
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 01 November 2019
Sec. Fungi and Their Interactions
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.02317
This article is part of the Research Topic Sensing and Signaling of the Environment by Fungi in Health and Disease View all 20 articles
 Maria Augusta C. Horta1
Maria Augusta C. Horta1 Nils Thieme1
Nils Thieme1 Yuqian Gao2
Yuqian Gao2 Kristin E. Burnum-Johnson2Carrie D. Nicora2Marina A. Gritsenko2Mary S. Lipton3Karthikeyan Mohanraj4
Kristin E. Burnum-Johnson2Carrie D. Nicora2Marina A. Gritsenko2Mary S. Lipton3Karthikeyan Mohanraj4 Leandro José de Assis5
Leandro José de Assis5 Liangcai Lin6
Liangcai Lin6 Chaoguang Tian6
Chaoguang Tian6 Gerhard H. Braus7
Gerhard H. Braus7 Katherine A. Borkovich8
Katherine A. Borkovich8 Monika Schmoll9Luis F. Larrondo10
Monika Schmoll9Luis F. Larrondo10 Areejit Samal4
Areejit Samal4 Gustavo H. Goldman5,11
Gustavo H. Goldman5,11 J. Philipp Benz1,11*
J. Philipp Benz1,11*Fungal plant cell wall degradation processes are governed by complex regulatory mechanisms, allowing the organisms to adapt their metabolic program with high specificity to the available substrates. While the uptake of representative plant cell wall mono- and disaccharides is known to induce specific transcriptional and translational responses, the processes related to early signal reception and transduction remain largely unknown. A fast and reversible way of signal transmission are post-translational protein modifications, such as phosphorylations, which could initiate rapid adaptations of the fungal metabolism to a new condition. To elucidate how changes in the initial substrate recognition phase of Neurospora crassa affect the global phosphorylation pattern, phospho-proteomics was performed after a short (2 min) induction period with several plant cell wall-related mono- and disaccharides. The MS/MS-based peptide analysis revealed large-scale substrate-specific protein phosphorylation and de-phosphorylations. Using the proteins identified by MS/MS, a protein-protein-interaction (PPI) network was constructed. The variance in phosphorylation of a large number of kinases, phosphatases and transcription factors indicate the participation of many known signaling pathways, including circadian responses, two-component regulatory systems, MAP kinases as well as the cAMP-dependent and heterotrimeric G-protein pathways. Adenylate cyclase, a key component of the cAMP pathway, was identified as a potential hub for carbon source-specific differential protein interactions. In addition, four phosphorylated F-Box proteins were identified, two of which, Fbx-19 and Fbx-22, were found to be involved in carbon catabolite repression responses. Overall, these results provide unprecedented and detailed insights into a so far less well known stage of the fungal response to environmental cues and allow to better elucidate the molecular mechanisms of sensory perception and signal transduction during plant cell wall degradation.
Fungi are cell factories with important functions for the bioeconomy since they often exhibit superior metabolic and secretory capabilities compared to bacterial and yeast-based production systems (Meyer et al., 2016). Furthermore, fungi are highly versatile in their ability to adjust their metabolism to diverse substrates. In the case of fungal utilization of complex plant biomass, the specific perception of the individual plant cell wall components is a necessary pre-requisite to adapt enzyme production and intracellular metabolism. These recognition reactions trigger a sophisticated series of intracellular signaling cascades in order to control and coordinate the cellular response. The fungal perception of plant cell wall polysaccharides in the presence of plant biomass is initiated by starvation-triggered de-repression of a large number of genes, leading to the production of low amounts of scouting enzymes responsible for plant cell wall depolymerization (van Munster et al., 2014). The resulting monomeric and/or small oligomeric soluble sugars are transferred into the cell via membrane-embedded transporters. Then these sugars (or their metabolic derivatives) can function as signaling molecules that activate signal cascades to promote specific metabolic adaptations (see e.g., Glass et al., 2013 and references therein). The recognition signals are responsible for the transcription and translation of specific genes, such as sugar transporters and enzymes necessary to depolymerize and catabolize the corresponding polysaccharide. However, the signaling reactions that occur between the first substrate contact and the gene activation by transcription factors are poorly described, making it difficult to predict good targets for strain engineering.
Reversible modifications in protein phosphorylation mediated by protein kinases or phosphatases represents a mechanism that is frequently employed by eukaryotic cells to transmit environmental cues and initiate signal transduction pathways, such as for transcription factor activation (Whitmarsh and Davis, 2000), cellular localization, protein stability, protein-protein interactions, DNA binding, and enzymatic activity (Pawson, 2007; Breitkreutz et al., 2010). Most cellular processes are in fact regulated by the reversible phosphorylation of proteins on serine (S), threonine (T), and tyrosine residues (Y) (Wart and Unit, 1993; Ficarro et al., 2002), including metabolism, movement, the circadian rhythms and many others (Wart and Unit, 1993; Cohen, 2000; Hurley et al., 2016).
At least 107 serine/threonine protein kinases and more than 30 nucleases and phosphatases are predicted in Neurospora crassa (Galagan et al., 2003; Borkovich et al., 2004; Miranda-Saavedra and Barton, 2007; Park et al., 2011), a filamentous ascomycete that has been used as a model system for genetics for many decades and has available a fully sequenced and well-annotated genome (Borkovich et al., 2004). Since N. crassa is furthermore a saprotroph growing robustly on complex lignocellulosic material (Tian et al., 2009; Seibert et al., 2016), we used this fungus in the current study to analyze to what extent post-translational protein modifications by phosphorylation are involved in metabolism-related signaling cascades leading to the perception of the individual polysaccharides (cellulose, hemicelluloses and pectin) within complex plant biomass. Little information is available to this end, however regarding the very early time points of the perception event previous work demonstrated the induction of phosphorylation events as early as 2 min after substrate addition (Xiong et al., 2014a; Nguyen et al., 2016). Many signaling pathways have already been implicated in the adjustment of fungal metabolism, which involve kinases or phosphatases.
The Mitogen-Activated Protein Kinase (MAP kinase) pathway, for example, represents a prototype of a signal distribution channel with phosphorylation being the major switch. In N. crassa, MAP kinases are known to be involved in cell-to-cell communication (Fischer et al., 2018), circadian regulation of cellular processes (Lamb et al., 2012; Bennett et al., 2013), phosphate signaling (Gras et al., 2013) as well as sensing of osmolarity and carbon (Huberman et al., 2017). Phosphorylation/dephosphorylation reactions involved in nutrient-sensing are furthermore central to the cyclic AMP (cAMP) signal transduction pathway (Aichinger et al., 1999; Lengeler et al., 2000; Ficarro et al., 2002). This pathway involves G-protein-coupled receptors (GPCRs), heterotrimeric G proteins, adenylate cyclase, cAMP-dependent protein kinases (PKA) (Xue et al., 1998; Lengeler et al., 2000; Lorenz et al., 2000) and phosphodiesterases. GPCRs are seven-helix transmembrane proteins anchored in the plasma membrane of eukaryotic cells, that are responsible for a diversity of functions, including environmental sensing, metabolism, immunity, growth, and development (Chini et al., 2013; Bock et al., 2014; Cabrera et al., 2015). However, how GPCR-mediated sugar sensing initiates the G-protein signaling reactions and then transduces this signal to the inside of the cell via heterotrimeric G proteins (Tesmer, 2010) is poorly understood. What is known is that cAMP, as the product of the adenylate cyclase, acts as an intracellular secondary messenger (Nauwelaers et al., 2006), regulating a variety of processes including plant cell wall degradation in Trichoderma, Penicillium, Aspergillus spp. and N. crassa (Farkas et al., 1990; Dong et al., 1995; Schuster et al., 2012). Low amounts of cAMP stimulate cellulase activity, whereas high levels of cAMP repress cellulase formation (Assis et al., 2015). It has been demonstrated in N. crassa that one Gα protein, GNA-1, associates with adenylate cyclase (CR-1) and is required for GTP-stimulatable activity (Kays et al., 2000). Some of the morphological aberrations of mutants in this pathway can be complemented by growth in the presence of exogenous cAMP (Kore-Eda et al., 1991; Kays et al., 2000).
Two-component regulatory systems, consisting of proteins with histidine kinase and/or response regulator domains are another system of widespread, ubiquitous signaling pathways in bacteria, slime molds, fungi and plants (Borkovich et al., 2004; Schaller et al., 2011). In fungi, all known histidine kinases are of the hybrid type (HHKs), containing both histidine kinase and response regulator domains within the same protein (Borkovich et al., 2004; Schaller et al., 2011). A histidine phosphotransfer protein (HPT) receives the phosphate from the response regulator domain of the HHK and in turn transfers this phosphate to an aspartate on a separate, terminal RR protein (Borkovich et al., 2004; Schaller et al., 2011). N. crassa possesses 11 HHKs, one HPT, two RRs (Borkovich et al., 2004) and one atypical RR, STK-12 (NCU07378) (Catlett et al., 2003; Park et al., 2011). Mutants lacking stk-12 have defects in development of aerial hyphae during conidiation (Park et al., 2011). The 11 N. crassa HHKs contain additional domains at their N-termini required for normal hyphal growth, resistance to osmotic stress and fungicides, conidial integrity, female fertility (Alex et al., 1996; Schumacher et al., 1997), normal conidiation, pigmentation and perithecial development during the sexual cycle (Froehlich et al., 2005; Barba-ostria et al., 2011). Evidence suggests that the HHK OS-1 (NCU02815) and RRG-1 act upstream of the OS-2/HOG-1 MAP kinase pathway (Jones et al., 2007) and are required for proper control of OS-2 phosphorylation during the circadian rhythm (Vitalini et al., 2007). RRG-2 possesses a Heat Shock transcription Factor (HSF) DNA binding motif and is required for normal sensitivity to oxidative stress (Banno et al., 2007).
Differential phosphorylation of key components in substrate perception was demonstrated for the cellulolytic transcription factors CLR-1, a major cellulase regulator in N. crassa (Coradetti et al., 2012, 2013) and XLR-1 (a cellulase/hemicellulase regulator conserved in fungi) (Samal et al., 2017) as well as a cellobionic acid transporter, CBT-1, after exposure to cellulose (Xiong et al., 2014a). Carbon catabolite repression (CCR) is another central regulatory mechanism involved in substrate perception and metabolic regulation found in a wide range of microbial organisms that ensures the preferential utilization of glucose over less favorable carbon sources (Aro et al., 2005; Vinuselvi et al., 2012) and therefore affects the fungal response to nutrient availability (Fernandez et al., 2012; Nguyen et al., 2016; Wang et al., 2017). Differential phosphorylation of CCR-related proteins after addition of different carbon sources was previously observed in a proteome analysis of Trichoderma reesei (Nguyen et al., 2016). In T. reesei, the phosphorylation of the transcription factor Cre1, a major mediator for CCR in fungi, at a serine in a conserved stretch within an acidic domain was furthermore found to regulate its DNA binding activity (Cziferszky et al., 2002).
Another protein class known to be involved in metabolic adjustments and CCR are F-Box proteins, as part of the SCF complex (Skp, Cullin, F-box containing complex) (Skowyra et al., 1997; Jonkers and Rep, 2009; Colabardini et al., 2012; Assis et al., 2018). This multi-protein E3 ubiquitin ligase complex catalyzes the ubiquitination of proteins destined for the proteasomal degradation process (Welchman et al., 2005; Assis et al., 2018) and is, therefore, paving the way for new cellular or metabolic states. F-box proteins are found in all eukaryotes and display a large variety of functions. In fungi, they are involved in the control of the cell division cycle, meiosis (Krappmann et al., 2006), mitochondrial connectivity, control of the circadian clock components, fungal development (Kress et al., 2012), virulence (Jöhnk et al., 2016), glucose sensing, as well as the induction of cellulolytic genes (Borkovich et al., 2004; Colabardini et al., 2012). Phosphorylation reactions are a central element of F-box protein-mediated degradation, since F-box target proteins are commonly first phosphorylated before being recognized and ubiquitinated (Jonkers and Rep, 2009). For A. nidulans, Fbx23 and Fbx47 were identified as being important for CCR, with the former working in a ubiquitin ligase complex that appears responsible for CreA transcription factor degradation under carbon catabolite-derepressing conditions (Assis et al., 2018). In T. reesei, the ubiquitin C-terminal hydrolase CRE2 and the E3 ubiquitin ligase LIM1 play a role in the regulation of cellulase gene expression (Gremel et al., 2008; Denton and Kelly, 2011; Glass et al., 2013). However, the targets of the ubiquitin pathway have not yet been identified, nor is it clear whether the ubiquitinated factor is destined for degradation by the proteasome or if ubiquitination causes a signal-specific activation (Welchman et al., 2005; Glass et al., 2013).
Despite the importance and widespread occurrence of phospho-modifications as laid out above, the identification of sites of protein phosphorylation is still a challenge (Ficarro et al., 2002), hampering the elucidation of its role in global regulatory pathways, such as during substrate perception. Making use of phospho-proteomics analysis by state-of-the-art mass-spectrometry, we aimed to overcome this limitation and determine the phosphorylation patterns during the initial 2 min of contact with new carbon sources. We thus identified a large number of such post-translational modifications in proteins related to the recognition process of plant cell wall compounds by N. crassa, such as transcription factors, GPCRs, kinases and F-box proteins. Subsequently, in silico methods enabled us to construct putative proteome-wide protein-protein interaction (PPI) maps. Physiological analyses of several components within the cAMP signaling pathway that have clear substrate-dependent phosphorylation changes indicates that they are involved in metabolic adjustments to new carbon-conditions, including CCR.
Overall, this dataset will serve as a benchmark for identification and prediction of phosphorylation reactions during the sensing and signaling of cellulosic carbon sources in filamentous fungi.
Flasks with N. crassa wild type strain (FGSC #2489) were pre-grown for 16 h on 2% sucrose medium, then washed three times with 1× Vogel’s salt solution (no carbon source added) for a total duration of 30 min before transferring to their respective carbon source: 1 mM galacturonic acid (D-GalA; Sigma Aldrich) plus 1 mM rhamnose (L-Rha; Sigma Aldrich), referred to as GalAR (D-GalA + L-Rha), 2 mM glucose (D-Glc; Sigma Aldrich), 2 mM xylose (D-Xyl; Sigma Aldrich), 0.5% cellobiose (Cel; Sigma Aldrich) and 2 mM 1,4-ß-D-glucosyl-D-mannose plus 1,4-ß-D-mannobiose (referred to as Glucomannodextrins or Gm; Megazyme). Samples for global proteome and phosphoproteome were incubated for additional 2 min. No carbon condition (NC) was incubated for 1 h after the medium switch (Figure 1A). More details about the experimental methodology for Mass Spectrometry can be found in the Supplementary Material (Supplementary Table S1).
