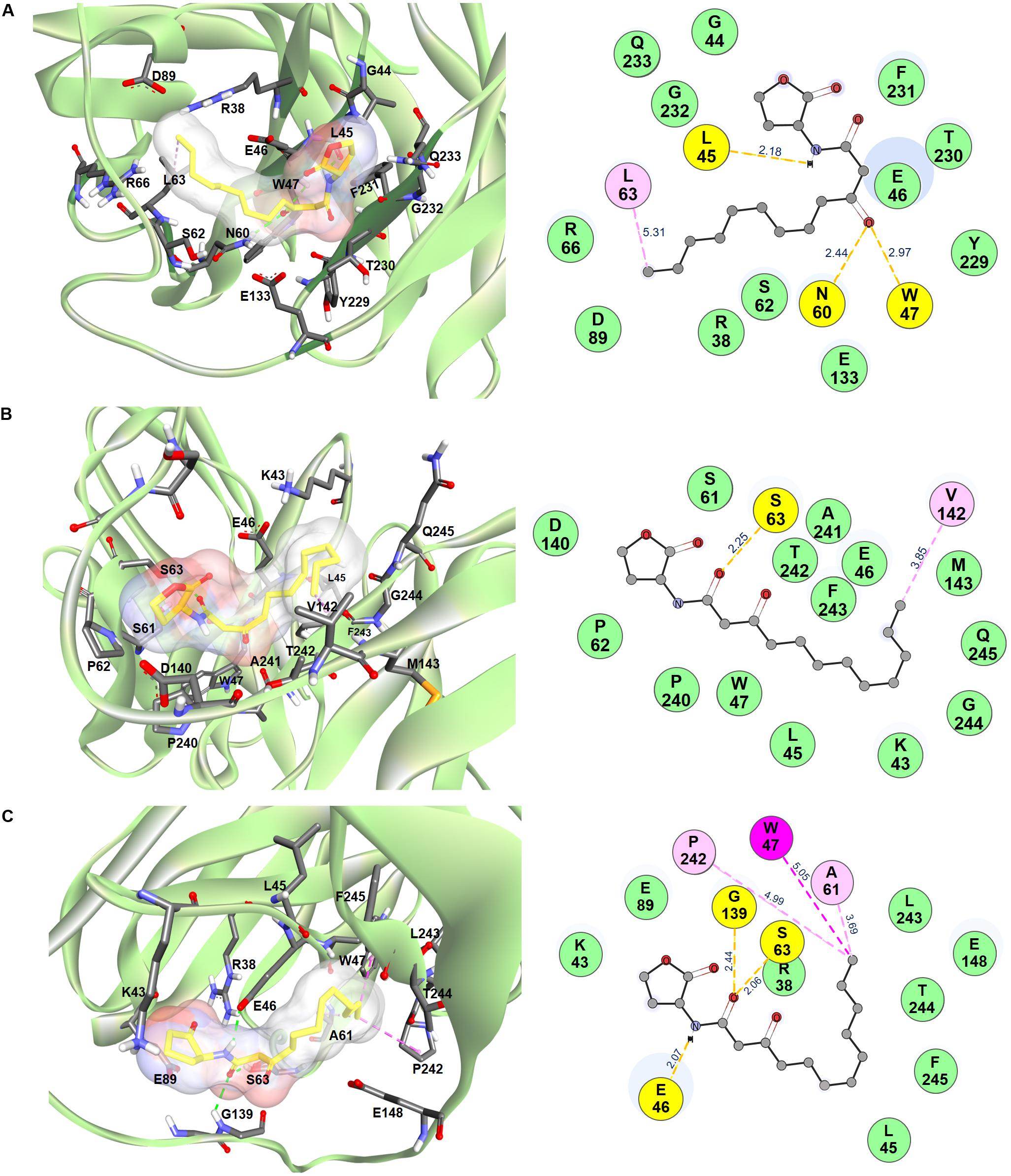
Figure 2. Computerized contact interfaces between 3O-C12-HSL and HuscFvs. (A–C) on the left side of the panels show interaction between 3D structures of 3O-C12-HSL (yellow stick) and green ribbons of (A) HuscFv-E44, (B) HuscFv-F15, (C) HuscFv-F19. Right side of the panels (A–C) show residues of the respective HuscFvs and the HuscFv-3O-C12-HSL interactive bonds (yellow, hydrogen bond; light pink, alkyl; magenta, Pi-alkyl; green, van der Waals force).
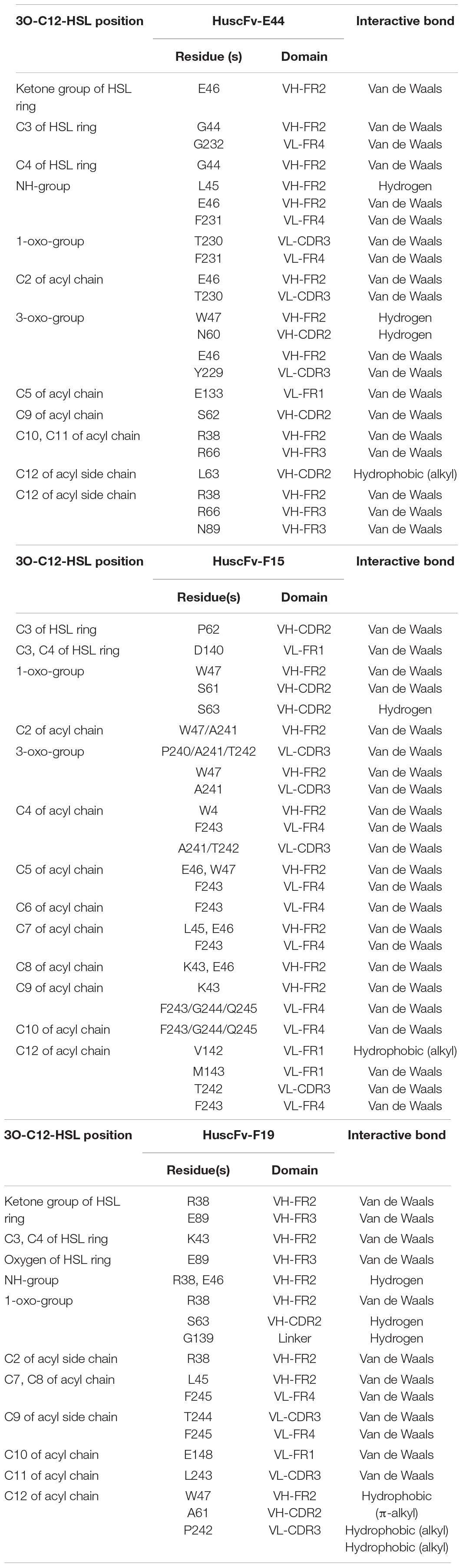
Table 1. Residues of Pseudomonas aeruginosa 3O-C12-HSL predicted to form contact interfaces with the effective HuscFv-E44, HuscFv-F15, and HuscFv-F19.
The HuscFv-E44 used residues from VH-CDR2 and VL-CDR3, as well as help from VH-FR2, VH-FR3, VL-FR1, and VL-FR4 to form contact interfaces with the functional groups of 3O-C12-HSL. The interactions were three hydrogen bonds between L45 of VH-FR2 with the NH group of the coordinated 3O-C12-HSL (2.18 Å) and N60 of VH-CDR2, and W47 of VH-FR2 with the 3O-C12-HSL carbonyl oxygen of 3-oxo-group of the acyl chain (2.97 and 2.44 Å, respectively). There was one hydrophobic interaction (alkyl) formed between L63 of VH-CDR2 and C12 of the acyl chain of AHL. HuscFv-E44 also used many residues in different domains to form contact interfaces via van der Waals forces with the 3O-C12-HSL, including S62 of VH-CDR2; Y229 and T230 of VL-CDR3; R38, G44, and E46 of VH-FR2; R66 and D89 of VH-FR3, E133 of VL-FR1; and F231, G232, and Q233 of VL-FR4 (Table 1).
HuscFv-F15 formed a hydrogen bond (2.25 Å) with the carbonyl oxygen of the 1-oxo-group of the fatty acid-like ligand through S63 of VH-CDR2. This antibody also used V142 of VL-FR1 to form hydrophobic contact (alkyl) with the C12 of the hapten acyl chain. Several other positions of the 3O-C12-HSL molecule have interacted via van der Waals forces with several residues of the HuscFv-F15 including S61 and P62 of VH-CDR2; P240, A241, and T242 of VL-CDR3; K43, L45, E46, and W47 of VH-FR2; D140 and M143 of VL-FR1; and F243, G244, and Q245 of VL-FR4.
