
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 15 November 2018
Sec. Food Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.02767
This article is part of the Research Topic New Knowledge of Food Microbiology in Asia View all 48 articles
 Shi Wu1
Shi Wu1 Jiahui Huang1
Jiahui Huang1 Qingping Wu1*
Qingping Wu1* Jumei Zhang1Feng Zhang1,2Xiaojuan Yang1
Jumei Zhang1Feng Zhang1,2Xiaojuan Yang1 Haoming Wu1
Haoming Wu1 Haiyan Zeng1
Haiyan Zeng1 Moutong Chen1
Moutong Chen1 Yu Ding3
Yu Ding3 Juan Wang4Tao Lei1Shuhong Zhang1Liang Xue1
Juan Wang4Tao Lei1Shuhong Zhang1Liang Xue1This study was to estimate the prevalence and characteristics of Staphylococcus aureus from 1,850 retail meat and meat products in China during July 2011 to June 2016. The samples were collected covering most provincial capitals in China, including 604 raw meat, 601 quick-frozen meat, and 645 ready-to-eat meat. Using the qualitative and quantitative methods, all 39 cities had S. aureus-positive samples, and S. aureus was detected in 35.0% (647/1,850) of the samples. The levels of S. aureus in retail meat showed that the MPN value of the majority of the positive samples ranged from 0.3 to 100 MPN/g. Twenty-four antibiotics were used to test all 868 S. aureus isolates for antibiotic susceptibility. Only 11 isolates (1.26%) were susceptible to all antibiotics, whereas most isolates (821/868, 94.6%) showed resistance or intermediary resistance to more than three or more antibiotics. Of these strains, 104 (12.0%) were resistant to more than 10 antibiotics. However, the most frequent resistance was observed to ampicillin (85.4%), followed by penicillin (84.6%), erythromycin (52.7%), tetracycline (49.3%), kanamycin (45.3%), telithromycin (30.1%), clindamycin (29.6%), streptomycin (21.1%), norfloxacin (20.4%), gentamicin (19.4%), fusidic acid (18.4%), ciprofloxacin (16.9%), chloramphenicol (13.1%), amoxycillin/clavulanic acid (11.0%), and others (<10%). 7.4% of isolates (62/868) were confirmed as methicillin-resistance S. aureus (MRSA). By molecular typing analysis, there were 164 spa types and 111 STs were identified, including 15 novel spa types and 65 newly STs by multilocus sequence typing (MLST) and spa typing. Despite the wide genetic diversity observed among the 868 isolates, a great proportion of the population belonged to finite number of major clones: ST1-t127 (93/868, 10.7%) and ST7-t091 (92/868, 10.6%), ST5-t002 (42/868, 4.8%), ST398-t034 (40/868, 4.6%), ST188-t034 (38/868, 4.4%), ST59-t437 (30/868, 3.5%), ST6-t701 (29/868, 3.3%), and ST9-t899 (27/868, 3.1%) in China. This study reflects S. aureus was readily detected in Chinese retail meat and meat products but the level were not very excessive. In this study, the high antibiotic resistance is alarming and raising public health concern. In additions, most of molecular types of isolates have been linked to human infections around the world, indicating that these types of S. aureus in China have a theoretical pathogenic potential.
Recent years, many pathogens were responsible for food safety. Staphylococcus aureus is recognized as one of the major foodborne pathogens in fresh and ready-to-eat products and responsible for various infections around world (Diep et al., 2006). It could grows at temperature between 15°C and 45°C and at NaCl concentrations as high as 15% (Behling et al., 2010). This bacterium multiplies quickly at room temperature to produce toxins that cause illness. Naturally, the distribution of S. aureus was ubiquity in the world, but the most important infection source of S. aureus was food. Every year, S. aureus involved in about 241,000 illnesses of foodborne disease in USA (Scallan et al., 2011; Wu et al., 2018). In 2013, there were 12.5% of foodborne bacterial outbreaks are caused by S. aureus in China, which showed third most frequently pathogen after Vibrio parahaemolyticus (27.8%) and Salmonella (23.1%) (Wei-Wei et al., 2018).
In the last decades, the spread use of antibiotics in bacteria is increasing the emergence of multidrug resistant strains (MDR), which showed great challenges to public health. As a formidable adaptive capacity strain, S. aureus can adapt to varying environmental conditions and rapidly become resistant to virtually all antibiotics (Mccallum et al., 2010). Nowadays, more and more MDR S. aureus were reported in food poisoning outbreaks and isolated from food product in previous researched (Sauer et al., 2008; Huang et al., 2009; Gharsa et al., 2012; Papadopoulos et al., 2018). Moreover, in recent years, methicillin-resistance S. aureus (MRSA) is attracting extensive attention. It usually showed multiple antimicrobial resistance and listed as one of 12 families of bacteria that pose the greatest threat to human health by WHO in 2017 (Govindaraj and Vanitha, 2018).
Nowadays, various of molecular subtyping approaches [e.g., pulsed-field gel electrophoresis (PFGE), multilocus sequence typing (MLST), and staphylococcal protein A (spa) typing] have been developed for the characterization of S. aureus. For epidemiology and evolutionary studies, as well as discriminatory power, it is important to produce unambiguous results that are readily comparable among different laboratories, and it is necessary to have a system for classifying the relationships among closely related strains to monitor changes and patterns in clonal lineages over time or space (O'Hara et al., 2016). Of these methods, MLST and spa typing showed highly clonal population structure identified for S. aureus in many previous study (Kanika et al., 2011; Fetsch et al., 2014; Basanisi et al., 2017). As a widely accepted method, MLST, which is based on DNA sequence and relied on analysis of relatively conserved genes that encode essential proteins, has proven very useful for epidemiology and evolutionary studies (Saunders and Holmes, 2007). spa typing is another efficient typing method for S. aureus, it based on sequencing of the polymorphic X region of the protein A gene (spa) (Hallin et al., 2009). It showed highly concordant with MLST. Some studies suggest that it is suitable for epidemiology and evolutionary investigations based on studies of European and international isolates (Koreen et al., 2004; Hallin et al., 2007; Strommenger et al., 2008). However, other studies question the use of a single locus method such as spa typing for epidemiologic investigations since recombination events might distort the underlying clonal relationships (O'Hara et al., 2016). Both two methods can assigned to MLST STs and spa types, which easily to compare with other laboratories in the world.
Of various food products surveyed, meat and meat products are widely known to be an important reservoir for S. aureus and involved in several outbreaks (Aydin et al., 2011; Hennekinne et al., 2012; Wang et al., 2013; Sallam et al., 2015). The investigate of S. aureus from meat can implement a system monitoring. In China, the qualitative and quantitative data of this bacterium in retail meat from different areas were limit. Therefore, the prevalence and levels of S. aureus from South China to North China was to investigate in retail meat and meat products in this study. After isolation and identification, antibiotic susceptibility test, as well as spa typing and MLST were used to determine the genetic background among the S. aureus isolates.
From July 2011 to June 2016, a total of 1,850 retail meat samples were collected from supermarkets, fairs, and farmers' markets. The samples, including 604 raw meat (bacon/sausage, poultry, pork, mutton, and beef), 601 quick-frozen meat (frozen dumpling/steamed stuffed bun, frozen poultry, frozen pork, frozen mutton, and frozen beef), and 645 ready-to-eat (RTE) meat (roast chicken/duck, salt-baked chicken, stewed meat sausage, and ham), were obtained from 39 cities, which covered most of provincial capitals of China (Figure 1). All collected samples were tightly sealed with sterile plastic wrap and placed in a cold box at a temperature lower than 4°C, then transported to an accredited laboratory and subjected to microbiological analysis within 24 h.
Figure 1. The location of sampling sites in China of this study.
The examination of S. aureus proceeded according to GB 4789.10-2010 of food microbiological (National Food Safety Standards of China) with slight modification. Briefly, 25 g of food sample was homogenized in 225 mL of tryptic soy broth with 10% sodium chloride (Huankai, Guangzhou, China). Subsequently, 1, 0.1, and 0.01-mL aliquots were transferred to tubes containing 9, 10, and 10 mL in triplicate with trypticase soy broth (Huankai) supplemented with 10% NaCl. Each tubes were incubated at 37°C for 48 h, respectively. A loopful of enrichment broth culture (10 μL) was streaked onto chromogenic S. aureus agar plates (Huankai) and incubated at 37°C for 24 h. One to four colonies with pink color were purified on nutrient agar medium. The purified colonies were analyzed via coagulase activity test by freeze-dried Rabbit Plasma (Huankai), and the API STAPH test strips (bio Merieux, Marcy-1'Etoile, France) was used. The MPN value was determined on the basis of the number of positive tube(s) in each of the three sets using the MPN table.
