
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 17 December 2018
Sec. Infectious Agents and Disease
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.03105
This article is part of the Research Topic Unraveling the Biology, Genetics, and Host/Environmental Interactions of Acinetobacter View all 17 articles
 Laura Fernandez-Garcia1†
Laura Fernandez-Garcia1† Antón Ambroa1†
Antón Ambroa1† Lucia Blasco1
Lucia Blasco1 Ines Bleriot1
Ines Bleriot1 Maria López1Rocio Alvarez-Marin2
Maria López1Rocio Alvarez-Marin2 Felipe Fernández-Cuenca3
Felipe Fernández-Cuenca3 Luis Martinez-Martinez4Jordi Vila5
Luis Martinez-Martinez4Jordi Vila5 Jesús Rodríguez-Baño3Jose Garnacho-Montero6Jose Miguel Cisneros2Alvaro Pascual3Jeronimo Pachón2,7
Jesús Rodríguez-Baño3Jose Garnacho-Montero6Jose Miguel Cisneros2Alvaro Pascual3Jeronimo Pachón2,7 German Bou1
German Bou1 Younes Smani2
Younes Smani2 Maria Tomás1*
Maria Tomás1*Acinetobacter baumannii (Ab) is one of the most important pathogens associated with nosocomial infections, especially pneumonia. Interest in the Quorum network, i.e., Quorum Sensing (QS)/Quorum Quenching (QQ), in this pathogen has grown in recent years. The Quorum network plays an important role in regulating diverse virulence factors such as surface motility and bacterial competition through the type VI secretion system (T6SS), which is associated with bacterial invasiveness. In the present study, we investigated 30 clinical strains of A. baumannii isolated in the “II Spanish Study of A. baumannii GEIH-REIPI 2000-2010” (Genbank Umbrella Bioproject PRJNA422585), a multicentre study describing the relationship between the Quorum network in A. baumannii and the development of pneumonia and associated bacteraemia. Expression of the aidA gene (encoding the AidA protein, QQ enzyme) was lower (P < 0.001) in strains of A. baumannii isolated from patients with bacteraemic pneumonia than in strains isolated from patients with non-bacteraemic pneumonia. Moreover, aidA expression in the first type of strain was not regulated in the presence of environmental stress factors such as the 3-oxo-C12-HSL molecule (substrate of AidA protein, QQ activation) or H2O2 (inhibitor of AidA protein, QS activation). However, in the A. baumannii strains isolated from patients with non-bacteraemic pneumonia, aidA gene expression was regulated by stressors such as 3-oxo-C12-HSL and H2O2. In an in vivo Galleria mellonella model of A. baumannii infection, the A. baumannii ATCC 17978 strain was associated with higher mortality (100% at 24 h) than the mutant, abaI-deficient, strain (carrying a synthetase enzyme of Acyl homoserine lactone molecules) (70% at 24 h). These data suggest that the QS (abaR and abaI genes)/QQ (aidA gene) network affects the development of secondary bacteraemia in pneumonia patients and also the virulence of A. baumannii.
Acinetobacter baumannii is a major cause of hospital-acquired infections associated with high mortality rates (Fuchs, 2016),s usually affecting patients in Intensive Care Units (ICU) (del Mar Tomas et al., 2005; Lee et al., 2017). In these patients, A. baumannii causes infections such as pneumonia or, to a lesser extent, serious infections of the bloodstream (around 10% of clinical isolates of A. baumannii cause bacteraemia) (Cisneros and Rodríguez-Baño, 2002; El Kettani et al., 2017).
The success of this bacterium as a nosocomial pathogen, has been attributed to the following factors, amongst others: (i) high genetic versatility, facilitating rapid adaptation to stressful or unfavorable situations (Gayoso et al., 2014; Trastoy et al., 2018); (ii) ability to acquire new genes horizontally by the acquisition of plasmids and phages (López et al., 2018); (iii) ability to persist for a long time on animate and inanimate surfaces (resistance to desiccation) (Gayoso et al., 2014), which is generally attributed to biofilm formation; (iv) resistance to antimicrobial agents, including broad-spectrum antibiotics such as carbapenems, colistin, and tigecycline (Fernández-Cuenca et al., 2015), as well as to disinfectants and biocides (Fernández-García et al., 2018); and (v) high virulence (colonization, invasiveness, and cytotoxicity) (Rumbo et al., 2014; Wong et al., 2017). These characteristics contribute to the fact that nosocomial outbreaks caused by A. baumannii are difficult to control and that therapeutic options to treat infections are scarce or non-existent (Fernández-Cuenca et al., 2013). In February, 2017, the World Health Organization (WHO) published a list of “priority pathogens.” The list includes antibiotic resistant bacteria, considered a serious threat to human health and for which new antibiotics are urgently needed, and is headed by carbapenem-resistant A. baumannii (Tacconelli et al., 2018).
The Quorum Sensing (QS) network is generally used by Gram-negative bacterial pathogens to regulate biological processes such as virulence, conjugation, resistance, biofilm formation (which also depends on other factors such as the lytic enzymes responsible for peptidoglycan recycling: Vijayakumar et al., 2016), motility and bacterial competition, via secretion systems (T6SS), which are associated with greater invasiveness (LaSarre and Federle, 2013; López et al., 2017a,b). Two proteins (AbaI /AbaR) identified in A. baumannii have been described as homologs of the LuxI/LuxR system found in Vibrio fischeri. This system comprises a signal or autoinducer molecule (acyl-homoserine lactone, AHL), an enzyme that synthesizes signaling molecules (AbaI) and a receptor protein activator of the QS (AbaR), which forms a complex with N-(3-hydroxydodecanoil)-L-homoserine lactone (3-OH-C12-HSL) to regulate virulence factors, biofilm formation, surface motility, and bacterial competence (T6SS) (Stacy et al., 2012). When a threshold concentration is reached, the AHL molecules present inside the cell are transported to its receptor (AbaR), putatively joining the lux-box, which is located 67 bp upstream of the ATG of AbaI, resulting in the synthesis of more AHL molecules (López et al., 2017b). The QS mechanism, on the other hand, acts naturally under environmental stress conditions such as the presence of bile salts in the gastrointestinal tract and H2O2 (ROS response) in the respiratory tract (López et al., 2017b).
A new enzyme (AidA) has recently been cloned in E. coli BL21 (DE3) and functionally characterized in clinical strains of A. baumannii capable of inhibiting their own QS (by Quorum Quenching) (López et al., 2017b). This enzyme acts by degrading signaling molecules such as N-(3-Oxo-dodecanoyl), L-homoserine lactone (3-Oxo-C12-HSL), and N-dodecanoyl-L-homoserine lactone (C12-HSL), as confirmed by observation of inhibition of motility, biofilm formation and other virulence factors associated with activation of the Quorum Sensing system (López et al., 2017b; Mayer et al., 2018). Other QQ enzymes have also recently been described in A. baumannii ATCC17978 (A1S_0383, A1S_2662, A1S_1876) (Mayer et al., 2018). Multiple QQ enzymes have been analyzed in diverse pathogens such as Pseudomonas aeruginosa (Zhang et al., 2011), Deinococcus radiodurans, Hyphomonas neptunium, Photorhabdus luminicencens, and Rhizobium spp. (Kalia et al., 2011; Krysciak et al., 2011).
