 Kevin Felipe Martho1
Kevin Felipe Martho1 Otávio J. B. Brustolini2
Otávio J. B. Brustolini2 Ana Tereza Vasconcelos2
Ana Tereza Vasconcelos2 Marcelo A. Vallim1
Marcelo A. Vallim1 Renata C. Pascon1*
Renata C. Pascon1*- 1Department of Biological Sciences, Campus Diadema, Universidade Federal de São Paulo, São Paulo, Brazil
- 2Laboratório Nacional de Computação Científica – LNCC, Labinfo – Laboratório de Bioinformática, Petrópolis, Brazil
Cryptococcus neoformans is an opportunist fungal pathogen that causes meningoencephalitis in immunocompromised patients. During infection, this basidiomycete yeast has to adapt to several adverse conditions, especially nutrient availability. The interruption on various amino acid biosynthetic pathways and on amino acid uptake causes reduced viability, inability to cope with various stresses, failure in virulence factors expression and avirulence in animal model of infection. The sulfur amino acid biosynthesis and uptake is an important feature for pathogen survival in vivo and in vitro. Our previous work demonstrates that C. neoformans Cys3 BZip transcription factor controls the gene expression in several steps of the sulfur assimilation and sulfur amino acid biosynthesis. Also, we have shown that Gpp2 phosphatase modulates Cys3 activity. In Saccharomyces cerevisiae Gpp2 is induced in response to hyper osmotic or oxidative stress and during diauxic shift. In this work, we will show that, in C. neoformans, Gpp2 is required to respond to stresses, mainly osmotic stress; also its transcription is induced during exposure to NaCl. Global transcriptional profile of gpp2Δ by RNAseq shows that CYS3 and other genes in the sulfur assimilation pathway are up regulated, which is consistent with our previous report, in which Gpp2 acts by avoiding Cys3 accumulation and nuclear localization. In addition, several transporters genes, especially amino acid permeases and oxidative stress genes are induced in the gpp2Δ strain; on the contrary, genes involved in glucose and tricarboxylic acid metabolism are down regulated. gpp2Δ strain fails to express virulence factors, as melanin, phospholipase, urease and has virulence attenuation in Galleria mellonella. Our data suggest that Gpp2 is an important factor for general pathogen adaptation to various stresses and also to the host, and perhaps it could be an interesting target for therapeutic use.
Introduction
Fungal pathogens can cause severe systemic mycoses which are difficult to treat (Garcia-Vidal et al., 2013). One of these pathogens, Cryptococcus neoformans, causes fatal meningitis in immunocompromised patients (Powderly, 1993; Limper et al., 2017). This basidiomycete yeast is commonly found in bird excreta and decomposing wood, but it can also colonize the animal host (Kronstad et al., 2012; Ballou and Johnston, 2017; Esher et al., 2018). In order to survive the transition from natural environment to the human host it must adapt quickly to changes, among which, nutrient and osmotic adaptation are essential to survival (Kronstad et al., 2012; Rutherford et al., 2019).
Our previous work showed that nutrient availability is important for virulence and stress resistance. Interruptions on amino acid biosynthesis or failure to uptake amino acids by permeases lead to growth arrest, defects in virulence factor and decreased virulence in mouse and Galleria mellonella animal model of infection (Yang et al., 2002; Pascon et al., 2004; Fernandes et al., 2015; Martho et al., 2016; Calvete et al., 2019). Recently, we showed the regulation of sulfur amino acid biosynthesis is essential for C. neoformans survival in vivo and in vitro (de Melo et al., 2019). Cys3 transcription factor, the major regulator of the sulfur uptake network, is part of a protein complex and physically interacts with calcineurin and Gpp2 phosphatase. In C. neoformans Cys3 is encountered in the nucleus at high protein levels during sulfur amino acid limitation. The addition of methionine and cysteine to the medium targets Cys3 protein to degradation. Calcineurin regulatory and catalytic subunits are important in order to maintain high levels of Cys3 in the nucleus and Gpp2 seems to be important to send Cys3 to degradation. Deletion of GPP2 gene causes abnormal accumulation of Cys3 protein (de Melo et al., 2019).
In Saccharomyces cerevisiae GPP2 is involved in glycerol biosynthesis (Norbeck et al., 1996). In this pathway a glycolytic intermediate, dihydroxyacetone-phosphate is converted into glycerol-3 phosphate by Glycerol-3-phosphate dehydrogenase encoded by GDP1 and GDP2 (Albertyn et al., 1994). GPP1 and GPP2 encode isoforms of glycerol-3-phosphate phosphatase; these proteins conduct redundant catalytic functions at the last step in this biosynthetic route, the conversion of glycerol-3-phosphate into glycerol (Larsson et al., 1993). GPP1 and GPP2 are induced under hyperosmotic shock, partially by the High Osmolarity Glycerol (Hog) pathway. GPP1, but not GPP2, is controlled by Protein Kinase A activity. The deletion of both genes renders yeast cells sensitive to oxidative stress (paraquat) and especially sensitive to osmotic shock (Pahlman et al., 2001).
Osmoadaptation is also controlled by the calcineurin complex in S. cerevisiae. In response to osmotic stress, Ca2+ stocks are released from the vacuole to the cytoplasm and functions as a second messenger that leads to dephosphorylation of Ena1 (P-type ATPase on the plasma membrane) facilitating Na and Li efflux, enabling yeast cells to grow on high levels of salt (Cunningham and Fink, 1994; Saxena and Sitaraman, 2016).
Calcineurin has been linked to osmotic stress and cell wall integrity response in C. neoformans (Kraus and Heitman, 2003; Fan et al., 2007). Also, the relationship between calcineurin complex and GPP2 identified in our previous work and others reinforces the idea that this phosphatase complex plays an important role on osmobalance (Park et al., 2016; Jung et al., 2018; de Melo et al., 2019).
In C. neoformans GPP2 (CNAG_01744) was first identified by a phosphoproteomics approach as a substrate of the calcineurin phosphatase and was later named HAD1 (Jung et al., 2018). It was hyper phosphorylated (two fold) when calcineurin was inactive (Park et al., 2016). C. neoformans GPP2 has been considered a target of the mitogen-activated protein kinase Hog1; it is more than two fold induced in wild type in response to osmotic shock and is repressed in a hog1Δ strain, suggesting that this gene is a homolog of S. cerevisiae glycerol-3-phosphate phosphatase (Ko et al., 2009). Collectively, these data indicate that GPP2 may be regulated by Hog1 and calcineurin.
In this work, we showed that a gpp2Δ strain is highly sensitive to cold shock, membrane, cell wall, alkaline and especially osmotic (NaCl and KCl) stresses. Proline and other amino acids are able to remediate growth arrest caused by osmotic stress. In addition, a global transcriptional profile of the gpp2Δ compared to wild type revealed that genes involved in oxidative stress, transmembrane transport and the sulfur amino acid biosynthetic network are induced in the mutant. The association of transcriptomic data and phenotypic analysis of the gpp2Δ strain suggests that, there is a connection between oxidative stress, sulfur uptake and osmotic stress response. Whereas, genes related to glucose and tricarboxylic acid metabolism and oxidation-reduction processes were repressed. These data lead us to verify the expression levels of amino acid permease genes, CYS3 and its target, SUL1, by quantitative real-time PCR.
Regarding virulence factors, the gpp2Δ strain is unable to produce phospholipase, urease and melanin. In vivo virulence assay in G. mellonella showed this strain is hypovirulent. In this paper, we propose that glycerol-3 phosphate phosphatase Gpp2 affects sulfur amino acid biosynthetic network, which in turn functions as a central hub which generates metabolites that are essential to counteract osmotic and oxidative stress. In connection with the virulence data this idea underlines the potential of the sulfur network for therapeutic use in C. neoformans.
Materials and Methods
Medium Composition, Growth Conditions and Materials
Supplementary Table 1 lists the primers used in this work. Growth of C. neoformans strains was carried out on YEPD (1% yeast extract, 2% bacto-peptone, 2% glucose); synthetic dextrose (SD) was made with yeast nitrogen base, YNB (0.67 g/L yeast nitrogen base with or w/o amino acid and ammonium sulfate, depending on experimental design, 2% glucose and 10 mM of each amino acid as sole nitrogen source); incubations were carried out at 22, 30 or 37°C on plates or on liquid medium with 150 rpm in a rotary shaker. Amino acids were supplemented at 10 or 20 mM depending on experimental design. Eugenol at 0.1 g/L (Sigma) was incorporated to the YEPD and SD.
Growth Rate on Amino Acids
Experiments to evaluate the growth on amino acid as sole nitrogen source was carried out on 96 well plates in 100 μL total volume of SD (YNB 1X), 2% dextrose and 10 mM of a single amino acid as sole nitrogen source, according to previous published protocol (Martho et al., 2016). In brief: fifteen amino acids were tested; cells were grown in YEPD at 30°C overnight, collected by centrifugation at 4000 rpm for 5 min and washed three times in sterile 1X PBS (Phosphate Buffered Saline). Intracellular nitrogen pools were exhausted by incubation of the washed cells in 1X PBS at 30°C with 150 rpm rotation for 2 h. After this period 200 cells were inoculated in each well containing a single amino acid as sole nitrogen source. All experiments were done in technical triplicates; plates were incubated at 30 and 37°C for 48 h. The OD600 was measured in a plate reader (Epoch 2 microplate reader, BioTek). A minimum of three biological replicates were done for all experiments. The following assay controls were used: inoculums were cultivated on medium with ammonium sulfate as nitrogen source (positive control) and without it (negative control) in the same condition described above. Statistical significance was calculated by one-way ANOVA with GraphPad Prism 7.0.
Strains and Genetic Manipulation
A reconstituted strain (CNU135) was created by introducing a PCR amplified wild type allele of GPP2 and a co-transforming plasmid pZPHyg (Idnurm et al., 2004) in CNU125 (gpp2Δ:NatR) described elsewhere (de Melo et al., 2019) by biolistic transformation (Toffaletti et al., 1993). Confirmation of the genetic modification was done by colony PCR and Southern blot (Supplementary Figure 1). All strains were constructed under the H99 background; therefore, it was used as the wild type control.