Serine 63 of VH-CDR2 and G139 of the HuscFv-F19 linker formed contact with the carbonyl oxygen of the 1-oxo-group of the 3O-C12-HSL via hydrogen bonds (2.06 and 2.44 Å, respectively). Hydrogen bonding also occurred between E46 of VH-FR2 and the NH-group of the HSL backbone (2.07 Å). The last carbon atom of the long acyl side chain of the 3O-C12-HSL formed π-alkyl hydrophobic interaction with W47 of VH-FR2 as well as the alkyl hydrophobic interaction with A61 of VH-CDR2 and P242 of VL-CDR3. In addition, the HuscFv-F19 formed van der Waals contacts with the 3O-C12-HSL by using L243 and T244 of VL-CDR3; R38, K43, and L45 of VH-FR2; E89 of VH-FR3; E148 of VL-FR1; and F245 of VL-FR4.
The results of the structural comparison of the HuscFvs with the quorum quenching mAb, RS2-1G9, and the intermolecular docking between the HuscFvs and the 3O-C12-HSL enticed us to test further the ability of HuscFvs to neutralize P. aeruginosa 3O-C12-HSL activities.
The huscfv inserts in the pCANTAB5E phagemids of the E. coli clones E44, F15, and F19 were sub-cloned into the pLATE52 vector. The DNA construct in the vector is shown in Figure 3A. The amplicon of DNA coding for 6 × His tagged-HuscFv formed a PCR amplicon band at ∼ 850 bp, as revealed on agarose gel (Figure 3B). The refolded and purified HuscFv-E44, HuscFv-F15, and HuscFv-F19, with molecular sizes of about 34 kDa, are shown in Figure 3C.
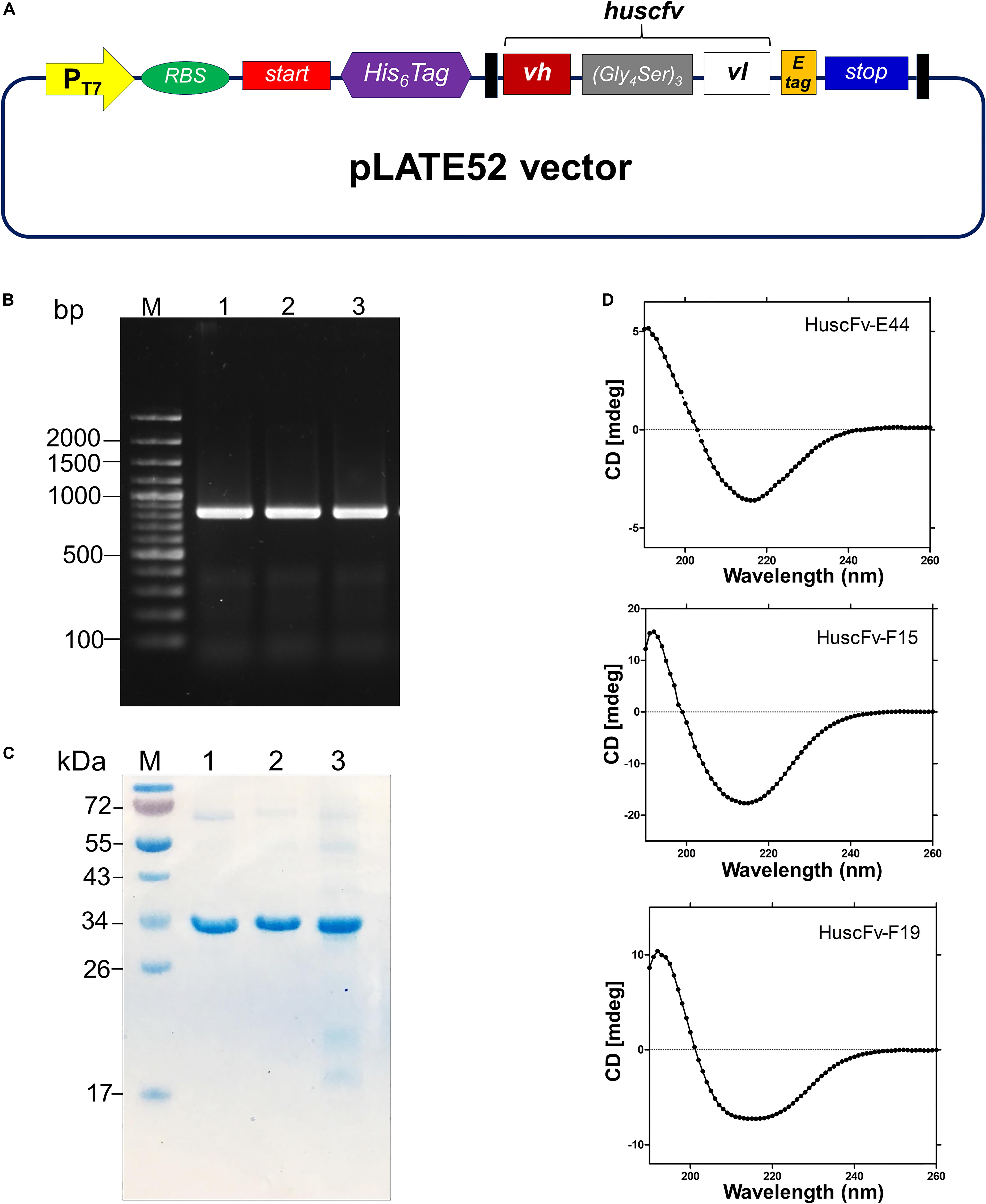
Figure 3. Production and characterization of HuscFvs to 3O-C12-HSL. (A) Schematic diagram of the inserted DNA construct in pLATE52 where the DNA sequence coding for HuscFv (vh-linker-vl) was flanked with DNA sequences of 6 × His at the 5′ end and E-tag at the 3′ end. (B) Amplicons of huscfv-LIC fragments (∼ 850 bp) for sub-cloning into pLATE52 vector. M, 100 bp-plus DNA ladder; 1–3, huscfv-LIC amplicons of three representatives transformed NiCo21(DE3) E. coli clones. Numbers at the left are DNA sizes in bp. (C) Stained SDS-PAGE-separated purified recombinant HuscFvs. M, protein standard; 1–3, purified HuscFv-E44, HuscFv-F15 and HuscFv-F19, respectively. Numbers at the left are protein masses in kDa. (D) CD spectra of the refolded HuscFv-E44, HuscFv-F15, and HuscFv-F19.
Secondary structures of the refolded HuscFvs were determined by far-UV CD spectroscopy. The far-UV CD spectra (190–260 nm) for all HuscFvs revealed their β-sheet structures, which shared a similar CD spectra pattern (Figure 3D). The antibody preparations did not form aggregates.
HeLa cells exposed to 2 μM of HuscFv-E44, HuscFv-F15, and HuscFv-F19 for 24 h showed more than 90% viability, which was not different from the cells in medium alone (p > 0.05) (Supplementary Figure S2) indicating biocompatibility of the HuscFvs to the representative mammalian cells.
The average percentages of apoptotic HeLa cells treated with 10, 25, 50, 75, and 100 μM of 3O-C12-HSL dissolved in 0.25% DMSO, from three independent experiments, were 6.67 ± 0.53, 10.08 ± 1.41, 20.85 ± 1.62, 36.37 ± 2.32, and 49.63 ± 2.51%, respectively, while the background apoptotic cells of the control (cells in culture medium) was 5.71 ± 0.59% (Figure 4A). Figure 4B shows the results of the flow cytometric analysis of apoptotic cells (stained with Annexin V/PI) from one representative experiment. The background apoptotic cells (% cell viability) with and without 0.25% DMSO in the culture medium were not different (Supplementary Figure S3).
