 Xianyun Sun
Xianyun Sun Fei Wang1,3
Fei Wang1,3 Chengcheng Hu
Chengcheng Hu Wei Xue
Wei Xue
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 10 April 2019
Sec. Fungi and Their Interactions
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.00750
This article is part of the Research Topic Sensing and Signaling of the Environment by Fungi in Health and Disease View all 20 articles
Conidiation and sexual development are critical for reproduction, dispersal and better-adapted survival in many filamentous fungi. The Neurospora crassa gene ada-6 encodes a Zn(II)2Cys6-type transcription factor, whose deletion resulted in reduced conidial production and female sterility. In this study, we confirmed the positive contribution of ada-6 to conidiation and sexual development by detailed phenotypic characterization of its deletion mutant and the complemented mutant. To understand the regulatory mechanisms of ADA-6 in conidiation and sexual development, transcriptomic profiles generated by RNA-seq from the Δada-6 mutant and wild type during conidiation and sexual development were compared. During conidial development, differential expressed genes (DEGs) between the Δada-6 mutant and wild type are mainly involved in oxidation-reduction process and single-organism metabolic process. Several conidiation related genes are positively regulated by ADA-6, including genes that positively regulate conidiation (fluffy and acon-3), and genes preferentially expressed during conidial development (eas, con-6, con-8, con-10, con-13, pcp-1, and NCU9357), as the expression of these genes were lower in the Δada-6 mutant compared to wild type during conidial development. Phenotypic observation of deletion mutants for other genes with unknown function down-regulated by ada-6 deletion revealed that deletion mutants for four genes (NCU00929, NCU05260, NCU00116, and NCU04813) produced less conidia than wild type. Deletion of ada-6 resulted in female sterility, which might be due to that ADA-6 affects oxidation-reduction process and transmembrane transport process, and positively regulates the transcription of pre-2, poi-2, and NCU05832, three key genes participating in sexual development. In both conidiation and the sexual development process, ADA-6 regulates the transcription of cat-3 and other genes participating in reactive oxygen species production according to RNA-seq data, indicating a role of ADA-6 in oxidative stress response. This was further confirmed by the results that deletion of ada-6 led to hypersensitivity to oxidants H2O2 and menadione. Together, these results proved that ADA-6, as a global regulator, plays a crucial role in conidiation, sexual development, and oxidative stress response of N. crassa.
Conidial production is critical for reproduction, dispersal and survival in many filamentous fungi. Sexual reproduction is a key feature that distinguishes eukaryotic organisms from prokaryotic organisms. It produces better-adapted progenies by driving genetic recombination and eliminating deleterious mutations (Ni et al., 2011; Heitman et al., 2013). Neurospora crassa is a multicellular ascomycete fungus in the family Sordariomycetes and has long been used as an excellent model organism for genetic and biochemical researches as well as the study of morphological development (Springer, 1993; Perkins and Davis, 2000; Davis and Perkins, 2002). From vegetative growth to conidiation or sexual reproduction, morphological changes were evident. Behind it, transcriptional levels of many genes are altered (Greenwald et al., 2010; Wang et al., 2012; Lehr et al., 2014). For example, 25% predicted genes in the genome of N. crassa are differentially expressed during conidiation (Greenwald et al., 2010), as well as during sexual development (Lehr et al., 2014). Transcription factors play important roles in activating or repressing gene expression in response to developmental signals. Thus, identification of transcription factors, which are crucial to conidial and sexual development, and characterization of their mechanisms are critical steps toward deeper understanding of how fungal morphogenesis is regulated.
Several transcription factors required for basal hyphae growth, asexual sporulation, and sexual development have been reported in N. crassa (Colot et al., 2006; Carrillo et al., 2017). Among identified 273 transcription factor genes, 33 genes were found specifically affect asexual development (Carrillo et al., 2017). Some of these genes have been extensively studied. For example, fl is required for the formation of major constriction chains (Matsuyama et al., 1974; Bailey and Ebbole, 1998). Overexpression of fl under the control of a heterologous promoter is sufficient to induce conidiation in a liquid medium which is unfavorable for conidiation (Bailey-Shrode and Ebbole, 2004). Some transcription factor genes, such as hsf-2/NCU08480 (Thompson et al., 2008) and chc-1/NCU00749 (Sun et al., 2011), were found to regulate the extent of conidiation or conidiation in response to environmental conditions. Deletion of hsf-2 does not affect hyphal growth and aerial hyphal development but dramatically reduces conidial yield (Thompson et al., 2008). CHC-1 is involved in CO2-mediated conidiation suppression, and chc-1 deletion results in earlier conidial formation than wild type, especially at a higher CO2 concentration (Sun et al., 2011). N. crassa is heterothallic with two mating types, designated mat a and mat A. Both of the mating types can form protoperithecia when nitrogen source is depleted (Davis and de Serres, 1970). Among identified transcription factor genes, ten of them were found to specifically affect sexual development (Carrillo et al., 2017). For example, ff-7/NCU04001 is required for initiation of sexual development, and deletion mutant of ff-7 does not produce protoperithecia, perithecia as well as ascopores (Colot et al., 2006; Carrillo et al., 2017); Deletion of bek-1/NCU00097 results in aberrant perithecia, which exhibit defective beaks and cannot produce ascopores (Colot et al., 2006; Carrillo et al., 2017).
In addition to these specific transcriptional regulators, some transcription factors were showed to participate in regulating both asexual sporulation and sexual development. Knockout mutants of 25 transcription factor genes display significant defects in both asexual sporulation and sexual development (Colot et al., 2006; Carrillo et al., 2017). Most of these genes were named as all development altered (ada) genes (Colot et al., 2006), including ada-1/NCU00499, ada-2/NCU02017, ada-3/NCU02896, ada-4/NCU03320, ada-5/NCU03931, ada-6/NCU04866 and ada-7/NCU09739. These transcription factors play crucial roles in fungal growth and development by regulating gene expression on a global scale. However, the regulatory roles of most of these newly found transcription factors in growth and development needs further confirmation, and their molecular mechanisms are not addressed. Among them, ada-6/NCU04866 is a representative gene, which codes a Zn(II)2Cys6 transcription factor (Borkovich et al., 2004). Deletion of ada-6 results in slower growth, dramatic reduction in conidiation and female infertility during sexual development (Colot et al., 2006), suggesting ADA-6 is an important transcription factor. However, the confirmation of its function is still required and their mechanism has not been investigated.
ADA-6 orthologs are widely distributed in filamentous fungi by sequence alignment1. The ortholog of ADA-6 in N. discreta has a predicted function involved in embryonic development2. In Aspergillus oryzae, the ADA-6 ortholog has predicted role in hyphal growth, positive regulation of secondary metabolite and sporocarp development during sexual reproduction3.
In this study, we confirmed the positive role of ADA-6 in growth and development by detailed phenotypic characterization of its deletion mutant and the complemented strain. By comparing transcriptomic profiles generated by RNA-seq from the Δada-6 mutant and wild type during conidiation and sexual development, as well as by analyzing the contribution of the genes or biological pathway influenced by ada-6 deletion, we explored the mechanisms by which ADA-6 promotes conidiation and sexual development. We also found that deletion of ada-6 causes hypersensitive to oxidants H2O2 and menadione. Together, our results proved that ADA-6 is a global regulator of conidiation, sexual development and oxidative stress response in N. crassa.
Most strains of N. crassa used in this study, including FGSC#4200 (wild type), FGSC#11022 (Δada-6/NCU04866; a), and knockout mutants for genes responsive to ada-6 deletion, were purchased from the Fungal Genetics Stock Center. All the strains were cultured at 28°C if it’s not mentioned.
Media used in this study include Vogel’s slant medium (1 × Vogel’s salts, 2% sucrose, and 1.5% Bacto Agar), Vogel’s plate medium (1 × Vogel’s salts, 2% glucose, and 0.75% Bacto Agar), liquid Vogel’s medium (1 × Vogel’s salts, 2% glucose), the agar medium for transformant regeneration (1 × Vogel’s salts, 1 M sorbitol, 1 × FGS, and 1.5% Bacto Agar), and the agar medium for filling race tubes [1 × Vogel’s salts, 2% carbon source (glucose, sucrose, xylose, xylan, or carboxymethyl cellulose sodium), and 1.5% Bacto Agar].
The plasmid pCB1532-ada6 used for complementation was created by inserting a 4547 bp DNA fragment, containing the ada-6 gene (2230 bp) flanked by a 1238 bp upstream regulatory region and a 1079 bp downstream region, into the plasmid pCB1532 which harbors a sulfonylurea resistant allele of the Magnaporthe grisea ILV1 as a selective marker (Sweigard et al., 1997). Briefly, the DNA fragment was amplified from the wild-type strain FGSC#4200 using primers Ada6F-EcoRI: GGAATTCGTAAAGTGACTGGAAGGTGG and Ada6R-HindIII: CCCAAGCTTATCAATAACATAACTGCCCCC (EcoRI and HindIII sites were underlined), digested by EcoRI and HindIII and ligated into plasmid pCB1532. The construct pCB1532-ada6 was transformed into the Δada-6 mutant FGSC#11022 according to the previously reported protoplast transformation method (Royer and Yamashiro, 1992). 15 μg/ml of chlorimuron ethyl (Sigma) was added to the top agar to inhibit the growth of non-transformed protoplasts. Obtained transformants were subjected to serial transfers on slants with 15 μg/ml chlorimuron ethyl to favor homokaryon formation (Ebbole and Sachs, 1990) and further verified by PCR.