In vitro and in vivo Virulence Tests
Stress tolerance and virulence factor analysis were done with wild type, gpp2Δ:NatR (CNU125 and CNU126) and reconstituted (CNU135) strains. Thermo tolerance was evaluated in rich medium YEPD and SD, with or without ammonium sulfate and amino acids at temperatures of 22, 30, and 37°C. Urea agar was used to determine urease activity (Christensen, 1946). Melanin and phospholipase were assayed according to previously published protocol at 30°C (Paliwal and Randhawa, 1978; Price et al., 1982). Capsule biosynthesis was induced in Sabouraud Broth diluted in MOPS (1:10) at 37°C in shaker (150 rpm) (Zaragoza and Casadevall, 2004). Samples for capsule size evaluation were collected at 24 h and stained with India ink and analyzed by light microscopy (Olympus BX51M). The measurements were performed in biological triplicates with the assistance of CellSens software (Olympus). The data was treated statistically using ANOVA (GraphPad Prism 7).
Multi-stress sensitivity was evaluated with YEPD or SD medium supplemented with 0.5, 0.75, and 1 M of NaCl or KCl. The cell wall and membrane integrity were evaluated on YEPD plus 0.5% Congo Red and YEPD plus 0.03% SDS, respectively. All plates were incubated at 22, 30 or 37°C depending on experimental design.
In vivo assays with G. mellonella were done according to previous published protocol (Mylonakis et al., 2005). In brief: mutant, wild type and complemented strains were inoculated into 5 mL of YEPD and incubated with orbital agitation 150 rpm for 16–18 h. Subsequently, suspensions were collected by centrifugation, washed twice in sterile 1X PBS and adjusted to 1 × 106 cell/mL in PBS supplemented with ampicillin (20 mg/kg body weight). Groups of 16 caterpillars with 200 mg of average weight were inoculated with 10 μL of the suspension with a Hamilton syringe in the last pro-paw. Thereafter, caterpillars were separated on glass Petri dishes (15 mm diameter) and incubated at 30 and 37°C during 8 days. They were monitored daily by observing spontaneous or induced movements with sterilized tweezers. The experiment was completed when the larvae died or formed cocoons.
qPCR
Total RNAs were obtained from wild type and mutant strains incubated overnight in liquid YEPD under 150 rpm agitation at 30°C. RNA extraction protocol was described before (Calvete et al., 2019). cDNA synthesis was done with 5 μg of total RNA, with RevertAid H minus First Strand cDNA synthesis kit (Thermo Scientific), Oligo dT and random hexamer primers. Real time PCR amplifications were made from diluted cDNA templates (1:10) with 600 nM target primers, 300 nM GPDH1 (Glyceraldehyde-3-phosphate dehydrogenase) internal control primers, and 1X SYBR Green (EvaGreen®). Quantification of the transcript levels was performed in StepOne thermo cycler (Applied Biosystems), normalizing gene was done with GPDH1 according to Livak and Schmittgen (2001). An analysis of variance was performed by Tukey’s multiple comparison test using GraphPad Prism 7.0 software, and p values lower than 0.05 were considered statistically significant.
Transcriptional Profile: RNA Extraction, Quantification, and RNAseq Analysis
All RNAseq experiments were performed in triplicates. Strains (mutant and wild type) were grown overnight in YEPD at 30°C with 150 rpm rotation. After this period, the cells were induced in liquid YEPD for 2 h (30°C) at 150 rpms. Total RNA was extracted as described above. RNA quantification, purity and quality were evaluated in NanoDrop spectrophotometer (Thermo Scientific, Waltham, MA, United States) and RNA Nano 6000 Assay Kit of the Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, United States). Libraries were made from 4 μg of total RNA according to instructions of the Illumina TruSeq Stranded mRNA Sample Prep LS Protocol. Library quantifications were performed by qPCR using the KAPA Illumina qPCR Quantification Kit. Quantification and validation of the libraries were done by quantitative PCR and they were diluted at the working concentration (2 ηM); libraries were denatured with sodium hydroxide solution at 0.1 N and then diluted to 20 (M and loaded into HiSeq sequencer with the v4 sequencing kit (2 × 100 cycles). Sequencing was performed at Centro de Genômica Funcional – ESALQ/USP, Piracicaba, Brazil.
The raw fastq files were filtered by the software BBDuk21 which is capable of quality-related trimming and filtering adapter-contaminant. Any Illumina adapters and reads with low quality (bellow Q30) were removed. The FASTQC2 created the overall quality of the sequencing reports. The filtered reads were mapped into the C. neoformans H99 genome sequence using the software STAR aligner (Dobin et al., 2013). The genome and annotation files were retrieved from the FungiDB database3. The R/Bioconductor package Rsubread/featureCounts (Liao et al., 2019) made the counting table for the posterior statistical analysis. The Differential Gene Expression (DGE) test was performed by the R/Bioconductor package DESeq2 (Love et al., 2014) with the method of shrink log-fold changes presented at the package apeglm (Zhu et al., 2018). The genes with adjusted p-value (corrected by false discovery rate – FDR) below 0.05 and log2 fold change above or below 1.0, were considered as differentially expressed (DEG). The Gene Ontology (GO) enrichment analysis was performed by the R/Bioconductor package GOstats (Falcon and Gentleman, 2007). The package Pathview (Luo and Brouwer, 2013) was used to visualize the KEGG pathways. The protein-protein interaction network was created using the STRING database (Szklarczyk et al., 2017).
Results
C. neoformans CNAG_01744 Encodes a Glycerol-3-Phosphate Phosphatase (GPP2)
Cryptococcus neoformans CNAG_01744 was identified in our earlier research by immune precipitation as part of a protein complex with the major sulfur amino acid biosynthesis transcription regulator Cys3 and the calcineurin catalytic and regulatory subunits Cna1 and Cnb1, respectively (de Melo et al., 2019). We named this gene as GPP2 based on the amino acid sequence similarity to S. cerevisiae GPP2, a glycerol-3-phosphate phosphatase (Pahlman et al., 2001).
Previously, this gene was also found in C. neoformans by phosphoproteomics approach as substrate for the calcineurin complex (Park et al., 2016) and later it was named as HAD1 (Halo Acid Dehalogenase hydrolase) due to the predicted molecular function (Jung et al., 2018). C. neoformans bears four genes that encode Halo Acid Dehalogenase hydrolase-like proteins (CNAG_06132, CNAG_01744, CNAG_6122, and CNAG_06698). BLAST similarity searches and amino acid sequence alignments indicated that CNAG_01744, CNAG_06122, and CNAG_06132 may be homologs of S. cerevisiae Gpp1 and Gpp2 (glycerol-3-phosphate phosphatase), which can also be classified as a HAD-like dehalogenase. However, the percentage of identity among these amino acid and S. cerevisiae GPP2 sequences are very close (29.9, 31.1, and 31.4% identity, respectively), suggesting that sequence similarities may not be the best tool to indicate which one of these genes is a homolog of Gpp2. Alternatively, glycerol-3-phosphate phosphatase (Gpp2) may be encoded by more than one gene, as it is the case in S. cerevisiae, in which GPP1 and GPP2 encode very similar proteins, that have the same catalytic function, but are differentially regulated according to the environmental condition (Pahlman et al., 2001). In this paper, among other data, we will present phenotypic features of gpp2Δ which suggest its role as an important genetic element involved in osmotic stress response. Also, we will refer to CNAG_01744 as GPP2.
GPP2 Is Involved in Cold Shock and Nutritional Stress
We tested several stress phenotypes associated to gpp2Δ strain relative to wild type and a reconstituted strain (CNU135) which was constructed in this work (Supplementary Figure 1). As shown in Figure 1A, deletion of GPP2 did not significantly changed growth in rich medium (YEPD) at 30 and 37°C. However, cold shock (22°C) caused severe growth arrest in gpp2Δ strains relative to wild type and reconstituted strains in YEPD (Figure 1A). There was also, growth deficiency in SD supplemented with the preferred nitrogen source (ammonium sulfate) and/or amino acids (Figure 1B), suggesting that, in this condition, the mutant strain may have some sort of nutritional deficiency. Interestingly, growth arrest in SD was reverted in the presence of 10 mM of proline as sole nitrogen source (Figure 1B).

Figure 1. Growth pattern of the gpp2Δ mutant (CNU125 and CNU 126) compared to wild type (H99) and reconstituted strain (CNU135) in (A) YEPD and three different temperatures (22, 30, and 37°C); (B) the same strains on Synthetic dextrose (SD) supplemented with ammonium sulfate and amino acids (tryptophan, histidine, and methionine), SD with ammonium sulfate only and SD supplemented with 10 mM of proline as the sole nitrogen source at 30°C. Spot dilutions are 1 × 105 to 1 cell at the last spot.
This result led us to investigate if other amino acids could rescue the low growth phenotype in SD. Table 1 shows the growth rate of two mutants compared to wild type in single amino acid as sole nitrogen source at 30 and 37°C in liquid medium. Besides proline, methionine, glutamine, asparagine, and aspartate could complement the growth defect in SD as sole nitrogen source, yielding growth rates that are not statistically different (NS) than wild type in the same amino acid as sole nitrogen source. It is noteworthy to mention that glutamic acid, raised growth rate to more than 70% compared to wild type in spite of a significant statistical difference between mutant and wild type (p < 0.001). It is important to mention that all these amino acids, except methionine, support growth in C. neoformans wild type to the same rate as the preferred nitrogen source (ammonium sulfate). Leucine as sole nitrogen source also improved the growth rate of the mutants to wild type levels (CNU125 100% and CNU126 97%, NS), but only at 37°C.
