 Tânia Aires
Tânia Aires Gerard Muyzer
Gerard Muyzer Ester A. Serrão
Ester A. Serrão Aschwin H. Engelen
Aschwin H. Engelen
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 09 January 2019
Sec. Aquatic Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.03283
The input of nutrients from anthropogenic sources is the leading cause of coastal eutrophication and is usually coupled with algal/seaweed blooms. Effects may be magnified in semi-enclosed systems, such as highly productive coastal lagoon ecosystems. Eutrophication and seaweed blooms can lead to ecosystem disruption. Previous studies have considered only one of these factors, disregarding possible interactive effects and the effect of the blooming species’ identity on sediment bacterial communities. We tested the effect of experimental nutrient loading and two common blooming seaweeds (Ulva rigida and Gracilaria vermiculophylla) in coastal lagoon sediments, on the structure of bacterial communities (using 16S rRNA amplicon sequencing) and corresponding putative functional potential (using PiCRUSt). At the Operational Taxonomic Unit (OTU) level, the addition of nutrients reduced bacterial community α-diversity and decreased the abundance of sulfate reducers (Desulfobacterales) compared to sulfur oxidizers/denitrifiers (Chromatiales and Campylobacterales), whereas this was not the case at the order level. Seaweed addition did not change bacterial α-diversity and the effect on community structure depended on the taxonomic level considered. The addition of Gracilaria increased the abundance of orders and OTUs involved in sulfate reduction and organic matter decomposition (Desulfobacterales, Bacteroidales, and Clostridiales, respectively), an effect which was also detected when only Ulva was added. Nutrients and the seaweeds combined only interacted for Ulva and nutrients, which increased known sulfide oxidizers and denitrifiers (order Campylobacterales). Seaweed enrichment affected putative functional profiles; a stronger increase of sulfur cycling KEGG pathways was assigned to nutrient-disturbed sediments, particularly with the seaweeds and especially Ulva. In contrast, nitrogen and sulfur cycle pathways showed a higher abundance of genes related to dissimilatory nitrate reduction to ammonium (DNRA) in Ulva+nutrients treatments. However, the other seaweed treatments increased the nitrogen fixation genes. Thiosulfate reduction, performed by sulfate-reducing bacteria, increased in seaweed treatments except when Ulva was combined with nutrients. In conclusion, the in situ addition of nutrients and the seaweeds to intertidal sediments affected the bacterial communities differently and independently. The predicted functional profile suggests a shift in relative abundances of putative pathways for nitrogen and sulfur cycles, in line with the taxonomic changes of the bacterial communities.
Coastal systems are affected by multiple anthropogenic stressors of both marine and terrestrial origin (Tallis et al., 2008), such as eutrophication. One of its main causes is the high input of phosphorus (P) and, more importantly, nitrogen (N) from agricultural fertilizer runoff and wastewater discharge (Nixon, 1995). This will, consequently, lead to elevated atmospheric N concentrations and increased atmospheric deposition of N (Holland et al., 2005), resulting in a dramatic rise of the amount of bioavailable N in natural habitats (Galloway and Cowling, 2002). In coastal marine systems, N is often a limiting nutrient (Taylor et al., 1995), but excessive nutrient inputs drive microbial communities to create biogeochemical feedbacks that promote the availability of N and P (Howarth et al., 2011). This positive feedback stimulates eutrophication and has negative effects, such as harmful or opportunistic blooms of seaweed (hereafter also referred as alga/algae) and the outgrowth of phytoplankton and epiphytic algae (Hauxwell et al., 2001). These blooms are the primary cause for oxygen depletion, which leads to hypoxia/anoxia with consequences for biodiversity (Isbell et al., 2013; Leff et al., 2015) and society (Leliaert et al., 2009).
Following seaweed-blooms, the decay of algal biomass increases oxygen consumption and results in anoxic conditions (Meyer-Reil and Köster, 2000). This, in turn, leads to a disruption in the ecosystem function and sediment biogeochemical cycling (Hale et al., 2016). Besides the “eutrophication-dependent” blooming seaweed, invasive algae may also proliferate due to a lack of grazing in the invaded areas (Engelen et al., 2011), which will later result in, mostly drifting, blooms that can lead to similar ecosystem disruption.
Bacteria play a determinant role in the biogeochemical cycles (Brune et al., 2000; Spring et al., 2000) of sulfur, nitrogen, and carbon, among others (Wetzel, 2000; Borsodi et al., 2003; Micsinai et al., 2003), and are responsible for nutrient transformation and organic matter decomposition (Zheng et al., 2014). Therefore, bacteria are crucial players in many ecosystem processes, such as the conversion of organic and inorganic compounds and, most importantly, the removal of anthropogenic nitrogen in coastal lagoons before its entry into the coastal ocean (Brin et al., 2010). Besides assimilation through sediments and into biological biomass and anammox, an essential process for nitrate removal is respiratory denitrification (Thamdrup, 2012). In contrast, dissimilatory nitrate reduction to ammonium (DNRA) retains nitrogen but in a more biologically available form (Tiedje, 1988). These two competing processes are mediated by bacteria and are dependent on the C/NO3- ratio (Burgin and Hamilton, 2007). Hence, identifying bacterial community shifts and their driving forces under common anthropogenic stresses is an essential step in predicting ecosystem responses and health during eutrophication and associated algal-bloom episodes.
The effects of eutrophication on biogeochemical cycles in sediments, at the chemical level, have been described by measuring the transformation rates of each final product (e.g., Sanz-Lázaro et al., 2015; Spivak and Ossolinski, 2016). Microbial processes have long been described and correlated with the degree of eutrophication in lakes (Chróst and Siuda, 2006). However, only in the last decade, advances in molecular techniques and the “omics” field allowed the expansion of our knowledge on the anthropogenic effects on coastal sediments through the study of their bacterial communities. For example, different levels of undifferentiated organic pollution revealed a consistent shift in microbial communities (Sinkko et al., 2013; Xiong et al., 2014), with bacterial taxa shifting to a Deltaproteobacteria-dominated guild, from which sulfate reducers prevailed. Other studies have reported contradictory results where sediment bacterial communities demonstrate resistance to nutrient inputs (Bowen et al., 2011), possibly by inducing dormancy in part of the community, which decreases diversity just on the active bacteria of the total community (Kearns et al., 2016). Hence, there are still large uncertainties regarding the prediction of the effects of eutrophication on sediment bacterial communities.
Algal blooms and bacterial communities are bidirectionally linked. Besides the pivotal role of bacteria in main biogeochemical processes and nutrient cycling, there is also evidence that they can regulate algal blooms (Kodama et al., 2006). Some planktonic bacterial groups can act either as growth stimulators (Anderson et al., 2012) or control factors. An example are the Cytophaga-Flavobacterium-Bacteroides (CFB), usually linked to the degradation of high molecular weight dissolved organic matter in the final stages of the blooms (Ishida et al., 1997; Jones et al., 2010). Consequently, bacterial communities are likely to change during a bloom (Grossart et al., 2005; Rooney-Varga et al., 2005; Chen et al., 2015), which makes it essential to understand how certain bacterial communities interact with algal blooms and determine the stage of the bloom at which this takes place. To date, most studies focused on the communities present in the water column or directly associated with the blooming seaweeds. However, earlier studies showed an increase of sulfur pools in eelgrass sediments by algal mats (Holmer and Nielsen, 2007), which might also be reflected in the sediment microbial community. Recently, Morrison and colleagues (2017) showed that bacterial communities from seaweed-enriched sediments, sourced from different inoculums, differed from this pre-enrichment community. Most of their distinctions were more inoculum-source than seaweed-species related but, in all of the treatments, the shifts were mainly toward the increase of bacterial lineages that are related to organic matter degradation. This suggests that algal blooms would lead to bacterial communities dominated by organic matter degraders and that algal identity is not of great importance.
Although nutrient inputs (which potentially affect sediment bacterial communities) are usually coupled with seaweed blooms (which itself triggers a shift in the bacterial community), their combined effects on sediment bacterial communities are still poorly understood. An important role of this interaction is, however, suggested by the observation that the composition of sediment bacterial communities only changed in response to nutrient fertilization in an area colonized by filamentous algae; possibly due to a shift in the carbon supply from those algae (Bowen et al., 2009). Moreover, changes in the bacterial community of both nutrient-poor and nutrient-rich (N and P naturally released by dreissenid mussels) freshwater sediments were shown to be associated with the presence of benthic algae, where bacterial taxa capable of degrading algal cellulose increased (Lee et al., 2015). Besides these few sources of evidence on the importance of nutrient and algal interactions for sediment bacterial communities, this topic strongly lacks further empirical and experimental evidence.
In this study, we conducted the first experimental field test of how nutrient input and algal loading (separately and combined) affect the sediment bacterial community structure in a coastal lagoon, and we predicted hypothetical functional consequences based on the resulting data. We used macroalgae (seaweeds) with different blooming strategies: Ulva rigida, whose blooms are driven by eutrophication (Fletcher, 1996; Gao et al., 2016), and Gracilaria vermiculophylla, which is invasive in the studied area, lacks natural predators, and often experiences blooms (Thomsen and McGlathery, 2007). These two seaweeds also present different structural profiles, with G. vermiculophylla showing a more ligneous/fibrous structure, highly resistant to anaerobic conditions and desiccation, in contrast with U. rigida, which has a thinner/fragile blade prone to desiccation, having a faster turnover in similar conditions. The distinct evolutionary histories of these two seaweeds are also noteworthy as they resulted in differentiated bacterial communities (Lachnit et al., 2011). We analyzed the bacterial community structure/composition using 16S rRNA gene amplicon sequencing on an Illumina MiSeq system in order to test three main hypotheses: (i) sediment bacterial community diversity will decrease and bacteria associated with nitrogen and sulfur cycles will change composition to respond to nutrient enrichment, (ii) seaweed enrichment will also affect bacteria related to nitrogen and sulfur cycles and decomposition of organic matter (carbon cycling), and (iii) the shifts in bacterial community will be cumulative (more evident) when the sediment is loaded with both nutrients and seaweed. By using two different seaweed species with different blooming strategies, we also aim to test the generality of the seaweed effects on bacterial communities. Last, we assess the predicted functional profiles and relate these to the main shifts in bacterial groups, linking them to possible responses to such environmental disturbances. This will contribute to a better understanding of the impact of eutrophication on its initial (nutrient increase) and secondary (algal bloom) stages, and the differentiated impact of different blooming algal species. This first experimental field test of effects of both nutrients and algal blooms reveals their individual and combined differentiated effects on bacterial communities in intertidal sediments.
The field experiment was performed during the summer of 2011 in the Ria Formosa lagoon, a warm temperate mesotidal system, located on the Atlantic coast of southern Portugal (37°N, 8°W). The lagoon is characterized by salt marshes, mudflats, and channels. It extends along the coast for about 55 km and is 6 km wide, with several connections to the Atlantic Ocean, ensuring water replacement during tidal cycles (1.5–3.0 m). Freshwater inflow is low and the salinity remains close to 36 (Machás et al., 2006). The experiment was conducted on a tidal flat with a vertical intertidal gradient of about 2 m. On average, Zostera noltii cover was 77%, with ∼8000 shoots/m2; 85 mm leaf length and above ground 219 gDW/m2, with a low abundance of seaweeds (19 gDW/m2) (DW – dry weight). Ambient nutrient levels at the experimental site were 1.0 μM NH3 (Ammonia), 0.6 μM PO4 (Phosphate), 1.0 DIN (dissolved inorganic nitrogen), and the average sediment organic matter was 3.6%DW during the experimental period. Seawater daily average temperature was 24.5°C (obtained from two HOBO loggers), and salinity stood at 36 psu, for the duration of the experiment.
Nutrient levels and seaweed abundances were manipulated in treatment plots of 0.25 m2, marked with zinc wire quadrats fixed on the sediment by two (about 30 cm long) zinc wire pegs in opposite corners, entirely open for natural intertidal conditions at the site. Quadrats were separate from each other by 1–1.5 m in blocks of 15 (1 per treatment), distanced by 5 m and set up on June 6th, 2011 (see Figure 1).