Hyphal extension of wild type and the Δada-6 mutant were analyzed in race tube. Briefly, one piece of mycelium mat (2 mm × 10 mm) for each strain was separately inoculated on one end of the race tube containing solid Vogel’s medium with different carbon source [2% glucose, 2% sucrose, 2% xylose, 2% xylan, and 2% carboxymethyl cellulose sodium (CMC-Na), respectively]. Inoculated race tubes were incubated at 28°C and the leading edge of the colony were marked every 24 h. The hyphal extension was then documented and measured by a ruler.
For conidiation analysis, mycelium for each strain was inoculated on Vogel’s slants and grown at 28°C with continuous light for 7 days. Conidia produced were washed by 5 ml distilled water and counted with a hemocytometer.
For protoperithecium and perithecium formation analysis, the mycelial mat or conidia suspension of the strain used as female parent was first inoculated on solid synthetic crossing medium with 0.1% sucrose and grown for 5 days under constant darkness at 25°C. Then the opposite mating-type strain was inoculated as male parent and incubated at 25°C for another 7 days under constant darkness. The protoperithecium and perithecium formation were checked and documented by an optical microscope equipped with a Zeiss CCD.
Genome-wide transcriptional profiles for wild type and the Δada-6 mutant during conidiation, and at the initiate stage of protoperithecium formation were obtained by RNA sequencing, while transcriptional profiles for vegetative growth were used as control. Briefly, N. crassa wild-type strain and the Δada-6 mutant were inoculated on Vogel’s plates covered with cellophane and grown at 28°C in darkness for 24 h. The mycelia were then transferred into 150-ml flasks containing 75 ml of liquid Vogel’s medium. Cultures were incubated at 28°C with constant agitation at 180 rpm for 18 h, and the mycelia were harvested by vacuum filtration. For conidial development analysis, the mycelial mats were transferred onto the surface of agar plates (9 cm) to induce conidial development at 28°C under constant light. Cultures were sampled at 12 h intervals. For sexual development analysis, the mycelial mats were inoculated on solid synthetic crossing medium with 0.1% sucrose and grown for 4 days under constant darkness at 25°C. Then, the mycelia were harvested and total RNA was extracted according to the standard TRIzol protocol (Invitrogen Corporation, Carlsbad, CA, United States).
RNA samples were sent to Beijing Genomics Institute (BGI) for RNA-seq analysis using the Illumina Hiseq2000 with a 50 bp single-end module (Illumina, San Diego, CA, United States). The obtained raw data was treated, mapped to N. crassa genome and transformed into expression value following standard BGI workflow. The gene expression level was calculated by using RPKM (Reads per kb per million reads). The differences in gene expression between samples was compared by comparing RPKM values (Grabherr et al., 2011), and those with fold change more than 2 (FDR < 0.001) were thought to be differentially expressed genes (DEGs). In addition, genes with RPKM less than 12 at all time points were thought to be low abundant transcripts and removed from the DEGs lists. The expressions of some genes, crucial for development and oxidative stress responses, were verified by time course experiment using real time PCR.
Samples were prepared as described above. Then mycelia were harvested and immediately frozen and ground into fine powder in liquid nitrogen. Total RNA was extracted and treated with DNase I to remove genomic DNA according to the standard TRIzol protocol (Invitrogen Corporation, Carlsbad, CA, United States). cDNA was prepared with a FastQuant RT Kit (with gDNase) (Tiangen, Beijing) according to the product’s instruction. qPCR was performed on a BIO-RAD CFX96TM Real-Time System (Bio-Rad, Hercules, CA, United States) with KAPA SYBR FAST qPCR mix (Kapa Biosystems, Wilmington, MA, United States) according to the product’s protocol. Each cDNA sample was analyzed in duplicate and at least three independent experiments were conducted. The average threshold cycle was used to calculate relative expression level according to 2-ΔΔCt method (Livak and Schmittgen, 2001). And the expression level was normalized to the level of β-tubulin. The primer pairs used for RT-qPCR assay were shown in Supplementary Table S1.
N. crassa wild-type strain and knockout mutants (ada-6, nox-1, cat-2, and cat-3) were separately inoculated onto ϕ90-mm plates (containing 15-ml liquid Vogel’s medium) and allowed to grow at 28°C in darkness for 24 h. The mycelial mat were punched and the round mat (ϕ2-mm) were inoculated on the center of plates (ϕ90-mm) with or without oxidant (H2O2 or menadione), and incubated at 28°C for 22 h (control), 32 h (25 μg/ml menadione) and 48 h (10 mM H2O2), respectively. Each test was duplicated and the experiment was independently repeated at least three times. The relative growth inhibition rates (mutant growth under oxidant stress was compared to wild type grown under oxidant stress and normalized by growth under non- oxidant condition) of each strain were calculated based on colony diameters after 22 h of incubation.
The phenotype of the Δada-6 mutant has been described by Colot et al. (2006). Deletion of ada-6 resulted in reduced hyphal growth and altered asexual and sexual development. However, the deletion of ada-6 only slightly affected colony growth. In race tubes containing solid Vogel’s medium with different carbon sources, the colony growth of the Δada-6 mutant were slightly slower than that of wild type: with 2% glucose, 2% sucrose, 2% xylose, 2% xylan, and 2% carboxymethyl cellulose sodium (CMC-Na) as carbon sources, the growth rate of the Δada-6 mutant was 6.5, 6.6, 5.6, 7.2, and 4.9 cm/day, respectively, while the growth rate of wild type was 8.5, 8.3, 7.5, 8.3, and 5.6 cm/day, respectively (Figure 1A).
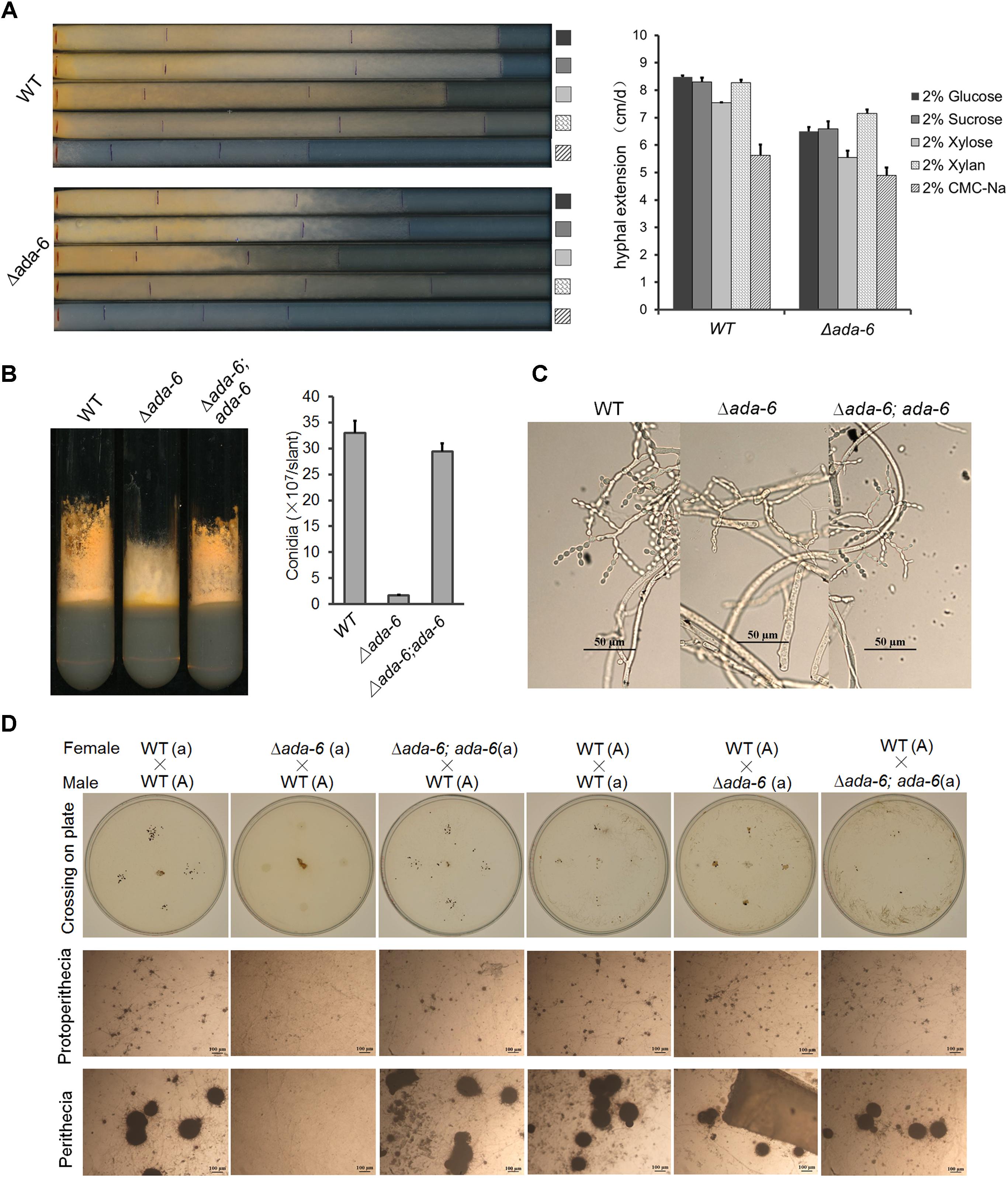
Figure 1. Deletion of ada-6 results in reduced hyphal growth, less conidial production, and female sterility in N. crassa. (A) Hyphal growth characterization of the ada-6 deletion mutant (Δada-6) and wild type (WT) grown with different carbon source. Strains were grown in race tube containing solid Vogel’s medium with different carbon sources. Inoculated race tubes were incubated at 28°C and the leading edge of the colony were marked every 24 h. The hyphal extension was then measured by a ruler. The means of hyphal extension rates from three race tubes are shown and standard deviations are indicated by error bars. (B) Conidiation characterization of wild type (WT), the ada-6 deletion mutant (Δada-6) and it’s complemented transformant (Δada-6; ada-6). Strains were grown in Vogel’s slants at 28°C with continuous light for 7 days and then imaged. Conidia produced on slants were counted with a hemocytometer. The means of conidial counts from three slants are shown and standard deviations are indicated by error bars. (C) Conidiophore structure of wild type (WT), the ada-6 deletion mutant (Δada-6) and it’s complemented transformant (Δada-6; ada-6). Bar, 50 μm. (D) Protoperithecium and perithecium formation by crossing of the ada-6 deletion mutant (Δada-6, a) with wild type (#2225, A). The ada-6 deletion mutant (Δada-6, a) or wild type (#2225, A) were used as female parent and first grown on solid crossing medium for 5 days under constant darkness at 25°C, then the opposite mating type strain was inoculated as male parent and incubated at 25°C for another 7 days under constant darkness. Protoperithecium and perithecium formation was checked and imaged.
