
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 28 February 2019
Sec. Infectious Agents and Disease
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.00305
 Pollyanna Stephanie Gomes1
Pollyanna Stephanie Gomes1 Scott Tanghe2
Scott Tanghe2 Julio Gallego-Delgado2
Julio Gallego-Delgado2 Luciana Conde1
Luciana Conde1 Leonardo Freire-de-Lima3Ana Carolina Lima4
Leonardo Freire-de-Lima3Ana Carolina Lima4 Célio Geraldo Freire-de-Lima3
Célio Geraldo Freire-de-Lima3 Josué da Costa Lima Junior4
Josué da Costa Lima Junior4 Otacílio Moreira4
Otacílio Moreira4 Paulo Totino4
Paulo Totino4 Ana Rodriguez2*
Ana Rodriguez2* Adriane Regina Todeschini5*
Adriane Regina Todeschini5* Alexandre Morrot1,6*
Alexandre Morrot1,6*Cerebral malaria (CM) is a clinical syndrome involving irreversible and lethal signs of brain injury associated to infection by parasites of the genus Plasmodium. The pathogenesis of CM derives from infection-induced proinflammatory cytokines associated with cytoadherence of parasitized red blood cells to brain microvasculature. Glycoconjugates are very abundant in the surface of Plasmodium spp., and are critical mediators of parasite virulence in host–pathogen interactions. Herein, we show that 6-Diazo-5-oxo-L-norleucine (DON) therapeutically used for blocking hexosamine biosynthetic pathway leads to recovery in experimental murine cerebral malaria. DON-induced protection was associated with decreased parasitism, which severely reduced Plasmodium transmission to mosquitoes. These findings point to a potential use of DON in combination therapies against malaria.
Malaria is a parasitic disease caused by the protozoan Plasmodium. The more severe clinical manifestation of the disease affects the microvasculature of the blood–brain barrier resulting in the cerebral form of disease (Polimeni and Prato, 2014). Cerebral malaria is associated with infection by Plasmodium falciparum, which affects over half a million individuals annually (World Health Organization, 2015), most of them being represented by children in sub-Saharan Africa (Idro et al., 2010). The use of animal models of infection has allowed us to accumulate knowledge about the pathophysiology of disease. Experimental murine models using C57BL/6 mice infected with Plasmodium berghei ANKA (PbA) are able to reproduce the clinical signs of human cerebral malaria (de Oca et al., 2013). Different studies have consistently shown the requirement for T lymphocytes in the pathogenesis of cerebral malaria (Renia et al., 2006) by demonstrating that infection in mice deficient in beta-2-microglobulin gene as well as mice lacking functional CD8+ T cells are resistant to experimental cerebral malaria (Belnoue et al., 2002).
Additional studies have shown that the trafficking of CD8+ T lymphocytes to the brain is a critical factor involved in the pathogenesis of experimental malaria after PbA infection (Howland et al., 2015a). Antigen-specific CD8+ T cells are able to recognize parasite antigens cross-presented via MHC class I by brain endothelium that is subject to cellular interaction with infected red blood cells (Howland et al., 2015b). The triggering of perforin-mediated endothelial cell dysfunction by activated effector CD8+ T cells is a determinant for disease pathogenesis (Jordan and Hunter, 2010). In vivo administration of 6-diazo-5-oxo-L-norleucine (DON) late in the course of infection, when the mice have already triggered endothelial dysfunction due to the action of CD8+ T cell responses, was able to rescue the clinical manifestation signs of cerebral malaria. These studies have suggested that DON treatment has a protective effect by decreasing function of activated effector CD8+ T cells (Gordon et al., 2015).
The inhibitory effect of DON in blocking the pathogenic role of CD8+T cells may account to its role in the rescue of mice with experimental cerebral malaria (Howland et al., 2015a). DON is a glutamine antagonist used as inhibitor of different glutamine-utilizing enzymes such as glutaminase, aminotransferases, and glutamine synthetase (Pinkus, 1977). Among those enzymes is the glutamine-fructose-6-phosphate transaminase (GFPT) (Ginsburg, 2006), the rate-limiting enzyme of the hexosamine biosynthetic pathway which transfers amino group from glutamine to the fructose-6-phosphate to form glucosamine-6-phosphate (GlcN6P). GFPT participates in the synthesis of uridine diphosphate N-acetylglucosamine (UDP-GlcNAc), used for biosynthesis of glycoproteins, glycosaminoglycans, proteoglycans, and glycolipids (Vasconcelos-Dos-Santos et al., 2015).
Hexosamine biosynthetic pathway is predicted to be essential in P. falciparum as it feeds the biosynthesis of glycosylphosphatidylinositol (GPI) anchors which are required for parasite survival and infectivity, thus contributing to malaria pathogenesis (Krishnegowda et al., 2005). Furthermore, GlcNAc can also be incorporated to short N-glycans composed of one or two residues of the sugar (Bushkin et al., 2010; Macedo et al., 2010; Samuelson and Robbins, 2015). N-linked glycosylation is essential for the parasite given that the N-glycosylation blocker tunicamycin arrested parasite development (Dieckmann-Schuppert et al., 1992). Those inhibitory effects of DON on the hexosamine biosynthetic pathway account for its antiparasitic activity of both in vitro and in vivo (Queen et al., 1990; Waknine-Grinberg et al., 2010). In the present study we investigated the importance of GFPT as a potential malarial transmission-blocking target required for successful development cycle of P. berghei ANKA within the vertebrate host.
Protocols for animal experimentation ere used in accordance with the guidelines for the animal welfare regulations set by the National Institutes of Health, United States. The study was approved by the Research Ethics Committee of Federal University of Rio de Janeiro (Protocol No. IMPPG040-07/16). Protocols for animal were approved by the Institutional Ethical Committees in accordance with international guidelines. All animal experimentation was performed in accordance with the terms of the Brazilian guidelines for animal welfare regulations.
C57Bl/6 mice and Swiss Webster (6–8 week-old females) were obtained from The Jackson Laboratories. C57Bl/6 mice were intraperitoneally infected (i.p.) with P. berghei ANKA expressing GFP (P. berghei ANKA) by injection of 1 × 106 PbA-infected red blood cells (PbARBC) in cerebral malaria experiments. This infection protocol was used for Swiss Webster mice were in transmission-blocking experiments upon infection with Plasmodium yoelli -luciferase. The parasitemia was determined by GIEMSA staining of peripheral blood smears during the course of infection. Infected mice were monitored for progression of CM signs using a 5-point clinical scoring system that rates mice from a score of 0 (no signs) to 5 (moribund) based on their appearance and behavior, as previously described (Plaimas et al., 2013). Mice weighing ∼20 g were intraperitoneally injected with DON (1.3 mg/kg), and/or GlcN (40 mg/kg) in 100 μL PBS, beginning the treatment at the first clinical symptoms of CM until day 11. In transmission-blocking experiments, Swiss Webster mice infected with 103 of luciferase-expressing P. yoelli were daily treated with doses of DON (0.5 mg/kg) or vehicle (saline) intraperitoneally administered, starting on day 4 after the infection.
Groups of five mice were infected with 103 blood-stage P. yoelli-luciferase by intraperitoneal injection. On day 7 post-infection, mice were anesthetized by isoflurane inhalation and injected with 150 mg/kg D-luciferin potassium salt substrate for bioimaging using IVIS imager (Lumina II in vivo Imaging System; Perkin-Elmer). Light intensity was measured in each mouse to determine the baseline of infection levels before treatment. Mice were then treated with DON administered daily by intraperitonial injection (0.5 mg/kg) and saline as vehicle (from days 4 to 7 post-infection). A group of negative-control mice were treated with vehicle alone. On day 7 post-infection, mice were imaged to determine the parasite burden.
