 Anaí Díaz1†
Anaí Díaz1† Pablo Villanueva1†
Pablo Villanueva1† Vicente Oliva1
Vicente Oliva1 Carlos Gil-Durán2
Carlos Gil-Durán2 Francisco Fierro3
Francisco Fierro3 Renato Chávez2*
Renato Chávez2* Inmaculada Vaca1*
Inmaculada Vaca1*- 1Departamento de Química, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
- 2Departamento de Biología, Facultad de Química y Biología, Universidad de Santiago de Chile, Santiago, Chile
- 3Departamento de Biotecnología, Universidad Autónoma Metropolitana, Unidad Iztapalapa, Mexico City, Mexico
Cold-adapted fungi isolated from Antarctica, in particular those belonging to the genus Pseudogymnoascus, are producers of secondary metabolites with interesting bioactive properties as well as enzymes with potential biotechnological applications. However, at genetic level, the study of these fungi has been hindered by the lack of suitable genetic tools such as transformation systems. In fungi, the availability of transformation systems is a key to address the functional analysis of genes related with the production of a particular metabolite or enzyme. To the best of our knowledge, the transformation of Pseudogymnoascus strains of Antarctic origin has not been achieved yet. In this work, we describe for the first time the successful transformation of a Pseudogymnoascus verrucosus strain of Antarctic origin, using two methodologies: the polyethylene glycol (PEG)-mediated transformation, and the electroporation of germinated conidia. We achieved transformation efficiencies of 15.87 ± 5.16 transformants per μg of DNA and 2.67 ± 1.15 transformants per μg of DNA for PEG-mediated transformation and electroporation of germinated conidia, respectively. These results indicate that PEG-mediated transformation is a very efficient method for the transformation of this Antarctic fungus. The genetic transformation of Pseudogymnoascus verrucosus described in this work represents the first example of transformation of a filamentous fungus of Antarctic origin.
Introduction
Antarctica, one of the most extreme ecosystems in the world, is inhabited by a myriad of microorganisms that have adapted to this harsh environment using several cellular and molecular strategies (Pearce, 2012). Among the microorganisms usually found in Antarctica, filamentous fungi are one of the most interesting groups. The number of fungal species recorded so far in the different Antarctic ecosystems studied is very high, suggesting a great diversity of fungi (Bridge and Spooner, 2012; Rosa et al., 2019). In addition, studies prospecting bioactivities in filamentous fungi isolated from Antarctica have gained much interest during last decades, showing that these fungi produce metabolites with interesting biological activities (Tian et al., 2017; Vaca and Chávez, 2019), as well as enzymes with novel properties, which could be useful for biotechnological purposes (Duarte et al., 2017, 2018).
Cold-adapted fungi from the genus Pseudogymnoascus are prevalent in most of the Antarctic environments (Ogaki et al., 2019; Rosa et al., 2019). To date, the species P. appendiculatus, P. destructans, P. pannorum, P. verrucosus, and P. vinaceous have been isolated from different Antarctic environments (Loque et al., 2010; Gomes et al., 2018; Alves et al., 2019; Kochkina et al., 2019; Rosa et al., 2019). Several studies have shown that extracts of these fungi produce bioactive metabolites with potential application, including antibacterial, antifungal, tripanocidal, herbicidal, and antitumoral activities (Furbino et al., 2014; Henríquez et al., 2014; Gonçalves et al., 2015; Gomes et al., 2018; Purić et al., 2018; Vieira et al., 2018). On the other hand, Pseudogymnoascus strains of Antarctic origin also have shown ability to produce cold-adapted enzymes with potential application in several industrial processes (Loperena et al., 2012; Poveda et al., 2018).
The aforementioned studies confirm the enormous biotechnological potential of Pseudogymnoascus strains of Antarctic origin. However, the future exploitation of this potential will require the development of suitable genetic tools. In this sense, and in order to study the genes responsible for the biosynthesis of secondary metabolites and enzymes from Pseudogymnoascus strains of Antarctic origin, an efficient transformation system is necessary. To the best of our knowledge, to date only an Agrobacterium tumefaciens-mediated transformation (ATMT) method has been used for the transformation of fungi from the genus Pseudogymnoascus. Zhang et al. (2015) used this method to perform the successful transformation of two different species: Pseudogymnoascus destructans, the etiologic agent of white-nose syndrome of hibernating bats, and Pseudogymnoascus pannorum, a species widely distributed in different environments.
ATMT has been described as a very laborious and time-consuming protocol that involves, among others, the construction of dual vectors for transformation and the previous transformation of bacteria A. tumefaciens (Rehman et al., 2016; Li et al., 2017). Thus, the development of alternative methods that could be simpler and faster is desirable and should be fostered (Rehman et al., 2016). Among alternative methods, polyethylene glycol (PEG)-mediated transformation has been highlighted as a simple and fast method, which has been successfully applied for the transformation of many fungal species (Rodríguez-Iglesias and Schmoll, 2015). On the other hand, electroporation has also been used for the transformation of several fungi. An additional advantage of this method is the possibility of using protoplasts, mycelia, or conidia, making electroporation potentially faster and simpler than protoplast transformation (Rivera et al., 2014).
Since several years, our team has been working in the study of the secondary metabolism and enzymes of Antarctic fungi, including species from the genus Pseudogymnoascus. As a result, we have described and purified novel secondary metabolites with interesting properties from these strains (Henríquez et al., 2014; Figueroa et al., 2015). In addition, we detected that these strains produce cold-adapted enzymes (Poveda et al., 2018). Unfortunately, at genetic level, the lack of a suitable transformation system hinders to go further in our research. To overcome this difficulty, we have addressed the development of transformation methods for Pseudogymnoascus strains of Antarctic origin. In this work, we have applied these developments, performing the successful transformation of an Antarctic strain, Pseudogymnoascus verrucosus, by using two different methodologies: PEG-mediated transformation and electroporation of germinated conidia. To the best of our knowledge, this is the first report of successful transformation in P. verrucosus, and more important, the first example of transformation of a filamentous fungus of Antarctic origin.
Materials and Methods
Fungal Strain, Culture Media, and Plasmid
The strain P. verrucosus FAE27 used in this work was previously isolated from an Antarctic marine sponge (Henríquez et al., 2014). The fungus was routinely kept on potato dextrose agar (PDA, Difco) and grown at 15°C. CM medium (6 g/l yeast extract, 6 g/l casein acid hydrolysate, 10 g/l sucrose) was used for growing the fungus in flasks.
The plasmid used for transformation experiments was pAN7-1, which contains the hygromycin B phosphotransferase (hph) gene as a dominant selectable marker under the control of gpdA promoter and trpC terminator from Aspergillus nidulans (Punt et al., 1987). The plasmid was purified from Escherichia coli using routine procedures and dialyzed against distilled water prior to use. In the case of experiments requiring linearized DNA, the plasmid was previously digested with HindIII.
Antibiotic Resistance Test of P. verrucosus FAE27
To determine the sensitivity of P. verrucosus FAE27 toward hygromycin B and phleomycin, 1 × 107 protoplasts (see section “Protoplast Isolation”) were added to 7.5 ml of molten overlay agar (CM medium plus 1% sorbitol and 0.8% agar) supplemented with different concentrations of hygromycin B (1, 2.5, 5, 10, and 20 μg/ml) or phleomycin (50, 100, 200, 500, and 700 μg/ml). Each mixture was quickly poured onto 7.5 ml of base agar (CM medium plus 1% sorbitol and 2%) on Petri dishes. Dishes were incubated during 15 days at 15°C. After this time, the presence of hygromycin-resistant or phleomycin-resistant colonies was evaluated. The same procedure was performed using 1 × 107 germinated conidia instead of protoplasts.