Based on these findings, in the present study, we examined the relationship between the global Quorum regulatory network (QS/QQ) mediated by the abaR (QS) and aidA (QQ) genes and the development of pneumonia and bacteraemia in clinical strains of A. baumannii isolated in the “II Spanish Study of A. baumannii GEIH-REIPI 2000-2010,” a multicentre study involving 45 Spanish hospitals and 246 patients. In addition, we used an in vivo infection model consisting of larvae of the wax moth Galleria mellonella to examine the relationship between the global QS/QQ and the development of mortality by a mutant abaI (QS)-deficient strain of A. baumannii (A. baumannii ATCC17978ΔabaI) relative to that of the wild-type A. baumannii ATCC17978 strain.
To carry out this study, we analyzed 30 clinical strains of A. baumannii from the 465 strains isolated in the “II Spanish Study of A. baumannii GEIH-REIPI 2000-2010” multicentre study (Genbank Umbrella Bioproject PRJNA422585). The multicentre study included 45 hospitals in Spain, in which new cases of colonization or infection by A. baumannii were analyzed between February and March 2010 (Villar et al., 2014). The 30 A. baumannii strains were all isolated from respiratory samples from patients with nosocomial pneumonia (n = 13: 6 with and 7 without bacteraemia) or A. baumannii colonization of the lower respiratory tract (n = 17) (Sánchez-Encinales et al., 2017). Molecular typing was performed by Multilocus Sequence Typing (MLST) (Mosqueda et al., 2014). In addition, we used a killing assay with the Galleria mellonella infection model and an A. baumannii ATCC17978ΔabaI mutant strain (identified by Castañeda-Tamez et al., 2018).
The main clinical study variables included demographics, underlying diseases, mechanical ventilation, tracheostomy, colonization of lower respiratory airways, bacteraemic pneumonia (Pn-B), non-bacteraemic pneumonia (Pn-NB) (Horan et al., 2008) and any cause of death during hospitalization.
To design the primers and probes of the QS genes and QQ enzymes, we analyzed the presence of QS genes (abaR and abaI) and the QQ enzyme (aidA) in A. baumannii ATCC 17978 (Genbank genome accession numbers CP000521.1 [CP018664.1]) and in 1000 A. baumannii genomes by consulting the “Integrated Microbial Genomes and Microbiomes” web page (https://img.jgi.doe.gov) and using nucleotide BLAST. The gene sequences used in the search were selected from the Acinetobacter baumannii ATCC 17978 genome. A threshold of 1e-50 was used as the limit for analysis of the nucleotide sequence, where the e-value was defined as the probability of random alignments with the same score. We also calculated the percentage presence of these genes in the genomes (Figure S1).
All clinical strains of A. baumannii were cultured on solid Luria-Bertani (LB) plates and incubated at 37° C for 24 h. One colony was removed and inoculated in liquid LB medium and incubated overnight at 37° C under stirring at 180 rpm. The inoculum was diluted (1:100) and allowed to grow until an optical density (OD600 nm) of 0.4–0.6 (corresponding to the logarithmic growth phase) was reached. The RNA was then extracted using the High Pure RNA Isolation kit (Roche, Germany) and the extract was treated with Dnase (Roche, Germany). The extracted RNA was subsequently quantified in a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies), and the concentration was adjusted to 50 ng/μl in order to yield efficiencies of 90-110% (Rumbo et al., 2013). All extractions were carried out in duplicate.
The 13 strains of A. baumannii, isolated from patients with pneumonia, were cultured on solid Luria-Bertani (LB) plates and incubated at 37°C for 24 h. One colony was then removed, inoculated in liquid LB medium and incubated overnight at 37°C under stirring at 180 rpm. The preinoculum was diluted (1:100) and allowed to grow until an optical density (OD600 nm) of 0.3 was reached. Aliquots of 10 μM of 3-Oxo-C12-HSL (QS-inactivating molecule by expression of the AidA protein) (Stacy et al., 2012; López et al., 2017b) and (10 μl) H2O2 were then added for 5 min (QS-activator by ROS response) (López et al., 2018). All controls were prepared by adding the same volumes of DMSO (dimethyl sulfoxide), 3-Oxo-C12-HSL and of sample, but with no H2O2. After incubation of the samples for 4 and 5 h in the presence of 3-Oxo-C12-HSL, to study the regulatory QS/QQ genes (abaR and aidA), as well as 5 min under H2O2 in static at 37°C, RNA was extracted using the High Pure RNA Isolation kit (Roche, Germany) and treated with Dnase. The extracted RNA was subsequently quantified as described above (Rumbo et al., 2013).
The studies were carried out with a Lightcycler 480 RNA MasterHydrolysis Probe (Roche, Germany), under the following conditions: reverse transcription at 63°C for 3 min, denaturation at 95°C for 30 s, followed by 45 cycles of 15 s at 95°C and 45 s at 60°C and, finally, cooling at 40°C for 30 s. The UPL primers and probes from conserved DNA regions identified by PCR (Universal Probe Library-Roche, Germany) used in the analysis are shown in Table 1.
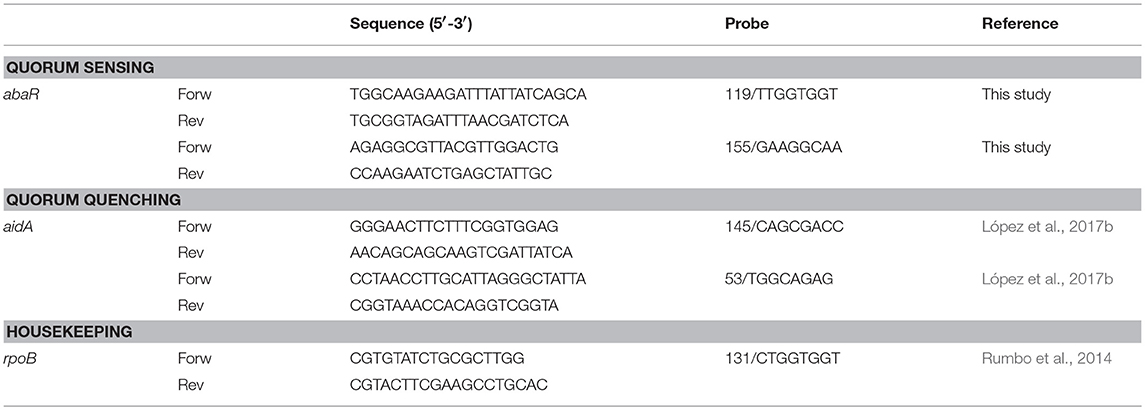
Table 1. Primers and Probes used in this study.