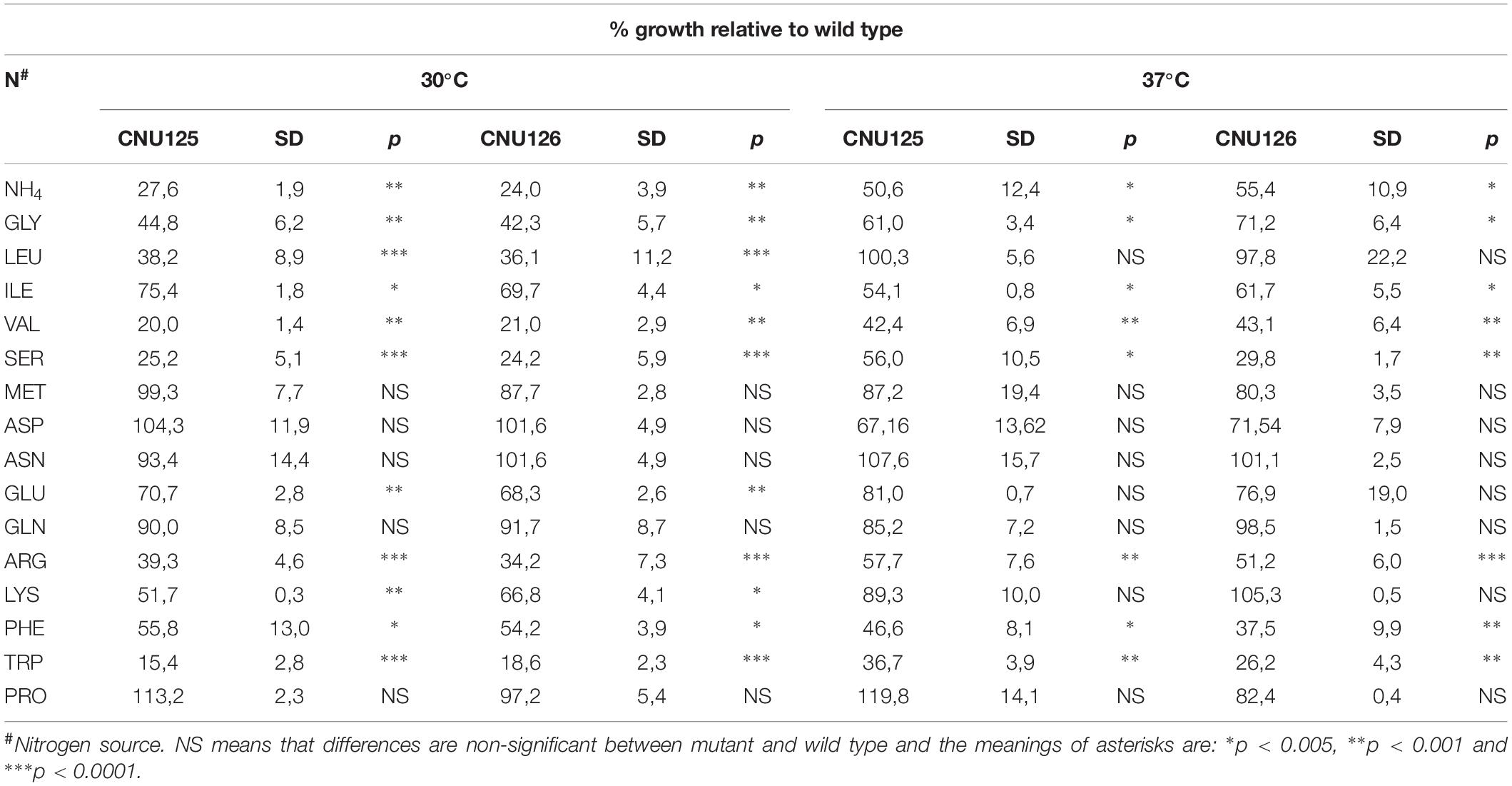
Table 1. The growth percentage of two independent gpp2Δ strains (CNU125 and CNU126) in 10 mM of each amino acid relative to wild type at 30°C and 37°C.
GPP2 Plays an Important Role in Membrane, Cell Wall and Osmotic Stress Responses
We confirm that GPP2 is required for membrane and cell wall integrity as showed before (Jung et al., 2018). As expected, we observed that our mutants are more sensitive to Congo red and SDS (Figure 2A). In addition, we also found out that gpp2Δ strains are more sensitive to alkaline stress (Figure 2B). Growth of gpp2 mutant was compared to wild type in the presence of hydrogen peroxide and only a slight difference was detected in YEPD. On SD, mutant and wild type were highly sensitive to even small concentrations of H2O2 (Supplementary Figure 2).

Figure 2. Growth rate of gpp2Δ mutant (CNU125 and CNU 126) compared to wild type (H99) and reconstituted strain (CNU135) in (A) YEPD, YEPD supplemented with 0.03% of SDS and YEPD supplemented with 0.5% of Congo red. (B) Growth rate of the same strains in different pHs (6.4, 7.0, and 8.0) at 30 and 37°C and (C) growth rate of the same strains in the presence of osmotic stress (0.75 and 1 M of NaCl and KCl) in YEPD and SD supplemented with the preferred nitrogen source; (D) growth rate of gpp2Δ mutants, wild type and reconstituted strain in SD, SD supplemented with 0.5 M NaCl and SD plus 0.5 M NaCl and 20 mM of proline (left panel), the same strains in YEPD, YEPD supplemented with 0 M NaCl and YEPD plus 0.5 M NaCl and glycerol (right panel); (E) effect of the temperature (22°C) on wild type, reconstituted and mutant strains in YEPD with and without glycerol. Spot dilutions are 1 × 105 to 1 cell at the last spot. (F) qPCR showing the induction of GPP2 transcript in the presence of 1 M NaCl relative to the control YEPD in wild type strain H99. Statistical significant differences were tested by Two-way ANOVA, ∗∗∗p < 0.001.
Saccharomyces cerevisiae Gpp2 is partially regulated by the Hog1 and is deeply involved in osmotic stress response (Norbeck et al., 1996; Pahlman et al., 2001). Therefore, we tested the hypothesis that gpp2Δ strains are hypersensitive to osmotic stress agents (NaCl and KCl) in rich YEPD and SD medium. As shown in Figure 2C, CNU125 and CNU126 have progressive growth impairment as the concentration of NaCl and KCl increases in YEPD and SD (0.75 and 1 M in YEPD and SD).
Since proline was able to rescue growth in SD as sole nitrogen source (Figure 1 and Table 1), we tested if it could also have any effect on growth rate in the presence of osmotic stress. As shown in Figure 2D, 20 mM proline as sole nitrogen source (but not 10 mM) was enough to relief the effects of 0.5 M of NaCl on growth in SD. The supplementation of glycerol was also sufficient to relief growth arrest caused by osmotic and cold shock (Figure 2E) in the gpp2Δ mutants. Since SD is a salt medium, it is possible that it represents an osmotic unbalanced condition for the gpp2 mutant and that is the reason why 10 mM of proline was enough to overcome growth arrest in SD, but not in SD plus 0.5 M NaCl. An increase concentration of proline (20 mM) was necessary to overcome the osmotic stress in SD medium supplemented with 0.5 M of NaCl. Proline was also able to suppress some of the osmotic effects of NaCl in YEPD, but not comparable to SD, probably because amino acid transport is more efficient in SD than YEPD, due to relief of nitrogen catabolite repression on permeases (Fernandes et al., 2015).
These results reinforce the idea that this gene encodes the glycerol-3-phosphate phosphatase (GPP2), which is responsible for generating glycerol in S. cerevisiae, an important osmolyte that helps the cell to overcome osmotic and cold stress. In the absence of glycerol, C. neoformans can compensate osmotic unbalance with proline.
In addition to the osmotic and cold shock phenotypes associated to gpp2 mutant, we tested if the transcription of GPP2 would be induced in the presence osmotic stress. Figure 2F shows that GPP2 is induced 7.1 fold in YEPD + 1 M NaCl relative to YEPD in H99. This result and the phenotypic characterization strongly indicate that the glycerol phosphate phosphatase encoded by GPP2 is involved in osmotic stress and probably is the homolog of S. cerevisiae GPP2.
GPP2 Is Important for Virulence in C. neoformans
Regarding the virulence factors, the gpp2Δ strains were unable to produce urease (Figure 3A), phospholipase (Figure 3B) and melanin (Figure 3C) compared to wild type and the reconstituted strain. However, capsule production was not significantly different from wild type (Figure 3D). gpp2Δ strain (CNU125) was hypovirulent in G. mellonella animal model (Figure 3E) which is consistent with the data generated by Jung in mouse model of infection (Jung et al., 2018).
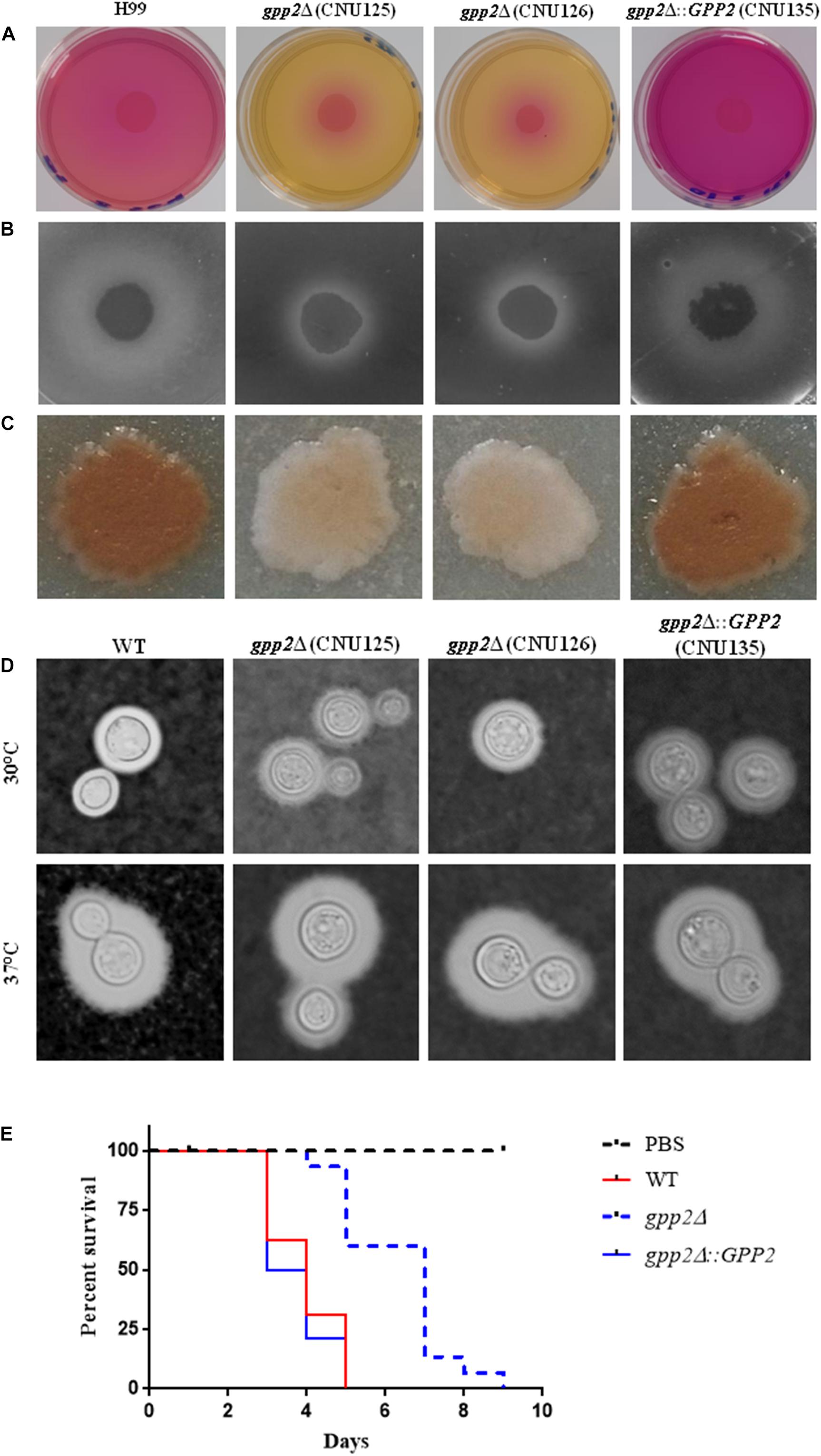
Figure 3. Virulence factor evaluation of the gpp2 mutant compared to wild type and reconstituted strains. (A) urease, (B) phospholipase, (C) melanin, (D) capsule, and (E) invertebrate animal model in G. mellonella.