The most dramatic effects caused by ada-6 deletion were the defects in asexual sporulation and sexual development. On slants containing solid Vogel’s medium with 2% sucrose, aerial hyphal growth of the Δada-6 mutant was only slightly shorter than that of wild type (Figure 1B), but the conidial production of the Δada-6 mutant was reduced by 93% as compared with that of wild type (Figure 1B). Unlike the deletion mutant of fl in which conidial development stops at the major constriction formation stage (Bailey and Ebbole, 1998), the Δada-6 mutant was capable to pass through all conidial development stages to produce mature conidia (Figure 1C).
To investigate the contribution of ADA-6 to sexual development, we analyzed the formation of protoperithecium and perithecium on solid synthetic crossing medium. When using the Δada-6 (a) as female parent and wild type FGSC#2225 (A) as male parent, only a very few and small protoperithecium formed but no perithecium was observed (Figure 1D). While using wild type 2225 (A) as female parent and the Δada-6 (a) as male parent, normal protoperithecium and perithecium were produced (Figure 1D). These results suggest that deletion of ada-6 resulted in female sterility.
To confirm the role of ADA-6 in growth and development, we generated a plasmid pCB1532-ada6 which carries the full length of ada-6 gene with its regulatory regions as described in Materials and Methods. By transforming this plasmid into the Δada-6 mutant FGSC#11022, complemented transformants (Δada-6; ada-6) were obtained and phenotypically compared the growth and development to wild type FGSC#4200 (WT). As expected, the complemented transformants (Δada-6; ada-6) displayed phenotypes resembling wild-type conidiation and sexual development (Figures 1B–D).
To understand the regulatory roles of ADA-6 in growth, conidiation and sexual development, transcriptomic profiles generated by RNA-seq from the Δada-6 mutant and wild type during conidial development or sexual development were compared. Among 9403 detected genes, 330, 441, and 1547 genes were found to be transcriptionally changed for more than two folds upon ada-6 deletion after conidiation induction for 0, 12, and 24 h, respectively (Supplementary Data S1). For sexual development, 1024 genes were found to be transcriptionally changed for more than two folds upon ada-6 deletion after 4 days of induction of sexual development (Supplementary Data S1).
To functionally understand the DEGs, functional classification and gene set enrichment analysis were conducted. In the vegetative growth stage (or conidiation induction for 0 h), DEGs between the Δada-6 mutant and wild type were mainly enriched in carbohydrate metabolic process (14 up- and 2 down-regulated genes), xylan catabolic process (5 up- and 2 down-regulated genes) and glucose import (6 up- and 2 down-regulated genes) (Supplementary Table S2). After 12 h of conidiation induction, DEGs between the Δada-6 mutant and wild type were mostly enriched in oxidation-reduction process (30 up- and 6 down-regulated genes) and transmembrane transport process (21 up- and 3 down-regulated genes) (Supplementary Table S2). While after 24 h of conidiation induction, DEGs between the Δada-6 mutant and wild type are mostly involved in oxidation-reduction process (107 up- and 111 down-regulated genes) and single-organism metabolic process (188 up- and 195 down-regulated genes) (Supplementary Table S2). During sexual development, DEGs between the Δada-6 mutant and wild type mostly participated in oxidation-reduction process (41 up- and 66 down-regulated genes) and transmembrane transport process (42 up- and 34 down-regulated genes) (Supplementary Table S2).
Under all tested conditions, the expression of 14 genes (cat-3, NCU02910, NCU03323, NCU04917, NCU5126, NCU05230, sut-28, NCU06170, NCU07088, NCU07095, cdt-2, NCU08223, pho-3, adh-9) were commonly increased and the expression of 13 genes (NCU00496, NCU00719, NCU05629, NCU05762, NCU05859, NCU06140, NCU08455, eas, gao-1, NCU09210, NCU10610, NCU11340, NCU17271) were commonly reduced in response to ada-6 deletion (Tables 1–4). Among these genes, seven genes (sut-28, cdt-2, NCU17271, NCU10610, NCU05762, NCU05230, NCU05126) encode proteins as integral components of plasma membrane, five genes (cat-3, gao-1, NCU09210, NCU05762, adh-9) are involved in oxidation-reduction process, and nine genes (NCU11340, NCU08455, NCU08223, NCU07088, NCU06170, NCU05859, NCU05629, NCU03323, NCU02310) encode proteins with unknown function (Table 1).
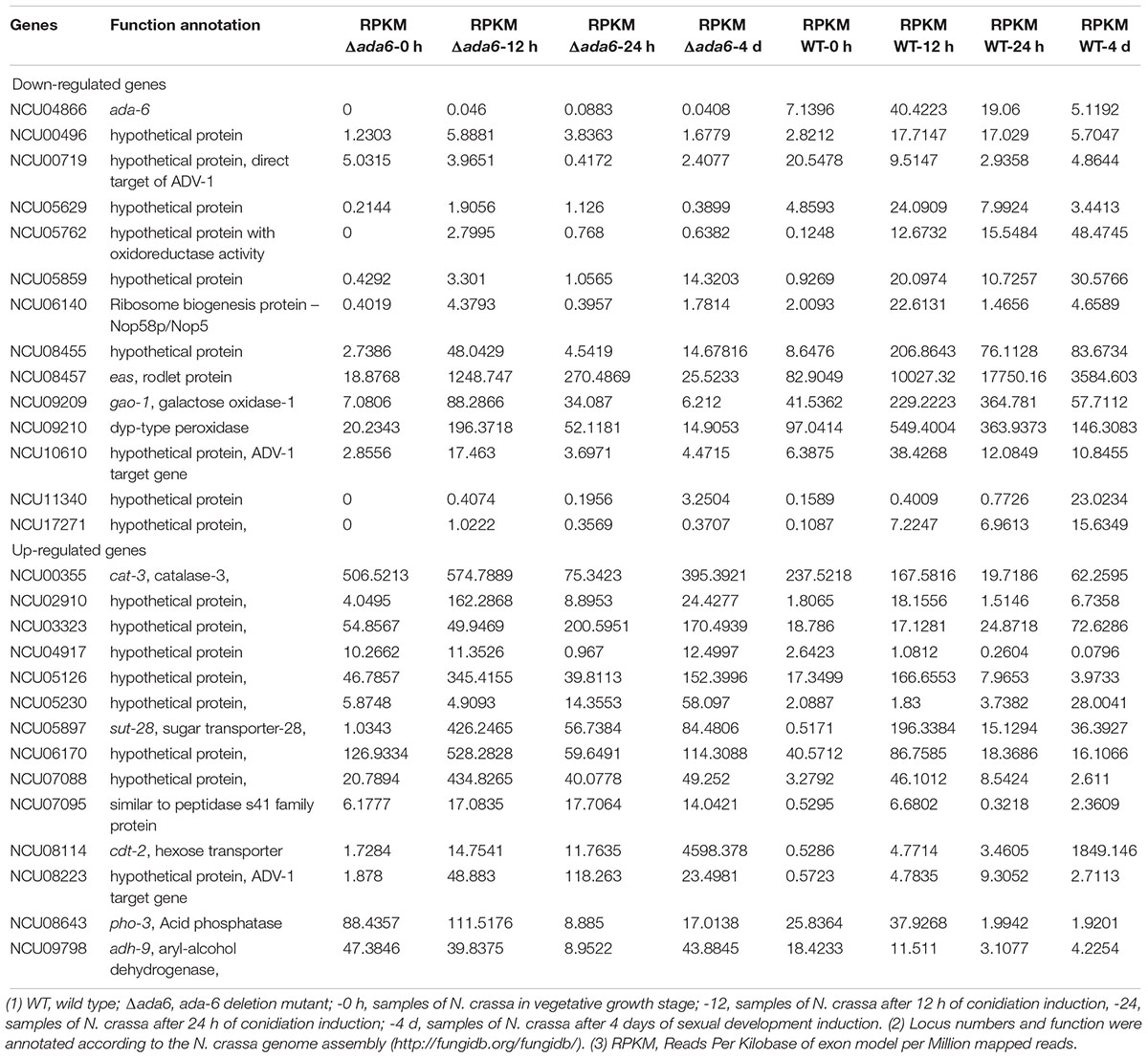
Table 1. Genes transcriptionally response to ada-6 deletion under all tested conditions.