All of the experiments were carried out in a final volume of 20 μl per well (18 μl of master mix and 2 μl of RNA). Each experiment was carried out in duplicate with two RNA extracts (50 ng/μl). For each strain, the expression of all genes, primers, and probes was normalized relative to the reference or housekeeping gene, rpoB, for RT-qPCR studies of Quorum sensing Primer sequences (5′-3′) with Taqman probes (Rumbo et al., 2013; López et al., 2017b). Analysis of the controls without reverse transcriptase confirmed the absence of DNA contamination.
The Galleria mellonella model was an adapted version of that developed by Peleg et al. (2009), Yang et al. (2015). The procedure was as follows: twelve G. mellonella larvae, acquired from TruLarvTM (Biosystems Technology, Exeter, Devon, UK), were each injected with 10 μl of a suspension of A. baumannii ATCC17978, or its isogenic deficient mutant A. baumannii ATCC17978ΔabaI, diluted in sterile phosphate buffer saline (PBS) and containing 8 × 104 CFU (± 0. 5 log). The injection was performed with a Hamilton syringe (volume 100 μl) (Hamilton, Shanghai, China). In addition, a control group of twelve larvae were injected with 10 μl of sterile PBS. After being injected, the groups of larvae were placed in Petri dishes and incubated in darkness at 37°C. The number of dead larvae was recorded twice a day (morning and afternoon) for 6 days. The larvae were considered dead when they showed no movement in response to touch (Peleg et al., 2009).
The gene expression studies were carried out in duplicate, and the data obtained were analyzed by Student's t-test, implemented with GraphPad Prism v.6 software (GraphPad Software Inc. San Diego, CA). The graphs were constructed using the GraphPad program, and the results were represented as means and their respective standard deviations.
The mortality curves corresponding to the in vivo Galleria mellonella infection model were constructed using GraphPad Prism v.6 and the data were analyzed using the Log-rank test (Mantel-Cox). In both cases, p-values < 0.05 were considered statistically significant, and the data were expressed as mean values.
The statistical analyses were applied to the following categorical variables: age, sex, immunosuppressive treatment, surgery, ICU stay, mechanical ventilation, tracheostomy, severe sepsis, septic shock, and expression of the Quorum genes in A. baumannii clinical strains (Bone et al., 1992). In addition, the severity of co-morbidities was assessed using the Charlson score (Charlson et al., 1987) and the McCabe score (McCabe and Jackson, 1962). Chi-square and Fisher tests were used in the univariate analysis of categorical variables. Continuous variables were analyzed using two-sample t-test or Mann Whitney, as appropriate. A logistic regression analysis was performed to identify factors independently associated with pneumonia and bacteraemia. Differences were considered significant at p < 0.05. All statistical analyses were performed using SPSS v.16.0 (SPSS Inc., Chicago, IL).
The Relative Expression (RE) of the abaR and aidA genes of the Quorum network (QS/QQ) was quantified by RT-qPCR analysis of the 17 isolates of A. baumannii from colonized patients and of the 13 isolates of A. baumannii from patients with pneumonia (Figure 1A). The mean values (of two biological replicates) are presented in Tables 2, 3. These values were first used to determine any significant differences between the two types of strains in terms of gene expression in the Quorum network.
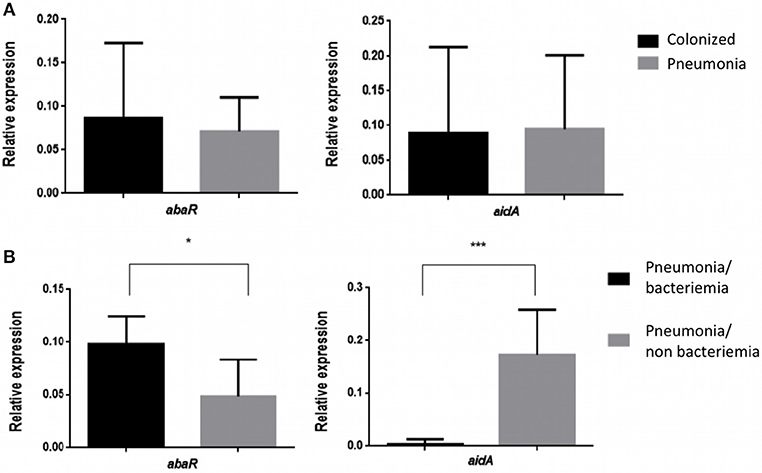
Figure 1. (A) Relative Expression of the abaR and aidA genes in strains of A. baumannii from patients colonized with A. baumannii and patients with pneumonia caused by A. baumannii. No significant differences (p > 0.05) were detected in either case. (B) Relative expression of the abaR and aidA genes in isolates of A. baumannii from patients with bacteraemic and non-bacteraemic pneumonia. *p-value < 0.05 and ***p-value < 0.001.
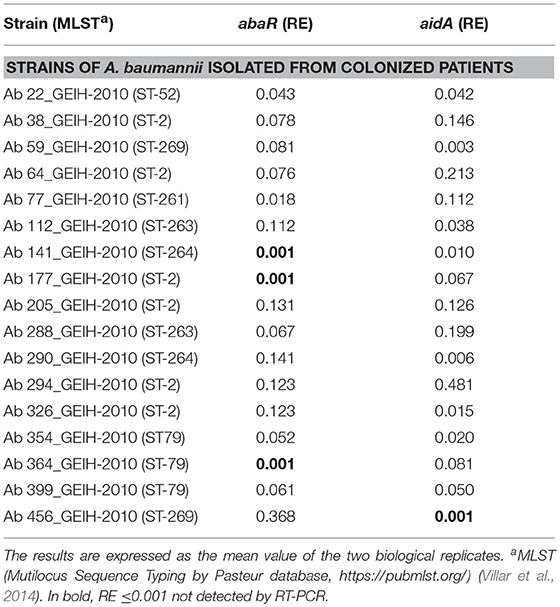
Table 2. Results of RT-qPCR analysis of the Relative Expression (RE) of the abaR and aidA genes (Quorum network genes) in the A. baumannii isolates from colonized patients.