Global Transcriptional Profile of the gpp2Δ Strain
In order to draw a broader picture on how GPP2 affects cellular metabolism, amino acid transport/biosynthesis, stress and the sulfur amino acids biosynthesis, the global transcription profile of gpp2Δ strain was done by RNAseq and compared to wild type. As a validation control of the experiment we detected that CNAG_01744 (encoding GPP2) is not expressed in the mutant, as expected. A total of 205 DEGs (Differentially Expressed Genes) were up regulated and 129 were down regulated in the mutant relative to wild type (Supplementary Tables 2, 3) considering a 1.0 log2Fold change as cutoff. Among the DEGs listed, many encode hypothetical proteins (126 up regulated 66 down regulated). Transport and sulfate assimilation are the main biological processes and molecular functions pointed out by GO analysis that appeared as up regulated two fold or more (Table 2 and Figures 4A,B and Supplementary Table 2).
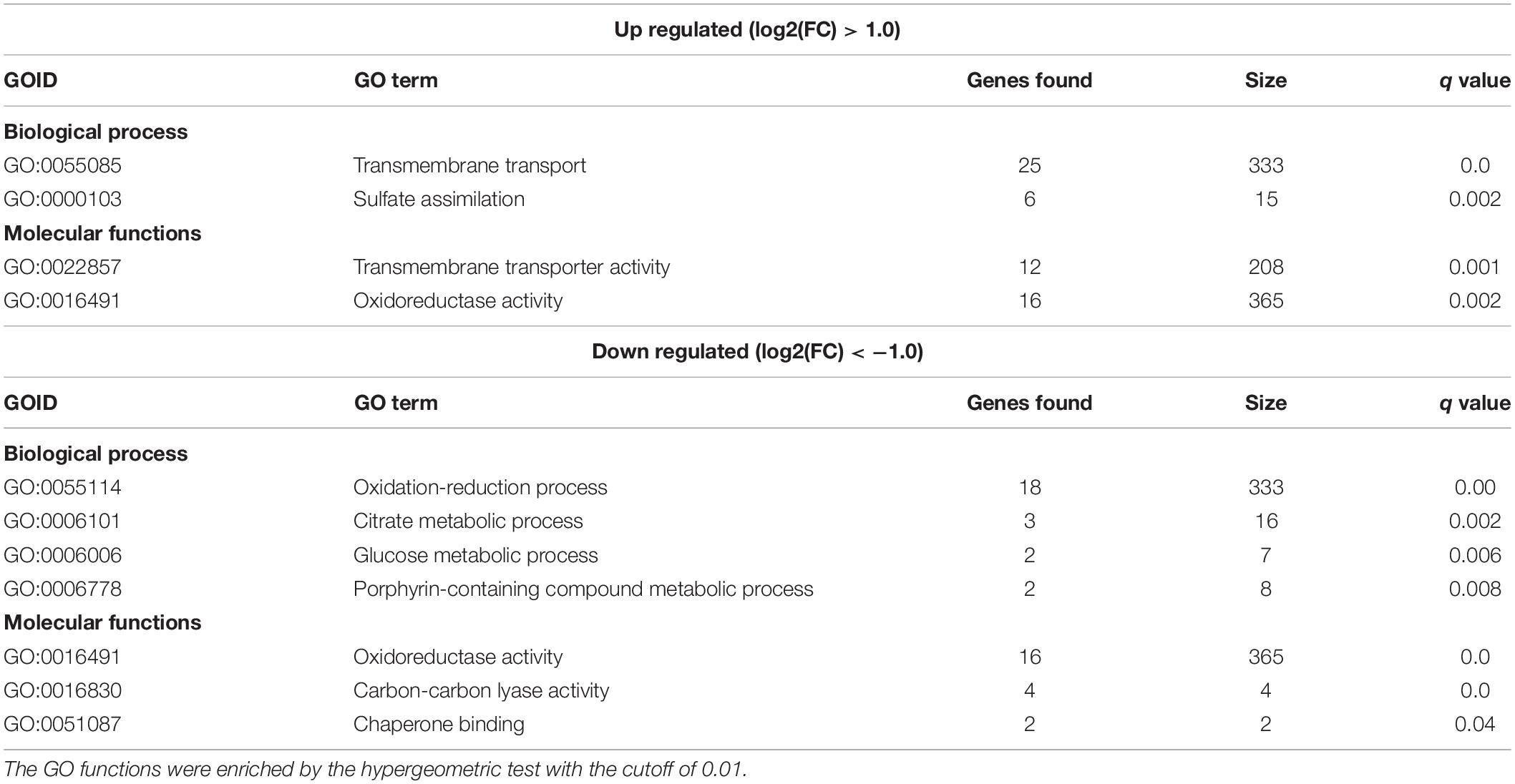
Table 2. List of the biological processes and molecular functions found to be differentially expressed (up and down regulated), according to Gene ontology (GO), in gpp2Δ mutant relative to wild type.
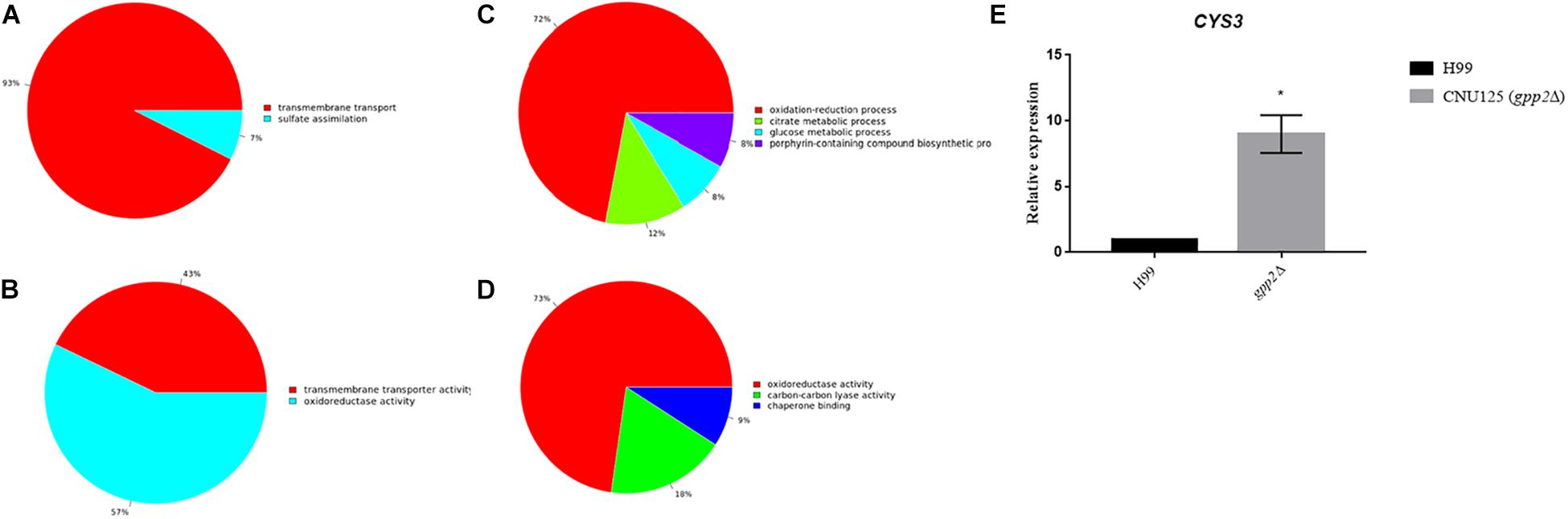
Figure 4. Graphs represent the DEGs found to be up (A,B) and down (C,D) regulated according to GO categories based on biological processes (A up regulated and C down regulated) and molecular function (B up regulated and D down regulated). The cutoff used was 1.0 Log2. (E) Graph shows CYS3 gene quantification by Real time qPCR in gpp2Δ strain relative to wild type. RNA was extracted from cultures growing in YEPD for 2 h at 30°C.
Among the transporters, sulfite (CNAG_00529), pantothenate (CNAG_00540), myo-inositol transporters (CNAG_00864 and CNAG_00867), amino acids permeases (CNAG_00597, 00728 and 01118), ion (CNAG_00979), phospholipid (CNAG_01055), and sugar transporters (CNAG_01683, 03772, 04474, and 06963) are examples of the most induced genes found in this analysis. Also, a major facilitator transporter super family gene (MFS, CNAG_06610) was up regulated. If we consider less than two fold change (0.5 log2Fold change) the number of induced transporters rises to 70 DEGs up regulated on the same categories described above. CNAG_01683, which was found to be 1.16 fold induced (log2), encodes a protein that shares 49% of amino acid sequence similarity to S. cerevisiae Stl1, a glycerol/H+ symporter that is thought to play a major role on glycerol transport. Its transcription is induced in the presence of glycerol and osmotic shock and it is repressed on glucose (Ferreira et al., 2005). The up regulation of these transporter genes in the gpp2Δ strain relative to the wild type indicates that small molecule translocation, such as amino acids and inositol, may function as a compensatory mechanism to the lack of glycerol, an important osmolyte.
A total of six genes of sulfur amino acid biosynthetic pathway (CNAG_02202, 03898, 04215, 04798, 06448, 02270), plus a sulfite transporter (CNAG_00529) and the arylsulfatase (CNAG_01498) were also induced in the mutant. MET17, encoding the cysteine synthase (CNAG_05028), was induced, however, less than two log2Fold change (0.66 fold induction p = 2.20 × 10e–08). These results are in agreement with our published data, in which, the deletion of GPP2 causes Cys3 protein to accumulate in higher levels compared to wild type, suggesting that Gpp2 is involved in the Cys3 degradation and consequently down regulation of the sulfur assimilation network (de Melo et al., 2019). In order to validate our transcriptome data regarding the up regulation of the sulfur uptake pathway in the GPP2 mutant, we calculated the expression level of CYS3 in CNU125 (gpp2Δ) by RT-qPCR. We found that Cys3 is up regulated nearly 8.9 fold in the mutant compared to wild type (Figure 4E). These results show that lack of GPP2 not only modulate CYS3 at the post translational levels, but also changes gene regulation on the sulfur assimilatory circuit.
