 Vijay K. Singh
Vijay K. Singh Avinash Mishra
Avinash Mishra Bhavanath Jha
Bhavanath Jha
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 07 June 2019
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.01269
This article is part of the Research Topic Beyond Antimicrobials: Non-Traditional Approaches to Combating Multidrug-Resistant Bacteria View all 24 articles
Bacterial cell-to-cell communication promotes biofilm formation and can potentially lead to multidrug resistance development. Quorum sensing inhibition (QSI) is an effective and widely employed strategy against biofilm formation. The extract from Exiguobacterium indicum SJ16, a gram-positive bacterium, isolated from the rhizosphere of Cyperus laevigatus showed significant anti-quorum sensing activity (about 99%) against the reference Chromobacterium violaceum CV026 strain without exerting any antibacterial effect. The potentially active QSI compound identified in the SJ16 extract was 3-Benzyl-hexahydro-pyrrolo[1, 2-a]pyrazine-1,4-dione. The SJ16 extract containing this active compound showed significant anti-quorum sensing activity against a model quorum sensing bacterium strain Pseudomonas aeruginosa PAO1 and a clinical isolate P. aeruginosa PAH by preventing biofilm formation without attenuating the cell growth within the biofilm. More specifically, the SJ16 extract changed the topography and architecture of the biofilm, thus preventing bacterial adherence and further development of the biofilm. Furthermore, it decreased virulence factors (rhamnolipid and pyocyanin), the bacterial motility, as well as the elastase, and protease activities in P. aeruginosa. Microarray analysis revealed the differential expression of quorum sensing regulatory genes. Based on these results, we herein propose a hypothetical model, characterizing the role of this QSI agent in the transcriptional regulation of quorum sensing in P. aeruginosa PAO1, demonstrating that this compound has significant drug-development potential. Further research is required to delineate its possible applications in therapeutics in the context of biofilm forming bacterial infections.
The diseases caused by pathogenic bacteria are controlled, prevented, and treated with a number of antibiotics which inhibit essential bacterial processes, such as cell wall synthesis, DNA replication, or protein biosynthesis. Antibiotics have long become the commonplace in our effort to tackle infectious diseases (Chu et al., 2014). However, extensive use of these agents creates an evolutionary pressure on bacteria which, in many cases, leads to antibiotics resistance. Therefore, alternative strategies with low potential for resistance emergence are required in order to combat pathogenic multidrug resistant bacteria (Chu et al., 2014; Singh et al., 2016b). Quorum sensing (QS) is directly involved in pathogenesis of infectious disease, by regulating the biofilm formation, the production of multiple virulence factors, as well as the motility of bacteria (Singh et al., 2017). In QS system, molecular cascades regulate the gene expression and determine the fate of bacterial biofilms (Ganin et al., 2015). In this context, gram-negative bacteria communicate through small diffusible molecules, such as acyl homoserine lactones, whereas gram-positive bacteria use autoinducer peptides for their communication (Galloway et al., 2011).
Biofilms are comprised of single or multiple microbial species and develop on different biotic and abiotic surfaces. In most of the cases, mixed species biofilms are predominant. However, single species biofilms are commonly associated with medical equipment-related infections, especially dental plaque and medical implants (O’Toole et al., 2000). Pseudomonas aeruginosa, a gram-negative bacterium, is considered to be the most important species of biofilm-forming bacteria. It can develop biofilms on a variety of biotic and abiotic surfaces; especially in immunocompromised patients (Driscoll et al., 2007). Moreover, this bacterium shows resistance to most of the conventional antibiotics, because it can form a biofilm matrix, which protects bacterial cells. Therefore, eradicating such infections poses a significant challenge (Driscoll et al., 2007; Lee and Zhang, 2015). QS regulatory networks control the virulence factors and biofilm formation in P. aeruginosa (Lee and Zhang, 2015). Therefore, utilization of anti-QS strategies could prove to be a promising way to tackle P. aeruginosa infections.
The rhizosphere is a region of soil adjacent to the plant roots that inhabits a number of microbes and facilitates various plant-microbe and microbe-microbe interactions. Many rhizospheric bacteria prevent the development of soilborne pathogens, while at the same time they protect the associated plants by activating the induced systemic resistance (Berendsen et al., 2012). Importantly, the rhizosphere supports different bacterial communities that exert QS and quorum quenching activities. Christiaen et al. (2011) isolated 59 bacterial species from 16 different environmental samples including plant rhizospheres and water, and found that 41 of them had anti-QS properties. Furthermore, Stenotrophomonas maltophilia and Delftia tsuruhatensis, isolated from the rhizosphere of Cyperus laevigatus showed anti-QS and anti-biofilm activities against P. aeruginosa (Singh et al., 2013, 2017). Moreover, a bacterium from the family of the Acinetobacter spp., isolated from the cucumber rhizosphere, inhibited the growth of Pseudomonas chlororaphis and Burkholderia glumae during co-cultivation by degrading acyl-homoserine lactones (AHLs), produced by these bacteria (Kang et al., 2004). Other AHL producing and degrading bacteria were isolated from the tobacco rhizosphere, including two newly identified species, Sphingopyxis witflariensis and Bosea thiooxidans, belong to the Bacillus α-proteobacteria family (D’Angelo-Picard et al., 2005). A three bacterial consortium comprised of Acinetobacter sp., Burkholderia sp. and Klebsiella sp. was isolated from the ginger rhizosphere, demonstrating significant AHL degrading activities and growth-inhibiting capabilities against P. aeruginosa and the plant pathogen Erwinia carotovora without affecting their planktonic growth (Chan et al., 2011). Last but not least, the AHL degrading bacterial consortia isolated from the potato rhizosphere showed significant biocontrol activity against Pectobacterium atrosepticum (Cirou et al., 2007). In most of these earlier studies, the above mentioned bacteria were isolated and characterized for their anti-QS properties; however, to the best of our knowledge there is no report, so far, on the isolation and characterization of the active compounds from these bacteria.
In the present study, a bacterium Exiguobacterium indicum SJ16 was isolated from the rhizosphere of a monocot C. laevigatus, amply growing in the coastal saline area and was tested for its anti-QS and anti-biofilm potential. The active fraction was collected, identified, and regulatory key genes were studied to elucidate a possible QSI mechanism.
Bacteria from the rhizosphere of C. laevigatus L. from the natural habitat of Bhavnagar, India (Latitude N 21°45.124”, Longitude E 72°13.579”) were isolated. Bacteria were screened by plate based bioassay, and two were found positive for anti-QS activity (Singh et al., 2017). The bacterial isolate SJ16, which showed anti-QS (but not antibacterial) activity was selected for further characterization. Genomic DNA of the bacterium was isolated, and the 16S rRNA gene was amplified by universal primers fD1-5′-AGA GTT TGA TCC TGG CTC AG -3′ and rP2-5′-ACG GCT ACC TTG TTA CGA CTT -3′ using optimized PCR conditions (Weisburg et al., 1991). The amplified PCR product was purified, sequenced (at Macrogen Inc., South Korea), analyzed and submitted to the NCBI GenBank. The phylogenetic analysis was performed using MEGA version 6.0 (Tamura et al., 2013), and a phylogenetic tree was inferred by neighbor-joining methods (Saitou and Nei, 1987). Bootstrap analysis was performed, and maximum composite likelihood algorithms were used for the determination of the evolutionary distances (Felsenstein, 1985; Tamura et al., 2004).
Fatty acid methyl ester (FAME) profiling of E. indicum SJ16 was carried out using Microbial Identification System coupled with gas chromatography (MIDI, Microbial ID; GC system-6850, Agilent Technologies, United States). The bacterium was grown for 24 h at 30°C on Tryptic soy yeast agar, FAMEs were prepared using MIDI manual, and peaks were identified with RTSBA6 6.10 database (Jha et al., 2015).
Bacterial culture (E. indicum SJ16) was grown for 48 h at 30°C in 500 ml of nutrient broth, kept in an incubator shaker with agitation at180 rpm. The culture was centrifuged for 15 min at 10,000 × g (4°C), the supernatant was collected, and filtered through 0.22 μm filter for the complete removal of remaining bacterial cells. The supernatant was extracted with ethyl acetate (equal volume), evaporated to dryness under vacuum and the dried residue finally dissolved in methanol.
The methanol extract of SJ16 was further fractionated by solid phase extraction method using different cartridges including anion exchanger DAE, cation mixed Plexa PCX, polar SI, and non-polar C18 (Agilent, United States). Each fraction was tested for quorum sensing inhibition (QSI) activity (Singh et al., 2017). The positive fraction (collected through the C18 cartridge with 40% methanol) showing a maximum zone of QSI was used for the subsequent studies and was also subjected to GC-MS (GC-2010, Shimadzu, Japan) analysis. The probable active compound was identified by comparing the mass spectra with the reference spectra library. The mass of the probable active compound was further confirmed by electrospray ionization mass spectrometry (ESI-MS; Q-Tof micro TM, Micromass, United Kingdom).
The anti-QS activity was tested (i) by a plate-based bioassay using Chromobacterium violaceum CV026 as a tester strain, methanol as a negative control and cinnamaldehyde (Sigma-Aldrich, United States) as a positive control, and (ii) by quantifying the violacein production. Both the plate-based bioassay and the violacein quantification, were performed as per our previously optimized methods (Singh et al., 2013).
The anti-biofilm formation assay was performed with different concentrations of bacterial (strain SJ16) extracts (0.2, 0.4, 0.6, 0.8, 1.0, and 1.2 mg/ml) using two P. aeruginosa strains, PAO1 as well as the PAH clinical isolate (provided as a courtesy from the Government Medical College, Bhavnagar; Goswami et al., 2011). Briefly, 200 μl bacterial culture (OD600 nm = 0.1) dilute from overnight grown PAO1 and PAH cultures were added to a 96-well polystyrene microtiter plate with different concentrations of the SJ16 extracts. The plates were allowed to grow for 24 h (at 37°C with 100 rpm), and growth was measured at 600 nm. The biofilm formation was assayed using our previously optimized crystal violet staining method (Kavita et al., 2014). The experiments were performed three times with five replicates each time.
Pseudomonas aeruginosa (strains PAO1 and PAH) were grown in a 24 well polystyrene plate with or without SJ16 extract to assess the development of biofilm on a sterilized glass coverslip (11 mm). In a 24-well polystyrene plate 1 ml of NB media containing bacterial culture at OD600 nm = 0.1 (diluted from overnight grown culture) was added to each well with or without (control) 1.0 mg/ml of SJ16 extract. One glass coverslip/well was submerged and the plate was incubated in static condition at 37°C. The effect of the SJ16 extract (1.0 mg/ml) on the P. aeruginosa cell viability within biofilm was examined at different time points (24, 48, and 72 h) under a fluorescence microscope using FilmTracer LIVE/DEAD Biofilm Viability Kit (Invitrogen, United States) (Singh et al., 2013). The bacterial cells within the biofilm were labeled with a fluorescent dye (propidium iodide and SYTO 9), were further processed according to the manufacturer’s instructions and were visualized under an epifluorescence microscope (Axio Imager, Carl Zeiss AG, Germany).