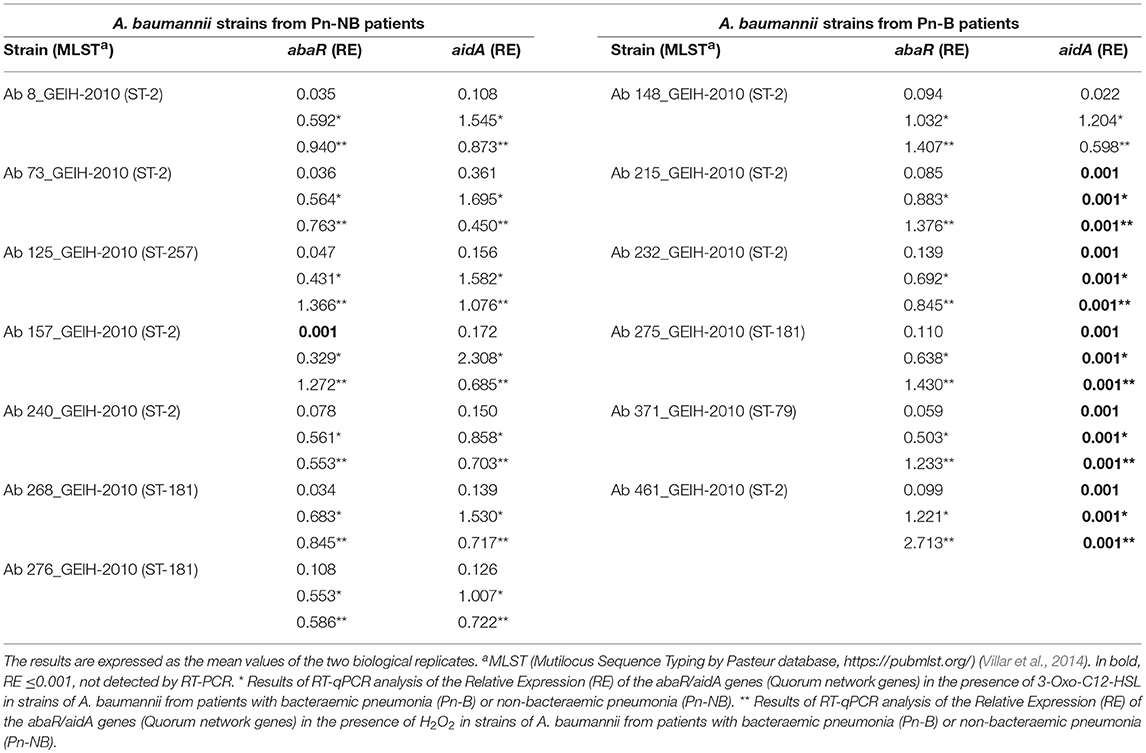
Table 3. Results of RT-qPCR analysis of the Relative Expression (RE) of the abaR and aidA genes (Quorum network genes) in the A. baumannii isolates from patients with bacteraemic pneumonia (Pn-B) or non-bacteraemic pneumonia (Pn-NB).
The results did not reveal any significant differences in the RE of the Quorum network genes (abaR, aidA) between clinical strains of A. baumannii isolated from colonized patients and strains of A. baumannii isolated from patients with pneumonia (0.086/0.094 vs. 0.071/0.095, p > 0.05).
We then proceeded to study the RE of the abaR and aidA genes in strains of A. baumannii from patients with pneumonia, differentiating the strains isolated from patients with bacteraemia (Pn-B) from those isolated from patients without bacteraemia (Pn-NB). The resulting graphs are shown below (Figure 1B). The findings reveal significant differences in the expression of the abaR and aidA genes between clinical strains of A. baumannii from patients with bacteraemic pneumonia (Pn-B) and those with non-bacteraemic pneumonia (Pn-NB). We observed that abaR gene was overexpressed in A. baumannii isolates from Pn-B patients relative to Pn-NB patients (0.047 vs. 0.097, p < 0.05). By contrast, the aidA gene was overexpressed in A. baumannii clinical strains in Pn-NB patients relative to Pn-B patients (0.173 vs. 0.0045, p < 0.001) (Figure 1B). Only one strain, Ab 148_GEIH-2010 (ST-2), isolated from Pn-B patients, showed an aidA gene profile different from the other isolates of this group, although the RE of this gene was lower (0.022) than that of isolates from Pn-NB patients.
The values of the RE of the abaR and aidA genes (Quorum network) in the presence of 3-Oxo-C12-HSL (Inhibition of the QS) and H2O2 (Activation of the QS), obtained by RT-qPCR of the 13 isolates of A. baumannii from patients with pneumonia (differentiated from Pn-NB) are shown in Table 3, expressed as the mean value of the two biological replicates. These values were then analyzed to determine any significant differences in the RE of the abaR/aidA (QS/QQ) genes between the different clinical isolates (Figures 2, 3).
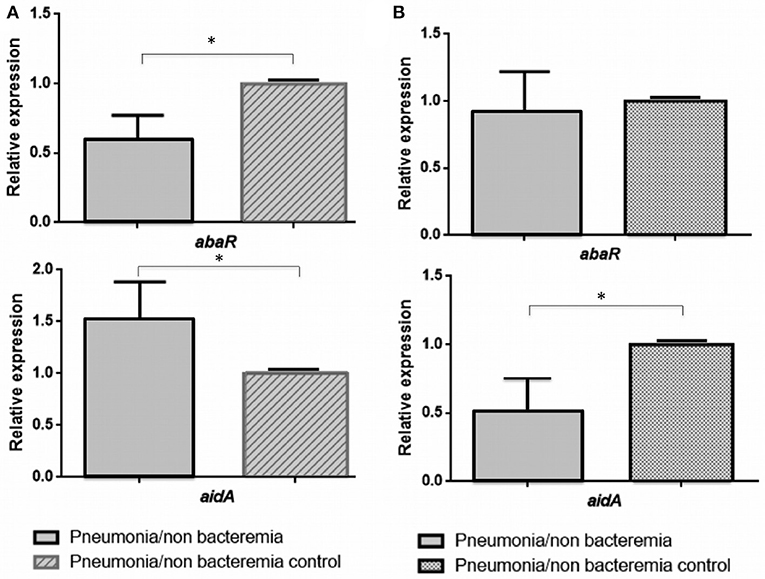
Figure 2. Relative expression of the abaR and aidA genes under 3-oxo-C12-HSL (A) and H2O2 (B) in isolates of A. baumannii from patients with non-bacteraemic pneumonia (Pn-NB). *p-value < 0.05.
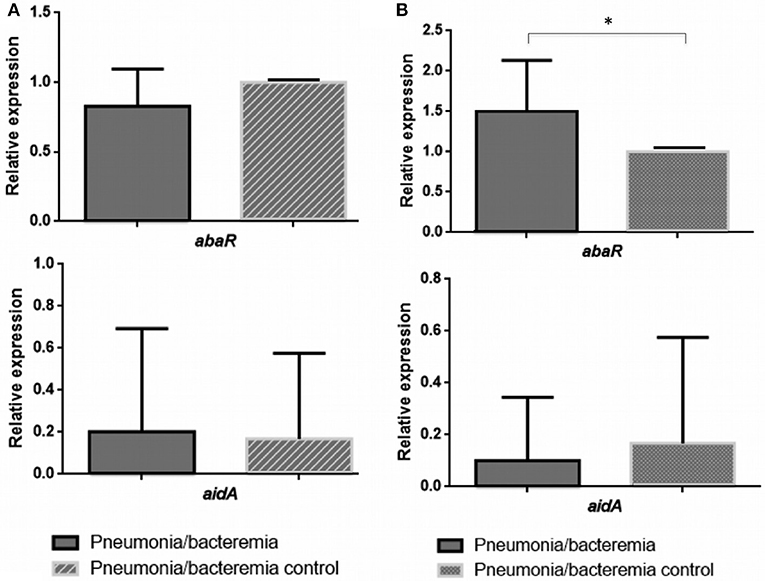
Figure 3. Relative expression of the abaR and aidA genes under 3-oxo-C12-HSL (A) and H2O2 (B) in isolates of A. baumannii from patients with bacteraemic pneumonia (Pn-B). *p-value < 0.05.