Among the down regulated DEGs, most of them are related to oxidation and reduction reaction, tricarboxylic, glucose and antibiotic metabolism (Table 2 and Figures 4C,D and Supplementary Table 3). This result is consistent with the role of Gpp2 as an important phosphatase involved in glycerol biosynthesis, which is not only important for osmoregulation, but also it is a key player in glycolysis and tricarboxylic acid metabolism (Possik et al., 2017).
Genes related to oxidative stress response were differentially regulated in the gpp2Δ, among them, the most induced are: catalase 2 (CNAG_05256 3.0 log2 Fold change), mitochondrial cytochrome C peroxidase (CNAG_01138 2.12 log2 Fold change) and glutathione transferase (CNAG_03848, 1.16 log2 Fold change).
Interestingly, three taurine dioxygenases (TauD) were induced in the mutant (CNAG_01542 and CNAG_06249 and CNAG_06876 at 4.0, 0.91 and 0.72 log2Fold change, respectively). This result suggests that the deletion of GPP2 not only derepresses sulfur uptake regulatory network through CYS3 expression, but also may induce the mobilization of sulfite from sulfonates, such as taurine. In S. cerevisiae, JLP1 is the only α-ketoglutarate dioxygenase known to release sulfite from isethionate and taurine (Hogan et al., 1999; Holt et al., 2017). Orthologs of JLP1 are genome linked to a predicted sulfur/sulfonate transporter in filamentous Eurotiomycete and Basidiomycota (Kensche et al., 2008). In S. cerevisiae the sulfur/sulfonate transporter is encoded by Soa1 gene, however, this gene pairing arrangement was lost in Ascomycetes (Holt et al., 2017). We analyzed if this genomic organization would be present in C. neoformans as well. Indeed, on chromosome 11 the arylsulfatase (CNAG_01498), sulfonate/sulfur transporter (CNAG_01499) and two taurine dioxygenases (CNAG_01500 and CNAG_01542) are clustered. Except for CNAG_01500, all other genes in this cluster are up regulated in the gpp2Δ strain.
It is important to mention that CNAG_01501, which is located between the two taurine dioxygenases genes, encodes a gamma-glutamyl cycle transferase (ChaC) involved in the biosynthesis of 5-oxoproline, an intermediate in the glutathione cycle (Kumar et al., 2012). This gene is not differentially regulated in gpp2Δ, but taken together these results suggests a connection between the osmotic stress, sulfur uptake and the glutathione cycle.
Amino Acid Permeases Regulation in gpp2Δ Strain
These results prompted us to validate the expression profile of the amino acid permeases in the gpp2 mutant. qPCR was applied to quantify the expression of the AAP genes, MUP1, and MUP3 in YEPD and during nitrogen starvation. The graph in Figure 5A shows that permease genes AAP4, AAP5, and AAP6 are significantly induced. During nitrogen starvation the expression level, in general, was higher compared to YEPD; notably permeases AAP2, AAP4, AAP5, AAP6, AAP8, and MUP1 had the highest induction (Figures 5A,B). The higher expression in SD compared to YEPD is due to the removal of nitrogen catabolite repression, which is known to induce permease gene expression (Fernandes et al., 2015; Martho et al., 2016; Calvete et al., 2019).
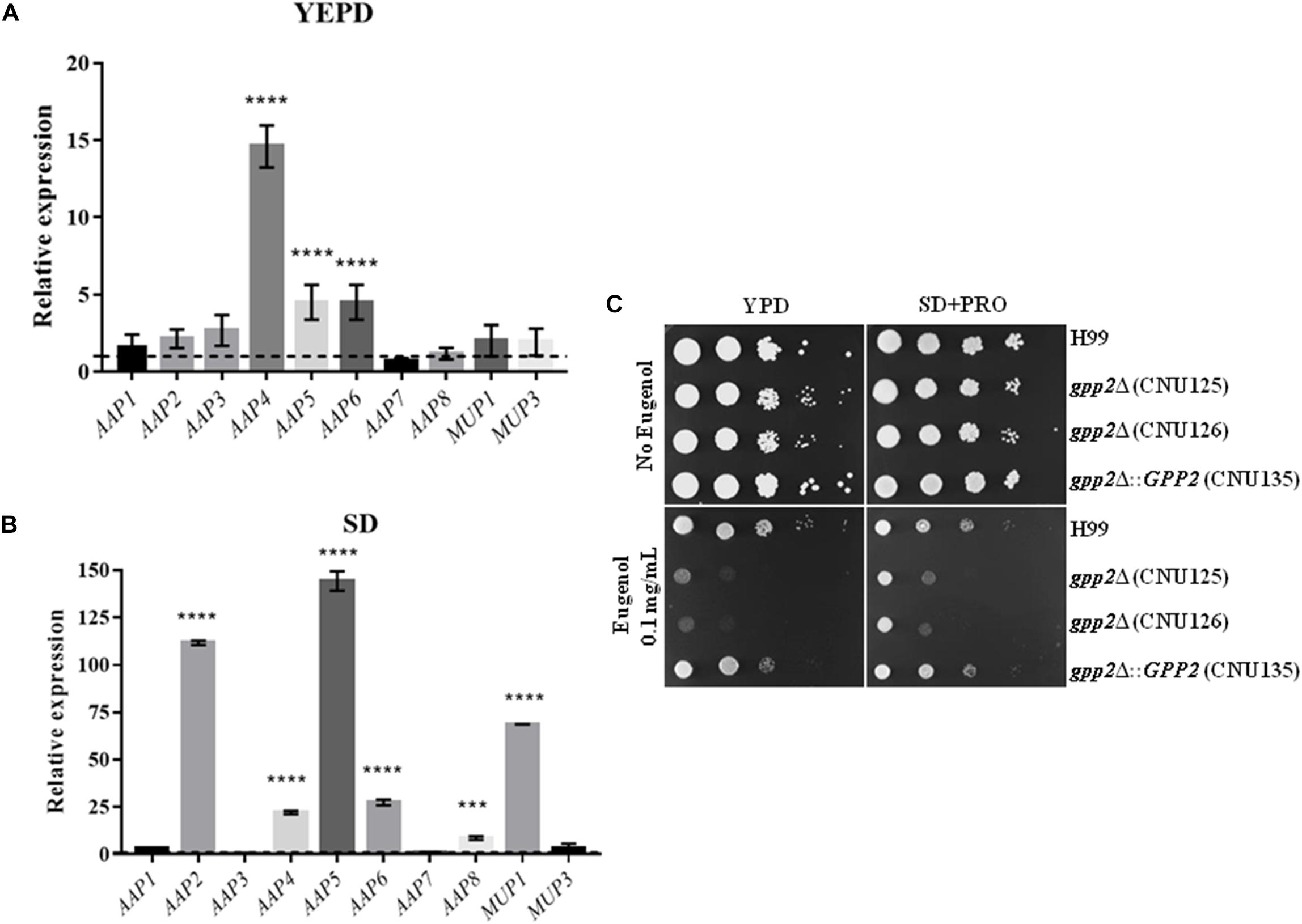
Figure 5. Expression of permease genes in the gpp2Δ mutant relative to wild type in (A) YEPD and (B) SD during nitrogen starvation by qPCR. Dotted line represents the expression level of the reference strain (wild type). Statistical significant differences were tested by Two-way ANOVA, ∗∗∗∗p < 0.0001 and ∗∗∗p < 0.001. (C) Growth of wild type, reconstituted (CNU135) and mutant strains (CNU125 and CNU 126) in YEPD without and with 0.1 g/L of eugenol (left panel) and the same strains in SD plus proline without and with 0.1 g/L eugenol (right panel).
Earlier, our group and other identified the amino acid permeases as the target of Eugenol, a natural product extracted from cloves that has antimicrobial activity (Darvishi et al., 2013; Martho et al., 2016). The deletion of permeases AAP4 and AAP5 renders C. neoformans resistant to eugenol, therefore, we reason that the up regulation of permease genes in gpp2Δ strain would confer sensitivity to eugenol, since its targets are over represented. As shown in Figure 5C, the gpp2Δ mutants are more sensitive to eugenol than wild type and reconstituted strains, as we predicted. The addition of proline to the medium was not able to compete with eugenol and promote a better growth.
At this point our data suggest that GPP2 is essential for glycerol generation, which seems to be the main osmolyte in C. neoformans. Without glycerol, cells have to uptake amino acids to achieve osmotic balance. The RNAseq data also suggest that lack of glycerol biosynthesis, and consequently the inability to cope even with small amounts of osmotic stress (SD medium), leads to activation of sulfur amino acid biosynthesis and oxidative stress response. These two metabolic pathways are deeply connected, since cysteine feeds the glutathione cycle, which is required to counter act oxidative stress. Since glycerol is the main osmolyte, sulfur amino acid biosynthesis and glutathione cycle would be required only in the absence of glycerol biosynthesis such as in the gpp2Δ strain. It is possible that sulfur amino acid and glutathione biosynthesis are not induced in the presence of osmotic stress, as long as glycerol is available in the cell. In order to test this hypothesis the expression levels of CYS3 and SUL1 were calculated in the presence of 1 M of NaCl in wild type. Interestingly, the expression levels of CYS3 and SUL1 were slightly down regulated (30% each) in the presence of osmotic stress as shown in Figures 6A,B, respectively, suggesting that, in fact, sulfur uptake network may be required only if glycerol is not available to counteract osmotic stress. However, further investigation will be necessary develop and confirm this hypothesis.