Groups of five Swiss Webster mice were infected with 103 P. yoelli-luciferase and both gametocytemia and parasitemia were detected at 4 days post-infection by blood smear (five mice per group in three independent experiments). Mice were then alternatively treated with DON administered daily by intraperitoneal injection (0.5 mg/kg). The vehicle control group was obtained by injecting saline only. At day 4 post-infection, mice were anesthetized with ketamine (3,500 mg/kg), xylazine (300 mg/kg) and groups of 100 Anopheles stephensi mosquitoes were allowed to feed on each group for 20 min with feeding disruption once every 5 min. The infected mosquitoes were then incubated at 18°C for 11 days to allow oocyte formation. Afterward, the mosquitoes were dissected and their midguts removed, and homogenized for parasite detection after incubation with D-Luciferin potassium salt (200 μg/ml). Luminescence was measured by using a microplateplate reader (PerkinElmer).
Total RNA was extracted from 1 mL blood using RNeasy®mini kit (QIAGEN). Samples were collected from infected and uninfected animals, and immediately mixed with 350 μL of lysis buffer (RLT buffer). RNA was extracted according to manufacturer’s protocol and resuspended in 30 μL of elution buffer. RNA was stored at -80°C until use. RNA samples were treated with DNAse I (Sigma-Aldrich), and the quantification was performed using a ND2000 Nanodrop (Thermo Fisher Scientific). The reverse transcription was performed using the Superscript IV ViloMastermix (Invitrogen), following manufacture’s instructions.
The comparative quantification of P. berghei gametocyte forms between treated (1,3 mg/kg/day of DON) versus control mice was performed by PbCCp3 mRNA detection in blood samples. In parallel, the PbHSP70 mRNA levels were also quantified to detect all the parasite evolutive forms (Lavazec et al., 2009). For real-time PCR assays, 2 μl cDNA were used in a final reaction volume of 20 μL, with 10 μL of Power SYBR Green PCR Master Mix 2X (Life Technologies, Foster City, CA, United States), 300 nM of each PbCCp3 (sense 5′-CTGCAGCTATTTATGATGGT, antisense 5′-TCATCACTTTCATCACCTTT) or PHSP70 (sense 51-AGAGAAGCAGCTGAAACAGC,antisense 5′-TCCCTTTAATAAATCATGGC) primers. PCR cycling conditions were used with first step at 95°C for 10 min, followed by 40 cycles at 95°C for 15 s and 60°C for 1 min. To check the primers specificity, melting curves were generated after the 40 cycles. To obtain the Threshold Cycle (Ct), threshold was set at 0.02. The comparative quantification was expressed as 2-ΔCt, where ΔCt = CtTreatedmouse – CtUntreatedmouse. RT-qPCR assays were performed in a ViiA7 Real Time PCR System (Applied Biosystems), in technical duplicates and biological triplicates.
The erythrocytic asexual stage of P. falciparum (3D7 strain) was cultured in 96 well plates for 96 h, in RPMI media containing 25 mM HEPES, 25 mM sodium bicarbonate, 10 μg/ml gentamycin, 0.5 mM hypoxanthine, at pH 6.75 in atmospheric conditions (5% CO2, 5% O2, and 90% N2). Alternative cultures were done in the presence of DON (50 μM). To set up the culture the parasites were synchronized using MACS cell separation column (MiltenyiBiotec) and the parasitemia was maintained below 5%. The parasite load was determined by GIEMSA staining of peripheral blood smear.
The effect of DON on parasite viability as well as development in P. falciparum cultures was examined by flow cytometry using rhodamine 123 (Invitrogen) as indicator of mitochondrial activity, as previously described (Totino et al., 2008). Briefly, cultures were washed in RPMI medium (RPMI-1640, 25 mM Hepes, 0.2% glucose, 23 mM sodium bicarbonate) and then incubated at 37°C for 5 min with 1 μg/ml rhodamine in RPMI. Rhodamine solution was removed by centrifugation and cultures were washed once before incubation for 30 min in complete medium alone. Afterward, cultures were ressuspended in fresh complete medium and analyzed in a flow cytometer (FACSVerse, Becton Dickinson) using 488-nm blue laser and 527/32 bandpass filter. Non-parasitized RBC samples stained with rodamine were used as negative control in flow cytometry analysis.
Statistical analyses were performed with GraphPad Prism 5 software. Statistical differences between mean values were evaluated by non-parametric Student’s t-test or analyzed with ANOVA and Bonferroni post hoc test. Results were expressed as mean ± standard deviation (SD), and differences between control and treated group were considered statistically significant when p ≤ 0.05.
We first analyzed the rates of parasite growth in cultures of P. falciparum-infected erythrocytes in the presence or absence of DON. Analysis of in vitro cultures of P. falciparum in erythrocyte cells showed a significant lower growth rate of the parasites in the presence of DON (IC50 = 1.6 μM) as compared to controls of erythrocytes supplemented with medium only (Figure 1A). Since DON is not a specific inhibitor of GFPT, as it inhibits other amidotransferases, we tested whether the addition of glucosamine (GlcN) could restore parasite growth by bypassing GFPT inhibition by DON. GlcN is easily transported by the glucose transporter systems, and then phosphorylated by hexokinase to glucosamine 6-phosphate. Glucosamine 6-phosphate directly enters the hexosamine biosynthetic pathway bypassing the key regulatory enzyme GFPT, resulting in an increase in UDP-GlcNAc. Indeed, the inhibition of GFPT by DON can be compensated by the exogenous addition of GlcN (Figure 1B). Our findings indicate that the inhibitory effect on the parasite growth in red blood cells cultured in presence of DON was reverted upon exogenous addition of glucosamine in infected erythrocytes, therefore indicating a requirement for the UDP-GlcNA chexosamine downstream product of GFPT in the parasite blood-stage cycle.