Scanning electron microscopy (SEM) and atomic force microscopy (AFM) were used to visualize the effect of the SJ16 extract on the topology of the biofilm developed by the P. aeruginosa strains. For the SEM, a previously described protocol was used (Andersson et al., 2009; Singh et al., 2013). In brief, biofilms of P. aeruginosa PAO1 and P. aeruginosa PAH were grown on glass coverslips submerged in nutrient broth in a 24-well polystyrene plate with (1.0 mg/ml) or without extract (control). The plate containing the culture and the coverslips were kept at 37°C for 24 h. After incubation, the planktonic culture was removed and coverslips were gently washed with 0.9% NaCl. The samples were treated with 2.5% glutaraldehyde for 20 min followed by 4% OsO4 for 30 min and dehydrated using ethanol gradient (10 to 95%) treatment for 10 min for each concentration. The dehydrated and dried biofilms were coated with gold and observed under a scanning electron microscope (SEM, LEO series VP1430, Germany). For the AFM, overnight grown cultures of P. aeruginosa PAO1 and P. aeruginosa PAH were diluted to reach an OD600 nm = 0.1 in NB broth, sterile glass cover slips were submerged in 24-well polystyrene plate with (1.0 mg/ml) or without extract (control). The plate was kept for 24 h at 37°C. Following this incubation period, the biofilm that developed on the glass coverslip was rinsed gently with phosphate buffer saline (pH 7.4) and was kept under the desiccators until completely dry. Finally, the biofilm was scanned under AFM (NT-MDT, Russia) in a semi-contact mode (Oh et al., 2009; Singh et al., 2013).
The swarming motility of PAO1 and PAH were tested in the presence (1.0 mg/ml) and absence of SJ16 extract. Overnight grown culture of P. aeruginosa PAO1 and P. aeruginosa PAH were diluted to OD600 nm = 1.0 and spotted on a plate containing BM2 swarming medium (62 mM PBS at pH 7, 2 mM MgSO4, 10 μM FeSO4, 0.4% glucose, 0.1% casamino acids and 0.5% agar) supplemented with 1.0 mg/ml SJ16 extract or without extract supplementation (control) (Overhage et al., 2007). The plates were incubated at 37°C for 24 h and swarming zones were observed.
The overnight grown culture of PAO1 and PAH were diluted to OD600 nm = 1.0 and spotted on a plate containing tryptone broth (10 g/l tryptone, 5 g/l NaCl and 0.3% agar) supplemented with 1.0 mg/ml SJ16 extract or without extract supplementation (control) (Rashid and Kornberg, 2000). The plates were incubated at 37°C and analyzed after 24 h.
The effect of the SJ16 bacterial extracts (1.0 mg/ml) on the production of virulence factors by reference P. aeruginosa strains (PAO1 and PAH) was studied by measuring the levels of pyocyanin and rhamnolipid, and by analyzing the elastase and protease activities. For pyocyanin assay, starter cultures of P. aeruginosa strains (PAO1 and PAH) were grown at 37 °C in an incubator shaker until OD600 nm = 3.0 and diluted to OD600 nm = 0.1 in PB medium (5 ml; 20 g/l peptone, 1.4 g/l MgCl2, and 10 g/l K2SO4). The tubes containing 5 ml PB medium (OD600 nm = 0.1) supplemented with 1.0 mg/ml SJ16 extract or without extract supplementation (control). Supernatants were collected by centrifuging cultures at 10,000 × g for 10 min; pyocyanin was extracted in 3 ml of chloroform followed by 1 ml of 0.2 N HCl and quantified spectrophotometrically at 520 nm (Essar et al., 1990). For rhamnolipid assay, P. aeruginosa strains PAO1 and PAH (OD600 nm = 0.1) were grown overnight at 37°C in NB medium, supplemented with 1.0 mg/ml SJ16 extract or without extract supplementation (control). The culture was centrifuged (10,000 × g for 10 min), the supernatant was collected and acidified to pH 2 (with HCl), and absorbance was measured at 570 nm (McClure and Schiller, 1992; Sarabhai et al., 2013).
To estimate the elastase activity, the supernatant (750 μl) from overnight grown (with or without SJ16 extract) cultures of P. aeruginosa (PAO1 and PAH) were incubated with elastin congo red solution (250 μl; 5 mg/ml in 0.1 M Tris–HCl pH 8;1 mM CaCl2) at 37°C for 16 h. Reaction-mixtures were centrifuged (3,000 × g for 10 min), the supernatant was collected, and the absorbance was read at 495 nm (Bjorn et al., 1979; Zhu et al., 2002).
For the protease activity, 2% azocasein solution was prepared in 50 mM phosphate buffer saline (pH 7). The supernatant (400 μl) from overnight grown (with or without SJ16 extract) cultures of P. aeruginosa (PAO1 and PAH) were incubated with an equal volume of azocasein solution (2%) at 37°C for 1 h. A measure of 500 μl of 10% trichloroacetic acid was added to stop the reaction. Reaction-mixtures were centrifuged (8,000 × g for 5 min) to remove residual azocasein and the absorbance of the supernatant was measured at 400 nm (Adonizio et al., 2008).
A single colony of the reference strain P. aeruginosa PAO1 was inoculated in a 5 ml NB tubes, was grown until OD600 nm 3.0 and was then diluted to OD600 nm = 0.1. Following dilution, P. aeruginosa PAO1 was grown in a tube containing 5 ml starter culture (OD600 nm = 0.1) with or without bacterial extracts (1.0 mg/ml) at 37°C for 24 h in a shaker incubator at 200 rpm (Singh et al., 2017). Planktonic cells were harvested by centrifuge at 12,000 × g for 5 min and total RNA was isolated using TRI reagent (Sigma, United States). Total RNA was analyzed on 2% agoras gel and quantified using ND-1000 spectrophotometer (Nanodrop Technologies, United States). Ten μg RNA, extracted from control and treated samples were converted to cDNA, fragmented and labeled using previously optimized method (Singh et al., 2016a). Labeled cDNAs were hybridized with genome array gene chip (Gene Chip P. aeruginosa PAO1 containing total 5,886 gene probes), washed, stained, and scanned (Scanner 3000 7G, Affymetrix, United States) (Singh et al., 2017). Scanned chips were processed and analyzed using the expression console and the transcriptome analysis console (Affymetrix, United States). Microarray analysis was performed in duplicate and the genes showing significant differences in fold-change expression (ANOVA p-value < 0.05) were considered for the study.
Statistical analysis was performed using GraphPad Prism software. One-way ANOVA followed by Tukey post hoc that was applied for the comparison of test samples and controls.
Previously, out of 56 bacterial axenic cultures which were obtained from the rhizosphere of C. laevigatus L., two axenic cultures (SJ01 and SJ16) showed anti-QS activity (Singh et al., 2017). The 16S rRNA gene sequence (accession no. KX130768) of isolate SJ16 showed 99% similarity with E. indicum with 100% query coverage; therefore, this isolate was designated as E. indicum SJ16. The phylogenetic analysis showed the taxonomic position of the strain SJ16 (Supplementary Figure S1). The whole-cell FAME profile of the E. indicum SJ16 bacterium demonstrated an abundance of the iso-C17:0 (17.39%), iso-C15:0 (14.79%) and anteiso-C13:0 (11.36%) fatty acids (Supplementary Figure S2).
All fractions, collected from the various SPE cartridges (anion exchanger DAE, cation mixed Plexa PCX, polar SI, and non-polar C18) were individually screened for anti-QS activity using a biosensor plate containing the tester strain C. violaceum CV026. The fraction collected from the C18 cartridge (with 40% methanol) showed a maximum zone of QSI. This fraction was subjected to GC-MS analysis, and the chromatogram showed a single predominant peak at the retention time 16.01 min (Figure 1). The detected peak resembled the 3-Benzyl-hexahydro-pyrrolo[1, 2-a]pyrazine-1,4-dione (C14H16N2O2; expected molecular mass 244.28) from the GC-MS library. The active fraction was also subjected to ESI-MS analysis, which showed a dominating spectral peak at m/z 249.29 (Figure 1). Thus, the experimental molecular mass of the active compound was determined to be 249.29, which is corresponding to the theoretical mass of the active fraction.
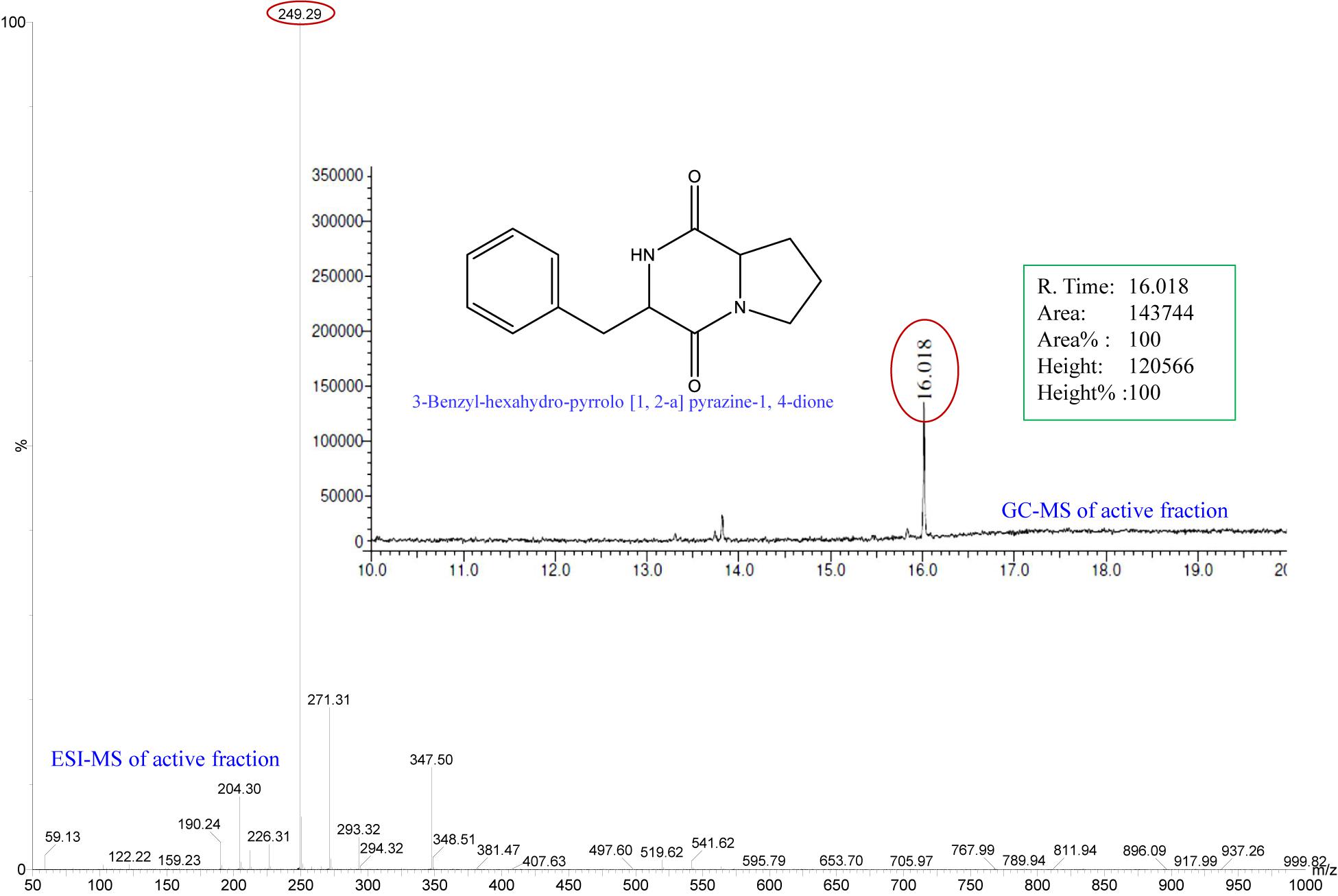
Figure 1. Analysis of active fraction showing QSI. GC chromatograms and ESI-MS/MS of the active fraction (C18-40) of E. indicum SJ16 extract showed resemblance to 3-Benzyl-hexahydro-pyrrolo[1,2-a]pyrazine-1,4-dione (redrawn by ChemBioDraw Ultra 12.0).