In the clinical strains of A. baumannii isolated from Pn-NB (Figure 2), we observed regulation of expression of the aidA gene in the presence of 3-Oxo-C12-HSL (overexpression, RE ≥ 1.5) (Figure 2A) and of H2O2 [underexpression, RE ≤ 0.5 (Figure 2B)]. Expression of the abaR gene decreased significantly in the presence of the 3-Oxo-C12-HSL molecule (RE ≤ 0.5, Figure 2A).
In the clinical strains A. baumannii isolated from Pn-B (Figure 3), expression of the aidA gene was not regulated in the presence of 3-Oxo-C12-HSL or H2O2. However, the abaR gene was overexpressed in the presence of H2O2 (RE ≥ 1.5, Figure 3B).
These results indicate that the isolates of A. baumannii from Pn-NB may harbor a functional AidA protein (QQ enzyme), in contrast to the isolates of A. baumannii from Pn-B, which did not have this functional protein. Therefore, in the A. baumannii strains isolated from Pn-B, overexpression of the abaR gene (activation of the QS) in the presence of H2O2 (ROS response) would enable the development of the virulence factors favoring invasiveness, such as type VI secretion system (T6SS) and motility.
Analysis of the risk factors associated with the development of pneumonia vs. colonization by clinical strains of A. baumannii revealed only one statistically significant variable, i.e., diabetes mellitus (Table 4).
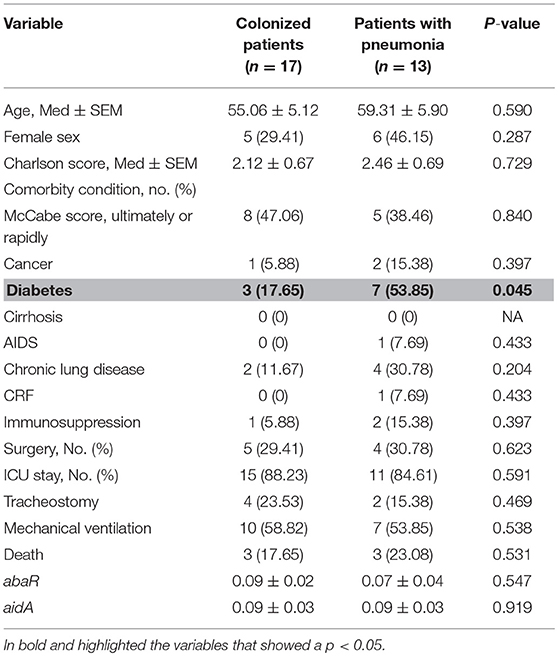
Table 4. Univariate analysis of risk factors associated with development of pneumonia relative to colonization by clinical strains of A. baumannii.
However, analysis of the risk factors associated with the development of bacteraemia in pneumonia caused by A. baumannii revealed underexpression of the aidA gene as the only statistically significant variable (p < 0.05) (Table 5).
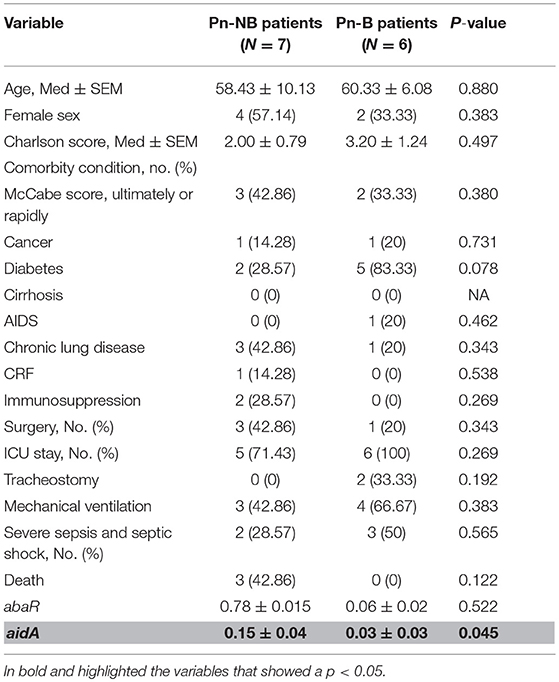
Table 5. Univariate analysis of risk factors associated with the development of bacteraemia in pneumonia caused by A. baumannii relative to the non-bacteraemic pneumonia control.
Injection of G. mellonella larvae with A. baumannii ATCC17978 at a concentration of 8 x 104 CFU/larva (± 0.5 log) caused 100% mortality after 24 h, whereas injection of the larvae with the same concentration of A. baumannii ATCC17978ΔabaI resulted in 70% mortality after 24 h (Figure 4; p < 0.05, Mantel-Cox analysis).
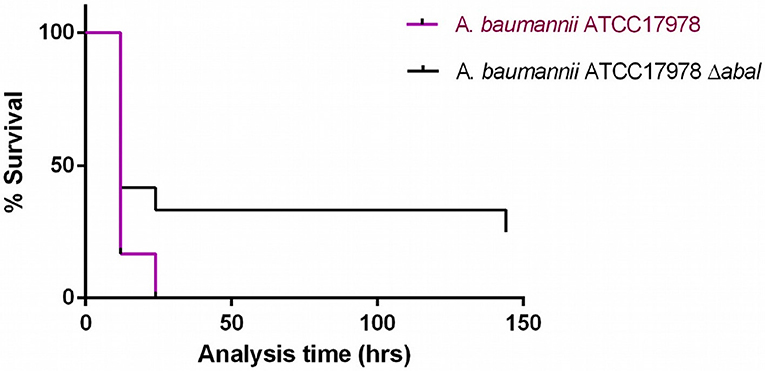
Figure 4. Survival curves for G. mellonella larvae injected with A. baumannii ATCC17978 reference strain and its isogenic derivative A. baumannii ATCC17978 ΔabaI. Data from a single representative assay. For simplicity, the control group is not included in this figure.
In this study, we analyzed the expression of Quorum network (QS/QQ) genes that differed between genomes of clinical isolates of A. baumannii, abaR and abaI (QS system) and aidA (QQ mechanism) in relation to clinical features of pneumonia and bacteraemia. Although other QQ enzymes have been described in A. baumannii ATCC 17978 (Mayer et al., 2018), these were not analyzed in the present study due to the lack of any differences between A. baumannii genomes.