All S. aureus isolates were tests for antibiotic susceptibility by the disk diffusion method using Mueller–Hinton agar (Bauer et al., 1966) and commercially available discs (Oxoid, UK). The antimicrobial agents used were amoxycillin/clavulanic acid, ampicillin, cefepime, cefoxitin, penicillin G, ceftazidime, amikacin, gentamicin, kanamycin, streptomycin, chloramphenicol, clindamycin, erythromycin, telithromycin, ciprofloxacin, norfloxacin, tetracycline, linezolid, trimethoprim/sulphamethoxazole 1:19, rifampicin, quinupristin/dalfopristin, teicoplanin, nitrofurantoin, and fusidic acid. MICs for linezolid resistant isolates by disk diffusion were also confirmed by the agar dilution method on Mueller–Hinton agar. The results were scored according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2015). Staphylococcus aureus ATCC25923 and Escherichia coli ATCC25922 were included as a control. The presence of the mecA/mecC gene was studied by PCR in all cefoxitin-resistant isolates (Perez-Roth et al., 2001; Stegger et al., 2012).
All isolates were analyzed by spa typing and MLST for genetic diversity analysis. Primers spa-1113f (5′-TAA AGA CGA TCC TTC GGT GAG C-3′) and spa-1514r (5′-CAG CAG TAG TGC CGT TTG CTT-3′) were used for spa amplification in each strains (Shopsin et al., 1999). The MLST scheme was followed using seven housekeeping genes: arcC, aroE, glpF, gmk, pta, tpi, and yqil. PCR products generated by both techniques were purified with a PCR purification kit (Qiagen, Genman) and were sequenced at an ABI 3730XL sequencer (Applied BioSystems). Spa types were assigned with the SpaServer website (http://spaserver2.ridom.de). The protocols was conducted using previous described primers (Enright et al., 2013). Sequence types (STs) were identified by consulting the S. aureus MLST database (https://pubmlst.org/saureus/). The clonal complex (CC) analysis was performed using the eBURST algorithm as previously described (Feil et al., 2004). The minimum spanning tree (MST) was constructed with Bionumerics 7.6 software (Applied Maths, Sint-Martens-Latem, Belgium).
For statistical analysis, the bacterial numbers were converted to base-10 logarithms. MPN values < 0.3 MPN/g were set to 0.15, and MPN values > 110 MPN/g were assigned the maximum value (Motes et al., 1998). The data of prevalence and levels of S. aureus-positive samples sorted by different food type and sampling sites were compared by using the analysis of variance (SPSS v21.0). Differences between the mean values were significant when P < 0.05.
In this survey, a total of 1,850 samples were collected in 39 Chinese cities. Overall, there were 647 samples showed coagulase positive and confirmed for S. aureus by API STAPH test. The average incidence of S. aureus was 35.0% (647/1,850), the MPN value of the majority of the positive samples ranged from 0.3 to 110 MPN/g, which showed the geometric mean was 10.35 MPN/g. Out of the 647 positive S. aureus samples, 33 (5.1%) exceeded 100 MPN/g, whereas 319 (49.3%) were below 1 MPN/g.
The analyzed food products were classified into three categories (raw meat, quick-frozen meat, and ready-to-eat meat), and the values of S. aureus contamination in each sample were determined (Table 1). Among the analyzed categories, raw meat was the most frequently contaminated with S. aureus, with a prevalence that reached 51.0%. The prevalence of S. aureus in raw poultry, raw mutton, raw beef, raw pork and bacon/sausage was 67.9, 54.5, 50.4, and 18.6%, respectively. The quick-frozen meat showed 43.4% of contamination for S. aureus in this survey, including 60.9% of positive samples in quick-frozen poultry, 50.0% in quick-frozen pork, 31.4% in quick-frozen beef, 30.9% in quick-frozen mutton, and 29.4% in quick-frozen dumpling. The prevalence of S. aureus in RTE meat was lower than that in other samples, which detected 12.2% of RTE samples were positive for S. aureus. S. aureus were detected in 25.0% (3/25) of RTE beef, 12.7% (22/173) of RTE pork, and 11.8% (54/456) of RTE poultry, whereas RTE mutton was free of S. aureus. Combining the MPN values, raw meat showed the highest levels of S. aureus (14.05 MPN/g), followed by RTE meat (9.11 MPN/g) and quick-frozen meat (6.37 MPN/g). Most of the S. aureus-positive samples exhibited less than 10 MPN/g, such as RTE poultry (9.89 MPN/g), quick-frozen poultry (7.37 MPN/g), pork (6.67 MPN/g), RTE pork (6.24 MPN/g), quick-frozen dumpling (4.99 MPN/g), quick-frozen mutton (1.96 MPN/g), quick-frozen pork (1.14 MPN/g), and bacon/sausage (0.59 MPN/g).
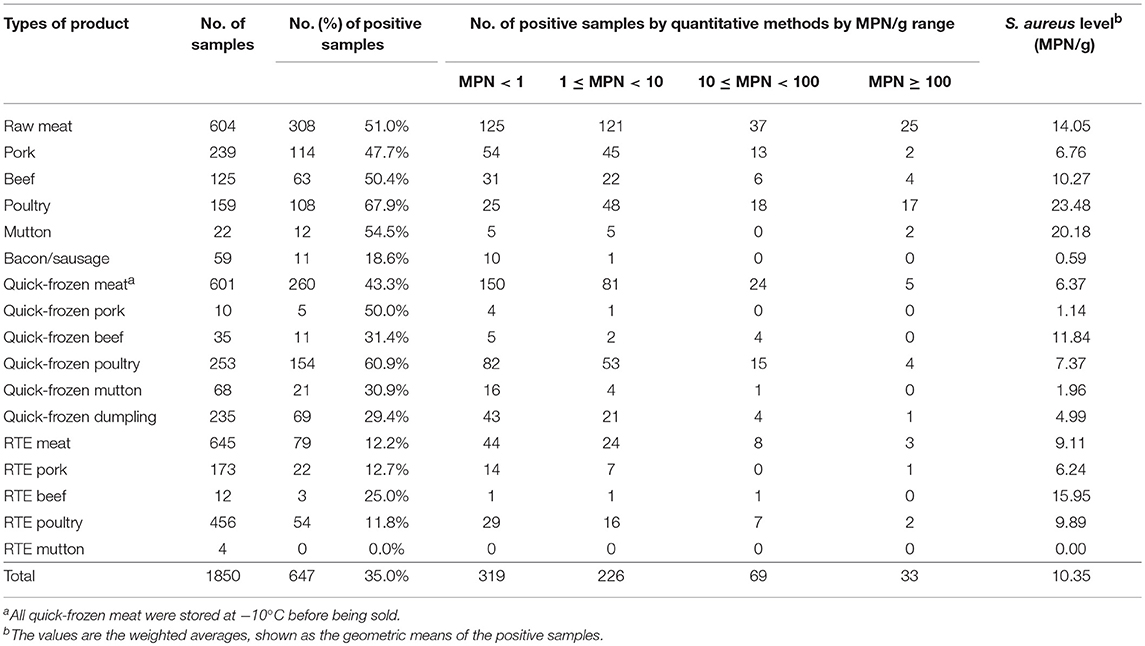
Table 1. Occurrence and levels of Staphylococcus aureus in different types of retail meat.
The distribution of S. aureus among different sampling sites is shown in Table 2. However, all of the 39 cities had S. aureus-positive samples, ranging from 16.7% in Shenzhen to 48.8% in Beihai. The prevalence of S. aureus exceeded 40% in one third of sampling sites. The highest contamination level in Beihai was 20.94 MPN/g, which showed 47.6% (10/21) of the positive samples were exceeded 10 MPN/g, whereas the lowest level in Changsha was only 0.43 MPN/g, which showed 80.0% (8/10) of the positive samples were less than 1 MPN/g. A total of 1,201 samples were collected from South China, with 35.3% prevalence, and 26 positive samples (26/424, 6.1%) had S. aureus densities exceeding the detectable level of 100 MPN/g. In North China, S. aureus was detected in 223 out of 649 samples (34.4%), of which only 7 samples (3.1%) of these samples had levels exceeding 100 MPN/g, whereas 127 samples (57.0%) yielded <1 MPN/g. Thus, there is no significantly difference (p > 0.05, χ2 test) between South China and North China.