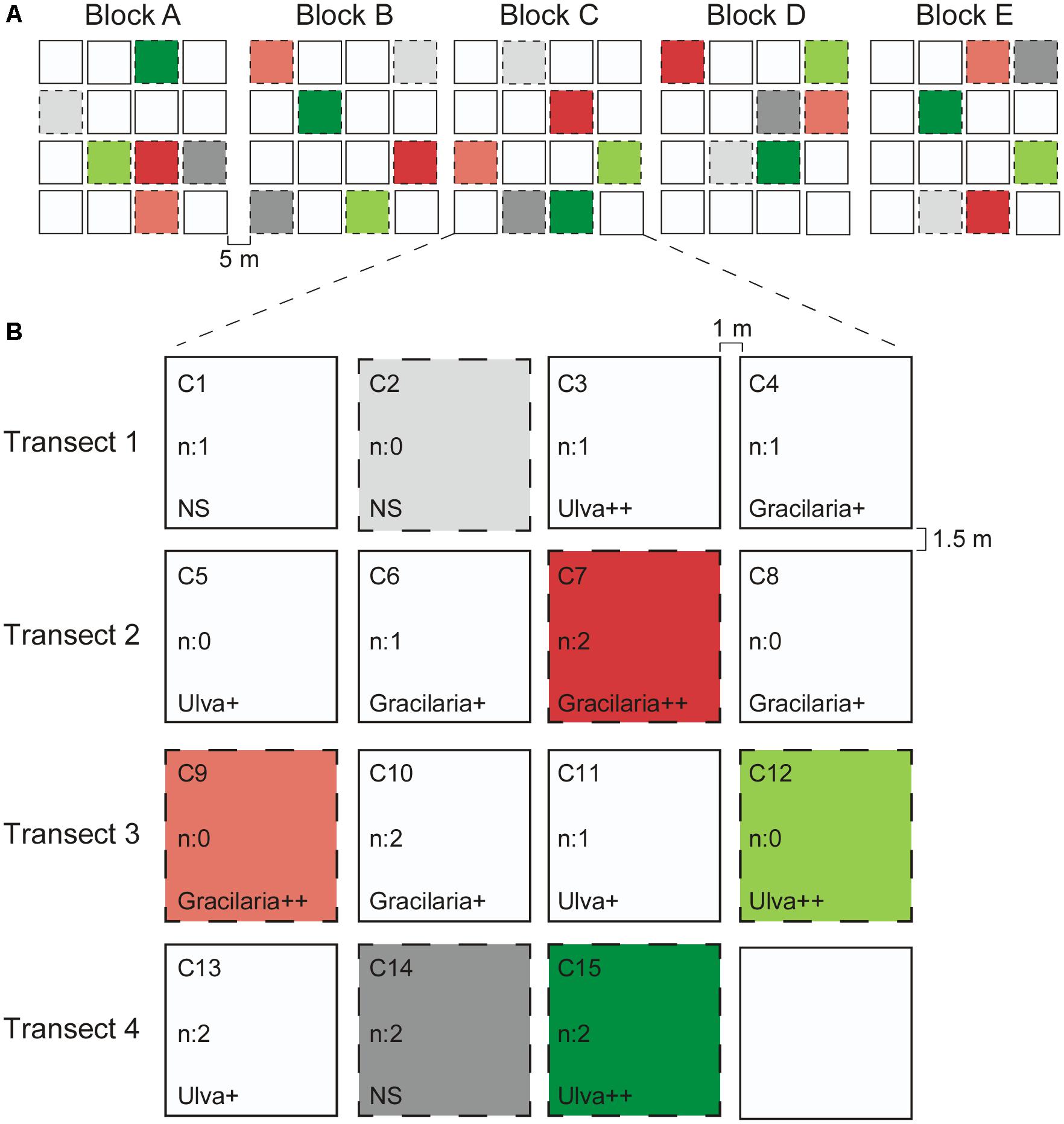
Figure 1. Experimental design. (A). Five blocks along a transect with 5 m in between the blocks. (B) Example of one of the blocks showing all the different treatment plots. Between two plots within a transect there is a distance of 1 m and the distance between two plots of adjacent transects is 1.5 m. The different treatments considered in this study are colored and are as follow: Light gray: n: 0 = no nutrients, NS = no seaweeds; Dark gray: n: 2 = 250 g N m-2 of nutrients added; Light red: n: 0 = no nutrients, Gracilaria++ = 3000 g WW m-2 of Gracilaria added; Dark red: n: 2 = 250 g N m-2 of nutrients added, Gracilaria++ = 3000 g WW m-2 of Gracilaria added; Light green: n: 0 = no nutrients, Ulva++ = 3000 g WW of Ulva m-2 added; Dark green: n: 2 = 250 g N m-2 of nutrients added, Ulva++ = 3000 g WW of Ulva m-2 added. Each block was composed by the same treatments and each one was sampled for each treatment totalling five replicates/treatment. No colored plots were not considered in this study.
The effect of eutrophication was tested with nutrient treatments consisting of (i) no nutrient (reference – N0) or (ii) 250 g slow diffusing N m-2 addition (N2). The nutrient addition was accomplished with a slow-release granulated fertilizer (Basacote Plus 3M, with 16% N, 8% P, and 12% K), which at the daily average seawater temperature during the experiment would release for about 3 months. To test the effects of the seaweeds, either (i) no seaweed (reference – SW0) or (ii) 3000 g m-2 (wet weight) of seaweed was added (for both species – Gracilaria, Ulva). In the Ria Formosa lagoon, both U. rigida and G. vermiculophylla are locally abundant; therefore, we used both seaweed species in the seaweed-enrichment treatment to test for species-specific effects. Overall, combinational treatments of two nutrient level treatments and two seaweed abundance treatments with two different seaweed species (Ulva and Gracilaria) were applied, and the positions of the treatment plots in a block were randomized. The seaweed biomass utilized was in accordance to biomasses found in the field (Cardoso et al., 2004) and used by others (Cardoso et al., 2004; Teichberg et al., 2010). The nutrient and seaweeds were added during low tide on June 13th, 2011.
The seaweeds (Ulva and Gracilaria) were collected in Quinta do Lago (Ria Formosa) 3 weeks before the start of the experiment and preserved in flow-through tanks at the CCMAR marine station. Before adding the algal biomass to the field, the seaweeds were cleaned of associated fauna. Treatments were randomly assigned to plots within blocks. Fertilizer granules were placed into the top centimeters of sediment and homogeneously distributed on the plot. If a plot had to be filled with both nutrients and seaweeds, the nutrients were put in first and, subsequently, the seaweeds were attached. Seaweed biomass was secured in plots by using five pegs per plot. Prior to the experiment, all plots were cleared of seaweed and all the plots received five pegs (including the no seaweed reference plot).
The experiment ran for 5 weeks (13th of June till 20th of July) during which photos of each plot were taken on the 17th and 30th of June and the 8th and 11th of July, to check whether the coated slow-releasing fertilizer granules and seaweeds were still in position. No other seaweed species than those added were observed in any of the plots during any of the surveys. The duration was chosen based on the commonly used time in drift algal-seagrass studies and reflects the short persistence time of drift algae in individual plots (Cardoso et al., 2004; Thomsen, 2010). By the end of the experiment, one replicate of 4 ml of the top few centimeters of sediment from each treatment was taken in cryovials from the correspondent plot in each of the five blocks (Figure 1) (totalling five replicates per treatment), flash frozen, and stored at -80°C until further processing. Seagrass root fragments (3.62 gDW ± 0.34 SE) were collected from each plot and these were processed for tissue nitrogen content as a proxy for plot-level nutrient status (Burkholder et al., 2007; Reynolds et al., 2018) using a Vario EL III elemental analyser (Elementar).
DNA was extracted with the PowerSoil® DNA Isolation Kit (MO BIO, Laboratories, Inc.) following the MO BIO Vortex Adapter protocol. The 16S rRNA gene V4 variable region was amplified by PCR using the primers 515F/806R (Caporaso et al., 2012) and processed for MiSeq Illumina sequencing by Research and Testing laboratories (Lubbock, TX, United States)1. Samples were amplified for sequencing using a forward and reverse fusion primer. The forward primer was constructed with the Illumina i5 adapter (5′-AATGATACGGCGACCACCGAGATCTACAC-3′), an eight bp barcode, a primer pad, and the 515F primer (GTGCCAGCMGCCGCGGTAA). The reverse fusion primer was constructed with the Illumina i7 adapter (5′-CAAGCAGAAGACGGCATACGAGAT-3′), an eight bp barcode, a primer pad, and the 806R primer (GGACTACHVGGGTWTCTAAT). Primer pads were designed to ensure the primer pad/primer combination had a melting temperature of 63–66°C, according to methods developed by the lab of Patrick Schloss2 (Kozich et al., 2013). Amplifications were performed in two independent 25 μl PCR reactions (which were pooled for the final assay) with Qiagen HotStar Taq master mix (Qiagen Inc., Valencia, CA, United States), 1 μl of each 5 μM primer, and 1 μl of template. Reactions were performed on ABI Veriti thermocyclers (Applied Biosystems, Carlsbad, CA, United States) under the following thermal profile: 95°C for 5 min, then 35 cycles of 94°C for 30 s, 54°C for 40 s, 72°C for 1 min, followed by one cycle of 72°C for 10 min. Samples were pooled together in equal proportions, based on their molecular weight and DNA concentrations. Calibrated Ampure XP beads were used to purify samples. DNA libraries were prepared by using an Illumina TruSeq DNA library preparation protocol.
Generated sequences were trimmed for quality and screened for a minimum read length of around 300 bp, sequences with ambiguous base calls were removed. Analyses were performed using the QIIME 1.8.0: Quantitative Insights Into Microbial Ecology (Caporaso et al., 2010) pipeline. Selected high-quality sequences were clustered into Operational Taxonomic Units (OTUs) within reads using the denovo OTU picking method. Representative sequences for each OTU were selected using the “most-abundant” method and OTU sequence alignment was carried out using PyNAST and Greengenes database v.13.8 (Caporaso et al., 2010). Taxonomic assignments were conducted through the UCLUST method with a 97% confidence threshold. Each OTU was assigned to its closest-matching description of taxon in the Greengenes database v.13.8 of 16S rRNA sequences (McDonald et al., 2012), and sequences were putatively assigned to a described taxon with a minimum threshold of 0.001 (default value). Eukaryote sequences (i.e., chloroplasts and mitochondria), as well as unassigned sequences, were excluded from the OTU table during downstream analyses. Due to the high variation in the number of high-quality sequences obtained among replicates (10311–124402 sequences), in the quality controlled OTU table the sample with the lowest number of sequences contained 10,311 reads. Therefore, all the samples were rarefied to this number of sequences for further statistical analyses (the rarefied table resulted in 18,278 unique OTUs and 289,576 sequences).
The statistical tests performed in this study were carried out using the rarefied data and considered significant at P < 0.05. Low abundance OTUs (singletons and doubletons) were removed from the dataset. The homogeneity of multivariate dispersions (based on the mean distance to group centroid for all groups within each factor) was tested using a resemblance-based permutation test (PERMDISP) (Anderson et al., 2006) before confirming differences in community structuring among sampling groups (β-Diversity) – by applying a permutational multivariate analysis of variance (PERMANOVA) using Bray–Curtis dissimilarity matrices from square-root transformed data.
For the β-diversity assessment, a multivariate analysis of microbial diversity using permutational analysis of variance (PERMANOVA) and a canonical analysis of principal coordinates (CAP) were performed. PERMANOVA tested for differences among samples with a priori factors: Different Treatments: Nutrients (N2) vs. No-nutrients (N0), Seaweeds (Ulva and Gracilaria) vs. No-seaweeds (SW0), and the interaction between both [Reference (N0SW0), N2SW0, N0Ulva, N0Gracilaria, N2Ulva, N2Gracilaria] and differences within seaweeds’ treatment [No seaweeds (SW0) vs. Gracilaria/Ulva, and Gracilaria vs. Ulva]. A canonical analysis of principal coordinates (CAP) tested the assignment/clustering of treatments interaction (Nutrients X Seaweeds) as an a priori factor. Discriminant vectors with a Pearson correlation >0.6 were considered for the characterization of the taxa discriminating the multivariate patterns. Since not all the OTUs were assigned to the same taxonomic level, only bacterial orders were considered in order to have a homogeneous characterization (at the same taxonomic level for each vector). SIMPER analyses were performed to determine which taxa contribute most to differences among groups. All analyses were performed using the software program PRIMER-E+PERMANOVA v.6 (Clarke and Warwick, 1994).
To investigate the shifts in the bacterial community at a higher taxonomic level and consistently present through the different replicates, we filtered the OTU table, at the order level, for OTUs that were present in at least 60% of the replicates. A bar chart was generated with those orders remaining after filtration and considering only the ones with a relative abundance of above 1.5%. The putative roles of the main OTUs/orders found for each of the treatments were inferred from the literature. A heat map was built, using STAMP (Parks et al., 2014), where the different treatments were clustered independently using Ward’s clustering method (with a 0.75 threshold) for the main orders’ relative abundances (as used for the bar chart above).
Estimates of α-diversity were based on evenly rarefied OTU level and most common orders abundance matrices and included the diversity and evenness indicator Shannon index and the richness and abundance Indexes Chao I and Observed OTUs, as calculated in QIIME. In order to maximize comparability with the analysis of β-diversity, treatment effects on α-diversity were examined using univariate PERMANOVA, based on Euclidean distances (as in Hartmann et al., 2015). Using the compare_alpha_diversity.py script on QIIME, boxplots were generated for a visualization of the α-diversity differences among groups.
In order to predict metagenomes, closed-reference OTU picking was performed using the subsampled reads against the GreenGenes reference database (version GG 13.5) using the pick_closed_reference_otus.py script from QIIME (Caporaso et al., 2010). This was done separately from the bacterial community composition analysis because PiCRUSt requires an older version of GreenGenes, as well as the closed reference OTU picking method, which would limit a more comprehensive and broad diversity analysis. PiCRUSt (Langille et al., 2013) was used online in the Galaxy environment following the PiCRUSt workflow from Huttenhower’s Lab3. This included 16S copy number normalization, metagenomes/function prediction, and function categorizing into KEGG pathways. General statistical analyses were performed using PERMANOVA following the same procedure as for the 16S rRNA gene analysis of the bacterial communities in PRIMER-E+PERMANOVA v.6 (Clarke and Warwick, 1994).
Based on the most important biogeochemical cycles in coastal sediments (i.e., sulfur and nitrogen cycles), candidate functional categories (KEGG classification, pathway category Level 3) were chosen and analyzed in detail for each treatment group. These analyses were performed with the software program STAMP v. 2.1.3 (Parks et al., 2014) using ANOVA and the multiple groups’ boxplot visualization for the Nutrient and Seaweed combined treatments. When statistical differences were found (p < 0.05), post hoc Turkey–Kramer tests were performed to identify the source of the variation. Besides, from the predicted metagenomes table calculated in PiCRUSt, the KEGG orthologues implicated in the different pathways of the sulfur and nitrogen cycle (according to the KEGG pathway database) were selected. The selected genes were pooled by pathway and plotted to compare relative abundances for each treatment.