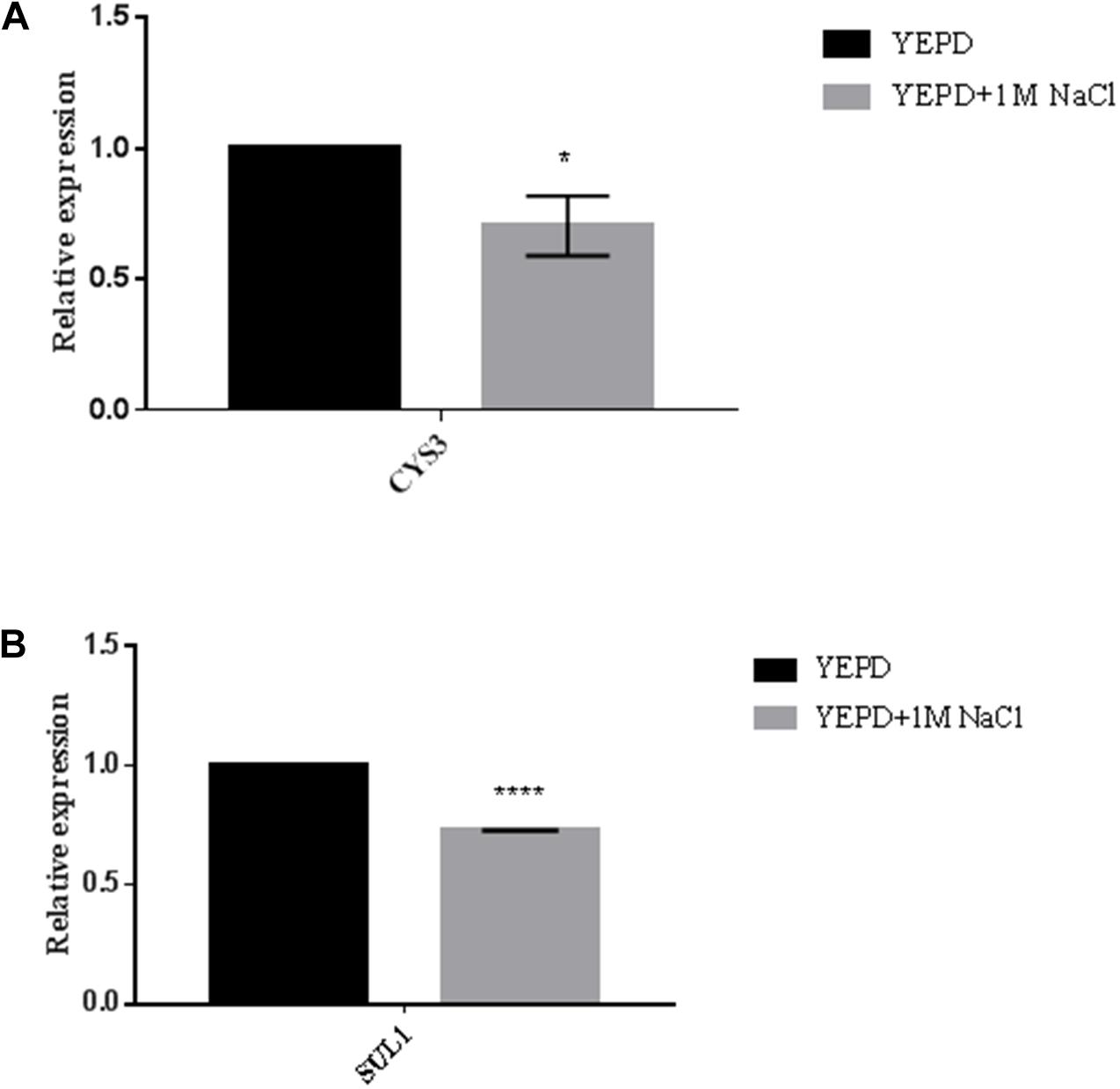
Figure 6. Expression pattern of CYS3 (A) and SUL1 (B) in wild type strain under osmotic stress by qPCR. YEPD was supplemented with 1 M of NaCl. Statistical significant differences were tested by Two-way ANOVA, ∗∗∗∗p < 0.0001 and ∗p < 0.05.
Discussion
The ability to adapt to sudden changes is a challenge encountered by many living organisms, specially microbes and plants. Since fungal pathogens occupy many different niches, they are vulnerable to nutritional, oxidative, cell wall and various types of stresses (Kronstad et al., 2012). Several of the genetic circuits which are triggered by these signals cross talk to each other eliciting responses that regulate many different genes producing a physiological conditions that allows survival even in most adverse conditions (Lin and Heitman, 2006; Brown et al., 2007; Lin, 2009; Li and Mody, 2010).
In this paper, we have explored several phenotypes of C. neoformans survival and virulence associated to the deletion of the glycerol-3-phospate phosphatase, which is responsible to carry the last step in the biosynthesis of glycerol, a major cellular osmolyte that helps the cells to counter act the effects of osmotic and cold shock (Norbeck et al., 1996). Also, we identified a link between GPP2 and the transcription of the sulfur amino acid biosynthetic pathway.
C. neoformans encodes 4 proteins with features of the Halo Acid Dehalogenases (HAD). In our previous work we identified one of them (CNAG_01744) and named it as GPP2 (de Melo et al., 2019). Among other reasons, this gene was designated so, due to its role on osmotic stress, which resembles phenotypes related to S. cerevisiae GPP2. Jung et al., also studied GPP2, but named it as HAD1 and described it as a genetic element involved in cell wall integrity (Jung et al., 2018); however, in S. cerevisiae HAD1 gene encodes an enzyme enrolled in the biosynthesis of nicotinic acid. Therefore, we maintained the name of the gene as GPP2 to avoid confusion.
Besides being transcriptionally induced by 1 M of NaCl, GPP2 is essential to counteract osmotic stress, membrane, cell wall and alkaline stresses. In our work we identified GPP2 as primordial to tolerate cold shock (Figures 1, 2). In S. cerevisiae it is well know that the Hog pathway is responsible for coordinating cold shock response (Hayashi and Maeda, 2006; Panadero et al., 2006). Ko and collaborators identified GPP2 as a target of Hog1 pathway (Ko et al., 2009) in C. neoformans. In this manuscript, we report for the first time a mutant that is sensitive to cold and osmotic stresses.
Another interesting feature of the gpp2Δ mutant is the low growth rate in SD in the presence of the preferred nitrogen source (ammonium sulfate), suggesting a nutritional defect. Curiously, growth arrest could be reverted by several single amino acids, as sole nitrogen source (Figure 2 and Table 1). One explanation for the observation is that certain amino acids improve growth rates in the gpp2Δ strains because they serve as osmolytes that compensate the lack of glycerol. As shown in this paper, proline was able to revert osmotic shock caused by NaCl confirming that this amino acids act as compatible solute in C. neoformans gpp2Δ mutant. In bacteria, archaea, and eukarya, proline and other amino acids function as compatible solute, protecting the cells against the osmotic stress (Kempf and Bremer, 1998).
The global transcriptional profile showed that transmembrane transport of several substrates into the cell is up regulated in the gpp2Δ. Among these transporters, amino acid permeases are highly induced (Figures 4, 5), suggesting that indeed amino acids may, at least partially, contribute to counteract osmotic stress. This result is in agreement with a previous transcriptome analysis reported in the literature in which amino acid permeases are induced in a Hog1 mutant (Ko et al., 2009). Also, the positive role of amino acids on osmotic stress is well known in many microorganisms and plants (Peddie et al., 1994; Zaprasis et al., 2015; Zou et al., 2016) The fact that amino acids rescued growth and permeases are induced in the gpp2 mutant argues that amino acids are important compatible solutes that combat osmotic stress in C. neoformans.
These results suggest that there is a link between osmotic stress, sulfur amino acid biosynthesis and amino acid uptake, which is depicted as a model in Figure 7. Since some amino acids can be considered osmolytes, it makes sense that an interruption on the biosynthesis of an important osmoregulator (glycerol) may activate the biosynthesis and uptake of alternative molecules that could serve as osmolytes, as an attempt to provide osmobalance to the cell. One possibility is that, accumulation of the Gpp2 substrate (glycerol-3-phosphate) signals the biosynthesis or uptake of other substances that promote osmotic balance, such as amino acids, which are capable of bypassing the growth arrest caused by osmotic shock in the gpp2 mutant (Figure 7).
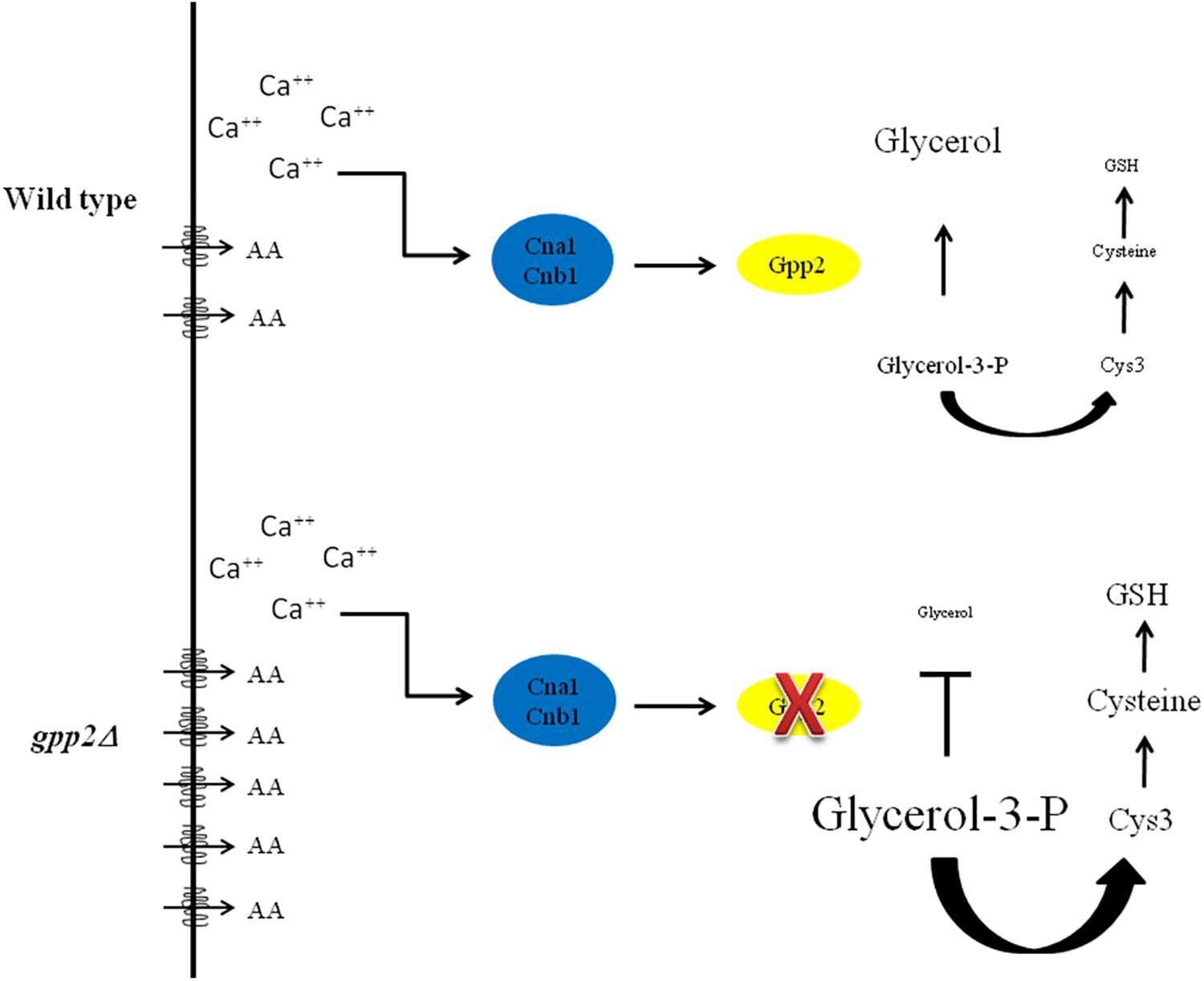
Figure 7. Model representing the main findings in this work. AA = amino acids; transmembrane drawing = amino acid permeases; Cna1 and Cnb1 = calcineurin catalytic and regulatory subunits respectively; GSH = glutathione.