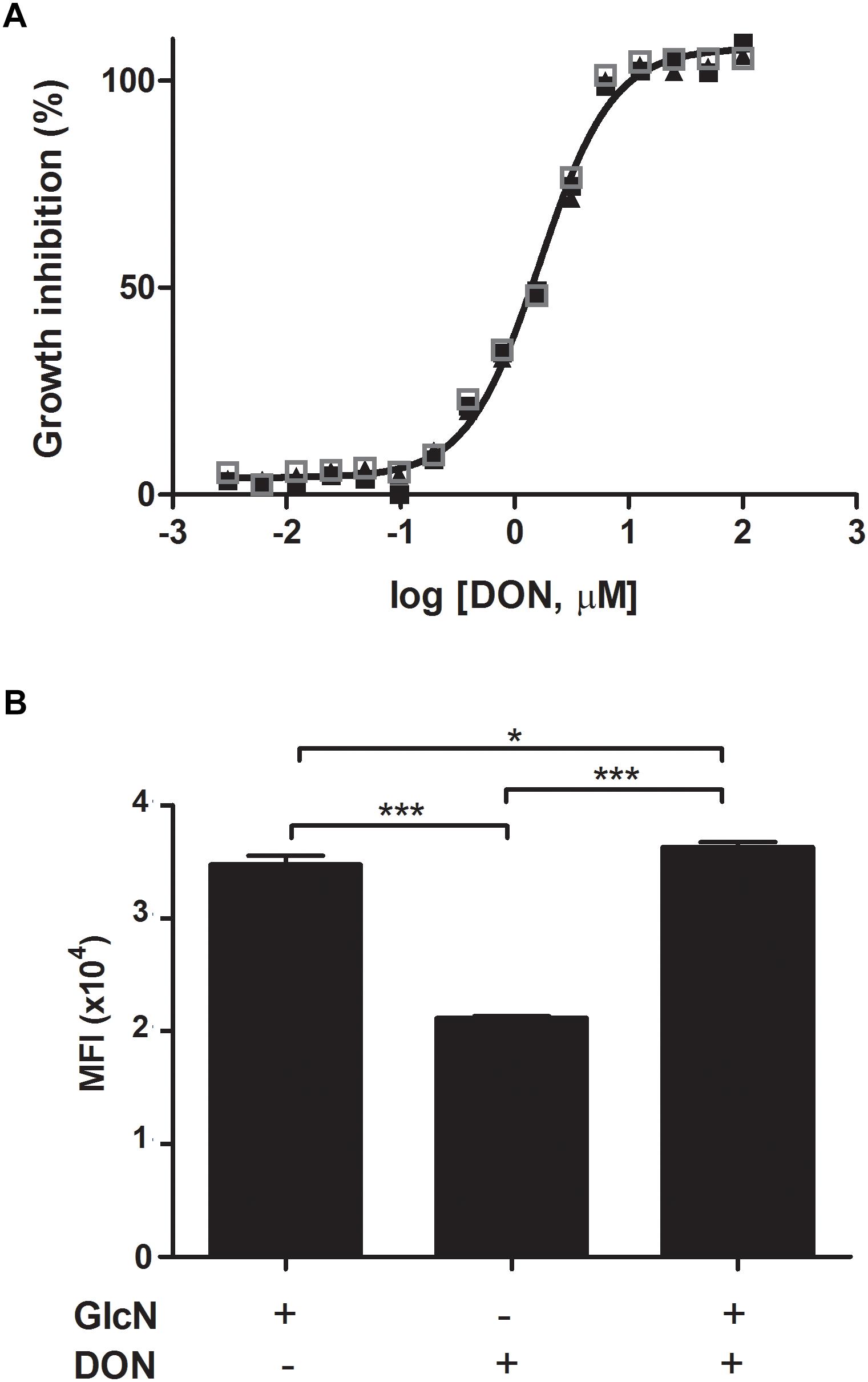
Figure 1. The hexosamine biosynthetic pathway is required for the growth of erythrocytic forms of Plasmodium parasites. (A) Plasmodium falciparum 3D7 strain parasites were incubated with fresh human erythrocytes in the absence or presence of increased doses of DON. Parasitemia was determined 4 days later. The IC50 value of DON (1.6 μM) was determined as the concentration that inhibits parasite growth by 50%. The concentration-dependent inhibitory dose-curve data was plotted as percentage inhibition normalized to normal infected-controls in the absence of inhibitor with applied curve fits calculated using GraphPad Prism. (B) The inhibitory effect of DON on P. falciparum 3D7 strain parasites incubated with erythrocytes was reverted by the addition of exogenous GlcN (125 μM) to the cultures and parasitemia was determined at day 4 pi. The results indicate a requirement of the hexosamine biosynthetic pathway for the parasite blood-stage growth. Data are means ± SD and represent the results of three independent experiments. Differences between groups are significant (∗p < 0.05, ∗∗∗p < 0.001).
To better understand the antimalarial effect of DON on blood-stage forms of Plasmodium parasites, we performed mitochondrial activity assay by flow cytometry using rhodamine 123 (Rho) to evaluate parasite viability as well as stage-specific parasite development (Figure 2A). Rho is incorporated by active mitochondria of living parasites in a proportional manner of the developmental stage, differentially staining early (Rho low RBCs) and late (Rho high RBCs) stages of intraerythrocytic development (Figure 2A, synchronized). Using this approach, we observed no effect of 1.6 μM DON on parasite viability in asynchronous P. falciparum culture treated for 4 h, as the number of Rho-positive RBCs and Rho fluorescence intensity (MFI) were similarly detected in control cultures, while 16 μM DON induced a slight, although statistically significant, decrease of Rho MFI at 4 h (Figure 2B). In contrast, both concentrations of DON (1.6 and 16 μM) notably inhibited the increase of Rho-positive RBC levels after 24 h of culture (Figure 2B), indicating that rather than an immediate cytotoxic effect on parasite viability, DON acts arresting plasmodial development.
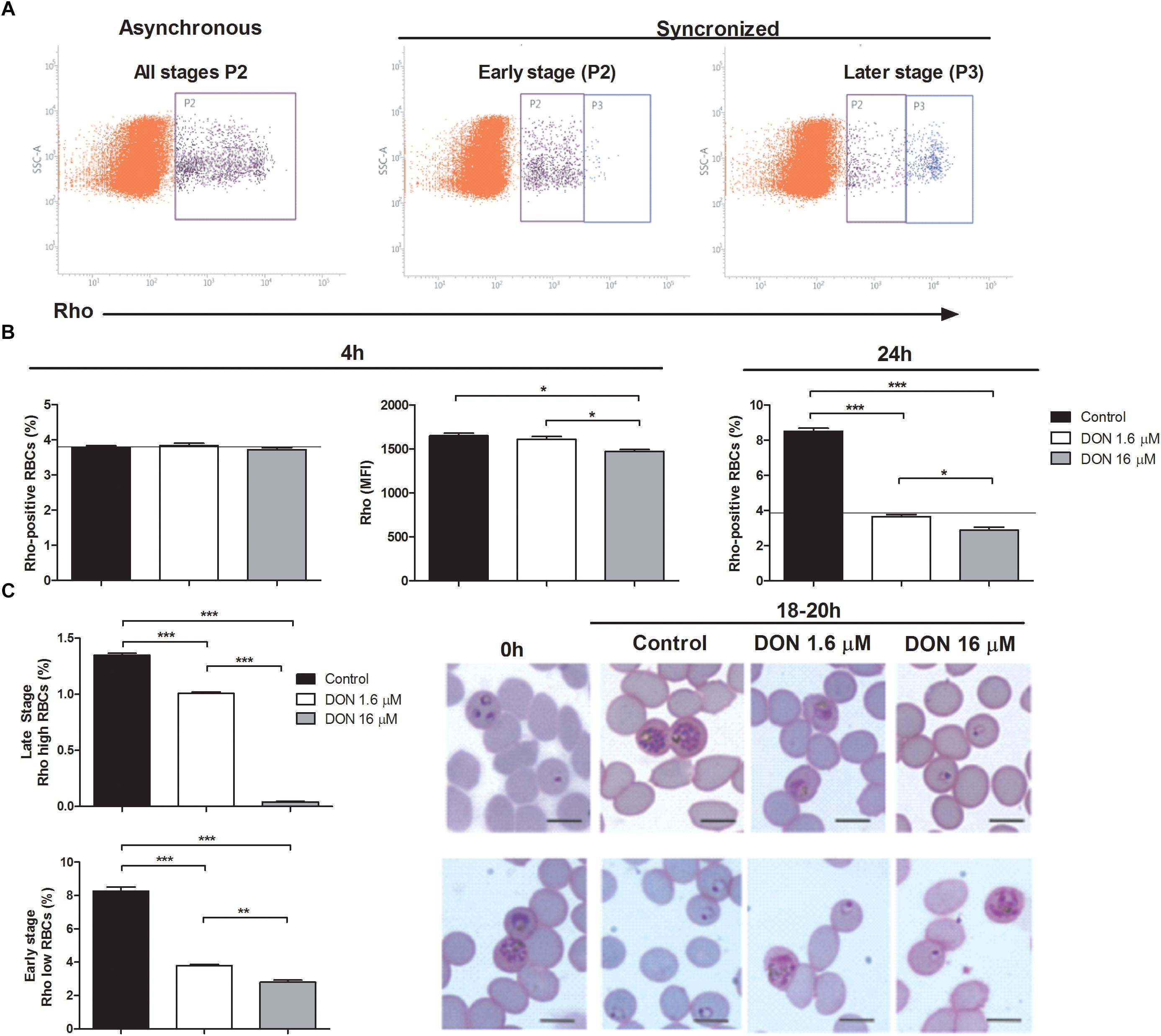
Figure 2. Effect of DON on parasite viability and stage-specific development measured by mitochondrial activity using rhodamine 123 (Rho) staining. (A) Representative flow cytometry analysis of mitochondrial activity in asynchronous (P2: Rho-positive RBCs) and synchronized P. falciparum culture enriched with early (P2: Rho low RBCs) or late (P3: Rho high RBCs) stages. (B) Asynchronous culture was incubated in absence (Control) or presence of DON (1.6 and 16 μM) and, then, parasite viability was evaluated after 4 h by measuring both percentage of Rho-positive RBCs (P2 in A, asynchronous) and Rho fluorescence intensity (MFI) in this RBCs population. Parasite growth was also estimated by determining percentage of Rho-positive RBCs after 24 h. The horizontal black lines represent initial parasitemia. (C) Stage-specific effect of DON was tested using synchronized cultures (0 h) in early (upper panel) or late (lower panel) stages, as shown in (A, synchronized). Cultures were incubated in absence (Control) or presence of DON (1.6 and 16 μM) and parasite development was evaluated after 18–20 h by determining the percentage of late (Rho high RBCs, upper panel) and early stages (Rho low RBCs; lower panel) in flow cytometry analysis. In addition, parasite morphology was examined by Giemsa-stained smears using light microscopy. Data are means ± SD and differences between groups are significant (∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001). Scale bars in (C) indicate 7 μm.