A clear white opaque zone was observed with the axenic culture and crude extract of the E. indicum strain SJ16 in the biosensor plate containing the reference strain C. violaceum CV026, result that was comparative to the positive control with cinnamaldehyde, a well-known QS inhibitor (Figure 2). The zone of inhibition was not detected with the extraction solvent methanol that was used as a negative control. As inhibition of violacein production is an indicator of QS activity, different concentrations (0.1–0.6 mg/ml) of the SJ16 extract were used to quantify the levels of this QSI marker. Indeed, the violacein production decreased concomitantly with increasing concentrations of the extract, and about 99% inhibition was achieved with 0.6 mg/ml of the SJ16 extract (Figure 2). Furthermore, the antibacterial assay, performed by using the disc diffusion technique, confirmed that the bacterial isolate SJ16 did not halt the growth or the viability of the reference strain C. violaceum CV026 (Supplementary Figure S3).
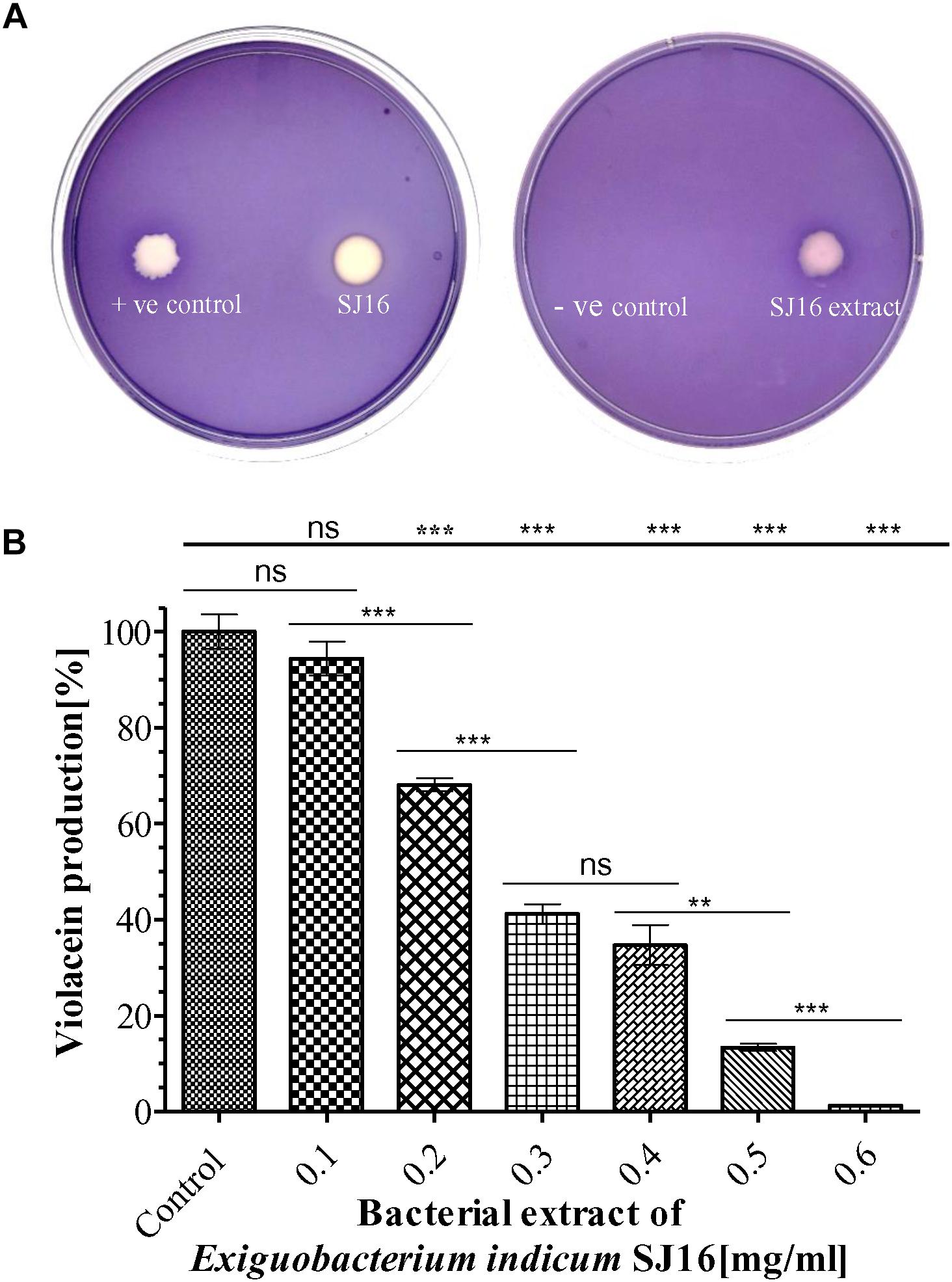
Figure 2. Anti-quorum sensing activity of SJ16. (A) The biosensor plates containing the reference strain C. violaceum CV026 were spotted with cinnamaldehyde (positive control), SJ16 (axenic culture), SJ16 extract, and methanol (negative control). A bright white opaque zone of inhibition showed anti-QS activity. (B) Different concentrations of bacterial extract (0.1–0.6 mg/ml) were used to quantify the inhibition of violacein production, an indicator of QS activity. Culture without extract was considered as a control. “∗∗” and “∗∗∗” Indicates significant differences from the control at P < 0.01 and P < 0.001, respectively.
The anti-biofilm activity of E. indicum SJ16 extract was performed with a different concentration (0.2–1.2 mg/ml) against PAO1 and PAH strains. The biofilm formation capability of both strains significantly decreased with increasing concentrations of the extract (Figure 3). About 50% inhibition of biofilm formation was noticed with 1.0 mg/ml extract. No significant effect of this compound was observed on the planktonic growth of either PAO1 or PAH even with highest extract concentrations used (0.2–1.2 mg/ml) (Figure 3).
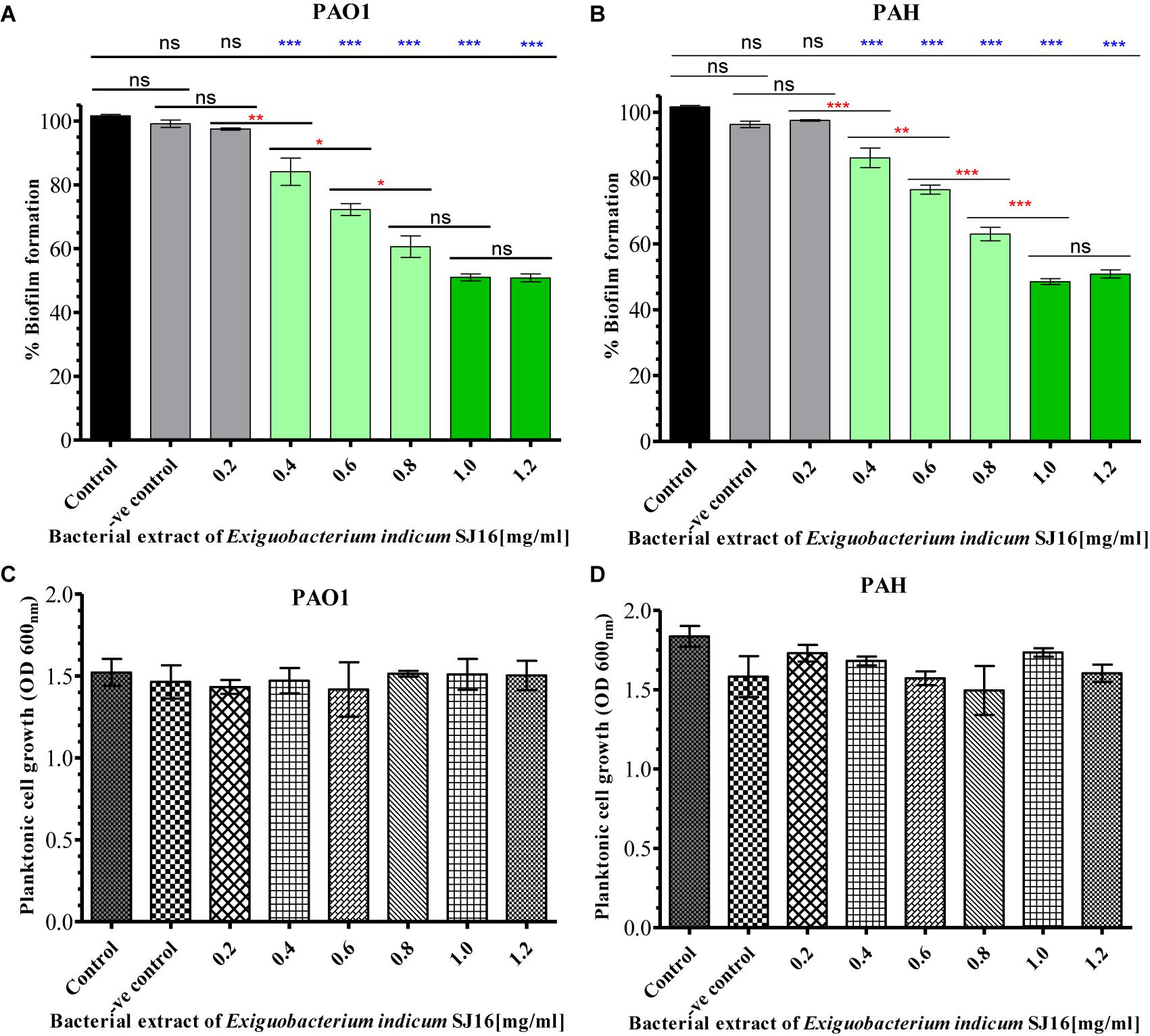
Figure 3. Effect of E. indicum SJ16 extract on biofilm formation and planktonic growth of P. aeruginosa. Different concentration of bacterial extracts (0.2–1.2 mg/ml) ware tested against the biofilm forming reference P. aeruginosa strains PAO1 (A) and PAH (B). The same concentrations of bacterial extracts were also tested for the inhibitory effect on the planktonic growth of PAO1 (C) and PAH (D) strains. Tests without extract and with methanol were considered as control and negative control, respectively. “∗”, “∗∗” and “∗∗∗” Indicates significant differences from the control at P < 0.05, P < 0.01, and P < 0.001, respectively.
