
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 29 October 2018
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.02587
 Mahbobeh Montazeri1,2
Mahbobeh Montazeri1,2 Saeed Mehrzadi3
Saeed Mehrzadi3 Mehdi Sharif1,4
Mehdi Sharif1,4 Shahabeddin Sarvi1,5
Shahabeddin Sarvi1,5 Asal Tanzifi1,2
Asal Tanzifi1,2 Sargis A. Aghayan6
Sargis A. Aghayan6 Ahmad Daryani1,5*
Ahmad Daryani1,5*Toxoplasma gondii (T. gondii) is a global protozoan parasite infecting up to one-third of the world population. Pyrimethamine (PYR) and sulfadiazine (SDZ) are the most widely used drugs for treatment of toxoplasmosis; however, several failure cases have been recorded as well; suggesting the existence of drug resistant strains. This review aims to give a systematic and comprehensive understanding of drug resistance in T. gondii including mechanisms of resistance and sites of drug action in parasite. Analogous amino acid substitutions in the Toxoplasma enzyme were identified to confer PYR resistance. Moreover, resistance to clindamycin, spiramycin, and azithromycin is encoded in the rRNA genes of T. gondii. However, T. gondii SDZ resistance mechanism has not been proved yet. Recently there has been a slight increase in SDZ resistance. That is why the majority of studies were carried out using SDZ. Six strains resistant to SDZ were found in clinical cases between 2013 and 2017 which among Brazilian T. gondii isolates, TgCTBr11, Ck3, and Pg1 were identified in human toxoplasmosis, as well as in livestock intended for human consumption. In conclusion, recent experimental studies in clinical cases have clearly shown that drug resistance in Toxoplasma is ongoing. Thus, establishing a more effective therapeutic scheme in the treatment of toxoplasmosis is critically needed. The emergence of T. gondii strains resistant to current drugs, reviewed here, represents a concern not only for treatment failure but also for increased clinical severity in immunocompromised patients. To improve the therapeutic outcome in patients, a greater understanding of the exact mechanisms of drug resistance in T. gondii should be developed. Thus, monitoring the presence of resistant parasites, in food products, would seem a prudent public health program.
Toxoplasma gondii (T. gondii) is a remarkably successful protozoan parasite that belongs to the phylum Apicomplexa (Tenter et al., 2000). It is estimated that up to one-third of the world's human population is infected with T. gondii (Montoya and Liesenfeld, 2004). In addition, toxoplasmosis is considered as the third most common food-borne parasitic infection requiring hospitalization (Vaillant et al., 2005).
Toxoplasma infection in humans mainly occurs through two ways: (1) ingestion of tissue cysts with raw or undercooked meat. (2) Consumption of oocysts with contaminated food, water, vegetables, fruits, etc. Congenital transmission from mother to fetus is also possible when a woman gets an infection during pregnancy (Moncada and Montoya, 2012; Sepúlveda-Arias et al., 2014).
In most immunocompetent people, infection with T. gondii is usually asymptomatic. But in immunocompromised or congenitally infected patients without proper treatment, severe diseases may occur (Moncada and Montoya, 2012; Wang et al., 2017). In a recent global meta-analysis study, high odds ratios (ORs) was reported for Toxoplasma infection in HIV/AIDS patients especially in Asia and Africa and in cancer patients in Asia (Wang et al., 2017). However, in South America, severe ocular toxoplasmosis is higher than in many other parts of the world (Glasner et al., 1992).
The population structure of T. gondii consists of three main clonal lineages; Type I (including a highly virulent RH strain), Type II (including ME49 and PRU, avirulent strains), and Type III (including avirulent strains like NED) (Howe and Sibley, 1995).
Type II is the predominant type of clonal lineage that infects humans and animals in Europe and in North America. However, more recent studies in South America have documented the discovery of genetically atypical (non-clonal) strains of T. gondii isolated from human patients, which caused much more dramatic clinical symptoms compared with their European counterparts. Thus, the number of strains, or isolates, not to be categorized as type I, II, and III clonal lineages has increased strongly, and has meanwhile outnumbered those who are conventionally categorized (Shwab et al., 2014). These atypical strains also influenced immunocompetent individuals, and there are reports of abortions in Toxoplasma-positive pregnant women due to atypical T. gondii strains. Phenotypically, atypical T. gondii field strains are completely different from their European counterparts and from laboratory-adapted strains used as models for studies on T. gondii biology and the efficacy of novel compounds in drug development programs (Shwab et al., 2014).
Recommended drugs for treatment or prophylaxis of toxoplasmosis are limited to combinations of pyrimethamine (PYR) and sulfadiazine (SDZ). Unfortunately, these drugs have severe side effects such as neutropenia, leucopenia, severe platelet count decrease, thrombocytopenia, and hypersensitivity reactions (Porter and Sande, 1992; Rajapakse et al., 2013; Montazeri et al., 2015). Additionally, these drugs are related to some uncommon reactions as well, including agranulocytosis, Stevens–Johnson syndrome, toxic epidermal necrolysis, and hepatic necrosis, which may be fatal in patients with toxoplasmosis (McLeod et al., 2006).