To confirm if the closed-reference OTU table presents the same trend for the main orders as the OTU table used for the bacterial community analysis, the procedure described above, for the bar chart construction, was applied to the closed-reference OTU table. That was used to strengthen the assumptions on putative functions made for the main OTUs/orders characterized by 16S barcoding (see above). The OTU table was then filtered by the orders that were more abundant (based on the bar chart graph) in each treatment group (Reference, N2SW0, N0Grac, N2Grac, N0Ulva, and N2Ulva). PiCRUSt analyses were then repeated to target sulfur metabolism (found to be the most shifting function) in these orders and to check if the functional profile followed the same pattern as the community profile. To investigate the carbon cycle, we directly selected the Level 3 KEGG pathways that were related to carbon utilization and present in the PiCRUSt list. The statistical analyses and visualization of the results were performed with STAMP (Parks et al., 2014).
Our raw dataset consisted of a total of 2,896,310 sequences. After quality control, and removal of chimeras, chloroplast, unassigned sequences, and singletons, a total of 1,114,886 high-quality sequences were retained for analysis (accessible through MG-RAST)4, consisting of a total of 21,812 unique OTUs (Supplementary Table S1).
Alpha diversity indices were calculated for each replicate of each treatment at both OTU and order levels (except for the Observed OTUs) and individual values are shown in Supplementary Table S2. Bacterial α-diversity was reduced by nutrient addition only when expressed by the Shannon index (Table 1 and Figure 2). Seaweed addition did not affect bacterial α-diversity according to any of the three indices used (Table 1).

Table 1. Results of PERMANOVA main test for the α-diversity indexes calculated: Shannon Diversity, ChaoI and Observed OTUs.
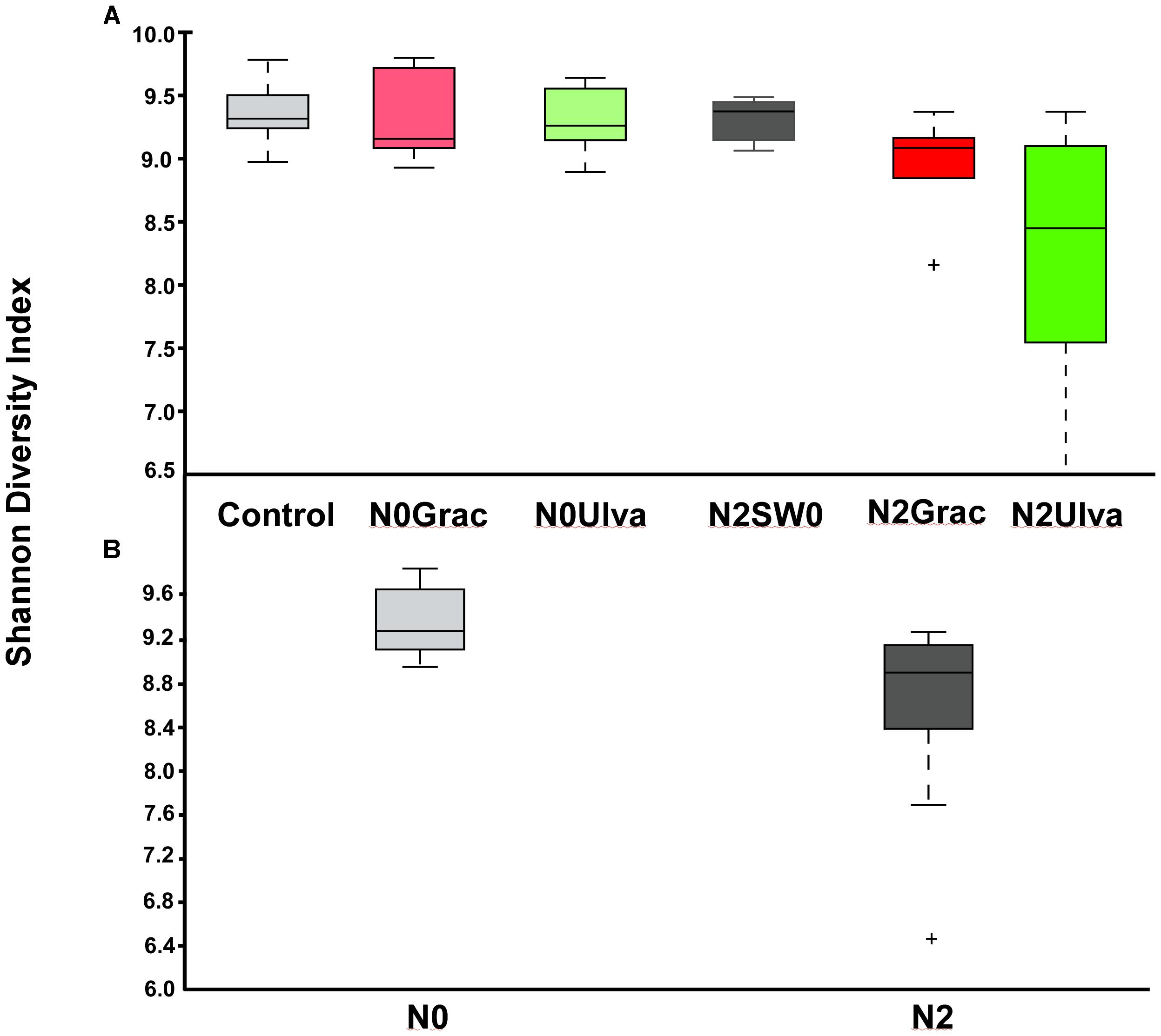
Figure 2. (A) Boxplot of the bacterial α-diversity using Shannon Diversity Index of intertidal sediment bacterial communities based on Illumina 16S rRNA amplicon sequencing for ambient conditions (Reference) and experimental conditions of nutrient (N) and seaweed (SW) addition. (B) Once significant differences were detected for nutrient addition, all non-nutrient vs. nutrient treatments were plotted together and new plots presented below the main plot. The line in the middle of each box is the median, crosses are outliers, and the whiskers represent error bars. From left to right: N0SW0 – No nutrients or seaweeds added (Reference), N0Grac – Only Gracilaria added, N0Ulva – Only Ulva added, N2SW0 – Only nutrients added, N2Grac – Both nutrients and Gracilaria added, N2Ulva – Both nutrients and Ulva added. Graphics were manipulated to both fit in the same figure. In addition, colors were changed so different treatment colors would match those of Figure 1.
At the OTU level, the bacterial community structure (abundance and composition) was affected by both seaweed and nutrient addition, independently of each other (Table 2A and Figure 3). For seaweed addition, Gracilaria strongly affected the structure of the bacterial community (SW0 vs. Gracilaria p = 0.024), whereas the Ulva addition had a marginal effect (SW0 vs. Ulva p = 0.079; Gracilaria vs. Ulva p = 0.405) (Table 2B). The Gracilaria addition (i) increased the average abundance of two Desulfobacterales order OTUs (denovo19603 and denovo36296) 20 and three times, respectively, (ii) increased the average abundance of a Clostridiales OTU by seven times (denovo1309), and (iii) doubled the average abundance of a Campylobacterales OTU (denovo60628). As such, increasing orders were involved in sulfate reduction and nitrogen fixation. In contrast, the average abundance of some specific OTUs of the order Thiohalorhabdales (denovo64034), Chromatiales (denovo6380), and Marinicellaceae (denovo60848) decreased 1.3 times with the addition of Gracilaria (SIMPER analysis, Supplementary Table S3A). Gracilaria addition also slightly increased the number of reads of methanogenic Archaea (p = 0.075), especially of Methanosarcinaceae and Methanomassiliicoccaceae members, and in that way affected Archaea composition (p = 0.015), whereas the nitrogen processing bacterial community was not driven by seaweed addition (p > 0.0359).
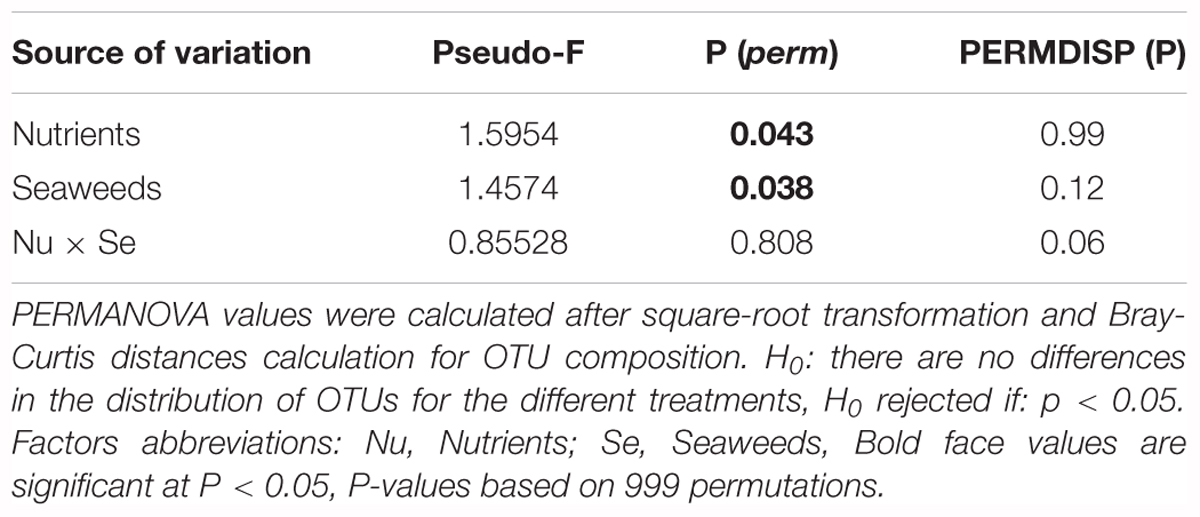
Table 2A. PERMANOVA main test for bacterial community β-diversity.
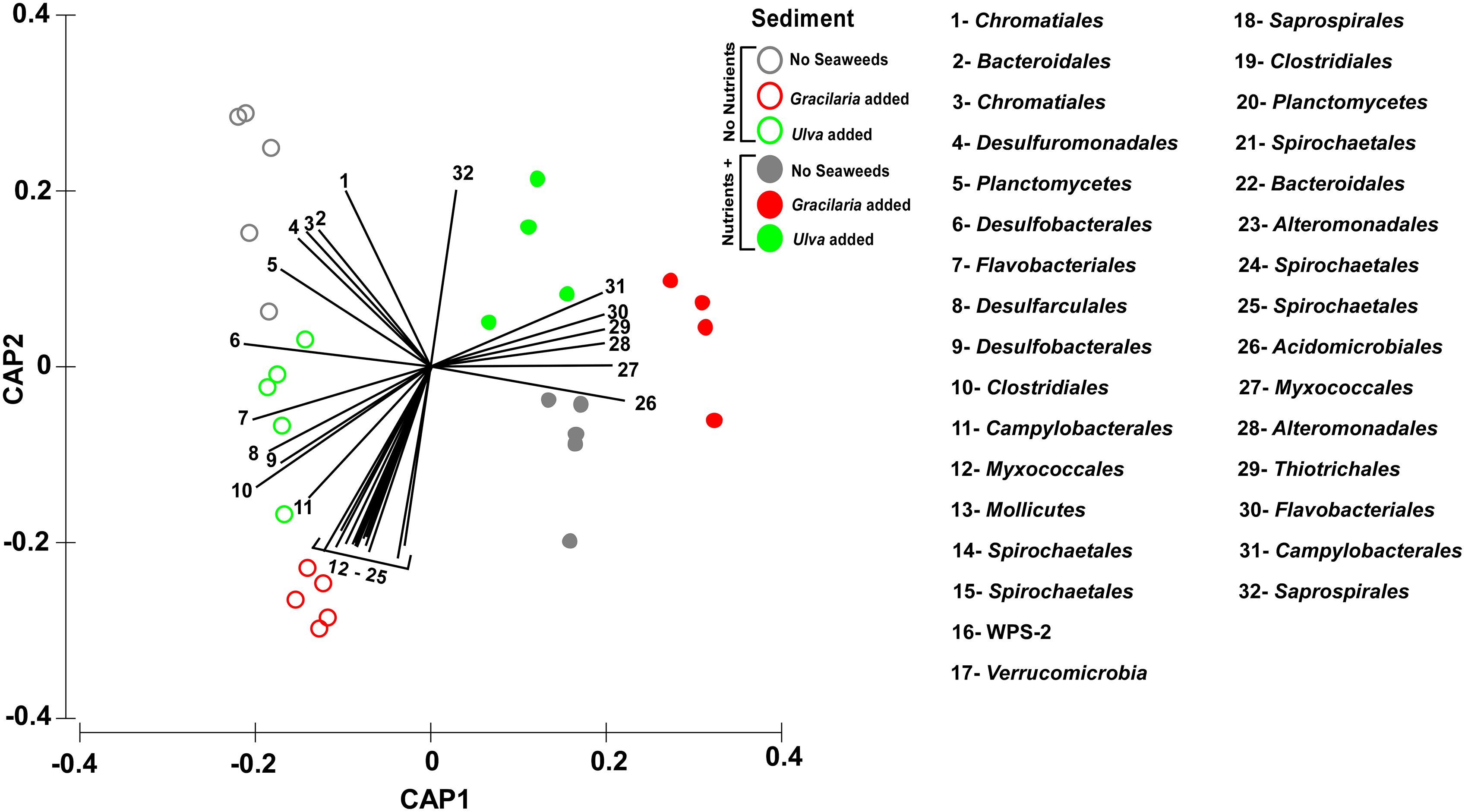
Figure 3. Canonical analysis of principal coordinates (CAP) ordination plot (based on Bray–Curtis of square-root transformed bacterial abundances) showing canonical axes that best discriminate the bacterial community assemblages across nutrient (open vs. closed circles) and seaweed addition (grey – no seaweeds, red – Gracilaria, green – Ulva). Order level OTUs with Pearson’s correlation >0.70 have been overlaid on the plot as vectors. Vector length corresponds to the strength of the correlation, with the circle representing the maximum correlation of 1. Image was manipulated by replacing the taxa names by numbers (with correspondence in lateral legend), in order to improve visualization. In addition, colors were changed so different treatment colors would match those of Figure 1.