The viability of the reference strains (P. aeruginosa PAO1 and PAH) within the biofilm was further analyzed with the SJ16 extract at 24, 48, and 72 h. The live cells of P. aeruginosa were labeled with SYTO 9, whereas dead cells were stained with propidium iodide using a Live/Dead staining kit. Live and dead cells produced green and red fluorescence, respectively, when visualized under an epi-fluorescence microscope (Figure 4A). P. aeruginosa (PAO1 and PAH) cells, treated with SJ16 extract were not tightly attached to the biofilm surface compared to the control. There was no difference between the control and treated biofilm when considering the cell viability whithin biofilm, and insignificant numbers of dead cells were detected in the biofilm at different time points (Figure 4A). Overall, these results indicate that the SJ16 extract does not exert any toxic effect on the growth or viability of the aforementioned reference strains, even after a longer duration, or higher concentrations of the compound, while at the same time it effectively inhibits the bacterial biofilm formation (Figure 4).
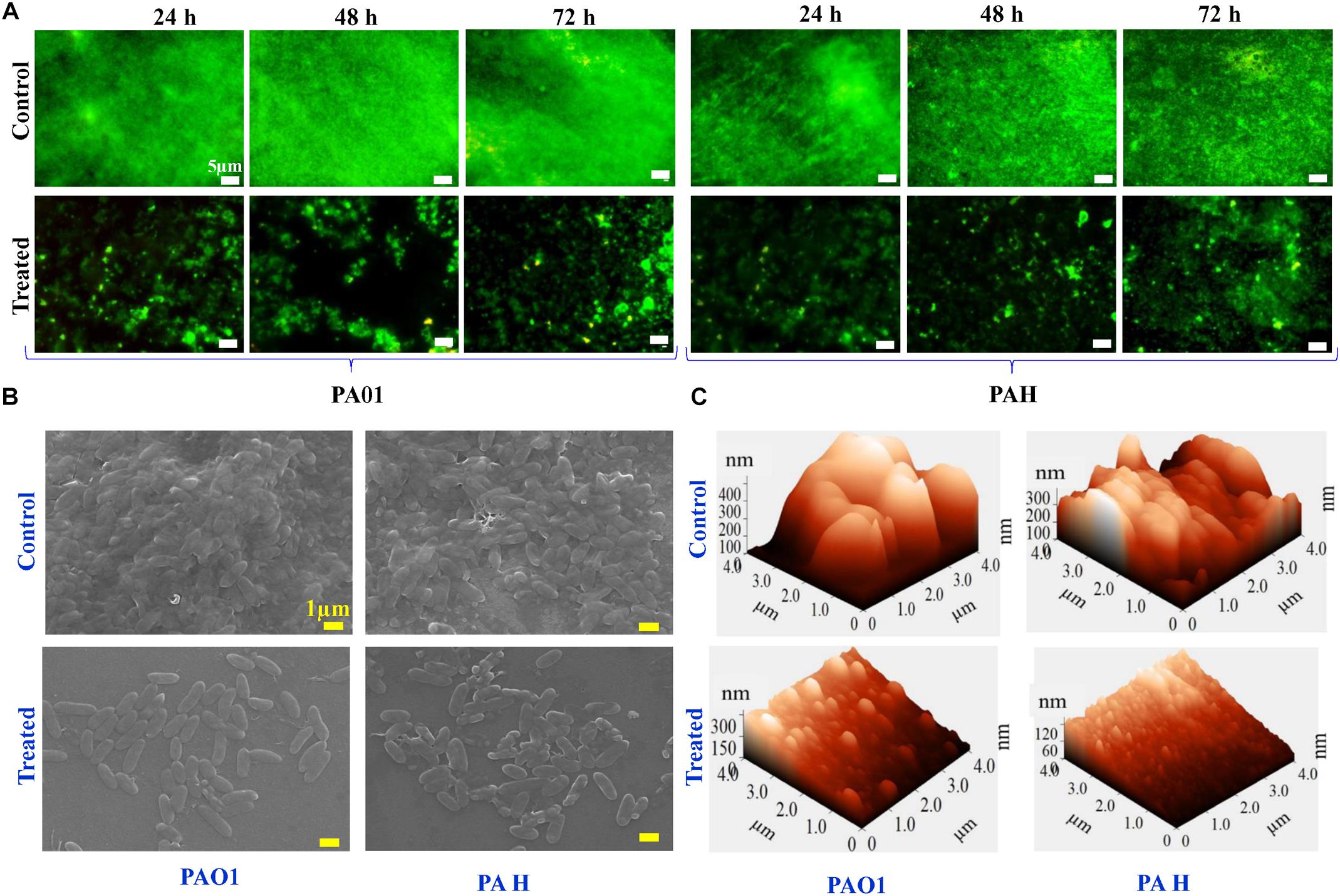
Figure 4. Effect of E. indicum SJ16 extract on cell viability, and topology of biofilms developed by P. aeruginosa. (A) The effect of the bacterial extract (1.0 mg/ml) on the viability of reference P. aeruginosa strains (PAO1 and PAH) in the biofilm was examined under an epifluorescence microscope at different time points (24, 48 and 72 h) and compared with control. The dead bacterial cells were labeled with propidium iodide whereas live cells were stained with SYTO 9, which produced red and green fluorescence, respectively. SEM (B) and AFM (C) images illustrate the effect of the bacterial extract (1.0 mg/ml) on biofilm formation and topology. (B) A well-grown biofilm along with adhering bacterial cells was observed in the control samples (normal biofilm developed by P. aeruginosa), whereas dispersed bacterial cells were observed in treated samples under SEM. Similarly, AFM (C) showed a disrupted surface topology and a height distribution profile of the biofilm developed by the reference P. aeruginosa strains in the presence of the bacterial extract compared to the control biofilm.
The effect of the SJ16 extract on the topology of the biofilm developed by the PAO1 and PAH was visualized under SEM and AFM. A biofilm along with adhering bacterial cells was developed by P. aeruginosa in the absence of SJ16 extract (control), whereas an immature biofilm with dispersed bacterial cells was observed when P. aeruginosa was grown with SJ16 extract (Figure 4B). Similarly, AFM micrographs showed changes in the topology of the biofilm developed by P. aeruginosa in the presence of SJ16 extract compared to normal (control) biofilm (Figure 4C). AFM showed poor biofilm adherence on the surface of the glass cover slip, while height distribution profile showed the average thickness of the biofilm developed in the presence of SJ16 extract was significantly reduced compared to the control biofilm (Figure 4C).
Bacterial motility and initial attachment of bacterial cells to surfaces are key prerequisites for biofilm formation. Therefore, the effect of the E. indicum SJ16 extract on the motility of reference P. aeruginosa strains (PAO1 and PAH) was studied. Our results show that the swarming and swimming motility of PAO1 and PAH are significantly inhibited in the presence of the SJ16 extract (Figure 5A).
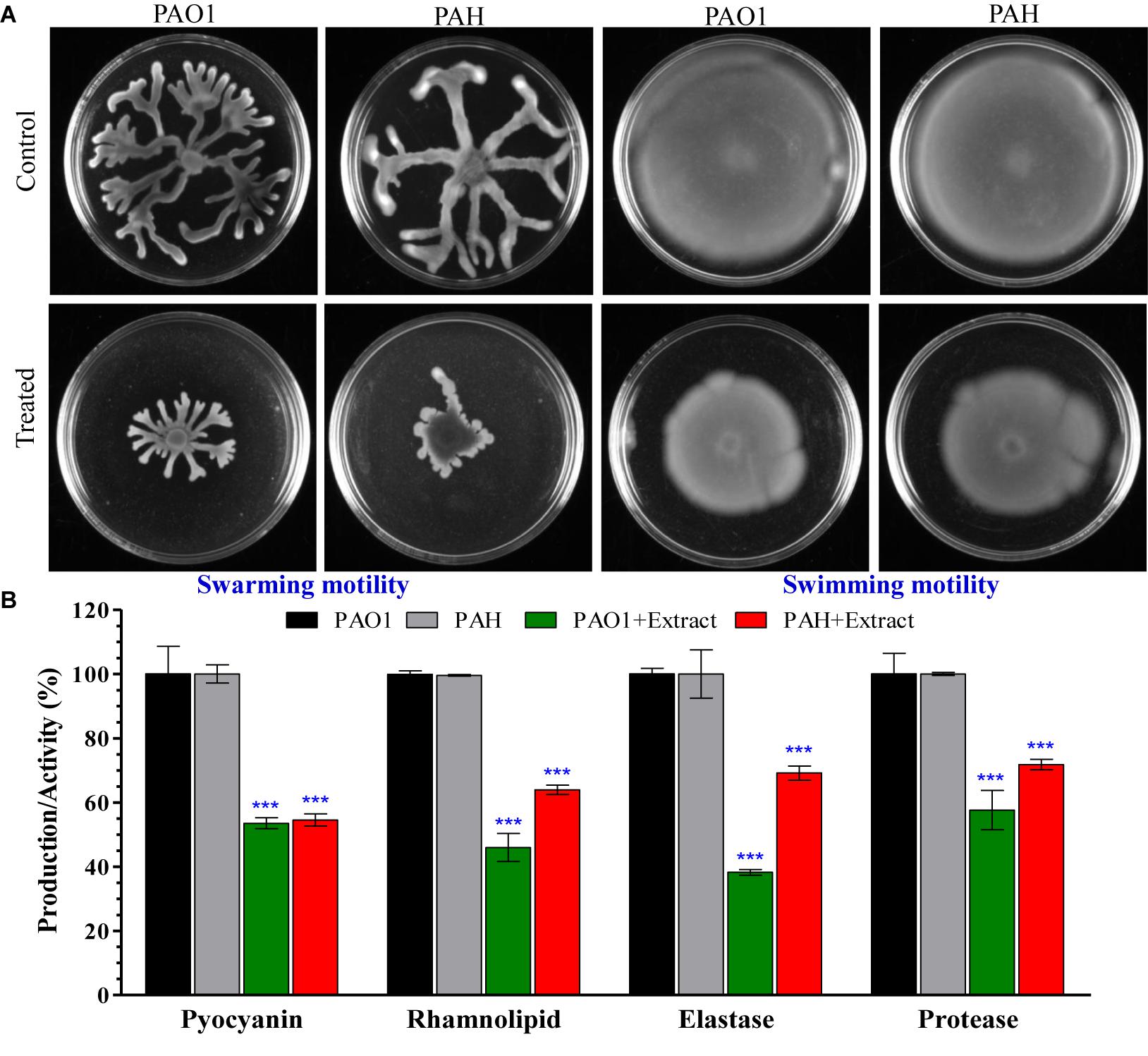
Figure 5. Effect of E. indicum SJ16 extract on cell motility, and the virulence factors of P. aeruginosa. (A) The effect of the bacterial extract on the swarming and swimming motility of reference P. aeruginosa strains (PAO1 and PAH). P. aeruginosa was spotted on a plate supplemented with 1.0 mg/ml extract or without extract supplementation (control). Plates were analyzed after incubation of 24 h at 37°C; (B) The effect of the bacterial extracts (SJ16; 1.0 mg/ml) was determined based on the production of virulence factors of the reference P. aeruginosa strains by quantifying pyocyanin and rhamnolipid, as well as by analyzing the elastase and protease activities. “∗∗∗” Indicates significant differences from the control at P < 0.001.