Polyethylene Glycol-Mediated Transformation of P. verrucosus FAE27
Protoplast Isolation
P. verrucosus FAE27 was cultured on PDA plates at 15°C for 15 days. Asexual spores (conidia) of one plate were harvested by gently scraping the agar with sterile distilled water. The resulting spore suspension was filtered through a sterile Miracloth filter (EMD Millipore, USA). The filtered spore suspension was inoculated in 50 ml of CM medium, and incubated at 15°C and 180 r.p.m. for 72 h. Mycelia obtained was collected by filtration in Miracloth filter and washed three times with 0.9% NaCl. Then, 1.5 g of harvested mycelia were aseptically transferred to an Erlenmeyer flask containing 10 ml of TPP solution (50 mM K2HPO4, 50 mM KH2PO4, 0.7 M KCl, pH 5.8). In parallel, 0.4 g of lysing enzymes from Trichoderma harzianum (Sigma-Aldrich, USA) were dissolved in 10 ml of TPP and filtered through 0.22 μm filter (Millex®GP, EMD Millipore, USA). This filtered solution was added to the Erlenmeyer flask containing resuspended mycelia, and the mixture was incubated at 28°C and 80 r.p.m. Protoplast release was checked every 30 min by counting in Neubauer chamber. Optionally, to increase the number of protoplasts released, 40 μl of β-glucuronidase solution (Sigma-Aldrich, USA) were added at 30 min, 1 h, and 2 h of incubation. Released protoplasts were carefully filtered two times, first using sterile Miracloth and then a sterile 40-μm nylon filter. The final solution containing protoplasts was centrifuged at 3,000 r.p.m. for 4 min, and the supernatant was discarded.
Polyethylene Glycol/CaCl2-Mediated Transformation
Protoplasts obtained were washed and centrifuged three times with KCM (0.5 M MES, 0.7 M KCl, 50 mM CaCl2, pH 5.8), and then resuspended with STC (20% sucrose, 10 mM Tris–HCl pH 8.0, 50 mM CaCl2) to 108 protoplasts/ml. About 100 μl of protoplasts suspension (1 × 107 protoplasts) were mixed with different amounts of plasmid DNA (3, 5, or 10 μg), 10 μl of 0.1 M aurintricarboxylic acid solution (Sigma-Aldrich, USA), and 10 μl of transformation solution PCM. Composition of PCM was 0.5 M MES, 50 mM CaCl2, and 50% PEG (PEG 3350, PEG 6000, or PEG 8000), pH 5.8.
Transformation mixtures were incubated on ice for 30 min, diluted with 500 μl of PCM, and incubated again at room temperature for additional 30 min. Then, 600 μl of STC and 3 ml of CM medium containing 1 M sorbitol (CM-sorbitol) were added. This mixture was incubated at 15°C and 80 r.p.m. during a protoplast recovery period of 24 h. After this time, 200–300 μl aliquots of transformation mixtures were added to 7.5 ml of molten overlay agar (CM medium, 1% sorbitol, 40 μg/ml hygromycin B, 0.8% agar). Mixture was quickly poured onto 7.5 ml of base agar (CM medium, 1% sorbitol, 40 μg/ml hygromycin B, 2% agar) on Petri dishes. After 15–20 days of incubation at 15°C, hygromycin-resistant colonies were visible. It is important to highlight that the temperature of molten agar must be around 40–42°C. Molten agar at higher temperatures dramatically reduces the number of transformants obtained, and at 45°C transformants are not obtained.
To verify viability of the protoplasts and effectiveness of the antibiotic selection during the experimental procedure, two controls were included: a transformation experiment without DNA and hygromycin B, and a transformation experiment without DNA but with hygromycin B.
Transformation Experiments Including Lithium Acetate
In some PEG-mediated transformations protocols previously published, calcium was replaced by other cations such as lithium (Mach, 2003; Olmedo-Monfil et al., 2004). Accordingly, we also performed PEG-mediated transformation experiments with solutions containing lithium acetate. In these experiments, we followed exactly the same protocol described for PEG/CaCl2-mediated transformation, changing one or all the solutions as follows:
1. Protoplasts obtained were washed and centrifuged three times with KLM (0.5 M MES, 0.7 M KCl, 50 mM lithium acetate, pH 5.8) and resuspended with STL (20% sucrose, 10 mM Tris-HCl pH 8.0, and 50 mM lithium acetate).
2. PCM was replaced by PLM. Composition of PLM was 0.5 M MES, 50 mM lithium acetate, and 50% PEG (PEG 3350, PEG 6000, or PEG 8000), pH 5.8.
3. After 30 min at room temperature, 600 μl of STL (not STC) were added.
Electroporation of Germinated Conidia of P. verrucosus FAE27
Preparation of Germinated Conidia
P. verrucosus FAE27 was cultured on PDA plates at 15°C for 15 days. Asexual spores from four Petri dishes were harvested by scraping their surfaces with 5 ml of potassium phosphate buffer (0.03 M potassium phosphate, 0.02% Tween 20, pH 7.0). Mycelia fragments were removed by filtering through sterile Miracloth. The filtered conidia were concentrated by centrifugation at 9,000 r.p.m. for 15 min, washed three times with deionized water, and suspended in water to 108 conidia/ml. Conidia suspension was kept at 4°C for 24 h to induce dormancy and thus obtain a synchronized germination. For germination, 50 ml of CM medium were inoculated with conidia at a final concentration of 1 × 107 conidia/ml, and incubated at 15°C and 180 r.p.m. during 19–20 h. After this time, 90–95% of spores were germinated.
Germinated conidia were centrifuged at 6,000 r.p.m. during 15 min, resuspended in 1 ml of ice-cold distilled water, and collected again by centrifugation during 15 min at 8,000 r.p.m. and 10°C. Then, the germinated conidia were resuspended to 1 × 108 conidia/ml in an ice-cold osmotic buffer: HEPES buffer (1 mM HEPES pH 7.5, 50 mM mannitol) or lithium acetate buffer (10 mM Tris-HCl pH 7.5, 270 mM sucrose, 1 mM lithium acetate).
Treatment of Germinated Conidia With Dithiothreitol
In some experiments, germinated conidia were treated with a DTT solution before their resuspension in osmotic buffer. DTT solution (50 mM potassium phosphate buffer, 25 mM DTT, pH 7) was prepared and sterilized by filtration. The conidia were resuspended in 1 ml of DTT solution and incubated at 15°C and 60 r.p.m. for 15 min to be subsequently centrifuged and resuspended in the osmotic buffer.
Electroporation of Germinated Conidia
Aliquots of 100 μl containing 1 × 107 germinated conidia were mixed with 3, 5, or 10 μg of plasmid DNA and kept on ice for 15 min before electroporation. Each transformation mixture was transferred into a 0.2 cm electroporation cuvette and subjected to electroporation using the Gene Pulser Xcell Total System (Bio-Rad Laboratories, USA). Each transformation mixture was electroporated by a single exponential pulse ranging between 0.8 and 2.0 kV/cm, whereas capacitance and resistance were kept constant at 25 μF and infinite Ω resistance, respectively. Experiments using capacitances of 10 or 50 μF, or lower resistances, were also performed.
Immediately after pulse, 1 ml of ice-cold CM-sorbitol medium was added to each cuvette. Each mixture was transferred to a sterile tube. Then, 1 ml of CM-sorbitol was added again and each mixture was incubated at 15°C for 24 h and 80 r.p.m. for recovery. Finally, each transformation mixture electroporated was poured on Petri dishes exactly as was described for protoplast transformation (see section “Polyethylene Glycol/CaCl2-Mediated Transformation”). Hygromycin-resistant colonies were visible after 15–20 days of incubation at 15°C.
For all the transformation experiments, the same controls described in PEG/CaCl2-mediated transformation were performed. In addition, controls where the pulse was omitted were included.
Stability Test for Hygromycin-Resistant Transformants
The mitotic stability of the transformants was assessed. For this purpose, 10 hygromycin-resistant transformants obtained by PEG-mediated transformation and 10 hygromycin-resistant transformants obtained by electroporation were randomly selected and transferred on plates with selective medium (CM medium containing 40 μg/ml hygromycin B). In order to obtain monosporic cultures of these transformants, asexual spores (conidia) were collected, diluted, and plated on selective media again. After growth, conidia from a single colony were transferred to non-selective medium (CM medium without hygromycin B) and incubated 2 weeks at 15°C. This last step was successively repeated three times. Finally, conidia growing on non-selective medium were transferred back to selective medium to confirm their resistance toward hygromycin B.