In clinical strains of A. baumannii isolated from patients with bacteraemic pneumonia (Pn-B), the abaR gene was overexpressed (p < 0.05). The AbaR protein was the receptor activator of the Quorum Sensing system (QS), and the aidA gene was not expressed. Moreover, we observed regulation of aidA gene expression in clinical strains of pneumonia-causing A. baumannii (non-bacteraemic pneumonia, Pn-NB) by the 3-Oxo-C12-HSL molecule (which is an AidA enzyme substrate in QQ activity) and H2O2 (an activator of the QS system). However, there was no difference in the expression of Quorum network genes between colonized and pneumonia patients, as previously described (Stones and Krachler, 2016).
On the other hand, clinical analysis of the risk factors associated with pneumonia caused by A. baumannii revealed diabetes mellitus as only statistically significant risk factor (Kim et al., 2014). In relation to bacteraemia in A. baumannii pneumonia (P < 0.05), underexpression of the aidA gene was also the only statistically significant variable (P < 0.05).
In several pathogens, such as Yersinia pseudotuberculosis, Proteus mirabilis, and Vibrio cholerae, the QS system is the main regulatory mechanism of bacterial competence via T6SS, which is involved in the invasiveness and motility that favor the development of bacteraemia (Zhang et al., 2011; Debnath et al., 2018; Jaskólska et al., 2018; Trastoy et al., 2018). Moreover, in 86% of ICU patients, gastrointestinal tract colonization by a clinical strain of A. baumannii led to development of bacteraemia caused by genetically similar strains (Thom et al., 2010). This implies that clinical isolates of A. baumannii most capable of surviving under stress conditions (such as the presence of bile salts in the gastrointestinal tract or H2O2 in the respiratory tract) (Zheng et al., 2018) may have a higher invasive capacity due to virulence factors, such as the type VI secretion system (T6SS), previously activated under stressful conditions. Motility is also a crucial virulence factor, allowing penetration of the bacteria into the host's body and subsequent colonization (Gellatly and Hancock, 2013). Previous studies have demonstrated the existence of a relationship between motility and the origin of the isolates. Indeed, blood isolates of A. baumannii have been found to be more mobile than sputum isolates (Vijayakumar et al., 2016). Interestingly, 67% of the clinical isolates of A. baumannii were non-mobile and all of them had the AidA protein and were of respiratory origin (López et al., 2017a,b). In addition, the aidA gene was not located in the genome of the only mobile strain (clone ST79/PFGE-HUI-1) isolated from blood and which was the origin of a bacteraemic outbreak (López et al., 2017a,b).
Finally, multiple studies carried out with the abaI mutant of the M2 strain of Acinetobacter nosocomialis have analyzed the role of the abaI gene (responsible for the synthesis of quorum sensing synthesizing molecules) in various virulence factors such as biofilm formation and motility. In both cases, abaI deficiency led to a decrease in biofilm production and motility (Niu et al., 2008; Bhargava et al., 2012). The mutant lacking abaI is believed to be less virulent than the wild strain. This result was confirmed in our study in which injection of G. mellonella larvae with the reference A. baumannii ATCC17978 strain caused higher mortality than injection with the mutant A. baumannii ATCC17978ΔabaI. Regarding the mortality of the reference strain (A. baumannii ATCC17978), similar effects have been observed in other studies, in which injection of G. mellonella larvae with the reference strain A. baumannii ATCC17978 resulted in rapid death. Mortality was significantly dependent on the number of cells injected. More than 75% of the larvae died in the first 48 h of injection with at least 3.7 × 105 CFU / larva, while very few of the larvae died after being injected with a concentration equal to or lower than 3.7 × 104 CFU/larva (p < 0.01) (Clemmer et al., 2011). The results regarding the mutant A. baumannii ATCC17978ΔabaI are consistent with those obtained in a study of Pseudomonas aeruginosa (Steindler et al., 2009) in which a mutant ΔrhLI ΔlasI (QS systems homologous to abaI) was obtained, demonstrating that inactivation of both QS systems leads to a significant reduction in pathogenicity (p < 0.01) when virulence factors are not activated, such as the type VI secretion system (T6SS) and motility (Jaskólska et al., 2018).
In conclusion, our findings suggest that the QS (abaR and abaI genes)/QQ (aidA gene) network plays a role in the development of bacteraemia in patients with pneumonia caused by A. baumannii. This is the first study reporting a relationship between reduced expression of this bacterial QQ enzyme gene (AidA protein) and bacteraemia. Further studies of this relationship in the same and other bacterial QQ enzymes would be of great interest.
LF-G, AA, LB, IB, ML and RA-M developed the experiments. FF-C, LM-M, JV, JR-B, JG-M, JMC, AP, JP, GB, and YS wrote the manuscript and provided the strains. MT led the experiments and manuscript redaction.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This study was funded by grant PI16/01163 awarded to MT within the State Plan for R+D+I 2013-2016 (National Plan for Scientific Research, Technological Development and Innovation 2008-2011) and co-financed by the ISCIII-Deputy General Directorate for Evaluation and Promotion of Research—European Regional Development Fund A way of Making Europe and Instituto de Salud Carlos III FEDER, Spanish Network for the Research in Infectious Diseases (REIPI, RD16/0016/0001, RD16/0016/0006, RD16/0016/0008, RD16/0016/0009, and RD16/0016/0010) and by the Study Group on Mechanisms of Action and Resistance to Antimicrobials, GEMARA (SEIMC, http://www.seimc.org/). MT was financially supported by the Miguel Servet Research Programme (SERGAS and ISCIII). LF-G was financially supported by a predoctoral fellowship from the Xunta de Galicia (GAIN, Axencia de Innovación).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.03105/full#supplementary-material
Bhargava, N., Sharma, P., and Capalash, N. (2012). N-acyl homoserine lactone mediated interspecies interactions between A. baumannii and P. aeruginosa. Biofouling. 28, 813–822. doi: 10.1080/08927014.2012.714372
Bone, R. C., Balk, R. A., Cerra, F. B., Dellinger, R. P., Fein, A. M., Knaus, W. A., et al. (1992). Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. The ACCP/SCCM Consensus Conference Committee. American College of Chest Physicians/Society of Critical Care Medicine. Chest H101, 1644–1655. doi: 10.1378/chest.101.6.1644
Castañeda-Tamez, P., Ramírez-Peris, J., Pérez-Velázquez, J., Kuttler, C., Jalalimanesh, A., Saucedo-Mora, M. Á., et al. (2018). Pyocyanin restricts social cheating in Pseudomonas aeruginosa. Front. Microbiol. 9:1348. doi: 10.3389/fmicb.2018.01348
Charlson, M. E., Pompei, P., Ales, K. L., and MacKenzie, C. R. (1987). A new method of classifying prognostic comorbidity in longitudinal studies: development and validation. J. Chronic Dis. 40, 373–383. doi: 10.1016/0021-9681(87)90171-8
Cisneros, J. M., and Rodríguez-Baño, J. (2002). Nosocomial bacteremia due to Acinetobacter baumannii: epidemiology, clinical features and treatment. Clin. Microbiol. Infect. 8, 687–693. doi: 10.1046/j.1469-0691.2002.00487.x
Clemmer, K. M., Bonomo, R. A., and Rather, P. N. (2011). Genetic analysis of surface motility in Acinetobacter baumannii. Microbiology 157, 2534–2544. doi: 10.1099/mic.0.049791-0
Debnath, I., Stringer, A. M., Smith, S. N., Bae, E., Mobley, H. L. T., Wade, J. T., et al. (2018). MrpJ directly regulates proteus mirabilis virulence factors, including fimbriae and type VI secretion, during urinary tract infection. Infect Immun. 86, e00388–18. doi: 10.1128/IAI.00388-18
del Mar Tomas, M., Cartelle, M., Pertega, S., Beceiro, A., Llinares, P., Canle, D., et al. (2005). Hospital outbreak caused by a carbapenem-resistant strain of Acinetobacter baumannii: patient prognosis and risk-factors for colonisation and infection. Clin. Microbiol. Infect. 11, 540–546. doi: 10.1111/j.1469-0691.2005.01184.x
El Kettani, A., Maaloum, F., Diawara, I., Katfy, K., Harrar, N., Zerouali, K., et al. (2017). Prevalence of Acinetobacter baumannii bacteremia in intensive care units of Ibn Rochd University Hospital, Casablanca. Iran J Microbiol. 9, 318–323.