Virulence activities of the P. aeruginosa strains (PAO1 and PAH), including elastase and protease activities, as well as the production of virulence factors (pyocyanin and rhamnolipid) accelerate biofilm formation. Our data indicate that the SJ16 extract considerably reduced the production of virulence factors, with pyocyanin production decreasing about 50% in both PAO1 and PAH compared to the untreated samples (Figure 5B). Similarly, rhamnolipid production was also significantly decreased by 55% in PAO1 and 37% in PAH in the presence of the SJ16 extract compared to control (Figure 5B). Furthermore, the SJ16 extract led to a drastic inhibition of the elastase and protease activities in the above-mentioned P. aeruginosa strains. Specifically, elastase activity was decreased by 62% in PAO1 and by 31% PAH compared to untreated samples (Figure 5B). Similarly, protease activity was reduced by about 40% in PAO1 and by 28% in PAH in the presence of the SJ16 extract compared to control (Figure 5B). Overall, our results (Figures 3–5) strongly demonstrate that the SJ16 extract inhibits the cell-to-cell communication and thus prevents P. aeruginosa from developing a robust biofilm.
A P. aeruginosa PAO1 genome array chip, containing 5,886 gene-probe sets, was used to study the effect of 3-Benzyl-hexahydro-pyrrolo[1, 2-a]pyrazine-1,4-dione on QS regulatory genes. Scattered plot analysis showed that 1,237 out of 5,886 genes (Array-Express accession E-MTAB-6829) were differentially expressed (Figure 6), with a two-fold (p < 0.05) up- (>2) or down-(<-2) regulation at minimum (Supplementary Table S1). More specifically, a total of 868 genes were down-regulated, whereas 369 genes were up-regulated in the presence of the compound. Among these, we identified a plethora of genes that are key players in bacterial QS network of P. aeruginosa and exert crucial functions in general metabolic pathways in bacteria (Table 1).
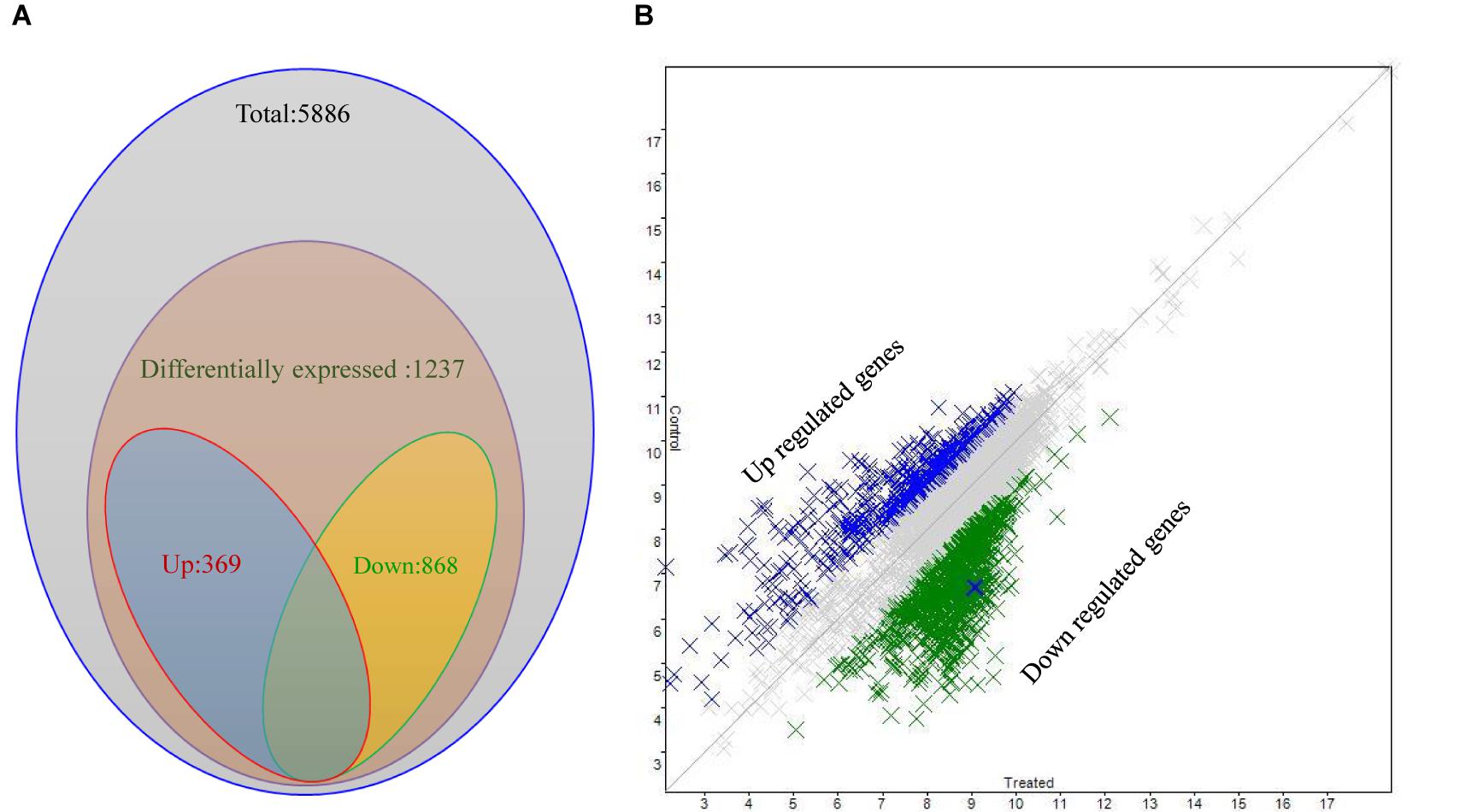
Figure 6. Differential expression analysis of P. aeruginosa PAO1 treated with SJ16 extract. (A) Venn diagram and (B) Scattered plot showing genes differentially expressed in P. aeruginosa PAO1 as determined by microarray.
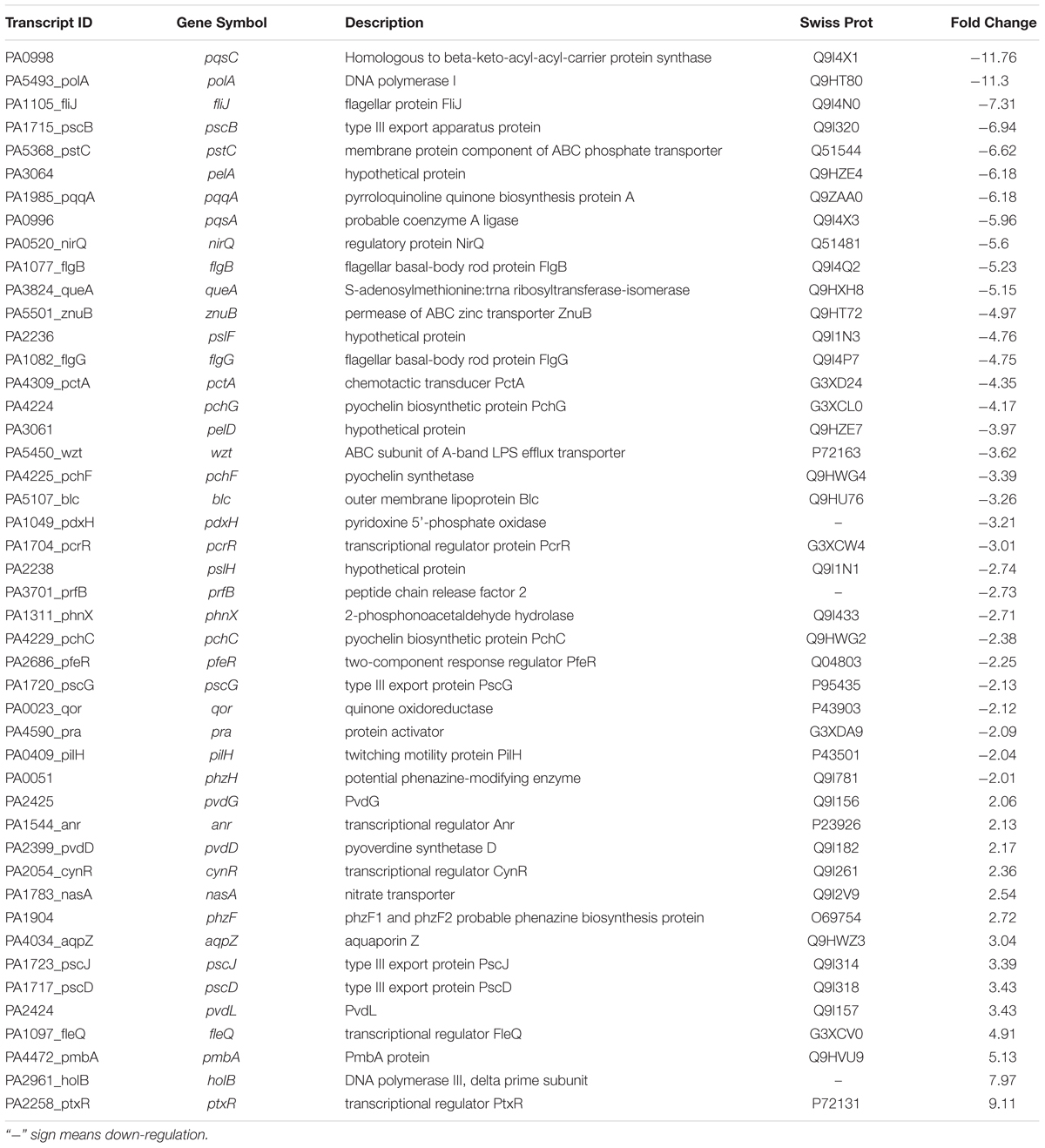
Table 1. Selected transcripts that are differentially expressed (up- or down- regulated) in P. aeruginosa PAO1, treated with the bacterial (E. indicum SJ16) active fraction compared to control (untreated PAO1 strain).