Table 2B. Results of PERMANOVA pair-wise test within the factor “Seaweeds” of OTU composition.
Although the nutrient addition did not affect the absolute N content and the C:N ratio of seagrass root fragments (p > 0.1600), it decreased the relative abundances of OTUs, predicted to be sulfate reducers (e.g., vectors 4, 5, 6, 8, 9, and 10 in Figure 3), whereas putative sulfur/sulfide oxidizers’ OTUs increased under nutrient loading conditions (vectors 29 and 31; Figure 3). The OTUs contributing most to this nutrient driven shift in community (recognized S-oxidizers and denitrifiers) increased when nutrients were added, with the most prominent being: Campylobacterales (denovo60628 and denovo42704) and Chromatiales (denovo64003), increasing 10, 17, and 9 times, respectively, (SIMPER analysis, Supplementary Table S3B). On the other hand, an OTU assigned to the sulfate reducer Desulfobacterales order (denovo19603) was reduced by 1.4 times in abundance by nutrient addition (SIMPER analysis, Supplementary Table S3B). The Desulfobacterales, also known as anaerobic N fixers, like Clostridiales and OTUs of both seaweed species, correlated negatively with the N addition (vectors 6, 9, 10, and 19 in Figure 3). For microaerophilic nitrogen-fixing Campylobacterales, two OTUs showed strongly contrasting correlations linked to nutrient addition (vectors 11 and 31 Figure 3) but coincided with Ulva addition.
Considering only abundant orders (those in proportions higher than 1.5% and present in at least 60% of the replicates to assure representability of the entire replication), bacterial α-diversity did not differ among treatments, regardless of the indexes used (Supplementary Table S4 – Shannon diversity index and Chao I).
For abundant orders, all plots without seaweed addition (irrespective of their nutrient treatment) displayed the same top six orders and equivalent proportions of putative sulfur/sulfide oxidizers (SO) and putative sulfate reducers (SR). Therefore, the bacterial community structure and composition were marginally affected by the nutrient addition (p = 0.056, Supplementary Table S5). Among the most abundant orders were Chromatiales (SO)- 11.2%, 12.3% (N2SW0); Desulfobacterales (SR)- 10.8%, 8.3% (N2SW0); Thiotrichales (SO)- 6.5%, 8.5% (N2SW0); and Alteromonadales (SR)- 5.5%, 5.1% (N2SW0) (Figure 4). Order level community structure/composition differed with seaweed treatment (p = 0.012, Supplementary Table S5A), with both seaweeds affecting sediment bacterial community equally (Gracilaria p = 0.008 and Ulva p = 0.016, Supplementary Table S5B). Desulfobacterales was the most abundant order in the seaweed addition treatments (N0Grac-23.9%, N2Grac-21.8%, and N0Ulva-23.8%), except in the N2Ulva treatment (9.2%) (Figure 4). Instead, Campylobacterales (SO) was conspicuously more abundant in the N2Ulva treatment (23.6%) compared with Ulva addition under ambient nutrient conditions (3.9%) and with the other treatments (Reference 1.9%, N2SW0 3.8%, N0Grac 3.4%, and N2Grac 8.2%) (Figure 4). The order Bacteroidales was the second most abundant only in seaweed-added sediments with ambient nutrient levels (8.5% for N0Grac and 7.7% for N0Ulva) but was also within the first orders for the combined nutrient and seaweed treatments (Figure 4). The order Clostridiales had an evident increase in Gracilaria-enriched sediments being very low to non-detectable in the other treatments (Reference- 0%, N2SW0- 0%, N0Grac-6.6%, N2Grac-4.2%, N0Ulva-1.5%, and N2Ulva-0%) (Figure 4). The heat map with samples clustering based on similarities given by the relative abundance of the main orders across all treatments shows a clear separation between the Gracilaria addition and the no seaweed addition treatments. Communities from plots where Ulva was added clustered either with the Gracilaria addition treatment (in the case of N0U), due to high abundances of Desulfobacterales (anaerobic N and S cyclers) or took an intermediate position between “no seaweeds” and the remaining “with seaweeds” treatments due to a unique high abundance of Campylobacterales (microaerophilic N fixers, Supplementary Figure S1; in the case of N2U). This position is more in agreement with OTU level beta-diversity multivariate analysis results that show an intermediate state of communities from the Ulva treatment, which is not statistically different from “no seaweeds” and Gracilaria treatments. Ambient plots without seaweed additions, as did the N2U treatment, showed distinct higher relative abundance of Chromatiales purpur sulfur bacteria known to contribute to P, S, and C cycling.
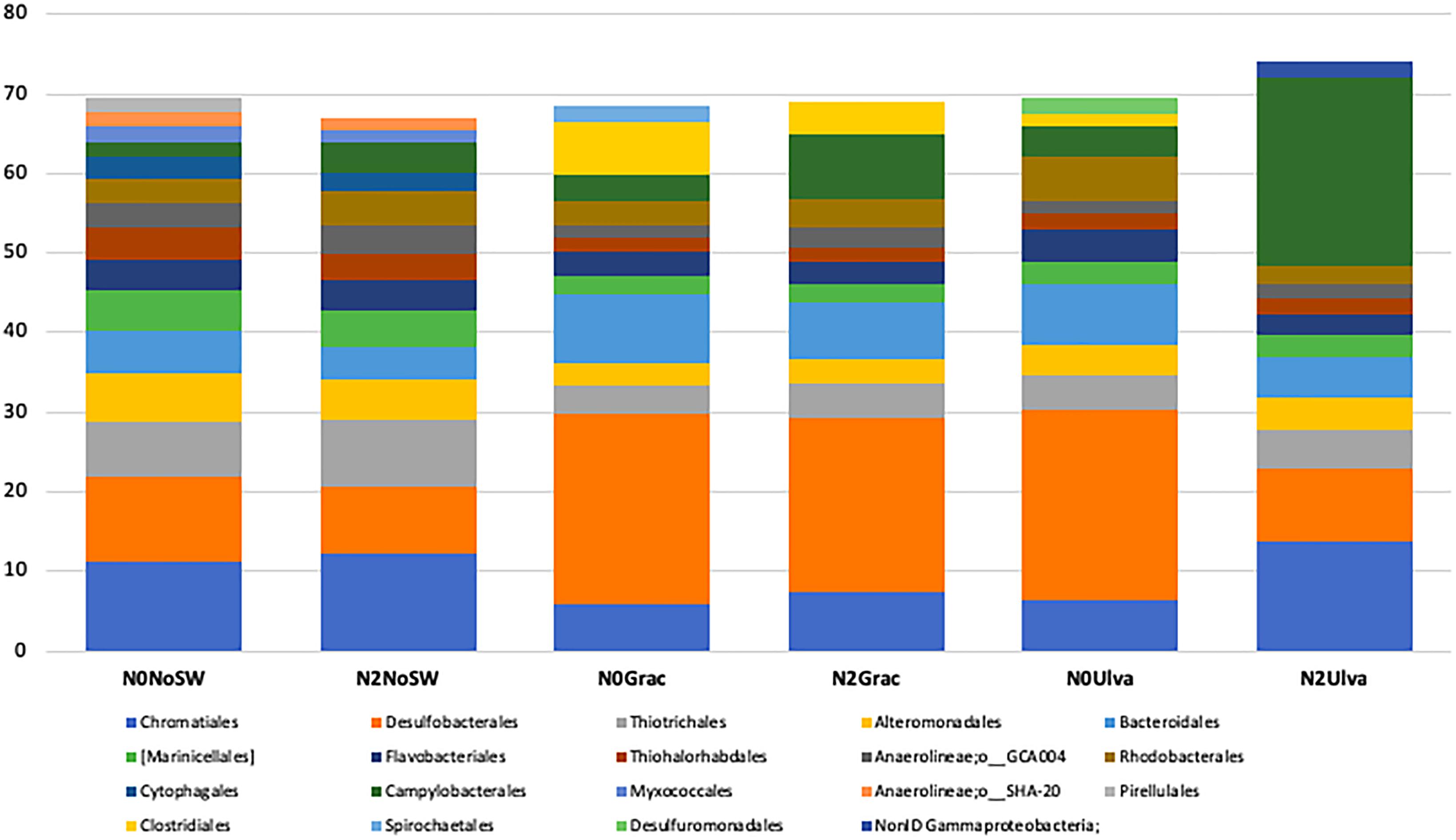
Figure 4. Mean bacterial community structure of sediments showing the most abundant (>1.5%) orders for each treatment, each represented by five replicate plots sampled, after filtering for the orders present in at least 60% of the replicates. Control – No seaweeds or nutrients added, N2SW0 – Only nutrients added, N0Grac – Only Gracilaria added, N2Grac – Both nutrients and Gracilaria added, N0Ulva – Only Ulva added, N2Ulva – Both nutrients and Ulva added.
The PiCRUSt functional predictions had NSTI (Nearest Sequenced Taxon Index) scores ranging from 0.12 to 0.18, with an overall mean of 0.15 ± 0.02, a value even lower than that observed in the reference sediment. This indicated small average phylogenetic distances between the obtained 16S amplicon sequences and those of fully sequenced genomes used for PiCRUSt inferences suggesting likely accurate predictions. Predicted human-related functions/diseases were removed from the final table of functions (Supplementary Table S6).
The overall predicted functions, in contrast with the bacterial community structure, were only affected by seaweed and not by the nutrient addition (Table 3A), with both Gracilaria (p = 0.011) and Ulva (p = 0.016) similarly affecting the predicted sediment functional profile (Table 3B). Surprisingly, differences in predictive functionality with both seaweed additions were driven by increased cell motility and signal transduction and decreased Tryptophan and fatty acid metabolism and Valine, Leucine, and Isoleucine degradation (SIMPER).
Table 3A. Results of PERMANOVA main test for PiCRUSt functional predictions.

Table 3B. Results of PERMANOVA pair-wise test within the factor “Seaweeds” on predicted functional profiles.
Although the overall functional predictions were only affected by the seaweed addition, the specific candidate KEGG categories related to the sulfur cycle were, in general, predicted to be more affected by the nutrient addition. Sulfur cycle-related sequences, such as the sulfur metabolism (p = 0.025) and the sulfur relay system (p = 0.004), were predicted to increase in sediments under the nutrient addition. This predicted increase was even more evident in the plots in which nutrients were combined with seaweeds, especially in the case of Ulva (Figures 5A–D). In contrast, nitrogen-cycling genes were predicted to be more, albeit insignificantly, affected by seaweed rather than the nutrient addition (Figure 5E), either linked to the increased variation in response among samples or reflecting the strong local effect of seaweeds.
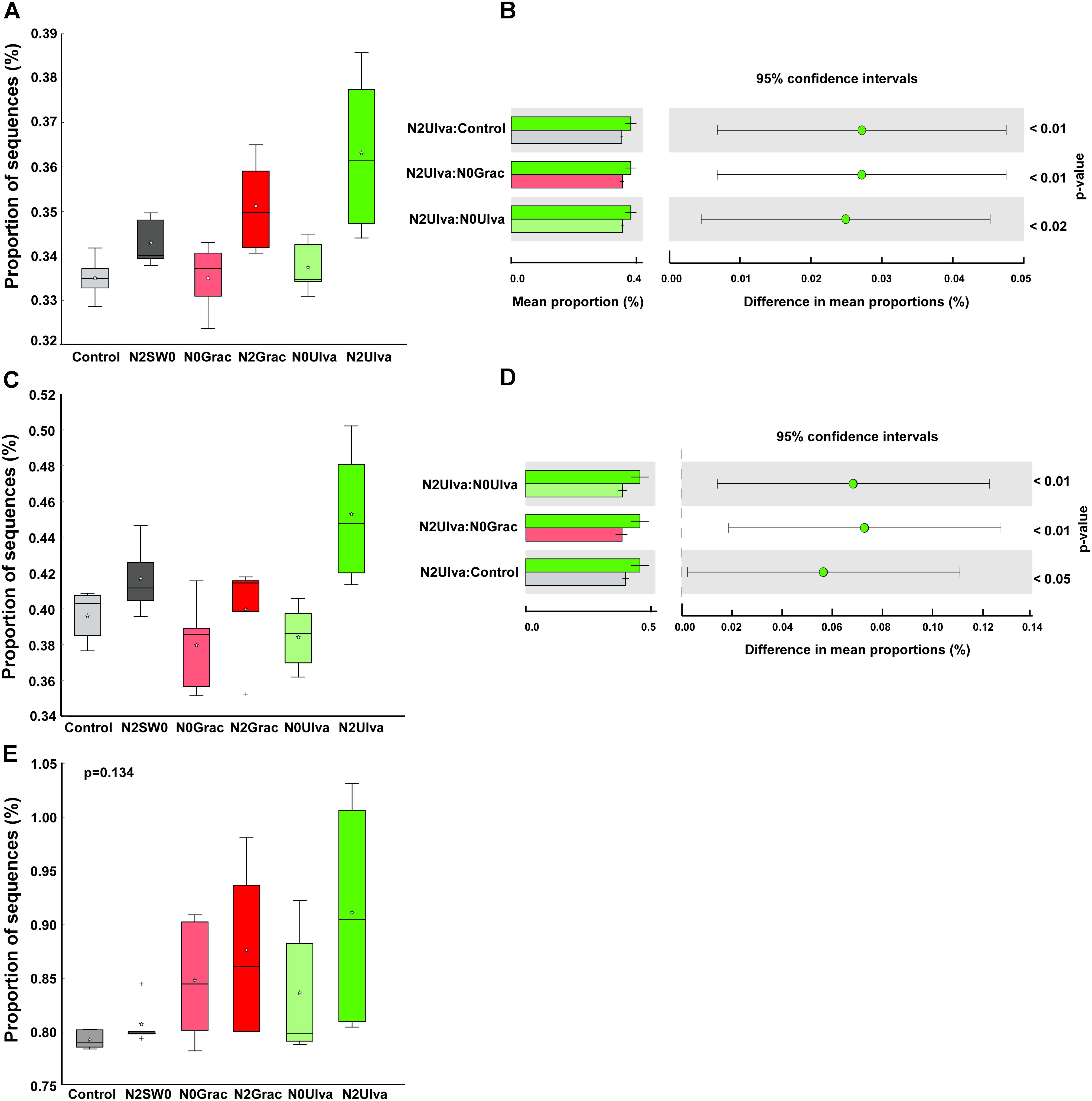
Figure 5. Box plot showing the distribution in the proportion of sequences assigned to the candidate functions: (A) Sulfur metabolism, (C) Sulfur relay system, and (E) Nitrogen metabolism. The top of the box indicates the third quartile, the bottom the first quartile, and the line in the middle is the median. Outliers are shown as crosses and the mean value as a star within the box. Right-side figures show Turkey–Kramer post hoc tests for each order where statistical differences were found (B) Sulfur metabolism, (D) Sulfur relay system, indicating the mean proportion of sequences within each treatment and the difference in mean proportions for each pair of treatment, with their corresponding pairwise p-value. Graphic colors were changed so different treatment colors would match those of Figure 1.