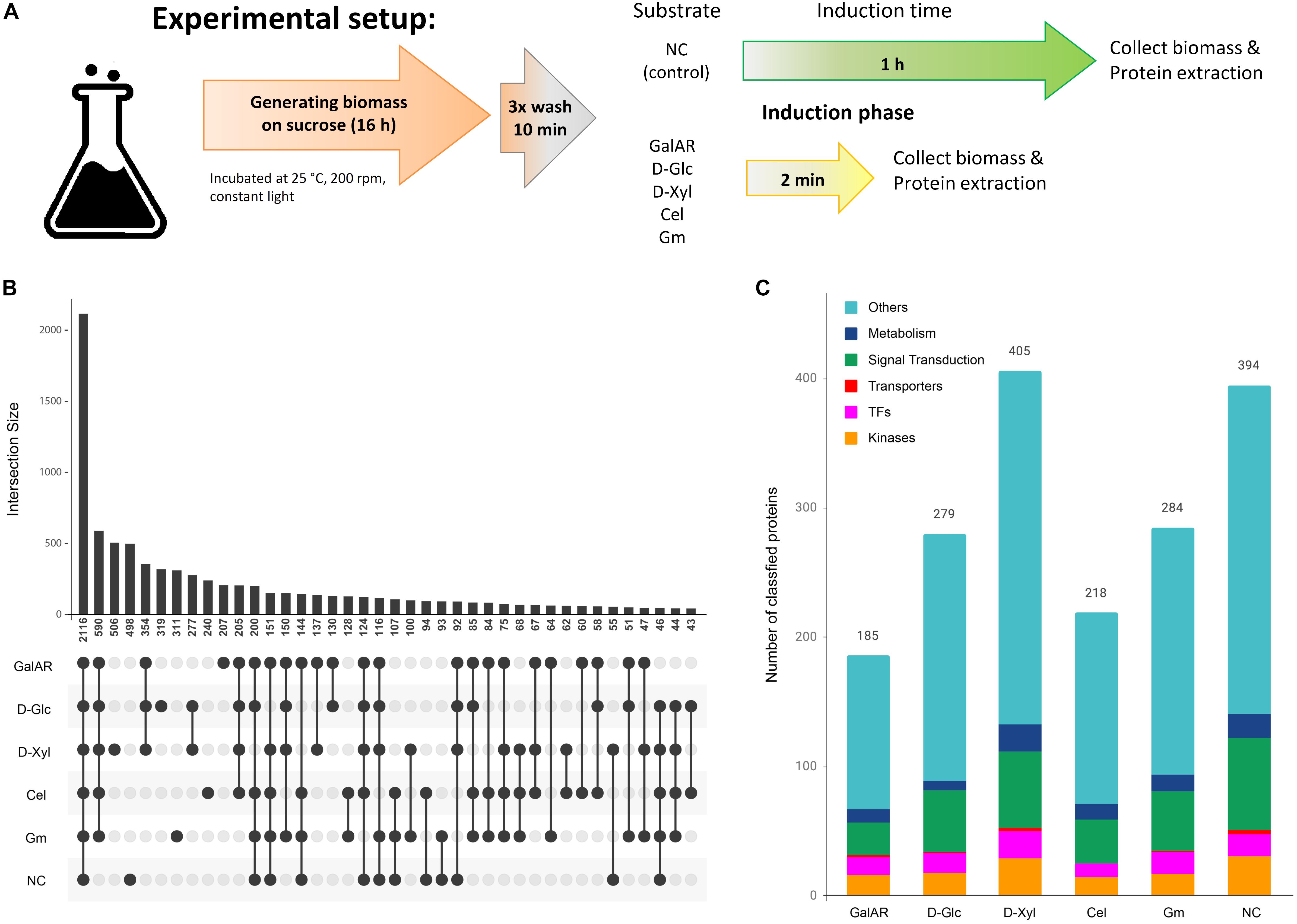
Figure 1. Proteome and phospho-proteome classification. (A) Experiment description. (B) Phospho-peptide distribution according to substrates. For this analysis, only the specific phospho-peptides (AScore > 13, P < 0.05) were considered. (C) Functional classification of proteins with specific phospho-peptides for each condition. GalAR, D-GalA + L-Rha; D-Glc, D-Glucose; D-Xyl, D-Xylose; Cel, cellobiose; Gm, Glucomannodextrins; NC, no carbon.
The samples were lysed, reduced, alkylated, and digested by trypsin. Digested samples were desalted using C18 solid phase extraction tubes (Supelco, St. Louis, MO, United States). The resulting peptide samples were concentrated and a bicinchoninic acid (BCA) assay (Thermo Scientific, Waltham, MA, United States) was performed to determine the peptide concentration and samples were diluted with nanopure water for MS analysis. 200 μg peptides aliquots for each sample were dried down for further IMAC enrichment, used for phosphoproteome analysis.
Immobilized Metal Affinity Chromatography (IMAC) enrichment of phospho-peptides follows a previously established protocol (Mertins et al., 2018). Peptides were reconstituted in 80% ACN/0.1% TFA and incubated with Fe3+-NTA agarose bead slurry. Then, the beads containing bound phospho-peptides were washed and desalted using the Stage Tips. After that, the phospho-peptides were eluted from the IMAC beads to the C18 membrane by washing the Stage Tip containing IMAC beads with 70 μl 500 mM phosphate buffer, pH 7.0 and 100 μl 1% FA before being eluted from the C18 membrane with 60 μl 50% ACN/0.1% FA. Eluted phospho-peptides were dried down and stored at −80°C until LC-MS/MS analysis.
The peptide and phospho-peptide samples were both analyzed using a nanoLC system (Waters NanoAcquity LC, Waters Corporation) coupled to a Q ExactiveTM Hybrid Quadrupole-OrbitrapTM Mass Spectrometer (Thermo Fisher Scientific) in data dependent mode. All mass spectrometry data were searched using MS-GF+ (Kim et al., 2008; Kim and Pevzner, 2014) to identify peptides by scoring MS/MS spectra against peptides derived from the whole protein sequence database. The MS-GF + results were filtered based on 1% false discovery rate (FDR) and less than 5-ppm mass accuracy. Based on spectral count MS data were considered only those peptides that were identified in at least 2 out of 4 experimental replicates with a positive spectrum, independent of the spectral count value. The phospho-peptide results were also searched using AScore algorithm (Gerber et al., 2006) to provide a probability-based score for each identified phospho-peptide. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository (Perez-Riverol et al., 2019) with the dataset identifier PXD013964 and 10.6019/PXD013964.
We constructed a system-wide network of the interactions between proteins found in the global proteome and phospho-proteome of N. crassa via prediction of functional interactions between the identified proteins, using the STRING database (Caccia et al., 2013; Szklarczyk et al., 2017). STRING database integrates and reassesses available experimental data from others curated databases, as well as information on structure, domains, function and similarity of proteins to predict functional protein–protein interactions or associations. In the STRING database, the predictions on protein-protein interactions are derived from the following sources: (i) systematic co-expression analysis, (ii) detection of shared selective signals across genomes, (iii) automated text-mining of the scientific literature and (iv) computational transfer of interaction knowledge between organisms based on gene orthology (Szklarczyk et al., 2017). While predicting interactions between N. crassa proteins using STRING database, we set the confidence score to be greater than 0.9 and the maximum of additional interactions to 10. Cytoscape was used for the visualization of the PPI network (Shannon et al., 2003). Supplementary Datasheet S1 lists the proteins used to construct the PPI network.
The WT and deletion strains were obtained from the FGSC1 (McCluskey et al., 2010). The F-box genes were named following A. nidulans (Colabardini et al., 2012; Assis et al., 2018): fbx-9 (spp-1, ΔNCU03658, FGSC#21255, Fbx9, AN10061), fbx-19 (ΔNCU08642, FGSC#15478, Fbx19, AN4510), fbx-20 (ΔNCU06250, FGSC#15629, Fbx20, AN4535) and fbx-22 (cdc4, ΔNCU05939, FGSC#13190, Fbx22, AN5517). The corresponding deletion strain for cr-1 (NCU08377) is FGSC#11514.
Neurospora crassa cultures were grown on Vogel’s minimal medium (VMM) (Vogel, 1956). Unless noted, 2% (w/v) sucrose was used as a carbon source. Strains were pre-grown on 3 ml VMM slants at 30°C dark for 48 h, then at 25°C in constant light for 4–10 days to stimulate conidia production. Conidia were inoculated into liquid media at 1 × 106 conidia/ml in individual triplicates and grown at 30°C in constant light and shaking (200 rpm) in 24-deep-well plates. VMM media containing 2% D-Xyl (Sigma Aldrich) as control and 0.2% (w/v) 2-Deoxy-D-glucose (2-DG; Sigma Aldrich) plus 2% D-Xyl were used to test 2-DG influence, evaluated after 4 days. To test allyl alcohol sensitivity, 1% D-Glc ± 100 mM of allyl alcohol (Sigma Aldrich) were incubated for 2 days.
For xylanase activity assays, the strains were grown in 1% fructose VMM during 24 h at 30°C, then transferred to 2% D-Glc plus 1% D-Xyl VMM, or 1% D-Xyl VMM for 3 days. The xylanase activity was measured in the supernatant using Azo-xylan (from Beechwood; Megazyme) as substrate. Xylanase assays were conducted by mixing 25 μl of culture supernatant with 200 μl of Azo-xylan in 50 μl 1 M sodium acetate, pH 5.0, filling up to a final reaction volume of 400 μl with acetate buffer, and incubated at 37°C for 10 min. Reactions were stopped by centrifugation at 7000 × g for 2 min and by addition of 1 ml of precipitation solution (95% v/v ethanol) to the reaction supernatants. The absorbance was measured at 590 nm.
To identify the extent of initial protein phosphorylations upon exposure to new carbon sources, we performed switch experiments, in which sucrose pre-grown N. crassa was first starved for 30 min and then incubated for 2 min with mono- and disaccharides that are known to act as signaling molecules for plant cell wall-related polysaccharides. This short retention period was deliberately chosen to allow N. crassa to take up inducer molecules, but ensure that the proteome does not have time to significantly change, so that all measured differences are due to post-translational changes as a reaction to nutrient availability and not due to changes in enzyme abundances. A large number of proteins and phospho-peptides were detected by LC-MS/MS analysis in samples from these short-exposure experiments for each induction condition (Supplementary Datasheet S2 lists the peptides and phospho-peptides identified in different experimental conditions). To aid in the confidence of the global phospho-peptide measurement, we required that peptides of interest to be quantified were present in at least 2 of the 4 biological replicates. Stringent alternative approaches to quantify proteins and phosphopeptide changes in relative abundance for Neurospora include in vivo SILAC labeling approaches (Baker et al., 2009) or isobaric labeling methods (Hurley et al., 2018). We also want to note that due to the presumed instability of histidine and aspartate phosphorylations during the enrichment strategy, we only searched for phosphorylation of serines (S), threonines (T) and tyrosines (Y) in our approach.
We identified 2116 peptides that were phosphorylated equally in all conditions, but also sets of phospho-peptides that were found to be phosphorylated specifically in only one condition: 207 in D-GalA + L-Rha, 240 in cellobiose, 311 in glucomannodextrins, 319 in D-Glc, 498 in no carbon, and 506 in D-Xyl (Figure 1B). We also studied the representation of the following functional classes among the proteins corresponding to these specific phospho-peptides: transcription factors (TFs), transporters, kinases, metabolism-associated proteins and proteins involved in signal transduction pathways (Figure 1C). The fact that a high number of these signaling-associated protein functions were present in the data sets (Table 1) strongly indicates that carbon sensing and signaling involves protein phosphorylation events as a central component in N. crassa.
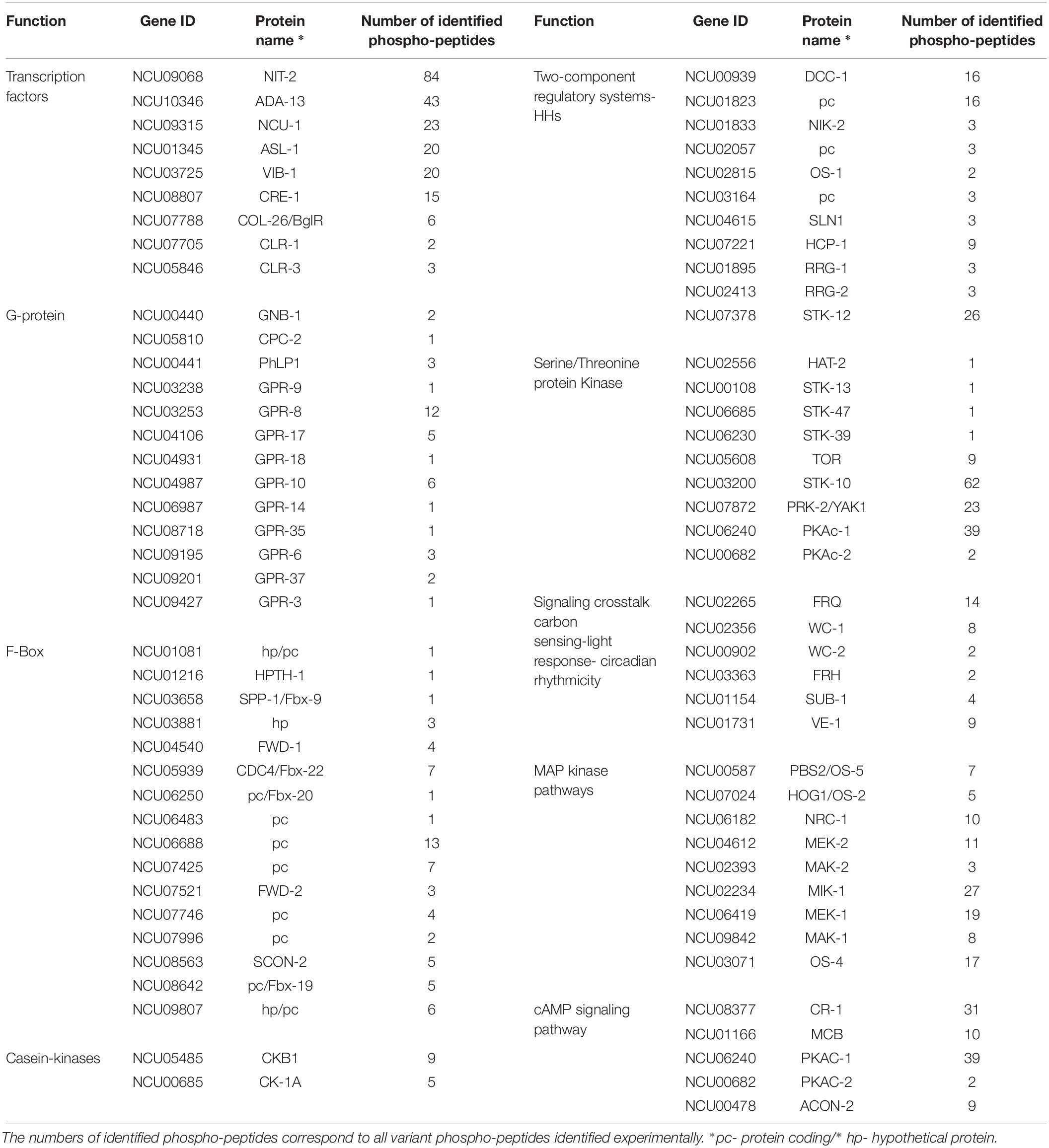
Table 1. Summary of discussed classes of genes, with the respective protein name.
A protein-protein interaction (PPI) network was constructed based on identified proteins in the proteomic and phospho-proteomic experiments, and Figure 2A visualizes this large-scale predicted network of interactions between N. crassa proteins present in the cell at the time point of the carbon switch. Thereafter we used the same functional classes (transcription factors, transporters, kinases, metabolism and signal transduction) to analyze the protein distribution across the network. The entire high-resolution network shows that proteins that belong to the same functional class have a tendency to cluster together (Supplementary Datasheet S1). Nevertheless, complex interactions between proteins belonging to different functional classes are also observed in this network, especially in the central position of the network.