Indeed, when we used early-stage synchronized culture to study parasite development under DON pressure, a decreased number of late stage parasites (Rho high RBCs) was detected in DON-treated cultures, with a remarkable effect of 16 μM DON (Figure 2C, upper panel). Parallel examination of such cultures by light microscopy showed that parasite development was arrested at ring and late trophozoite stages at the 16 and 1.6 μM concentration, respectively. Moreover, a significant reduction of newly invaded early forms was also observed after incubating late-stage cultures in the presence of DON, which induced appearance of late-stage parasites displaying abnormal morphology, as evidenced by microscopy analysis (Figure 2C, lower panel). These results indicates that DON acts arresting stage-specific development of Plasmodium parasites in a dose-dependent manner, while invariably affects late-stage maturation and, consequently, the increase of parasitemia.
To access the inhibitory effect of DON on malaria erythrocytic stage development in the experimental murine model of infection, CB57BL/6 mice treated with DON (1.3 mg/kg) was injected intraperitoneally, administered consecutively every day, during the course of infection with 103 blood-stage P. berghei ANKA parasites. RT-qPCR analysis by targeting PbHSP70 mRNA, which is constitutively expressed in all the P. berghei evolutive forms (Figure 3), confirmed a reduction in the blood-stage parasitemia at day 6 post-infection. In addition, the detection of P. berghei gametocyte forms, by accessing the PbCCp3 mRNA levels, indicated significantly lower levels of transcripts (59,96 times) from DON-treated group as compared to control (vehicle) group (Figure 3).
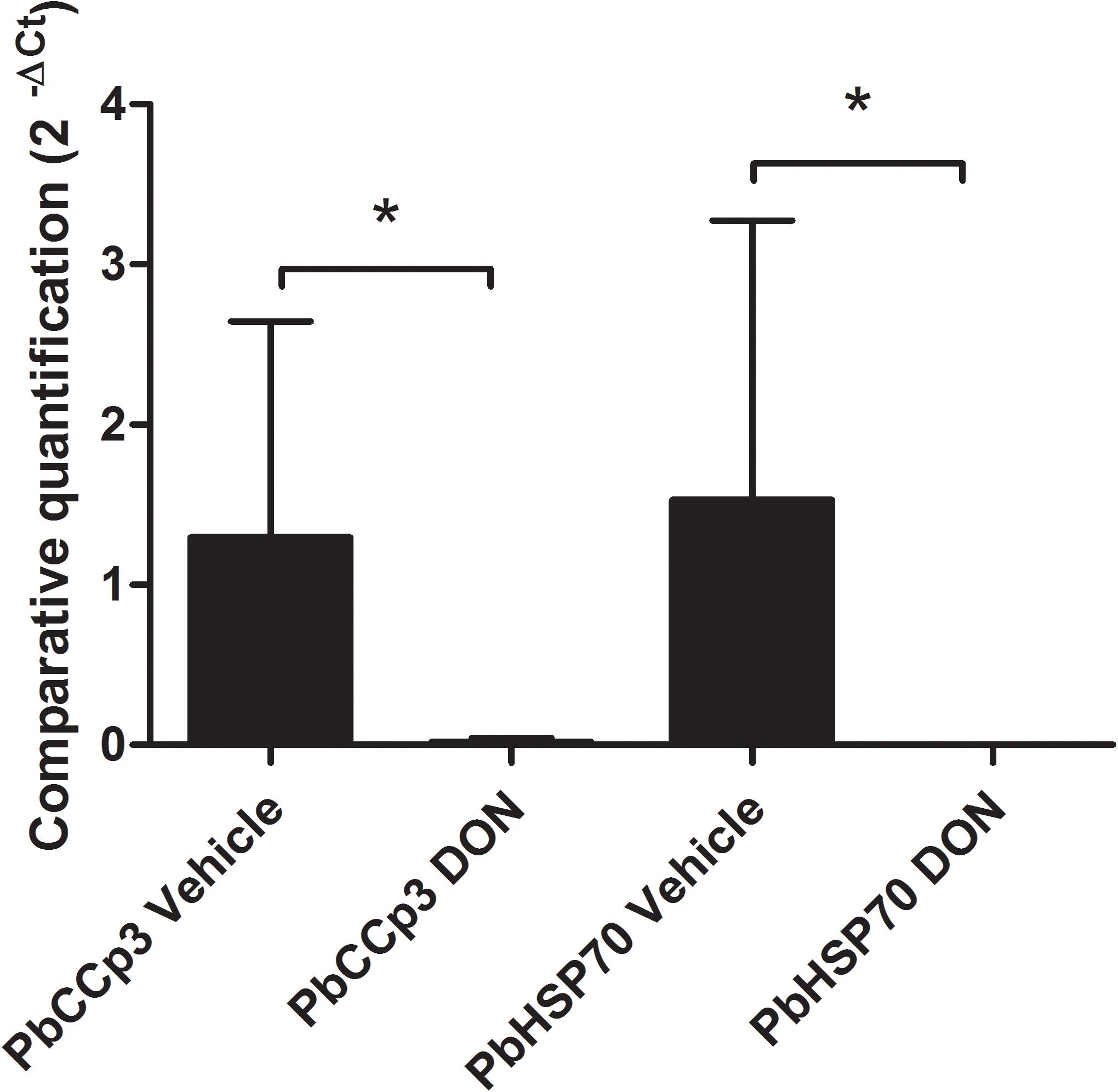
Figure 3. Inhibitory effect of DON on malaria erythrocytic stage development. CB57BL/6 mice were injected intraperitoneally with DON (1.3 mg/kg), administered consecutively every day, during the course of infection with 103 blood-stage Plasmodium berghei ANKA parasites. Gene expression levels by RT-qPCR at day 6 post-infection allow the detection of gametocytes and asexual erythrocytic forms of P. berghei. It was possible to verify the presence of circulating both forms in the whole blood of the animals used in the study and confirm the anti-parasitic action of the drug. Statistical analysis was performed using ΔCt values and we observed that treated animals showed significantly higher parasite density than untreated animals as regards to asexual and sexual forms of P. berghei. Data are means ± SD and differences between groups are significant (∗p < 0.05).