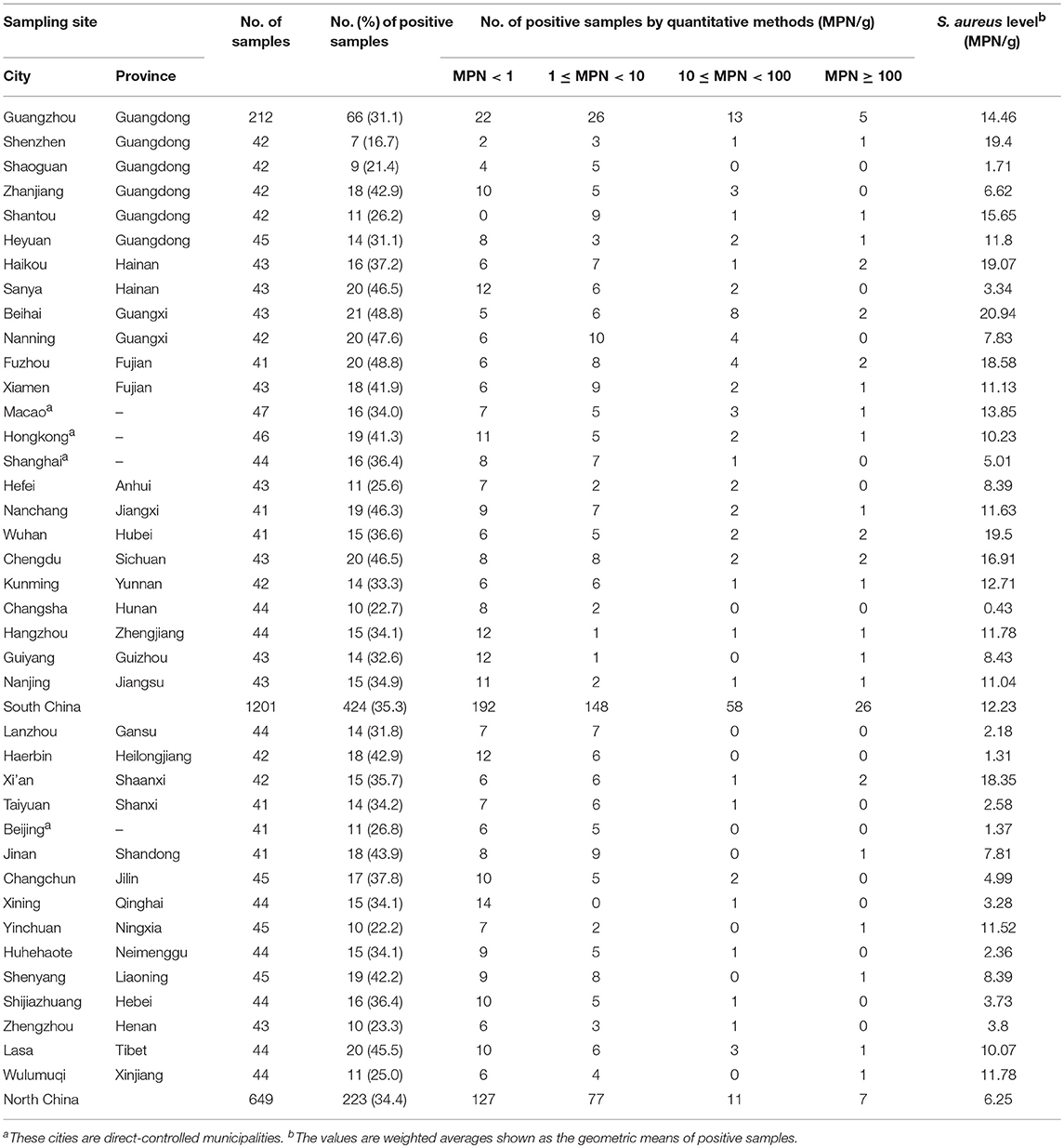
Table 2. Prevalence and levels of Staphylococcus aureus at different sampling sites.
A total of 868 isolates (1–2 isolates per sample) were collected from the retail meat and meat products, consisting of 415 isolates from raw meat, 354 isolates from quick-frozen meat, and 99 isolates from RTE meat. The results of antibiotic susceptibility testing of all isolates are shown in Table 3. Overall, only 11 isolates (1.26%) were susceptible to all 24 tested antibiotics, whereas most isolates (821/868, 94.6%) showed resistance or intermediary resistance to more than three or more antibiotics. Of these isolates, 104 isolates (12.0%) were resistant to more than 10 antibiotics. For 24 antibiotics, 85.4% of isolates were resistant to ampicillin, followed by penicillin G (84.6%), erythromycin (52.7%), tetracycline (49.3%), kanamycin (45.3%), telithromycin (30.1%), clindamycin (29.6%), streptomycin (21.1%), norfloxacin (20.4%), gentamicin (19.4%), fusidic acid (18.4%), ciprofloxacin (16.9%), chloramphenicol (13.1%), amoxycillin/clavulanic acid (11.0%), and others (<10%). There is no significant difference (p > 0.05, χ2 test) between the resistances of types of meat samples for most antimicrobials tested. In total, 95.5, 92.1, and 94.0% of the S. aureus isolates from raw meat, quick-frozen meat, and RTE meat were resistant to at least one antimicrobial. In this study, it was found 62 MRSA isolates (7.14%) which showed resistant to cefoxitin and carried mecA genes, including 34 isolates (8.2%) from raw meat, 20 isolates (5.6%) from quick-frozen meat, and 8 isolates (8.1%) from RTE meat. The MRSA isolates were resistant to most selected β-lactams and other antibiotics, and 77.4% (48/62) of them were resistant to more than 10 antibiotics.
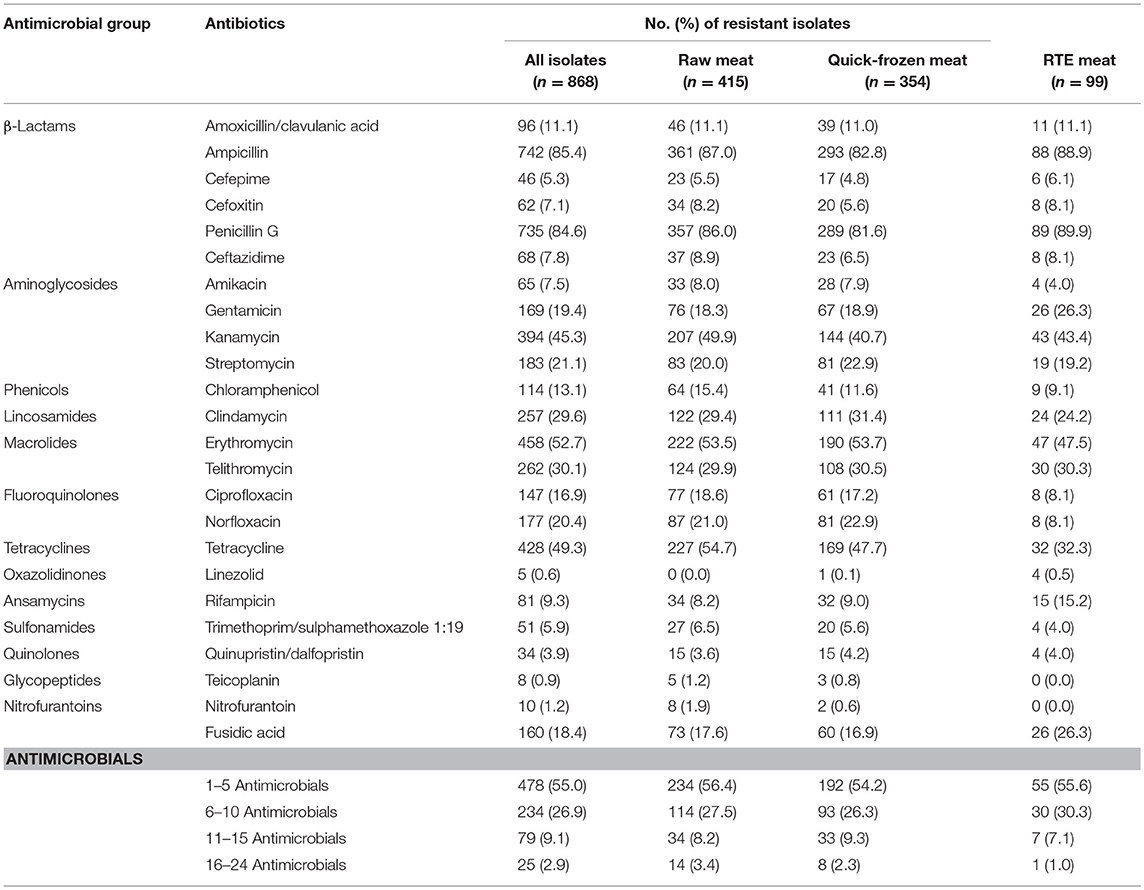
Table 3. Antimicrobial resistance of Staphylococcus aureus isolated from retail meat and meat product in China.
By the spa typing methods, we identified 164 different spa types including 15 novel spa types that were not in the Ridom StaphType database among the 868 isolates tested. Four isolates could not be typed (Table 4). The most commonly observed spa types were t091 (113/868, 13.0%) and t127 (104/868, 12%), followed by t002 (49/868, 5.7%), t189 (47/868, 5.4%), t034 (46/868, 5.3%), t701 (33/868, 3.8%), t437 (30/868, 3.5%), t899 (29/868, 3.3%), t796 (27/868, 3.1%), t084 (19/868, 2.2%), t3092 (15/868, 1.7%), t085 (14/868, 1.6%), t164 (14/868, 1.6%), t1376 (13/868, 1.5%), t213 (9/868, 1.0%), and other spa types (<1.0%).

Table 4. The CCs, STs, and spa types of the 868 S. aureus isolated from retail meat and meat products.
Using MLST there were 111 different sequence types (STs) were detected among the 868 S. aureus isolates analyzed, including 65 newly assigned STs which showed most of them was a single-locus variant (refer to Table 4). Based on eBURST analysis, the 111 STs were grouped into 20 clonal complexes (CCs) and 23 singletons (Table 4). Most CCs (CC1, CC7, CC15, CC5, CC188, CC398, CC6, and CC59) consisted of a central prevalent genotype associated with several much less-frequent single-locus variants (SLVs). The common allelic profiles (STs) was ST7 (151/868, 17.4%), followed by ST1 (124/868, 14.3%), ST5 (73/868, 8.4%), ST398 (59/868, 6.8%), ST188 (48/868, 5.53%), ST6 (42/868, 4.8%), ST59 (42/868, 4.8%), ST88 (41/868, 4.7%), ST15 (35/868, 4.0%), ST9 (30/868, 3.5%), ST72 (18/868, 2.1%), ST25 (15/868, 1.7%), ST943 (14/868, 1.6%), ST12 (12/868, 1.4%), and other STs (<1.0%). Most of the novel STs were represented by either two or one strains.