Molecular Analysis of Hygromycin-Resistant Transformants
Genomic DNA from the hygromycin-resistant transformants was extracted using the protocol described by Navarrete et al. (2009). The presence of hph gene in DNA samples was detected by PCR using primers pgpdRNAi (GCTACATCCATACTCCATCC) and P4-sh2hindIII (AGTCGTAAGCTTAATGTGTGTCCTGTAGGCTT). PCR conditions were as follows: initial denaturation at 94°C for 3 min followed by 30 cycles at 94°C for 40 s, annealing at 55°C for 40 s and extension at 72°C for 2 min. The expected amplicon size was 1,689 bp.
Results
Sensitivity of P. verrucosus FAE27 Toward Different Antibiotics
Before transformation experiments, the sensitivity of P. verrucosus FAE27 toward hygromycin B and phleomycin was tested. We selected these antibiotics because they are widely used as selection markers in fungal transformation (Ruiz-Díez, 2002). Typically, phleomycin inhibits growth of filamentous fungi at concentrations of 25–150 μg/ml. In our case, we did not observe inhibition of P. verrucosus FAE27 growth using phleomycin, even at concentration as high as 700 μg/ml.
In the case of hygromycin B, the growth of P. verrucosus FAE27 was inhibited at 20 μg/ml (Figure 1). This result agrees with those observed in P. destructans and P. pannorum (Zhang et al., 2015; see section “Discussion”). Therefore, hereafter we used 40 μg/ml of hygromycin B, equivalent to two fold the MIC determined, for the selection of transformants.
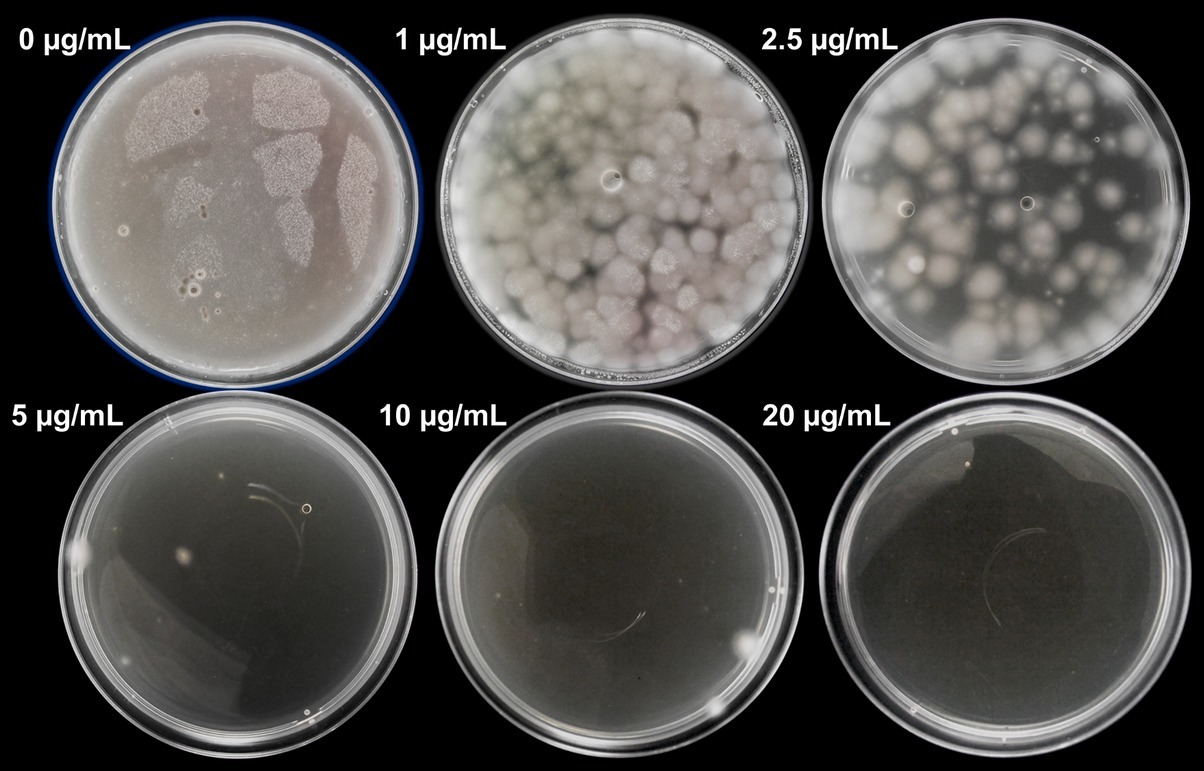
Figure 1. Sensitivity of protoplasts of P. verrucosus FAE27 toward hygromycin B. CM medium was supplemented with different concentrations of hygromycin B (0–20 μg/ml) and incubated for 15 days at 15°C. As can be seen, 20 μg/ml of hygromycin B completely inhibited fungal growth. The same experiment with identical results was performed using germinated conidia (data not shown).
Polyethylene Glycol-Mediated Transformation of P. verrucosus FAE27
Generation of Protoplasts
The first step in PEG-mediated transformation is the generation of protoplasts. For this purpose, fungal cell wall should be digested using hydrolytic enzymes. To the best of our knowledge, preparation of protoplasts has not been reported in any Pseudogymnoascus strain so far. In our case, we tested whether the use of lysing enzymes from Trichoderma harzianum, which are widely used to produce fungal protoplasts, provides good release of protoplasts from P. verrucosus FAE27. We observed that the incubation of mycelia with lysing enzymes yielded 3.82 × 108 ± 0.86 × 108 protoplasts after 3 h, enough to perform around 20 transformation experiments. This value was estimated considering 1 × 107 protoplasts for each transformation experiment, amount optimal in most protocols of fungal PEG-mediated transformation (Liu and Friesen, 2012; Wang et al., 2018).
In addition to lysing enzymes, it has been described that the supplementation with β-glucuronidase improves protoplasts release from fungi (Wiebe et al., 1997). In P. verrucosus FAE27, we observed that the joint use of lysing enzymes and β-glucuronidase produced 9.26 × 108 ± 1.36 × 108 protoplasts, two to three times more protoplasts than lysing enzymes alone. Thus, and depending on protoplasts required by the user, the use of β-glucuronidase could be considered optional.
Effect of Plasmid DNA Concentration, Recovery Time of Protoplasts, Molecular Weight of Polyethylene Glycol, and Metal Ion in Polyethylene Glycol-Mediated Transformation
For PEG-mediated transformation of fungi, researchers usually use DNA amounts ranging from 2 to 12 μg (Fincham, 1989; Wang et al., 2018). In our case, we tested three DNA amounts within this range (Table 1). As observed in Table 1, the increase of DNA amount from 3 to 5 μg increased the number of transformants obtained. However, in terms of transformation efficiency, this increase was not statistically significant. On the other hand, when DNA amount was increased up to 10 μg, the number of transformants increased around nine times, while transformation efficiencies increased around four times (Table 1). Therefore, for the rest of experiments, concentration of plasmid DNA was set at 10 μg. Figure 2 shows hygromycin-resistant transformants of P. verrucosus FAE27 obtained after a typical transformation experiment. Interestingly, the linearization of the plasmid negatively affected the efficiency of the transformation. Thus, and after several experiments using linearized plasmid, transformants were not obtained (Table 1).
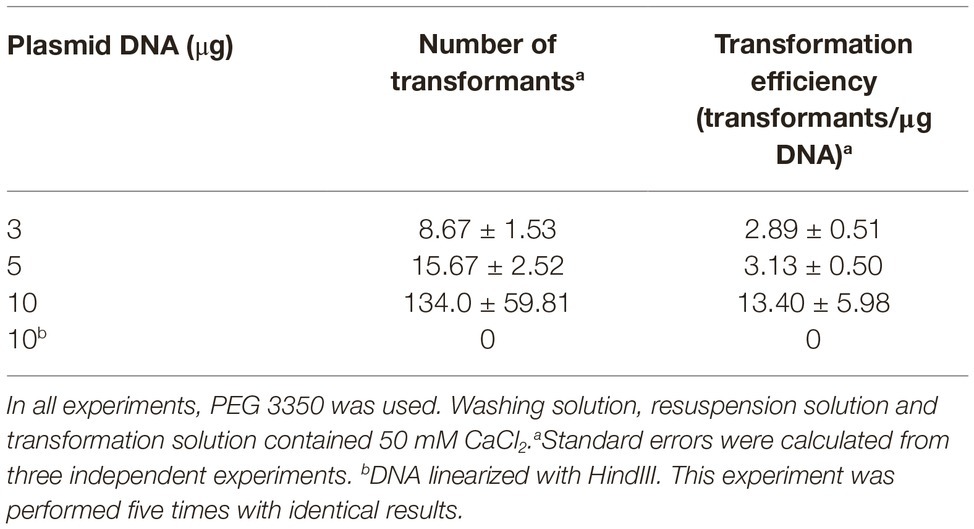
Table 1. Effect of plasmid DNA amount on transformation efficiencies in PEG-mediated transformation of Pseudogymnoascus verrucosus FAE27.