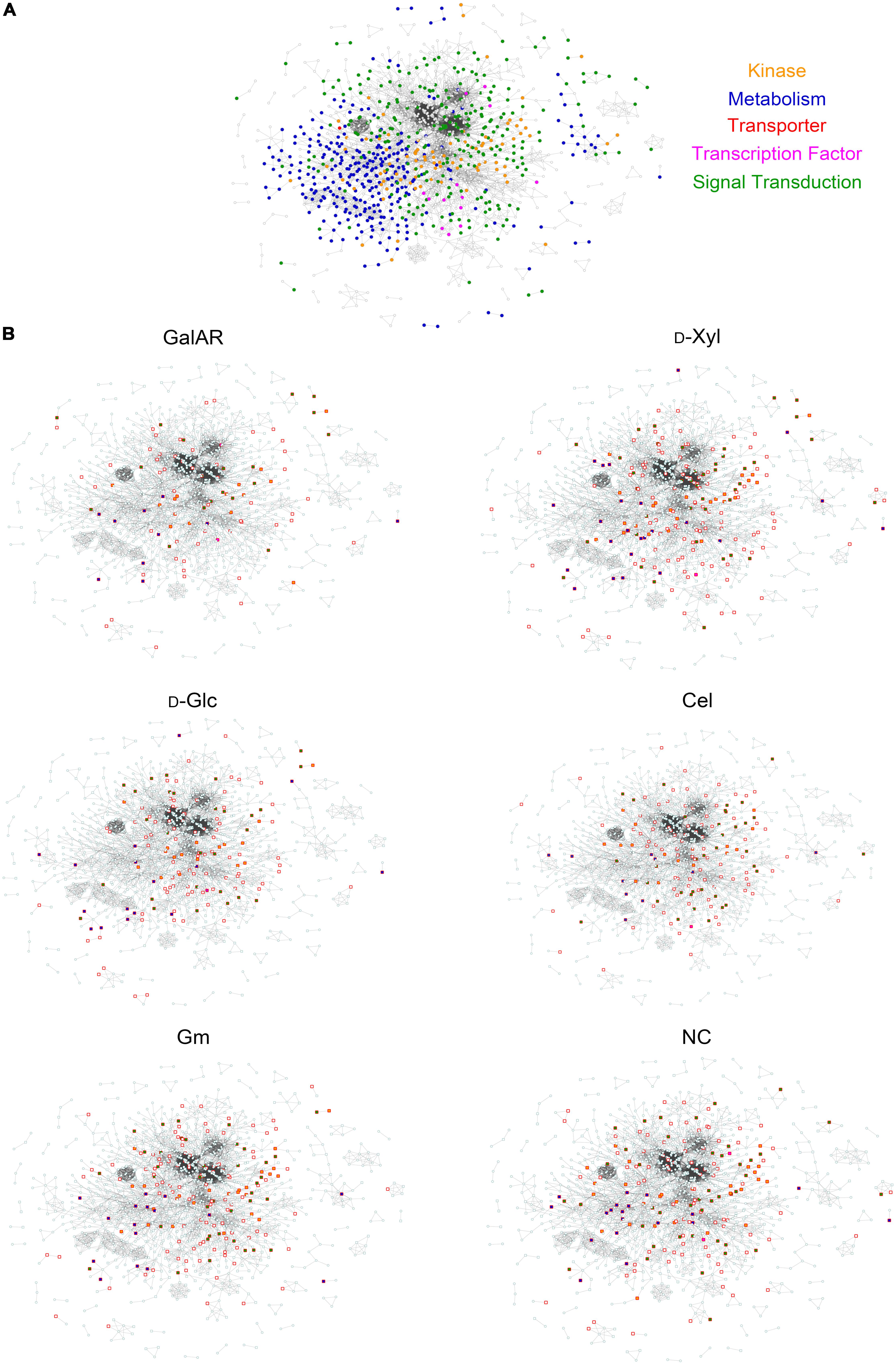
Figure 2. Protein-Protein Interaction network. (A) The PPI network of all proteins identified by proteomics and phospho-proteomics in our experiments. Functional categories related to signaling are color-coded: green, signal transduction; orange, kinases; blue, metabolism; pink, transcription factors; red, transporters. (B) Visualization of proteins in the PPI with specific phospho-peptides (independent of AScores) found in each experimental condition. Squares represent phosphorylated proteins, circles non-phosphorylated proteins. Red border-colored squares represent proteins phosphorylated specifically at the displayed condition and when filled, the proteins belong to one of the functional categories described in panel (A). GalAR (D-GalA + L-Rha), D-Glc (D-Glucose), D-Xyl (D-Xylose), Cel (cellobiose), Gm (Glucomannodextrins), NC (No carbon).
To visualize the influence of substrates on this PPI network, the proteins that were identified to be phosphorylated under each condition were highlighted within the network (Figure 2B). Interestingly, it is possible to observe differences in the profiles of phosphorylation for each substrate induction in Figure 2B. Specifically, the changes in distribution of highlighted protein nodes between conditions delineate the changing phosphorylation patterns that occur in response to each carbon source substrate (Figure 2B).
An earlier study had estimated the number of TFs in Neurospora to be 182 (Borkovich et al., 2004), a number that has been recognized as larger in recent analyses (Weirauch et al., 2014; Carrillo et al., 2017). Thus, we took a conservative list comprising 253 TFs included in the previous studies and inquired for the presence of phospho-sites in the corresponding proteins. One hundred-thirty TFs exhibited one or more phospho-peptides (Supplementary Datasheet S2), with numbers going from one up to several dozens of modified residues per TF. Thus, for example, NIT-2 (NCU09068) exhibited a total of 84 different phospho-peptides, which represents a total of 18 in vivo unambiguously modified S and T (with Ascore > 13: meaning a p-value for phosphosite localization <0.05; i.e., their precise localization within the peptide is confirmed) plus many more for which the localization is less certain. Of these, several are clustered in regions neighboring the GATA-type DNA binding domain (aa positions 742–792) including several that are phosphorylated in different residues. Determining which of the sites (many of which appear conserved in NIT2/AreaA homologs) could be playing regulatory roles, is an interesting question lying ahead.
Another TF exhibiting the second highest number of modifications is NCU10346 (ADA-13). This protein, with a predicted MYB DNA binding domain (PF00249), showed a total number of 43 phospho-peptides, comprising at least 23 phosphorylation sites.
Twenty-seven other TFs presented many identified phospho-peptides each, among which were NUC-1 (NCU09315), ASL-1 (NCU01345) and VIB-1 (NCU03725), all transcription factors important in mounting responses to changes in environmental conditions. Thus, for example VIB-1 has been associated with glucose sensing and carbon catabolite repression (Xiong et al., 2014b), playing a key role under cellulolytic conditions. The different VIB-1 phospho-peptides revealed seven different phosphorylated residues, one of which also corresponds to a Y (Y107), the latter being modified only upon contact with D-GalA + L-Rha and Glucomannodextrins. T366, located less than 20 aa downstream from the VIB-1 DNA-binding domain, appears modified only in D-Xyl.
CRE-1 (NCU08807), the yeast Mig1 ortholog that plays a key role in carbon catabolite repression, showed six clearly identifiable phosphorylated residues (with Ascore > 13) in four different regions of the protein (represented by five different peptides) including the extreme N- (S6, S11) and C-terminus. While four phospho-sites (S6, S11, S211, and T366) appeared modified in all conditions, some modifications were clearly condition-specific, such as S362 (only on cellobiose, Glucomannodextrins and without carbon source) and S209 (exclusively de-phosphorylated in cellobiose). Which phosphorylations may be modifying CRE-1 activity in Neurospora under different culture conditions remain to be determined.
COL-26/BglR (NCU07788) is a TF that has also been involved in glucose sensing/metabolism, regulation of starch degradation as well as the integration of carbon and nitrogen metabolism (Xiong et al., 2014b, 2017). Six versions of two different phospho-peptides reveal four unique phosphorylation sites arranged in two pairs (S79/S83 and S674/S676). Interestingly, in glucose and D-GalA + L-Rha, all of the phospho-sites can be observed, while in no carbon condition only S676 was found to be modified.
The cellulase transcription factor CLR-1 (NCU07705) is phosphorylated at two sites, of which S59 phosphorylation was detected under all conditions tested. Phosphorylation of T120, which lies close to the Zn2-Cys6 binuclear cluster domain and within the predicted recognition sequence of a cAMP-dependent protein kinase (PKA) phosphorylation site, occurred specifically in the presence of Glucomannodextrins and D-Xyl. This latter phospho-site had already been detected in a previous study [next to S108, which was not identified here (Xiong et al., 2014a)] to be more abundant on cellulose vs. no carbon, but found to be dispensable for activation (for a direct comparison of all peptides from that study and ours, please refer to Supplementary Datasheet S2). Of CLR-2 (NCU08042) and XLR-1 (NCU06971), which are also important regulators of cellulase gene transcription in N. crassa, no phospho-peptides were detected, peptides of these proteins were also not found in the proteomics analysis, indicating a dominating transcriptional kind of regulation, as known for CLR-2 (Coradetti et al., 2012).
Although not a transcription factor, CLR-3 (NCU05846) is still highly relevant for cellulase regulation (Huberman et al., 2017) and was found to be phosphorylated in two regions and three potential sites (Supplementary Datasheet S2). Interestingly, phosphorylation of T713 within a predicted casein kinase II phosphorylation site was specific for the absence of carbon.
Eight out of the 11 HHKs present in N. crassa (Borkovich et al., 2004) were found to be phosphorylated under at least one condition, including DCC-1/NCU00939, NCU01823, NIK-2/NCU01833, NCU02057, OS-1/NCU02815, NCU03164, NCU04615, and NCU07221 (Supplementary Datasheet S2). Of note, the majority of these HHKs (5) were proteins with at least one PAS/PAC domain: DCC-1, NIK-2, NCU02057, NCU03164, and NCU07221. Four HHKs (DCC-1, NCU01823, NIK-2 and NCU07221) were phosphorylated under all conditions tested. NCU03164 was phosphorylated under every condition except D-Glc and D-Xyl. NCU04615 is unique in only being phosphorylated in the presence of Glucomannodextrins, while this is the only condition under which OS-1 and NCU02057 were not phosphorylated.
HPT-1 (NCU01489) was not phosphorylated under any conditions in this study. RRG-1 (NCU01895) was phosphorylated under all conditions, with three sites in total. RRG-2 (NCU02413) also had two phosphorylation sites and was not phosphorylated during growth on D-Xyl. The atypical RR STK-12 (NCU07378) had 12 regions of phosphorylation and was modified under all conditions. There were two different regions of phosphorylation on protein kinase domain (around S892, T893, and S903) and one in the AGC kinase, C-terminal domain (S1238) of STK-12.
The MAPKKK SskB/OS-4 (NCU03071), representing the first step in the osmolarity pathway, was found to be phosphorylated at S409 (and potentially S407) in response to D-Glc and Glucomannodextrins, with an additional ambiguous position for cellobiose. Phosphorylation of S1223, S153, and S77 was independent of the applied conditions. Response to cellobiose and no carbon resulted in phosphorylation of T1210, while D-Xyl as well as the D-GalA + L-Rha caused phosphorylation of S123. Interestingly, Glucomannodextrins and D-Xyl elicited phosphorylation of an SSTT-motif (S1343-T1346) within a potential PEST domain (Rechsteiner et al., 1989), suggesting a role for protein degradation in Glucomannodextrins and D-Xyl dependent carbon signaling. Although the AScore for these phosphorylation sites was below 13 (above the threshold of p = 0.05; i.e., their precise localization within the peptide is not confirmed) phosphorylation of the respective peptides still indicates a relevance of the PEST domain modification.
In the MAPKK PBS2/OS-5 (NCU00587), condition-independent phosphorylation occurs at serines 219 and 346, while phosphorylation of S482 was only detected in the absence of carbon. The predicted PKC phosphorylation site at S63 is phosphorylated upon recognition of Glucomannodextrins, D-Glc and D-GalA + L-Rha. Phosphorylation of the serines at positions 77 and other sites adjacent to this was found to be condition-dependent for all conditions except no carbon indicating that this area may be a hot spot for post-translational regulation of carbon response in this kinase. HOG1/OS-2 (NCU07024), the MAP kinase of the osmolarity pathway, showed only two phosphorylated sites, one being a tyrosine. For Y173, phosphorylation was found under all conditions tested. However, phosphorylation of the adjacent T171 obviously required recognition of a carbon source, as no phosphorylated peptide was detected in the absence of carbon.
For NRC-1 (NCU06182), the MAPKKK of the pheromone response pathway, several phospho-peptides were detected. Six phosphorylated amino acids reside within predicted casein kinase phosphorylation sites and one in a PKC phosphorylation site. The serines at positions 49 and 558 were phosphorylated under all conditions tested. The predicted PKC phosphorylation site at S189 appears to be starvation-specific (no carbon) and T872 phosphorylation was only detected on cellobiose. In contrast to the osmolarity pathway, specific phosphorylations were also detected for D-GalA + L-Rha in combination with other carbon sources for NRC-1. In case of the MAPKK MEK-2 (NCU04612), we found phospho-peptides supporting the presence of both protein variants, with phosphorylation of T490 of variant 2 being present under all conditions tested. Despite detection of several phosphorylated sites, only few carbon-specific ones were found: while D-Xyl specifically elicited phosphorylation of S212, which is close to the active site, the serine at position 351 became phosphorylated upon sensing of D-Xyl, D-Glc and D-GalA + L-Rha. The MAP kinase of the pheromone response pathway, MAK-2 (NCU02393), only showed two phosphorylation sites, of which the tyrosine at position 182 was phosphorylated under all conditions tested. However, the adjacent T180 was phosphorylated only in the presence of D-Xyl and D-Glc.
In the cell integrity pathway, the MAPKKK MIK-1 (NCU02234) was found to be broadly phosphorylated under different conditions with up to 27 detected different phospho-peptides with 13 variant residues, albeit some have uncertain localization scores within peptides. In many cases, phosphorylation was not condition-dependent, but both for D-Glc (with S1215, a putative PKC site and S864, a putative casein kinase II site) and D-Xyl (with S1460, a putative cAMP-dependent site) specific sites were obvious. Phosphorylation of S450 was specific to the absence of carbon. Additionally, the region between S477 and S487 appeared to be strongly phosphorylated upon detection of carbon and may hence be a hotspot for carbon dependent regulation. Interestingly, this region overlaps with a poor PEST motif (Rechsteiner and Rogers, 1996), which hints at the involvement of protein degradation of MEK-1 (NCU06419) in regulation. Similarly, the region between amino acids 758 and 762 contains up to four phosphorylated sites specific for different conditions except “no carbon” and may as well represent such a hotspot.
In the MAPKK MEK-1, phospho-peptides were detected for different conditions, suggesting carbon-dependent phosphorylations also for this protein. Additionally, specific phosphorylations were detected for Glucomannodextrins on amino acids T92, S135, S139, and S184. The clustered serines at positions S130, S135 and S139 were phosphorylated in different combinations in various conditions and could thus represent an important area for carbon sensing. As also this region overlaps with a potential PEST motif, a contribution of protein stability or degradation in carbon sensing is further supported.