The transcriptome analysis showed that oxidative stress response may be active in the gpp2Δ. In support of this finding, Ko and collaborators (Ko et al., 2009) detected an increase in the expression of genes related to this type of stress in C. neoformans hog1Δ. Also, in S. cerevisiae, GPP1, and GPP2 are both involved in oxidative stress response, in addition to osmotic stress (Pahlman et al., 2001). Osmotic, nutritional and oxidative conditions are diverse signals which trigger specific stress response pathways; however, one signal may elicit more than one stress response pathway (Taymaz-Nikerel et al., 2016). Often, osmotic stress signals lead to osmotic and oxidative stress responses, exactly as we see in this case; one interpretation is that osmotic stress also generates oxidative stress, which must be mitigated for survival (Figure 7).
Our previous data and the present one showed that GPP2 is important for Cys3 protein modulation (de Melo et al., 2019) and allowed us to assemble a model to explain the facts observed here (Figure 7). In this paper, we provided further evidence that gpp2 deletion caused an up regulation of several genes in the sulfur uptake pathway and sulfur amino acid biosynthesis. Ko and collaborators also found two genes up regulated in this pathway in a hog1 deletion mutant (Ko et al., 2009). It is possible that up regulation of the sulfur uptake pathway leads to cysteine increase which would reflect in a higher ability to counter act oxidative stress through glutathione synthesis (Figure 7). The fact that taurine dioxygenases are up regulated indicate that sulfite is been generated from sulfonate and could increase the cysteine pools and consequently glutathione. In plants, the generation of intermediate metabolites in the sulfur amino acid biosynthetic routes, especially hydrogen sulfite, alleviates osmotic and oxidative stress by accumulation of osmolytes and synthesis of antioxidant enzymes (Anjum et al., 2017; Khan et al., 2017; Zhou et al., 2018). In fact, the sulfur metabolism has been considered as a defense system against biotic and abiotic stresses in plants (Rausch and Wachter, 2005).
In this paper, we revealed that GPP2 is (i) involved in osmotic stress response, (ii) is required for cold shock and transcriptionally induced by salt; (iii) it affects the expression of amino acid permeases and oxidative stress genes, and (v) it also influences the sulfur amino acid biosynthetic pathway. Cell wall integrity and ability to cope with alkaline condition are also affected by GPP2 function. Expression of virulence factors was highly reduced by GPP2 deletion, which in conjunction with the inability to cope with stresses, led to hypovirulence in G. mellonella animal model. The connections found in this paper between GPP2, the various types of stress and its link with the sulfur metabolism, which is a metabolic process that has been well proved to be linked to virulence, open a window of opportunities for therapeutical possibilities in pathogenic fungi.
Data Availability Statement
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
Author Contributions
KM performed the data collection and analysis. AV and OB performed the bioinformatics analysis (RNAseq). MV provided the financial support and wrote the manuscript. RP contributed with financial support, study design, data analysis, and manuscript writing and editing.
Funding
This work was supported by the Fundac̨ão de Amparo à Pesquisa do Estado de São Paulo to RP (2016/14542-8) and MV (2015/04400-9). KM was supported by the Fundac̨ão de Amparo à Pesquisa do Estado de São Paulo scholarship grant (2018/20390-1). AV and OB were supported by CNPq (303170/2017-4) and FAPERJ (26/202.903/20).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02728/full#supplementary-material
Footnotes
- ^ https://jgi.doe.gov/data-and-tools/bbtools/bb-tools-user-guide/bbduk-guide
- ^ https://www.bioinformatics.babraham.ac.uk/projects/fastqc
- ^ https://fungidb.org
References
Albertyn, J., Hohmann, S., Thevelein, J. M., and Prior, B. A. (1994). GPD1, which encodes glycerol-3-phosphate dehydrogenase, is essential for growth under osmotic stress in Saccharomyces cerevisiae, and its expression is regulated by the high-osmolarity glycerol response pathway. Mol. Cell Biol. 14, 4135–4144. doi: 10.1128/mcb.14.6.4135
Anjum, S. A., Ashraf, U., Tanveer, M., Khan, I., Hussain, S., Shahzad, B., et al. (2017). Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 8:69. doi: 10.3389/fpls.2017.00069
Ballou, E. R., and Johnston, S. A. (2017). The cause and effect of cryptococcus interactions with the host. Curr. Opin. Microbiol. 40, 88–94. doi: 10.1016/j.mib.2017.10.012
Brown, S. M., Campbell, L. T., and Lodge, J. K. (2007). Cryptococcus neoformans, a fungus under stress. Curr. Opin. Microbiol. 10, 320–325. doi: 10.1016/j.mib.2007.05.014
Calvete, C. L., Martho, K. F., Felizardo, G., Paes, A., Nunes, J. M., Ferreira, C. O., et al. (2019). Amino acid permeases in Cryptococcus neoformans are required for high temperature growth and virulence; and are regulated by Ras signaling. PLoS One 14:e0211393. doi: 10.1371/journal.pone.0211393
Christensen, W. B. (1946). Urea decomposition as a means of differentiating proteus and paracolon cultures from each other and from Salmonella and Shigella Types. J. Bacteriol. 52, 461–466.
Cunningham, K. W., and Fink, G. R. (1994). Calcineurin-dependent growth control in Saccharomyces cerevisiae mutants lacking PMC1, a homolog of plasma membrane Ca2+ ATPases. J. Cell Biol. 124, 351–363. doi: 10.1083/jcb.124.3.351
Darvishi, E., Omidi, M., Bushehri, A. A., Golshani, A., and Smith, M. L. (2013). The antifungal eugenol perturbs dual aromatic and branched-chain amino acid permeases in the cytoplasmic membrane of yeast. PLoS One 8:e76028. doi: 10.1371/journal.pone.0076028
de Melo, A. T., Martho, K. F., Roberto, T. N., Nishiduka, E. S., Machado, J., Brustolini, O. J. B., et al. (2019). The regulation of the sulfur amino acid biosynthetic pathway in Cryptococcus neoformans: the relationship of Cys3, calcineurin, and Gpp2 phosphatases. Sci. Rep. 9:11923. doi: 10.1038/s41598-019-48433-5
Dobin, A., Davis, C. A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., et al. (2013). STAR: ultrafast universal RNA-seq aligner. Bioinformatics 29, 15–21. doi: 10.1093/bioinformatics/bts635
Esher, S. K., Zaragoza, O., and Alspaugh, J. A. (2018). Cryptococcal pathogenic mechanisms: a dangerous trip from the environment to the brain. Mem. Inst. Oswaldo Cruz 113:e180057. doi: 10.1590/0074-02760180057
Falcon, S., and Gentleman, R. (2007). Using GOstats to test gene lists for GO term association. Bioinformatics 23, 257–258. doi: 10.1093/bioinformatics/btl567
Fan, W., Idnurm, A., Breger, J., Mylonakis, E., and Heitman, J. (2007). Eca1, a sarcoplasmic/endoplasmic reticulum Ca2+-ATPase, is involved in stress tolerance and virulence in Cryptococcus neoformans. Infect. Immun. 75, 3394–3405. doi: 10.1128/iai.01977-06
Fernandes, J. D., Martho, K., Tofik, V., Vallim, M. A., and Pascon, R. C. (2015). The role of amino acid permeases and tryptophan biosynthesis in Cryptococcus neoformans Survival. PLoS One 10:e0132369. doi: 10.1371/journal.pone.0132369
Ferreira, C., van, F., Voorst, Martins, A., Neves, L., Oliveira, R., et al. (2005). A member of the sugar transporter family, Stl1p is the glycerol/H+ symporter in Saccharomyces cerevisiae. Mol. Biol. Cell 16, 2068–2076. doi: 10.1091/mbc.e04-10-0884
Garcia-Vidal, C., Viasus, D., and Carratala, J. (2013). Pathogenesis of invasive fungal infections. Curr. Opin. Infect. Dis. 26, 270–276. doi: 10.1097/QCO.0b013e32835fb920
Hayashi, M., and Maeda, T. (2006). Activation of the HOG pathway upon cold stress in Saccharomyces cerevisiae. J. Biochem. 139, 797–803. doi: 10.1093/jb/mvj089
Hogan, D. A., Auchtung, T. A., and Hausinger, R. P. (1999). Cloning and characterization of a sulfonate/alpha-ketoglutarate dioxygenase from Saccharomyces cerevisiae. J. Bacteriol. 181, 5876–5879.
Holt, S., Kankipati, H., De Graeve, S., Van Zeebroeck, G., Foulquie-Moreno, M. R., Lindgreen, S., et al. (2017). Major sulfonate transporter Soa1 in Saccharomyces cerevisiae and considerable substrate diversity in its fungal family. Nat. Commun. 8:14247. doi: 10.1038/ncomms14247
Idnurm, A., Reedy, J. L., Nussbaum, J. C., and Heitman, J. (2004). Cryptococcus neoformans virulence gene discovery through insertional mutagenesis. Eukaryot Cell 3, 420–429. doi: 10.1128/ec.3.2.420-429.2004
Jung, W. H., Son, Y. E., Oh, S. H., Fu, C., Kim, H. S., Kwak, J. H., et al. (2018). Had1 is required for cell wall integrity and fungal virulence in Cryptococcus neoformans. G3 8, 643–652. doi: 10.1534/g3.117.300444
Kempf, B., and Bremer, E. (1998). Uptake and synthesis of compatible solutes as microbial stress responses to high-osmolality environments. Arch. Microbiol. 170, 319–330.