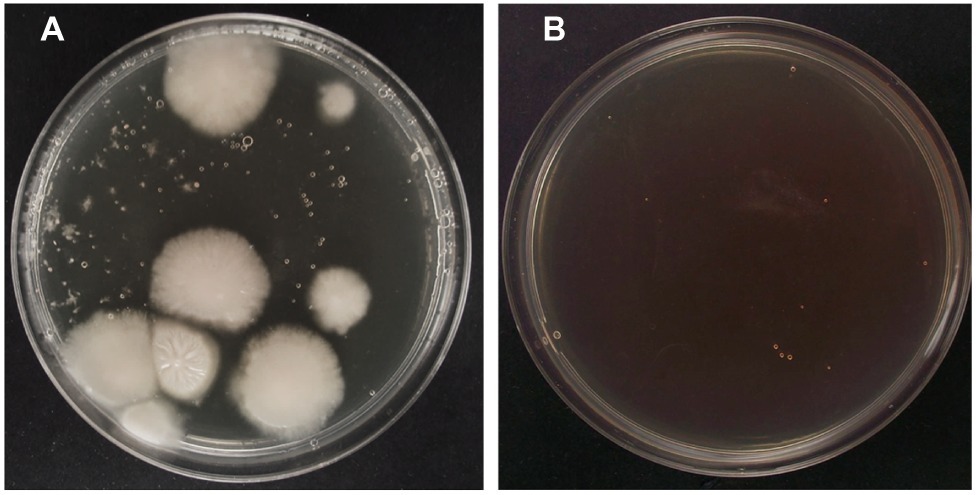
Figure 2. Successful transformation of P. verrucosus FAE27 by using PEG-mediated transformation method. (A) Transformation experiment using plasmid pAN7-1. Colonies observed after 20 days of growth on selective plates with 40 μg/ml hygromycin B. (B) Control transformation experiment without pAN7-1. Note the absence of fungal colonies.
Regarding time of regeneration of protoplasts after transformation, we used recovery times of 3 and 24 h. Transformants were obtained only after 24 h of recovery, so this time was used for all the experiments.
In PEG-mediated transformation of fungi, the components of the solutions used are important. Typical PEG-mediated transformation protocols include a washing solution which is used during the recovery process of the protoplasts, a solution for resuspension of protoplasts prior to transformation, and a transformation solution which is added together with DNA to perform the transformation. The main components of these solutions are an osmotic stabilizer to maintain the integrity of protoplasts during the procedure, calcium ions to produce the uptake of exogenous DNA by the cell, and PEG, which promotes cell agglomeration making DNA uptake easier. In particular, it has been observed that the molecular weight of the PEG is a key factor influencing the efficiency of transformation, so this parameter should be optimized in each fungal species (Rodríguez-Iglesias and Schmoll, 2015). Table 2 shows the effect of PEG 3350, PEG 6000, and PEG 8000 in the transformation efficiencies of P. verrucosus FAE27. As can be observed, best results were achieved by using PEG 3350. When molecular weight of PEG was increased, transformation efficiencies were drastically diminished and indeed, when PEG 8000 was used, transformants were not obtained (Table 2).
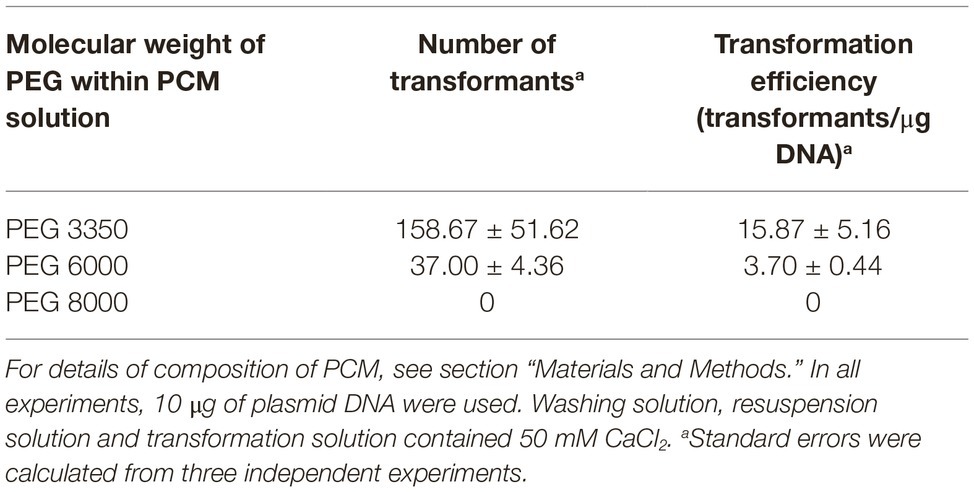
Table 2. Transformation efficiencies of PEG/CaCl2-mediated transformation of Pseudogymnoascus verrucosus FAE27, using PEG of different molecular weights.
As mentioned, typical PEG-mediated transformation protocols use calcium ions to facilitate DNA uptake. However, in some fungi, it has been described that good transformation yields may be obtained using solutions containing lithium ion and PEG (Mach, 2003; Olmedo-Monfil et al., 2004). Accordingly, we tested the effect of changing CaCl2 by lithium acetate in the last step of PEG/CaCl2-mediated transformation (Table 3). Our results indicate that this change is not favorable for the PEG-mediated transformation of P. verrucosus FAE27, producing a diminished transformation efficiency of 23% with respect to CaCl2. At this point it should be highlighted that the use of PEG 6000 or PEG 8000, together with lithium acetate, did not yield any transformant in several experiments performed (Table 3), indicating a joint detrimental effect of high molecular weight PEG and lithium acetate.
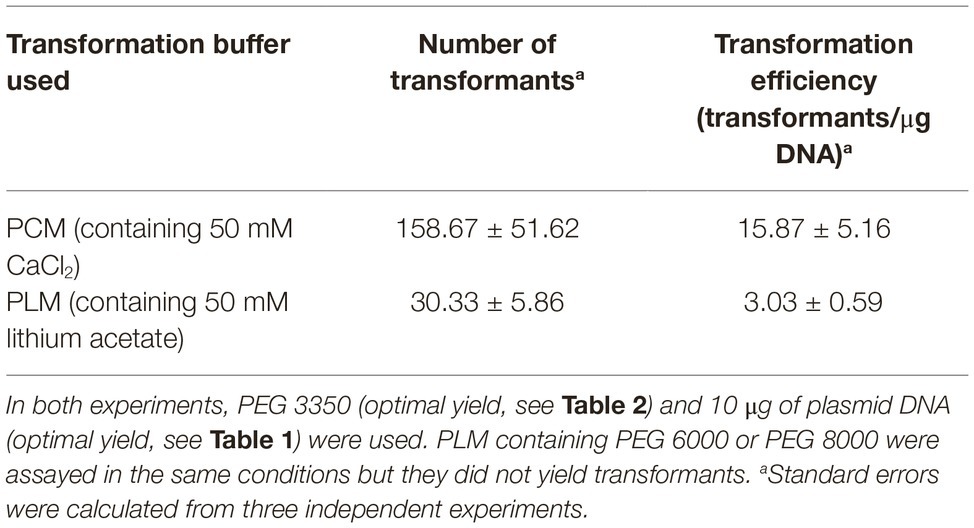
Table 3. Effect of changing PCM by PLM in the last step of PEG/CaCl2-mediated transformation of Pseudogymnoascus verrucosus FAE27.
Similar negative results were observed when we performed experiments where CaCl2 was replaced by lithium acetate in all the solutions used, that is, a PEG/lithium acetate-mediated transformation. After several attempts, we never obtained transformants by using this method, even when PEG 3350 was used.