Our results indicating an inhibitory effect of DON on blood-stage P. falciparum allowed us to investigate a possible effect of this inhibitor on the infection in vivo. For this purpose, we first used an experimental model of murine cerebral malaria established by P. berghei ANKA infection in C57BL/6 mice which reproduces the critical pathological features of human cerebral malaria. Recent studies have shown that the therapeutic administration of DON in the late phase of murine cerebral malaria, from day 6 post-infection of P. berghei ANKA infection in CB57BL/6 mice, has an inhibitory effect on parasitemia, further showing that the glutamine analog is able to prevent pathogenic CD8+ T cell responses (Gordon et al., 2015). In the current study, our results demonstrate that when DON (1.3 mg/kg) was intraperitoneally injected beginning at day 6 post-infection and administered consecutively every day, during 4 days, there was a significant reduction of parasitemia in comparison to controls treated with saline alone (Figure 4A). All infected control animals developed neurological signs associated with murine cerebral malaria, showing a 100% mortality around day 7 post-infection. However, the animals treated with DON displayed a survival rate of 100% at 9 days post-infection (Figure 4B). It is worth of note that mice treated with DON and GlcN present a survival curve similar to the control (untreated mice), in which 80% died up to day 7 post-infection, and all died by day 8 post-infection (Figure 4B). These results clearly showed that GlcN reverts DON effect on mice survival, reinforcing the role of hexosamine biosynthetic pathway in parasite infection.
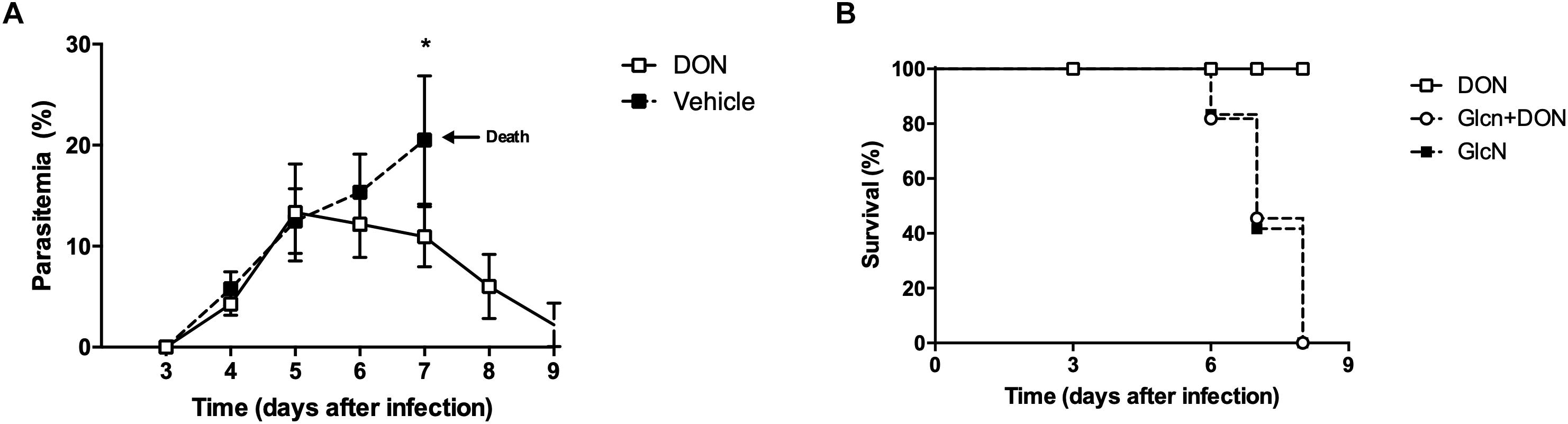
Figure 4. Inhibition of GFPT in the onset of malaria blood-stage infection attenuates the parasitemia and establishment of disease pathology. C57BL/6 mice were infected with 103 blood-stage P. berghei ANKA by intraperitoneal injection. DON (1.3 mg/kg) and/or GlcN (40 mg/kg) in 200 μL saline were i.p. daily after the day 5 pi. The control group was injected with saline. Results from (A) peripheral blood parasitemia (B) and Kaplan–Meier survival plots were obtained from three independent experiments. The results shown are combined from three independent experiments performed with five mice per group. Data are means ± SD and differences between groups are significant (∗p < 0.05).
We then investigated the ability of DON to act as a potential malarial transmission-blocking agent in view of its inhibitory effect on Plasmodium blood-stage development. For this purpose, we used the outbred Swiss Webster mouse model of cerebral malaria, as these are commonly used for testing vaccines and drugs that are expected to be effective in genetically heterogeneous background populations (Martins et al., 2009). Infection of these mice with P. berghei ANKA rapidly increased parasitemia but did not induce signs of cerebral malaria allowing the study of mouse-mosquito transmission. Corroborating our findings with CB57BL/6 mice, we detected a significant reduction in parasitemia indexes in infected Swiss mice treated with DON as compared to controls groups (Figure 5A). Analysis by chemiluminescence detection of luciferase-tagged P. berghei ANKA parasites in vivo (Figures 5B,C) confirmed a reduction in the blood-stage parasitemia.
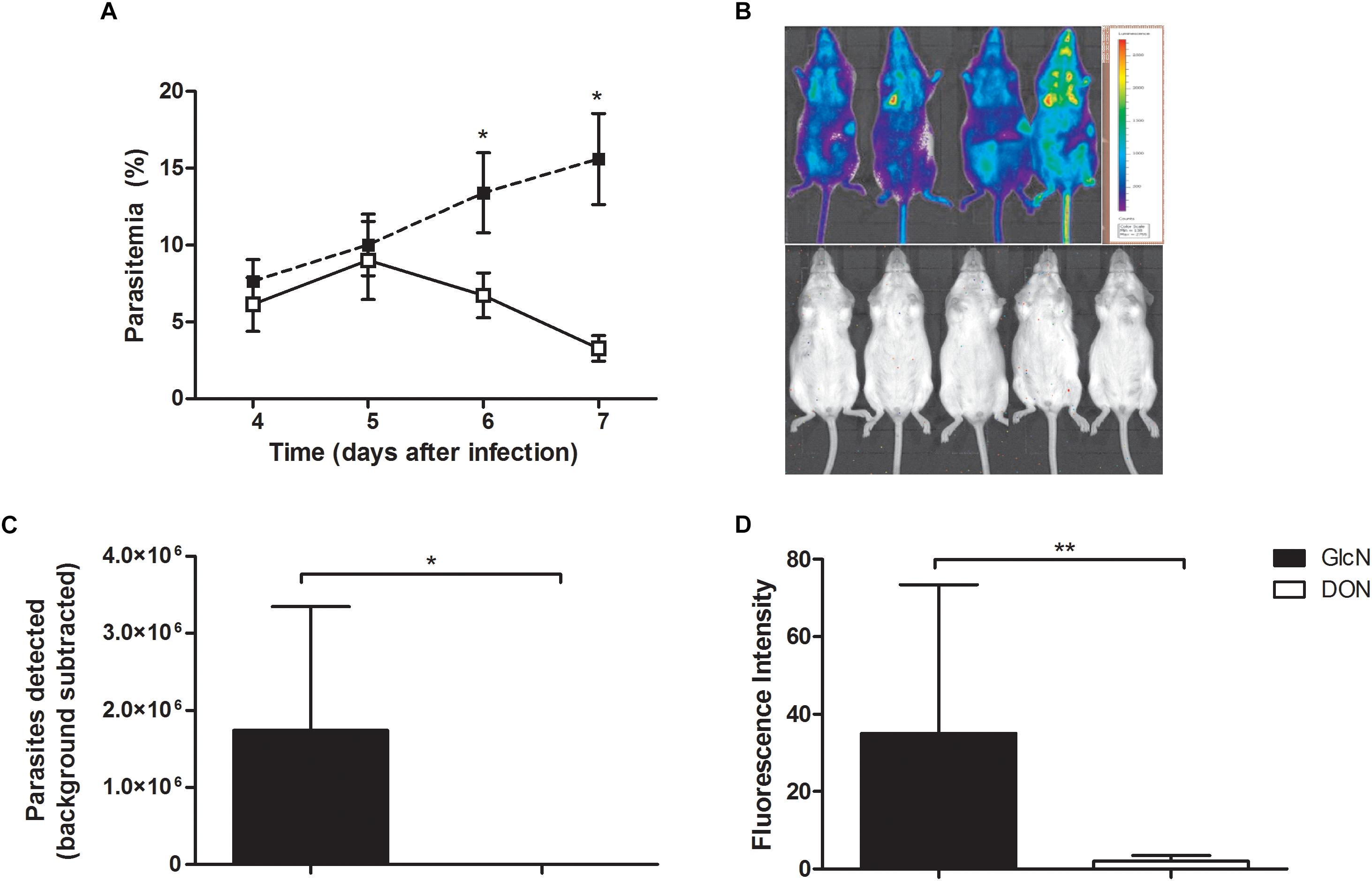
Figure 5. Potential effect of DON as malarial transmission-blocking agent preventing Plasmodium developmental cycle. Swiss Webster mice infected with 5 × 10-5 with luciferase-expressing P. berghei ANKA were treated with daily doses of DON (0.5 mg/kg) or vehicle (saline) as control from days 4 to 7 post-infection, administered intraperitoneally twice per day. Afterward, mice were injected (i.p.) with 150 mg/kg body weight of D-luciferin potassium salt substrate for bioimaging detection of asexual blood-stage parasites in vivo using IVIS imager (Lumina II In Vivo Imaging System; Perkin-Elmer). (A) Peripheral blood parasitemia were obtained from three independent experiments. The results are shown as means ± SE combined from three independent experiments performed with five mice per group. (B) Rainbow scale analysis of luciferase-tagged P. berghei ANKA parasite burden as representative bioluminescent images. (C) Quantification of the total luminescence intensity from whole body imaging of mice (n = 5 mice per group). (D) Transmission-blocking effect of DON in the sexual phase of the parasite life cycle. Infected Swiss Webster mice were alternatively treated with daily doses of DON (1.3 mg/kg) or vehicle as control from the beginning of infection (day 0) and 4 days later, experimental mouse groups (n = 5) were used to assay for the oocysts formation in the vector. Female Anopheles stephensi mosquitos (100 per experimental mouse group) were allowed to feed for 20 min and after 11 days later their maintenance at 18°C, the oocyst formation was determined from macerated midguts (n = 20 midguts per group) with addition of D-Luciferin potassium salt (200 μg/ml) for fluorescence intensity detection. Data are means ± SD and differences between groups are significant (∗p < 0.05, ∗∗p < 0.01).