Phosphorylation of the MAP kinase MAK-1 (NCU09842) was found to occur on up to five sites, two of which being tyrosines. Thereby, phosphorylation of Y189 occurred independently of the conditions tested, while Y185 was only phosphorylated in response to D-Glc (and potentially D-Xyl and Glucomannodextrins, but with low AScores) (Supplementary Datasheet S2). T187 became phosphorylated in response to plant cell wall-related carbon sources, but not in the absence of carbon.
Previous studies showed that there are 107 serine/threonine protein kinases in the N. crassa genome (Borkovich et al., 2004; Park et al., 2011). According to our results, seventy-three out of the 107 serine/threonine protein kinases previously described were found to be phosphorylated under at least one condition. It is worth noting that HAT-2 (NCU02556) is unique in only being phosphorylated in presence of cellobiose, implying that this modification may be important for cellulose utilization. Furthermore, STK-13 (NCU00108) and STK-47 (NCU06685) were only phosphorylated in response to Glucomannodextrins and D-GalA + L-Rha, respectively. Besides that, STK-39 (NCU06230) appears modified only in starvation condition.
In eukaryotes, protein kinase A (PKA) and the phosphatidylinositol 3-kinase TOR (NCU05608) are two major kinases involved in regulation of cell growth in response to nutritional signals (Kunz et al., 1993; Roosen et al., 2005). Yak1 and Sch9 are two downstream targets of these kinase cascades (Lv et al., 2015). Recent investigation showed that TrSch9 and TrYak1 play important roles in cellulase biosynthesis in T. reesei (Lv et al., 2015). Our results demonstrated that STK-10 (NCU03200), the TrSch9 ortholog in N. crassa, exhibited the highest number of modifications upon recognition of cellobiose. This protein showed a total of 12 phosphorylated phospho-peptides (in 62 variants), which represents 22 in vivo unambiguously modified S, T and Y, suggesting that these sites may be playing regulatory roles. Similarly, PRK-2/YAK1 (NCU07872), the homolog of T. reesei TrYak1, showed 13 distinguishable phosphorylated residues (23 variants), represented in 12 different phospho-peptides under cellobiose condition. Thus, determining the function of these phosphorylation sites will be a worthwhile target for an increased scientific understanding of cellulase gene expression in filamentous fungi.
There are two catalytic subunits of PKA, namely PKAc-1 (NCU06240) and PKAc-2 (NCU00682), in N. crassa. Previous studies showed that PKAc-1 works as the major PKA in N. crassa, while deletion of pkac-2 does not affect fungal morphology, suggesting that these two isoforms may play distinct roles (Banno et al., 2005). According to our results, PKAc-1 was found to be phosphorylated under all tested conditions, while PKAc-2 was phosphorylated only in the presence of D-Glc and cellobiose (for more details, see section “CR-1 Phosphosites Identification”).
We analyzed the six G-protein subunits (Gα proteins GNA-1/NCU06493, GNA-2/NCU06729, GNA-3/NCU05206; Gβ proteins GNB-1/NCU00440 and CPC-2/NCU05810 and Gγ subunit GNG-1/NCU00041) (Muller et al., 1995; Yang et al., 2002), prosducin and 43 GPCRs, looking for phospho-sites in our dataset (Supplementary Datasheet S2). Of the six G-proteins, only the Gβ subunits GNB-1 and CPC-2 (RACK1 homolog) were phosphorylated. GNB-1 was phosphorylated under all conditions in its extreme N-terminus, while S157 of CPC-2 was found to be phosphorylated only with Glucomannodextrins or without any carbon source.
The phosducin like-protein PhLP1 (NCU00441), which acts as a co-chaperone for β and γ subunits (Willardson and Tracy, 2012) is phosphorylated at S120 and S129 independent of the carbon source. The serines at position 41 and 47 are specifically phosphorylated in presence of Glucomannodextrins.
We furthermore detected 33 distinctive phospho-peptides for 10 GPCRs. This set contains NCU03238 (GPR-9), NCU03253 (GPR-8), NCU04106 (GPR-17), NCU04931 (GPR-18), NCU04987 (GPR-10), NCU06987 (GPR-14), NCU08718 (GPR-35), NCU09195 (GPR-6), NCU09201 (GPR-37), and NCU09427 (GPR-3). Four of these (GPR-17, 18, 35, and 37) are Pth11-related GPCRs, a large class previously implicated in sensing/degradation of cellulose in N. crassa (15). GPR-17 and GPR-35 were phosphorylated under all conditions, with an extra phospho-peptide observed only in the presence of D-Xyl for GPR-17 and during starvation (no carbon source) for GPR-35. GPR-9 was phosphorylated in D-Glc, D-GalA + L-Rha and D-Xyl. GPR-18 was phosphorylated in the presence of D-GalA + L-Rha, cellobiose and Glucomannodextrins, but not D-Glc or starvation, suggesting that this GPCR is phosphorylated only during growth on plant cell wall substrates. Results from RNAseq analysis showed that gpr-18 is also among the most highly expressed of the Pth11-class GPCRs during growth on avicel (Cabrera et al., 2015).
The 10 GPCRs were variously phosphorylated in three different regions of the proteins. GPR-3, GPR-8, GPR-14, GPR-17, GPR-18, GPR-35, and GPR-37 are phosphorylated on residues in the intracellular C-terminal region. The mPR-Like/PAQR GPCRs GPR-9 and GPR-10 are phosphorylated on the first extracellular domain, while Stm1-like GPR-6 is phosphorylated in the large second intracellular loop region.
Even though Neurospora was grown under constant light conditions for the present study, the data reveal abundant phosphorylations present in clock components. Crosstalk between gene regulation in response to different nutritional conditions and the light response pathway is extensive (Stappler et al., 2017; Schmoll, 2018) which was also shown in N. crassa (Sancar et al., 2012; Schmoll et al., 2012). FREQUENCY (FRQ; NCU02265) is a key component of the Neurospora circadian clock, allowing precise daily control of several processes, including metabolism (Montenegro-Montero et al., 2015; Hurley et al., 2016). Under day-night regimes, FRQ is known to be subjected to extensive phosphorylations (>75 unambiguous S/T sites) throughout its daily cycle, many of which are key in determining the proper pace of the clock (Baker et al., 2009; Tang et al., 2009; Larrondo et al., 2015). Herein, we have confirmed the phosphorylated status of 12 of these residues (seven with AScores > 13), including the previously described T289 (Tang et al., 2009). Out of the known sites, many appear to be phosphorylated under many conditions, while others appear to be modified only upon certain media transitions (Supplementary Datasheet S2). Interestingly, the new in vivo determined phospho-site appears to be modified only under D-Glc. However, since we did not control tightly for sampling under identical time-of-day conditions, some of the observed changes might have been affected by the clock. Nevertheless, most of the sites observed in FRQ in this study have been shown to be in vitro substrates for CKI or/and CKII activity, such as S28, S145, T289, S462, S683, S685, and S950 (Tang et al., 2009).
Two other essential clock components, and also key actors in light-responses, are the transcription factors WC-1 (NCU02356) and WC-2 (NCU00902), which also impact regulation of plant cell wall degrading enzymes (Schmoll et al., 2012). WC-1 and WC-2 act as a heterodimer, forming the White Collar Complex (WCC). Our data verify discrete post-translational modifications of WC-2 in S433, a residue shown impact WCC activity and therefore circadian and light-responses (Sancar et al., 2009). Indeed, recently S433 was shown to be part of a phosphorylation cluster in WC-2, required for properly closing the circadian feedback loop (Wang et al., 2019). In the case of WC-1, three different regions were found to be phosphorylated, with one region (S1005-S1015) displaying multiple phosphorylations, particularly in D-Glc condition. These sites are located few residues distal from other phosphorylatable residues known to modulate circadian function of WC-1 (He et al., 2005; Wang et al., 2015, 2019). Some of the sites in WC-1 were recently shown to be phosporylable in vitro by CDK-1 (S831) or MAPK (S831 and S1015), without the need of a priming phosphorylation events (Wang et al., 2019). Interestingly, no phospho-peptides were detected for the LOV/PAS domain protein VVD (NCU03967) (Schafmeier and Diernfellner, 2011) which is involved in photoadaptation, despite the presence of several predicted phosphorylation sites and detection of the protein under all tested conditions. Finally, FRH (FRQ-interacting RNA helicase; NCU03363) is another key core-clock component, which in vivo studies have shown to display only one phosphorylation site (S21) (Tang et al., 2009). Herein, we observed such phosphorylation in all analyzed conditions.
The transcription factor SUB-1 (NCU01154) is predominantly known for its function in development in diverse fungi (Colot et al., 2006; Bazafkan et al., 2017), but has been shown to be connected to regulation of light response and chromatin remodeling by the white collar complex (Chen et al., 2009; Sancar et al., 2015). In SUB-1, we detected one unambiguous cellobiose-specific phosphorylation site (S381). S247 was phosphorylated under all conditions tested, while S143 was phosphorylated except upon recognition of cellobiose and in the absence of carbon, suggesting a starvation-triggered switch.
Also for VE-1 (Velvet; NCU01731), functions in development and secondary metabolism are best known (Bayram et al., 2008; Bayram and Braus, 2011), although this regulator also impacts plant cell wall degradation and light response (Bayram and Braus, 2011; Aghcheh et al., 2014). For VE-1 we detected one constant phospho-modification (S455) and two regions with potentially carbon sensing-relevant phosphorylations, one from T114 to S119, which overlaps with a poor PEST domain and a second one from T262 to T267. Interestingly, these regions contain two cellobiose-specific phosphorylations (T114 and T267, the latter with an Ascore only slightly below our threshold of 13.
The finding of clear condition-dependent phosphorylation patterns for both the photoreceptors and their regulators, such as SUB-1 or the casein kinases (see below) adds further support to the crosstalk between the reaction to carbon sources in the environment and sensing of light as well as circadian rhythmicity.
In N. crassa, casein kinases are mainly known for their function in the circadian clock (see above) with important impact on FRQ phosphorylation and stability (Diernfellner and Schafmeier, 2011), and are also know in other fungi (Al Quobaili and Montenarh, 2012; Apostolaki et al., 2012). N. crassa has one casein kinase II catalytic subunit and two casein kinase regulatory subunit as well as one casein kinase I (Borkovich et al., 2004).
The casein kinase II catalytic subunit alpha (CKA; NCU03124) did not display any detectable phosphorylation. In the casein kinase II regulatory subunit beta-1 (CKB1; NCU05485) however, S291 was found phosphorylated under all conditions tested, while S78, S101, S105, S106 and S223 show carbon-dependent phosphorylation specific to certain carbon sources (S101 and S105 for example to cellobiose and no carbon conditions). For the second casein kinase regulatory beta subunit (CKB2; NCU02754), we could not find any phosphorylated peptides, although the protein was detected under all tested conditions. The C-terminus of casein kinase I (CK-1A; NCU00685) on the other hand is phosphorylated at S313 and S320 (and potentially S312; low Ascore) independent of the condition tested, while S340 (and potentially S338; low Ascore) show carbon-specific phosphorylations including the absence of a carbon source.
The genome of N. crassa is predicted to encode for 42 F-Box proteins. For 16 of these, we were able to detect phospho-peptides under at least one condition. Since homologs for some of these phosphorylated proteins had been implicated in regulatory functions of D-Glc utilization and CCR in A. nidulans (Assis et al., 2018), the respective N. crassa deletion strains were tested in more detail. First, we tested the capability of the strains to grow on different carbon sources (Figure 3A). Particularly the deletion strain for Fbx-22 (NCU05939; CDC4) displayed an aberrant growth phenotype, with improved growth, compared to WT, on D-Glc and glucomannan, while displaying reduced growth on arabinan, xylan, and pectin. The deletion strain for Fbx-19 (NCU08642) displayed reduced growth on arabinan and pectin.

Figure 3. F-Box screening. (A) Growth phenotypes of the F-box mutants on glucose (D-Glc), xylan, arabinan, glucomannan and pectin. (B) The mutant strains were grown on 1% xylose (D-Xyl) plates. 2-DG resistance: deletion strains were grown on 2% D-Xyl as control and 2% D-Xyl + 0.2% 2-DG. Allyl alcohol (AA)–sensitivity: deletion strains were grown on 1% D-Glc as control and 1% D-Glc + 100 mM AA. (C) Xylanase activities of F-box deletion strains of N. crassa and A. nidulans. Error bars represent standard deviation from triplicate experiments. Statistical differences were calculated between the mutants and WT within each treatment using t-test (∗p < 0.05, ∗∗p < 0.001).
To test for a potential role in CCR, we next evaluated the resistance of WT and the mutants to 2-deoxy-glucose (2-DG) and allyl alcohol (AA) (Figure 3B and Supplementary Image S1). 2-DG is a non-metabolizable analogue of D-Glc that is often used to indicate impaired glucose repression in filamentous fungi (Allen et al., 1989; Xiong et al., 2014b; Assis et al., 2018). When functional, 2-DG is phosphorylated and activates CCR, resulting in the inability of the strain to grow on alternative carbon sources. When mutant strains are impaired in CCR, however, they become resistant to 2-DG. Of the four tested strains, the Δfbx-22 mutant showed some 2-DG resistance when 2% D-Xyl was used as an alternative carbon source in the presence of 2-DG (Figure 3B). AA is also used for assessing CCR, as reported for M. oryzae (Fernandez et al., 2012) and A. nidulans (Colabardini et al., 2012; Assis et al., 2018). When CCR is impaired, alcohol dehydrogenases are expressed despite the presence of D-Glc and will convert AA into the toxic compound acrolein. Thus, strains with impaired CCR exhibit AA sensitivity (Xiong et al., 2014b). As observed, the mutants Δfbx-9 and Δfbx-20 were insensitive, similar to WT. The mutants Δfbx-19 and Δfbx-22, on the other hand, were sensitive to AA (Figure 3B). These data indicate that F-Box proteins Fbx-19 and particularly Fbx-22 might be involved in the correct signaling of, or adjustment to, conditions of high D-Glc concentrations.
The same AA-sensitivity with reduced growth and conidiation was also described for both homologous genes in A. nidulans (Assis et al., 2018). The deletion of these genes in the A. nidulans Δfbx19 and Δfbx22 strains furthermore led to reduced xylanase activity compared to the WT on D-Xyl (Figure 3C). In summary, these phenotypes strongly suggest an at least partially conserved defect of CCR control for Fbx-22 as well as potentially Fbx-19.