The KEGG genes specifically selected for nitrogen and sulfur cycles and pooled according to the different pathways (for samples pooled according to treatments), showed the presence of seven different pathways (Supplementary Table S7 and Figure 6). The nitrogen cycle pathways were: nitrogen fixation, nitrification, assimilatory nitrate reduction, and dissimilatory nitrate reduction – DNRA; and for the sulfur cycle: thiosulfate reduction, dissimilatory sulfate reduction, and assimilatory sulfate reduction. Statistically, the differences among treatments were not significant (p > 0.121; Supplementary Table S7), although some conspicuous predicted trends in some of the pathways for specific treatments are worth noting (Figure 6). Nitrogen fixation was predicted to show an elevated trend in seaweed treatments except for Ulva in combination with nutrients, in which the DNRA level seemed elevated (Figure 6). Concerning the sulfur cycle, both dissimilatory and assimilatory sulfate reduction were predicted to show similar relative abundances for all the treatments (Figure 6) and only thiosulfate reduction seemed to be predicted to display a higher relative abundance, of this pathway’s genes, for all the seaweed treatments except for Ulva combined with nutrients (Figure 6). Unfortunately, the PiCRUSt tool does not provide information on SOX genes (coding for thiosulfate oxidation – the main pathway in the sulfur cycle performed by sulfur-oxidizing bacteria), representing a large gap in the interpretation of the putative processes through our experiment.
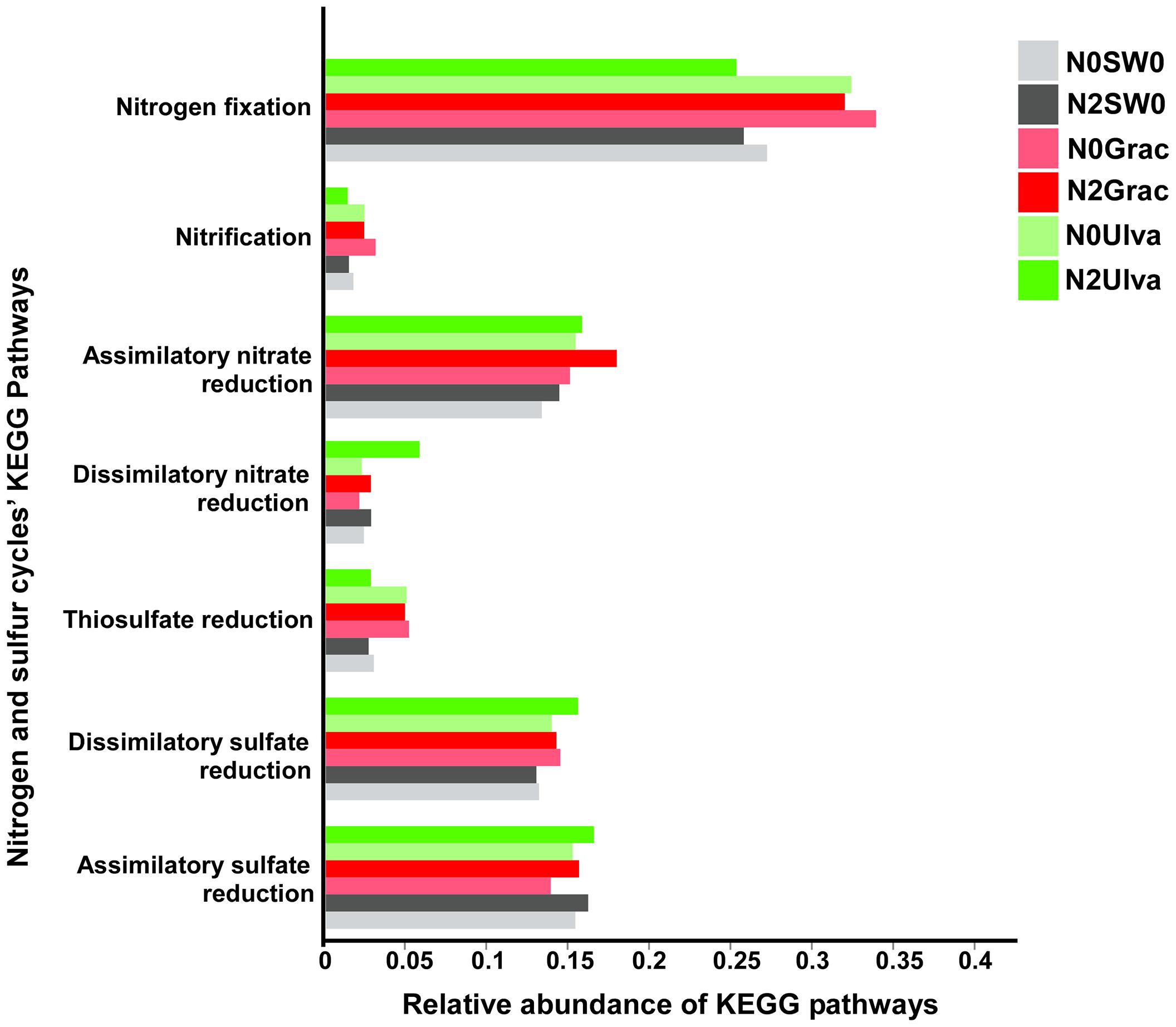
Figure 6. Relative abundance of nitrogen and sulfur cycles KEGG pathways for each treatment (with samples from the same treatment pooled together). Graphic colors were changed so different treatment colors would match those of Figure 1.
At the order level, the closed reference PiCRUSt input table, used for functional inference, corresponded well with the table used for the bacterial community analysis. The readings associated with each of the most abundant orders were analyzed in PiCRUSt, targeting sulfur metabolism (the putative functional category of these orders). The treatment groups followed a similar trend (in terms of relative abundance) when comparing the most abundant orders with the proportion of sequences related to sulfur metabolism (Supplementary Figure S2). For example, Campylobacterales was only conspicuously abundant in the treatments where seaweeds (Ulva) and nutrients were combined (Figure 3 for OTUs, and Figure 4 for order level). The predicted proportion of sulfur metabolism related sequences of Campylobacterales was higher in nutrient addition and combined nutrient and seaweed treatments independently of the seaweed used (p = 0.007; Supplementary Figures S2A,B). Although some Chromatiales were strongly correlated with ambient sediments (vector 3 in Figure 3), the proportion of their predicted sulfur metabolism genes increased with the nutrient addition and they seemed to decrease with the seaweed addition under ambient nutrient conditions (p = 0.0001; Supplementary Figures S2C,D). For Desulfobacterales, the variation in the predicted proportion of sulfur metabolism genes increased with the addition of seaweeds (p = 0.254; Supplementary Figure S2E). As mentioned above, PiCRUSt did not provide information on SOX genes for the family function prediction.
For the carbon-related pathways found in the putative functional profile of PiCRUSt (Supplementary Table S6), we considered the different carbohydrate metabolism and carbon fixation pathways in prokaryotes (Supplementary Table S8). Similar to the nitrogen and sulfur cycles, no differentiation among treatments was detected (PERMANOVA p > 0.7553) with no conspicuous changes noted across treatments (Supplementary Figure S3). It is, however, worth noting that predicted carbon fixation by prokaryotes was the process contributing most to possible differences with an approximate 25% increase in the seaweed treatments, whereas these processes seemed to be decreased upon nutrient addition.
This study provides an important novel contribution to the current understanding of effects of combined nutrient and macroalgal loads on the variation of sediment bacterial communities, having identified the main taxonomical shifts and putative functional differences. Eutrophication episodes are associated with algal blooms, but so far, the cumulative (or not) effects of these two coupled ecosystem disruptors on sediment bacterial communities have barely been addressed. Here, for the first time to our knowledge, both nutrients and algal blooms (including the two natural blooming species) were experimentally manipulated in the field, in order to assess their individual and combined differentiation effects on bacterial communities in intertidal sediments. Our results clearly show that although eutrophication and algal blooms go hand in hand, they predominantly affected sediment bacterial communities differently and independently of one another. This will help us to better understand the impact of eutrophication in its different phases (nutrient increasing and consequent algal bloom) and the differentiated impact of different blooming algal species.
The nutrient load affected the sediment bacterial community by decreasing diversity (alpha diversity) and changing its taxonomic composition (beta diversity), with the latter in agreement with previous studies (e.g., Campbell et al., 2010; Fierer et al., 2012; Guevara et al., 2014; Chen et al., 2015; Lawes et al., 2016; Liu et al., 2018). A low level of microbial diversity/richness is usually associated with an unhealthy and/or disturbed ecosystem (Allison and Martiny, 2008). Eutrophication through increasing nutrients is known to disrupt the local microbial structure through a cascade of processes that leads to unbalanced redox processes (Meyer-Reil and Köster, 2000). A pronounced shift in the microbial structure was evident in treatments where both seaweed and fertilizer were added.
Nutrient and seaweed addition affected the bacterial community composition in sediments independently. The seaweed effect was dependent on the seaweed species used, with Gracilaria causing a more pronounced community shift. Ulva loading resulted in a bacterial community that was not different from no seaweed and Gracilaria addition. However, Ulva in combination with the nutrient addition recorded a marked characteristic shift in the microbial structure.
Bacterial community composition in both sediment treatments, comparing only nutrient loading (without seaweeds), was rather similar, with the top six orders in close proportions and diversity remaining unchanged. This indicates that an increased amount of nutrients leads to bacterial community changes at a finer scale/lower phylogenetic level that is not reflected at higher taxonomic levels. Something similar was found by others authors (Koyama et al., 2014), which suggests the high plasticity of some taxonomic groups under a moderate disturbance. Kearns et al. (2016) also hypothesized that excess N would only favor a small number of taxa that are capable of responding to a nutrient increase.
However, where seaweeds were added, the differences were significant and evident even at the order level, as the addition of either of the seaweeds significantly changed the bacterial community similarly, irrespective of the seaweed species.
The bacterial community composition of the plots where Ulva was added along with nutrients was different when compared to the other seaweed-treated plots (including Ulva without nutrients). Together with the results at the OTU level, where the Ulva addition resulted in an “intermediate” community, this might reflect the different bloom strategies between these two seaweed species, as well as a fast turnover of Ulva biomass. These two fast-growing seaweeds have different blooming strategies, but both are recognized to profit from global eutrophication and its collateral effects as, for example, ocean acidification (Valiela et al., 1997; Young and Gobler, 2016). However, although Gracilaria can also benefit from nutrient increasing by increased growing rates (Ye et al., 2013), Ulva grows faster with high nutrient concentrations (Wallace and Gobler, 2014; Young and Gobler, 2016). Moreover, there are structural differences, where Gracilaria presents a much more robust conformation. These differences and potential repercussion in bacterial communities of the sediments will be discussed further.
Overall, the results showed that the most abundant taxa in all treatments were related to Proteobacteria and to sulfate-reducing and sulfur/sulfide-oxidizing (S-oxidizers) bacteria. This suggests, as predicted, an active sulfur cycle under the sediment conditions of this ecosystem. The vectors plotted and separating the two main “nutrient vs. non-nutrient” clusters suggest an increase of sulfur oxidizers when nutrients are added (Figure 3). This tendency was also followed by the OTUs that contributed most to this difference in similarity percentages analysis. OTUs affiliated to the orders Campylobacterales (denitrifiers) and Chromatiales (both recognized as S-oxidizers) increased in nutrient-loaded sediments (especially those combined with seaweeds). In coastal sediments affected by anoxic and sulfidic conditions, DNRA can be the predominant nitrate reduction pathway (Brunet and Garcia-Gil, 1996; Christensen et al., 2000; McGlathery et al., 2007) in which S-oxidizing bacteria play a crucial role (Christensen et al., 2000; An and Gardner, 2002; Song et al., 2014). DNRA might be favored over denitrification at high organic C loading (Hardison et al., 2015), such as might be the case during joint nutrient and seaweed loading (however, denitrification was using PiCrust not found in our data).
At the order level, nutrient loading alone was not reflected in a changing bacterial community. Two of the most abundant orders, Chromatiales and Thiotrichales (most of them affiliated to Thiotrichaceae family), were also the two orders that slightly increased in abundance when nutrients were added (Figure 3). These two orders are known dominant sulfur oxidizers in salt marsh sediments (Thomas et al., 2014) and are recognized for their potential roles in coupled N and S cycles (Ganesh et al., 2015), which could grant them an advantage when nutrient concentrations increase.