We next investigated the potential effect of DON on Plasmodium transmission from vertebrate to mosquito. Swiss Webster mice were infected with 5 × 10-5 luciferase-expressing P. berghei ANKA and either treated or not with daily doses of DON (1.3 mg/kg) administered intraperitoneally from the beginning of infection (day 0). At day 4 post-infection, mice were anesthetized and Anopheles stephensi mosquitoes were allowed to feed on each mice group. Following incubation for 11 days, the levels of luciferase-expressing parasites in mosquitos’ midguts were estimated using D-Luciferin potassium salt. Our results demonstrated that, in fact, DON was able to significantly reduce the intensity of parasite transmission compared to control animals not treated with the inhibitor (Figure 5D), thus also indicating an effect of this inhibitor on development of sexual blood stages of malaria parasites.
Malaria is an infectious disease transmitted by mosquitoes and caused by parasitic protozoa of the genus Plasmodium (Cox, 2010). More severe cases of the disease may involve neurological complications and encephalopathies characteristic of cerebral malaria (Trampuz et al., 2003). These clinical manifestations are a consequence of the pathological activation of responses mostly triggered by CD8+ T lymphocytes directed against the parasite at the brain-endothelial interfaces (Howland et al., 2015a). Malaria prevention methods include mosquito eradication and use of medications able to target the parasite virulence (Ramirez et al., 2009). The prevalence of malaria in endemic area presupposes the combination of several factors, including high human population density, high population density of Anopheles mosquitoes, and a high rate of transmission between humans and mosquitoes (Martens and Hall, 2000). When any of these factors is reduced significantly, the parasite is expected to eventually disappear from the endemic regions as it occurred in North America, Europe and regions of the Middle East (Shiff, 2002).
There are a number of drugs available for malaria prevention in endemic areas. Many of these drugs are also used in the treatment of the disease (White, 2004). In cases where the parasite is still sensitive, chloroquine can be used. However, several species of Plasmodium are resistant to one or more drugs, so it is often necessary to resort to other drugs or combinations of drugs (White, 1999). Among these are mefloquine, doxycycline or the combination of atovaquone and proguanil (Gomes and Morrot, 2016). Ideally, drugs would be designed to act not only on prevention by acting directly on the parasite growth but also acting on the clinical symptoms by reducing disease severity. In this regard, important considerations are given to targets of carbohydrate metabolism and glycoconjugate biosynthesis pathways. For instance, GPI anchors are abundantly expressed in the surface of Plasmodium spp. and are determinants in the induction of parasite virulence in host–pathogen interactions and pathological responses to infection (Gazzinelli et al., 2014), while N-linked glycosylation seems to be essential to parasite grown (Dieckmann-Schuppert et al., 1992).
Previous work showed that glutamine analog DON reduced parasite growth in cultures of P. falciparum-infected erythrocytes (Waknine-Grinberg et al., 2010). Recently it was also shown that the activation of pathogenic CD8+ T cell responses in the course of cerebral malaria are dependent on glutamine metabolism and in vivo administration of DON in the late-phase of experimental cerebral malaria was able to inhibit the pathogenic CD8+ T cells responses thus rescuing the clinical manifestation signs of disease (Gordon et al., 2015). In the present study, we show that when administered to susceptible mouse models early in the onset of malaria blood-stage parasite infection, DON significantly decreased the levels of parasitemia thus reducing P. berghei ANKA lethality in CB57BL/6 mice. We demonstrated, for the first time, that the effect of DON in experimental malaria is due to inhibition of the aminotransferase activity of rate-limiting enzyme of the hexosamine biosynthetic pathway, GFPT.
Our findings demonstrated a requirement for GFPT activity for growth of erythrocytic forms of Plasmodium parasites. In vitro culture of P. falciparum in erythrocyte cells showed a significant inhibition of parasite growth in the presence of DON, which arrested parasite development at early (16 μM) and late trophozoite (1.6 μM) stages as well as induced abnormal morphology in mature forms. The inhibitory effect of DON was reverted upon exogenous addition of GlcN in infected erythrocytes, thus indicating a requirement for the UDP-GlcNAc downstream product of the hexosamine biosynthetic pathway in the parasite blood-stage cycle. In this line of evidence, coadministration of GlcN to DON treated mice reverts DON effect on mice survival, restoring high parasitemia and mortality of untreated mice. Moreover, our results indicate that the sensitivity of blood-stage forms of Plasmodium parasites to DON and GlcN, which reflects the importance of the parasite hexosamine biosynthetic pathway, is not restricted to dividing asexual forms, since our findings indicated that mice treated with DON presented significantly lower parasitemias of both asexual and gametocyte forms when compared with non-treated animals. In this regard, it has been shown that in erythrocytes infected with P. falciparum there is an increase of the glutamine influx as compared to normal cells that may reflect a possible increment of the hexosamine biosynthetic pathway needed for parasite blood-stage growth and cycle differentiation (Srivastava et al., 2016). Besides, studies have shown that gametocytes and gametes rely mainly on glycolysis but also display dependence on glutamine metabolism (Srivastava et al., 2016).