In conclusion, high efficiencies in PEG-mediated transformation of P. verrucosus FAE27 were successfully achieved using 10 μg of non-linearized DNA, and solutions containing low molecular weight PEG and CaCl2.
Transformation of P. verrucosus FAE27 by Electroporation of Germinated Conidia
Optimization of Electroporation Conditions
Capacitance and field strength are critical parameters to obtain high transformation efficiencies in electroporation. Regarding capacitance, we initially tested three different capacitance values: 10, 25, and 50 μF. After several experiments, we observed that transformants were obtained only when 25 μF were used. On the contrary, experiments performed with capacitances of 10 or 50 μF did not give positive results. Therefore, the rest of experiments were done with capacitance value set at 25 μF.
In addition, we tested voltages in the range of 0.8–2.0 kV. Transformants were never obtained using high voltages of 1.5 and 2.0 kV (Table 4). Conversely, transformants of P. verrucosus FAE27 were obtained using voltages of 0.8, 1.0, and 1.25 kV (Table 4), corresponding to field strengths of 4, 5, and 6.25 kV/cm, respectively. We found that a voltage of 1.0 kV produced the highest electrotransformation efficiency, while pulses of 0.8 and 1.25 kV reduced the efficiency. In summary, the highest transformation efficiencies were obtained with 25 μF and 5 kV/cm.
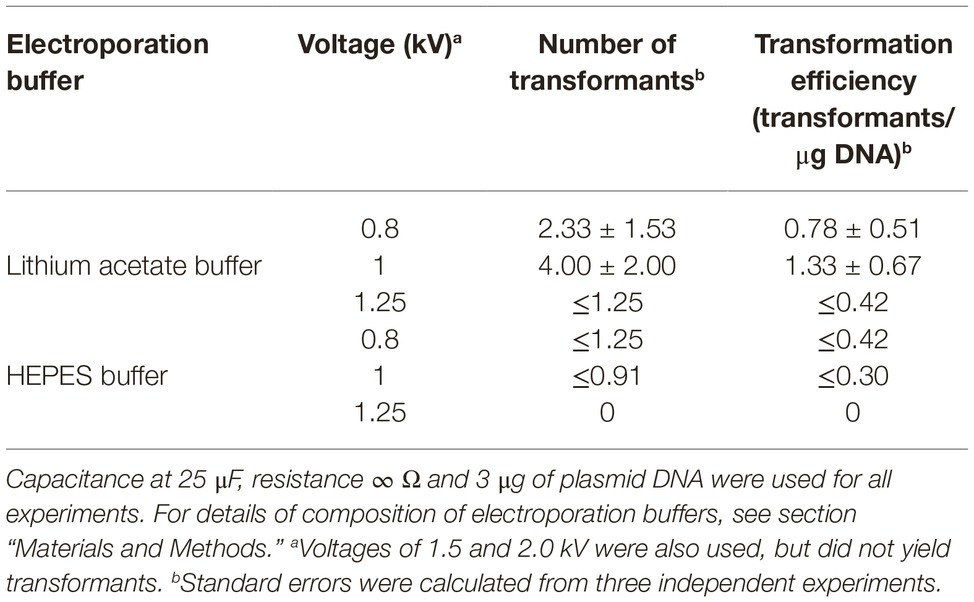
Table 4. Effect of voltage and electroporation buffer on efficiencies of electroporation of germinated conidia of Pseudogymnoascus verrucosus FAE27.
On the other hand, the permeability of the cellular membrane depends on electroporation buffer composition (Dermol et al., 2016). Buffers commonly used in fungal transformation by electroporation are HEPES buffer (Chakraborty et al., 1991; Cruz-Ramón et al., 2019) and lithium acetate buffer (Xu et al., 2014; Vela-Corcía et al., 2015). We analyzed the effect of both buffers in the transformation efficiencies of P. verrucosus FAE27. Table 4 shows that higher transformation efficiencies were obtained using lithium acetate buffer in the three voltages assayed.
Effect of Amount of DNA and Treatment of Germinated Conidia With Dithiothreitol in Electrotransformation Efficiencies
In general, amount of DNA used in electroporation protocols ranges between 1 and 10 μg (Chakraborty et al., 1991; Dantas-Barbosa et al., 1998). We used three amounts of DNA within this range (Table 5). We observed that highest transformation efficiencies were achieved using 3 μg of plasmid (1.00 ± 0.33 transformants/μg DNA, Table 5). In addition, we observed that the linearization of the plasmid positively affected the efficiency of the transformation, which was increased to 2.67 ± 1.15 transformants/μg DNA (Table 5).

Table 5. Effect of plasmid DNA amount on efficiencies of electroporation of germinated conidia of Pseudogymnoascus verrucosus FAE27.
In previous reports (Meilhoc et al., 1990; Kawai et al., 2010; Cruz-Ramón et al., 2019), it has been described that DTT, a reagent that reduces disulfide bonds, promotes the weakening of fungal cell wall, thus facilitating DNA uptake in electroporation protocols. However, in our case, we observed that the treatment of germinated conidia with DTT had a negative effect on transformation efficiency, yielding 0.44 ± 0.19 transformants/μg DNA, around a 66% reduction in efficiency as compared to the control experiment without DTT.
Mitotic Stability and Molecular Analysis of Transformants
The 20 randomly selected hygromycin-resistant transformants had high stability. After cultivation on non-selective medium for three successive times, they were able to grow well on selective plates containing hygromycin B. Importantly, these hygromycin-resistant transformants did not show any visible defect of growth as compared to wild-type P. verrucosus FAE27.
Finally, the presence of hph gene in genomic DNA of the 20 randomly selected hygromycin-resistant transformants was verified by using PCR. Figure 3 shows all these DNA samples yielded the expected amplicon of 1,689 bp, whereas amplicon was not obtained using genomic DNA from untransformed P. verrucosus. These results confirm the successful genetic transformation of P. verrucosus FAE27.
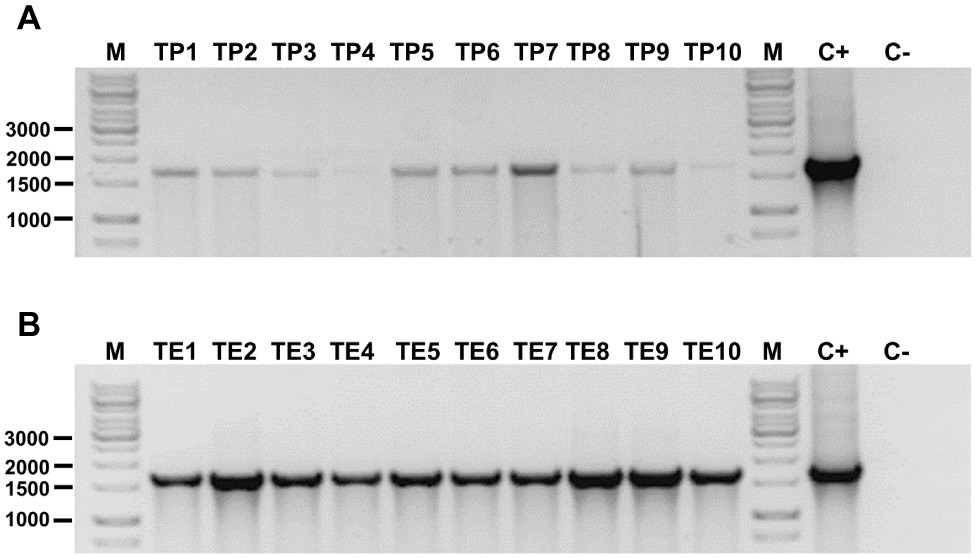
Figure 3. Detection of hph gene in transformants of P. verrucosus obtained by PEG-mediated transformation (A) or by electroporation of germinated conidia (B). In each case, 10 transformants were analyzed. Lanes TP1 to TP10 and TE1 to TE10: Amplicon of 1,689 bp belonging to the hph gene conferring resistance to hygromycin B to the transformants. Lane C+: Positive control of amplification using pure pAN7-1. Lane C−: Negative control using genomic DNA from untransformed P. verrucosus FAE27. Lane M: Molecular-size marker (GeneRuler 1 kb DNA Ladder, Thermo Scientific). Relevant molecular weights are indicated.