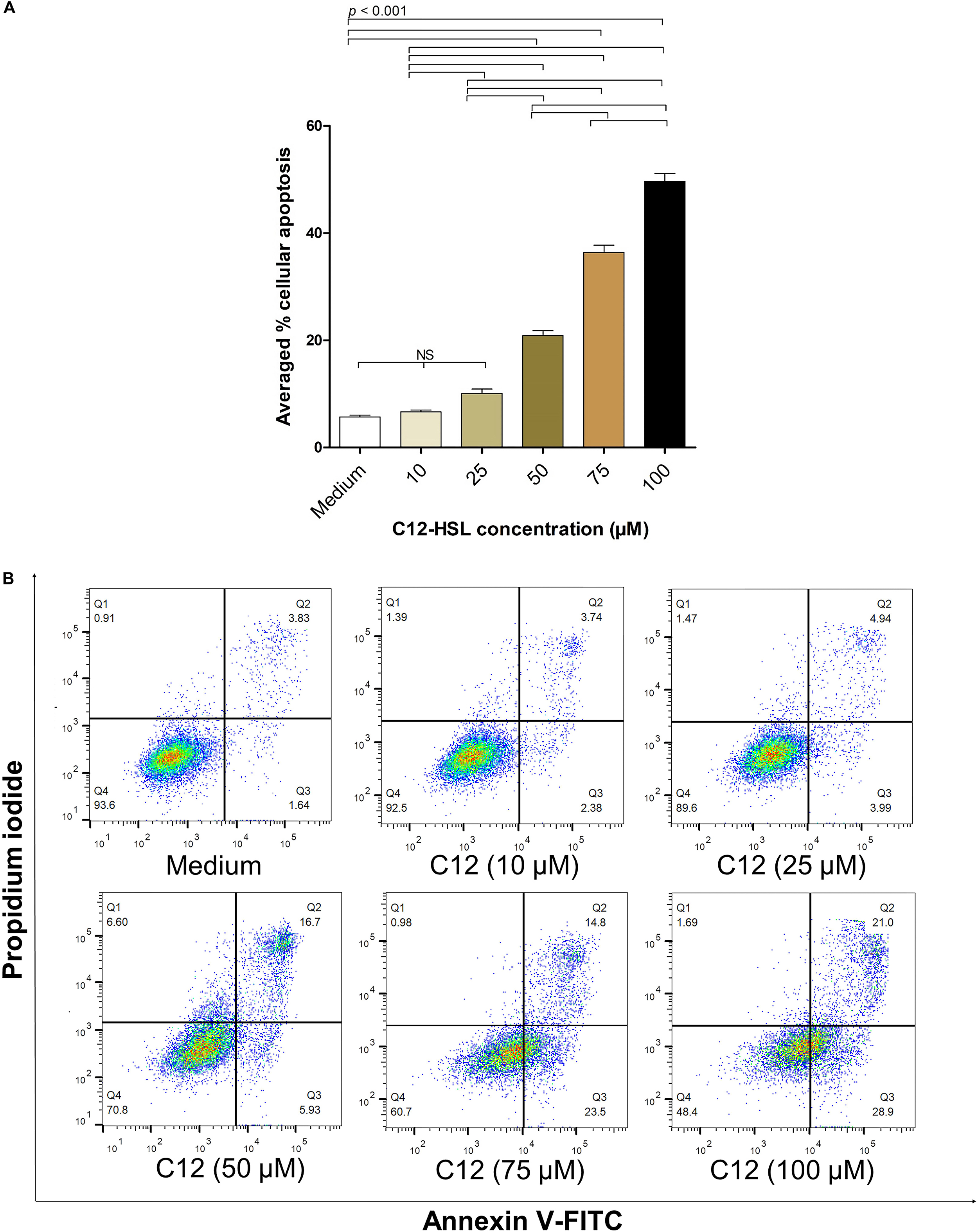
Figure 4. The percentages of apoptotic HeLa cells exposed to different concentrations of 3O-C12-HSL. (A) Bar graph of average percentages (means ± SD) of apoptotic cells from three independent experiments after exposure to 10, 25, 50, 75, and 100 μM of C12-HSL, compared with the cells in medium alone. (B) The density plots of HeLa cells after treatment with 10, 25, 50, 75, and 100 μM of C12-HSL, and control cells stained by Annexin V-FITC/PI and subjected to flow cytometric analysis (representative of three independent experiments). The cytotoxic activity of the 3O-C12-HSL was dose-dependent. Q1, necrotic cells (Annexin V negative/PI positive); Q2, late apoptotic cells (Annexin V positive/PI positive); Q3, early-apoptotic cells (Annexin V positive/PI negative), and Q4, viable cells (Annexin V negative/PI negative).
The percentages of apoptotic HeLa cells exposed to 50 μM of HuscFv-bound 3O-C12-HSL (0.25, 0.5, 1.0, and 1.2 μM of individual HuscFvs) were significantly decreased compared with those without HuscFvs (Table 2 and Figure 5). The HuscFvs of all three E. coli clones could neutralize 3O-C12-HSL, leading to reduced HeLa-cell apoptosis.
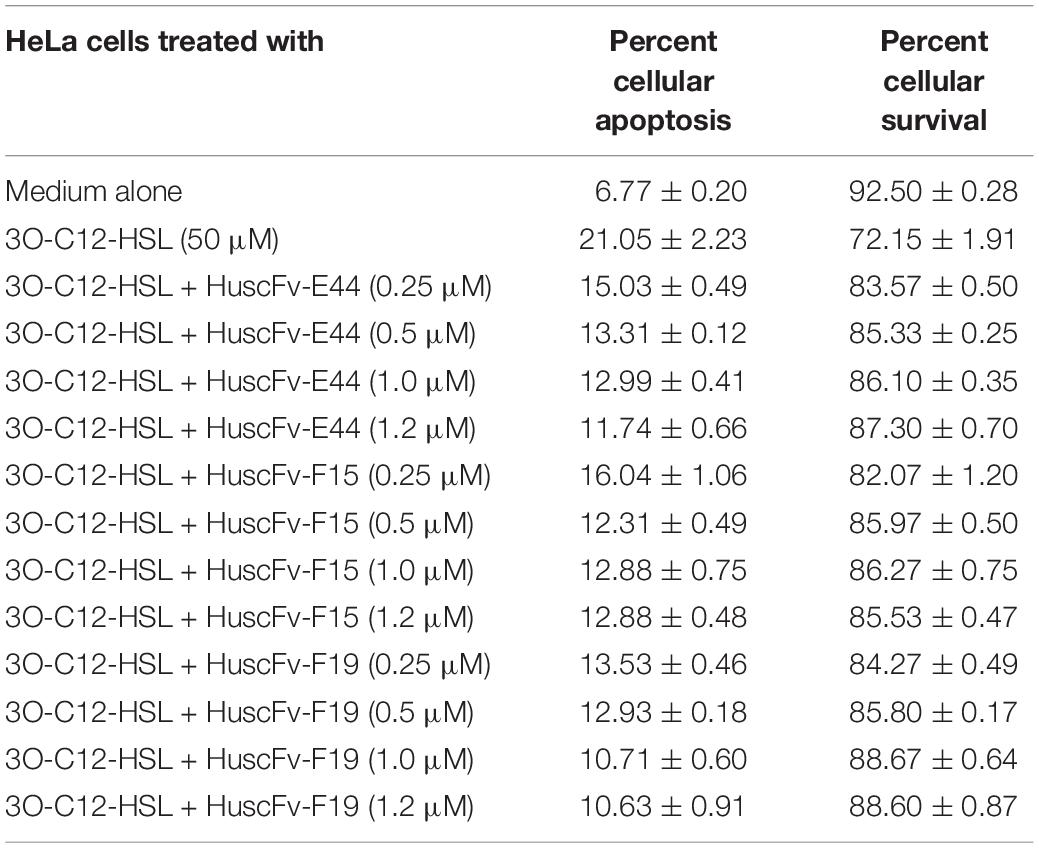
Table 2. Flow cytometric results evaluating the efficacy of HuscFvs using Annexin V-FITC/PI staining for 3O-C12-HSL-mediated cell apoptosis.

Figure 5. HuscFvs rescue HeLa cells from 3O-C12-HSL-mediated apoptosis. (A) The percentages of apoptotic cells of different treatment conditions, i.e., HeLa cells in medium alone, cells exposed to 50 μM 3O-C12-HSL (C12), cells added with 3O-C12-HSL mixed with various amounts of HuscFvs. (B) Density plots of flow cytometric analysis of doubly stained HeLa cells as in (A) (representative of one of the three reproducible experiments). The percent apoptotic cells caused by the 3O-C12-HSL was reduced significantly in the presence of HuscFvs.
Exposure of HeLa cells with 50 μM 3O-C12-HSL for 18 h resulted in 3.79 ± 0.52% of apoptotic cells in the hypodiploid DNA peak (sub-G1 population, which were apoptotic cells) as determined by flow cytometric analysis of the PI-stained cellular DNA. The numbers of cells with a hypodiploid DNA peak induced by the 3O-C12-HSL bound by the HuscFv-E44, HuscFv-F15, and HuscFv-F19, were decreased to 2.58 ± 0.10, 2.71 ± 0.10, and 1.79 ± 0.11%, respectively. The cells in medium alone had 1.04 ± 0.04% apoptotic cells (Figure 6). The results of the sub-G0/G1 analysis were conformed to those of the Annexin V/PI binding assay data.