Except for the combination of Ulva and nutrients, all the sediments loaded with seaweeds (regardless of the species) showed a high increase of bacteria belonging to the order Desulfobacterales. These, predominant in seaweed-enriched sediments (Morrison et al., 2017), are sulfate-reducers that oxidize organic compounds and produce hydrogen sulfide as the metabolic end product (Lamers et al., 2013). During algal blooms, the organic matter deposited in the sediments results in intense microbial activity due to the high decomposition rates (Fenchel and Jørgensen, 1977). The upper layer of sediments will become anoxic along with the release of toxic sulfide resulting from the metabolism of these abundant sulfate reducing bacteria. The addition of algal organic matter to the sediment may have become advantageous to this strictly anaerobic, metabolically diverse group of bacteria. These groups also have larger genomes relative to other sulfate-reducing bacteria, which allow a faster response to stronger environmental perturbations and are stimulated by the release of carbon compounds (Kearns et al., 2016).
A puzzling effect was the different response of the nutrient and Ulva combination plots, in which there was an increase in the abundance of Campylobacterales order instead of Desulfobacterales. The order Campylobacterales is the second most abundant in treatments with Gracilaria and nutrients combined, although the difference is three times higher for Ulva. As Campylobacterales (Epsilonproteobacteria) are hardly found on seaweeds (Hollants et al., 2012), it is highly unlikely that their presence in these treatments comes from the seaweeds. In addition, bacterial algal cell wall degraders are from other groups, mainly Alpha- and Gammaproteobacteria and members of the CFB group possessing specific sugar-degrading enzymes such as agarases, carrageenases, and alginases and for Ulva specifically ulvan lyase (Hollants et al., 2012). As mentioned above, these seaweeds have different blooming behaviors and tissue conformation. Both take advantage of high nutrient concentrations, but Ulva grows faster than Gracilaria (Young and Gobler, 2016). This difference in growth rates, allied with a less robust conformation in Ulva, may result in asynchronous decomposition rates and timings, which can be mirrored in the bacterial community of the sediments sampled in the same time frame for all the treatments. Also, due to its higher uptake capacity, Ulva holds a high nutrient content (C:N ratio), usually reflecting environmental increments (Sjöö and Mörk, 2009), which causes it to decompose more rapidly (Teichberg et al., 2010) than Gracilaria (Enriquez et al., 1993). The taxonomic differences found among Gracilaria and Ulva plots do not reflect the differences found between the associated communities of these two seaweeds (Lachnit et al., 2011). Observations during the experiment showed that Ulva was the fastest-disappearing seaweed from the plots. Campylobacterales (here mostly affiliated to the genus Sulfurimonas, see Supplementary Table S1) are mostly sulfide oxidizers commonly found in pelagic marine redox-clines (Campbell et al., 2006). Following Ulva’s faster decomposition and the putative high production of sulfides during the process, the bloom would collapse allowing some oxygenation of the sediments. Accounting for a large percentage of denitrification genes, Campylobacterales may also be responsible for the removal of the nitrates, through their use for sulfide oxidation (Llorens-Marès et al., 2015). Liu et al. (2011) analyzed the bacterial communities in the seawater during Ulva blooms and detected Campylobacterales as the dominant bacterial clade when the bloom was collapsing and the algal material was decaying. This suggests that this group of bacteria might be taking advantage of the high concentration of nutrients and organic matter in decomposition.
The order Bacteroidales also increased in abundance in the plots where seaweeds were added independently of their nutrient status. Members of this order include efficient decomposers/digesters of dead organic matter, which are usually found in sludge, contributing to the degradation of proteins (Rivière et al., 2009) and have been found in seaweed enriched sediments (Morrison et al., 2017). Coupled with their decomposition skills, Bacteroidales, are also able to couple the electron flow from organic matter to reduce nitrate to ammonium (Llorens-Marès et al., 2015).
It is also important to mention that the order Clostridiales was among the most abundant orders for Gracilaria enriched treatments and was also present (although much less abundant) in no-nutrient Ulva enriched sediments. Clostridiales are strict anaerobes, known to mediate the degradation of complex carbohydrate polymers with the capacity for cellulose degradation (Vanwonterghem et al., 2014). They have been found associated with fermentation of recalcitrant plant cell wall polysaccharides and have been found in environments with high plant biomass turnover rates (van der Lelie et al., 2012). The elevated presence of this order in seaweed-enriched sediments, with a notably higher proportion in Gracilaria plots, is in line with the high amount of organic matter in these plots, and the harder tissue structure and the stronger burial survival of Gracilaria compared to Ulva. Gracilaria is, in general, also (partly) buried in the mud in the Ria Formosa, which might actually stimulate anaerobic conditions required for Clostridiales members. G. vermiculophylla is highly resistant to burial, desiccation, and grazing and growth is not very dependent on nutrient levels (Thomsen and McGlathery, 2007), which might explain the similar behavior of bacterial communities under both nutrient treatments within the Gracilaria addition and the strong differences with Ulva enrichment treatments. The fact that this order was not detected in the treatment where Ulva was added along with nutrients, is consistent with the hypothesis of an already advanced decomposition in these plots. In future experiments, physicochemical and geochemical parameters should be monitored continuously (e.g., oxygen levels, C:N ratios, decomposition rates, organic matter content) to assess possible relationships with the bacterial community guilds.
PiCRUSt can be used as an exploratory tool to help to decide on a more expensive high-depth metagenomic sequencing approach, due to the demonstrated consistency in predicting functional diversity based on phylogenetic affiliation (e.g., Staley et al., 2014), with good results demonstrated for soil and sediments (Langille et al., 2013). However, being aware of these prediction limitations and possible flaws, as well as the controversy around the use of this tool, this discussion will be general and limited to raising pertinent hypotheses. Therefore, we only focused on candidate pathways from the sulfur cycle, coupled with the nitrogen cycle, which is one of the most important biogeochemical cycles in coastal sediments (Rysgaard et al., 1996).
Most of the work done so far suggests that, as a result of disturbance, sediment bacterial communities change their functional profile along with community structure (as reviewed by Allison and Martiny, 2008). Most recently it has been found that, even when bacterial composition remains unaltered through community resilience, the functional capacity is adapted by a shift in the portion of actually active bacteria (Kearns et al., 2016).
Statistical differences found for predicted metagenomes are more comparable with those achieved with the main order levels than with the OTU level. The overall (all the pathways considered) functional profiles predictions are distinct for seaweed-enriched sediments but do not differ between treatments with the seaweed species. However, significance in seaweed treatments may reflect functional instability among treatments, related to the different decomposition timings discussed above. Functional profiles of chosen candidate pathways show that the number of reading assigned to the sulfur cycle increased in the nutrient treatments and, more sharply, when seaweeds are combined. Sulfur cycle stimulation has been reported in coastal lagoons, particularly sulfate reduction rates in anoxic conditions caused by organic enrichments (Holmer et al., 2004; Duarte et al., 2005). Seaweed-enriched sediments can have an upregulation of dsr genes, which encode proteins responsible for dissimilatory sulfate reduction (Morrison et al., 2017). The readings assigned to the nitrogen cycle followed a similar pattern as those of the sulfur cycle. Seaweed addition is likely the determinant factor and its effect is enhanced when combined with nutrients. These two cycles are coupled and their dynamics allows the efficiency of this ecosystem service. However, as the amount of sulfide can be six times higher than that of ammonium in coastal lagoons, the abundance of sulfur-related genes is expected to be higher (Laverock et al., 2011), in agreement with our results. Sulfate reduction is considered the most important anaerobic respiratory process occurring in coastal lagoons (Caumette et al., 1996).
Zooming into sulfur and nitrogen cycles specific pathways corroborates some of the hypotheses advanced for the bacterial community taxonomical shifts. The relative abundances of reads assigned to DNRA were higher in the treatment where Ulva was combined with nutrients. The lack of SOX genes in the PiCRUSt database did not allow us to infer sulfur oxidation activity, although it is known that sulfur oxidation is one of the pathways used by some DNRA species (Burgin and Hamilton, 2007). The higher abundance of the sulfur-oxidizers/denitrifiers – Campylobacterales- in this treatment, probably reflects the increase of these processes in the N2Ulva treatment. In contrast, nitrogen-fixation is higher in all seaweed treatments except for N2Ulva. An experiment in nitrogen-rich coastal surface waters demonstrated a higher abundance of nifH genes in surface waters covered with massive macroalgal canopies (Zhang et al., 2015). Surprisingly, the relative abundance of reads putatively assigned to sulfate-reduction (dissimilatory and assimilatory) was equivalent for all treatments. However, those putatively assigned to thiosulfate-reduction (also performed by sulfate-reducing bacteria) seemed higher in seaweed-treated sediments (except N2Ulva), which is in line with the higher abundance of sulfate-reducing bacteria (Desulfobacterales) in these treatments.
In the Carbon cycling category, the relative abundance of sequences associated with carbon-fixation pathways in prokaryotes slightly increased in the seaweed-treated plots. That might be related to the seaweed degradation in the seaweed-added plots and the consequent fixation of the released CO2 by sediment bacterial community. This study constitutes a scaffold to be considered in further analyses targeting specific genomic and/or transcriptomic pathways that are required to validate our hypothesis. These analyses, paired with physical/geochemical data, would allow a more complete picture of the taxonomical and functional shifting in sediments under coupled environmental stress.
In this experimental field study, nutrient and seaweed levels were not kept constant throughout the experimental duration. The use of slow-release fertilizer and a pulse loading of seaweeds aimed to simulate natural field conditions of this lagoon. Seaweed blooms on intertidal banks are variable both in time and space due to the ecological processes, but also due to a to lack of hard substrate for fixation and to variable tidal currents. After 5 weeks at the point of sampling, most seaweed biomass was lost from the experimental plots and, as such, our results mostly represent the post-bloom conditions. Although effects of nutrient and seaweed addition were experimentally tested, we did not assess the temporal changes in bacterial communities, which likely changed during the duration of the experiment. In addition, the effects documented are a combination of indistinguishable direct and indirect treatment effects on the entire ecosystem present. Both treatments may not only affect the bacterial communities directly but probably also affect eukaryotic fauna in and on sediments. Infauna activity, for example, directly affects the oxygenations of sediments by bioturbation, biogeochemical cycling, as well as the processing of seaweed biomass, and this has consequences for the bacterial community structure and function in sediments (Herbert, 1999). However, the objective was not to identify the mechanisms, but rather to assess the net effects of nutrient and seaweed addition on bacterial communities in the field. Despite all this natural variability that takes place in field conditions, the experimental design ensures that the differences between treatments can be attributed to the factors tested, and not to other sources of variability that were affecting all plots equally.
Although we are very cautious in the use of PICRUSt for providing functional insights from our nonhuman model systems, the low NSTI values and the good match between the main taxonomical and predictive functional differences provide confidence in the used approach in this study. Gene expression data would have provided more accurate functional insights but would probably also be more specific for the time and conditions of sampling.
Environmental metadata could have provided us with clues regarding the specific environmental changes that our treatments would have caused, concerning nutrient conditions for example. Rather than multiple samplings across the duration of the experiments to adjust for strong spatial variability in intertidal environments, we opted for a more time-integrated approach using the C:N ratios of macrophyte biomass in each plot. Unfortunately, the obtained biomass showed no differences among treatments, despite documented internal N pool and environmental dissolved inorganic nitrogen concentrations (Teichberg et al., 2010). We believe this is more a consequence of the used method, partly due to the turnover rate of biomass during the experimental 5 weeks than the treatment considering the strong effects on the bacterial community. Redox, pH, oxygen, and sulfide electrode profiles would have been extremely valuable to correlate with community changes.
Eutrophication and macroalgal blooms are challenging for coastal marine management. Although both lead to elevated microbial activity resulting in anoxia with ecosystem and social implications, this study demonstrated that nutrient and seaweed enrichment drive microbial communities in sediment into different compositions and structure. Algal blooms could be considered to be nature’s solution for buffering short term eutrophication by transferring inorganic into organic and subsequently “slowly” recycling these compounds into the system by different microbial communities specialized in degrading complex compounds. Overall, however, it seems that the natural solution in the form of macroalgal blooms has a larger impact on sediment microbial communities than nutrient loading does. For coastal management, our findings show that reducing eutrophication sources will be most effective. Macroalgae can be effectively used for nutrient capturing, but they should be harvested in time. In addition, introduced Gracilaria seems to affect sediment microbial community composition and functioning, as well as biogeochemical cycling, differently from blooms of native Ulva. Sediment microbial communities could provide considerable information as indicators of ecosystem status, similarly to macro-organism indices.
Both nutrient and seaweed additions to sediments have the net capacity to change (directly or indirectly) the microbial structure of coastal sediments and act mostly independently of one another. Seaweed addition effects were more pronounced than nutrient addition effects and were species-dependent. The most important bacterial community changes (regarding taxa abundance) occurred among bacteria involved in nitrogen and carbon, but especially in sulfur cycling.
AE designed the study and was responsible for the field experiment execution and final sampling. TA extracted DNA, amplified 16S rRNA genes and performed all in silico analysis. TA, GM, ES, and AE drafted the manuscript. All authors approved the final manuscript.
This study received Portuguese national funds from FCT – Foundation for Science and Technology through projects UID/Multi/04326/2019 and /2016, Seas-Era project grant “Invasea” (SEAS-ERA/0001/2012) and scholarships to AE (SFRH/BPD/107878/2015) and TA (SFRH/BPD/116774/2016). GM was financially supported by the research priority area Systems Biology of the University of Amsterdam.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer LK and handling Editor declared their shared affiliation.