During both conidiation and sexual development, the most seriously affected biological process by ada-6 deletion is oxidation-reduction reaction. After conidiation induction for 12 and 24 h, 36 and 218 genes, respectively, which involved in oxidation-reduction process, were differentially expressed between the Δada-6 mutant and wild type (Supplementary Data S1). The expressions of some crucial genes were verified by time course experiment using real time PCR (Figure 2). During the conidiation induction, the transcriptional levels of 15 genes, involved in oxidation-reduction reaction, were increased in wild type. However, their transcriptional levels in the Δada-6 mutant were obviously lower than those in wild type (Table 2). Most of these 15 DGEs encode oxidase or dehydrogenase, including NCU08856 (myo-inositol oxygenase), NCU05858 (fatty acid oxygenase), NCU09209 (galactose oxidase), NCU10015 (methanesulfonate monooxygenase), NCU04474 (sulfite oxidase), NCU01853 (choline dehydrogenase), NCU03893 (short-chain dehydrogenase/reductase SDR), NCU02287 (acyl-CoA dehydrogenase-1), and three peroxidase encoding genes (NCU09210, cat-1 and cat-4) (Table 2). There were 13 genes in wild type were down-regulated during conidiation, but their transcriptional levers in the Δada-6 mutant were higher than those in wild type. These 13 DGEs include three oxidase encoding genes (NCU06402, NCU04983 and NCU01546), two peroxidase encoding genes (cat-3 and NCU10051), two dehydrogenase encoding genes (NCU09798 and NCU01754), and two hydrolase encoding genes (NCU01720 and NCU05969), etc. (Table 2).
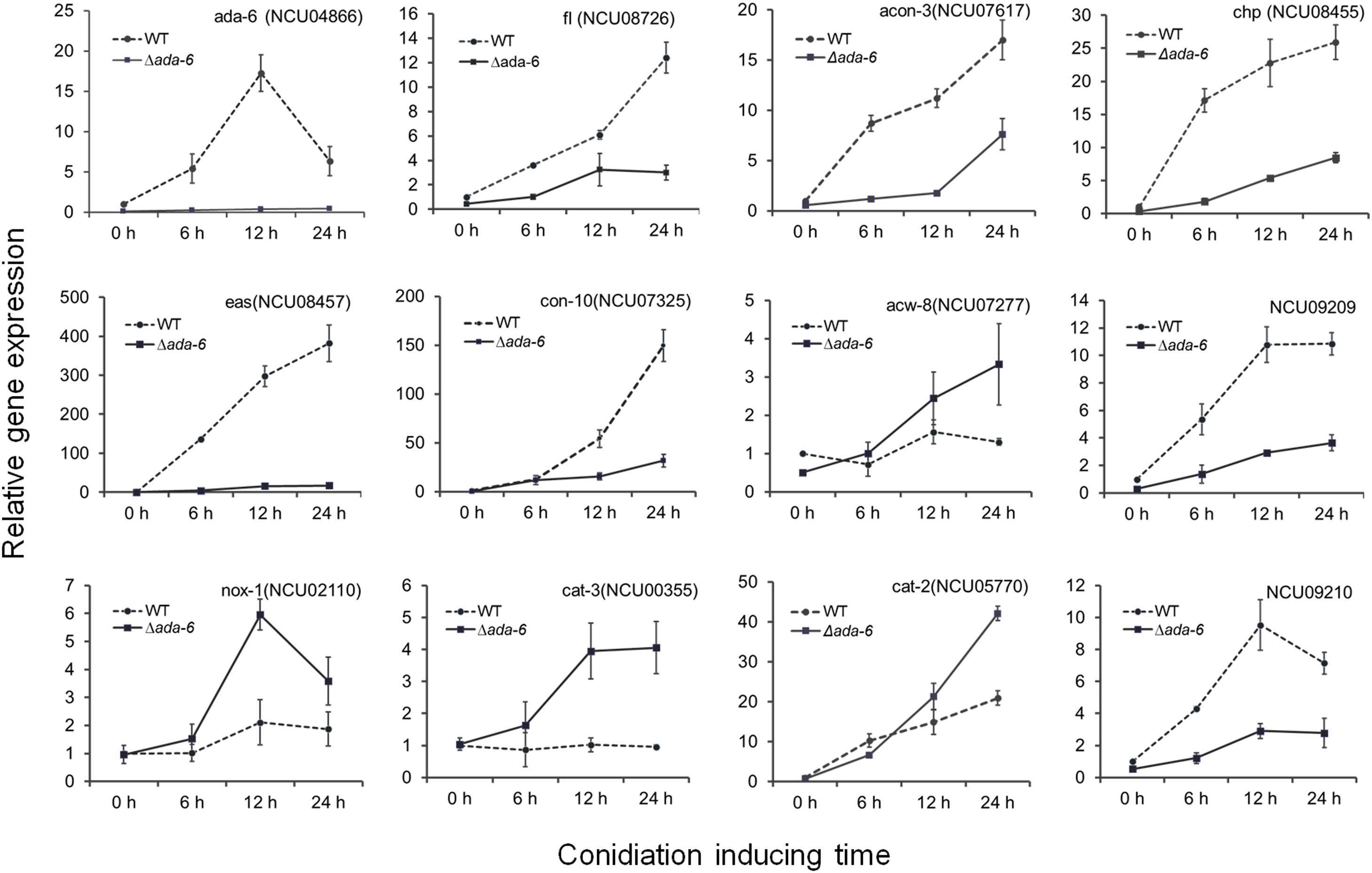
Figure 2. The expressions of some DGEs, crucial for development and oxidative stress responses, were determined by time course experiment using RT-qPCR. Wild type (WT) and the ada-6 deletion mutant (Δada-6) were inoculated on Vogel’s plates and allowed to grow at 28°C in darkness for 24 h. The mycelia were then transferred into 150-ml flasks containing 75 ml of liquid Vogel’s medium (2% sucrose). Cultures were incubated at 28°C with constant agitation at 180 rpm for 18 h, and the mycelia were harvested by vacuum filtration and transferred onto the surface of agar plates (9 cm) to induce conidial development at 28°C under constant light. Cultures were sampled after induction for 6, 12, and 24 h. The total RNA was extracted and transcriptional levels of indicated genes were analyzed by RT-qPCR.
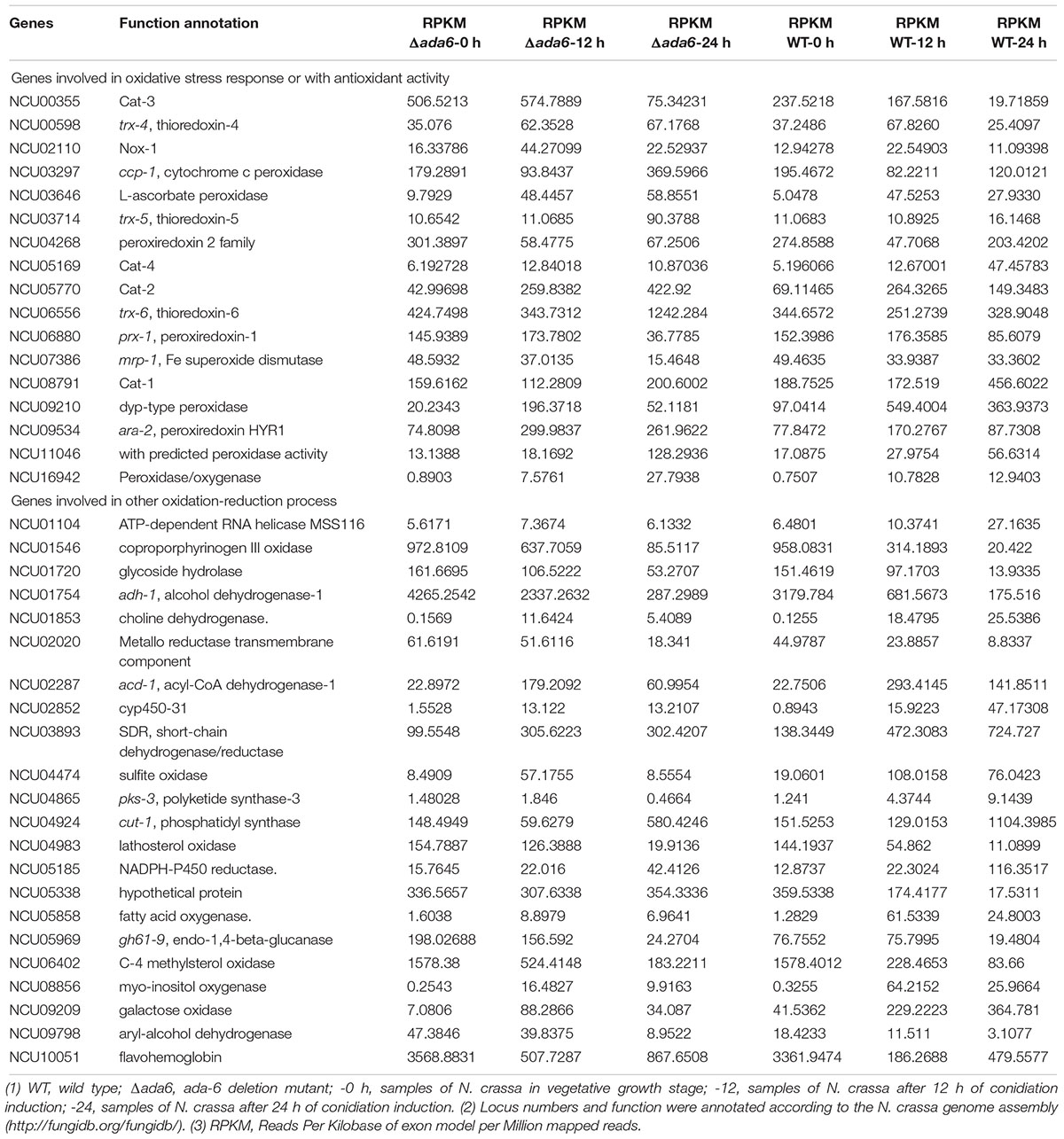
Table 2. Transcriptional responses to ada-6 deletion by the genes involved in oxidation-reduction process.