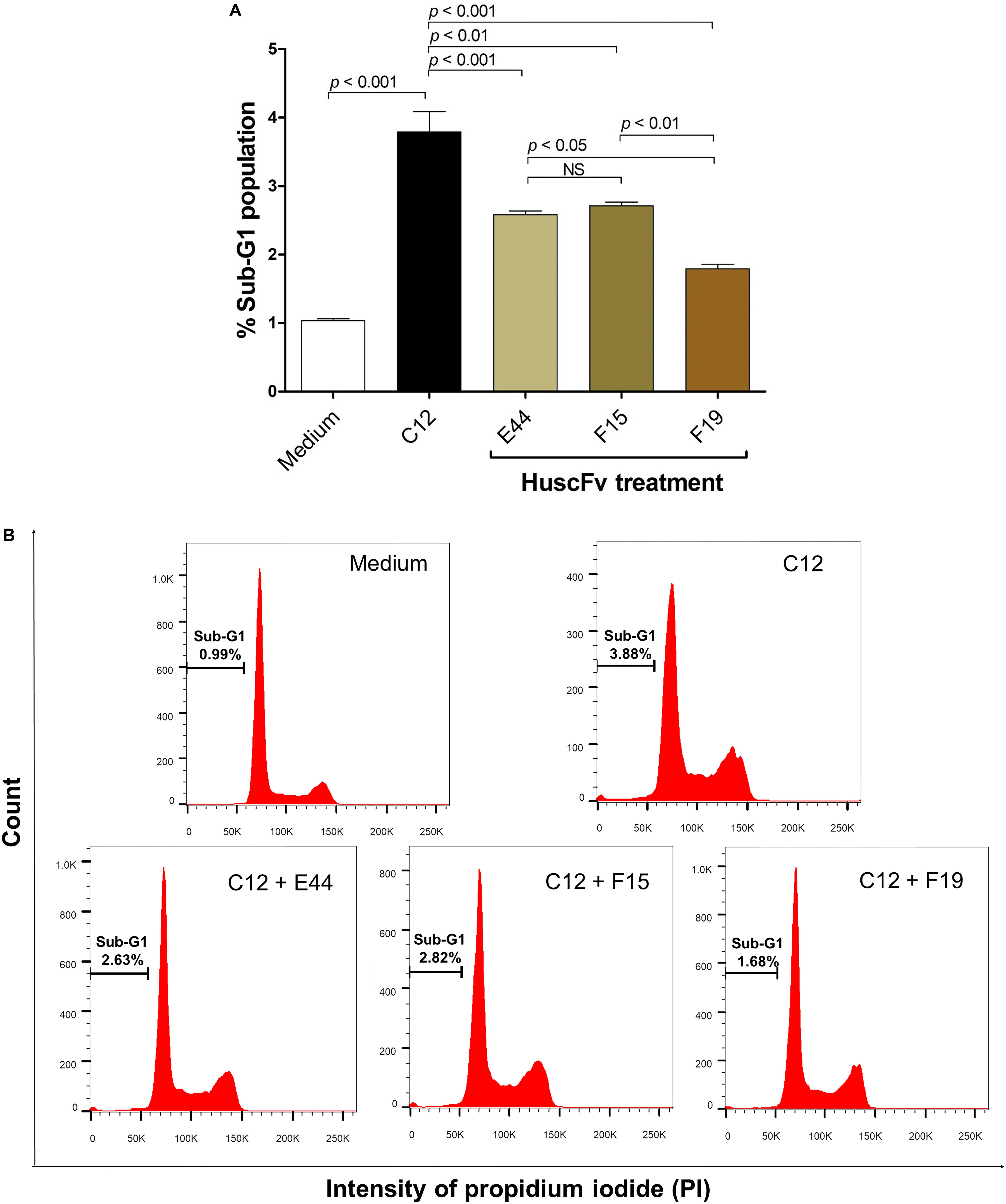
Figure 6. Reduction of sub-G1 arrest population of 3O-C12-HSL-exposed cells by HuscFvs. (A) Percent sub-G1 population (mean ± SD of triplicate individual experiments) after different treatments. (B) Histograms of cell cycle distribution. The percentage of apoptotic cells in the hypodiploid DNA peak (sub-G1 population) of each treatment is indicated in each plot.
DAPI staining and confocal microscopy were used to observe the intact HeLa nuclei and nuclear DNA damage induced by the 3O-C12-HSL and the HuscFv-bound 3O-C12-HSL (Figure 7). Intact nuclei of normal HeLa cells were stained weakly by the dye (Figure 7A), while the fragmented nuclei of the 3O-C12-HSL-exposed cells were stained brightly (Figure 7B). Damage to the nuclear DNA was reduced in cells exposed to HuscFv-F19-bound 3O-C12-HSL, as shown by the dimly stained nuclei (Figure 7C).
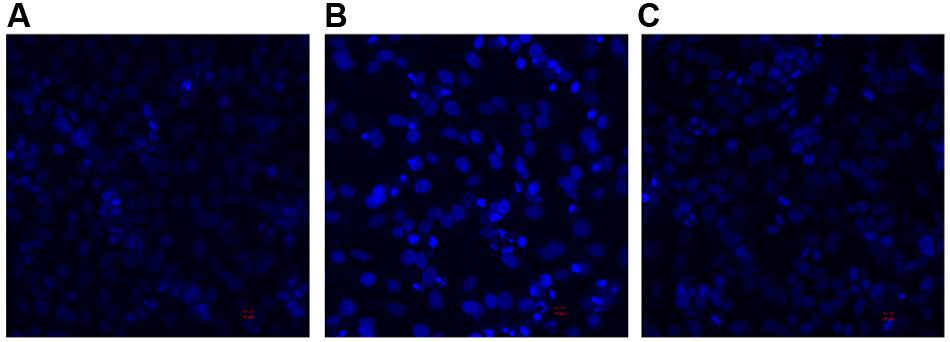
Figure 7. Appearances of DAPI stained-nuclei of HeLa cells after treatment with (A) Medium alone (B) 3O-C12-HSL (50 μM) and (C) mixture of 3O-C12-HSL (50 μM) and HuscFv-F19 (2 μM) for 18 h (original magnification 200×).
Transmission electron microscopy was used to study mitochondrial changes of the HeLa cells after exposure to the 3O-C12-HSL and HuscFv-bound 3O-C12-HSL, using the cells in medium alone as a normal control. As shown in Figures 8A,B, the mitochondria of the normal cells revealed an intact mitochondrial subcellular structure. In contrast, mitochondria of the cells treated with 50 μM of 3O-C12-HSL for 18 h exhibited a swollen appearance, with single or multiple distensions of the intercellular matrix in association with severe loss of cristae and double membranes (Figures 8C,D). The pathological changes of the mitochondria were ameliorated in the cells exposed to HuscFv-F19-bound 3O-C12-HSL (representative), i.e., mild mitochondrial swelling and more cristae (Figures 8E,F), compared with the 3O-C12-HSL-exposed cells.
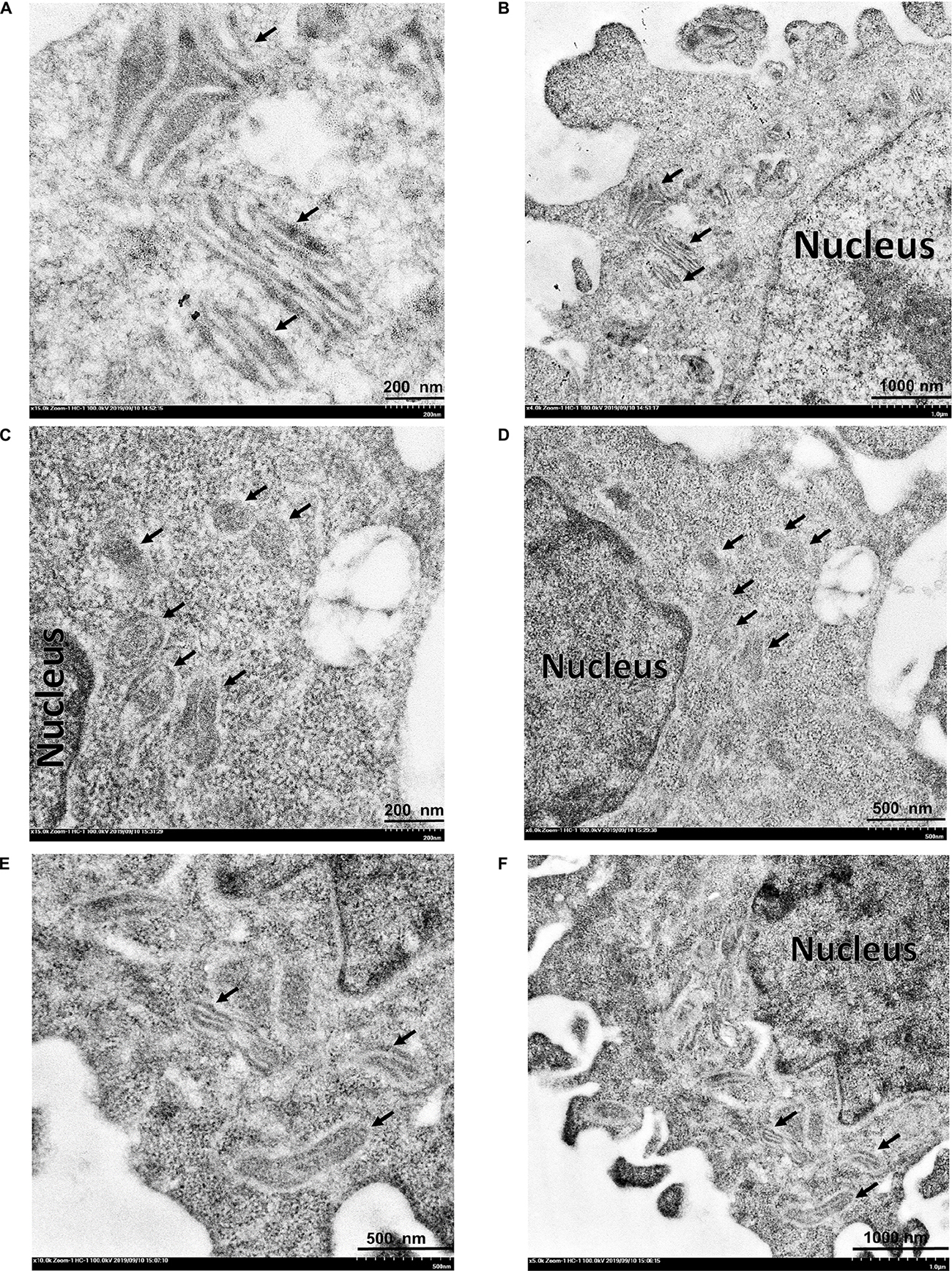
Figure 8. Transmission electron micrographs of mitochondrial ultrastructure. (A,B) normal mitochondria of HeLa cells. (C,D) HeLa cells treated with 3O-C12-HSL (50 μM). (E,F) HeLa cells treated with a mixture of 50 μM 3O-C12-HSL and 2 μM HuscFv-F19. Black arrows indicate mitochondria of the HeLa cells. Mitochondria of the cells exposed to the 3O-C12-HSL were swollen, characterized by a size increment with decreased electron density of the crista-free matrix.