Discussion
In filamentous fungi, the most widely methods used for transformation are ATMT, PEG-mediated protoplast transformation, and electroporation (Li et al., 2017). To date, only ATMT has been used for the transformation of Pseudogymnoascus species (Zhang et al., 2015, 2018). On the contrary, and despite PEG-mediated transformation systems have been used to transform successfully a great variety of fungi (Rodríguez-Iglesias and Schmoll, 2015), to date this method has not been tested in Pseudogymnoascus. The PEG-mediated transformation method developed in this work for P. verrucosus FAE27 yielded an average of 159 transformants per transformation experiment (Table 2). As comparison, ATMT method yielded an average of 60 transformants in P. destructans, and 120 transformants in P. pannorum for each transformation experiment (Zhang et al., 2015, 2018). Thus, the number of transformants obtained with our PEG-mediated transformation method compares positively with those obtained by ATMT method, making our method an alternative that could be applied to other Pseudogymnoascus species.
Before transformation experiments, we evaluated the sensitivity of P. verrucosus FAE27 to hygromycin B and phleomycin. Our results showed that hygromycin B, at concentration of 20 μg/ml, completely inhibited the growth of P. verrucosus FAE27. Hygromycin B was previously used as selection marker for P. destructans and P. pannorum, which were inhibited at concentration of 100 μg/ml (Zhang et al., 2015). Thus, P. verrucosus FAE27 seems to be more sensitive to hygromycin B than P. destructans and P. pannorum. On the other hand, we observed that P. verrucosus FAE27 was able to grow in medium containing up to 700 μg/ml of phleomycin, which suggest that this fungus is essentially resistant to this antibiotic. These results suggest that for the future development of transformation systems in other Pseudogymnoascus species, hygromycin should be considered as the first option.
PEG-mediated transformation requires the generation of a suitable amount of protoplasts. In P. verrucosus FAE27, the digestion with lysing enzymes released in the order of 108 protoplasts per gram and a half of mycelia, which is similar to the protoplasts obtained in other fungi successfully transformed (Liu and Friesen, 2012; Li et al., 2017). Interestingly, this number was dramatically increased by the addition of β-glucuronidase, a fact that has also been observed in other fungi (Wiebe et al., 1997).
Two essential components of solutions used in PEG-mediated transformation are PEG and metal cation. PEG induces disorder charges on the cytomembrane surface of protoplasts, thus altering membrane permeability and facilitating the entry exogenous nucleic acids into the cells (Fincham, 1989). In PEG-mediated transformations of fungi, in general it has been observed that low molecular weight PEG has superior performance than high molecular weight PEG, although this variable has to be optimized case-by-case (Becker and Lundblad, 2001). In our case, and in agreement with literature, we observed that in P. verrucosus FAE27, PEG of low molecular weight (PEG 3350) produced markedly better transformation efficiencies of protoplasts than high molecular weight PEG. Regarding metal ions, PEG-mediated transformation solutions usually contain calcium ions, which are thought opening channels in the cytomembrane, thereby facilitating the entry of exogenous DNA into protoplasts (Li et al., 2017). On the other hand, and although not much is known about the effect of lithium ions on membranes, it has been observed that these ions can alter membranes allowing DNA uptake into the cells (Jakobsson et al., 2017). In our experiments, we found that the replacement of calcium ions by lithium ions in the transformation solution produced a drastic reduction in transformation efficiencies. Indeed, the replacement of calcium by lithium in all the transformation solutions did not yield transformants. During the course of these experiments with lithium, we observed that control dishes of protoplasts viability showed around 1% of protoplasts regeneration, a very low level of regeneration as compared with experiments performed with calcium ions, where around 40% of protoplasts regenerated. Although more research is needed, these results suggest that the detrimental effect of lithium ions could be related with the lack of stability of the protoplasts in solutions containing lithium, or their inability to perform cell wall regeneration after transformation process.
We also identified other two key parameters that critically influenced the transformation efficiencies of P. verrucosus FAE27: temperature of molten agar, and time of recovery of transformants. Regarding the temperature of molten agar, most fungal transformation protocols described in literature specify that transformed protoplasts or electroporated conidia should be mixed with molten agar at 50°C (Gangavaram et al., 2009; Liu and Friesen, 2012; Cruz-Ramón et al., 2019). However, in our case, we never obtained transformants of P. verrucosus FAE27 using molten agar at temperatures over 45°C. On the contrary, good transformation efficiencies were always obtained with molten agar not exceeding 42°C. Important, and although our protocol of electroporation of germinated conidia had lower efficiencies (Tables 4, 5), the precautions regarding the temperature of molten agar also apply to this protocol. Taken together, these results suggest that both protoplasts and germinated conidia of P. verrucosus FAE27 are very sensitive to high temperatures. P. verrucosus FAE27 has a temperature range of growth between 4 and 25°C with optimal growth at 15°C (Henríquez et al., 2014; Poveda et al., 2018), which agrees with this observation. However, it should be highlighted that some mesophilic fungi also requires molten agar at low temperature to successfully obtain transformants (Cho et al., 2013; Roth and Chilvers, 2019), so this is not a specific requirement of cold-adapted fungi.
After transformation, transformants need a time of incubation for the regeneration of the cell wall before plating on selection medium (Liu and Friesen, 2012). In most protocols involving mesophilic strains, this recovery time ranges between 2 and 5 h (Liu and Friesen, 2012; Xu et al., 2014). Accordingly, we initially attempted recovery times of 3 h, but we did not obtain transformants. We reasoned that due to its cold-adapted character, P. verrucosus FAE27 probably has slower growth velocity than mesophilic fungi and needs longer recovery times. Indeed, the growth velocity of P. verrucosus is 2.4 mm/day at 15°C, which is markedly lower than the growth velocity of several mesophilic fungal species, which are in a range of 4–10 mm/day (Sautour et al., 2002; Segers et al., 2016). Consequently, we attempted a longer time of recovery of 24 h with successful results. This longer recovery time was applied to PEG-mediated transformation and the electroporation of germinated conidia methods.
In some PEG-mediated transformation protocols, the use of linear plasmids is recommended to increase the frequency of transformation. However, results obtained are variable and depend on the fungal species used (Poyedinok and Blume, 2018). In our case, we observed that the linearization of plasmid, in combination with PEG-mediated transformation, did not yield transformants (Table 1). Other authors have observed significant decrease in transformation efficiencies when a linear vector is used (Herrera-Estrella et al., 1990). It has been argued that during PEG-mediated transformation, linear vectors would be more susceptible to degradation by exonucleases (Lopes et al., 2008), which would negatively affect the transformation efficiencies.
Finally, although we obtained transformants of P. verrucosus by electroporation of germinated conidia, this method had comparatively, a lower efficiency. Our results show that the electroporation protocol yielded an average of four transformants per experiment. Low frequencies of transformation by electroporation have been previously observed in other fungal genera (Rivera et al., 2014). These low frequencies of transformation can be attributed to different causes. One of these may be a low viability of conidia after being submitted to electrical pulses. In our experiments, we observed that after a pulse of 1.0 kV, the viability of the germinated conidia was around 38%. Other researchers have obtained high transformation efficiencies with similar levels of conidia viability (Sánchez and Aguirre, 1996; Cruz-Ramón et al., 2019), so this would not be a key factor determining the low efficiencies observed during electrotransformation of P. verrucosus. In addition, we observed that an increase in the amount of DNA used also did not increase transformation efficiencies (Table 5). Although the mechanism of electroporation of germinated conidia still remains unclear (Li et al., 2017), it has been suggested that cell wall of filamentous fungi acts as a natural barrier to uptake of DNA during electroporation (Chakraborty et al., 1991). Accordingly, we thought that probably, the cell wall of germinated conidia of P. verrucosus is a natural barrier to the uptake of DNA that the electroporation conditions used in this work cannot completely overcome. Anyway, and despite the lower efficiency of the electrotransformation method, its easiness and speed make it a good option for functional analyses where few transformants are required, such as gene overexpression.
In summary, this study describes for the first time the successful transformation of a filamentous fungus of Antarctic origin, P. verrucosus. The transformation protocols here developed will enable the genetic analysis of Pseudogymnoascus strains of Antarctic origin.