In the adenylate cyclase CR-1 (NCU08377), a central component of the cAMP signaling pathway known to be involved in nutritional sensing (Terenzi et al., 1979; Li and Borkovich, 2006; Wang et al., 2017), we identified 18 unambiguous phosphorylated residues on 31 variant peptides (Figure 4A and Supplementary Datasheet S2). Moreover, we found support for both isoforms of CR-1 with specific phosphorylation in both variants. CR-1 is heavily phosphorylated in both variants under different conditions. Phosphorylation of S67 was specific for cellobiose while T885 was specifically phosphorylated with D-GalA + L-Rha in variant 2. Glucose-specific phosphorylation only occurred in variant 1 at T191 and in the absence of carbon at S161 and S163 (only variant 1). For D-Xyl, specific phosphorylation occurred at T159 in variant 1 and around positions and in a different region at variant 2. Besides the support for two variants of adenylate cyclase by confirmation at the peptide level, these phosphorylation patterns indicate specific functions in carbon sensing and signaling of individual compounds for both variants. Consequently, N. crassa likely distinguishes these carbon sources and triggers specific responses also in terms of alternative splicing and coordinated phosphorylation for their utilization or degradation. Besides CR-1 as the core component, the cAMP pathway further comprises the functionalities of phosphodiesterases, which degrade cAMP in response to environmental signals, and protein kinase A (PKA; see also section “Serine/Threonine Protein Kinases are Variably Phosphorylated Under Different Carbon Conditions”). MCB (NCU01166), the regulatory subunit of PKA, showed phosphorylation in two regions of the N-terminus. In most cases, phosphorylation occured upon sensing of several C-sources or constitutively (e.g., T38, T40). However, phosphorylation of S41 was specific for recognition of D-Xyl and cellobiose. Phosphorylation of the catalytic subunit PKAC-1 (NCU06240) occured predominantly in two areas of the protein: from T19 to T32, containing eight putative phosphorylated sites, and from T377 to Y386. Both regions are overlapping with potential or poor PEST regions. In many cases, multiple parallel phosphorylations occured on several of the tested carbon sources. In contrast to PKAC-1, the second catalytic subunit of protein kinase A, PKAC-2 (NCU00682) showed only two phosphorylated amino acids. Albeit both having low AScores, phosphorylation of Y255 specifically occured in the presence of cellobiose and phosphorylation of T260 was specific for recognition of D-Glc.

Figure 4. Carbon source-dependent changes of CR-1 interactions. (A) Phospho-site distribution of CR-1 in response to substrate conditions. Picture shows all possible sites of phosphorylation. GalAR (D-GalA + L-Rha), D-Glc (D-Glucose), D-Xyl (D-Xylose), Cel (cellobiose), Gm (Glucomannodextrins), NC (No carbon). Colored circles represent sites with Ascore > 13, and gray circles with AScore < 13. (B) Set of proteins directly interacting with CR-1. Colors according to functional categories: green, signal transduction; orange, kinases; blue, metabolism. (C) Sub-PPI-networks of CR-1 nodes represent proteins that interact with CR-1, those peptides were found specific phosphorylated at condition. Colors according to functional categories described in panel (B). (D) Mycelial biomass (dry weight) of the Δcr-1 mutant relative to on D-Glc, xylan, arabinan, glucomannan and pectin. Error bars represent the standard deviation from triplicate experiments. Statistical differences were calculated using t-test (∗∗∗p < 10E-5).
The high affinity phosphodiesterase ACON-2 (NCU00478) showed phosphorylation at several sites, in most cases relatively unspecific with respect to the carbon source (particularly S727 and S218). Nevertheless, carbon-specific phosphorylation was observed for Glucomannodextrins at T226 and D-Xyl as well as no carbon at S797, a site that overlaps with a predicted cAMP-dependent phosphorylation site. The low affinity phosphodiesterase PUR-1 (NCU00237) was not detected in the proteome.
Within the PPI, CR-1 is predicted to interact with 67 different proteins that form a complex PPI sub-network with a total of 265 connections (Figure 4B). For the visualization of potential carbon source-specific interactions around CR-1, we extracted from the entire network the phosphorylated CR-1-interacting proteins at each condition (Figure 4C). The sub-networks show the different possibilities of phosphorylation cascades that CR-1 might be involved in according to each induction condition. The predicted interactions are in line with the previously mentioned critical function of this gene in the regulation of growth and utilization of carbon sources (Kore-Eda et al., 1991; Lee et al., 1998). This is exemplified by the enrichment analysis (Ruepp et al., 2004) of specific functional categories within the proteins of the CR-1 sub-networks (Supplementary Datasheet S3). For the sub-network from D-Xyl-induction, the most significantly enriched class was protein kinase (30.01.05.01; p-value 1.7E-16), while cAMP/cGMP-mediated signal transduction (30.01.09.07; p-value 7.7E-13) was dominating the Glucomannodextrins-induced subset (next to protein kinases, but with lower p-value: 1.0E-09). For the Cellobiose-induced subset, the top 7 classes were related to activities in the nucleus, such as DNA/RNA synthesis, modification and control (42.10.03, 42.10, 11.02.03, 10.01.09.05, 10.01.09, 11.02, 11.02.03.04, respectively; p-values 3.9E-13, 2.3E-12, 1.5E-11, 1.9E-11, 2.4E-11, 2.9E-11, 7.6E-11), followed by signaling pathways (30.01.09.07, 30.05.01.12.05, 30.05.01.12; p-values 7.7E-11, 3.6E-10, 1.5E-09). In the D-Glc sub-network, nucleus regulation, metal binding and serine/threonine kinase activity were overrepresented (42.10, 42.10.03, 16.17, 30.01.05.01.06; p-values 1.7E-10, 7.4E-10, 7.4E-10, 9.2E-10) plus the protein kinase class, cAMP/cGMP-mediated signal transduction and MAPKKK cascade (30.01.05.01, 30.01.09.07, 30.01.05.01.03; p-values 5.8E-9, 7.8E-9, 9.5E-9, indeed, a connection between MAPK and PKA pathways was described in A. fumigatus (Assis et al., 2015). The induction condition that seemed to trigger a high number of different pathways related to signaling and phosphorylation was D-GalA + L-Rha. In this condition, receptor-mediated signaling (30.05.01; p-value 5.5E-14), protein kinase (30.01.05.01; p-value 4.2E-13) and phosphate metabolism (01.04; p-value 3.43E-12) were dominating.
The analysis of the growth phenotype of the respective Δcr-1 deletion strain on different substrates demonstrated a substantially decreased capacity to degrade complex substrates (Figure 4D), further corroborating the role of CR-1 as a central hub in the cAMP signaling pathways involved in the sensing and response to environmental conditions.
Numerous studies have indicated that rapid and reversible protein phosphorylations are involved in many perception pathways of environmental conditions across the tree of life (Cohen, 2000; Whitmarsh and Davis, 2000; Hornbeck et al., 2004; Sugiyama et al., 2008; Nakagami et al., 2010; Lenoir et al., 2018). Also in N. crassa, differential phosphorylation during growth on crystalline cellulose, sucrose and carbon-free medium has been described (Xiong et al., 2014a). In the present study, we were specifically interested in the initial events of signaling at the onset of perception. For this purpose, we chose to study an extremely early time point (only 2 min after induction), which would be too short for substantial changes on either the transcriptome or proteome levels (Nguyen et al., 2016), but would give us an exclusive insight into rapid post-translational changes. To allow for a fast signal perception, we utilized soluble mono- and oligosaccharides that are known to act as inducing molecules in fungi, signaling the presence of the respective major plant cell wall polysaccharides: cellobiose for cellulose (Znameroski et al., 2012), D-Xyl for xylan (Sun et al., 2012), D-galacturonic acid and L-rhamnose for pectin (Vries et al., 2002; Thieme et al., 2017), and Glucomannodextrins for mannan (Ogawa et al., 2012). D-Glc and no carbon (starvation) were used as further controls. This extended set of induction conditions allowed us for the first time to identify signaling events that are specific for each of the cues derived from the individual components of a plant cell wall.
To better visualize the complex interplay of phosphorylation reactions, we constructed a protein-protein interaction (PPI) network with all detected proteins, exhibiting 12,066 predicted interactions based on available information gathered from STRING (Szklarczyk et al., 2017). The protein interaction prediction can infer physical and functional protein interactions from the genomic context (Shoemaker and Panchenko, 2007; Browne et al., 2010). PPI networks have already been used to provide insights into protein function and association with complex diseases (Kovács et al., 2019), fungal secretion pathways (Petranovic et al., 2013), identification and association of protein phenotypes (Hu et al., 2011) as well as domain interactions (Browne et al., 2010). Our PPI network represents the first view of possible N. crassa protein interactions at the early stage of substrate recognition. While the principal protein classes that putatively participate in the initial stages of substrate recognition (such as kinases, transcription factors, etc.) were found to be clustered across the network, numerous interactions between proteins of different functions are also represented. These interactions include many variances in phosphorylated proteins that can be hypothesized to be part of signaling pathways leading to metabolic adaptations. Detecting the vast changes of phosphorylation patterns across the network in response to different carbon sources strongly indicates that phosphorylation reactions are a central part of the environmental response of N. crassa that can be specifically adjusted to the composition of the substrate.
For transcription factors, kinases and other signaling components, such as CR-1, G-proteins and GPCRs, the phosphorylation profile is clearly affected by the substrate composition, providing novel insights into several well-known signaling pathways. For example, the differential phosphorylation of VIB-1, that acts upstream of CRE-1 (Xiong et al., 2014b), could be a mode of action how this TF is activated or inactivated and thus be involved in the modulation of the activity of CRE-1, potentially supporting the lignocellulolytic response. Further downstream in this regulatory cascade, the cellulase transcription factor CLR-1 shows condition-dependent phosphorylation as well. The regulatory impact of CLR-1 on the transcription factor CLR-2 (Coradetti et al., 2012) could be modulated by this phosphorylation and hence adjusted to the available substrate. As CLR-1 binds to its target promoters also under non-inducing conditions (Coradetti et al., 2012; Craig et al., 2015), the observed phosphorylation in presence of Glucomannodextrins and D-Xyl could help to keep CLR-1 de-activated in non-inducing situations.
To a large extent, our experiments enable the analysis of the phosphorylation reactions at amino acid resolution (reliably for all phospho-peptides with AScores > 13, meaning a p-value for the phospho-site localization <0.05). Regarding the G-proteins, RACK1-homologs have been demonstrated to function as an alternative Gβ in several fungi and are well-known for their additional role as scaffolding proteins for numerous processes in eukaryotes (Ron et al., 2013; Zhang et al., 2016). In N. crassa, the RACK1 homolog cross-pathway control-2 (CPC-2) is required for proper expression of numerous amino acid biosynthetic enzymes in response to starvation for a single amino acid (Muller et al., 1995). Of interest, one of two residues that are phosphorylated by the WNK8 kinase in plant RACK1 proteins is T162 in the fourth WD40 repeat (Chen, 2015; Urano et al., 2015), just three residues away from the N. crassa phospho-site, which is also found during carbon starvation (no carbon condition). Genetic evidence suggests that phosphorylation of RACK1 on S122 and T162 negatively regulates its functions (Urano et al., 2015).
In terms of Gβ, studies in mammals have shown that morphine stimulation leads to phosphorylation and adenylate cyclase activity (Chakrabarti and Gintzler, 2003). Although the causative phospho-site has not been identified, phospho-proteomics studies have identified phosphorylated S residues in the N-terminus at positions 2, 72 and 74 (Chakravorty and Assmann, 2018), analogous to the N-terminal phosphorylation observed in this study. In S. cerevisiae, the Gβ Ste4p is phosphorylated, but on a T and S residue near the C-terminus of the protein (Chakravorty and Assmann, 2018). This region is not conserved in Gβ subunits from mammals or plants, and yeast mutants lacking both phosphorylation sites have relatively subtle defects in cell polarization during the pheromone response (Chakravorty and Assmann, 2018). In the crystal structure of the mammalian Gαβγ heterotrimer, the N-termini of the Gβ and Gγ form interacting α-helices are important for dimerization (Moreira, 2014). Thus, it is intriguing that Gβ proteins from both N. crassa and mammals, but not S. cerevisiae, are phosphorylated in this region, perhaps influencing the association of the Gβ and Gγ subunits.
Also for the histidine kinase two-component systems, the identified phospho-sites within the domain structure of the proteins provide novel information that might be helpful to better understand their molecular setup. We found that a few of the N. crassa HHKs had at least one S/T phosphorylation site within a conserved motif in the protein. S/T phosphorylation of a plant HHK has been demonstrated for the cytokinin receptor AHK2 in Arabidopsis thaliana (Sugiyama et al., 2008; Nakagami et al., 2010; Dautel et al., 2016). One of these studies found that T4, S596 and T740 were phosphorylated, the latter two residues within the HK domain of AHK2 (Dautel et al., 2016). In N. crassa, two of the seven detected phospho-sites in DCC-1 (NCU00939) were identified within the RR domain, with S1338 and S1359 being phosphorylated under all conditions. NCU01823 was phosphorylated in different regions with two sites in the Orc1-like AAA ATPase domain, S722 and S723. Regarding NIK-2 (NCU01833), one phospho-peptide was detected at the extreme 3′ end of the first PAS/PAC domain being phosphorylated under all conditions. NCU03164 displayed four phosphorylation sites and is the only HHK with a site (S966) in the HK domain, being phosphorylated in presence of D-GalA + L-Rha, cellobiose, Glucomannodextrins and no carbon.
With the MAP kinase pathway, we evaluated a prototypical threefold phosphorylation cascade impacting osmosensing, pheromone response and cell wall integrity. According to Huberman et al. (2017), a relevant connection between carbon sensing and osmolarity pathway was described in N. crassa. The osmosensing (OS) MAPK pathway has been shown to act as a negative regulator of cellulase production in N. crassa, with the suggestion that osmolarity serves as a proxy for soluble sugars (Huberman et al., 2017). The Hog1 MAPK and several upstream components of the osmosensing signaling cascade are also required for normal expression of cellulases in Trichoderma (Wang et al., 2018). We found changes in phosphorylation across the whole MAP kinase pathway, with particularly heavy phosphorylation in the MAP KKK proteins. In contrast, MAP kinases HOG1, FUS3, and SLT2 contained only few phosphorylated sites. Carbon source-specific phosphorylation was detected for all the pathways, indicating that carbon sensing involves modification of all three MAP kinase pathways. Moreover, for several regions with increased phosphorylation frequency, we detected overlaps with PEST domains (Rechsteiner et al., 1996). Consequently, not only specific phosphorylation, but also protein degradation is likely to play a role in carbon sensing via the MAP kinase pathways. In many cases phospho-peptides with one phosphorylation or two phosphorylations were detected for MAP kinases, while multiple phosphorylation sites occurred in MAPKK and MAPKKK proteins. Hence, the conditions were selected correctly, i. e. during the process of signal recognition, in which the cascade is postulated to mediate the phosphorylation through the cascade from the MAPKKK (abundant phosphorylation) to the MAP kinase (moderate phosphorylation).
Another important class of proteins that also seems under regulation by phosphorylation are the F-Box proteins. Normally, F-box proteins are substrate receptors that recruit phosphorylated substrates, whereas phosphorylation of the F-box protein itself (due to lack of substrate) often results in autoubiquitination (by the SCF) and self-destruction. However, phosphorylation of fungal F-box proteins can also affect other cellular functions, such as localization (Jöhnk et al., 2016). This study reveals specific phosphorylation of Fbx-22 (CDC-4) and a strong phenotype in the respective deletion strain, indicating that this gene has a critical function in CCR regulation that appears partly conserved in N. crassa and A. nidulans. The visualization of the Fbx-22 interaction network predicts that the protein might interact with two different clusters of proteins depending on the substrate conditions (Supplementary Image S2). It remains to be determined how these interactions can influence the transcriptional response in the context of CCR regulation, showing a complex coordination of CCR by multiple signals.