Hexosamine biosynthetic pathway is controlled by the rate-limiting enzyme GFPT which belongs to the aminotransferase family (Ginsburg, 2006) and generates UDP-GlcNAc, by de novo pathway. UDP-GlcNAc can be also obtained by the salvage pathway, perhaps due to the action of hexokinase that catalyzes the phosphorylation of glucosamine (GlcN) to GlcN-6-P, which then enters the same route as the de novo pathway (Sanz et al., 2013). UDP-GlcNAc is a donor substrate used in the production of short N-linked glycans and GPI anchors, both are essentials for parasite survival (Krishnegowda et al., 2005). Inhibition of N-linked glycosylation by tunicamycin, which catalyzes the transfer of GlcNAc-1P from UDP-GlcNAc to dolichol phosphate in the first step of N-linked glycoprotein synthesis (Vasconcelos-Dos-Santos et al., 2015), is lethal for the parasite (Naik et al., 2001). Inhibition of P. falciparum GPI by Glucosamine (GlcN) constrains the growth of the parasite in a dose-dependent manner. GlcN specifically arrested the maturation of trophozoites, a stage at which the parasite synthesizes all of its GPI anchor pool, and had no effect during the parasite growth from rings to early trophozoites and from late trophozoites to schizonts and merozoites (Naik et al., 2003).
Besides, addition of GlcNAc to P. falciparum culture is one of the most used method to obtain gametocytes stages in vitro (Ponnudurai et al., 1986; Fivelman et al., 2007). Our findings demonstrating the blood-stage parasite dependence of the GFPT enzyme may indicate the use of the glutamine metabolism through the hexosamine biosynthetic pathway. The findings indicating that DON is able to act directly on the growth and differentiation of the blood forms of the Plasmodium parasites as well as attenuate the pathology of the host inflammatory responses emphasizing its importance in the design of new therapies against malaria. Drugs able to reduce the parasite blood-stage cycle, affecting the sexual stages of Plasmodium parasites should have an immediate impact on mosquito infection (oocyst prevalence and intensity) and the transmission intensity of vectors in the endemic areas of malaria (Dyer and Day, 2000). In this regards, the therapeutic use of analogs of DON should provide a potential target for its use as transmission-blocking drugs against malaria.
PG, AL, OM, and PT conducted the experiments. PG, AL, OM, and PT acquired the data. PG, LC, AL, and OM analyzed the data. PG, ST, JG-D, JLJ, PT, AR, AT, and AM designed the research studies. LF-d-L, CF-d-L, OM, PT, AR, AT, and AM provided the reagents. PG, JLJ, OM, PT, and AM wrote the manuscript. All the authors have read and approved the final manuscript.
This work was supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico do Brasil (CNPq), Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and FIOCRUZ (PAEF II, IOC-023-FIO-18-2-47).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank the Real Time PCR Platform from Fiocruz (RPT09A) for the support on the real time PCR experiments.
Belnoue, E., Kayibanda, M., Vigario, A. M., Deschemin, J. C., van Rooijen, N., Viguier, M., et al. (2002). On the pathogenic role of brain-sequestered alphabeta CD8+ T cells in experimental cerebral malaria. J. Immunol. 169, 6369–6375. doi: 10.4049/jimmunol.169.11.6369
Bushkin, G. G., Ratner, D. M., Cui, J., Banerjee, S., Duraisingh, M. T., Jennings, C. V., et al. (2010). Suggestive evidence for darwinian selection against asparagine-linked glycans of Plasmodium falciparum and Toxoplasma gondii. Eukaryot. cell 9, 228–241. doi: 10.1128/EC.00197-09
Cox, F. E. (2010). History of the discovery of the malaria parasites and their vectors. Parasit. Vectors 3:5. doi: 10.1186/1756-3305-3-5
de Oca, M. M., Engwerda, C., and Haque, A. (2013). Plasmodium berghei ANKA (PbA) infection of C57BL/6J mice: a model of severe malaria. Methods Mol. Biol. 1031, 203–213. doi: 10.1007/978-1-62703-481-4_23
Dieckmann-Schuppert, A., Hensel, J., and Schwarz, R. T. (1992). Studies on the effect of tunicamycin on erythrocytic stages of Plasmodium falciparum. Biochem. Soc. Trans. 20:184S. doi: 10.1042/bst020184s
Dyer, M., and Day, K. P. (2000). Commitment to gametocytogenesis in Plasmodium falciparum. Parasitol. Today 16, 102–107. doi: 10.1016/S0169-4758(99)01608-7
Fivelman, Q. L., McRobert, L., Sharp, S., Taylor, C. J., Saeed, M., Swales, C. A., et al. (2007). Improved synchronous production of Plasmodium falciparum gametocytes in vitro. Mol. Biochem. Parasitol. 154, 119–123. doi: 10.1016/j.molbiopara.2007.04.008
Gazzinelli, R. T., Kalantari, P., Fitzgerald, K. A., and Golenbock, D. T. (2014). Innate sensing of malaria parasites. Nat. Rev. Immunol. 14, 744–757. doi: 10.1038/nri3742
Ginsburg, H. (2006). Progress in in silico functional genomics: the malaria metabolic pathways database. Trends Parasitol. 22, 238–240. doi: 10.1016/j.pt.2006.04.008
Gomes, P. S., and Morrot, A. (2016). Therapeutic approaches blocking glycan synthesis as targeting strategy for malaria. Curr. Clin. Pharmacol. 12, 26–30. doi: 10.2174/1574884711666161220152827
Gordon, E. B., Hart, G. T., Tran, T. M., Waisberg, M., Akkaya, M., Kim, A. S., et al. (2015). Targeting glutamine metabolism rescues mice from late-stage cerebral malaria. Proc. Natl. Acad. Sci. U.S.A. 112, 13075–13080. doi: 10.1073/pnas.1516544112
Howland, S. W., Claser, C., Poh, C. M., Gun, S. Y., and Renia, L. (2015a). Pathogenic CD8+ T cells in experimental cerebral malaria. Semin. Immunopathol. 37, 221–231. doi: 10.1007/s00281-015-0476-6
Howland, S. W., Poh, C. M., and Renia, L. (2015b). Activated brain endothelial cells cross-present malaria antigen. PLoS Pathog. 11:e1004963. doi: 10.1371/journal.ppat.1004963
Idro, R., Marsh, K., John, C. C., and Newton, C. R. (2010). Cerebral malaria: mechanisms of brain injury and strategies for improved neurocognitive outcome. Pediatr. Res. 68, 267–274. doi: 10.1203/00006450-201011001-00524
Jordan, K. A., and Hunter, C. A. (2010). Regulation of CD8+ T cell responses to infection with parasitic protozoa. Exp. Parasitol. 126, 318–325. doi: 10.1016/j.exppara.2010.05.008
Krishnegowda, G., Hajjar, A. M., Zhu, J., Douglass, E. J., Uematsu, S., Akira, S., et al. (2005). Induction of proinflammatory responses in macrophages by the glycosylphosphatidylinositols of Plasmodium falciparum: cell signaling receptors, glycosylphosphatidylinositol (GPI) structural requirement, and regulation of GPI activity. J. Biol. Chem. 280, 8606–8616. doi: 10.1074/jbc.M413541200