Kensche, P. R., Oti, M., Dutilh, B. E., and Huynen, M. A. (2008). Conservation of divergent transcription in fungi. Trends Genet. 24, 207–211. doi: 10.1016/j.tig.2008.02.003
Khan, M. N., Mobin, M., Abbas, Z. K., and Siddiqui, M. H. (2017). Nitric oxide-induced synthesis of hydrogen sulfide alleviates osmotic stress in wheat seedlings through sustaining antioxidant enzymes, osmolyte accumulation and cysteine homeostasis. Nitric Oxide 68, 91–102. doi: 10.1016/j.niox.2017.01.001
Ko, Y. J., Yu, Y. M., Kim, G. B., Lee, G. W., Maeng, P. J., Kim, S., et al. (2009). Remodeling of global transcription patterns of Cryptococcus neoformans genes mediated by the stress-activated HOG signaling pathways. Eukaryot Cell 8, 1197–1217. doi: 10.1128/EC.00120-09
Kraus, P. R., and Heitman, J. (2003). Coping with stress: calmodulin and calcineurin in model and pathogenic fungi. Biochem. Biophys. Res. Commun. 311, 1151–1157. doi: 10.1016/s0006-291x(03)01528-6
Kronstad, J., Saikia, S., Nielson, E. D., Kretschmer, M., Jung, W., Hu, G., et al. (2012). Adaptation of Cryptococcus neoformans to mammalian hosts: integrated regulation of metabolism and virulence. Eukaryot Cell 11, 109–118. doi: 10.1128/EC.05273-11
Kumar, A., Tikoo, S., Maity, S., Sengupta, S., Sengupta, S., Kaur, A., et al. (2012). Mammalian proapoptotic factor ChaC1 and its homologues function as gamma-glutamyl cyclotransferases acting specifically on glutathione. EMBO Rep. 13, 1095–1101. doi: 10.1038/embor.2012.156
Larsson, K., Ansell, R., Eriksson, P., and Adler, L. (1993). A gene encoding sn-glycerol 3-phosphate dehydrogenase (n.d.) complements an osmosensitive mutant of Saccharomyces cerevisiae. Mol. Microbiol. 10, 1101–1111. doi: 10.1111/j.1365-2958.1993.tb00980.x
Li, S. S., and Mody, C. H. (2010). Cryptococcus. Proc. Am. Thorac. Soc. 7, 186–196. doi: 10.1513/pats.200907-063AL
Liao, Y., Smyth, G. K., and Shi, W. (2019). The R package rsubread is easier, faster, cheaper and better for alignment and quantification of RNA sequencing reads. Nucleic Acids Res. 47:e47. doi: 10.1093/nar/gkz114
Limper, A. H., Adenis, A., Le, T., and Harrison, T. S. (2017). Fungal infections in HIV/AIDS. Lancet Infect. Dis. 17, e334–e343. doi: 10.1016/S1473-3099(17)30303-1
Lin, X. (2009). Cryptococcus neoformans: morphogenesis, infection, and evolution. Infect. Genet. Evol. 9, 401–416. doi: 10.1016/j.meegid.2009.01.013
Lin, X., and Heitman, J. (2006). The biology of the Cryptococcus neoformans species complex. Annu. Rev. Microbiol. 60, 69–105.
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome. Biol. 15:550.
Luo, W., and Brouwer, C. (2013). Pathview: an R/bioconductor package for pathway-based data integration and visualization. Bioinformatics 29, 1830–1831. doi: 10.1093/bioinformatics/btt285
Martho, K. F., de Melo, A. T., Takahashi, J. P., Guerra, J. M., Santos, D. C., Purisco, S. U., et al. (2016). Amino acid permeases and virulence in Cryptococcus neoformans. PLoS One 11:e0163919. doi: 10.1371/journal.pone.0163919
Mylonakis, E., Moreno, R., El Khoury, J. B., Idnurm, A., Heitman, J., Calderwood, S. B., et al. (2005). Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immun. 73, 3842–3850. doi: 10.1128/iai.73.7.3842-3850.2005
Norbeck, J., Pahlman, A. K., Akhtar, N., Blomberg, A., and Adler, L. (1996). Purification and characterization of two isoenzymes of DL-glycerol-3-phosphatase from Saccharomyces cerevisiae. identification of the corresponding GPP1 and GPP2 genes and evidence for osmotic regulation of Gpp2p expression by the osmosensing mitogen-activated protein kinase signal transduction pathway. J. Biol. Chem. 271, 13875–13881. doi: 10.1074/jbc.271.23.13875
Pahlman, A. K., Granath, K., Ansell, R., Hohmann, S., and Adler, L. (2001). The yeast glycerol 3-phosphatases Gpp1p and Gpp2p are required for glycerol biosynthesis and differentially involved in the cellular responses to osmotic, anaerobic, and oxidative stress. J. Biol. Chem. 276, 3555–3563. doi: 10.1074/jbc.m007164200
Paliwal, D. K., and Randhawa, H. S. (1978). A rapid pigmentation test for identification of Cryptococcus neoformans. Antonie Van Leeuwenhoek 44, 243–246. doi: 10.1007/bf00643226
Panadero, J., Pallotti, C., Rodriguez-Vargas, S., Randez-Gil, F., and Prieto, J. A. (2006). A downshift in temperature activates the high osmolarity glycerol (HOG) pathway, which determines freeze tolerance in Saccharomyces cerevisiae. J. Biol. Chem. 281, 4638–4645. doi: 10.1074/jbc.m512736200
Park, H. S., Chow, E. W., Fu, C., Soderblom, E. J., Moseley, M. A., Heitman, J., et al. (2016). Calcineurin targets involved in stress survival and fungal virulence. PLoS Pathog. 12:e1005873. doi: 10.1371/journal.ppat.1005873
Pascon, R. C., Ganous, T. M., Kingsbury, J. M., Cox, G. M., and McCusker, J. H. (2004). Cryptococcus neoformans methionine synthase: expression analysis and requirement for virulence. Microbiology 150, 3013–3023. doi: 10.1099/mic.0.27235-0
Peddie, B. A., Lever, M., Hayman, C. M., Randall, K., and Chambers, S. T. (1994). Relationship between osmoprotection and the structure and intracellular accumulation of betaines by Escherichia coli. FEMS Microbiol. Lett. 120, 125–131. doi: 10.1016/0378-1097(94)00188-x
Possik, E., Madiraju, S. R. M., and Prentki, M. (2017). Glycerol-3-phosphate phosphatase/PGP: role in intermediary metabolism and target for cardiometabolic diseases. Biochimie 143, 18–28. doi: 10.1016/j.biochi.2017.08.001
Price, M. F., Wilkinson, I. D., and Gentry, L. O. (1982). Plate method for detection of phospholipase activity in Candida albicans. Sabouraudia 20, 7–14. doi: 10.1080/00362178285380031
Rausch, T., and Wachter, A. (2005). Sulfur metabolism: a versatile platform for launching defence operations. Trends Plant. Sci. 10, 503–509. doi: 10.1016/j.tplants.2005.08.006
Rutherford, J. C., Bahn, Y. S., van den Berg, B., Heitman, J., and Xue, C. (2019). Nutrient and stress sensing in pathogenic yeasts. Front. Microbiol. 10:442. doi: 10.3389/fmicb.2019.00442
Saxena, A., and Sitaraman, R. (2016). Osmoregulation in Saccharomyces cerevisiae via mechanisms other than the high-osmolarity glycerol pathway. Microbiology 162, 1511–1526. doi: 10.1099/mic.0.000360
Szklarczyk, D., Morris, J. H., Cook, H., Kuhn, M., Wyder, S., Simonovic, M., et al. (2017). The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 45, D362–D368. doi: 10.1093/nar/gkw937
Taymaz-Nikerel, H., Cankorur-Cetinkaya, A., and Kirdar, B. (2016). Genome-wide transcriptional response of saccharomyces cerevisiae to stress-induced perturbations. Front. Bioeng. Biotechnol. 4:17. doi: 10.3389/fbioe.2016.00017
Toffaletti, D. L., Rude, T. H., Johnston, S. A., Durack, D. T., and Perfect, J. R. (1993). Gene transfer in Cryptococcus neoformans by use of biolistic delivery of DNA. J. Bacteriol. 175, 1405–1411. doi: 10.1128/jb.175.5.1405-1411.1993
Yang, Z., Pascon, R. C., Alspaugh, A., Cox, G. M., and McCusker, J. H. (2002). Molecular and genetic analysis of the Cryptococcus neoformans MET3 gene and a met3 mutant. Microbiology 148, 2617–2625. doi: 10.1099/00221287-148-8-2617
Zaprasis, A., Bleisteiner, M., Kerres, A., Hoffmann, T., and Bremer, E. (2015). Uptake of amino acids and their metabolic conversion into the compatible solute proline confers osmoprotection to Bacillus subtilis. Appl. Environ. Microbiol. 81, 250–259. doi: 10.1128/AEM.02797-14
Zaragoza, O., and Casadevall, A. (2004). Experimental modulation of capsule size in Cryptococcus neoformans. Biol. Proc. Online 6, 10–15. doi: 10.1251/bpo68
Zhou, Z. H., Wang, Y., Ye, X. Y., and Li, Z. G. (2018). Signaling molecule hydrogen sulfide improves seed germination and seedling growth of maize (Zea mays L.) under high temperature by inducing antioxidant system and osmolyte biosynthesis. Front. Plant Sci. 9:1288. doi: 10.3389/fpls.2018.01288
Zhu, A., Ibrahim, J. G., and Love, M. I. (2018). Heavy-tailed prior distributions for sequence count data: removing the noise and preserving large differences. Bioinformatics 35, 2084–2092. doi: 10.1093/bioinformatics/bty895
Keywords: C. neoformans, glycerol-3-phosphate phosphatase, GPP2, sulfur amino acid biosynthesis, oxidative stress, fungal virulence
Citation: Martho KF, Brustolini OJB, Vasconcelos AT, Vallim MA and Pascon RC (2019) The Glycerol Phosphatase Gpp2: A Link to Osmotic Stress, Sulfur Assimilation and Virulence in Cryptococcus neoformans. Front. Microbiol. 10:2728. doi: 10.3389/fmicb.2019.02728
Received: 13 September 2019; Accepted: 08 November 2019;
Published: 26 November 2019.
Edited by:
Jorge Amich, The University of Manchester, United KingdomReviewed by:
Thorsten Heinekamp, Leibniz Institute for Natural Product Research and Infection Biology – Hans Knoell Institute (HKI), GermanyMélissa Caza, The University of British Columbia, Canada
Guanggan Hu, University of British Columbia, Canada
Copyright © 2019 Martho, Brustolini, Vasconcelos, Vallim and Pascon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renata C. Pascon, UmVuYXRhLnBhc2NvbkBnbWFpbC5jb20=