In a retrospective review, 62% of patients treated with PYR, SDZ, and leucovorin showed a high rate of toxication and a number of side effects which required a change in the therapeutic regimen in 44% of patients (Porter and Sande, 1992).
Drugs, such as azithromycin, clarithromycin, spiramycin, atovaquone, dapsone, and cotrimoxazole (trimethoprim-sulfamethoxazole), have also been used to treat clinical toxoplasmosis; however, they are poorly tolerated and have no effect on the bradyzoite form of the parasite (Montazeri et al., 2017b, 2018).
Also, there have been several reports on failures of the long-term treatment of toxoplasmic encephalitis, chorioretinitis, and congenital toxoplasmosis with antifolate, particularly among AIDS patients (Jacobson et al., 1996; Bossi et al., 1998; Villena et al., 1998. Hence, there is controversy whether these failures are related to pharmacological parameters (drug intolerance, poor compliance, and malabsorption) and/or to the development of drug-resistant parasites or a lower susceptibility of the T. gondii strain (Meneceur et al., 2008). T. gondii parasite has an exceptional adaptive potential which renders it “resistant,” but the mechanism of resistance, or adaption, has not been completely elucidated (Kropf et al., 2012).
In a previous study by Ouellette, the basic mechanisms of parasite drug resistance in malaria, leishmaniasis, sleeping sickness, and common helminthiases were evaluated (Ouellette, 2001). In another study by McFadden et al. resistance was investigated as a tool to investigate old and new drug action sites in Toxoplasma parasite (McFadden et al., 2001). However, previous studies have demonstrated that drug resistance in T. gondii is not yet a major problem in human population (McFadden et al., 2001; Sims, 2009); recently, studies have focused on finding safe drugs with novel mechanisms of action for toxoplasmosis that are both efficacious and nontoxic for patients (Alday and Doggett, 2017; Montazeri et al., 2017a; Daryani et al., 2018).
It should be noted that various studies have been published reporting drug resistance in T. gondii (Table 1). The fact that drug resistant forms of T. gondii strains can contribute to human disease could raise a concern for treatment failure in the future (Silva et al., 2017). This review is focused on the available knowledge, encompassing information on anti-Toxoplasma drug resistance including mechanisms of resistance and drug target in parasite.
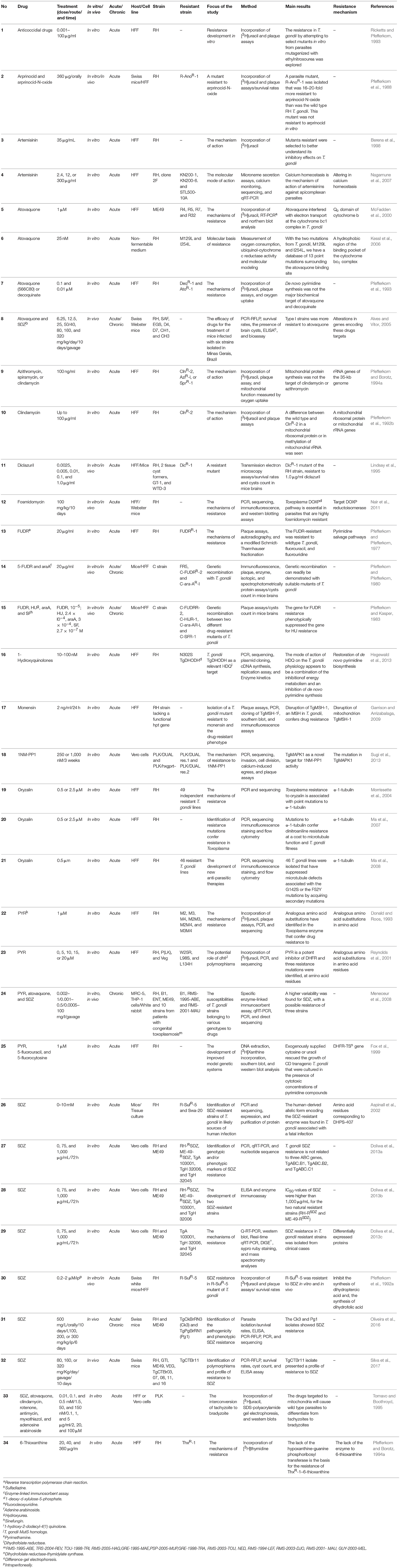
Table 1. List of the studies that evaluated drug resistance in T. gondii.
Clinically, acute toxoplasmosis is usually treated with a combination of PYR and SDZ. These drugs inhibit important enzymes for pyrimidine biosynthesis in the parasite [dihydrofolate reductase (DHFR) and dihydropteroate synthase (DHPS)] and have a remarkable synergistic activity against parasite survival and replication. DHFR is also present in humans so that the treatment with DHFR inhibitors may induce a folate deficiency state, which is probably responsible for hematological side effects and embryopathies (Rajapakse et al., 2013). Therefore, in order to limit adverse hematological events, these treatments are administered with folinic acid (Alday and Doggett, 2017).