Lavazec, C., Moreira, C. K., Mair, G. R., Waters, A. P., Janse, C. J., Templeton, T. J., et al. (2009). A nalysis of mutant Plasmodium berghei parasites lacking expression of multiple PbCCp genes. Mol. Biochem. Parasitol. 163, 1–7. doi: 10.1016/j.molbiopara.2008.09.002
Macedo, C. S., Schwarz, R. T., Todeschini, A. R., Previato, J. O., and Mendonca-Previato, L. (2010). Overlooked post-translational modifications of proteins in Plasmodium falciparum: N- and O-glycosylation – a review. Mem. Inst. Oswaldo Cruz 105, 949–956. doi: 10.1590/S0074-02762010000800001
Martens, P., and Hall, L. (2000). Malaria on the move: human population movement and malaria transmission. Emerg. Infect. Dis. 6, 103–109. doi: 10.3201/eid0602.000202
Martins, Y. C., Smith, M. J., Pelajo-Machado, M., Werneck, G. L., Lenzi, H. L., Daniel-Ribeiro, C. T., et al. (2009). Characterization of cerebral malaria in the outbred Swiss Webster mouse infected by Plasmodium berghei ANKA. Int. J. Exp. Pathol. 90, 119–130. doi: 10.1111/j.1365-2613.2008.00622.x
Naik, R. S., Krishnegowda, G., and Gowda, D. C. (2003). Glucosamine inhibits inositol acylation of the glycosylphosphatidylinositol anchors in intraerythrocytic Plasmodium falciparum. J. Biol. Chem. 278, 2036–2042. doi: 10.1074/jbc.M208976200
Naik, R. S., Venkatesan, M., and Gowda, D. C. (2001). Plasmodium falciparum: the lethal effects of tunicamycin and mevastatin on the parasite are not mediated by the inhibition of N-linked oligosaccharide biosynthesis. Exp. Parasitol. 98, 110–114. doi: 10.1006/expr.2001.4616
Pinkus, L. M. (1977). Glutamine binding sites. Methods Enzymol. 46, 414–427. doi: 10.1016/S0076-6879(77)46049-X
Plaimas, K., Wang, Y., Rotimi, S. O., Olasehinde, G., Fatumo, S., Lanzer, M., et al. (2013). Computational and experimental analysis identified 6-diazo-5-oxonorleucine as a potential agent for treating infection by Plasmodium falciparum. Infect. Genet. Evol. 20, 389–395. doi: 10.1016/j.meegid.2013.09.019
Polimeni, M., and Prato, M. (2014). Host matrix metalloproteinases in cerebral malaria: new kids on the block against blood-brain barrier integrity? Fluids Barriers CNS 11:1. doi: 10.1186/2045-8118-11-1
Ponnudurai, T., Lensen, A. H., Meis, J. F., and Meuwissen, J. H. (1986). Synchronization of Plasmodium falciparum gametocytes using an automated suspension culture system. Parasitology 93(Pt 2), 263–274. doi: 10.1017/S003118200005143X
Queen, S. A., Jagt, D. L., and Reyes, P. (1990). In vitro susceptibilities of Plasmodium falciparum to compounds which inhibit nucleotide metabolism. Antimicrob. Agents Chemother. 34, 1393–1398. doi: 10.1128/AAC.34.7.1393
Ramirez, J. L., Garver, L. S., and Dimopoulos, G. (2009). Challenges and approaches for mosquito targeted malaria control. Curr. Mol. Med. 9, 116–130. doi: 10.2174/156652409787581600
Renia, L., Potter, S. M., Mauduit, M., Rosa, D. S., Kayibanda, M., Deschemin, J. C., et al. (2006). Pathogenic T cells in cerebral malaria. Int. J. Parasitol. 36, 547–554. doi: 10.1016/j.ijpara.2006.02.007
Samuelson, J., and Robbins, P. W. (2015). Effects of N-glycan precursor length diversity on quality control of protein folding and on protein glycosylation. Semin. Cell Dev. Biol. 41, 121–128. doi: 10.1016/j.semcdb.2014.11.008
Sanz, S., Bandini, G., Ospina, D., Bernabeu, M., Mariño, K., Fernández-Becerra, C., et al. (2013). Biosynthesis of GDP-fucose and other sugar nucleotides in the blood stages of Plasmodium falciparum. J. Biol. Chem. 288, 16506–16517. doi: 10.1074/jbc.M112.439828
Shiff, C. (2002). Integrated approach to malaria control. Clinical Microbiol. Rev. 15, 278–293. doi: 10.1128/CMR.15.2.278-293.2002
Srivastava, A., Philip, N., Hughes, K. R., Georgiou, K., MacRae, J. I., Barrett, M. P., et al. (2016). Stage-specific changes in plasmodium metabolism required for differentiation and adaptation to different host and vector environments. PLoS Pathog. 12:e1006094. doi: 10.1371/journal.ppat.1006094
Totino, P. R., Daniel-Ribeiro, C. T., Corte-Real, S., and Fátima Ferreira-da-Cruz, F. (2008). Plasmodium falciparum: erythrocytic stages die by autophagic-like cell death under drug pressure. Exp. Parasitol. 118, 478–486. doi: 10.1016/j.exppara.2007.10.017
Trampuz, A., Jereb, M., Muzlovic, I., and Prabhu, R. M. (2003). Clinical review: severe malaria. Crit. Care 7, 315–323. doi: 10.1186/cc2183
Vasconcelos-Dos-Santos, A., Oliveira, I. A., Lucena, M. C., Mantuano, N. R., Whelan, S. A., Dias, W. B., et al. (2015). Biosynthetic machinery involved in aberrant glycosylation: promising targets for developing of drugs against cancer. Front. Oncol. 5:138. doi: 10.3389/fonc.2015.00138
Waknine-Grinberg, J. H., Nicholas, H., Bentura-Marciano, A., McQuillan, J. A., Ho-Wai, C., Wing-Chi, C., et al. (2010). Artemisone effective against murine cerebral malaria. Malar. J. 9:227. doi: 10.1186/1475-2875-9-227
White, N. (1999). Antimalarial drug resistance and combination chemotherapy. Philos. Trans. R. Soc. Lond. B Biol. Sci. 354, 739–749. doi: 10.1098/rstb.1999.0426
White, N. J. (2004). Antimalarial drug resistance. J. Clin. Invest. 113, 1084–1092. doi: 10.1172/JCI21682
Keywords: Plasmodium falciparum, glycobyology, cerebral malaria, treatment strategies, parasites
Citation: Gomes PS, Tanghe S, Gallego-Delgado J, Conde L, Freire-de-Lima L, Lima AC, Freire-de-Lima CG, Lima Junior JdC, Moreira O, Totino P, Rodriguez A, Todeschini AR and Morrot A (2019) Targeting the Hexosamine Biosynthetic Pathway Prevents Plasmodium Developmental Cycle and Disease Pathology in Vertebrate Host. Front. Microbiol. 10:305. doi: 10.3389/fmicb.2019.00305
Received: 17 March 2018; Accepted: 05 February 2019;
Published: 28 February 2019.
Edited by:
Renato A. Mortara, Federal University of São Paulo, BrazilReviewed by:
Sandra Marcia Muxel, University of São Paulo, BrazilCopyright © 2019 Gomes, Tanghe, Gallego-Delgado, Conde, Freire-de-Lima, Lima, Freire-de-Lima, Lima Junior, Moreira, Totino, Rodriguez, Todeschini and Morrot. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Rodriguez, QW5hLlJvZHJpZ3VlekBueXVtYy5vcmc= Adriane Regina Todeschini, YWRyaWFuZXRAYmlvZi51ZnJqLmJy Alexandre Morrot, YWxleGFuZHJlLm1vcnJvdEBpb2MuZmlvY3J1ei5icg==; bW9ycm90QG1pY3JvLnVmcmouYnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.