Combining the STs and spa types, there are 248 different ST-spa types in this research (Table 4). However, most isolates demonstrated high consistent between STs and spa types, such as ST1-t127 (93/868, 10.7%), ST7-t091 (92/868, 10.6%), ST5-t002 (42/868, 4.8%), ST398-t034 (40/868, 4.6%), ST188-t189 (38/868, 4.4%), ST59-t437 (30/868, 3.5%), ST6-t701 (29/868, 3.3%), and ST9-t899 (27/868, 3.1%), and other types. The different sampling regions or food types possibly affected which genotypes were found; for example, Sta353, Sta3802C1, and Sta4117A1 were isolated from different places and belonged to the same ST but were divided into different spa types (ST30-t021, ST30-t964, and ST30-t018). Some strains had the same spa type but differing STs (e.g., t127-ST1, t127-ST12, and t127-ST59).
A phylogenetic tree based on the 7 concatenated MLST sequences shows the relatedness between the strains. The STs of S. aureus isolates were further analyzed based on the sampling sites and food sources (Figures 2A,B). All STs were fell into three different cladogram (designated as A, B, and C) which originate from CC7. Most novel were single-locus variant. Cladogram A contained CC6, CC5, CC1, CC188, ST97, ST20, ST25 and some relevant single-locus variant; Cladogram B included CC398, CC59, ST522, ST30 and some relevant single-locus variant; Cladogram C contained ST72, ST8, ST630, and ST692. It could be found that three different cladogram were evolved in directions, but distribute in different cities and different food sources, which showed high genetic diversity of S. aureus in China. The quick-frozen meat and raw meat were the major types in most of the CCs, whereas RTE meat was dominant for CC72. Interestingly, CC72 was only identified in S. aureus isolated from South China (Guangzhou, Macao, Hong Kong and Haikou).
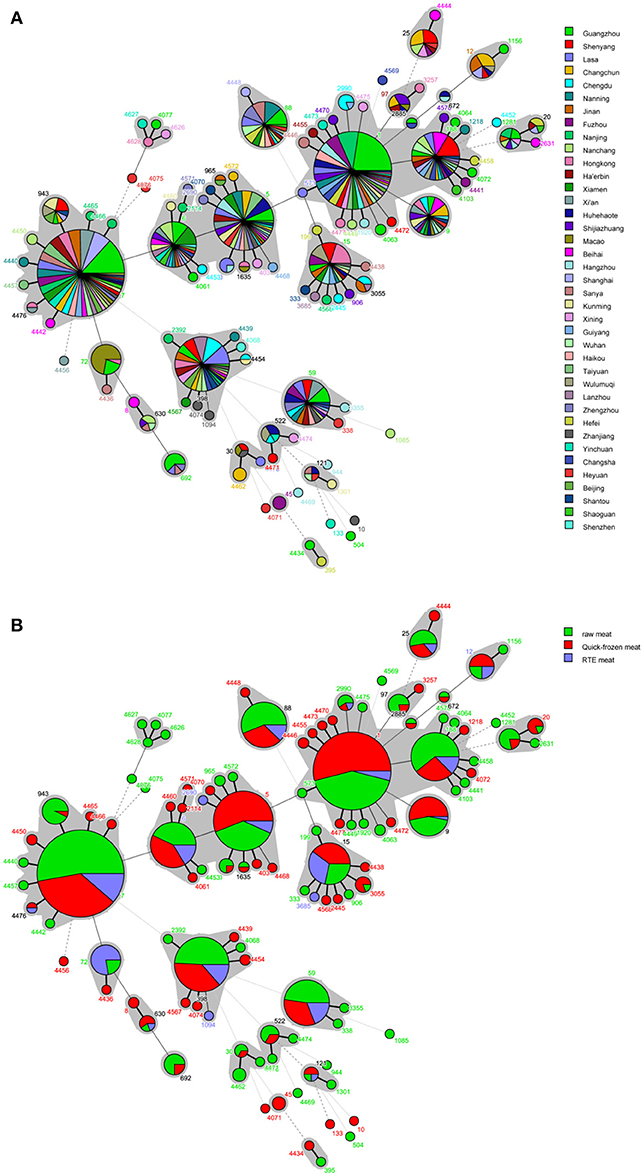
Figure 2. Minimum spanning tree of MLST data for 868 S. aureus isolates. Each circle represents one ST, the size of which is related to the number of strains within this ST, while the branch numbers account for the number of allele differences between connected STs. The colors in the circles in (A) represent the sampling locations, and the colors in the circles in (B) represent the food sources. The gray zones surrounding some STs indicate that these genotypes belong to the same clonal complex.
Recent years, although many studies about the prevalence of S. aureus in retail food have been reported in China (Wang et al., 2012, 2013; Zhang et al., 2013; Chao et al., 2015), the investigation of qualitative and quantitative in national level are lacking. Therefore, a more comprehensively investigation for the incidence of S. aureus isolated from retail meat foods is needed. In this study, samples collected covered all provincial capitals of China, and the food types were also more diversification. Moreover, in conjunction with resistance properties and molecular types, it is better to understand the genetic background of these strains.
Overall, 35% of retail meat and meat products were positive for S. aureus. Over the 39 cities, S. aureus were both confirmed that ranging from 16.7% in Shenzhen to 48.8% in Beihai. This result suggests that the contamination of S. aureus was common in retail meat and meat product in China. This study also determined the contamination level of retail meat in China, showing that the MPN values of the majority of the positive samples between 0.3 and 110 MPN/g, resulting in a average value of 10.35 MPN/g. Among the 647 positive samples, 33 (5.1%) samples had levels beyond 100 MPN/g, whereas 319 (49.3%) had levels below 1 MPN/g. To our knowledge, a limit of 100 cfu/g had been proposed for S. aureus levels in raw meat and RTE meat in China (National Population and Family Planning Commission of the People's Republic of China, 2014). Thus, as a whole, the contamination levels of S. aureus in raw meat in China were not very excessive.
Compared with studies from other countries, the presence of S. aureus in retail meat was higher than the study of Ge et al. (2017) who reported that 27.9% of retail meat samples in the USA were positive for S. aureus, but lower than (Tang et al., 2017)'s research who reported that 68% of samples in Denmark were positive. In other researches, the prevalence of S. aureus in food ranged from 4 to 76.47% (Bakr et al., 2004; Kitai et al., 2005; van Loo et al., 2007; Buyukcangaz et al., 2013; Jackson et al., 2013; Bunnoeng et al., 2014; Dhup et al., 2015). Attribute to the sample sizes, sample types, and geographic locations of investigation, it is maybe the reason for these differences. Based on food categories, S. aureus was found in 51.0%. 43.35 and 12.2% of samples of raw meat, quick-frozen meat, and RTE meat, respectively. This result indicated that the processing can reduce the contamination of S. aureus. There was a previous study which summarized by Ou et al. (2017) in 2017, it was collected 39 studies of the prevalence rates of S. aureus contamination by different types of raw meat products across different periods, regions, and sampling locations and found that the prevalence of S. aureus contamination in chicken products was highest in Asian. In this study, it was similar to this research result, which showed the prevalence of S. aureus contamination in 67.9% of raw poultry and 60.9% of quick-frozen poultry. Besides, the prevalence of S. aureus in raw meat and quick-frozen meat was significantly higher than that in RTE meat (p < 0.001, χ2 test), however, the differences were not significant in the quantitative level (p > 0.05, χ2 test). At present, freezing is an excellent way to preserve food quality and to minimize the incidence of foodborne pathogens (Kumurya, 2013; Wu et al., 2015). Therefore, the cross contamination from environments (such as human, water or transportations) may be the major reason for the presence of S. aureus, which suggests that the Chinese food safety regulators should further improve their hygiene and supervision.
The resistance of S. aureus in this study was higher than that of other food pathogens such as Salmonella, Listeria monocytogenes, Vibrio parahaemolyticus in our previous study (Shi et al., 2015; Wu et al., 2015; Xie et al., 2016; Yang et al., 2016). However, many reports have detected antibiotic-resistant strains of S. aureus in various food products (Normanno et al., 2007; Basanisi et al., 2017; Rong et al., 2017), our data were similar to their results. In the current study, we detected only 11 isolates (1.26%) that were susceptible to all 24 tested antibiotics. The isolates of resistance or intermediary resistance to more than three or more antibiotics were reached 94.6% (821/868). Of which, 104 isolates (12.0%) were resistant to more than 10 antibiotics. In China, (Xing et al., 2014) revealed that 98.4% of S. aureus isolates in ready-to-eat food were resist to more than one antimicrobial agent and 58.6% of isolates resist to more than three antimicrobials, while (Hao et al., 2015) found that 35.9% of S. aureus isolated from quick-frozen dumpling were resistant to less than or equal to 3 kinds of drugs. In Wang et al. (2017) reported that 97.6% of S. aureus isolates from retail food showed resistance in one or more antimicrobial agents. The prevalence of resistance to clindamycin, gentamicin, penicillin, erythromycin, tetracycline, and ciprofloxacin is in agreement with our data. In addition, resistance to kanamycin, erythromycin, and tetracycline was common in our study. As important supplements for animal feed as growth promoters and for the treatment of human disease (Adzitey et al., 2013; Rasha et al., 2018), it maybe the major reason for more and more resistant strains have been commonly observed.