Visualization of the entire network can show differences in phosphorylation patterns across conditions, but cannot give a precise idea about the proteins involved in the interactions. We therefore decided to do a detailed analysis of the interactions of CR-1, since this enzyme is essential to the cAMP-dependent protein kinase pathway that is involved in cellulase and xylanase production (Assis et al., 2015). Especially for CR-1, the phosphorylation patterns changed drastically across the conditions. Interestingly, a major “hot spot” of phosphorylation appears to lie between two LRR (leucine-rich repeat) regions (Figure 4A). This motif is present in proteins with diverse functions and provides a versatile structural framework for the formation of protein-protein (Kobe and Deisenhofer, 1995; Kobe and Kajava, 2001) and ligand binding sites (Liu et al., 2017). As the structure of LRR motifs can lead to a conformational flexibility necessary to the protein interaction (Kobe and Deisenhofer, 1995), the phosphorylation in between these regions might also affect the binding properties by influencing the conformational structure of the binding sites. The phosphorylation of Ser/The residues between subdomains of LRR-RLK (leucine-rich repeat receptor-like protein kinase) was also previously described as being essential for catalytic activation of some protein kinases (Bojar et al., 2014; Liu et al., 2017).
Similarly intriguing, the identifiable sub-networks of CR-1 were found to have a specific design according to the interacting proteins with condition-specific phosphorylations. Extending the previously described importance of adenylate cyclase in development and growth (Kore-Eda et al., 1991; Ivey and Hoffman, 2005; Nauwelaers et al., 2006), our results therefore indicate that differently phosphorylated forms of CR-1 can interact with many other proteins and trigger specific phosphorylation cascades, depending on the composition of the medium.
The present study reveals fundamental changes in global protein phosphorylation during the initial moments of substrate recognition in N. crassa. The identification of protein phosphorylation followed by the prediction of protein-protein interactions allowed us to visualize the condition-specific phospho-protein distribution within the network. Many of the identified phospho-proteins are associated with sensing and signaling functions in the cell and thus appear to be involved in the regulatory networks governing the specific metabolic adaptation to plant cell wall polysaccharide degradation. Our protein-protein-interaction network analysis highlights that these changing patterns might lead to vast alterations in the respective sub-networks, as is the case for the adenylate cyclase CR-1, identified as a central hub with variances in phosphorylation patterns in the tested conditions. In many cases, the analysis provided amino acid level resolution, allowing to make assumptions about carbon source-specific structure-function relationships. Overall, the taken approach can therefore greatly help to identify novel functional interactions related to substrate signaling, and the experimental data can serve as a resource for future efforts to elucidate the underlying molecular mechanisms.
The datasets generated for this study have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD013964 and 10.6019/PXD013964.
MH, NT, CT, LL, AS, and JB conceived the study, and each designed and supervised substantial parts of it. MH, NT, YG, KB-J, CN, MG, ML, and LA performed the experiments and acquired the data. All authors helped to analyze and interpret the data, read and approved the final manuscript. MH, NT, KB, MS, LL, LFL, and JB drafted the manuscript, which was critically revised by LA, CT, GB, AS, and GG.
JB was supported by the DFG grant # BE6069/3-1. LFL was supported by CONICYT/FONDECYT grant # 1171151. LA was supported by 2014/FAPESP grants # 2016/007870-9 and # 2017/23624-0. LL and CT were supported by the National Natural Science Foundation of China, grant # 31761133018. KB acknowledges the NIGMS GM068087 and GM086565, and NIFA Hatch Project #CAR-PPA-6980-H. MH thanks Prof. Anete Pereira de Souza (UNICAMP, Brazil) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Computational Biology Program (CAPES Foundation, Brazil) for the post-doctoral fellowship (88881.161048/2017-01). GG acknowledges support by the Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP, Brazil), the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil), and the Technical University of Munich – Institute for Advanced Study (TUM-IAS), funded by the German Excellence Initiative. LFL thanks for support by the iBio, Iniciativa Científica Milenio (MINECON, Chile), CONICYT/FONDECYT (Chile), and the Howard Hughes Medical Institute (United States).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We are grateful for excellent technical and computational assistance by Petra Arnold, Sabrina Paulus, Olaf Strehl, and Yuxin Zhang (TUM). We would like to thank N. Louise Glass (UC Berkeley, LBNL) and Qian Liu (TIB, China) for critical reading of the manuscript. We also acknowledge the Fungal Genetics Stock Center (FGSC; Manhattan, KS, United States) for providing their services and strain collection. A portion of this research was performed by using Environmental Molecular Sciences Laboratory (EMSL), a national scientific user facility sponsored by the DOE’s Office of Biological and Environmental Research located at Pacific Northwest National Laboratory (PNNL). PNNL is a multi-program national laboratory operated by the Battelle for the Department of Energy (DOE) under Contract DE-AC05-76RLO 1830.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02317/full#supplementary-material
Supplementary Datasheet S2 contains an interactive tool to filter the detected phosphopeptides.
Aghcheh, R. K., Nemeth, Z., Atanasova, L., Fekete, E., Paholcsek, M., Sandor, E., et al. (2014). The VELVET A Orthologue VEL1 of Trichoderma reesei regulates fungal development and is essential for cellulase gene expression. PloS One 9:e112799. doi: 10.1371/journal.pone.0112799
Aichinger, C., Kahmann, R., Feldbrügge, M., and Kahmann, R. (1999). The MAP kinase Kpp2 regulates mating and pathogenic development in Ustilago maydis. Mol. Microbiol. 34, 1007–1017. doi: 10.1046/j.1365-2958.1999.01661.x
Al Quobaili, F., and Montenarh, M. (2012). CK2 and the regulation of the carbohydrate metabolism. Metabolism 61, 1512–1517. doi: 10.1016/j.metabol.2012.07.011
Alex, L. A., Borkovich, K. A., and Simon, M. I. (1996). Hyphal development in Neurospora crassa: involvement of a two-component histidine kinase. Proc. Natl. Acad. Sci. U.S.A. 93, 3416–3421. doi: 10.1073/pnas.93.8.3416
Allen, K. E., Mcnally, M. T., Lowendorf, H. S., Slayman, C. W., and Free, S. J. (1989). Deoxyglucose-resistant mutants of Neurospora crassa: isolation, mapping, and biochemical characterization. J. Bacteriol. 171, 53–58. doi: 10.1128/jb.171.1.53-58.1989
Apostolaki, A., Harispe, L., Calcagno-Pizarelli, A. M., Vangelatos, I., Sophianopoulou, V., and Arst, H. N. Jr. (2012). Aspergillus nidulans CkiA is an essential casein kinase I required for delivery of amino acid transporters to the plasma membrane. Mol. Microbiol. 84, 530–549. doi: 10.1111/j.1365-2958.2012.08042.x
Aro, N., Pakula, T., and Penttilä, M. (2005). Transcriptional regulation of plant cell wall degradation by filamentous fungi. FEMS Microbiol. Rev. 29, 719–739. doi: 10.1016/j.femsre.2004.11.006
Assis, L. J., De Ries, L. N. A., Savoldi, M., Reis, T. F., Brown, N. A., and Goldman, G. H. (2015). Aspergillus nidulans protein kinase A plays an important role in cellulase production. Biotechnol. Biofuels 8, 1–20. doi: 10.1186/s13068-015-0401-1
Assis, L. J., Ulas, M., Ries, L. N. A., Ramli, N. A. M. El, Sarikaya-Bayram, O., Braus, G. H., et al. (2018). Regulation of Aspergillus nidulans CreA-mediated catabolite repression by the F-Box Proteins Fbx23 and Fbx47. mBio 9:e00840-18. doi: 10.1128/mBio.00840-18
Baker, C. L., Kettenbach, A. N., Loros, J. J., Gerber, S. A., and Dunlap, J. C. (2009). Quantitative proteomics reveals a dynamic interactome and phase-specific phosphorylation in the Neurospora circadian clock. Mol. Cell 34, 354–363. doi: 10.1016/j.molcel.2009.04.023
Banno, S., Noguchi, R., Yamashita, K., Fukumori, F., Kimura, M., Yamaguchi, I., et al. (2007). Roles of putative His-to-Asp signaling modules HPT-1 and RRG-2, on viability and sensitivity to osmotic and oxidative stresses in Neurospora crassa. Curr. Genet 51, 197–208. doi: 10.1007/s00294-006-0116-8
Banno, S., Ochiai, N., Noguchi, R., Kimura, M., Yamaguchi, I., Kanzaki, S., et al. (2005). A catalytic subunit of cyclic AMP-dependent protein kinase, PKAC-1, regulates asexual differentiation in Neurospora crassa. Genes Genet. Syst. 80, 25–34. doi: 10.1266/ggs.80.25
Barba-ostria, C., Lledías, F., and Georgellis, D. (2011). The Neurospora crassa DCC-1 protein, a putative histidine kinase, is required for normal sexual and asexual development and carotenogenesis. Eukaryot. Cell 10, 1733–1739. doi: 10.1128/EC.05223-11
Bayram, Ö, and Braus, G. H. (2011). Coordination of secondary metabolism and development in fungi: the velvet family of regulatory proteins. FEMS 36, 1–24. doi: 10.1111/j.1574-6976.2011.00285.x
Bayram, Ö, Krappmann, S., Seiler, S., Vogt, N., and Braus, G. H. (2008). Neurospora crassa ve-1 affects asexual conidiation. Fungal. Genet. Biol. 45, 127–138. doi: 10.1016/j.fgb.2007.06.001
Bazafkan, H., Beier, S., Stappler, E., Böhmdorfer, S., Oberlerchner, J. T., Sulyok, M., et al. (2017). SUB1 has photoreceptor dependent and independent functions in sexual development and secondary metabolism in Trichoderma reesei. Mol. Biol. 106, 742–759. doi: 10.1111/mmi.13842
Bennett, L. D., Beremand, P., Thomas, T. L., and Bell-Pedersen, D. (2013). Circadian activation of the mitogen-activated protein kinase MAK-1 facilitates rhythms in clock-controlled genes in Neurospora crassa. Eukaryot. Cell 12, 59–69. doi: 10.1128/EC.00207-12
Bock, A., Kostenis, E., Tränkle, C., Lohse, M. J., and Mohr, K. (2014). Pilot the pulse: controlling the multiplicity of receptor dynamics. Trends Pharmacol. Sci. 35, 630–638. doi: 10.1016/j.tips.2014.10.002
Bojar, D., Martinez, J., Santiago, J., Rybin, V., Bayliss, R., and Hothorn, M. (2014). Crystal structures of the phosphorylated BRI1 kinase domain and implications for brassinosteroid signal initiation. Plant J. 78, 31–43. doi: 10.1111/tpj.12445
Borkovich, K. A., Alex, L. A., Yarden, O., Freitag, M., Turner, G. E., Read, N. D., et al. (2004). Lessons from the genome sequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 68, 1–108. doi: 10.1128/MMBR.68.1.1
Breitkreutz, A., Choi, H., Sharom, J. R., Boucher, L., Neduva, V., Larsen, B., et al. (2010). A global protein kinase and phosphatase interaction network in yeast. Science 328, 1043–1047.
Browne, F., Zheng, H., Wang, H., and Azuaje, F. (2010). From experimental approaches to computational techniques: a review on the prediction of protein-protein interactions. Adv. Artif. Intell. 2010, 15. doi: 10.1155/2010/924529
Cabrera, I. E., Pacentine, I. V., Lim, A., Guerrero, N., Krystofova, S., Li, L., et al. (2015). Global analysis of predicted G Protein 2 coupled receptor genes in the filamentous fungus, Neurospora crassa. Mol. Gen. Genet. 248, 2729–2743. doi: 10.1534/g3.115.020974
Caccia, D., Dugo, M., Callari, M., and Bongarzone, I. (2013). Bioinformatics tools for secretome analysis. Biochim. Biophys. Acta 1834, 2442–2453. doi: 10.1016/j.bbapap.2013.01.039
Carrillo, A. J., Schacht, P., Cabrera, I. E., Blahut, J., Prudhomme, L., Dietrich, S., et al. (2017). Functional profiling of transcription factor genes in Neurospora crassa. G3 7, 2945–2956. doi: 10.1534/g3.117.043331
Catlett, N. L., Yoder, O. C., and Turgeon, B. G. (2003). Whole-genome analysis of two-component signal transduction genes in fungal pathogens. Eukaryot. Cell 2, 1151–1161. doi: 10.1128/EC.2.6.1151
Chakrabarti, S., and Gintzler, A. R. (2003). Phosphorylation of G is augmented by chronic morphine and enhances G stimulation of adenylyl cyclase activity. Mol. Brain Res. 119, 144–151. doi: 10.1016/j.molbrainres.2003.09.002
Chakravorty, D., and Assmann, S. M. (2018). G protein subunit phosphorylation as a regulatory mechanism in heterotrimeric G protein signaling in mammals, yeast, and plants. Biochem. J. 475, 3331–3357. doi: 10.1042/BCJ20160819
Chen, C., Ringelberg, C. S., Gross, R. H., Dunlap, J. C., and Loros, J. J. (2009). Genome-wide analysis of light-inducible responses reveals hierarchical light signalling in Neurospora. EMBO J. 28, 1029–1042. doi: 10.1038/emboj.2009.54
Chen, J. (2015). Phosphorylation of RACK1 in plants. Plant Signal. Behav. 10:e1022013. doi: 10.1080/15592324.2015.1022013
Chini, B., Parenti, M., Poyner, D. R., and Wheatley, M. (2013). G-Protein-coupled receptors: from structural insights to functional mechanisms. Biochem. Soc. Trans. 41, 135–136. doi: 10.1042/BST20120344
Cohen, P. (2000). The regulation of protein function by multisite phosphorylation – a 25 year update. Trends Biochem. Sci. 25, 596–601. doi: 10.1016/S0968-0004(00)01712-6
Colabardini, A. C., Humanes, A. C., Gouvea, P. F., Savoldi, M., Goldman, M. H. S., von Zeska Kress, M. R., et al. (2012). Molecular characterization of the Aspergillus nidulans fbxA encoding an F-box protein involved in xylanase induction. Fungal Genet. Biol. 49, 130–140. doi: 10.1016/j.fgb.2011.11.004
Colot, H. V., Park, G., Turner, G. E., Ringelberg, C., Crew, C. M., Litvinkova, L., et al. (2006). A high-throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. PNAS 103, 10352–10357. doi: 10.1073/pnas.0601456103
Coradetti, S. T., Craig, J. P., Xiong, Y., Shock, T., Tian, C., and Glass, N. L. (2012). Conserved and essential transcription factors for cellulase gene expression in ascomycete fungi. Proc. Natl. Acad. Sci. U.S.A. 109, 7397–7402. doi: 10.1073/pnas.1200785109
Coradetti, S. T., Xiong, Y., and Glass, N. L. (2013). Analysis of a conserved cellulase transcriptional regulator reveals inducer-independent production of cellulolytic enzymes in Neurospora crassa. Microbiol. Open 2, 595–609. doi: 10.1002/mbo3.94
Craig, J. P., Coradetti, S. T., Starr, T. L., and Glass, N. L. (2015). Direct target network of the Neurospora crassa plant cell wall. mBio 6:e01452-15. doi: 10.1128/mBio.01452-15.Editor
Cziferszky, A., Mach, R. L., and Kubicek, C. P. (2002). Phosphorylation positively regulates DNA binding of the carbon catabolite repressor Cre1 of Hypocrea jecorina (Trichoderma reesei). J. Biol. Chem. 277, 14688–14694. doi: 10.1074/jbc.M200744200
Dautel, R., Wu, X. N., Heunemann, M., Schulze, W. X., and Harter, K. (2016). The sensor histidine kinases AHK2 and AHK3 proceed into multiple serine / threonine / tyrosine phosphorylation pathways in Arabidopsis thaliana. Mol. Plant 9, 182–186. doi: 10.1016/j.molp.2015.10.002
Denton, J. A., and Kelly, J. M. (2011). Disruption of Trichoderma reesei cre2, encoding an ubiquitin C-terminal hydrolase, results in increased cellulase activity. BMC Biotechnol. 11:103. doi: 10.1186/1472-6750-11-103
Diernfellner, A. C. R., and Schafmeier, T. (2011). Phosphorylations: making the Neurospora crassa circadian clock tick. FEBS J. 585, 1461–1466. doi: 10.1016/j.febslet.2011.03.049
Dong, W., Yinbo, Q., and Peiji, G. (1995). Regulation of cellulase synthesis in mycelial fungi: participation of ATP and cyclic AMP. Biotechnol. Lett. 17, 593–598. doi: 10.1007/BF00129384
Farkas, V., Gresik, M., Kolarova, N., Sulova, Z., and Sestak, S. (1990). “Biochemical and physiological changes during photoinduced conidiation and derepression of cellulase synthesis in Trichoderma,” in Trichoderma Reesei Cellulases: Biochemistry, Genetics, Physiology and Application, eds C. P. Kubicek, and Royal Society of Chemistry, (Cambridge: Royal Society of Chemistry), 139–155.