Pseudomonas aeruginosa 3O-C12-HSL not only regulates virulence factors of the bacteria, but also causes inflammation in the infecting host by the induction of pro-inflammatory cytokine and chemokine synthesis (Smith et al., 2002). The 3O-C12-HSL killed mammalian cells through programmed cell death, i.e., an apoptotic mechanism at concentrations ranging from 10 to 100 μM by rapidly triggering depolarization of mitochondrial membrane potential and release of cytochrome c into cytosol, which activates the caspase cascades (Sultan and Sokolove, 2001; Tateda et al., 2003; Kravchenko et al., 2006; Schwarzer et al., 2012; Tao et al., 2016, 2018). The apoptotic cells manifest mitochondrial permeability transition (MPT), caspase activation, nuclear fragmentation, phosphatidylserine externalization, and cell shrinkage with apoptotic bodies (Wyllie et al., 1980; Cummings and Schnellmann, 2004). Mitochondrial swelling, depolarization, and membrane permeability are the key markers of the MPT that indicates mitochondria-stimulated programmed cell death in the pathogenesis of several diseases. Upon response to external stimuli or oxidative stress, the cells undergo continuous opening of permeability transition pores (PTP) in the mitochondrial inner membrane, which augments colloidal osmotic pressure in the matrix together with mitochondrial membrane depolarization, resulting in mitochondrial swelling (Chapa-Dubocq et al., 2018) followed by rupture of the mitochondrial outer membrane and release of cytochrome c into the cytosol and activation of caspase cascades (Petronilli et al., 2001). The stimulated caspase-3 activates endogenous endonuclease, which cleaves nuclear DNA (Zhang and Ming, 2000). Cells with apoptotic fragmented DNA or sub-G1 population are used as a marker of apoptosis (Riccardi and Nicoletti, 2006). In this study, 3O-C12-HSL produced a significant dose-dependent increment in mammalian cell death by inducing apoptosis, which validates previous notions on the cytotoxicity of P. aeruginosa QS substance.
Deletion of lasI or lasI and rhlI diminished the lung-colonization ability of P. aeruginosa in a mouse model of acute pneumonitis (Smith et al., 2002). P. aeruginosa mutants with defective QS are known to have less virulence and be more susceptible to antibiotic treatments and host immunity than the respective wild-type (Hentzer et al., 2003). As such, P. aeruginosa QS systems are attractive targets for direct-acting therapeutic agents, of which the expected treatment consequences are mitigation of the severity of the bacteria-associated diseases (Penesyan et al., 2015). During the past decades, several groups of P. aeruginosa QS inhibitors/modulators have been identified: small chemical molecules, i.e., AHL analogs (phenylpropionyl homoserine lactones and phenyloxyacetyl homoserine lactones of the N-aryl homoserine lactone library) (Geske et al., 2008), N-acyl cyclopentylamides (Ishida et al., 2007), halogenated furanone compound (Hentzer et al., 2002), other furanone derivatives (Kim et al., 2008), aspirin (El-Mowafy et al., 2014), and itaconimides and citraconimides (Fong et al., 2018); and natural inhibitors, such as secondary metabolites of the Australian marine macroalgae, Delisea pulchra (Givskov et al., 1996), patulin and penicillic acid from extracts of Penicillium species (Rasmussen et al., 2005), an organosulfur compound found in garlic extracts, named Ajoene (Jakobsen et al., 2012), and derivatives of ellagic acid (dilactone of hexahydroxydiphenic acid) from black or chebulic myrobalan, Terminalia chebula Retz (Sarabhai et al., 2013). Unfortunately, these compounds are relatively toxic to mammalian cells, which limits their therapeutic use (Ni et al., 2009). Recently, natural plant-derived compounds, trans-cinnamaldehyde (CA), and salicylic acid (SA) have been shown to effectively downregulate both las and rhl QS systems, reduce the production of extracellular virulence factors, i.e., protease, elastase, and pyocyanin, and reduce biofilm formation, concomitantly with repressed rhamnolipid gene expression (Ahmed et al., 2019). However, the sole use of QS inhibitors at high concentrations to eradicate bacterial infection completely is of legitimate concern due to potential toxicity (Shreaz et al., 2016).
Passive immunization has been used as an intervention for post-exposure morbidity and/or treatment of diseases since the late 18th century (Keller and Stiehm, 2000). An antibody molecule uses multiple amino acid residues in several CDRs (sometimes with the help of FRs) to form multiple non-covalent bonds with the target molecule, thus, making it difficult for pathogens to create antibody escape mutants, compared with small molecular drugs/inhibitors, from which resistant variants emerge rather easily and frequently. Therapeutic antibodies may be in the form of intact molecules (two antigen-binding sites with Fc fragment- when the bioactivities of the Fc are required for effectiveness) or merely smaller antibody fragments, i.e., F(ab′)2, Fab, scFv, or single domain (VH, VHH) with higher tissue penetrating ability than the intact four-chain counterpart when the Fc is dispensable. For P. aeruginosa infection, specific murine mAb, RS2-1G9, directed toward bacterial 3O-C12-HSL has been generated for use as an immunotherapeutic agent (Kaufmann et al., 2006, 2008). This murine antibody displayed the cytoprotective effect of 3O-C12-HSL-exposed host cells (Kaufmann et al., 2006, 2008; Debler et al., 2007). In addition, sheep-mouse chimeric mAb recognized native AHL protected mice from lethal P. aeruginosa infection (Palliyil et al., 2014). Nevertheless, while these 3O-C12-HSL-specific antibodies have therapeutic potential, their immunogenicity in human recipients, with possible adverse consequences, such as serum sickness, should be of concern.