In this study, we also identified 62 MRSA isolates by the disk diffusion method and mecA/C detection. It showed 7.14% of MRSA in S. aureus isolates from retail meat. Most of the MRSA strains were resist to β-lactams and other antibiotics which showed 77.4% (48/62) were resistant to more than 10 antibiotics. This result as similar with the results of research by Wang et al. (2017) and Hao et al. (2015) in China. In fact, the prevalence of MRSA varies greatly by geographical location in retail meats. For example, MRSA was present in 1.9% of 3,520 retail meats in the USA (Ge et al., 2017), 0.5% (13/2,810) in Korea (Kim et al., 2015), 1.6% (5/318) in Spain, 13% in Denmark (19/145) (Tang et al., 2017), 11.9% in the Netherlands (264/2,217), and 24.8% in Canada (655/2,640) (Narvaez et al., 2016). Sample size, geographic area and collection period may responsible for the differences observed (Ge et al., 2017).
The genetic types of 868 S. aureus isolates acquired from raw meat and meat products in China were characterized by MLST and spa typing. Comparing the STs and spa types, it seems that spa typing had a superior distinguishability than MLST. In this study, more than one spa type was detected in the major of STs. This result is consistent with those of previous studies by Hata et al. (2010) and Song et al. (2015). In our study, despite the wide genetic diversity observed among the 868 S. aureus meat isolates, a great proportion of the population belonged to finite number of major clones: ST1, ST7, ST5, ST398, ST188, ST6, ST59, ST88, ST15, and ST9 (74.3%, 645/868) were the dominant STs in S. aureus from retail meat. In concordant with MLST types, t127, t091, t002, t189, t034, t701, t437, t899, t796, t084, t3092, t085, t164, and t1376 (63.7%, 553/868) were the dominant spa types. According to the S. aureus MLST database (https://pubmlst.org/bigsdbdb=pubmlst_saureus_isolates&page=profiles), there were 448 strains of ST1, 54 strains of ST7, 3,993 strains of ST5, 1,412 strains of ST398, 183 strains of ST188, 82 strains of ST6, 172 strains of ST59, 108 strains of ST88, 485 strains of ST15, and 50 strains of ST9 were demonstrating, the origins of them covered US, Canada, Poland, The Netherlands, Denmark, Australia, Bulgaria, Italy, Malaysia, Ivory Coast, France, Japan, Switzerland, and many other countries around world. Most of them isolated from the blood, pus, nasal swab, or skin swab. Beside, these types of S. aureus isolates have been relevant to a variety of clinical infections (Grundmann et al., 2010; Neocleous et al., 2010; Lozano et al., 2011; Valentindomelier et al., 2011; He et al., 2013; Suhaili et al., 2016). it indicates that these types of S. aureus strains have a theoretical pathogenic potential.
In Denmark, Tang et al. (2017) reported that the most frequent MRSA spa type and MSSA spa type were both CC398-t034. In USA, 25.9% of MRSA were identified as ST398-t034 (Hanson et al., 2011). In Germany, Feßler et al. (2011) also found 28 of 32 food related MRSA were CC398. In fact, CC398 strains are usually found in retail chicken, turkey, pork, and beef (Lozano et al., 2009; Huber et al., 2010; Argudín et al., 2011; Verhegghe et al., 2013), and have been perceived as a livestock-associated pathogens. Interestingly, we only found 7 isolates that were MRSA ST398, whereas MSSA ST398 was a common clone (52 isolates) in retail meat, and meat products from China in this study. Currently, infections caused by MSSA ST398 have been reported in humans and shown to cause infections more frequently than MRSA ST398 (Valentindomelier et al., 2011; Mediavilla et al., 2012; David et al., 2013; Li et al., 2015). In addition, the staphylococcal cassette chromosome mec (SCCmec) element types were diverse in MRSA ST398 (Song et al., 2015). In Price et al. (2012) collected 89 CC398 S. aureus isolates (including MSSA and MRSA) from animals and humans spanning 19 countries and four continents by whole-genome sequence typing and found that livestock-associated MRSA CC398 originated as MSSA ST398 in humans. The lineage appears to have undergone a rapid radiation in conjunction with the jump from humans to livestock, where it subsequently acquired tetracycline and methicillin resistance (Price et al., 2012). Therefore, when antibiotic selection in connection with food animal production, it maybe raise the potential chance of MSSA CC398 to acquire the SCCmec cassette (Song et al., 2015), however, further studies should be elucidate the possible rules for regarding MSSA ST398 as well.
In the present study, the majority types of MSSA was ST1-t127 (93/868, 10.7%) and ST7-t091 (92/868, 10.6%). In fact, ST7-t091 and ST1-t127 isolates have been reported as the fourth and sixth most prevalent clone, comprising both MSSA and MRSA strains, isolated from human invasive infections in 26 European countries (Grundmann et al., 2010). In Franco et al. (2011) demonstrated that t127/ST1 isolates can be assigned to two genetically different clusters (porcine and human) and hypothesized that ST1-t127 strains could represent another lineage of livestock-associated pathogens. On the other hand, CC59-t437 (41.9%, 26/62) was the predominant types in retail meat MRSA in China, followed by ST9-t899 (27.4%, 17/62). CC59-t437 is the predominant Asian community-associated MRSA (CA-MRSA) lineage, ranging from 35.8 to 76.7% with CA-MRSA in China (Yang et al., 2017). In Yang et al. (2017) were collected S. aureus strains in Beijing Children's hospital from respiratory tract, skin, and soft tissue, sterile sites in 104 children cases and found 61.7% of CA-MRSA was ST59-SCCmec IV-t437. Except China, ST59 was also reported in Vietnam, Japan, Australia, and other counties for CA-MRSA infection (Tang et al., 2007; Coombs et al., 2010; Higuchi et al., 2010). Therefore, further research will be proceed and found the reason of why ST59-t437 was the predominant types of MRSA isolates in retail meat in our study China. Besides, ST9-t899 was consider that the predominant S. aureus and MRSA genotype in pigs and related workers in Asia (Chuang and Huang, 2015). According to the previous studies, there were many studies have been reported the prevalence of this type of MRSA in Taiwan, Hong Kong, Malaysia, Thailand, and other countries from livestock (Neela et al., 2009; Graveland et al., 2011; Larsen et al., 2012; Lo et al., 2012). Thus, the majority types are different than those in studies in other counties. However, as well as livestock-associated lineages and clinical S. aureus clones, it revealed that these strains were widespread from retail meat and meat products in China. The pathogenic potential of these strains from retail meat food in China should not be ignored.
In conclusion, we report a wide scale and systematic investigation of S. aureus from retail meat and meat products in China, it supplements the nationwide qualitative and quantitative data of the prevalence and levels of S. aureus. The contamination of S. aureus was common in retail meat in China, but the levels of S. aureus were not very excessive. The prevalence rate of S. aureus in raw meat and quick-frozen meat was significantly higher than that in RTE meat. Most S. aureus isolates exhibited resistance to a variety of antimicrobials. By molecular typing analysis showed that these isolates had a high genetic diversity. The majority of these types have been linked to human infections worldwide, indicating that S. aureus strains of these types in retail meat related S. aureus in China have at least a theoretical pathogenic potential. However, the frequently types in our study are different than those in studies in other counties. Thus, further studies may need to elucidate the potential origins of these strains in China.
QW, JZ, SW, and TL conceived and designed the experiments. JH and FZ performed the experiments. SW, HW, and HZ analyzed the data. XY, LX, MC, YD, and SZ contributed reagents, materials, analysis tools. SW, JW, and QW contributed to the writing of the manuscript.
We would like to acknowledge the financing support of the National Key R&D Program of China (2016YFD0401204), National Natural Science Foundation of China (No. 31801657), China's Post-doctoral Science Fund (2017M612623), and GDAS' Special Project of Science and Technology Development (2017GDASCX-0201, 2017GDASCX-0817).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Adzitey, F., Ali, G. R. R., Huda, N., Cogan, T., and Corry, J. (2013). Prevalence, antibiotic resistance and genetic diversity of Listeria monocytogenes isolated from ducks, their rearing and processing environments in Penang, Malaysia. Food Control 32, 607–614. doi: 10.1016/j.foodcont.2012.12.016
Argudín, M. A., Tenhagen, B. A., Fetsch, A., Sachsenröder, J., Käsbohrer, A., Schroeter, A., et al. (2011). Virulence and resistance determinants of german Staphylococcus aureus ST398 isolates from nonhuman sources. Appl. Environ. Microbiol. 77, 3052–3060. doi: 10.1128/AEM.02260-10
Aydin, A., Sudagidan, M., and Muratoglu, K. (2011). Prevalence of staphylococcal enterotoxins, toxin genes and genetic-relatedness of foodborne Staphylococcus aureus strains isolated in the Marmara Region of Turkey. Int. J. Food Microbiol. 148, 99–106. doi: 10.1016/j.ijfoodmicro.2011.05.007
Bakr, W. M., Fawzi, M., and Hashish, M. H. (2004). Detection of coagulase positive staphylococci in meat products sold in Alexandria using two different media. J. Egypt Public Health Assoc. 79, 31–42.