We thank all those who contributed directly or indirectly to this work, especially Geert Akkermans for initiating this experiment and maintaining the conditions, RTL for all post-sequencing hints and tips, and João Neiva and two reviewers for their comments and suggestions.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.03283/full#supplementary-material
Allison, S. D., and Martiny, J. B. H. (2008). Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. U.S.A. 105, 11512–11519. doi: 10.1073/pnas.0801925105
An, S., and Gardner, W. S. (2002). Dissimilatory nitrate reduction to ammonium (DNRA) as a nitrogen link, versus denitrification as a sink in a shallow estuary (Laguna Madre/Baffin Bay, Texas). Mar. Ecol. Prog. Ser. 237, 41–50. doi: 10.3354/meps237041
Anderson, D. M., Cembella, A. D., and Hallegraeff, G. M. (2012). Progress in understanding harmful algal blooms: paradigm shifts and new technologies for research, monitoring, and management. Ann. Rev. Mar. Sci. 4, 143–176. doi: 10.1146/annurev-marine-120308-081121
Anderson, M. J., Ellingsen, K. E., and McArdle, B. H. (2006). Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 9, 683–693. doi: 10.1111/j.1461-0248.2006.00926.x
Borsodi, A. K., Makk, J., Moln, P., and Andrea, M. (2003). Algological and bacteriological investigations on reed periphyton in Lake Velencei, Hungary. Hydrobiologia 506-509, 549–557. doi: 10.1023/b:hydr.0000008591.89558.75
Bowen, J. L., Crump, B. C., Deegan, L. A., and Hobbie, J. E. (2009). Salt marsh sediment bacteria: their distribution and response to external nutrient inputs. ISME J. 3, 924–934. doi: 10.1038/ismej.2009.44
Bowen, J. L., Ward, B. B., Morrison, H. G., Hobbie, J. E., Valiela, I., Deegan, L. A., et al. (2011). Microbial community composition in sediments resists perturbation by nutrient enrichment. ISME J. 5, 1540–1548. doi: 10.1038/ismej.2011.22
Brin, L. D., Valiela, I., Goehringer, D., and Howes, B. (2010). Nitrogen interception and export by experimental salt marsh plots exposed to chronic nutrient addition. Mar. Ecol. Prog. Ser. 400, 3–17. doi: 10.3354/meps08460
Brune, A., Frenzel, P., and Cypionka, H. (2000). Life at the oxic - anoxic interface: microbial activities and adaptations. FEMS Microbiol. Rev. 24, 691–710. doi: 10.1111/j.1574-6976.2000.tb00567.x
Brunet, R. C., and Garcia-Gil, L. J. (1996). Sulfide-induced dissimilatory nitrate reduction to ammonia in anaerobic freshwater sediments. FEMS Microbiol. Ecol. 21, 131–138. doi: 10.1016/0168-6496(96)00051-7
Burgin, A. J., and Hamilton, S. K. (2007). Have we overemphasized the role of denitrification in aquatic ecosystems? A review of nitrate removal pathways. Front. Ecol. Environ. 5, 89–96. doi: 10.1890/1540-9295(2007)5[89:HWOTRO]2.0.CO;2
Burkholder, J. M., Tomasko, D. A., and Touchette, B. W. (2007). Seagrasses and eutrophication. J. Exp. Mar. Biol. Ecol. 350, 46–72. doi: 10.1016/j.jembe.2007.06.024
Campbell, B. J., Engel, A. S., Porter, M. L., and Takai, K. (2006). The versatile 𝜖-proteobacteria: key players in sulphidic habitats. Nat. Rev. Microbiol. 4, 458–468. doi: 10.1038/nrmicro1414
Campbell, B. J., Polson, S. W., Hanson, T. E., Mack, M. C., and Schuur, E. A. (2010). The effect of nutrient deposition on bacterial communities in Arctic tundra soil. Environ. Microbiol. 12, 1842–1854. doi: 10.1111/j.1462-2920.2010.02189.x
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). Correspondence QIIME allows analysis of high- throughput community sequencing data Intensity normalization improves color calling in SOLiD sequencing. Nat. Publ. Gr. 7, 335–336. doi: 10.1038/nmeth0510-335
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Huntley, J., Fierer, N., et al. (2012). Ultra-high-throughput microbial community analysis on the illumina HISEQ and MISEQ platforms. ISME J. 6, 1621–1624. doi: 10.1038/ismej.2012.8
Cardoso, P. G., Pardal, M. A., Raffaelli, D., Baeta, A., and Marques, J. C. (2004). Macroinvertebrate response to different species of macroalgal mats and the role of disturbance history. J. Exp. Mar. Bio. Ecol. 308, 207–220. doi: 10.1016/j.jembe.2004.02.018
Caumette, P., Castel, J., and Herbert, R. A. (1996). “Coastal Lagoon Eutrophication and Anaerobic Processes (C.L.E.A.N.): Nitrification, denitrification, and nitrate ammonification in sediments of two coastal lagoons in Southern France,” in Developments in Hydrobiology (Dordrecht: Kluwer Academic Publishers), 133–141.
Chen, N., Yang, J.-S., Qu, J.-H., Li, H.-F., Liu, W.-J., Li, B.-Z., et al. (2015). Sediment prokaryote communities in different sites of eutrophic Lake Taihu and their interactions with environmental factors. World J. Microbiol. Biotechnol. 31, 883–896. doi: 10.1007/s11274-015-1842-1
Christensen, P. B., Rysgaard, S., Sloth, N. P., Dalsgaard, T., and Schwærter, S. (2000). Sediment mineralization, nutrient fluxes, denitrification and dissimilatory nitrate reduction to ammonium in an estuarine fjord with sea cage trout farms. Aquat. Microb. Ecol. 21, 73–84. doi: 10.3354/ame021073
Chróst, R. J., and Siuda, W. (2006). Microbial production, utilization, and enzymatic degradation of organic matter in the upper trophogenic layer in the pelagial zone of lakes along a eutrophication gradient. Limnol. Oceanogr. 51, 749–762. doi: 10.4319/lo.2006.51.1_part_2.0749
Clarke, K. R., and Warwick, R. M. (1994). Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. Plymouth: PRIMER-E.
Duarte, C. M., Holmer, M., and Marbà, N. (2005). “Plant–microbe interactions in seagrass meadows,” in Interactions Between Macro- and Microorganisms in Marine Sediments, eds E. Kristensen, R. R. Haese, and J. E. Kostka (Washington, DC: American Geophysical Union), 31–60.
Engelen, A. H., Henriques, N., Monteiro, C., and Santos, R. (2011). Mesograzers prefer mostly native seaweeds over the invasive brown seaweed Sargassum muticum. Hydrobiologia 669, 157–165. doi: 10.1007/s10750-011-0680-x
Enriquez, S., Duarte, C., and Sand-Jensen, K. (1993). Patterns in decomposition rates among photosynthetic organisms: the importance of detritus C: NP content. Oecologia 94, 457–471. doi: 10.1007/BF00566960
Fenchel, T. M., and Jørgensen, B. B. (1977). “Detritus food chains of aquatic ecosystems: the role of bacteria,” in Advances in Microbial Ecology, ed. M. Alexander (Boston, MA: Springer), 1–58. doi: 10.1007/978-1-4615-8219-9_1
Fierer, N., Lauber, C. L., Ramirez, K. S., Zaneveld, J., Bradford, M. A., and Knight, R. (2012). Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 6, 1007–1017. doi: 10.1038/ismej.2011.159
Fletcher, R. L. (1996). “The occurrence of green tides - a review,” in Marine Benthic Vegetation: Recent Changes and the Effects of Eutrophication, eds W. Schramm and P. H. Nienhuis (Berlin: Springer), 7–43. doi: 10.1007/978-3-642-61398-2_2
Galloway, J. N., and Cowling, E. B. (2002). Reactive nitrogen and the world: 200 years of change. Ambio J. Hum. Environ. 31, 64–71. doi: 10.1579/0044-7447-31.2.64
Ganesh, S., Bristow, L. A., Larsen, M., Sarode, N., Thamdrup, B., and Stewart, F. J. (2015). Size-fraction partitioning of community gene transcription and nitrogen metabolism in a marine oxygen minimum zone. ISME J. 9, 2682–2696. doi: 10.1038/ismej.2015.44
Gao, G., Clare, A. S., Rose, C., and Caldwell, G. S. (2016). Eutrophication and warming-driven green tides (Ulva rigida) are predicted to increase under future climate change scenarios. Mar. Pollut. Bull. 114, 439–447. doi: 10.1016/j.marpolbul.2016.10.003
Grossart, H. P., Levold, F., Allgaier, M., Simon, M., and Brinkhoff, T. (2005). Marine diatom species harbour distinct bacterial communities. Environ. Microbiol. 7, 860–873. doi: 10.1111/j.1462-2920.2005.00759.x
Guevara, R., Ikenaga, M., Dean, A. L., Pisani, C., and Boyer, J. N. (2014). Changes in sediment bacterial community in response to long-term nutrient enrichment in a subtropical seagrass-dominated estuary. Microb. Ecol. 68, 427–440. doi: 10.1007/s00248-014-0418-1
Hale, S. S., Cicchetti, G., and Deacutis, C. F. (2016). Eutrophication and hypoxia diminish ecosystem functions of benthic communities in a New England estuary. Front. Mar. Sci. 3:249. doi: 10.3389/fmars.2016.00249
Hardison, A. K., Algar, C. K., Giblin, A. E., and Rich, J. J. (2015). Influence of organic carbon and nitrate loading on partitioning between dissimilatory nitrate reduction to ammonium (DNRA) and N2 production. Geochim. Cosmochim. Acta 164, 146–160. doi: 10.1016/j.gca.2015.04.049
Hartmann, M., Frey, B., Mayer, J., Mäder, P., and Widmer, F. (2015). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. doi: 10.1038/ismej.2014.210
Hauxwell, J., Cebrian, J., Furlong, C., and Valiela, I. (2001). Macroalgal canopies contribute to eelgrass (Zostera marina) decline in temperate estuarine ecosystems. Ecology 82, 1007–1022. doi: 10.1890/0012-96582001082
Herbert, R. A. (1999). Nitrogen cycling in coastal marine ecosystems. FEMS Microbiol. Rev. 23, 563–590. doi: 10.1111/j.1574-6976.1999.tb00414.x
Holland, E. A., Braswell, B. H., Sulzman, J., and Lamarque, J.-F. (2005). Nitrogen deposition onto the United States and Western Europe: synthesis of observations and models. Ecol. Appl. 15, 38–57. doi: 10.1890/03-5162
Hollants, J., Leliaert, F., De Clerck, O., and Willems, A. (2012). What can we learn from sushi: a review on seaweed-bacterial associations. FEMS Microbiol. Ecol. 83, 1–16. doi: 10.1111/j.1574-6941.2012.01446.x
Holmer, M., Duarte, C. M., Boschker, H. T. S., and Barrón, C. (2004). Carbon cycling and bacterial carbon sources in pristine and impacted Mediterranean seagrass sediments. Aquat. Microb. Ecol. 36, 227–237. doi: 10.3354/ame036227
Holmer, M., and Nielsen, R. M. (2007). Effects of filamentous algal mats on sulphide invasion in eelgrass (Zostera marina). J. Exp. Mar. Bio. Ecol. 353, 245–252. doi: 10.1016/j.jembe.2007.09.010
Howarth, R., Chan, F., Conley, D. J., Garnier, J., Doney, S. C., Marino, R., et al. (2011). Coupled biogeochemical cycles: eutrophication and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 9, 18–26. doi: 10.1890/100008
Isbell, F., Reich, P. B., Tilman, D., Hobbie, S. E., Polasky, S., and Binder, S. (2013). Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity. Proc. Natl. Acad. Sci. U.S.A. 110, 11911–11916. doi: 10.1073/pnas.1310880110
Ishida, Y., Yoshinaga, I., Kim, M. C., and Uchida, A. (1997). “Possibility of bacterial control of harmful algal blooms,” in Progress in Microbial Ecology, eds P. S. M. T. Martins, J. M. Tiedje, L. C. N. Hagler, and J. Dobereiner (Brazil: SBM),495–500.