Fernandez, J., Wright, J. D., Hartline, D., Quispe, C. F., Madayiputhiya, N., and Wilson, R. A. (2012). Principles of carbon catabolite repression in the rice blast fungus: Tps1, Nmr1-3, and a MATE – family pump regulate glucose metabolism during infection. PLoS Genet. 8:e1002673. doi: 10.1371/journal.pgen.1002673
Ficarro, S. B., McCleland, M. L., Stukenberg, P. T., Burke, D. J., Ross, M. M., Shabanowitz, J., et al. (2002). Phosphoproteome analysis by mass spectrometry and its application to Saccharomyces cerevisiae. Nat. Biotechnol. 20, 301–305. doi: 10.1038/nbt0302-301
Fischer, M. S., Wu, V. W., Lee, J. E., O’Malley, R. C., and Glass, N. L. (2018). Regulation of cell-to-cell communication and cell wall integrity by a network of map kinase pathways and transcription factors in Neurospora crassa. Genetics 209, 489–506. doi: 10.1534/genetics.118.300904
Froehlich, A. C., Noh, B., Vierstra, R. D., Loros, J., and Dunlap, J. C. (2005). Genetic and molecular analysis of phytochromes from the filamentous fungus Neurospora crassa. Eukaryot. Cell 4, 2140–2152. doi: 10.1128/EC.4.12.2140
Galagan, J., Calvo, S., Borkovich, K., Selker, E., Read, N., Jaffe, D., et al. (2003). The genome sequence of the filamentous fungus Neurospora crassa. Nature 422, 859–868. doi: 10.1038/nature01554
Gerber, S. A., Rush, J., Gygi, S. P., Beausoleil, S. A., and Ville, J. (2006). A probability-based approach for high-throughput protein phosphorylation analysis and site localization. Nat. Biotech. 24, 1285–1292. doi: 10.1038/nbt1240
Glass, N. L., Schmoll, M., Cate, J. H. D., and Coradetti, S. (2013). Plant cell wall deconstruction by ascomycete fungi. Ann. Rev. Microbiol. 67, 477–498. doi: 10.1146/annurev-micro-092611-150044
Gras, D. E., Persinoti, G. F., Peres, N. T. A., Martinez-Rossi, N. M., Tahira, A. C., Reis, E. M., et al. (2013). Transcriptional profiling of Neurospora crassa Δmak-2 reveals that mitogen-activated protein kinase MAK-2 participates in the phosphate signaling pathway. Fungal Genet. Biol. 60, 140–149. doi: 10.1016/j.fgb.2013.05.007
Gremel, G., Dorrer, M., and Schmoll, M. (2008). Sulphur metabolism and cellulase gene expression are connected processes in the filamentous fungus Hypocrea jecorina (anamorph Trichoderma reesei). BMC Microbiol. 8:174. doi: 10.1186/1471-2180-8-174
He, Q., Shu, H., Cheng, P., Chen, S., Wang, L., and Liu, Y. (2005). Light-independent phosphorylation of WHITE COLLAR-1 regulates its function in the Neurospora circadian negative feedback loop. J. Biol. Chem. 280, 17526–17532. doi: 10.1074/jbc.M414010200
Hornbeck, P. V., Chabra, I., Kornhauser, J. M., Skrzypek, E., and Zhang, B. (2004). PhosphoSite: a bioinformatics resource dedicated to physiological protein phosphorylation. Proteomics 4, 1551–1561. doi: 10.1002/pmic.200300772
Hu, L., Huang, T., Liu, X., and Cai, Y. (2011). Predicting protein phenotypes based on protein-protein interaction network. PLoS One 6:e17668. doi: 10.1371/journal.pone.0017668
Huberman, L. B., Coradetti, S. T., and Glass, N. L. (2017). Network of nutrient-sensing pathways and a conserved kinase cascade integrate osmolarity and carbon sensing in Neurospora crassa. PNAS 114, E8665–E8674. doi: 10.1073/pnas.1707713114
Hurley, J. M., Loros, J. J., and Dunlap, J. C. (2016). The circadian system as an organizer of metabolism. Fungal Genet. Biol. 90, 39–43. doi: 10.1016/j.fgb.2015.10.002
Hurley, J. M., Jankowski, M. S., Santos, H., Crowell, A. M., Fordyce, S. B., Zucker, J. D., et al. (2018). Circadian proteomic analysis uncovers mechanisms of post-transcriptional regulation in metabolic pathways. Cell Syst. 7, 613.e5–626.e5. doi: 10.1016/j.cels.2018.10.014
Ivey, F. D., and Hoffman, C. S. (2005). Direct activation of fission yeast adenylate cyclase by the Gpa2 G alpha of the glucose signaling pathway. PNAS 102, 6180–6113.
Jöhnk, B., Bayram, Ö, Abelmann, A., Heinekamp, T., Mattern, D. J., Brakhage, A. A., et al. (2016). SCF ubiquitin ligase F-box protein Fbx15 controls nuclear co-repressor localization, stress response and virulence of the human pathogen Aspergillus fumigatus. PLoS Pathog. 12:e1005899. doi: 10.1371/journal.ppat.1005899
Jones, C. A., Greer-Phillips, S. E., and Borkovich, K. A. (2007). The response regulator RRG-1 functions upstream of a mitogen-activated protein kinase pathway impacting asexual development, female fertility, osmotic stress, and fungicide resistance in Neurospora crassa. Mol. Biol. Cell 18, 2123–2136. doi: 10.1091/mbc.E06
Jonkers, W., and Rep, M. (2009). Lessons from fungal F-Box proteins. Eukaryot. Cell 8, 677–695. doi: 10.1128/EC.00386-08
Kays, A. M., Rowley, P. S., Baasiri, R. A., and Borkovich, K. A. (2000). Regulation of conidiation and adenylyl cyclase levels by the G alpha Protein GNA-3 in Neurospora crassa. Mol. Cell. Biol. 20, 7693–7705. doi: 10.1128/mcb.20.20.7693-7705.2000
Kim, S., Gupta, N., and Pevzner, P. A. (2008). Spectral probabilities and generating functions of tandem mass spectra: a strike against decoy databases. J. Proteome Res. 7, 3354–3363. doi: 10.1021/pr8001244
Kim, S., and Pevzner, P. A. (2014). MS-GF+ makes progress towards a universal database search tool for proteomics. Nat. Commun. 5:5277. doi: 10.1038/ncomms6277
Kobe, B., and Deisenhofer, J. (1995). A structural basis of the interactions between leucine-rich repeats and protein ligands. Nat. Lett. 374, 183–186. doi: 10.1038/374183a0
Kobe, B., and Kajava, A. V. (2001). The leucine-rich repeat as a protein recognition motif. Curr. Opin. Struc. Biol. 11, 725–732. doi: 10.1016/s0959-440x(01)00266-4
Kore-Eda, S., Murayama, T., and Uno, I. (1991). Isolation and Characterization of the adenylate cyclase structural gene of Neurospora crassa. Jpn. J. Gnet. 66, 317–334. doi: 10.1266/jjg.66.317
Kovács, I. A., Luck, K., Spirohn, K., Wang, Y., Pollis, C., Schlabach, S., et al. (2019). Network-based prediction of protein interactions. Nat. Commun. 10, 1–8. doi: 10.1038/s41467-019-09177-y
Krappmann, S., Jung, N., Medic, B., Busch, S., Prade, R. A., and Braus, G. H. (2006). The Aspergillus nidulans F-box protein GrrA links SCF activity to meiosis. Mol. Microbiol. 61, 76–88. doi: 10.1111/j.1365-2958.2006.05215.x
Kress, M. R. Z., Harting, R., Bayram, Ö, Christmann, M., Irmer, H., Valerius, O., et al. (2012). The COP9 signalosome counteracts the accumulation of cullin SCF ubiquitin E3 RING ligases during fungal development. Mol. Microbiol. 83, 1162–1177. doi: 10.1111/j.1365-2958.2012.07999.x
Kunz, J., Deuter-Reinhard, M., Hall, M. N., Schneider, U., Movva, N. R., and Henriquez, R. (1993). Target of rapamycin in yeast, TOR2, is an essential phosphatidylinositol kinase homolog required for G1 progression. Cell 73, 585–596. doi: 10.1016/0092-8674(93)90144-f
Lamb, T. M., Finch, K. E., and Bell-Pedersen, D. (2012). The Neurospora crassa OS MAPK pathway-activated transcription factor ASL-1 contributes to circadian rhythms in pathway responsive clock-controlled genes. Fungal Genet. Biol. 49, 180–188. doi: 10.1016/j.fgb.2011.12.006
Larrondo, L. F., Olivares-Yañez, C., Baker, C. L., Loros, J. J., and Dunlap, J. C. (2015). Decoupling circadian clock protein turnover from circadian period determination. Science 347:1257277. doi: 10.1126/science.1257277
Lee, I. H., Walline, R. G., and Plamann, M. (1998). Apolar growth of Neurospora crassa leads to increased secretion of extracellular proteins. Mol. Microbiol. 29, 209–218. doi: 10.1046/j.1365-2958.1998.00923.x
Lengeler, K. B., Davidson, R. C., D’souza, C., Harashima, T., Shen, W.-C., Wang, P., et al. (2000). Signal transduction cascades regulating fungal development and virulence. Microbiol. Mol. Biol. Rev. 64, 746–785. doi: 10.1128/MMBR.64.4.746-785.2000
Lenoir, M., Ustunel, C., Rajesh, S., Kaur, J., Moreau, D., Gruenberg, J., et al. (2018). Phosphorylation of conserved phosphoinositide binding pocket regulates sorting nexin membrane targeting. Nat. Commun. 9:993. doi: 10.1038/s41467-018-03370-1
Li, L., and Borkovich, K. A. (2006). GPR-4 is a predicted G-protein-coupled receptor required for carbon source-dependent asexual growth and development in Neurospora crassa. Eukaryot. cell 5, 1287–1300. doi: 10.1128/ec.00109-06
Liu, P., Du, L., Huang, Y., Gao, S., and Yu, M. (2017). Origin and diversification of leucine-rich repeat receptor-like protein kinase (LRR-RLK) genes in plants. BMC Evol. Biol. 17:47. doi: 10.1186/s12862-017-0891-5
Lorenz, M. C., Pan, X., Harashima, T., Cardenas, M. E., Xue, Y., Hirsch, J. P., et al. (2000). The G protein-coupled receptor Gpr1 Is a nutrient sensor that regulates pseudohyphal differentiation in Saccharomyces cerevisiae. Genetics 154, 609–622.