Fernández-Cuenca, F., Tomás, M., Caballero-Moyano, F. J., Bou, G., Martínez-Martínez, L., Vila, J., et al. (2015). Reduced susceptibility to biocides in Acinetobacter baumannii: association with resistance to antimicrobials, epidemiological behaviour, biological cost and effect on the expression of genes encoding porins and efflux pumps. J. Antimicrob. Chemother. 70, 3222–3229. doi: 10.1093/jac/dkv262
Fernández-Cuenca, F., Tomás-Carmona, M., Caballero-Moyano, F., Bou, G., Martínez-Martínez, L., Vila, J., et al. (2013). [In vitro activity of 18 antimicrobial agents against clinical isolates of Acinetobacter spp.: multicenter national study GEIH-REIPI-Ab 2010]. Enferm. Infecc. Microbiol. Clin. 31, 4–9. doi: 10.1016/j.eimc.2012.06.010
Fernández-García, L., Fernandez-Cuenca, F., Blasco, L., Lopez-Rojas, R., Ambroa, A., Lopez, M., et al. (2018). Relationship between tolerance and persistence mechanisms in Acinetobacter baumannii strains with AbkAB Toxin-Antitoxin system. Antimicrob Agents Chemother. 62, e00250–18. doi: 10.1128/AAC.00250-18
Fuchs, R. P. (2016). Tolerance of lesions in E. coli: Chronological competition between Translesion Synthesis and Damage Avoidance. DNA Repair 44, 51–58. doi: 10.1016/j.dnarep.2016.05.006
Gayoso, C. M., Mateos, J., Méndez, J. A., Fernández-Puente, P., Rumbo, C., Tomás, M., et al. (2014). Molecular mechanisms involved in the response to desiccation stress and persistence in Acinetobacter baumannii. J. Proteome Res. 13, 460–476. doi: 10.1021/pr400603f
Gellatly, S. L., and Hancock, R. E. (2013) Pseudomonas aeruginosa: new insights into pathogenesis host defenses. Pathog. Dis. 67, 1590–173. doi: 10.1111/2049-632x.12033
Horan, T. C., Andrus, M., and Dudeck, M. A. (2008). CDC/NHSN surveillance definition of health care-associated infection and criteria for specific types of infections in the acute care setting. Am. J. Infect. Control. 36, 309–332. doi: 10.1016/j.ajic.2008.03.002
Jaskólska, M., Stutzmann, S., Stoudmann, C., and Blokesch, M. (2018). QstR-dependent regulation of natural competence and type VI secretion in Vibrio cholerae. Nucleic Acids Res. 46, 10619–10634. doi: 10.1093/nar/gky717
Kalia, V. C., Raju, S. C., and Purohit, H. J. (2011). Genomic analysis reveals versatile organisms for quorum quenching enzymes: acyl-homoserine lactone-acylase and -lactonase. Open Microbiol. J. 5, 1–13. doi: 10.2174/1874285801105010001
Kim, T., Chong, Y. P., Park, S. Y., Jeon, M. H., Choo, E. J., Chung, J. W., et al. (2014). Risk factors for hospital-acquired pneumonia caused by carbapenem-resistant Gram-negative bacteria in critically ill patients: a multicenter study in Korea. Diagn. Microbiol. Infect. Dis. 78, 457–461. doi: 10.1016/j.diagmicrobio.2013.08.011
Krysciak, D., Schmeisser, C., Preuss, S., Riethausen, J., Quitschau, M., Grond, S., et al. (2011). Involvement of multiple loci in quorum quenching of autoinducer I molecules in the nitrogen-fixing symbiont Rhizobium (Sinorhizobium) sp. strain NGR234. Appl. Environ. Microbiol. 77, 5089–5099. doi: 10.1128/AEM.00112-11
LaSarre, B., and Federle, M. J. (2013). Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 77, 73–111. doi: 10.1128/MMBR.00046-12
Lee, C. R., Lee, J. H., Park, M., Park, K. S., Bae, I. K., Kim, Y. B., et al. (2017). Biology of Acinetobacter baumannii: pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell. Infect. Microbiol. 7:55. doi: 10.3389/fcimb.2017.00055
López, M., Blasco, L., Gato, E., Perez, A., Fernandez-Garcia, L., Martinez-Martinez, L., et al. (2017a). Response to bile salts in clinical strains of Acinetobacter baumannii lacking the AdeABC efflux pump: virulence associated with quorum sensing. Front. Cell. Infect. Microbiol. 7:143. doi: 10.3389/fcimb.2017.00143
López, M., Mayer, C., Fernandez-Garcia, L., Blasco, L., Muras, A., Ruiz, F. M., et al. (2017b). Quorum sensing network in clinical strains of A. baumannii: AidA is a new quorum quenching enzyme. PLoS ONE 12:e0174454. doi: 10.1371/journal.pone.0174454
López, M., Rueda, A., Florido, J. P., Blasco, L., Fernandez-Garcia, L., Trastoy, R., et al. (2018). Evolution of the Quorum network and the mobilome (plasmids and bacteriophages) in clinical strains of Acinetobacter baumannii during a decade. Sci. Rep. 8:2523. doi: 10.1038/s41598-018-20847-7
Mayer, C., Muras, A, Romero, M., López, M., Tomás, M., and Otero, A. (2018). Multiple quorum quenching enzymes are active in the nosocomial pathogen Acinetobacter baumannii ATCC17978. Front. Cell Infect. Microbiol. 8:310. doi: 10.3389/fcimb.2018.00310
McCabe, W. R., and Jackson, G. G. (1962). Gram-negative bacteremia. I. Etiology and ecology. Arch Int Med. 110, 847–855.