The genus Exiguobacterium is comprised of 16 species, and E. indicum was isolated from meltwater of the Hamta glacier (Himalayan mountain ranges) of India (Chaturvedi and Shivaji, 2006). The present study is the second report on the isolation of E. indicum from a rhizosphere of the coastal area after Susilowati et al. (2015), who isolated this bacteria from rice rhizosphere and reported its plant growth promotion trait. To the best of our knowledge, there is no available study, so far reporting the anti-QS activity of the genus Exiguobacterium. We herein for the first time demonstrate the inhibitory effect of E. indicum on bacterial quorum-sensing biofilm formation. In this study, the E. indicum SJ16 extract showed QSI activity (without any antibacterial activity; Supplementary Figure S3) against the reference strain C. violaceum CV026 on biosensor plates. This was also demonstrated by reduced levels of violacein in the violacein quantification assay (99% inhibition with 0.6 mg/ml SJ16 extract), which was a dose-dependent (Figure 2). A high concentration of methanolic extract (4 mg/ml) of an edible plant, Melicope lunuankenda, inhibited the response of C. violaceum CV026 to N-hexanoyl homoserine lactone (Tan et al., 2012). Similarly, Rodrigues et al. (2016) have shown that another plant methanol extract (Eugenia uniflora) showed inhibition of violacein production in C. violaceum, reaching up to 96% at the highest concentration tested. We have previously demonstrated a 98% decrease of violacein production in a dose-dependent manner with 3.7 mg/ml extract of S. maltophilia, as well as a 95% reduction with 0.1 mg/ml of D. tsuruhatensis extract, both isolated from the rhizosphere of C. laevigatus (Singh et al., 2013, 2017).
The treatment of biofilm-associated infections is extremely challenging, as biofilm-forming bacteria are commonly resistant to a broad spectrum of antibiotics (Høiby et al., 2010). P. aeruginosa, an opportunistic pathogen that causes severe infections in immunocompromised patients possesses a regulatory gene cascade that controls quorum-sensing and thus, biofilm formation and virulence factors productions. Quorum-sensing inhibition with potent QSI compounds shows promise in tackling biofilm formation in the setting of bacterial infections (Kalia, 2013; Singh et al., 2017; Quecán et al., 2018). In the field of drug discovery, extracts from natural products, especially those of microbial origin, have always been a tremendous source effective novel therapeutics (Gillespie et al., 2002; Courtois et al., 2003). These compounds have the ability to interfere with the QS system, inhibit the expression of virulence factors and prevent biofilm formation. In the present study, the E. indicum SJ16 extract robustly inhibits the biofilm formation of two reference P. aeruginosa strains (PAO1 and PAH) by modulating virulence factors (Figure 5), without affecting the planktonic growth of these bacteria (Figure 3). It is possible that these observations are the effect of the active compound within the extract that hinder the P. aeruginosa QS regulatory cascade (Table 1). These results are in accordance to our previous work describing the quorum quenching and anti-biofilm forming activities of the extracts from S. maltophilia BJ01 and D. tsuruhatensis SJ01 that we isolated from the rhizosphere of C. laevigatus (Singh et al., 2013, 2017).
In this study, the active compound that we herein describe was fractionated and identified as 3-Benzyl-hexahydro-pyrrolo[1,2-a]pyrazine-1,4-dione with a theoretical mass of 244.28 (RT-16.018) and an experimental mass of 249.28 (m/z) (Figure 1). Interestingly, a similar to this molecule, Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro compound that was extracted from the newly identified species of Streptomyces mangrovisoli showed significant antioxidant and free-radical scavenging activities (Ser et al., 2015). Furthermore, the Hexahydropyrrolo[1,2-a]pyrazine-1,4-dione compound, isolated from the Shewanella sp. Lzh-2 possessed algicidal activity against several cyanobacterial and algal strains (Li et al., 2014). Moreover, the compound Pyrrolo[1,2-a]pyrazine-1,4-dione,hexahydro-3-(phenylmethyl) compound, obtained from the Streptomyces sp. VITPK9 showed anticandidal activity against Candida albicans, C. krusei, and C. tropicalis (Sanjenbam et al., 2014). Additionally, the Pyrrolo[2,1-c][1,4]benzodiazepine compound was demonstrated to be an effective anti-tumor drugs (Cipolla et al., 2009), while the recently described hordenine compound that was obtained from sprouting barley showed significant QSI and anti-biofilm effects against P. aeruginosa (Zhou et al., 2018). Taken together, compounds that are similar to the one described in our study present great potential as bioactive molecules with diverse functions including antimicrobial, anti-cancer, antioxidant and now, anti-QS activity.
As stated above, the SJ16 extract inhibits the biofilm formation of the reference P. aeruginosa strains PAO1 and PAH in a concentration-dependent manner without affecting their planktonic growth (Figure 3). Furthermore, epi-fluorescence microscopy confirmed the viability of P. aeruginosa cells within the biofilm (Figure 4A). Thus, the possibility of an inhibitory effect of the SJ16 extract on the growth of these reference strains (PAO1 and PAH) was ruled out. The SEM and AFM images suggest an alteration in the topology of the biofilm in the samples supplemented with the extract, compared to their corresponding controls, while a growing biofilm with dispersed bacterial cells was developed by P. aeruginosa strains in the presence of the SJ16 extract (Figures 4B,C). Disruption of biofilm architecture is a promising strategy to inhibit biofilm formation of drug-resistant P. aeruginosa strains. Importantly, in order for the bacteria to form a biofilm, they need to attach to a surface or substratum. Bacterial motility is crucial in the effort of the microbes to reach the substratum. Once attached to the surface, the bacteria spread all around via swarming and swimming type of motility, ultimately leading to biofilm formation over the surface (O’May and Tufenkji, 2011). Inhibition of the swarming and swimming motility of PAO1 and PAH was observed in the presence of the SJ16 extract (Figure 5A). It is possible that the SJ16 extract may also have the capability to block the initial attachment of P. aeruginosa by preventing the bacterial motility toward the surface. This strategy could potentially open new avenue in the effort to halt bacterial spreading and thus minimize the ability of microbes to form biofilms (Singh et al., 2017).
The P. aeruginosa pathogenicity depends on the ability of this microbe to produce virulence factors. Pyocyanin, rhamnolipids, elastase, and protease are the key virulence factors which are highly expressed by P. aeruginosa during QS, infection and biofilm formation (Driscoll et al., 2007; Sarabhai et al., 2013). Elastase and protease are involved in the early steps of host colonization by the bacterial cells, pyocyanin is crucial for the demonstration of P. aeruginosa virulence, while rhamnolipids facilitate the bacterial motility. The combined effect of the afore-mentioned microbial factors eventually lead to biofilm formation (O’May and Tufenkji, 2011; Sarabhai et al., 2013). Elastase and protease activities, as well as the production of pyocyanin and rhamnolipid of both P. aeruginosa strains PAO1 and PAH were significantly reduced following SJ16 extract supplementation (Figure 5B), demonstrating the ability of the extract to attenuate the P. aeruginosa virulence functions, largely regulated by the las and rhl regulatory gene cascades (De Kievit and Iglewski, 2000). Similar results were obtained with the extract of Terminalia chebula and D. tsuruhatensis (Sarabhai et al., 2013, Singh et al., 2017).
The microarray analysis revealed that at least 1,237 genes were differentially expressed following SJ16 extract supplementation, 868 out of which were down-regulated, whereas 369 were up-regulated (Supplementary Table S1 and Figure 6). These microarray results demonstrated that a plethora of genes affected by the SJ16 extract are involved in QS regulation and biofilm formation. More specifically, these genes are crucial in the control of bacterial QS, virulence, motility, cell metabolism, cell wall synthesis, and transcriptional regulation, while some others encode hypothetical proteins (Table 1). The expression profile of cells treated with the SJ16 extract containing the active 3-Benzyl-hexahydro-pyrrolo[1,2-a]pyrazine-1,4-dione compound showed that this agent downregulates genes responsible for the synthesis of the flagellar protein, the type III export apparatus protein, the flagellar basal-body rod protein, the pyochelin biosynthetic protein, the phenazine-modifying enzyme, the type III export protein, the twitching motility protein, as well as some hypothetical proteins. The extract also represses the expression of gene(s) which are involved in the biosynthesis of transcriptional regulators/ activators of QS network. These genes are controlled by QS systems and are closely associated with the pathogenicity of P. aeruginosa (Wagner et al., 2003). Similar results were obtained when P. aeruginosa PAO1 cells were treated with 1,2-benzenedicarboxylic acid, diisooctyl ester (Singh et al., 2017).
Based on the results that we herein report, we adopted a hypothetical model that elucidates the transcriptional regulation of P. aeruginosa virulence mediated by the by E. indicum SJ16 extract (Figure 7). We hypothesized that the identified active compound, 3-Benzyl-hexahydro-pyrrolo[1,2-a]pyrazine-1,4-dione lowers the expression of key regulatory genes including, pqsA, pqsC, pscB, pstC, pqqA, and qor (Table 1), in addition to the differential expression of other genes (Supplementary Table S1). Alteration in the transcript expression of these genes leads to lower levels of HHQ. Thus, it is possible that 3-Benzyl-hexahydro-pyrrolo[1, 2-a]pyrazine-1,4-dione modulates the function of the pqs transcriptional regulatory system, hence controlling the production of MvfR (also known as PqsR). Inhibition of the MvfR system decreases the production of QS activators and signaling molecules, thus regulating the las and rhl transcription cascades. Subsequently, rhl leads lower levels of pyocyanin and rhamnolipid, decreased protease and elastase activities as well as reduced bacterial motility (Déziel et al., 2005). Our results indicate that the active compound present in the SJ16 extract decreases the virulence activity through modulating the pqs transcription regulation system. The proposed model is a schematic representation of the regulatory mechanism, illustrated based on the available literature. However, a comprehensive study is required to confirm the precise role of the identified compound, 3-Benzyl-hexahydro-pyrrolo[1,2-a]pyrazine-1,4-dione in the QSI regulation mechanism as well as its interacting partners.