Oxidation-reduction reaction is involved in many processes. The foremost is oxidative stress response pathway, which affect fungal growth, development and stress responses. During the conidiation induction, 15 genes involved in oxidative stress response or with antioxidant activity were differentially expressed between the Δada-6 mutant and wild type (Table 2). After 24 h of conidiation induction, the transcriptional levels of 11 genes (nox-1, cat-2, cat-3, NCU03646, NCU09534, NCU16942, ara-2, NCU16942, ccp-1, NCU11046, trx-4, trx-5 and trx-6) in the Δada-6 mutant were obviously higher than those in wild type. Among these 11 genes, the transcriptional levels of nox-1 and cat-3 in the Δada-6 mutant were higher than those in wild type by both 12 and 24 h after conidiation induction (Table 2 and Figure 2). NOX-1 participates in reactive oxygen species (ROS) production, and CAT-3 activity increases during exponential growth and is induced under various stress conditions (Chary and Natvig, 1989; Peraza and Hansberg, 2002). The gene cat-2 encodes catalase-2, which is mainly found in aerial hyphae and conidia (Peraza and Hansberg, 2002). The coding products of NCU03646, NCU09534, NCU16942, ara-2, NCU16942, ccp-1, NCU11046 have peroxidase activity, and trx-4, trx-5 and trx-6 encode thioredoxin. All these genes are induced by ROS or other stresses (Circu and Aw, 2010; Matsuo and Yodoi, 2013). The higher transcription of these 11 genes in the Δada-6 mutant than those in wild type may suggest that the Δada-6 mutant produce more ROS than wild type during the conidiation induction. For the genes cat-1, cat-4, NCU07386 (mrp-1, Fe superoxide dismutase), NCU09210 (dyp-type peroxidase), NCU04268 (peroxiredoxin 2), and NCU06880 (prx-1, peroxiredoxin-1), their transcriptional levels in the Δada-6 mutant were obviously lower than those in wild type after 24 h of conidiation induction (Table 2). The transcriptional levels of cat-1 and cat-4 were increased in wild type but not changed in the Δada-6 mutant during the late period of conidiation (Table 2). CAT-1 is highly abundant in conidia and function mainly in condia germination (Peraza and Hansberg, 2002; Wang et al., 2007), CAT-4 is located in the cytosol (Schliebs et al., 2006), and both CAT-4 and dyp-type peroxidase (NCU09210) were induced during the conidiation (Greenwald et al., 2010). As expression of these genes is correlated with conidiation, the lower transcription of the cat-1, cat-4, and NCU09210 in the Δada-6 mutant compared with those in wild type is consistent with the less sporulation phenotype of the Δada-6 mutant.
During sexual development, 107 genes involved in oxidation-reduction process were found differentially expressed between the Δada-6 mutant and wild type (Supplementary Table S2 and Table 4). After 4 days of sexual development induction, the transcriptional levels of 61 genes in wild type were increased or not changed, but were obviously lower in the Δada-6 mutant than in wild type. While the transcriptional levels of 26 genes were obviously higher in the Δada-6 mutant than in wild type. Among these 107 genes, 15 genes are involved in oxidative stress response or with antioxidant activity (Table 4). After 4 days of sexual development induction, the transcriptional levels of 7 genes (nox-1, nox-2, nox-R, cat-3, NCU03151, NCU08114, and NCU07966) in the Δada-6 mutant were obviously higher than those in wild type. NOX-1, NOX-2, and NOX-R participate in ROS production, and CAT-3 activity increases during exponential growth and is induced under various stress conditions (Chary and Natvig, 1989; Peraza and Hansberg, 2002). NCU03151 encodes a peroxisomal membrane protein, and NCU08114 (cdt-2) encodes a hexose transporter. The higher expression of these seven genes in the Δada-6 mutant suggests that the Δada-6 mutant may be still metabolically active and produced more ROS than wild type after 4 days of sexual development induction. For the genes sod-1, cat-1, cat-4, NCU09210, NCU07966, NCU05858, NCU11286, and NCU03651, their transcriptional levels in the Δada-6 mutant were obviously lower than those in wild type (Table 4). As expression of these genes is correlated with conidiation, the lower expression of these eight genes in the Δada-6 mutant than in wild type is consistent with their phenotypic characteristics, as asexual development is companied with sexual development after 4 days of sexual development induction.
Many genes are associated with conidiation in N. crassa. Among the genes positively regulating conidiation, acon-3 and fl were differentially expressed between the Δada-6 mutant and wild type (Table 3). The result was verified by time course experiment, in which the expression profiles of acon-3 and fl were analyzed by real time PCR (Figure 2). During the conidiation induction, the transcriptional level of acon-3 and fl in wild type were significantly increased after conidiation induction. At 12 h, the acon-3 and fl transcriptional level was 11.2 times and 6.1 times higher than that at the initial time point, respectively. However, the transcriptional increases of acon-3 and fl were only 1.7-fold and 3-fold in the Δada-6 mutant after 12 h induction. Similar results were found at 24 h of conidiation induction. The transcriptional levels of acon-3 and fl in the Δada-6 mutant were lower than those in wild type during the entire experiment (Figure 2).
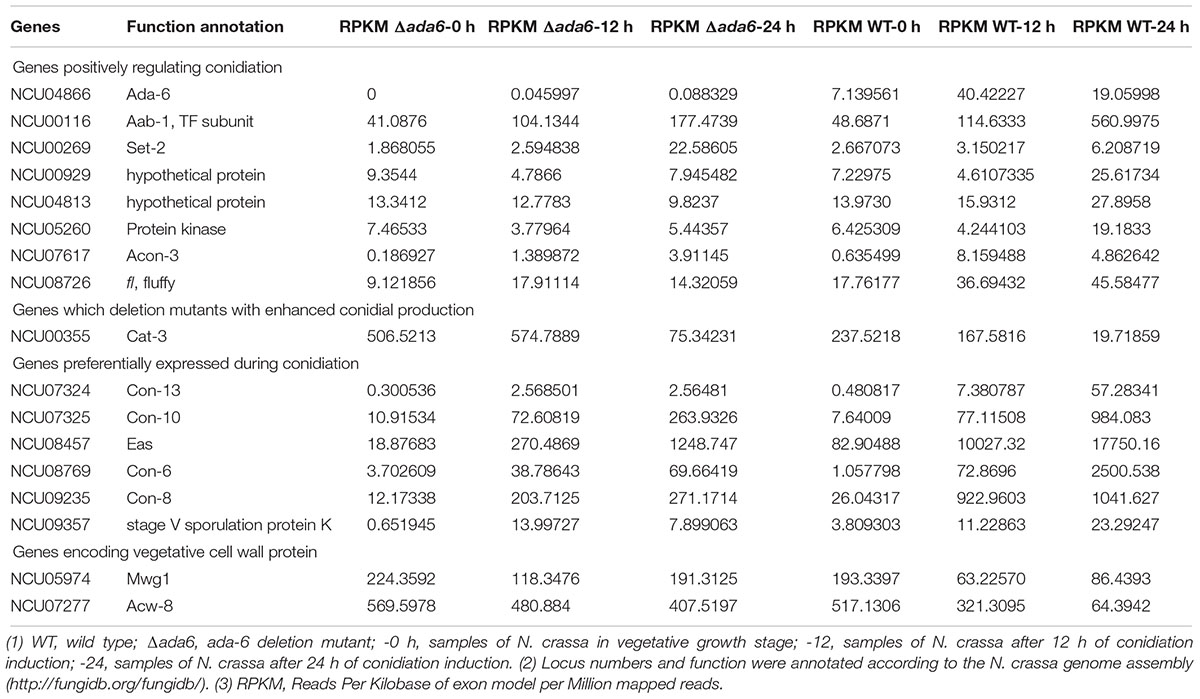
Table 3. Transcriptional response to ada-6 deletion by the genes involved in conidiation and vegetative cell wall development.
For DEGs with unknown functions, we analyzed conidiation of the corresponding gene deletion mutants grown on Vogel’s slants and found that mutants for four genes, including NCU00929, NCU05260, NCU00116, and NCU04813, displayed reduced conidial production (Figure 3). NCU05260 encodes a protein kinase, NCU00116 encodes a CCAAT-binding transcription factor subunit AAB-1 (Chen et al., 1998), and both NCU00929 and NCU04813 encode hypothetical protein. The transcriptional levels of these four genes were increased dramatically at 24 h after conidiation induction, but their transcriptional levels were lower in the Δada-6 mutant than those in wild type (Table 3).