Basanisi, M. G., Bella, G. L., Nobili, G., Franconieri, I., and Salandra, G. L. (2017). Genotyping of methicillin-resistant Staphylococcus aureus (MRSA) isolated from milk and dairy products in South Italy. Food Microbiol. 62, 141–146. doi: 10.1016/j.fm.2016.10.020
Bauer, A., Kirby, W., Sherris, J. C., and Turck, M. (1966). Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 45:493. doi: 10.1093/ajcp/45.4_ts.493
Behling, R. G., Eifert, J., Erickson, M. C., Gurtler, J. B., Kornacki, J. L., Line, E., et al. (2010). “Selected pathogens of concern to industrial food processors: infectious, toxigenic, toxico-infectious, selected emerging pathogenic bacteria,” in Principles of Microbiological Troubleshooting in the Industrial Food Processing Environment, ed J. L. Kornacki (New York, NY: Springer), 5–61.
Bunnoeng, N., Themphachana, M., Pewleang, T., Kongpheng, S., Singkhamanan, K., Saengsuwan, P., et al. (2014). High prevalence and molecular characterization of methicillin-resistant Staphylococcus aureus isolated from retailed meats, south Thailand. Int. Food Res. J. 21, 569–576.
Buyukcangaz, E., Velasco, V., Sherwood, J. S., Stepan, R. M., Koslofsky, R. J., and Logue, C. M. (2013). Molecular typing of Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) isolated from animals and retail meat in North Dakota, United States. Foodborne Pathog. Dis. 10, 608–617. doi: 10.1089/fpd.2012.1427
Chao, G., Bao, G., Cao, Y., Yan, W., Wang, Y., Zhang, X., et al. (2015). Prevalence and diversity of enterotoxin genes with genetic background of Staphylococcus aureus isolates from different origins in China. Int. J. Food Microbiol. 211, 142–147. doi: 10.1016/j.ijfoodmicro.2015.07.018
Chuang, Y. Y., and Huang, Y. C. (2015). Livestock-associated meticillin-resistant Staphylococcus aureus in Asia: an emerging issue? Int. J. Antimicrob. Agents 45, 334–340. doi: 10.1016/j.ijantimicag.2014.12.007
Coombs, G. W., Monecke, S., Ehricht, R., Slickers, P., Pearson, J. C., Tan, H.-L., et al. (2010). Differentiation of clonal complex 59 community-associated methicillin-resistant Staphylococcus aureus in Western Australia. Antimicrobial Agents and Chemotherapy 54, 1914–1921. doi: 10.1128/AAC.01287-09
David, M. Z., Siegel, J., Lowy, F. D., Zychowski, D., Taylor, A., Lee, C. J., et al. (2013). Asymptomatic carriage of sequence type 398, spa type t571 methicillin-susceptible staphylococcus aureus in an urban jail: a newly emerging, transmissible pathogenic strain. J. Clin. Microbiol. 51, 2443–2447. doi: 10.1128/JCM.01057-13
Dhup, V., Kearns, A. M., Pichon, B., and Foster, H. A. (2015). First report of identification of livestock-associated MRSA ST9 in retail meat in England. Epidemiol. Infect. 143, 2989–2992. doi: 10.1017/S0950268815000126
Diep, B. A., Gill, S. R., Chang, R. F., Phan, T. H., Chen, J. H., Davidson, M. G., et al. (2006). Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 367, 731–739. doi: 10.1016/S0140-6736(06)68231-7
Enright, M. C., Day, N. P. J., Davies, C. E., Peacock, S. J., and Spratt, B. G. (2013). Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 51, 306–310. doi: 10.1128/JCM.02421-12
Feil, E. J., Li, B. C., Aanensen, D. M., Hanage, W. P., and Spratt, B. G. (2004). eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J. Bacteriol. 186, 1518–1530. doi: 10.1128/JB.186.5.1518-1530.2004
Feßler, A. T., Kadlec, K., Hassel, M., Hauschild, T., Eidam, C., Ehricht, R., et al. (2011). Characterization of methicillin-resistant Staphylococcus aureus isolates from food and food products of poultry origin in Germany. Appl. Environ. Microbiol. 77, 7151–7157. doi: 10.1128/AEM.00561-11
Fetsch, A., Contzen, M., Hartelt, K., Kleiser, A., Maassen, S., Rau, J., et al. (2014). Staphylococcus aureus food-poisoning outbreak associated with the consumption of ice-cream. Int. J. Food Microbiol. 187, 1–6. doi: 10.1016/j.ijfoodmicro.2014.06.017
Franco, A., Hasman, H., Iurescia, M., Lorenzetti, R., Stegger, M., Pantosti, A., et al. (2011). Molecular characterization of spa type t127, sequence type 1 methicillin-resistant Staphylococcus aureus from pigs. J. Antimicrob. Chemother. 66, 1231–1235. doi: 10.1093/jac/dkr115
Ge, B., Mukherjee, S., Hsu, C. H., Davis, J. A., Tran, T. T. T., Yang, Q., et al. (2017). MRSA and multidrug-resistant Staphylococcus aureus in U.S. retail meats, 2010–2011. Food Microbiol. 62, 289–297. doi: 10.1016/j.fm.2016.10.029
Gharsa, H., Ben, S. K., Lozano, C., Gómezsanz, E., Klibi, N., Ben, S. R., et al. (2012). Prevalence, antibiotic resistance, virulence traits and genetic lineages of Staphylococcus aureus in healthy sheep in Tunisia. Vet. Microbiol. 156, 367–373. doi: 10.1016/j.vetmic.2011.11.009
Govindaraj, A. V., and Vanitha, A. (2018). WHO global priority pathogens list on antibiotic resistance: an urgent need for action to integrate one health data. Perspect. Public Health 138, 87–88. doi: 10.1177/1757913917743881
Graveland, H., Duim, B., Van, D. E., Heederik, D., and Wagenaar, J. A. (2011). Livestock-associated methicillin-resistant Staphylococcus aureus in animals and humans. Int. J. Med. Microbiol. 301, 630–634. doi: 10.1016/j.ijmm.2011.09.004
Grundmann, H., Aanensen, D. M., van den Wijngaard, C. C., Spratt, B. G., Harmsen, D., and Friedrich, A. W. (2010). Geographic distribution of Staphylococcus aureus causing invasive infections in Europe: a molecular-epidemiological analysis. PLoS Med. 7:e1000215. doi: 10.1371/journal.pmed.1000215
Hallin, M., Deplano, A., Denis, O., De, M. R., De, R. R., and Struelens, M. J. (2007). Validation of pulsed-field gel electrophoresis and spa typing for long-term, nationwide epidemiological surveillance studies of Staphylococcus aureus infections. J. Clin. Microbiol. 45:127. doi: 10.1128/JCM.01866-06
Hallin, M., Friedrich, A. W., and Struelens, M. J. (2009). spa typing for epidemiological surveillance of Staphylococcus aureus. Methods Mol. Biol. 551, 189–202. doi: 10.1007/978-1-60327-999-4_15
Hanson, B. M., Dressler, A. E., Harper, A. L., Scheibel, R. P., Wardyn, S. E., Roberts, L. K., et al. (2011). Prevalence of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus (MRSA) on retail meat in Iowa. J. Infect. Public Health 4, 169–174. doi: 10.1016/j.jiph.2011.06.001
Hao, D., Xing, X., Li, G., Wang, X., Zhang, M., Zhang, W., et al. (2015). Prevalence, toxin gene profiles, and antimicrobial resistance of Staphylococcus aureus isolated from quick-frozen dumplings. J. Food Protect. 78, 218–223. doi: 10.4315/0362-028X.JFP-14-100
Hata, E., Katsuda, K., Kobayashi, H., Uchida, I., Tanaka, K., and Eguchi, M. (2010). Genetic variation among Staphylococcus aureus strains from bovine milk and their relevance to methicillin-resistant isolates from humans. J. Clin. Microbiol. 48, 2130–2139. doi: 10.1128/JCM.01940-09
He, W., Chen, H., Zhao, C., Zhang, F., Li, H., Wang, Q., et al. (2013). Population structure and characterisation of Staphylococcus aureus from bacteraemia at multiple hospitals in China: association between antimicrobial resistance, toxin genes and genotypes. Int. J. Antimicrob. Agents 42, 211–219. doi: 10.1016/j.ijantimicag.2013.04.031
Hennekinne, J. A., De Buyser, M. L., and Dragacci, S. (2012). Staphylococcus aureus and its food poisoning toxins: characterization and outbreak investigation. Fems Microbiol. Rev. 36, 815–836. doi: 10.1111/j.1574-6976.2011.00311.x
Higuchi, W., Hung, W. C., Takano, T., Iwao, Y., Ozaki, K., Isobe, H., et al. (2010). Molecular characteristics of the Taiwanese multiple drug-resistant ST59 clone of Panton-Valentine leucocidin-positive community-acquired methicillin-resistant Staphylococcus aureus from pediatric cellulitis. J. Infect. Chemother. 16, 144–149. doi: 10.1007/s10156-010-0029-9
Huang, B., Deng, Z., Tan, M., and Liao, Y. (2009). Contamination status, staphylococcal enterotoxin-producing property and drug resistance of Staphylococcus aureus isolated from food products. Chin. J. Health Lab. Technol. 19, 1380–1382.