Nowadays, any engineered fully human antibody format can be generated in vitro using phage display technology, invented by Nobel laureate, George Pearson Smith (Smith, 1985) as a biological tool (Santajit et al., 2019). The target antigens, such as proteins or peptides, attached to a carrier surface, e.g., fixed cell, plastic bead, or well of an ELISA plate, can be used as bait to fish out phage clones that display recombinant antibodies binding to the antigen from an antibody phage display library (Kulkeaw et al., 2009). Suppressor E. coli, such as strain HB2151 transfected with antigen-bound phages, when grown in appropriate conditioned medium, produces antigen-specific antibodies, and these antibodies can be isolated from the bacterial lysate/homogenate (Glab-Ampai et al., 2017; Santajit et al., 2019). Nevertheless, attachment of the small molecular haptens, like 3O-C12-HSL, to solid surfaces (as well as retaining the native configuration of the molecule) for conventional phage biopanning, is a relatively complicated process compared with proteins or peptides. Therefore, in this study, an alternative method was used to produce fully human scFvs (HuscFvs) to the synthetic P. aeruginosa 3O-C12-HSL based on the principle of antibody polyspecificity and antigenic molecular mimicry, i.e., completely unrelated molecules can share common receptors, possibly through similar structural and/or chemical features involved in recognition and binding (Wing, 1995; Tapryal et al., 2013). A repertoire of E. coli clones carrying recombinant huscfv-phagemids was previously retrieved from a HuscFv phage display library (Kulkeaw et al., 2009) by panning with P. aeruginosa exotoxin A (Santajit et al., 2019). Moreover, because the previously produced murine mAb, RS2-1G9, has been known as the P. aeruginosa quorum quencher, we used computerized antibody structure superimposition to select the bacterial derived-HuscFvs that shared structural homology with the murine mAb RS2-1G9 antigen-binding site. Using this method, the HuscFvs of three phagemid-transformed E. coli clones (E44, F15, and F19) showed high and satisfactory degrees of molecular similarity to the mAb RS2-1G9 antigen-binding site. Besides, these HuscFvs could neutralize the cytotoxic effects of the 3O-C12-HSL in the induction of cellular apoptosis. The HuscFv bound-3O-C12-HSL had a reduced capacity to mediate mitochondrial swelling, diminishing DNA damage and reducing sub-G1 arrest population of exposed cells. Unfortunately, the amount of C12-HSL inside the HeLa cells with and without HuscFv treatments were not measured; therefore, it is not known whether the HuscFvs could prevent HSL from entering the cells. Although the actual 3O-C12-HSL neutralizing mechanism of the HuscFvs needs laboratory investigation, the predicted structural complexes between the QS (ligand) and the HuScFvs (receptors) indicated that the latter used several residues in different CDRs and FRs to interact non-covalently with the target, including van der Waals’ forces, hydrophobic interactions, and hydrogen bonds. These interactions might render the disarming of the bacterial toxic molecule through C12-HSL signal interference, which would mitigate bacterial disease severity. This perspective needs further testing of the HuscFvs on P. aeruginosa phenotypes both in vitro (bacterial culture), such as expression of the QS controlled virulence factors, as well as in the in vivo model of bacterial infection.
The engineered human single-chain variable fragments that attenuated the potent cytotoxicity of the P. aeruginosa quorum sensing molecule, 3O-C12-HSL, were generated successfully through the molecular basis of antibody polyspecificity and antigenic mimicry. The fully human antibody fragments rescued mammalian cells from the 3O-C12-HSL-mediated mitochondrial injuries, DNA damage, and cellular apoptosis in vitro. They should be tested further by step-by-step in vivo toward the clinical application as a sole or an adjunct therapy for the failing antibiotic treatment of P. aeruginosa infections.
All datasets generated for this study are included in the article/Supplementary Material.
NI and WC conceived the project, analyzed the data, and edited the manuscript. SS did most of the experiments, drafted the manuscript, and prepared the figures. KM helped SS with flow cytometric analysis. WS advised SS on recombinant HuscFv production. PP supervised SS on fluorescence staining and confocal microscopy. SA performed the electron microscopy. NS, OR, and MC-N helped NI and WC to analyze the data and made comments. All authors critically reviewed the manuscript and gave final approval for publication.
This work was co-supported by the RSA scholar grant (RSA5980048) to NI, NSTDA Chair Professor grant (P-1450624) funded by the Crown Property Bureau of Thailand to WC, and Royal Golden Jubilee Ph.D. scholarship grant (PHD/0073/2558) to SS from the Thailand Research Fund.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We acknowledge the Center of Research Excellence on Therapeutic Proteins and Antibody Engineering, Department of Parasitology, and Biomedical Research Unit, Department of Research, Faculty of Medicine Siriraj Hospital, for their technical support.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01172/full#supplementary-material
Abhinandan, K. R., and Martin, A. C. (2008). Analysis and improvements to Kabat and structurally correct numbering of antibody variable domains. Mol. Immunol. 45, 3832–3839. doi: 10.1016/j.molimm.2008.05.022
Ahmed, S. A. K. S., Rudden, M., Smyth, T. J., Dooley, J. S., Marchant, R., and Banat, I. M. (2019). Natural quorum sensing inhibitors effectively downregulate gene expression of Pseudomonas aeruginosa virulence factors. Appl. Microbiol. Biotechnol. 103, 3521–3535. doi: 10.1007/s00253-019-09618-0
Bleves, S., Soscia, C., Nogueira-Orlandi, P., Lazdunski, A., and Filloux, A. (2005). Quorum sensing negatively controls type III secretion regulon expression in Pseudomonas aeruginosa PAO1. J. Bacteriol. 187, 3898–3902. doi: 10.1128/jb.187.11.3898-3902.2005
Boontham, P., Robins, A., Chandran, P., Pritchard, D., Cámara, M., Williams, P., et al. (2008). Significant immunomodulatory effects of Pseudomonas aeruginosa quorum-sensing signal molecules: possible link in human sepsis. Clin. Sci. 115, 343–351. doi: 10.1042/cs20080018
Chapa-Dubocq, X., Makarov, V., and Javadov, S. (2018). Simple kinetic model of mitochondrial swelling in cardiac cells. J. Cell. Physiol. 233, 5310–5321. doi: 10.1002/jcp.26335
Cummings, B. S., and Schnellmann, R. G. (2004). Measurement of cell death in mammalian cells. Curr. Protoc. Pharmacol. 25, 12–18.
Debler, E. W., Kaufmann, G. F., Kirchdoerfer, R. N., Mee, J. M., Janda, K. D., and Wilson, I. A. (2007). Crystal structures of a quorum-quenching antibody. J. Mol. Biol. 368, 1392–1402. doi: 10.1016/j.jmb.2007.02.081
Defoirdt, T., Boon, N., and Bossier, P. (2010). Can bacteria evolve resistance to quorum sensing disruption? PLoS Pathog. 6:e1000989. doi: 10.1371/journal.ppat.1000989
Duan, K., and Surette, M. G. (2007). Environmental regulation of Pseudomonas aeruginosa PAO1 Las and Rhl quorum-sensing systems. J. Bacteriol. 189, 4827–4836. doi: 10.1128/jb.00043-07
Eberhard, A., Burlingame, A. L., Eberhard, C., Kenyon, G. L., Nealson, K. H., and Oppenheimer, N. J. (1981). Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 20, 2444–2449. doi: 10.1021/bi00512a013
El-Mowafy, S. A., El Galil, K. H. A., El-Messery, S. M., and Shaaban, M. I. (2014). Aspirin is an efficient inhibitor of quorum sensing, virulence and toxins in Pseudomonas aeruginosa. Microb. Pathog. 74, 25–32. doi: 10.1016/j.micpath.2014.07.008
Fong, J., Mortensen, K. T., Nørskov, A., Qvortrup, K., Yang, L., Tan, C. H., et al. (2018). Itaconimides as novel quorum sensing inhibitors of Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 8:443. doi: 10.3389/fcimb.2018.00443
Forli, S., Huey, R., Pique, M. E., Sanner, M. F., Goodsell, D. S., and Olson, A. J. (2016). Computational protein–ligand docking and virtual drug screening with the AutoDock suite. Nat. Protoc. 11, 905–919. doi: 10.1038/nprot.2016.051
Geske, G. D., Mattmann, M. E., and Blackwell, H. E. (2008). Evaluation of a focused library of N-aryl L-homoserine lactones reveals a new set of potent quorum sensing modulators. Bioorg. Med. Chem. Lett. 18, 5978–5981. doi: 10.1016/j.bmcl.2008.07.089
Givskov, M., de Nys, R., Manefield, M., Gram, L., Maximilien, R. I. A., Eberl, L. E. O., et al. (1996). Eukaryotic interference with homoserine lactone-mediated prokaryotic signalling. J. Bacteriol. 178, 6618–6622. doi: 10.1128/jb.178.22.6618-6622.1996
Glab-Ampai, K., Chulanetra, M., Malik, A. A., Juntadech, T., Thanongsaksrikul, J., Srimanote, P., et al. (2017). Human single chain-transbodies that bound to domain-I of non-structural protein 5A (NS5A) of hepatitis C virus. Sci. Rep. 7:15042.