Jones, K. L., Mikulski, C. M., Barnhorst, A., and Doucette, G. J. (2010). Comparative analysis of bacterioplankton assemblages from Karenia brevis bloom and nonbloom water on the west Florida shelf (Gulf of Mexico, USA) using 16S rRNA gene clone libraries. FEMS Microbiol. Ecol. 73, 468–485. doi: 10.1111/j.1574-6941.2010.00914.x
Kearns, P. J., Angell, J. H., Howard, E. M., Deegan, L. A., Stanley, R. H., and Bowen, J. L. (2016). Nutrient enrichment induces dormancy and decreases diversity of active bacteria in salt marsh sediments. Nat. Commun. 7:12881. doi: 10.1038/ncomms12881
Kodama, M., Doucette, G. J., and Green, D. H. (2006). Relationships between bacteria and harmful algae. Ecol. Harmful Algae 189, 243–255. doi: 10.1007/978-3-540-32210-8_19
Koyama, A., Wallenstein, M. D., Simpson, R. T., and Moore, J. C. (2014). Soil bacterial community composition altered by increased nutrient availability in arctic tundra soils. Front. Microbiol. 5:516. doi: 10.3389/fmicb.2014.00516
Kozich, J. J., Westcott, S. L., Baxter, N. T., Highlander, S. K., and Schloss, P. D. (2013). Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 79, 5112–5120. doi: 10.1128/AEM.01043-13
Lachnit, T., Meske, D., Wahl, M., Harder, T., and Schmitz, R. (2011). Epibacterial community patterns on marine macroalgae are host-specific but temporally variable. Environ. Microbiol. 13, 655–665. doi: 10.1111/j.1462-2920.2010.02371.x
Lamers, L. P., Govers, L. L., Janssen, I. C., Geurts, J. J., Van der Welle, M. E., Van Katwijk, M. M., et al. (2013). Sulfide as a soil phytotoxin-a review. Front. Plant Sci. 4:268. doi: 10.3389/fpls.2013.00268
Langille, M., Zaneveld, J., Caporaso, J. G., McDonald, D., Knights, D., Reyes, J., et al. (2013). Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 31, 814–821. doi: 10.1038/nbt.2676
Laverock, B., Gilbert, J. A., Tait, K., Osborn, A. M., and Widdicombe, S. (2011). Bioturbation: impact on the marine nitrogen cycle. Biochem. Soc. Trans. 39, 315–320. doi: 10.1042/BST0390315
Lawes, J. C., Neilan, B. A., Brown, M. V., Clark, G. F., and Johnston, E. L. (2016). Elevated nutrients change bacterial community composition and connectivity: high throughput sequencing of young marine biofilms. Biofouling 32, 57–69. doi: 10.1080/08927014.2015.1126581
Lee, P. O., McLellan, S. L., Graham, L. E., and Young, E. B. (2015). Invasive dreissenid mussels and benthic algae in Lake Michigan: characterizing effects on sediment bacterial communities. FEMS Microbiol. Ecol. 91, 1–12. doi: 10.1093/femsec/fiu001
Leff, J. W., Jones, S. E., Prober, S. M., Barberán, A., Borer, E. T., Firn, J. L., et al. (2015). Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. U.S.A. 112, 10967–10972. doi: 10.1073/pnas.1508382112
Leliaert, F., Zhang, X., Ye, N., Malta, E. J., Engelen, A. H., Mineur, F., et al. (2009). Research note: identity of the Qingdao algal bloom. Phycol. Res. 57, 147–151. doi: 10.1111/j.1440-1835.2009.00532.x
Liu, M., Dong, Y., Zhao, Y., Zhang, G., Zhang, W., and Xiao, T. (2011). Structures of bacterial communities on the surface of Ulva prolifera and in seawaters in an Ulva blooming region in Jiaozhou Bay, China. World J. Microbiol. Biotechnol. 27, 1703–1712. doi: 10.1007/s11274-010-0627-9
Liu, S., Jiang, Z., Deng, Y., Wu, Y., Zhang, J., Zhao, C., et al. (2018). Effects of nutrient loading on sediment bacterial and pathogen communities within seagrass meadows. Microbiologyopen 7:e600. doi: 10.1002/mbo3.600
Llorens-Marès, T., Yooseph, S., Goll, J., Hoffman, J., Vila-Costa, M., Borrego, C. M., et al. (2015). Connecting biodiversity and potential functional role in modern euxinic environments by microbial metagenomics. ISME J. 9, 1648–1661. doi: 10.1038/ismej.2014.254
Machás, R., Santos, R., and Peterson, B. (2006). Elemental and stable isotope composition of Zostera noltii (Horneman) leaves during the early phases of decay in a temperate mesotidal lagoon. Estuar. Coast. Shelf Sci. 66, 21–29. doi: 10.1016/j.ecss.2005.07.018
McDonald, D., Price, M. N., Goodrich, J., Nawrocki, E. P., DeSantis, T. Z., Probst, A., et al. (2012). An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 6, 610–618. doi: 10.1038/ismej.2011.139
McGlathery, K. J., Sundbäck, K., and Anderson, I. C. (2007). Eutrophication in shallow coastal bays and lagoons: the role of plants in the coastal filter. Mar. Ecol. Prog. Ser. 348, 1–18. doi: 10.3354/meps07132
Meyer-Reil, L. A., and Köster, M. (2000). Eutrophication of marine waters: effects on benthic microbial communities. Mar. Pollut. Bull. 41, 255–263. doi: 10.1016/S0025-326X(00)00114-4
Micsinai, A., Borsodi, A. K., Csengeri, V., Horváth, A., Oravecz, O., Nikolausz, M., et al. (2003). Rhizome-associated bacterial communities of healthy and declining reed stands in Lake Velencei, Hungary. Hydrobiologia 506–509, 707–713. doi: 10.1023/b:hydr.0000008590.37567.fa
Morrison, J. M., Murphy, C. L., Baker, K., Elshahed, M. S., Corresp, N. H. Y., Zamor, R., et al. (2017). Microbial communities mediating algal detritus turnover under anaerobic conditions. PeerJ 5:e2803. doi: 10.7717/peerj.2803
Nixon, S. W. (1995). Coastal marine eutrophication: a definition, social causes, and future concerns. Ophelia 41, 199–219. doi: 10.1080/00785236.1995.10422044
Parks, D. H., Tyson, G. W., Hugenholtz, P., and Beiko, R. G. (2014). STAMP: statistical analysis of taxonomic and functional profiles. Bioinformatics 30, 3123–3124. doi: 10.1093/bioinformatics/btu494
Reynolds, P. L., Stachowicz, J. J., Hovel, K., Bostrom, C., Boyer, K., Cusson, M., et al. (2018). Latitude, temperature, and habitat complexity predict predation pressure in eelgrass beds across the Northern Hemisphere. Ecology 99, 29–35. doi: 10.1002/ecy.2064
Rivière, D., Desvignes, V., Pelletier, E., Chaussonnerie, S., Guermazi, S., Weissenbach, J., et al. (2009). Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge. ISME J. 3, 700–714. doi: 10.1038/ismej.2009.2
Rooney-Varga, J. N., Giewat, M. W., Savin, M. C., Sood, S., Legresley, M., and Martin, J. L. (2005). Links between phytoplankton and bacterial community dynamics in a coastal marine environment. Microb. Ecol. 49, 163–175. doi: 10.1007/s00248-003-1057-0
Rysgaard, S., Risgaard-Petersen, N., and Sloth, N. P. (1996). “Nitrification, denitrification, and nitrate ammonification in sediments of two coastal lagoons in Southern France,” in Coastal Lagoon Eutrophication and ANaerobic Processes (C.L.E.AN.): Nitrogen and Sulfur Cycles and Population Dynamics in Coastal Lagoons A Research Programme of the Environment Programme of the EC (DG XII), eds P. Caumette, J. Castel, and R. Herbert (Dordrecht: Springer), 133–141. doi: 10.1007/978-94-009-1744-6_11
Sanz-Lázaro, C., Valdemarsen, T., and Holmer, M. (2015). Effects of temperature and organic pollution on nutrient cycling in marine sediments. Biogeosciences 12, 4565–4575. doi: 10.5194/bg-12-4565-2015
Sinkko, H., Lukkari, K., Sihvonen, L. M., Sivonen, K., Leivuori, M., Rantanen, M., et al. (2013). Bacteria contribute to sediment nutrient release and reflect progressed eutrophication-driven hypoxia in an organic-rich continental sea. PLoS One 8:e67061. doi: 10.1371/journal.pone.0067061
Sjöö, G. L., and Mörk, E. (2009). Tissue nutrient content in Ulva spp. (Chlorophyceae) as bioindicator for nutrient loading along the coast of East Africa. Open Environ. Biol. Monit. J. 2, 11–17. doi: 10.2174/1875040000902010011
Song, B., Lisa, J. A., and Tobias, C. R. (2014). Linking DNRA community structure and activity in a shallow lagoonal estuarine system. Front. Microbiol. 5:460. doi: 10.3389/fmicb.2014.00460
Spivak, A. C., and Ossolinski, J. (2016). Limited effects of nutrient enrichment on bacterial carbon sources in salt marsh tidal creek sediments. Mar. Ecol. Prog. Ser. 544, 107–130. doi: 10.3354/meps11587
Spring, S., Schulze, R., Overmann, J., and Schleifer, K. H. (2000). Identification and characterization of ecologically significant prokaryotes in the sediment of freshwater lakes: molecular and cultivation studies. FEMS Microbiol. Rev. 24, 573–590. doi: 10.1016/S0168-6445(00)00046-2
Staley, C., Gould, T. J., Wang, P., Phillips, J., Cotner, J. B., and Sadowsky, M. J. (2014). Core functional traits of bacterial communities in the Upper Mississippi River show limited variation in response to land cover. Front. Microbiol. 5:414. doi: 10.3389/fmicb.2014.00414
Tallis, H., Ferdaña, Z., and Gray, E. (2008). Linking terrestrial and marine conservation planning and threats analysis. Conserv. Biol. 22, 120–130. doi: 10.1111/j.1523-1739.2007.00861.x
Taylor, D., Nixon, S., Granger, S., and Buckley, B. (1995). Nutrient limitation and the eutrophication of coastal lagoons. Mar. Ecol. Prog. Ser. 127, 235–244. doi: 10.3354/meps127235
Teichberg, M., Fox, S. E., Olsen, Y. S., Valiela, I., Martinetto, P., Iribarne, O., et al. (2010). Eutrophication and macroalgal blooms in temperate and tropical coastal waters: nutrient enrichment experiments with Ulva spp. Glob. Chang. Biol. 16, 2624–2637. doi: 10.1111/j.1365-2486.2009.02108.x
Thamdrup, B. (2012). New pathways and processes in the global nitrogen cycle. Annu. Rev. Ecol. Evol. Syst. 43, 407–428. doi: 10.1146/annurev-ecolsys-102710-145048
Thomas, F., Giblin, A. E., Cardon, Z. G., and Sievert, S. M. (2014). Rhizosphere heterogeneity shapes abundance and activity of sulphur-oxidizing bacteria in vegetated salt marsh sediments. Front. Microbiol. 5:309. doi: 10.3389/fmicb.2014.00309
Thomsen, M. S. (2010). Experimental evidence for positive effects of invasive seaweed on native invertebrates via habitat-formation in a seagrass bed. Aquat. Invasions 5, 341–346. doi: 10.3391/ai.2010.5.4.02
Thomsen, M. S., and McGlathery, K. J. (2007). Stress tolerance of the invasive macroalgae Codium fragile and Gracilaria vermiculophylla in a soft-bottom turbid lagoon. Biol. Invasions 9, 499–513. doi: 10.1007/s10530-006-9043-3
Tiedje, J. M. (1988). “Ecology of denitrification and dissimilatory nitrate reduction to ammonium,” in Biology of Anaerobic Microorganisms, ed. A. J. B. Zehnder (New York, NY: John Wiley and Sons), 179–244.
Valiela, I., Mcclelland, J., Hauxwell, J., Behr, P. J., Hersh, D., and Foreman, K. (1997). Macroalgal blooms in shallow estuaries: controls and ecophysiological and ecosystem consequences. Limnol. Oceanogr. 42, 1105–1118. doi: 10.4319/lo.1997.42.5_part_2.1105
van der Lelie, D., Taghavi, S., McCorkle, S. M., Li, L. L., Malfatti, S. A., Monteleone, D., et al. (2012). The metagenome of an anaerobic microbial community decomposing poplar wood chips. PLoS One 7:e36740. doi: 10.1371/journal.pone.0036740
Vanwonterghem, I., Jensen, P. D., Dennis, P. G., Hugenholtz, P., Rabaey, K., and Tyson, G. W. (2014). Deterministic processes guide long-term synchronised population dynamics in replicate anaerobic digesters. ISME J. 8, 2015–2028. doi: 10.1038/ismej.2014.50
Wallace, R. B., and Gobler, C. J. (2014). Factors controlling blooms of microalgae and macroalgae (Ulva rigida) in a eutrophic, urban estuary: Jamaica Bay, NY, USA. Estuaries Coast. 38, 519–533. doi: 10.1007/s12237-014-9818-1
Wetzel, R. G. (2000). Freshwater ecology: changes, requirements, and future demands. Limnology 1, 3–9. doi: 10.1007/s102010070023
Xiong, J., Ye, X., Wang, K., Chen, H., Hu, C., Zhu, J., et al. (2014). Biogeography of the sediment bacterial community responds to a nitrogen pollution gradient in the East China sea. Appl. Environ. Microbiol. 80, 1919–1925. doi: 10.1128/AEM.03731-13
Ye, C. P., Zhang, M. C., Zhao, J. G., Yang, Y. F., and Zuo, Y. (2013). Photosynthetic response of the macroalga, Gracilaria lemaneiformis (Rhodophyta), to various N and P levels at different temperatures. Int. Rev. Hydrobiol. 98, 245–252. doi: 10.1002/iroh.201301435
Young, C. S., and Gobler, C. J. (2016). Ocean acidification accelerates the growth of two bloom-forming macroalgae. PLoS One 11:e0155152. doi: 10.1371/journal.pone.0155152
Zhang, X., Song, Y., Liu, D., Keesing, J. K., and Gong, J. (2015). Macroalgal blooms favor heterotrophic diazotrophic bacteria in nitrogen-rich and phosphorus-limited coastal surface waters in the Yellow Sea. Estuar. Coast. Shelf Sci. 163, 75–81. doi: 10.1016/j.ecss.2014.12.015
Keywords: marine sediments, microbiome, 16S amplicon sequencing, eutrophication, algal bloom
Citation: Aires T, Muyzer G, Serrão EA and Engelen AH (2019) Seaweed Loads Cause Stronger Bacterial Community Shifts in Coastal Lagoon Sediments Than Nutrient Loads. Front. Microbiol. 9:3283. doi: 10.3389/fmicb.2018.03283
Received: 19 April 2018; Accepted: 17 December 2018;
Published: 09 January 2019.
Edited by:
Alison Buchan, University of Tennessee, Knoxville, United StatesReviewed by:
Wenli Chen, Huazhong Agricultural University, ChinaCopyright © 2019 Aires, Muyzer, Serrão and Engelen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tânia Aires, dGFuaWFpcmVzQGdtYWlsLmNvbQ== Aschwin H. Engelen, YWVuZ2VsZW5AdWFsZy5wdA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.