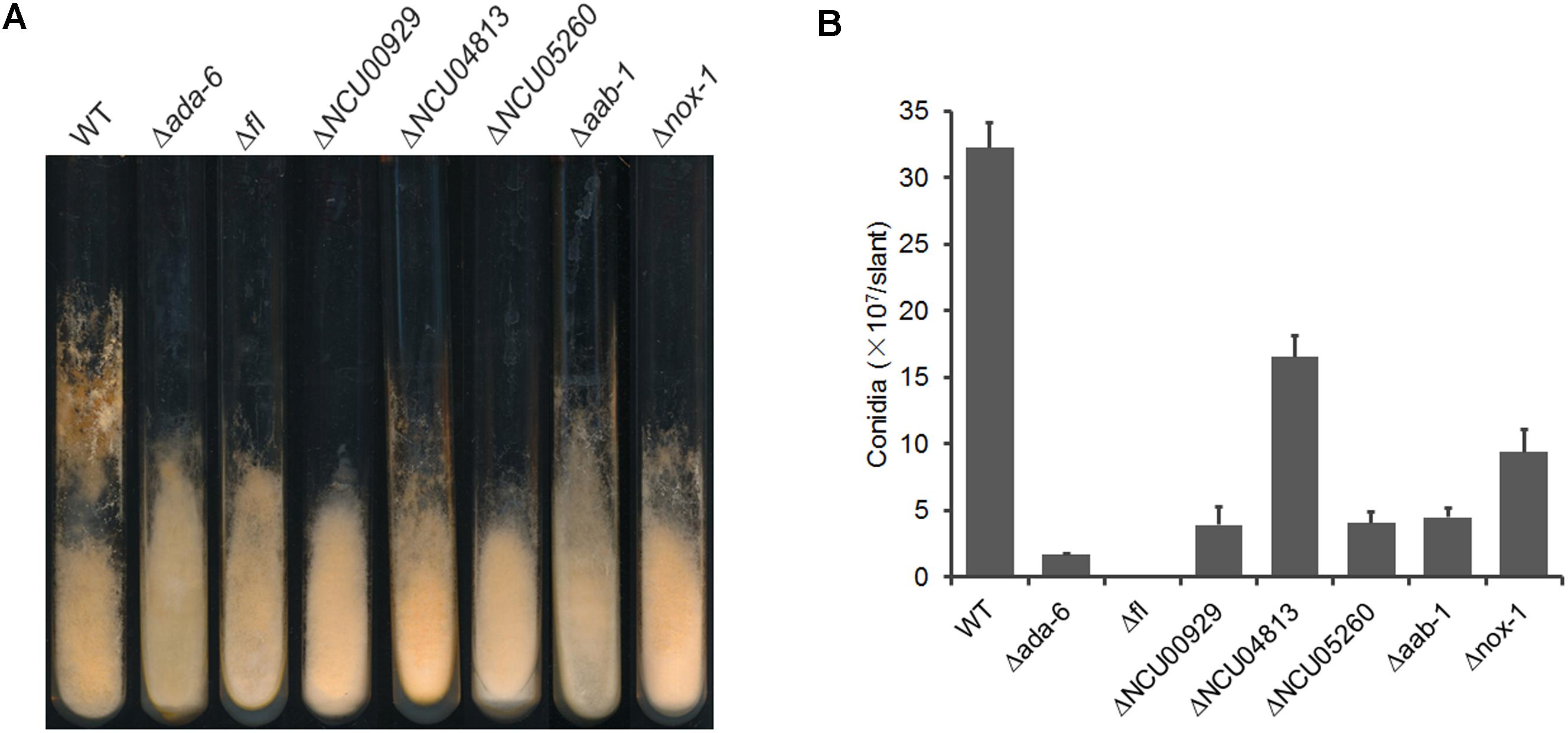
Figure 3. Conidial production of the knockout mutants of genes regulated by ADA-6. (A) Wild type and knockout mutants were inoculated and grown on Vogel’s slants at 28°C with constant darkness for 1 day, and then transferred to constant light for another 6 days. Conidial production of each strain was documented as images. (B) Conidial production was measured as number of conidia per slant. Standard deviations from three replicates were marked by error bars.
Some conidiation related genes, including eas, con-6, con-8, con-10, con-13, and NCU09357 (encoding stage V sporulation protein K), were highly expressed in wild type during the late period of conidiation (Table 3 and Figure 2). However, the transcriptional levels of these genes in the Δada-6 mutant were much lower than those in wild type in the mid-late period of conidiation (Table 3 and Figure 2). This result is consistent with the phenotype of reduced conidial production in the Δada-6 mutant. After the 24 h of conidiation induction, the wild type had produced many spores, while the Δada-6 mutant produced only a few spores (data not shown). In consistent with this phenotype, the genes encoding cell wall proteins in vegetative hyphae, including acw-8 and mwg1 (Maddi et al., 2009), were higher expressed in the Δada-6 mutant than those in wild type in the mid-late period of conidiation (Table 3 and Figure 2).
Some genes negatively influence conidial development and their deletion resulted in earlier or enhanced conidial production (Michán et al., 2003; Sun et al., 2011, 2012). Among these genes, cat-3 was differentially expressed between the Δada-6 mutant and wild type (Table 3 and Figure 2). As shown in Figure 2, the expression of cat-3 was not increased during conidiation in wild type. However, the transcriptional level of cat-3 in the Δada-6 mutant was higher than those in wild type during the entire period of conidiation induction.
A large number of N. crassa genes have been identified to be required for sexual development. Among them, only five genes (app, poi-2, pre-2, fbm-1, and NCU05832) were found differentially expressed between the Δada-6 mutant and wild type during sexual development according to RNA-seq data (Table 4). NCU05832 encodes a methyltransferase, whose homologue in A. fumigatus negatively regulates sexual sporulation, and its deletion resulted in formation of a cellular spore4. The transcription of NCU05832 was not induced in wild type, but induced in the Δada-6 mutant during sexual development. After 4 days of sexual development induction, the transcriptional level of NCU05832 in the Δada-6 mutant was 142% higher than that of wild type (Table 4). This result indicates that NCU05832 is negatively regulated by ADA-6.
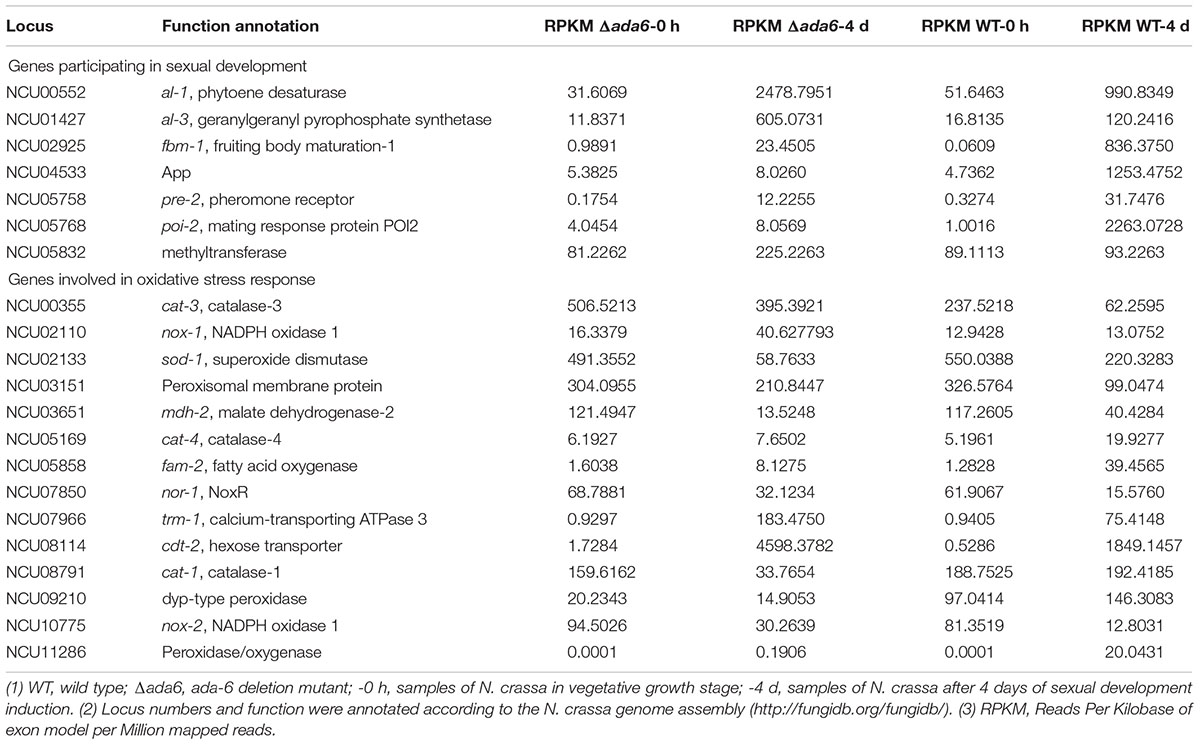
Table 4. Transcriptional response to ada-6 deletion by the genes involved in sexual development and oxidative stress response.
The app (abundant perithecial protein) is an indicator of sexual development, and its transcripts occur only after the onset of sexual development (Nowrousian et al., 2007). pre-2 (NCU005758) is a pheromone receptor encoding gene, and plays a vital role during mating in N. crassa (Kim and Borkovich, 2006). poi-2 is essential for differentiation of female reproduction structures and perithecial development (Kim and Nelson, 2005). fbm-1 encodes fruiting body maturation-1, whose homologue in N. discreta is Kynurenine 3-monooxygenase and related to flavoprotein monooxygenases5. The transcripts of app, poi-2, pre-2, and fbm-1 increased during sexual development, but were obviously lower in the Δada-6 mutant than those in wild type. After 4 days of sexual development induction, transcriptional level of app, poi-2, pre-2, and fbm-1 in the Δada-6 was only 0.6, 0.4, 38.5, and 2.8%, respectively, of that in wild type (Table 4). This result suggests that ADA-6 positively regulates the transcription of pre-2 and poi-2, which promote sexual development. The lower expression of app and fbm-1 in the Δada-6 mutant might be the consequence of female sterility of the Δada-6 mutant.
During both conidiation and sexual development, RNA-seq data suggest that ADA-6 regulate the transcription of cat-3 and genes participating in ROS production. If it is true, deletion of ada-6 might cause an alteration in the sensitivity to oxidants. To confirm this, we inoculated knockout mutants (ada-6, nox-1, cat-2, and cat-3) and wild type on plates with or without oxidants (10 mM H2O2 or 25 μg/ml menadione), and the relative growth inhibition rates of each strain were calculated after 22 h of incubation. The Δcat-3 strain displayed hypersensitivity phenotype to H2O2 (Figure 4A): the relative growth inhibition of the Δcat-3 strain (87%) is higher than that of wild type (70%) (Figure 4B), while, the sensitivity of the Δcat-3 mutant to menadione was similar to that of wild type (Figure 4). This result is consistent with previous report (Michán et al., 2003). The sensitivity of the Δcat-2 mutant to both H2O2 and menadione was similar to wild type, and the Δnox-1 mutant showed slight hypersensitive to menadione and similar sensitive to H2O2 with wild type (Figure 4). The Δada-6 mutant was more sensitive than wild type to both H2O2 and menadione (Figure 4A). On the plates with 10 mM H2O2, the growth of the Δada-6 mutant and wild type was inhibited by 75 and 70%, respectively. On the plates with 25 μg/ml menadione, the growth of the Δada-6 mutant and wild type was inhibited by 63 and 58%, respectively. Above results, together with RNA-seq data suggest that ADA-6 might play a role in oxidative stress response.