Huber, H., Koller, S., Giezendanner, N., Stephan, R., and Zweifel, C. (2010). Prevalence and characteristics of meticillin-resistant Staphylococcus aureus in humans in contact with farm animals, in livestock, and in food of animal origin, Switzerland, 2009. Euro Surveill. 15:19542. doi: 10.5167/uzh-41122
Jackson, C. R., Davis, J. A., and Barrett, J. B. (2013). Prevalence and characterization of methicillin-resistant Staphylococcus aureus isolates from retail meat and humans in Georgia. J. Clin. Microbiol. 51, 1199–1207. doi: 10.1128/JCM.03166-12
Kanika, B., Wang, X., Susan, D., Marcus, Z., Da, R. L., and Zhang, Y. (2011). Methicillin-resistant Staphylococcus aureus in retail meat, detroit, Michigan, USA. Emerg. Infect. Dis. 17, 1135–1137. doi: 10.3201/eid/1706.101905
Kim, Y. J., Oh, D. H., Song, B. R., Heo, E. J., Lim, J. S., Moon, J. S., et al. (2015). Molecular characterization, antibiotic resistance, and virulence factors of methicillin-resistant Staphylococcus aureus strains isolated from imported and domestic meat in Korea. Foodborne Pathog. Dis. 12, 390–398. doi: 10.1089/fpd.2014.1885
Kitai, S., Shimizu, A., Kawano, J., Sato, E., Nakano, C., Kitagawa, H., et al. (2005). Prevalence and characterization of Staphylococcus aureus and enterotoxigenic Staphylococcus aureus in retail raw chicken meat throughout Japan. J. Vet. Med. Sci. 67, 269–274. doi: 10.1292/jvms.67.269
Koreen, L., Ramaswamy, S. V., Graviss, E. A., Naidich, S., Musser, J. M., and Kreiswirth, B. N. (2004). spa typing method for discriminating among Staphylococcus aureus isolates: implications for use of a single marker to detect genetic micro- and macrovariation. J. Clin. Microbiol. 42, 792. doi: 10.1128/JCM.42.2.792-799.2004
Kumurya, A. S. (2013). Loss of the meca gene during storage of methicillin-resistant Staphylococcus aureus isolates in Northwestern Nigeria Acad. J. 5, 410–415. doi: 10.5897/JPHE12.105
Larsen, J., Imanishi, M., Hinjoy, S., Tharavichitkul, P., Duangsong, K., Davis, M. F., et al. (2012). Methicillin-resistant Staphylococcus aureus ST9 in pigs in thailand. PLoS ONE 7:e31245. doi: 10.1371/journal.pone.0031245
Li, G., Wu, C., Wang, X., and Meng, J. (2015). Prevalence and characterization of methicillin susceptible Staphylococcus aureus ST398 isolates from retail foods. Int. J. Food Microbiol. 196, 94–97. doi: 10.1016/j.ijfoodmicro.2014.12.002
Lo, Y. P., Wan, M. T., Chen, M. M., Su, H. Y., Lauderdale, T. L., and Chou, C. C. (2012). Molecular characterization and clonal genetic diversity of methicillin-resistant Staphylococcus aureus of pig origin in Taiwan. Compar. Immunol. Microbiol. Infect. Dis. 35, 513–521. doi: 10.1016/j.cimid.2012.05.001
Lozano, C., Aspiroz, C., Lasarte, J. J., Gómez-Sanz, E., Zarazaga, M., and Torres, C. (2011). Dynamic of nasal colonization by methicillin-resistant Staphylococcus aureus ST398 and ST1 after mupirocin treatment in a family in close contact with pigs. Compar. Immunol. Microbiol. Infect. Dis. 34, e1–e7. doi: 10.1016/j.cimid.2010.06.006
Lozano, C., López, M., Gómezsanz, E., Ruizlarrea, F., Torres, C., and Zarazaga, M. (2009). Detection of methicillin-resistant Staphylococcus aureus ST398 in food samples of animal origin in Spain. J. Antimicrob. Chemother. 64, 1325–1326. doi: 10.1093/jac/dkp378
Mccallum, N., Bergerbächi, B., and Senn, M. M. (2010). Regulation of antibiotic resistance in Staphylococcus aureus. Int. J. Med. Microbiol. 300, 118–129. doi: 10.1016/j.ijmm.2009.08.015
Mediavilla, J. R., Chen, L., Uhlemann, A. C., Hanson, B. M., Rosenthal, M., Stanak, K., et al. (2012). Methicillin-susceptible Staphylococcus aureus ST398, New York and New Jersey, USA. Emerg. Infect. Dis. 18, 700–702. doi: 10.3201/eid1804.111419
Motes, M., DePaola, A., Cook, D., Veazey, J., Hunsucker, J., Garthright, W., et al. (1998). Influence of water temperature and salinity onvibrio vulnificus in northern gulf and atlantic coast oysters (Crassostrea virginica). Appl. Environ. Microbiol. 64, 1459–1465.
Narvaez, B. C., Toufeer, M., Weese, S. J., Diarra, M. S., Deckert, A. E., Reidsmith, R., et al. (2016). Prevalence of methicillin-resistant Staphylococcus aureus in canadian commercial pork processing plants. J. Appl. Microbiol. 120, 770–780. doi: 10.1111/jam.13024
National Population and Family Planning Commission of the People's Republic of China. (2014). The Standard of limit for foodborne pathogens-GB29921-2013. Capit. J. Publ. Health 2, 34–36.
Neela, V., Zafrul, A. M., Mariana, N. S., Belkum, A. V., Yun, K. L., and Rad, E. G. (2009). Prevalence of ST9 methicillin-resistant Staphylococcus aureus among pigs and pig handlers in Malaysia. J. Clin. Microbiol. 47, 4138–4140. doi: 10.1128/JCM.01363-09
Neocleous, C., Damani, A., Gerogianni, I., Gourgoulianis, K., and Petinaki, E. (2010). Necrotizing pneumonia in Greece caused by a USA400 (ST1) Staphylococcus aureus harboring SSCmec type V. Infection 38, 76–77. doi: 10.1007/s15010-009-9199-8
Normanno, G., La, S. G., Dambrosio, A., Quaglia, N. C., Corrente, M., Parisi, A., et al. (2007). Occurrence, characterization and antimicrobial resistance of enterotoxigenic Staphylococcus aureus isolated from meat and dairy products. Int. J. Food Microbiol. 115, 290–296. doi: 10.1016/j.ijfoodmicro.2006.10.049
O'Hara, F. P., Suaya, J. A., Ray, G. T., Baxter, R., Brown, M. L., Mera, R. M., et al. (2016). spa Typing and multilocus sequence typing show comparable performance in a macroepidemiologic study of Staphylococcus aureus in the United States. Microb. Drug Resist. 22, 88–96. doi: 10.1089/mdr.2014.0238
Ou, Q., Peng, Y., Lin, D., Bai, C., Zhang, T., Lin, J., et al. (2017). A meta-analysis of the global prevalence rates of Staphylococcus aureus and methicillin-resistant, S. aureus contamination of different raw meat products. Journal of Food Protection 80, 763–774. doi: 10.4315/0362-028X.JFP-16-355
Papadopoulos, P., Papadopoulos, T., Angelidis, A. S., Boukouvala, E., Zdragas, A., Papa, A., et al. (2018). Prevalence of Staphylococcus aureus and of methicillin-resistant S. aureus (MRSA) along the production chain of dairy products in north-western Greece. Food Microbiol. 69, 43–50. doi: 10.1016/j.fm.2017.07.016
Perez-Roth, E., Claverie-Martin, F., Villar, J., and Mendez-Alvarez, S. (2001). Multiplex PCR for simultaneous identification of Staphylococcus aureus and detection of methicillin and mupirocin resistance. J. Clin. Microbiol. 39, 4037–4041. doi: 10.1128/JCM.39.11.4037-4041.2001
Price, L. B., Stegger, M., Hasman, H., Aziz, M., Larsen, J., Andersen, P. S., et al. (2012). Staphylococcus aureus CC398: host adaptation and emergence of methicillin resistance in livestock. Mbio 3, e00305–00311. doi: 10.1128/mBio.00305-11
Rasha, I. M., Mohamed, A. A., and Heba, M. A. (2018). Virulence and antimicrobial susceptibility profile of Listeria monocytogenes isolated from frozen vegetables available in the Egyptian market. Afr. J. Microbiol. Res. 12, 218–224. doi: 10.5897/AJMR2018.8794
Rong, D., Xu, M., Wu, Q., Zhang, J., and Yu, S. (2017). Prevalence, virulence genes, antimicrobial susceptibility, and genetic diversity of Staphylococcus aureus from retail aquatic products in China. Front. Microbiol. 8:714. doi: 10.3389/fmicb.2017.00714
Sallam, K. I., Abd-Elghany, S. M., Elhadidy, M., and Tamura, T. (2015). Molecular characterization and antimicrobial resistance profile of methicillin-resistant Staphylococcus aureus in retail chicken. J. Food Protect. 78, 1879–1884. doi: 10.4315/0362-028X.JFP-15-150
Sauer, P., Síla, J., Stosová, T., Vecerová, R., Hejnar, P., Vágnerová, I., et al. (2008). Prevalence of genes encoding extracellular virulence factors among meticillin-resistant Staphylococcus aureus isolates from the University Hospital, Olomouc, Czech Republic. J. Med. Microbiol. 57, 403–410. doi: 10.1099/jmm.0.47413-0
Saunders, N. A., and Holmes, A. (2007). Multilocus sequence typing (MLST) of Staphylococcus aureus. Methods Mol. Biol. 391, 71–85. doi: 10.1007/978-1-59745-468-1_6
Scallan, E., Hoekstra, R. M., Angulo, F. J., Tauxe, R. V., Widdowson, M.-A., Roy, S. L., et al. (2011). Foodborne illness acquired in the United States—major pathogens. Emerg. Infect. Dis. 17, 7–15. doi: 10.3201/eid1701.P11101
Shi, W., Wu, Q., Zhang, J., Chen, M., and Yan, Z. (2015). Prevalence, antibiotic resistance and genetic diversity of Listeria monocytogenes isolated from retail ready-to-eat foods in China. Food Control 47, 340–347. doi: 10.1016/j.foodcont.2014.07.028
Shopsin, B., Gomez, M., Montgomery, S. O., Smith, D. H., Waddington, M., Dodge, D. E., et al. (1999). Evaluation of protein A gene polymorphic region DNA sequencing for typing of Staphylococcus aureus strains. J. Clin. Microbiol. 37, 3556–3563.