Hentzer, M., Riedel, K., Rasmussen, T. B., Heydorn, A., Andersen, J. B., Parsek, M. R., et al. (2002). Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 148, 87–102. doi: 10.1099/00221287-148-1-87
Hentzer, M., Wu, H., Andersen, J. B., Riedel, K., Rasmussen, T. B., Bagge, N., et al. (2003). Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J. 22, 3803–3815. doi: 10.1093/emboj/cdg366
Ishida, T., Ikeda, T., Takiguchi, N., Kuroda, A., Ohtake, H., and Kato, J. (2007). Inhibition of quorum sensing in Pseudomonas aeruginosa by N-acyl cyclopentylamides. Appl. Environ. Microbiol. 73, 3183–3188. doi: 10.1128/aem.02233-06
Jacobi, C. A., Schiffner, F., Henkel, M., Waibel, M., Stork, B., Daubrawa, M., et al. (2009). Effects of bacterial N-acyl homoserine lactones on human Jurkat T lymphocytes-OdDHL induces apoptosis via the mitochondrial pathway. Int. J. Med. Microbiol. 299, 509–519. doi: 10.1016/j.ijmm.2009.03.005
Jakobsen, T. H., van Gennip, M., Phipps, R. K., Shanmugham, M. S., Christensen, L. D., Alhede, M., et al. (2012). Ajoene, a sulfur-rich molecule from garlic, inhibits genes controlled by quorum sensing. Antimicrob. Agents Chemother. 56, 2314–2325. doi: 10.1128/aac.05919-11
Jittavisutthikul, S., Seesuay, W., Thanongsaksrikul, J., Thueng-in, K., Srimanote, P., Werner, R. G., et al. (2016). Human transbodies to HCV NS3/4A protease inhibit viral replication and restore host innate immunity. Front. Immunol. 7, 318. doi: 10.3389/fimmu.2016.00318
Kalia, V. C., Wood, T. K., and Kumar, P. (2014). Evolution of resistance to quorum-sensing inhibitors. Microb. Ecol. 68, 13–23.
Kaufmann, G. F., Park, J., Mee, J. M., Ulevitch, R. J., and Janda, K. D. (2008). The quorum quenching antibody RS2-1G9 protects macrophages from the cytotoxic effects of the Pseudomonas aeruginosa quorum sensing signaling molecule N-3-oxo-dodecanoyl-homoserine lactone. Mol. Immunol. 45, 2710–2714. doi: 10.1016/j.molimm.2008.01.010
Kaufmann, G. F., Sartorio, R., Lee, S. H., Mee, J. M., Altobell, L. J., Kujawa, D. P., et al. (2006). Antibody interference with N-acyl homoserine lactone-mediated bacterial quorum sensing. J. Am. Chem. Soc. 128, 2802–2803. doi: 10.1021/ja0578698
Keller, M. A., and Stiehm, E. R. (2000). Passive immunity in prevention and treatment of infectious diseases. Clin. Microbiol. Rev. 13, 602–614. doi: 10.1128/cmr.13.4.602
Kim, C., Kim, J., Park, H. Y., Park, H. J., Lee, J. H., Kim, C. K., et al. (2008). Furanone derivatives as quorum-sensing antagonists of Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 80, 37–47.
Kim, S., Thiessen, P. A., Bolton, E. E., Chen, J., Fu, G., Gindulyte, A., et al. (2015). PubChem substance and compound databases. Nucleic Acids Res. 44, D1202–D1213.
Kravchenko, V. V., Kaufmann, G. F., Mathison, J. C., Scott, D. A., Katz, A. Z., Wood, M. R., et al. (2006). N-(3-oxo-acyl) homoserine lactones signal cell activation through a mechanism distinct from the canonical pathogen-associated molecular pattern recognition receptor pathways. J. Biol. Chem. 281, 28822–28830. doi: 10.1074/jbc.m606613200
Krzyżek, P. (2019). Challenges and limitations of anti-quorum sensing therapies. Front. Microbiol. 10:2473. doi: 10.3389/fmicb.2019.02473
Kulkeaw, K., Sakolvaree, Y., Srimanote, P., Tongtawe, P., Maneewatch, S., Sookrung, N., et al. (2009). Human monoclonal ScFv neutralize lethal Thai cobra. Naja kaouthia, neurotoxin. J. Proteomics 72, 270–280.
Laskowski, R. A., MacArthur, M. W., Moss, D. S., and Thornton, J. M. (1993). PROCHECK: a program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 26, 283–291. doi: 10.1107/s0021889892009944
Li, L., Hooi, D., Chhabra, S. R., Pritchard, D., and Shaw, P. E. (2004). Bacterial N-acylhomoserine lactone-induced apoptosis in breast carcinoma cells correlated with down-modulation of STAT3. Oncogene 23, 4894–4902. doi: 10.1038/sj.onc.1207612
Liu, Y. C., Chan, K. G., and Chang, C. Y. (2015). Modulation of host biology by Pseudomonas aeruginosa quorum sensing signal molecules: messengers or traitors. Front. Microbiol. 6:1226. doi: 10.3389/fmicb.2015.01226
López-Jácome, L. E., Garza Ramos-Martínez, G., Hernández-Durán, M., Franco-Cendejas, R., Loarca, D., Romero-Martínez, D., et al. (2019). AiiM lactonase strongly reduces quorum sensing controlled virulence factors in clinical strains of Pseudomonas aeruginosa isolated from burned patients. Front. Microbiol. 10:2657. doi: 10.3389/fmicb.2019.02657
Maeda, T., García-Contreras, R., Pu, M., Sheng, L., Garcia, L. R., Tomás, M., et al. (2012). Quorum quenching quandary: resistance to antivirulence compounds. ISME J. 6, 493–501. doi: 10.1038/ismej.2011.122
Malhotra, S., Hayes, D., and Wozniak, D. J. (2019). Cystic fibrosis and Pseudomonas aeruginosa: the host-microbe interface. Clin. Microbiol. Rev. 32:e00138-18.
Miyairi, S., Tateda, K., Fuse, E. T., Ueda, C., Saito, H., Takabatake, T., et al. (2006). Immunization with 3-oxododecanoyl-L-homoserine lactone-protein conjugate protects mice from lethal Pseudomonas aeruginosa lung infection. J. Med. Microbiol. 55, 1381–1387. doi: 10.1099/jmm.0.46658-0
Moradali, M. F., Ghods, S., and Rehm, B. H. (2017). Pseudomonas aeruginosa lifestyle: a paradigm for adaptation, survival, and persistence. Front. Cell. Infect. Microbiol. 7:39. doi: 10.3389/fcimb.2017.00039
Nguyen, M. N., Tan, K. P., and Madhusudhan, M. S. (2011). CLICK—topology-independent comparison of biomolecular 3D structures. Nucleic Acids Res. 39, W24–W28.
Ni, N., Li, M., Wang, J., and Wang, B. (2009). Inhibitors and antagonists of bacterial quorum sensing. Med. Res. Rev. 29, 65–124. doi: 10.1002/med.20145
O’Connor, G., Knecht, L. D., Salgado, N., Strobel, S., Pasini, P., and Daunert, S. (2015). Whole-cell biosensors as tools for the detection of quorum-sensing molecules: uses in diagnostics and the investigation of the quorum-sensing mechanism. Adv. Biochem. Eng. Biotechnol. 154, 181–200. doi: 10.1007/10_2015_337
Palliyil, S., and Broadbent, I. D. (2009). Novel immunotherapeutic approaches to the treatment of infections caused by Gram-negative bacteria. Curr. Opin. Pharmacol. 9, 566–570. doi: 10.1016/j.coph.2009.07.007
Palliyil, S., Downham, C., Broadbent, I., Charlton, K., and Porter, A. J. (2014). High-sensitivity monoclonal antibodies specific for homoserine lactones protect mice from lethal Pseudomonas aeruginosa infections. Appl. Environ. Microbiol. 80, 462–469. doi: 10.1128/aem.02912-13
Pearson, J. P., Gray, K. M., Passador, L., Tucker, K. D., Eberhard, A., Iglewski, B. H., et al. (1994). Structure of the autoinducer required for expression of Pseudomonas aeruginosa virulence genes. Proc. Natl. Acad. Sci. U.S.A. 91, 197–201. doi: 10.1073/pnas.91.1.197
Pearson, J. P., Passador, L., Iglewski, B. H., and Greenberg, E. P. (1995). A second N-acylhomoserine lactone signal produced by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 92, 1490–1494. doi: 10.1073/pnas.92.5.1490
Penesyan, A., Gillings, M., and Paulsen, I. T. (2015). Antibiotic discovery: combatting bacterial resistance in cells and in biofilm communities. Molecules 20, 5286–5298. doi: 10.3390/molecules20045286
Petronilli, V., Penzo, D., Scorrano, L., Bernardi, P., and Di Lisa, F. (2001). The mitochondrial permeability transition, release of cytochrome c and cell death correlation with the duration of pore openings in situ. J. Biol. Chem. 276, 12030–12034. doi: 10.1074/jbc.m010604200
Preston, M. J., Seed, P. C., Toder, D. S., Iglewski, B. H., Ohman, D. E., Gustin, J. K., et al. (1997). Contribution of proteases and LasR to the virulence of Pseudomonas aeruginosa during corneal infections. Infect. Immun. 65, 3086–3090. doi: 10.1128/iai.65.8.3086-3090.1997
Rasamiravaka, T., and El Jaziri, M. (2016). Quorum-sensing mechanisms and bacterial response to antibiotics in P. aeruginosa. Curr. Microbiol. 73, 747–753. doi: 10.1007/s00284-016-1101-1
Rasmussen, T. B., and Givskov, M. (2006). Quorum-sensing inhibitors as anti-pathogenic drugs. Int. J. Med. Microbiol. 296, 149–161. doi: 10.1016/j.ijmm.2006.02.005
Rasmussen, T. B., Skindersoe, M. E., Bjarnsholt, T., Phipps, R. K., Christensen, K. B., Jensen, P. O., et al. (2005). Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 151, 1325–1340. doi: 10.1099/mic.0.27715-0
Riccardi, C., and Nicoletti, I. (2006). Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat. Protoc. 1, 1458–1461. doi: 10.1038/nprot.2006.238
Ritchie, A. J., Whittall, C., Lazenby, J. J., Chhabra, S. R., Pritchard, D. I., and Cooley, M. A. (2007). The immunomodulatory Pseudomonas aeruginosa signalling molecule N-(3-oxododecanoyl)-l-homoserine lactone enters mammalian cells in an unregulated fashion. Immunol. Cell. Biol. 85, 596–602. doi: 10.1038/sj.icb.7100090
Rutherford, S. T., and Bassler, B. L. (2012). Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb. Perspect. Med. 2:a012427. doi: 10.1101/cshperspect.a012427
Sadikot, R. T., Blackwell, T. S., Christman, J. W., and Prince, A. S. (2005). Pathogen-host interactions in Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 171, 1209–1223.