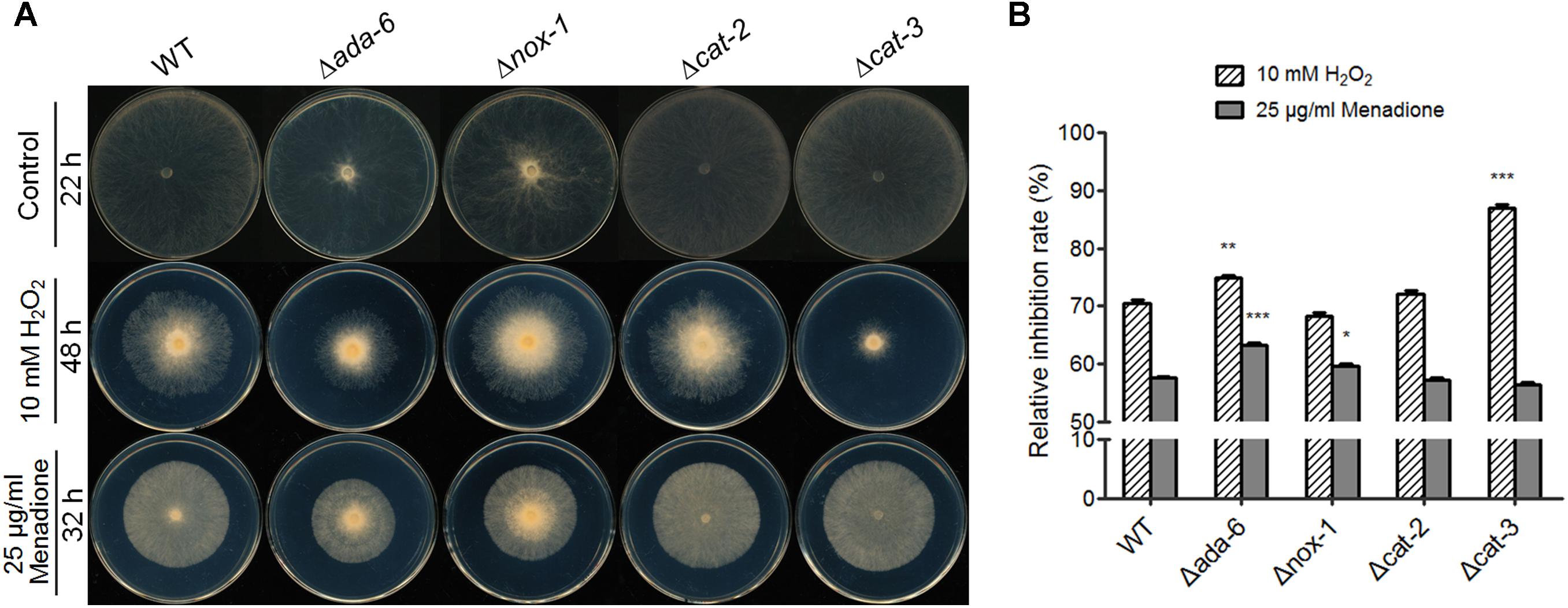
Figure 4. Susceptibility test of N. crassa wild type and knockout mutants (ada-6, nox-1, cat-2, and cat-3) to oxidants. (A) Susceptibility test of the strains to oxidants. Wild type and the mutants were inoculated in ϕ90-mm plates (containing 15-ml liquid Vogel’s medium) and allowed to grow at 28°C in darkness for 24 h. The mycelial mat were punched and the round mat with ϕ4-mm were inoculated in the center of plates (ϕ-90 mm) with or without oxidant (H2O2 or menadione), and incubated at 28°C for indicated time, respectively. Each test was duplicated and the experiment was independently repeated at least three times. (B) Relative growth inhibition rates were calculated based on colony diameters after 22 h of incubation. Values from three replicates were used for a statistical analysis. Means of the inhibition rates are shown, and standard deviations are marked with error bars. Differences between the mutants and the WT were statistically analyzed by the analysis of variance. Values with P < 0.001, 0.001 < P < 0.01, and 0.01 < P < 0.05 are marked with ∗∗∗, ∗∗, and ∗, respectively.
Global transcription factors control fungal growth, development and stress responses by regulating gene transcription on a global scale. Several transcription factors encoding genes named as all development altered (ada) were previously identified, and their deletion resulted in significant defects in basal hyphal growth, asexual sporulation, and sexual development (Colot et al., 2006; Carrillo et al., 2017). One of them is ada-6 (NCU04866), whose deletion resulted in slower growth, dramatic reduction in conidiation and female infertility (Colot et al., 2006). However, the regulatory role of ADA-6 in growth and development needs further confirmation by mutant complementation, and its mechanism has not been addressed. Here we first confirmed the positive role of ADA-6 in growth and development by detailed phenotypic characterization of its deletion mutant and the complemented mutant. Then, by comparing transcriptomic profiles and functional analysis of genes influenced by ada-6 deletion, we explored the mechanisms by which ADA-6 promotes conidiation and sexual development. Our results demonstrate that ADA-6 might play a role in conidiation by regulating oxidation-reduction process and single-organism metabolic process. Moreover, ADA-6 positively regulates the transcription of fluffy and acon-3, two key genes required for the initiation of asexual sporulation by controlling the formation of major constriction chains (Matsuyama et al., 1974; Springer and Yanofsky, 1989; Bailey and Ebbole, 1998; Bailey-Shrode and Ebbole, 2004; Chung et al., 2011). Deletion of ada-6 also resulted in female sterility. ADA-6 regulates some genes associated with sexual development, including pre-2, poi-2, and NCU05832, three key genes required for sexual development (Kim and Nelson, 2005; Kim and Borkovich, 2006).
Oxidation-reduction reaction is involved in many pathways. The foremost of these is oxidative stress response pathway, which affects fungal growth, development and stress responses. ADA-6 regulates the transcription of cat-3 and genes participating in ROS production according to RNA-seq data, indicating a role of ADA-6 in oxidative stress response. This was confirmed by the hypersensitivity phenotype of the Δada-6 mutant to oxidants H2O2 and menadione.
Numerous studies have found the role of ROS in the regulation of conidiation and sexual development (Michán et al., 2003; Aguirre et al., 2005; Cano-Domínguez et al., 2008). In N. crassa, formation of conidia from growing hyphae includes three morphogenetic developmental stages: growing hyphae to adherent mycelium, adherent mycelium to aerial hyphae, and aerial hyphae to conidia. A hyperoxidant state develops at the start of these morphogenetic transitions (Hansberg et al., 1993; Toledo et al., 1995). Oxidative stress due to lack of CAT-3 induces hyphal adhesion, and development of more aerial hyphae and conidia (Michán et al., 2003). NOX-1, NOX-2, and NOX-R participates in ROS production, and NOX-1 elimination results in complete female sterility, decreased asexual development, and reduction of hyphal growth in N. crassa (Cano-Domínguez et al., 2008). All these studies indicate that ROS, whose accumulation is induced by eliminating ROS-decomposion (CAT-3) or activating ROS-generation (NOX-1), is a critical cell differentiation signal promoting conidiation and sexual development. In our study, ADA-6 regulates the transcription of oxidative stress related genes, including nox-1, cat-3, etc., during both conidiation and sexual development. Based on above result, we speculate that more ROS may accumulate in cells of the Δada-6 mutant during development or stress response process. This was further confirmed by the results that deletion of ada-6 resulted in hypersensitive to oxidative stress inducers H2O2 and menadione. Based on previous studies, this ROS accumulation can promote conidiation and sexual development in N. crassa (Hansberg et al., 1993; Toledo et al., 1995; Cano-Domínguez et al., 2008). However, the Δada-6 mutant still exhibited reduced conidial production and female sterility. These results indicate that regulation of development and oxidative stress response by ADA-6 may be independent.
In summary, our study showed that ADA-6, as a global regulator, plays positive roles in conidiation and sexual development, and regulates oxidative stress response of N. crassa. Combining transcriptomic profiles and functional assay, we explored the mechanisms by which ADA-6 promotes conidiation and sexual development, and regulates oxidative stress response. This work has augmented our knowledge of the functions and mechanisms of global regulators that influence growth, development and stress responses in filamentous fungi.
XS designed the study and wrote the manuscript. XS, FW, NL, and CH performed the main experiments. SL, BL, WX, and ZZ contributed to the data analysis and the data interpretation.
This project was supported by grants 31370024 (to XS) from National Natural Science Foundation of China.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00750/full#supplementary-material
Aguirre, J., Rios-Momberg, M., Hewitt, D., and Hansberg, W. (2005). Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 13, 111–118. doi: 10.1016/j.tim.2005.01.007
Bailey, L. A., and Ebbole, D. J. (1998). The fluffy gene of Neurospora crassa encodes a Gal4p-type C6 zinc cluster protein required for conidial development. Genetics 148, 1813–1820.