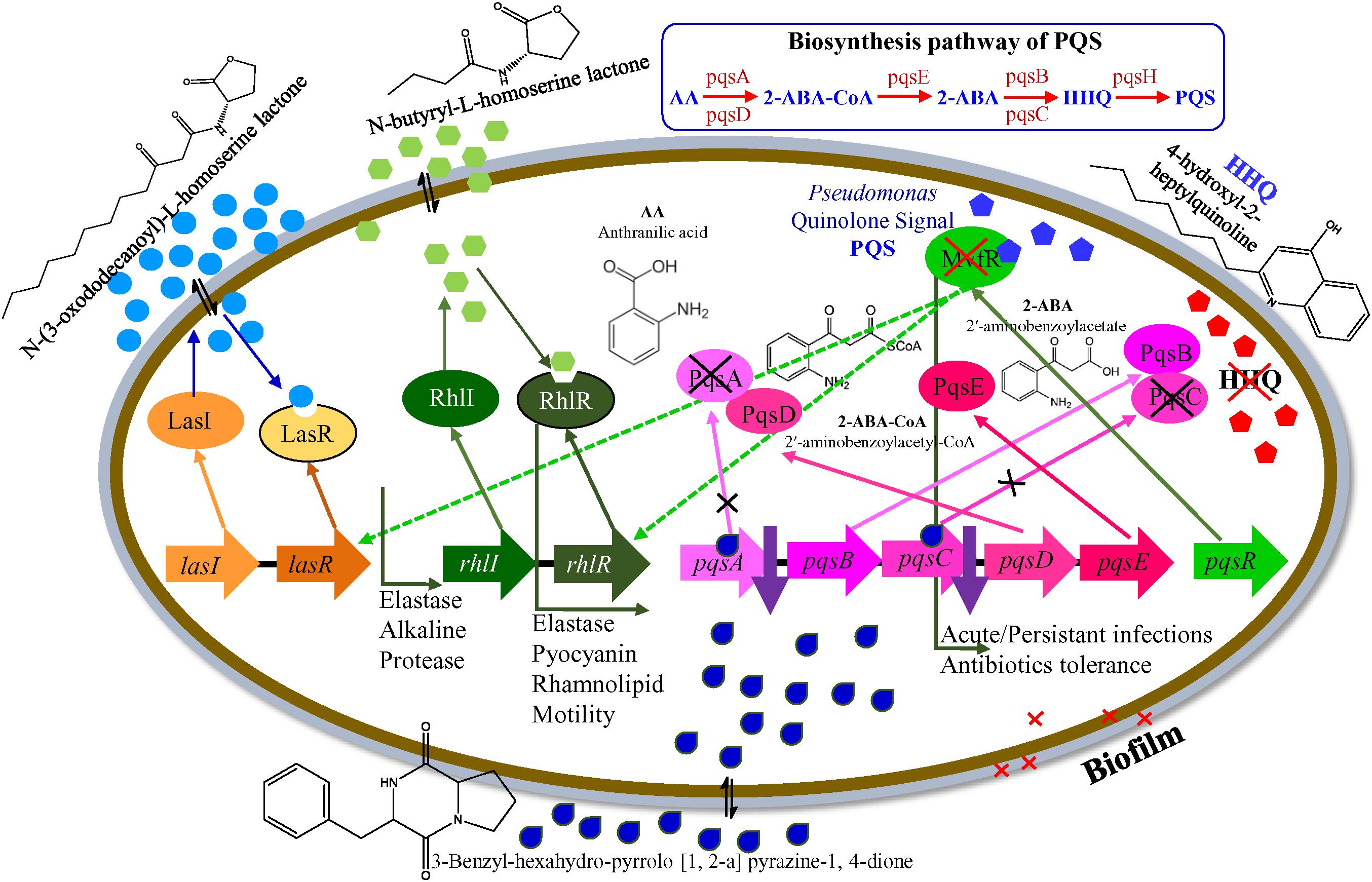
Figure 7. A hypothetical model for the transcriptional regulation of QSI in P. aeruginosa PAO1.
The QS regulates virulence factor activities and biofilm formation, and disrupting the QS mechanism is an important strategy to inhibit pathogenicity of P. aeruginosa strains. Anti-QS compounds provides a useful tool in the effort to tackle infections caused by pathogenic bacteria. E. indicum SJ16, a microbe isolated from the rhizosphere of C. laevigatus showed promising anti-QS and anti-biofilm activities, without exhibiting any anti-bacterial properties. The 3-Benzyl-hexahydro-pyrrolo[1, 2-a]pyrazine-1,4-dione compound present in the SJ16 extract was identified as a potentially active agent inhibiting the biofilm formation of two references P. aeruginosa strains, PAO1 and PAH by decreasing their swimming and swarming motility and by regulating the production of virulence factors such as pyocyanin, rhamnolipid, elastase, and protease. Furthermore, it is possible that this compound controls the pqs QS system, thus regulating the bacterial QS mechanism as indicated by our proposed inhibitory model. Overall, our results indicate that the SJ16 extract did not have any toxic effect on the growth and viability of the reference P. aeruginosa strains (PAO1 and PAH), even after longer incubation periods. On the contrary, it exhibits a strong inhibitory effects on the microbial motility, on the production of virulence factors as well as on biofilm formation. Importantly, our data indicate that the SJ16 extract is able to disrupt the cell-to-cell communication (QSI) by modulating a key component of the molecular cascade regulating the P. aeruginosa of QS systems (las, rhl, and pqs). Therefore, the identified compound has great potential for drug development in our efforts to enrich our antimicrobial armamentarium. Further research is necessary to explore and determine its pharmaceutical applications.
All authors conceived and designed the experiments. VS performed the experiments. VS and AM analyzed the data and wrote the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to gratefully acknowledge Prof. Anton Hartmann (Helmholtz Zentrum, München, Germany) for providing the reference strain C. violaceum CV026. The authors are also thankful to the Government Medical College, Bhavnagar (India) for providing the clinical isolate P. aeruginosa PAH. The Analytical and Environmental Science Division and Centralized Instrument Facility of the Institute is duly acknowledged for running the samples. Prof. Uelinton Manoel Pinto, University of Sao Paulo, Brazil and Dr. Marianna Almpani, Harvard Medical School, Boston, United States are kindly acknowledged for their language editing and proofreading. CSIR-CSMCRI Communication No. PRIS-103/2018.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01269/full#supplementary-material
Adonizio, A., Kong, K. F., and Mathee, K. (2008). Inhibition of quorum sensing controlled virulence factor production in Pseudomonas aeruginosa by south florida plant extracts. Antimicrob. Agents Chemother. 52, 198–203. doi: 10.1128/AAC.00612-07
Andersson, S., Dalhammar, G., Land, C., and Rajarao, G. (2009). Characterization of extracellular polymeric substances from denitrifying organism Comamonas denitrificans. Appl. Microbiol. Biotechnol. 82, 535–543. doi: 10.1007/s00253-008-1817-3
Berendsen, R. L., Pieterse, C. M. J., and Bakker, P. A. (2012). The rhizosphere microbiome and plant health. Trends Plant Sci. 17, 478–486. doi: 10.1016/j.tplants.2012.04.001
Bjorn, M. J., Sokol, P. A., and Iglewski, B. H. (1979). Influence of iron on yields of extracellular products in Pseudomonas aeruginosa cultures. J. Bacteriol. 138, 193–200.
Chan, K., Atkinson, S., Mathee, K., Sam, C., Chhabra, S. R., Camara, M., et al. (2011). Characterization of N-acylho- moserine lactone-degrading bacteria associated with the Zingiber officinale (ginger) rhizosphere: co-existence of quorum quenching and quorum sensing Acinetobacter and Burkholderia. BMC Microbiol. 11:51. doi: 10.1186/1471-2180-11-51
Chaturvedi, P., and Shivaji, S. (2006). Exiguobacterium indicum sp. nov., a psychrophilic bacterium from the Hamta glacier of the Himalayan mountain ranges of India. Int. J. Syst. Evol. Microbiol. 56, 2765–2770. doi: 10.1099/ijs.0.64508-0
Christiaen, S. E., Brackman, G., Nelis, H. J., and Coenye, T. (2011). Isolation and identification of quorum quenching bacteria from environmental samples. J. Microbiol. Methods 87, 213–219. doi: 10.1016/j.mimet.2011.08.002
Chu, W., Zhou, S., Zhu, W., and Zhuang, X. (2014). Quorum quenching bacteria Bacillus sp. QSI-1 protect zebrafish (Danio rerio) from Aeromonas hydrophila infection. Sci. Rep. 4:5446. doi: 10.1038/srep05446
Cipolla, L., Araújo, A. C., Airoldi, C., and Bini, D. (2009). Pyrrolo [2, 1-c] [1, 4] benzodiazepine as a scaffold for the design and synthesis of anti-tumour drugs. AntiCancer Agents Med. Chem. 9, 1–31. doi: 10.2174/187152009787047743
Cirou, A., Diallo, S., Kurt, C., Latour, X., and Faure, D. (2007). Growth promotion of quorum-quenching bacteria in the rhizosphere of Solanum tuberosum. Environ. Microbiol. 9, 1511–1522. doi: 10.1111/j.1462-2920.2007.01270.x
Courtois, S., Cappellano, C. M., Ball, M., Francou, F. X., Normand, P., Helynck, G., et al. (2003). Recombinant environmental libraries provide access to microbial diversity for drug discovery from natural products. Appl. Environ. Microbiol. 69, 49–55. doi: 10.1128/AEM.69.1.49-55.2003
D’Angelo-Picard, C., Faure, D., Penot, I., and Dessaux, Y. (2005). Diversity of N-acyl homoserine lactone-producing and degrading bacteria in soil and tobacco rhizosphere. Environ. Microbiol. 7, 1796–1808. doi: 10.1111/j.1462-2920.2005.00886.x
De Kievit, T. R., and Iglewski, B. H. (2000). Bacterial quorum sensing in pathogenic relationships. Infect. Immun. 68, 4839–4849. doi: 10.1128/IAI.68.9.4839-4849.2000
Déziel, E., Gopalan, S., Tampakaki, A. P., Lépine, F., Padfield, K. E., Saucier, M., et al. (2005). The contribution of MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing circuitry regulation: multiple quorum sensing-regulated genes are modulated without affecting lasRI, rhlRI or the production of N-acyl-l-homoserine lactones. Mol. Microbiol. 55, 998–1014. doi: 10.1111/j.1365-2958.2004.04448.x
Driscoll, J. A., Brody, S. L., and Kollef, M. H. (2007). The epidemiology, pathogenesis, and treatment of Pseudomonas aeruginosa infections. Drugs 67, 351–368. doi: 10.2165/00003495-200767030-00003
Essar, D. W., Eberly, L., Hadero, A., and Crawford, I. P. (1990). Identification and characterization of genes for a second anthranilate synthase in Pseudomonas aeruginosa: interchangeability of the two anthranilate synthases and evolutionary implications. J. Bacteriol. 172, 884–900. doi: 10.1128/JB.172.2.884-900.1990
Felsenstein, J. (1985). Confidence limits on phylogenesis: an approach using the bootstrap. Evolution 39, 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Galloway, W. R., James, T. H., Bowden, S. D., Welch, M., and Spring, D. R. (2011). Quorum sensing in gram-negative bacteria: small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem. Rev. 111, 28–67. doi: 10.1021/cr100109t
Ganin, H., Yardeni, E. H., and Kolodkin-Gal, I. (2015). “Biofilms: maintenance, development, and disassembly of bacterial communities are determined by QS cascades,” in Quorum Sensing vs Quorum Quenching: A Battle with No End in Sight, ed. V. Kalia (New Delhi: Springer), 23–37. doi: 10.1007/978-81-322-1982-8_3
Gillespie, D. E., Brady, S. F., Bettermann, A. D., Cianciotto, N. P., Liles, M. R., Rondon, M. R., et al. (2002). Isolation of antibiotics turbomycin A and B fromametagenomic library of soil microbial DNA. Appl. Environ. Microbiol. 68, 4301–4306. doi: 10.1128/AEM.68.9.4301-4306.2002