Mosqueda, N., Gato, E., Roca, I., Lopez, M., de Alegria, C. R., Fernandez Cuenca, F., et al. (2014). Characterization of plasmids carrying the blaOXA-24/40 carbapenemase gene and the genes encoding the AbkA/AbkB proteins of a toxin/antitoxin system. J. Antimicrob. Chemother. 69, 2629–2633. doi: 10.1093/jac/dku179
Niu, C., Clemmer, K. M., Bonomo, R. A., and Rather, P. N. (2008). Isolation and characterization of an autoinducer synthase from Acinetobacter baumannii. J. Bacteriol. 190, 3386–3392. doi: 10.1128/JB.01929-07
Peleg, A. Y., Jara, S., Monga, D., Eliopoulos, G. M., Moellering, R. C., and Mylonakis, E. (2009). Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. Antimicrob. Agents Chemother. 53, 2605–2609. doi: 10.1128/AAC.01533-08
Rumbo, C., Gato, E., López, M., Ruiz de Alegría, C., Fernández-Cuenca, F., Martínez-Martínez, L., et al. (2013). Contribution of efflux pumps, porins, and β-lactamases to multidrug resistance in clinical isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 57, 5247–5257. doi: 10.1128/AAC.00730-13
Rumbo, C., Tomás, M., Fernández Moreira, E., Soares, N. C., Carvajal, M., Santillana, E., et al. (2014). The Acinetobacter baumannii Omp33-36 porin is a virulence factor that induces apoptosis and modulates autophagy in human cells. Infect. Immun. 82, 4666–4680. doi: 10.1128/IAI.02034-14
Sánchez-Encinales, V., Álvarez-Marín, R., Pachón-Ibá-ez, M. E., Fernández-Cuenca, F., Pascual, A., Garnacho-Montero, J., et al. (2017). Overproduction of Outer Membrane Protein A by Acinetobacter baumannii as a risk factor for nosocomial pneumonia, bacteremia, and mortality rate increase. J. Infect. Dis. 215, 966–974. doi: 10.1093/infdis/jix010
Stacy, D. M., Welsh, M. A., Rather, P. N., and Blackwell, H. E. (2012). Attenuation of quorum sensing in the pathogen Acinetobacter baumannii using non-native N-Acyl homoserine lactones. ACS Chem. Biol. 7, 1719–1728. doi: 10.1021/cb300351x
Steindler, L., Bertani, I., De Sordi, L., Schwager, S., Eberl, L., and Venturi, V. (2009). LasI/R and RhlI/R quorum sensing in a strain of Pseudomonas aeruginosa beneficial to plants. Appl. Environ. Microbiol. 75, 5131–5140. doi: 10.1128/AEM.02914-08
Stones, D. H., and Krachler, A. M. (2016). Against the tide: the role of bacterial adhesion in host colonization. Biochem. Soc. Trans. 44, 1571–1580. doi: 10.1042/BST20160186
Tacconelli, E., Carrara, E., Savoldi, A., Harbarth, S., Mendelson, M., Monnet, D. L., et al. (2018). Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327. doi: 10.1016/S1473-3099(17)30753-3
Thom, K. A., Hsiao, W. W., Harris, A. D., Stine, O. C., Rasko, D. A., and Johnson, J. K. (2010). Patients with Acinetobacter baumannii bloodstream infections are colonized in the gastrointestinal tract with identical strains. Am. J. Infect. Control. 38, 751–753. doi: 10.1016/j.ajic.2010.03.005
Trastoy, R., Manso, T., Fernández-García, L., Blasco, L., Ambroa, A., Pérez Del Molino, M. L., et al. (2018). Mechanisms of bacterial tolerance and persistence in the gastrointestinal and respiratory environments. Clin. Microbiol. Rev. 31:e00023–18. doi: 10.1128/CMR.00023-18
Vijayakumar, S., Rajenderan, S., Laishram, S., Anandan, S., Balaji, V., and Biswas, I. (2016). Biofilm formation and motility depend on the nature of the Acinetobacter baumannii clinical isolates. Front Public Health. 4:105. doi: 10.3389/fpubh.2016.00105
Villar, M., Cano, M. E., Gato, E., Garnacho-Montero, J., Miguel Cisneros, J., Ruíz de Alegría, C., et al. (2014). Epidemiologic and clinical impact of Acinetobacter baumannii colonization and infection: a reappraisal. Medicine 93, 202–210. doi: 10.1097/MD.0000000000000036
Wong, D., Nielsen, T. B., Bonomo, R. A., Pantapalangkoor, P., Luna, B., and Spellberg, B. (2017). Clinical and pathophysiological overview of acinetobacter infections: a century of challenges. Clin. Microbiol. Rev. 30, 409–447. doi: 10.1128/CMR.00058-16
Yang, H., Chen, G., Hu, L., Liu, Y., Cheng, J., Li, H., et al. (2015). In vivo activity of daptomycin/colistin combination therapy in a Galleria mellonella model of Acinetobacter baumannii infection. Int. J. Antimicrob. Agents 45, 188–191. doi: 10.1016/j.ijantimicag.2014.10.012
Zhang, W., Xu, S., Li, J., Shen, X., Wang, Y., and Yuan, Z. (2011). Modulation of a thermoregulated type VI secretion system by AHL-dependent quorum sensing in Yersinia pseudotuberculosis. Arch. Microbiol. 193, 351–363. doi: 10.1007/s00203-011-0680-2
Keywords: quorum, sensing/quenching, pneumonia, bacteraemia, Acinetobacter
Citation: Fernandez-Garcia L, Ambroa A, Blasco L, Bleriot I, López M, Alvarez-Marin R, Fernández-Cuenca F, Martinez-Martinez L, Vila J, Rodríguez-Baño J, Garnacho-Montero J, Cisneros JM, Pascual A, Pachón J, Bou G, Smani Y and Tomás M (2018) Relationship Between the Quorum Network (Sensing/Quenching) and Clinical Features of Pneumonia and Bacteraemia Caused by A. baumannii. Front. Microbiol. 9:3105. doi: 10.3389/fmicb.2018.03105
Received: 22 September 2018; Accepted: 30 November 2018;
Published: 17 December 2018.
Edited by:
Maria Soledad Ramirez, California State University, Fullerton, United StatesReviewed by:
Adriana S. Limansky, CONICET Instituto de Biología Molecular y Celular de Rosario (IBR), ArgentinaCopyright © 2018 Fernandez-Garcia, Ambroa, Blasco, Bleriot, López, Alvarez-Marin, Fernández-Cuenca, Martinez-Martinez, Vila, Rodríguez-Baño, Garnacho-Montero, Cisneros, Pascual, Pachón, Bou, Smani and Tomás. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Tomás, bWEuZGVsLm1hci50b21hcy5jYXJtb25hQHNlcmdhcy5lcw==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.