Data Availability Statement
All datasets generated for this study are included in the article/supplementary material.
Author Contributions
IV, RC, and FF designed the experiment. AD, PV, VO and CG-D performed all laboratory analysis and evaluated the data. IV and RC drafted the manuscript.
Funding
This work was financed by a grant Fondecyt 1150894. RC was supported by DICYT-USACH. VO and CG-D have received doctoral fellowship CONICYT-PFCHA/Doctorado Nacional/2018-21181056 and CONICYT-PFCHA/Doctorado Nacional/2014-63140056, respectively.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Felipe Vega and María Inés Polanco for their excellent technical assistance.
References
Alves, I. M. S., Gonçalves, V. N., Oliveira, F. S., Schaefer, C. E., Rosa, C. A., and Rosa, L. H. (2019). Diversity, distribution, and pathogenic potential of cultivable fungi present in rocks of south Shetlands archipelago, maritime Antarctica. Fungal Biol. 23, 327–336. doi: 10.1007/s00792-019-01086-8
Becker, D. M., and Lundblad, V. (2001). Introduction of DNA into yeast cells. Curr. Protoc. Mol. Biol. 27, 13.7.1–13.7.10. doi: 10.1002/0471142727.mb1307s27
Bridge, P. D., and Spooner, B. M. (2012). Non-lichenized Antarctic fungi: transient visitors or members of a cryptic ecosystem? Fungal Ecol. 5, 381–394. doi: 10.1016/j.funeco.2012.01.007
Chakraborty, B., Patterson, N. A., and Kapoor, M. (1991). An electroporation-based system for high-efficiency transformation of germinated conidia of filamentous fungi. Can. J. Microbiol. 37, 858–863. doi: 10.1139/m91-147
Cho, Y., Srivastava, A., and Nguyen, C. (2013). “Improved transformation method for Alternaria brassicicola and its applications” in Laboratory protocols in fungal biology: Current methods in fungal biology. eds. V. K. Gupta and M. G. Tuohy (New York: Springer), 395–401.
Cruz-Ramón, J., Fernández, F. J., Mejía, A., and Fierro, F. (2019). Electroporation of germinated conidia and young mycelium as an efficient transformation system for Acremonium chrysogenum. Folia Microbiol. 64, 33–39. doi: 10.1007/s12223-018-0625-0
Dantas-Barbosa, C., Araújo, E. F., Moraes, L. M., Vainstein, M. H., and Azevedo, M. O. (1998). Genetic transformation of germinated conidia of the thermophilic fungus Humicola grisea var. thermoidea to hygromycin B resistance. FEMS Microbiol. Lett. 169, 185–190. doi: 10.1111/j.1574-6968.1998.tb13316.x
Dermol, J., Pakhomova, O. N., Pakhomov, A. G., and Miklavčič, D. (2016). Cell electrosensitization exists only in certain electroporation buffers. PLoS One 11:e0159434. doi: 10.1371/journal.pone.0159434
Duarte, A., Blanco Barato, M., Suzigan Nobre, F., Polezel, D., Oliveira, T., Aparecida dos Santos, J., et al. (2018). Production of cold-adapted enzymes by filamentous fungi from King George Island, Antarctica. Polar Biol. 41, 2511–2521. doi: 10.1007/s00300-018-2387-1
Duarte, A. W. F., Dos Santos, J. A., Vianna, M. V., Vieira, J. M. F., Mallagutti, V. H., Inforsato, F. J., et al. (2017). Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 38, 600–619. doi: 10.1080/07388551.2017.1379468
Figueroa, L., Jiménez, C., Rodríguez, J., Areche, C., Chávez, R., Henríquez, M., et al. (2015). 3-Nitroasterric acid derivatives from an Antarctic sponge-derived Pseudogymnoascus sp. fungus. J. Nat. Prod. 78, 919–923. doi: 10.1021/np500906k
Furbino, L. E., Godinho, V. M., Santiago, I. F., Pellizari, F. M., Alves, T. M., Zani, C. L., et al. (2014). Diversity patterns, ecology and biological activities of fungal communities associated with the endemic macroalgae across the Antarctic peninsula. Microb. Ecol. 67, 775–787. doi: 10.1007/s00248-014-0374-9
Gangavaram, L., Mchunu, N., Ramakrishnan, P., Singh, S., and Permaul, K. (2009). Improved electroporation-mediated non-integrative transformation of Thermomyces lanuginosus. J. Microbiol. Methods 77, 159–164. doi: 10.1016/j.mimet.2009.01.025
Gomes, E. C. Q., Godinho, V. M., Silva, D. A. S., de Paula, M. T. R., Vitoreli, G. A., Zani, C. L., et al. (2018). Cultivable fungi present in Antarctic soils: taxonomy, phylogeny, diversity, and bioprospecting of antiparasitic and herbicidal metabolites. Extremophiles 22, 381–393. doi: 10.1007/s00792-018-1003-1
Gonçalves, V. N., Carvalho, C. R., Johann, S., Mendes, G., Alves, T. M. A., Zani, C. L., et al. (2015). Antibacterial, antifungal and antiprotozoal activities of fungal communities present in different substrates from Antarctica. Polar Biol. 38, 1143–1152. doi: 10.1007/s00300-015-1672-5
Henríquez, M., Vergara, K., Norambuena, J., Beiza, A., Maza, F., Ubilla, P., et al. (2014). Diversity of cultivable fungi associated with Antarctic marine sponges and screening for their antimicrobial, antitumoral and antioxidant potential. World J. Microbiol. Biotechnol. 30, 65–76. doi: 10.1007/s11274-013-1418-x
Herrera-Estrella, A., Goldman, G. H., and Van Montagu, M. (1990). High-efficiency transformation system for the biocontrol agents, Trichoderma spp. Mol. Microbiol. 4, 839–843. doi: 10.1111/j.1365-2958.1990.tb00654.x
Jakobsson, E., Arguello-Miranda, O., Chiu, S.-W., Fazal, Z., Kruczek, J., Núñez-Corrales, S., et al. (2017). Towards a unified understanding of lithium action in basic biology and its significance for applied biology. J. Membr. Biol. 250, 587–604. doi: 10.1007/s00232-017-9998-2
Kawai, S., Hashimoto, W., and Murata, K. (2010). Transformation of Saccharomyces cerevisiae and other fungi: methods and possible underlying mechanism. Bioeng. Bugs. 1, 395–403. doi: 10.4161/bbug.1.6.13257
Kochkina, G. A., Ivanushkina, N. E., Lupachev, A. V., Starodumova, I. P., Vasilenko, O. V., and Ozerskaya, S. M. (2019). Diversity of mycelial fungi in natural and human-affected Antarctic soils. Polar Biol. 42, 47–64. doi: 10.1007/s00300-018-2398-y
Li, D., Tang, Y., Lin, J., and Cai, W. (2017). Methods for genetic transformation of filamentous fungi. Microb. Cell Factories 16:168. doi: 10.1186/s12934-017-0785-7
Liu, Z., and Friesen, T. (2012). Polyethylene glycol (PEG)-mediated transformation in filamentous fungal pathogens. Methods Mol. Biol. 835, 365–375. doi: 10.1007/978-1-61779-501-5_21
Loperena, L., Soria, V., Varela, H., Lupo, S., Bergalli, A., Guigou, M., et al. (2012). Extracellular enzymes produced by microorganisms isolated from maritime Antarctica. World J. Microbiol. Biotechnol. 28, 2249–2256. doi: 10.1007/s11274-012-1032-3
Lopes, F. J. F., de Queiroz, M. V., Lima, J. O., Silva, V. A. O., and de Araújo, E. F. (2008). Restriction enzyme improves the efficiency of genetic transformations in Moniliophthora perniciosa, the causal agent of witches’ broom disease in Theobroma cacao. Braz. Arch. Biol. Technol. 51, 27–34. doi: 10.1590/S1516-89132008000100004
Loque, C. P., Medeiros, A. O., Pellizzari, F. M., Oliveira, E. C., Rosa, C. A., and Rosa, L. H. (2010). Fungal community associated with marine macroalgae from Antarctica. Polar Biol. 33, 641–648. doi: 10.1007/s00300-009-0740-0
Mach, R. L. (2003). “Transformation and gene manipulation in filamentous fungi: an overview” in Handbook of fungal biotechnology. 2nd Edn. ed. D. K. Arora (New York: Marcel Dekker Inc), 109–120.