Goswami, N. N., Trivedi, H. R., Goswami, A. P. P., Patel, T. K., and Tripathi, C. B. (2011). Antibiotic sensitivity profile of bacterial pathogens in postoperative wound infections at a tertiary care hospital in Gujarat, India. J. Pharmacol. Pharmacother. 2:158. doi: 10.4103/0976-500X.83279
Høiby, N., Bjarnsholt, T., Givskov, M., Molin, S., and Ciofu, O. (2010). Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. 35, 322–332. doi: 10.1016/j.ijantimicag.2009.12.011
Jha, B., Singh, V. K., Weiss, A., Hartmann, A., and Schmid, M. (2015). Zhihengliuella somnathii sp. nov., a halotolerant actinobacterium from the rhizosphere of a halophyte Salicornia brachiata. Int. J. Syst. Evol. Microbiol. 65, 3137–3142. doi: 10.1099/ijsem.0.000391
Kalia, V. C. (2013). Quorum sensing inhibitors: an overview. Biotechnol. Adv. 31, 224–245. doi: 10.1016/j.biotechadv.2012.10.004
Kang, B. R., Lee, J. H., Ko, S. J., Lee, Y. H., Cha, J. S., Cho, B. H., et al. (2004). Degradation of acyl-homoserine lactone molecules by Acinetobacter sp. strain C1010. Can. J. Microbiol. 50, 935–941. doi: 10.1139/w04-083
Kavita, K., Singh, V. K., Mishra, A., and Jha, B. (2014). Characterisation and anti-biofilm activity of extracellular polymeric substances from Oceanobacillus iheyensis. Carbohydr. Polym. 101, 29–35. doi: 10.1016/j.carbpol.2013.08.099
Lee, J., and Zhang, L. (2015). The hierarchy quorum sensing network in Pseudomonas aeruginosa. Protein Cell 6, 26–41. doi: 10.1007/s13238-014-0100-x
Li, Z., Lin, S., Liu, X., Tan, J., Pan, J., and Yang, H. (2014). A freshwater bacterial strain, Shewanella sp. Lzh-2, isolated from Lake Taihu and its two algicidal active substances, hexahydropyrrolo [1, 2-a] pyrazine-1, 4-dione and 2, 3-indolinedione. Appl. Microbiol. Biotechnol. 98, 4737–4748. doi: 10.1007/s00253-014-5602-1
McClure, C. D., and Schiller, N. L. (1992). Effects of Pseudomonas aeruginosa rhamnolipids on human monocyte-derived macrophages. J. Leukoc. Biol. 51, 97–102. doi: 10.1002/jlb.51.2.97
Oh, Y., Lee, N., Jo, W., Jung, W., and Lim, J. (2009). Effects of substrates on biofilm formation observed by atomic force microscopy. Ultramicroscopy 109, 874–880. doi: 10.1016/j.ultramic.2009.03.042
O’May, C., and Tufenkji, N. (2011). The swarming motility of Pseudomonas aeruginosa is blocked by cranberry proanthocyanidins and other tannin-containing materials. Appl. Environ. Microbiol. 77, 3061–3067. doi: 10.1128/AEM.02677-10
O’Toole, G., Kaplan, H. B., and Kolter, R. (2000). Biofilm formation as microbial development. Annu. Rev. Microbiol. 54, 49–79. doi: 10.1146/annurev.micro.54.1.49
Overhage, J., Lewenza, S., Marr, A. K., and Hancock, R. E. (2007). Identification of genes involved in swarming motility using a Pseudomonas aeruginosa PAO1 Mini-Tn5-lux mutant library. J. Bacteriol. 189, 2164–2169. doi: 10.1128/JB.01623-06
Quecán, B. X. V., Rivera, M. L. C., and Pinto, U. M. (2018). “Bioactive phytochemicals targeting microbial activities mediated by quorum sensing,” in the Biotechnological Applications of Quorum Sensing Inhibitors. ed. V. Kalia (Singapore: Springer), 397–416. doi: 10.1007/978-981-10-9026-4_19
Rashid, M. H., and Kornberg, A. (2000). Inorganic polyphosphate is needed for swimming, swarming, and twitching motilities of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 97, 4885–4890. doi: 10.1073/pnas.060030097
Rodrigues, A. C., Zola, F. G., Ávila Oliveira, B. D., Sacramento, N. T. B., da Silva, E. R., Bertoldi, M. C., et al. (2016). Quorum quenching and microbial control through phenolic extract of Eugenia uniflora fruits. J. Food Sci. 81, M2538–M2544. doi: 10.1111/1750-3841.13431
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. doi: 10.1093/oxfordjournals.molbev.a040454
Sanjenbam, P., Gopal, J. V., and Kannabiran, K. (2014). Isolation and identification of anticandidal compound from Streptomyces sp. VITPK9. Appl. Microbiol. Biotechnol. 50, 492–499. doi: 10.1134/S0003683814050081
Sarabhai, S., Sharma, P., and Capalash, N. (2013). Ellagic acid derivatives from terminalia chebula retz. Downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS One 8:e53441. doi: 10.1371/journal.pone.0053441
Ser, H. L., Palanisamy, U. D., Yin, W. F., Malek, S. N. A., Chan, K. G., Goh, B. H., et al. (2015). Presence of antioxidative agent, Pyrrolo [1, 2-a] pyrazine-1, 4-dione, hexahydro-in newly isolated Streptomyces mangrovisoli sp. nov. Front. Microbiol. 6:854. doi: 10.3389/fmicb.2015.00854
Singh, V. K., Kavita, K., Prabhakaran, R., and Jha, B. (2013). Cis-9-octadecenoic acid from the rhizospheric bacterium Stenotrophomonas maltophilia BJ01 shows quorum quenching and anti-biofilm activities. Biofouling 29, 855–867. doi: 10.1080/08927014.2013.807914
Singh, V. K., Mishra, A., and Jha, B. (2016b). “Marine bacterial extracellular polymeric substances: characteristics and applications,” in Marine Glycobiology: Principles and Applications. ed. S. Kim (Boca Raton, FL: CRC Press), 369–377. doi: 10.1201/9781315371399-27
Singh, V. K., Mishra, A., Haque, I., and Jha, B. (2016a). A novel transcription factor-like gene SbSDR1 acts as a molecular switch and confers salt and osmotic endurance to transgenic tobacco. Sci. Rep. 6:31686. doi: 10.1038/srep31686
Singh, V. K., Mishra, A., and Jha, B. (2017). Anti-quorum sensing and anti-biofilm activity of Delftia tsuruhatensis extract by attenuating the quorum sensing-controlled virulence factor production in Pseudomonas aeruginosa. Front. Cell. Infect. Microbiol. 7:337. doi: 10.3389/fcimb.2017.00337
Susilowati, D. N., Sudiana, I. M., Mubarik, N. R., and Suwanto, A. (2015). Species and functional diversity of rhizobacteria of rice plant in the coastal soils of Indonesia. Indones. J. Agric. Sci. 16, 39–50. doi: 10.21082/ijas.v16n1.2015.p39-50
Tamura, K., Nei, M., and Kumar, S. (2004). Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. U.S.A. 101, 11030–11035. doi: 10.1073/pnas.0404206101
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Tan, L. Y., Yin, W. F., and Chan, K. G. (2012). Silencing quorum sensing through extracts of Melicope lunuankenda. Sensors 12, 4339–4351. doi: 10.3390/s120404339
Wagner, V. E., Bushnell, D., Passador, L., Brooks, A. I., and Iglewski, B. H. (2003). Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J. Bacteriol. 185, 2080–2095. doi: 10.1128/JB.185.7.2080-2095.2003
Weisburg, W. G., Barns, S. M., Pelletier, D. A., and Lane, D. J. (1991). 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173, 697–703. doi: 10.1128/jb.173.2.697-703.1991
Zhou, J. W., Luo, H. Z., Jiang, H., Jian, T. K., Chen, Z. Q., and Jia, A. Q. (2018). Hordenine, a novel quorum sensing inhibitor and anti-biofilm agent against Pseudomonas aeruginosa. J. Agric. Food Chem. 66, 1620–1628. doi: 10.1021/acs.jafc.7b05035
Keywords: anti-biofilm, anti-quorum, biofilm, Exiguobacterium, Pseudomonas, quorum quenching, quorum sensing
Citation: Singh VK, Mishra A and Jha B (2019) 3-Benzyl-Hexahydro-Pyrrolo[1,2-a]Pyrazine-1,4-Dione Extracted From Exiguobacterium indicum Showed Anti-biofilm Activity Against Pseudomonas aeruginosa by Attenuating Quorum Sensing. Front. Microbiol. 10:1269. doi: 10.3389/fmicb.2019.01269
Received: 09 November 2018; Accepted: 22 May 2019;
Published: 07 June 2019.
Edited by:
Natalia V. Kirienko, Rice University, United StatesReviewed by:
Bartolome Moya Canellas, University of Florida, United StatesCopyright © 2019 Singh, Mishra and Jha. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Avinash Mishra, YXZpbmFzaEBjc21jcmkucmVzLmlu; YXZpbmFzaG1pc2hyYTExQHJlZGlmZm1haWwuY29t; YXZpbmFzaG1pc2hyYS5jc21jcmlAZ21haWwuY29t; Bhavanath Jha, amhhLmJoYXZhbmF0aEBnbWFpbC5jb20=
†Present address: Vijay K. Singh, Department of Microbiology and Immunobiology, Harvard Medical School, Boston, MA, United States; Department of Surgery, Massachusetts General Hospital, Boston, MA, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.