Santajit, S., Seesuay, W., Mahasongkram, K., Sookrung, N., Ampawong, S., Reamtong, O., et al. (2019). Human single-chain antibodies that neutralize Pseudomonas aeruginosa-exotoxin A-mediated cellular apoptosis. Sci. Rep. 9, 1–15.
Sarabhai, S., Sharma, P., and Capalash, N. (2013). Ellagic acid derivatives from Terminalia chebula Retz. downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS One 8:e53441. doi: 10.1371/journal.pone.0053441
Schwarzer, C., Fu, Z., Patanwala, M., Hum, L., Lopez-Guzman, M., Illek, B., et al. (2012). Pseudomonas aeruginosa biofilm-associated homoserine lactone C12 rapidly activates apoptosis in airway epithelia. Cell. Microbiol. 14, 698–709. doi: 10.1111/j.1462-5822.2012.01753.x
Shiner, E. K., Terentyev, D., Bryan, A., Sennoune, S., Martinez-Zaguilan, R., Li, G., et al. (2006). Pseudomonas aeruginosa autoinducer modulates host cell responses through calcium signalling. Cell. Microbiol. 8, 1601–1610. doi: 10.1111/j.1462-5822.2006.00734.x
Shreaz, S., Wani, W. A., Behbehani, J. M., Raja, V., Irshad, M., Karched, M., et al. (2016). Cinnamaldehyde and its derivatives, a novel class of antifungal agents. Fitoterapia 112, 116–131. doi: 10.1016/j.fitote.2016.05.016
Silva Filho, L. V. R. F., Ferreira, F. D. A., Reis, F. J. C., Britto, M. C. A. D., Levy, C. E., Clark, O., et al. (2013). Pseudomonas aeruginosa infection in patients with cystic fibrosis: scientific evidence regarding clinical impact, diagnosis, and treatment. J. Bras. Pneumol. 39, 495–512. doi: 10.1590/s1806-37132013000400015
Smith, G. P. (1985). Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science 228, 1315–1317. doi: 10.1126/science.4001944
Smith, R. S., Harris, S. G., Phipps, R., and Iglewski, B. (2002). The Pseudomonas aeruginosa quorum-sensing molecule N-(3-oxododecanoyl) homoserine lactone contributes to virulence and induces inflammation in vivo. J. Bacteriol. 184, 1132–1139. doi: 10.1128/jb.184.4.1132-1139.2002
Soto-Aceves, M. P., Cocotl-Yañez, M., Merino, E., Castillo-Juárez, I., Cortés-López, H., González-Pedrajo, B., et al. (2019). Inactivation of the quorum-sensing transcriptional regulators LasR or RhlR does not suppress the expression of virulence factors and the virulence of Pseudomonas aeruginosa PAO1. Microbiology 165, 425–432. doi: 10.1099/mic.0.000778
Sultan, A., and Sokolove, P. M. (2001). Free fatty acid effects on mitochondrial permeability: an overview. Arch. Biochem. Biophys. 386, 52–61. doi: 10.1006/abbi.2000.2195
Tang, H. B., DiMango, E., Bryan, R., Gambello, M., Iglewski, B. H., Goldberg, J. B., et al. (1996). Contribution of specific Pseudomonas aeruginosa virulence factors to pathogenesis of pneumonia in a neonatal mouse model of infection. Infect. Immun. 64, 37–43. doi: 10.1128/iai.64.1.37-43.1996
Tao, S., Luo, Y., He, B., Liu, J., Qian, X., Ni, Y., et al. (2016). Paraoxonase 2 modulates a proapoptotic function in LS174T cells in response to quorum sensing molecule N-(3-oxododecanoyl)-L-homoserine lactone. Sci. Rep. 6:28778.
Tao, S., Niu, L., Cai, L., Geng, Y., Hua, C., Ni, Y., et al. (2018). N-(3-oxododecanoyl)-l-homoserine lactone modulates mitochondrial function and suppresses proliferation in intestinal goblet cells. Life Sci. 201, 81–88. doi: 10.1016/j.lfs.2018.03.049
Tapryal, S., Gaur, V., Kaur, K. J., and Salunke, D. M. (2013). Structural evaluation of a mimicry-recognizing paratope: plasticity in antigen–antibody interactions manifests in molecular mimicry. J. Immunol. 191, 456–463. doi: 10.4049/jimmunol.1203260
Tateda, K., Ishii, Y., Horikawa, M., Matsumoto, T., Miyairi, S., Pechere, J. C., et al. (2003). The Pseudomonas aeruginosa autoinducer N-3-oxododecanoyl homoserine lactone accelerates apoptosis in macrophages and neutrophils. Infect. Immun. 71, 5785–5793. doi: 10.1128/iai.71.10.5785-5793.2003
Trott, O., and Olson, A. J. (2010). AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 31, 455–461.
Wagner, V. E., Bushnell, D., Passador, L., Brooks, A. I., and Iglewski, B. H. (2003). Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J. Bacteriol. 185, 2080–2095. doi: 10.1128/jb.185.7.2080-2095.2003
Waters, C. M., and Goldberg, J. B. (2019). Pseudomonas aeruginosa in cystic fibrosis: a chronic cheater. Proc. Natl. Acad. Sci. U.S.A. 116, 6525–6527.
Wing, M. G. (1995). The molecular basis for a polyspecific antibody. Clin. Exp. Immunol. 99, 313–315. doi: 10.1111/j.1365-2249.1995.tb05551.x
Wyllie, A. H., Kerr, J. R., and Currie, A. R. (1980). Cell death: the significance of apoptosis. Int. Rev. Cytol. 68, 251–306. doi: 10.1016/s0074-7696(08)62312-8
Xu, D., and Zhang, Y. (2011). Improving the physical realism and structural accuracy of protein models by a two-step atomic-level energy minimization. Biophys. J. 101, 2525–2534. doi: 10.1016/j.bpj.2011.10.024
Yang, J., Yan, R., Roy, A., Xu, D., Poisson, J., and Zhang, Y. (2015). The I-TASSER suite: protein structure and function prediction. Nat. Methods 12, 7–8. doi: 10.1038/nmeth.3213
Keywords: Pseudomonas aeruginosa, quorum sensing, N-3-oxo-dodecanoyl-L-homoserine lactone (3O-C12-HSL), apoptosis, human scFv
Citation: Santajit S, Seesuay W, Mahasongkram K, Sookrung N, Pumirat P, Ampawong S, Reamtong O, Chongsa-Nguan M, Chaicumpa W and Indrawattana N (2020) Human Single-chain Variable Fragments Neutralize Pseudomonas aeruginosa Quorum Sensing Molecule, 3O-C12-HSL, and Prevent Cells From the HSL-mediated Apoptosis. Front. Microbiol. 11:1172. doi: 10.3389/fmicb.2020.01172
Received: 18 December 2019; Accepted: 07 May 2020;
Published: 24 June 2020.
Edited by:
Marco Rinaldo Oggioni, University of Leicester, United KingdomReviewed by:
Song Lin Chua, The Hong Kong Polytechnic University, Hong KongCopyright © 2020 Santajit, Seesuay, Mahasongkram, Sookrung, Pumirat, Ampawong, Reamtong, Chongsa-Nguan, Chaicumpa and Indrawattana. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nitaya Indrawattana, bml0YXlhLmluZEBtYWhpZG9sLmFjLnRo
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.