Bailey-Shrode, L., and Ebbole, D. J. (2004). The fluffy gene of Neurospora crassa is necessary and sufficient to induce conidiophore development. Genetics 166, 1741–1749. doi: 10.1534/genetics.166.4.1741
Borkovich, K. A., Alex, L. A., Yarden, O., Freitag, M., Turner, G. E., Read, N. D., et al. (2004). Lessons from the genome sequence of Neurospora crassa: tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 68, 1–108. doi: 10.1128/MMBR.68.1.1-108.2004
Cano-Domínguez, N., Alvarez-Delfin, K., Hansberg, W., and Aguirre, J. (2008). NADPH oxidases NOX-1 and NOX-2 require the regulatory subunit NOR-1 to control cell differentiation and growth in Neurospora crassa. Eukaryot.Cell 7, 1352–1361. doi: 10.1128/EC.00137-08
Carrillo, A. J., Schacht, P., Cabrera, I. E., Blahut, J., Prudhomme, L., Dietrich, S., et al. (2017). Functional profiling of transcription factor genes in Neurospora crassa. G3 7, 2945–2956. doi: 10.1534/g3.117.043331
Chary, P., and Natvig, D. O. (1989). Evidence for three differentially regulated catalase genes in Neurospora crassa: effects of oxidative stress, heat shock, and development. J. Bacteriol. 171, 2646–2652. doi: 10.1128/jb.171.5.2646-2652.1989
Chen, H., Crabb, J. W., and Kinsey, J. A. (1998). The neurospora aab-1 gene encodes a CCAAT binding protein homologous to yeast HAP5. Genetics 148, 123–130.
Chung, D. W., Greenwald, C., Upadhyay, S., Ding, S., Wilkinson, H. H., Ebbole, D. J., et al. (2011). acon-3, the Neurospora crassa ortholog of the developmental modifier, medA, complements the conidiation defect of the Aspergillus nidulans mutant. Fungal Genet. Biol. 48, 370–376. doi: 10.1016/j.fgb.2010.12.008
Circu, M. L., and Aw, T. Y. (2010). Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 48, 749–762. doi: 10.1016/j.freeradbiomed.2009.12.022
Colot, H. V., Park, G., Turner, G. E., Ringelberg, C., Crew, C. M., Litvinkova, L., et al. (2006). A high-throughput gene knockout procedure for neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. U.S.A. 103, 10352–10357. doi: 10.1073/pnas.0601456103
Davis, R. H., and de Serres, F. J. (1970). Genetic and microbiological research techniques for Neurospora crassa. Method Enzymol. 17, 79–143. doi: 10.1016/0076-6879(71)17168-6
Davis, R. H., and Perkins, D. D. (2002). Neurospora: a model of model microbes. Nat. Rev. Genet. 3, 397–403. doi: 10.1038/nrg797
Ebbole, D. J., and Sachs, M. S. (1990). A rapid and simple method for isolation of Neurospora crassa homokaryons using microconidia. Fungal Genet. Newslett. 37, 17–18. doi: 10.4148/1941-4765.1472
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1038/nbt.1883
Greenwald, C. J., Kasuga, T., Glass, N. L., Shaw, B. D., Ebbole, D. J., and Wilkinson, H. H. (2010). Temporal and spatial regulation of gene expression during asexual development of Neurospora crassa. Genetics 186, 1217–1230. doi: 10.1534/genetics.110.121780
Hansberg, W., de Groot, H., and Sies, H. (1993). Reactive oxygen species associated with cell differentiation in Neurospora crassa. Free Radic. Biol. Med. 14, 287–293. doi: 10.1016/0891-5849(93)90025-P
Heitman, J., Sun, S., and James, T. Y. (2013). Evolution of fungal sexual reproduction. Mycologia 105, 1–27. doi: 10.3852/12-253
Kim, H., and Borkovich, K. A. (2006). Pheromones are essential for male fertility and sufficient to direct chemotropic polarized growth of trichogynes during mating in Neurospora crassa. Eukaryot. Cell 5, 544–554. doi: 10.1128/EC.5.3.544-554.2006
Kim, H., and Nelson, M. A. (2005). Molecular and functional analyses of poi-2, a novel gene highly expressed in sexual and perithecial tissues of Neurospora crassa. Eukaryot.Cell 4, 900–910. doi: 10.1128/EC.4.5.900-910.2005
Lehr, N. A., Wang, Z., Li, N., Hewitt, D. A., López-Giráldez, F., Trail, F., et al. (2014). Gene expression differences among three Neurospora species reveal genes required for sexual reproduction in Neurospora crassa. PLoS One 9:e110398. doi: 10.1371/journal.pone.0110398
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using Real-time quantitative PCR and the 2-ΔΔCt method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Maddi, A., Bowman, S. M., and Free, S. J. (2009). Trifluoromethanesulfonic acid-based proteomic analysis of cell wall and secreted proteins of the ascomycetous fungi Neurospora crassa and Candida albicans. Fungal Genet. Biol. 46, 768–781. doi: 10.1016/j.fgb.2009.06.005
Matsuo, Y., and Yodoi, J. (2013). Extracellular thioredoxin: a therapeutic tool to combat inflammation. Cytokine Growth Factor Rev. 24, 345–353. doi: 10.1016/j.cytogfr.2013.01.001
Matsuyama, S. S., Nelson, R. E., and Siegel, R. W. (1974). Mutations specifically blocking differentiation of macroconidia in Neurospora crassa. Dev. Biol. 41, 278–287. doi: 10.1016/0012-1606(74)90306-6
Michán, S., Lledías, F., and Hansberg, W. (2003). Asexual development is increased in Neurospora crassa cat-3-null mutant strains. Eukaryot. Cell 2, 798–808. doi: 10.1128/EC.2.4.798-808.2003
Ni, M., Feretzaki, M., Sun, S., Wang, X., and Heitman, J. (2011). Sex in fungi. Annu. Rev. Genet. 45, 405–430. doi: 10.1146/annurev-genet-110410-132536
Nowrousian, M., Piotrowski, M., and Kück, U. (2007). Multiple layers of temporal and spatial control regulate accumulation of the fruiting body-specific protein APP in Sordaria macrospora and Neurospora crassa. Fungal Genet. Biol. 44, 602–614. doi: 10.1016/j.fgb.2006.09.009
Peraza, L., and Hansberg, W. (2002). Neurospora crassa catalases, singlet oxygen and cell differentiation. Biol. Chem. 383, 569–575. doi: 10.1515/BC.2002.058
Perkins, D. D., and Davis, R. H. (2000). Neurospora at the millennium. Fungal Genet. Biol. 31, 153–167. doi: 10.1006/fgbi.2000.1248
Royer, J. C., and Yamashiro, C. T. (1992). Generation of transformable spheroplasts from mycelia, macroconidia, microconidia and germinating ascospores of Neurospora crassa. Fungal Genet. Newslett. 39, 76–79. doi: 10.4148/1941-4765.1440
Schliebs, W., Würtz, C., Kunau, W.-H., Veenhuis, M., and Rottensteiner, H. (2006). A eukaryote without catalase-containing microbodies: Neurospora crassa exhibits a unique cellular distribution of its four catalases. Eukaryot. Cell 5, 1490–1502. doi: 10.1128/EC.00113-06
Springer, M. L. (1993). Genetic control of fungal differentiation: the three sporulation pathways of Neurospora crassa. BioEssays 15, 365–374. doi: 10.1002/bies.950150602
Springer, M. L., and Yanofsky, C. (1989). A morphological and genetic analysis of conidiophore development in Neurospora crassa. Gene. Dev. 3, 559–571. doi: 10.1101/gad.3.4.559
Sun, X., Yu, L., Lan, N., Wei, S., Yu, Y., Zhang, H., et al. (2012). Analysis of the role of transcription factor VAD-5 in conidiation of Neurospora crassa. Fungal Genet. Biol. 49, 379–387. doi: 10.1016/j.fgb.2012.03.003
Sun, X., Zhang, H., Zhang, Z., Wang, Y., and Li, S. (2011). Involvement of a helix-loop-helix transcription factor CHC-1 in CO2-mediated conidiation suppression in Neurospora crassa. Fungal Genet. Biol. 48, 1077–1086. doi: 10.1016/j.fgb.2011.09.003
Sweigard, J., Chumley, F. G., Carroll, A., Farrall, L., and Valent, B. (1997). A series of vectors for fungal transformation. Fungal Genet. Newslett. 44, 52–53. doi: 10.4148/1941-4765.1287
Thompson, S., Croft, N. J., Sotiriou, A., Piggins, H. D., and Crosthwaite, S. K. (2008). Neurospora crassa heat shock factor 1 is an essential gene; a second heat shock factor-like gene, hsf2, is required for asexual spore formation. Eukaryot.Cell 7, 1573–1581. doi: 10.1128/EC.00427-07
Toledo, I., Rangel, P., and Hansberg, W. (1995). Redox imbalance at the start of each morphogenetic step of Neurospora crassa conidiation. Arch. Biochem. Biophys. 319, 519–524. doi: 10.1006/abbi.1995.1326
Wang, N., Yoshida, Y., and Hasunuma, K. (2007). Loss of catalase-1 (Cat-1) results in decreased conidial viability enhanced by exposure to light in Neurospora crassa. Mol. Genet. Genomics 277, 13–22. doi: 10.1007/s00438-006-0170-4
Keywords: conidiation, ada-6, sexual development, oxidative stress response, Neurospora crassa
Citation: Sun X, Wang F, Lan N, Liu B, Hu C, Xue W, Zhang Z and Li S (2019) The Zn(II)2Cys6-Type Transcription Factor ADA-6 Regulates Conidiation, Sexual Development, and Oxidative Stress Response in Neurospora crassa. Front. Microbiol. 10:750. doi: 10.3389/fmicb.2019.00750
Received: 06 December 2018; Accepted: 26 March 2019;
Published: 10 April 2019.
Edited by:
Neil Andrew Brown, University of Bath, United KingdomReviewed by:
Kap-Hoon Han, Woosuk University, South KoreaCopyright © 2019 Sun, Wang, Lan, Liu, Hu, Xue, Zhang and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xianyun Sun, c3VueHlAaW0uYWMuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.