Song, M., Bai, Y., Xu, J., Carter, M. Q., Shi, C., and Shi, X. (2015). Genetic diversity and virulence potential of Staphylococcus aureus isolates from raw and processed food commodities in Shanghai. Int. J. Food Microbiol. 195, 1–8. doi: 10.1016/j.ijfoodmicro.2014.11.020
Stegger, M., Andersen, P. S., Kearns, A., Pichon, B., Holmes, M. A., Edwards, G., et al. (2012). Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecA LGA251. Clin. Microbiol. Infect. 18, 395–400. doi: 10.1111/j.1469-0691.2011.03715.x
Strommenger, B., Braulke, C., Heuck, D., Schmidt, C., Pasemann, B., Nübel, U., et al. (2008). spa typing of Staphylococcus aureus as a frontline tool in epidemiological typing. J. Clin. Microbiology 46, 574–581. doi: 10.1128/JCM.01599-07
Suhaili, Z., Lean, S. S., Mohamad, N. M., Rachman, A. R. A., Desa, M. N. M., and Yeo, C. C. (2016). Draft genome sequence of Staphylococcus aureus KT/312045, an ST1-MSSA PVL positive isolated from pus sample in East Coast Malaysia. Genomics Data 9, 111–112. doi: 10.1016/j.gdata.2016.07.002
Tang, C. T., Tho, N. D., Hoa, N. T., Phuong, N. T. M., Le, V. T., Diep, T. S., et al. (2007). An outbreak of severe infections with community-acquired MRSA carrying the panton-valentine leukocidin following vaccination. PLoS ONE 2:e822. doi: 10.1371/journal.pone.0000822
Tang, Y., Larsen, J., Kjeldgaard, J., Andersen, P. S., Skov, R., and Ingmer, H. (2017). Methicillin-resistant and -susceptible Staphylococcus aureus from retail meat in Denmark. Int. J. Food Microbiol. 249, 72–76. doi: 10.1016/j.ijfoodmicro.2017.03.001
The Clinical and Laboratory Standards Institute (CLSI) (2015). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. Approved Standard-M02-A12. Wayne, PA: The Clinical and Laboratory Standards Institute.
Valentindomelier, A. S., Girard, M., Bertrand, X., Violette, J., François, P., Donnio, P. Y., et al. (2011). Methicillin-susceptible ST398 Staphylococcus aureus responsible for bloodstream infections: an emerging human-adapted subclone? PLoS ONE 6:e28369. doi: 10.1371/journal.pone.0028369
van Loo, I. H., Diederen, B. M., Savelkoul, P. H., Woudenberg, J. H., Roosendaal, R., Van, B. A., et al. (2007). Methicillin-resistant Staphylococcus aureus in meat products, the Netherlands. Emerg. Infect. Dis. 13, 1753–1755. doi: 10.3201/eid1311.070358
Verhegghe, M., Pletinckx, L. J., Crombé, F., Vandersmissen, T., Haesebrouck, F., Butaye, P., et al. (2013). Methicillin-resistant Staphylococcus aureus (MRSA) ST398 in pig farms and multispecies farms. Zoonoses Public Health 60, 366–374. doi: 10.1111/zph.12007
Wang, W., Baloch, Z., Jiang, T., Zhang, C., Peng, Z., Li, F., et al. (2017). Enterotoxigenicity and antimicrobial resistance of Staphylococcus aureus isolated from retail food in China. Front. Microbiol. 8:2256. doi: 10.3389/fmicb.2017.02256
Wang, X., Meng, J., Zhang, J., Zhou, T., Zhang, Y., Yang, B., et al. (2012). Characterization of Staphylococcus aureus isolated from powdered infant formula milk and infant rice cereal in China. Int. J. Food Microbiol. 153, 142–147. doi: 10.1016/j.ijfoodmicro.2011.10.030
Wang, X., Tao, X., Xia, X., Yang, B., Xi, M., Meng, J., et al. (2013). Staphylococcus aureus and methicillin-resistant Staphylococcus aureus in retail raw chicken in China. Food Control 29, 103–106. doi: 10.1016/j.foodcont.2012.06.002
Wei-Wei, L. I., Zhu, J. H., Zhen, S. Q., Liang, X. C., Jiang, Y. Y., Ning, L. I., et al. (2018). Analysis of foodborne disease outbreaks in China mainland in 2011. Chin. J. Food Hygiene. 30, 283–288.
Wu, S., Huang, J., Wu, Q., Zhang, F., Zhang, J., Lei, T., et al. (2018). Prevalence and characterization of Staphylococcus aureus isolated from retail vegetables in China. Front. Microbiol. 9:1263. doi: 10.3389/fmicb.2018.01263
Wu, S., Wu, Q., Zhang, J., Chen, M., Yan, Z. A., and Hu, H. (2015). Listeria monocytogenes prevalence and characteristics in retail raw foods in China. PLoS ONE 10:e0136682. doi: 10.1371/journal.pone.0136682
Xie, T., Xu, X., Wu, Q., Zhang, J., and Cheng, J. (2016). Prevalence, molecular characterization, and antibiotic susceptibility of vibrio parahaemolyticus from ready-to-eat foods in China. Front. Microbiol. 7:549. doi: 10.3389/fmicb.2016.00549
Xing, X., Li, G., Zhang, W., Wang, X., Xia, X., Yang, B., et al. (2014). Prevalence, antimicrobial susceptibility, and enterotoxin gene detection of Staphylococcus aureus isolates in ready-to-eat foods in Shaanxi, People's Republic of China. J. Food Protect. 77, 331–334. doi: 10.4315/0362-028X.JFP-13-301
Yang, X., Huang, J., Wu, Q., Zhang, J., Liu, S., Guo, W., et al. (2016). Prevalence, antimicrobial resistance and genetic diversity of Salmonella isolated from retail ready-to-eat foods in China. Food Control 60, 50–56. doi: 10.1016/j.foodcont.2015.07.019
Yang, X., Qian, S., Yao, K., Wang, L., Liu, Y., Dong, F., et al. (2017). Multiresistant ST59-SCCmecIV-t437 clone with strong biofilm-forming capacity was identified predominantly in MRSA isolated from Chinese children. BMC Infect. Dis. 17:733. doi: 10.1186/s12879-017-2833-7
Keywords: S. aureus, retail meat, antimicrobial resistance, MLST, spa typing
Citation: Wu S, Huang J, Wu Q, Zhang J, Zhang F, Yang X, Wu H, Zeng H, Chen M, Ding Y, Wang J, Lei T, Zhang S and Xue L (2018) Staphylococcus aureus Isolated From Retail Meat and Meat Products in China: Incidence, Antibiotic Resistance and Genetic Diversity. Front. Microbiol. 9:2767. doi: 10.3389/fmicb.2018.02767
Received: 17 July 2018; Accepted: 29 October 2018;
Published: 15 November 2018.
Edited by:
Qingli Dong, University of Shanghai for Science and Technology, ChinaReviewed by:
Haike Antelmann, Freie Universität Berlin, GermanyCopyright © 2018 Wu, Huang, Wu, Zhang, Zhang, Yang, Wu, Zeng, Chen, Ding, Wang, Lei, Zhang and Xue. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qingping Wu, d3VxcDIwM0AxNjMuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.