Meilhoc, E., Masson, J. M., and Teissie, J. (1990). High efficiency transformation of intact yeast cells by electric field pulses. Nat. Biotechnol. 8, 223–227. doi: 10.1038/nbt0390-223
Navarrete, K., Roa, A., Vaca, I., Espinosa, Y., Navarro, C., and Chávez, R. (2009). Molecular characterization of the niaD and pyrG genes from Penicillium camemberti, and their use as transformation markers. Cell. Mol. Biol. Lett. 14, 692–702. doi: 10.2478/s11658-009-0028-y
Ogaki, M. B., de Paula, M. T., Ruas, D., Pellizzari, F. M., García-Laviña, C. X., and Rosa, L. H. (2019). “Marine fungi associated with Antarctic macroalgae” in The ecological role of micro-organisms in the Antarctic environment. ed. S. Castro-Sowinski (Cham: Springer Polar Sciences, Springer), 239–255.
Olmedo-Monfil, V., Cortés-Penagos, C., and Herrera-Estrella, A. (2004). Three decades of fungal transformation: key concepts and applications. Methods Mol. Biol. 267, 297–313. doi: 10.1385/1-59259-774-2:297
Pearce, D. A. (2012). “Extremophiles in Antarctica: life at low temperatures” in Adaption of microbial life to environmental extremes. eds. H. Stan-Lotter and S. Fendrihan (Vienna: Springer), 99–131.
Poveda, G., Gil-Durán, C., Vaca, I., Levicán, G., and Chávez, R. (2018). Cold-active pectinolytic activity produced by filamentous fungi associated with Antarctic marine sponges. Biol. Res. 51:28. doi: 10.1186/s40659-018-0177-4
Poyedinok, N. L., and Blume, Y. B. (2018). Advances, problems, and prospects of genetic transformation of fungi. Cytol. Genet. 52, 139–154. doi: 10.3103/S009545271802007X
Punt, P. J., Oliver, R. P., Dingemanse, M. A., Pouwels, P. H., and van den Hondel, C. A. (1987). Transformation of Aspergillus based on the hygromycin B resistance marker from Escherichia coli. Gene 56, 117–124. doi: 10.1016/0378-1119(87)90164-8
Purić, J., Vieira, G., Cavalca, L. B., Sette, L. D., Ferreira, H., Vieira, M. L. C., et al. (2018). Activity of Antarctic fungi extracts against phytopathogenic bacteria. Lett. Appl. Microbiol. 66, 530–536. doi: 10.1111/lam.12875
Rehman, L., Su, X., Guo, H., Qi, X., and Cheng, H. (2016). Protoplast transformation as a potential platform for exploring gene function in Verticillium dahlia. BMC Biotechnol. 16:57. doi: 10.1186/s12896-016-0287-4
Rivera, A. L., Magaña-Ortíz, D., Gómez, L. M., Fernández, F., and Loske, A. (2014). Physical methods for genetic transformation of fungi and yeast. Phys Life Rev 11, 184–203. doi: 10.1016/j.plrev.2014.01.007
Rodríguez-Iglesias, A., and Schmoll, M. (2015). “Transformation for genome manipulation in fungi” in Genetic transformation systems in fungi. eds. M. van den Berg and K. Maruthachalam (Cham: Springer), 21–40.
Rosa, L. H., Zani, C. L., Cantrell, C. L., Duke, S. O., Van Dijck, P., Desideri, A., et al. (2019). “Fungi in Antarctica: diversity, ecology, effects of climate change, and bioprospection for bioactive compounds” in Fungi of Antarctica. ed. L. H. Rosa (Cham: Springer), 1–17.
Roth, M. G., and Chilvers, M. I. (2019). A protoplast generation and transformation method for soybean sudden death syndrome causal agents Fusarium virguliforme and F. brasiliense. Fungal Biol. Biotechnol. 6:7. doi: 10.1186/s40694-019-0070-0
Ruiz-Díez, B. (2002). Strategies for the transformation of filamentous fungi. J. Appl. Microbiol. 92, 189–195. doi: 10.1046/j.1365-2672.2002.01516.x
Sánchez, O., and Aguirre, J. (1996). Efficient transformation of Aspergillus nidulans by electroporation of germinated conidia. Fungal Genet. Newsl. 43, 48–51.
Sautour, M., Mansur, C., Divies, C., Bensoussan, M., and Dantigny, P. (2002). Comparison of the effects of temperature and water activity on growth rate of food spoilage moulds. J. Ind. Microbiol. Biotechnol. 28, 311–315. doi: 10.1038/sj.jim.7000248
Segers, F. J., van Laarhoven, K. A., Huinink, H. P., Adan, O. C., Wösten, H. A., and Dijksterhuis, J. (2016). The indoor fungus Cladosporium halotolerans survives humidity dynamics markedly better than Aspergillus niger and Penicillium rubens despite less growth at lowered steady-state water activity. Appl. Environ. Microbiol. 82, 5089–5098. doi: 10.1128/AEM.00510-16
Tian, Y., Li, Y.-L., and Zhao, F.-C. (2017). Secondary metabolites from polar organisms. Mar. Drugs 15:28. doi: 10.3390/md15030028
Vaca, I., and Chávez, R. (2019). “Bioactive compounds produced by Antarctic filamentous fungi” in Fungi of Antarctica. ed. L. H. Rosa (Cham: Springer), 265–283.
Vela-Corcía, D., Romero, D., Torés, J. A., De Vicente, A., and Pérez-García, A. (2015). Transient transformation of Podosphaera xanthii by electroporation of conidia. BMC Microbiol. 15:20. doi: 10.1186/s12866-014-0338-8
Vieira, G., Purić, J., Morão, L. G., Dos Santos, J. A., Inforsato, F. J., Sette, L. D., et al. (2018). Terrestrial and marine Antarctic fungi extracts active against Xanthomonas citri subsp. Citri. Lett. Appl. Microbiol. 67, 64–71. doi: 10.1111/lam.12890
Wang, M., Zhang, J., Wang, L., Han, L., Zhang, X., and Feng, J. (2018). Optimization of production conditions for protoplasts and polyethylene glycol-mediated transformation of Gaeumannomyces tritici. Molecules 23:1253. doi: 10.3390/molecules23061253
Wiebe, M., Novákova, M., Miller, L., Blakebrough, M., Robson, G., and Trinci, A. P. J. (1997). Protoplast production and transformation of morphological mutants of the Quorn® myco-protein fungus, Fusarium graminearum A3/5, using the hygromycin B resistance plasmid pAN7-1. Mycol. Res. 101, 871–877. doi: 10.1017/S0953756296003425
Xu, S., Zhou, Z., Du, G., Zhou, J., and Chen, J. (2014). Efficient transformation of Rhizopus delemar by electroporation of germinated spores. J. Microbiol. Methods 103, 58–63. doi: 10.1016/j.mimet.2014.05.016
Zhang, T., Ping, R., De Jesus, M., Chaturvedi, V., and Chaturvedi, S. (2018). Green fluorescent protein expression in Pseudogymnoascus destructans to study its abiotic and biotic lifestyles. Mycopathologia 183, 805–814. doi: 10.1007/s11046-018-0285-2
Keywords: transformation, Pseudogymnoascus, protoplasts, electroporation, Antarctica
Citation: Díaz A, Villanueva P, Oliva V, Gil-Durán C, Fierro F, Chávez R and Vaca I (2019) Genetic Transformation of the Filamentous Fungus Pseudogymnoascus verrucosus of Antarctic Origin. Front. Microbiol. 10:2675. doi: 10.3389/fmicb.2019.02675
Edited by:
Stefanie Pöggeler, University of Göttingen, GermanyReviewed by:
Skander Elleuche, Miltenyi Biotec, GermanyDaniela Nordzieke, University of Göttingen, Germany
Copyright © 2019 Díaz, Villanueva, Oliva, Gil-Durán, Fierro, Chávez and Vaca. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renato Chávez, renato.chavez@usach.cl; Inmaculada Vaca, inmavaca@uchile.cl
†These authors have contributed equally to this work