Lv, X., Zhang, W., Chen, G., and Liu, W. (2015). Trichoderma reesei Sch9 and Yak1 regulate vegetative growth, conidiation, and stress response and induced cellulase production. J. Microbiol. 53, 236–242. doi: 10.1007/s12275-015-4639-x
McCluskey, K., Wiest, A., and Plamann, M. (2010). The fungal genetics stock center: a repository for 50 years of fungal genetics research. J. Biosci. 35, 119–126. doi: 10.1007/s12038-010-0014-6
Mertins, P., Tang, L. C., Krug, K., Clark, D. J., Gritsenko, M. A., Chen, L., et al. (2018). Reproducible workflow for multiplexed deep-scale proteome and phosphoproteome analysis of tumor tissues by liquid chromatography – mass spectrometry. Nat. Protoc. 13, 1632–1661. doi: 10.1038/s41596-018-0006-9
Meyer, V., Andersen, M. R., Brakhage, A. A., Braus, G. H., Caddick, M. X., Cairns, T. C., et al. (2016). Current challenges of research on filamentous fungi in relation to human welfare and a sustainable bio-economy: a white paper. Fungal Biol. Biotechnol. 3:6. doi: 10.1186/s40694-016-0024-8
Miranda-Saavedra, D., and Barton, G. J. (2007). Classification and functional annotation of eukaryotic protein kinases. Proteins 68, 893–914. doi: 10.1002/prot.21444
Montenegro-Montero, A., Canessa, P., and Larrondo, L. F. (2015). “Around the fungal clock: recent advances in the molecular study of circadian clocks in neurospora and other fungi,” in Advances in Genetics, Vol. 92, eds T. Friedmann, J. C. Dunlap, and S. F. Goodwin, (Cambridge, MA: Academic Press.), 107–184. doi: 10.1016/bs.adgen.2015.09.003
Moreira, I. S. (2014). Structural features of the G-protein/GPCR interactions. Biochim. Biophys. Acta 1840, 16–33. doi: 10.1016/j.bbagen.2013.08.027
Muller, F., Kruger, D., Sattlegger, E., Hoffmann, B., Ballario, P., Kanaan, M., et al. (1995). The cpc-2 gene of Neurospora crassa encodes a protein entirely composed of WD-repeat segments that is involved in general amino acid control and female fertility. Mol. Gen. Genet. 248, 162–173. doi: 10.1007/bf02190797
Nakagami, H., Sugiyama, N., Mochida, K., Daudi, A., Yoshida, Y., Toyoda, T., et al. (2010). Large-scale comparative phosphoproteomics identifies conserved phosphorylation sites in plants. Plant Physiol. 153, 1161–1174. doi: 10.1104/pp.110.157347
Nauwelaers, D., Thevelein, J. M., Versele, M., Peeters, T., Louwet, W., and Gelade, R. (2006). Kelch-repeat proteins interacting with the G alpha protein Gpa2 bypass adenylate cyclase for direct regulation of protein kinase A in yeast. PNAS 103, 13034–13039. doi: 10.1073/pnas.0509644103
Nguyen, E. V., Imanishi, S. Y., Haapaniemi, P., Yadav, A., Saloheimo, M., Corthals, G. L., et al. (2016). Quantitative site-specific phosphoproteomics of Trichoderma reesei signaling pathways upon induction of hydrolytic enzyme production. J. Proteome Res. 15, 457–467. doi: 10.1021/acs.jproteome.5b00796
Ogawa, M., Kobayashi, T., and Koyama, Y. (2012). ManR, a novel Zn(II)2Cys6 transcriptional activator, controls the β-mannan utilization system in Aspergillus oryzae. Fungal Genet. Biol. 49, 987–995. doi: 10.1016/j.fgb.2012.09.006
Park, G., Servin, J. A., Turner, G. E., Altamirano, L., Colot, H. V., Collopy, P., et al. (2011). Global analysis of serine-threonine protein kinase genes in Neurospora crassa. Eukaryot. Cell 10, 1553–1564. doi: 10.1128/EC.05140-11
Pawson, T. (2007). Dynamic control of signaling by modular adaptor proteins. Curr. Opin. Cell Biol. 19, 112–116. doi: 10.1016/j.ceb.2007.02.013
Perez-Riverol, Y., Csordas, A., Bai, J., Bernal-Llinares, M., Hewapathirana, S., Kundu, D. J., et al. (2019). The PRIDE database and related tools and resources in 2019: improving support for quantification data. Nucleic Acids Res. 47, D442–D450. doi: 10.1093/nar/gky1106
Petranovic, D., Bordel, S., Nielsen, J., Feizi, A., and Tobias, O. (2013). Genome-Scale Modeling of the Protein Secretory Machinery in Yeast. PLos One 8:e63284. doi: 10.1371/journal.pone.0063284
Rechsteiner, M., and Rogers, S. (1996). PEST sequences and regulation by proteolysis. Trends Biochem. Sc.i 21, 267–271. doi: 10.1016/0968-0004(96)10031-1
Ron, D., Adams, D. R., Baillie, G. S., Long, A., Connor, R. O., and Kiely, P. A. (2013). RACK (1) to the future – a historical perspective. Cell Commun. Sign. 11, 53. doi: 10.1186/1478-811X-11-53
Roosen, J., Engelen, K., Marchal, K., Mathys, J., Griffioen, G., Cameroni, E., et al. (2005). PKA and Sch9 control a molecular switch important for the proper adaptation to nutrient availability. Mol. Microbiol. 55, 862–880. doi: 10.1111/j.1365-2958.2004.04429.x
Ruepp, A., Zollner, A., Maier, D., Albermann, K., Hani, J., Mannhaupt, G., et al. (2004). The FunCat, a functional annotation scheme for systematic classification of proteins from whole genomes. Nucleic Acids Res. 32, 5539–5545. doi: 10.1093/nar/gkh894
Samal, A., Craig, J. P., Coradetti, S. T., Benz, J. P., Eddy, J. A., Price, N. D., et al. (2017). Network reconstruction and systems analysis of plant cell wall deconstruction by Neurospora crassa. Biotechnol. Biofuels 10, 1–21. doi: 10.1186/s13068-017-0901-2
Sancar, C., Ha, N., Yilmaz, R., Tesorero, R., and Fisher, T. (2015). Combinatorial control of light induced chromatin remodeling and gene activation in Neurospora. PLoS Genet. 11:e1005105. doi: 10.1371/journal.pgen.1005105
Sancar, G., Sancar, C., and Brunner, M. (2012). Metabolic compensation of the Neurospora clock by a glucose-dependent feedback of the circadian repressor CSP1 on the core oscillator. Genes Dev. 26, 2435–2442. doi: 10.1101/gad.199547.112
Sancar, G., Sancar, C., Brunner, M., and Schafmeier, T. (2009). Activity of the circadian transcription factor white collar complex is modulated by phosphorylation of SP-motifs. FEBS Lett. 583, 1833–1840. doi: 10.1016/j.febslet.2009.04.042
Schafmeier, T., and Diernfellner, A. C. R. (2011). Light input and processing in the circadian clock of Neurospora. FEBS Lett. 585, 1467–1473. doi: 10.1016/j.febslet.2011.03.050
Schaller, G. E., Shiu, S., and Armitage, J. P. (2011). Two-component systems and their co-option for eukaryotic signal transduction review. Curr. Biol. 21, R320–R330. doi: 10.1016/j.cub.2011.02.045
Schmoll, M. (2018). Regulation of plant cell wall degradation by light in Trichoderma. Fungal Biol. Biotechnol. 5, 1–20. doi: 10.1186/s40694-018-0052-7
Schmoll, M., Tian, C., Sun, J., Tisch, D., and Glass, N. L. (2012). Unravelling the molecular basis for light modulated cellulase gene expression - the role of photoreceptors in Neurospora crassa. BMC Genomics 13:127. doi: 10.1186/1471-2164-13-127
Schumacher, M. M., Enderlin, C. S., and Selitrennikoff, C. P. (1997). The osmotic-1 locus of Neurospora crassa encodes a putative histidine kinase similar to osmosensors of bacteria and yeast. Curr. Microbiol. 34, 340–347. doi: 10.1007/s002849900193
Schuster, A., Tisch, D., Seidl-Seiboth, V., Kubicek, C. P., and Schmoll, M. (2012). Roles of protein kinase A and adenylate cyclase in light-modulated cellulase regulation in Trichoderma reesei. Appl. Environ. Microbiol. 78, 2168–2178. doi: 10.1128/AEM.06959-11
Seibert, T., Thieme, N., and Benz, J. P. (2016). “The Renaissance of Neurospora crassa: How a Classical Model System is Used for Applied Research,” in Gene Expression Systems in Fungi: Advancements and Applications. Fungal Biology, eds M. Schmoll, and C. Dattenböck, (Cham: Springer).
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303.metabolite
Shoemaker, B. A., and Panchenko, A. R. (2007). Protein and domain interaction partners methods for predicting protein interaction partners. PLoS Comput. Biol. 3:e43. doi: 10.1371/journal.pcbi.0030043
Skowyra, D., Craig, K. L., Tyers, M., Elledge, S. J., and Harper, J. W. (1997). F-Box proteins are receptors that recruit phosphorylated substrates to the SCF ubiquitin-ligase complex. Cell Press 91, 209–219. doi: 10.1016/s0092-8674(00)80403-1
Stappler, E., Dattenböck, C., Tisch, D., and Schmoll, M. (2017). Analysis of light- and carbon-specific transcriptomes implicates a class of G- protein-coupled receptors in cellulose sensing. mSphere 2, e00089-17. doi: 10.1128/mSphere.00089-17
Sugiyama, N., Nakagami, H., Mochida, K., Daudi, A., Tomita, M., and Shirasu, K. (2008). Large-scale phosphorylation mapping reveals the extent of tyrosine phosphorylation in Arabidopsis. Mol. Syst. Biol. 4:193. doi: 10.1038/msb.2008.32
Sun, J., Tian, C., Diamond, S., and Glass, N. L. (2012). Deciphering transcriptional regulatory mechanisms associated with hemicellulose degradation in Neurospora crassa. Eukaryot. Cell 11, 482–493. doi: 10.1128/EC.05327-11
Szklarczyk, D., Morris, J. H., Cook, H., Kuhn, M., Wyder, S., Simonovic, M., et al. (2017). The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 45, D362–D368. doi: 10.1093/nar/gkw937
Tang, C.-T., Li, S., Long, C., Cha, J., Huang, G., Li, L., et al. (2009). Setting the pace of the Neurospora circadian clock by multiple independent FRQ phosphorylation events. PNAS 106, 10722–10727. doi: 10.1073/pnas.0904898106
Terenzi, H. F., Jorge, J. A., Roselino, J. E., and Migliorini, R. H. (1979). Adenylyl cyclase deficient cr-1 (Crisp) mutant of Neurospora crassa: cyclic AMP-dependent nutritional deficiencies. Arch. Microbiol. 123, 251–258. doi: 10.1007/bf00406658
Tesmer, J. J. G. (2010). The quest to understand heterotrimeric G protein signaling. Nat. Struct. Mol. Biol. 17, 650–652. doi: 10.1038/nsmb0610-650.The
Thieme, N., Wu, V. W., Dietschmann, A., Salamov, A. A., Wang, M., Johnson, J., et al. (2017). The transcription factor PDR-1 is a multi-functional regulator and key component of pectin deconstruction and catabolism in Neurospora crassa. Biotechnol. Biofuels 10, 1–21. doi: 10.1186/s13068-017-0807-z
Tian, C., Beeson, W. T., Iavarone, A. T., Sun, J., Marletta, M. A., Cate, J. H. D., et al. (2009). Systems analysis of plant cell wall degradation by the model filamentous fungus Neurospora crassa. PNAS 106, 22157–22162. doi: 10.1073/pnas.0906810106
Urano, D., Czarnecki, O., Wang, X., Jones, A. M., and Chen, J. G. (2015). Arabidopsis receptor of activated C Kinase1 phosphorylation by WITH NO LYSINE8 KINASE 1. Plant Physiol. 167, 507–516. doi: 10.1104/pp.114.247460
van Munster, J. M., Daly, P., Delmas, S., Pullan, S. T., Blythe, M. J., Malla, S., et al. (2014). The role of carbon starvation in the induction of enzymes that degrade plant-derived carbohydrates in Aspergillus niger. Fungal Genet. Biol. 72, 34–47. doi: 10.1016/j.fgb.2014.04.006
Vinuselvi, P., Kim, M. K., Lee, S. K., and Ghim, C. (2012). Rewiring carbon catabolite repression for microbial cell factory. BMB 45, 59–70. doi: 10.5483/BMBRep.2012.45.2.59
Vitalini, M. W., Paula, R. M., De Goldsmith, C. S., Jones, C. A., Borkovich, K. A., and Bell-pedersen, D. (2007). Circadian rhythmicity mediated by temporal regulation of the activity of p38 MAPK. PNAS 104, 18223–18228. doi: 10.1073/pnas.0704900104
Vogel, H. J. (1956). A convenient growth medium for Neurospora crassa. Microb. Genet. Bull. 13, 42–47.
Vries, R. P., De Jansen, J., Aguilar, G., Par, L., Joosten, V., Wu, F., et al. (2002). Expression profiling of pectinolytic genes from Aspergillus niger. FEBS Lett. 530, 41–47. doi: 10.1016/s0014-5793(02)03391-4
Wang, B., Kettenbach, A. N., Zhou, X., Loros, J. J., and Dunlap, J. C. (2019). The phospho-code determining circadian feedback loop closure and output in Neurospora. Mol. Cell 74, 771.e–784.e. doi: 10.1016/j.molcel.2019.03.003
Wang, B., Li, J., Gao, J., Cai, P., Han, X., and Tian, C. (2017). Identification and characterization of the glucose dual-affinity transport system in Neurospora crassa: pleiotropic roles in nutrient transport, signaling, and carbon catabolite repression. Biotechnol. Biofuels 10, 1–22. doi: 10.1186/s13068-017-0705-4
Wang, B., Zhou, X., Loros, J. J., and Dunlap, J. C. (2015). Alternative use of DNA binding domains by the neurospora white collar complex dictates circadian regulation and light responses. Mol. Cell. Biol. 36, 781–793. doi: 10.1128/mcb.00841-15
Wang, Z., An, N., Xu, W., Zhang, W., Meng, X., Chen, G., et al. (2018). Functional characterization of the upstream components of the Hog1-like kinase cascade in hyperosmotic and carbon sensing in Trichoderma reesei. Biotechnol. Biofuels 11, 1–17. doi: 10.1186/s13068-018-1098-8
Wart, V., and Unit, P. (1993). On target with a new mechanism for the regulation of protein phosphorylation. Trends Biochem. Sci. 18, 172–177. doi: 10.1016/0968-0004(93)90109-z
Weirauch, M. T., Yang, A., Albu, M., Cote, A., Montenegro-, A., Drewe, P., et al. (2014). Determination and inference of eukaryotic transcription factor sequence specificity. Cell 158, 1431–1443. doi: 10.1016/j.cell.2014.08.009.Determination
Welchman, R. L., Gordon, C., and Mayer, R. J. (2005). Ubiquitin and ubiquitin-like proteins as multifunctional signals. Nat. Rev. Mol. Cell Biol. 6, 599–609. doi: 10.1038/nrm1700
Whitmarsh, A. J., and Davis, R. J. (2000). Regulation of transcription factor function by phosphorylation. Cell. Mol. Life Sci. 57, 1172–1183. doi: 10.1007/pl00000757
Willardson, B. M., and Tracy, C. M. (2012). “Chaperone-Mediated Assembly of G Protein Complexes,” in GPCR Signalling Complexes – Synthesis, Assembly, Trafficking and Specificity, eds D. J. Dupré, T. E. Hébert, and R. Jockers, (Dordrecht: Springer), 131–153. doi: 10.1007/978-94-007-4765-4_8
Xiong, Y., Coradetti, S. T., Li, X., Gritsenko, M. A., Clauss, T., Petyuk, V., et al. (2014a). The proteome and phosphoproteome of Neurospora crassa in response to cellulose, sucrose and carbon starvation. Fungal Genet. Biol. 72, 21–33. doi: 10.1016/j.fgb.2014.05.005
Xiong, Y., Sun, J., and Glass, N. L. (2014b). VIB1, a link between glucose signaling and carbon catabolite repression, is essential for plant cell wall degradation by Neurospora crassa. PLoS Genet. 10:e1004500. doi: 10.1371/journal.pgen.1004500
Xiong, Y., Wu, V. W., Lubbe, A., Qin, L., Deng, S., Kennedy, M., et al. (2017). A fungal transcription factor essential for starch degradation affects integration of carbon and nitrogen metabolism. PLos Genet. 13:e1006737. doi: 10.1371/journal.pgen.1006737
Xue, Y., Batlle, M., and Hirsch, J. P. (1998). GPR1 encodes a putative G protein-coupled receptor that associates with the Gpa2p GAlfa subunit and functions in a Ras-independent pathway. EMBO J. 17, 1996–2007. doi: 10.1093/emboj/17.7.1996
Yang, Q., Poole, S. I., and Borkovich, K. A. (2002). A G-Protein beta subunit required for sexual and vegetative development and maintenance of normal G alpha protein levels in Neurospora crassa. Eukaryot. Cell 1, 378–390. doi: 10.1128/EC.1.3.378
Zhang, X., Jain, R., and Li, G. (2016). Roles of rack1 proteins in fungal pathogenesis. BioMed Res. Int. 2016, 1–8. doi: 10.1155/2016/4130376
Keywords: Neurospora crassa, signal transduction, substrate recognition, lignocellulose degradation, fungi, phosphorylation, proteomics
Citation: Horta MAC, Thieme N, Gao Y, Burnum-Johnson KE, Nicora CD, Gritsenko MA, Lipton MS, Mohanraj K, de Assis LJ, Lin L, Tian C, Braus GH, Borkovich KA, Schmoll M, Larrondo LF, Samal A, Goldman GH and Benz JP (2019) Broad Substrate-Specific Phosphorylation Events Are Associated With the Initial Stage of Plant Cell Wall Recognition in Neurospora crassa. Front. Microbiol. 10:2317. doi: 10.3389/fmicb.2019.02317
Received: 15 May 2019; Accepted: 23 September 2019;
Published: 01 November 2019.
Edited by:
Cristina Silva Pereira, New University of Lisbon, PortugalReviewed by:
Zheng Wang, Yale University, United StatesCopyright © 2019 Horta, Thieme, Gao, Burnum-Johnson, Nicora, Gritsenko, Lipton, Mohanraj, de Assis, Lin, Tian, Braus, Borkovich, Schmoll, Larrondo, Samal, Goldman and Benz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: J. Philipp Benz, YmVuekBoZm0udHVtLmRl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.