However, these pathways are not essential for T. gondii viability, as studies showed that in resistant strains of T. gondii to 5-fluorodeoxyuridine, adenosine arabinoside, and 6-thioxanthine, the purine and pyrimidine analogs, isolated in vitro were viable despite defects in the crucial nucleotide synthesis enzymes (Pfefferkorn and Pfefferkorn, 1977, 1978; Pfefferkorn and Borotz, 1994b).
Reynolds et al. (2001) reported that using in vitro mutagenesis, single-point mutations in T. gondii DHFR-TS (dihydrofolate reductase-thymidylate synthase) (e.g., W25R, L98S, and L134H) can produce drug resistance in RH strain parasites compared with type II and type III strains. In addition, using site directed mutagenesis and transgenic experiments several mutations were induced in the DHFR-TS gene related to resistance to PYR. The T83N mutation was found to probably confer resistance to PYR (Donald and Roos, 1993). Resistance is even increased when T83N mutation is associated with mutation of S36R and F245S (Reynolds et al., 2001). Meneceur et al. reported variability in the susceptibilities of T. gondii strains to PYR, with no clear evidence of drug resistance and no relationship with strain genotype or defined mutations in drug target genes (Meneceur et al., 2008).
Given that PYR resistance is differently marked among various strains of T. gondii; it can provide new insights into potential sources of treatment failures and possible drug resistance mechanisms.
Sulfonamides, in conjunction with PYR, are a mainstay of toxoplasmosis treatment, although AIDS patients are unable to tolerate this treatment. The first experimentally induced drug-resistance was resistance to sulfamethoxazole, when the parasite was exposed to sub-lethal doses of the drug for long periods (Sander and Midtvedt, 1971; Luft and Remington, 1992; Reynolds and Roos, 1998). In a study by Pfefferkorn et al. (1992a), researchers induced resistance in RH strain using chemical mutagenesis and growing parasites in environments with gradually increased SDZ concentrations. Sulfamethoxazole-resistant strain (R-SulR-5) appeared to be more resistant than the parental RH strain. Further study on R-SulR-5 confirmed previous findings that this strain is sulfonamide resistant with an IC50 value near 5 mM like another SDZ resistant strain Swa-20 which was isolated from patients with clinical toxoplasmosis (Aspinall et al., 2002. In Aspinall et al. (Aspinall et al., 2002) study, the presence of one mutation at positions 407 of DHPS was associated with sulfonamides resistance by direct sequencing of PCR products (Aspinall et al., 2002). This mutation was also retrieved in laboratory induced R-SulR-5 (Pfefferkorn et al., 1992a). As, mutation 407 was not identified in five T. gondii Brazilian isolates obtained from newborns with congenital toxoplasmosis (Silva et al., 2017), a larger number of atypical isolates of T. gondii must be evaluated to confirm these results.
Meneceur et al. (Meneceur et al., 2008) isolated three strains from clinical cases containing: TgA 103001, previously described as B1 (Type I strain), TgH 32006, previously described as RMS-1995-ABE (Type II strain), and TgH 32045, previously described as RMS-2001-MAU (Type II variant strain) which were detected as resistant to SDZ. Doliwa et al. (2013c) found 44% over-expressed proteins in resistant T. gondii strains. The virulence-associated rhoptry protein, ROP2A, was found in greater abundance in both naturally resistant Type II strains TgH 32006 and TgH 32045. Totally, 31 proteins were identified which are differentially modulated between SDZ resistant and sensitive strains of T. gondii according to their genotype using proteomics approach. Recently, two SDZ resistant strains were developed called RH-RSDZ and ME-49-RSDZ in vitro (Doliwa et al., 2013b). Also, other studies analyzed genotypic and/or phenotypic markers of resistance in T. gondii (Doliwa et al., 2013a).
In Oliveira et al. (Oliveira et al., 2016) study, Ck3 and Pg1 T. gondii isolates showed SDZ resistance in samples collected from livestock intended for human consumption. Monitoring the presence of resistant parasites, particularly in food products, would seem a prudent public health measure (Sims, 2009).
Silva et al. (2017) have confirmed the existence of a Brazilian T. gondii isolate, TgCTBr11, isolated from newborns infected with congenital toxoplasmosis, which is resistant to SDZ. Despite the large number of polymorphisms identified in the DHPS gene, no association was found between the profile of susceptibility to SDZ and the virulence-phenotype and genotype of the parasite. However, the mutation in the DHPS gene is known to confer resistance in T. gondii and has demonstrated cross-resistance to several sulfonamides including SDZ and sulfamethoxazole. Until now, T. gondii SDZ resistance has not been related to genetic mutations in DHPS in all clinical isolates. Based on these findings, the range of resistance to sulfonamide is greater than PYR or atovaquone.
Atovaquone is a substituted hydroxynaphthoquinone compound that is being used clinically for the treatment of T. gondii infections against chronic bradyzoite stage via mitochondrial electron transport chain inhibition (Kovacs, 1992; Tomavo and Boothroyd, 1995); however, atovaquone prophylaxis and treatment failure was reported in hematopoietic cell transplant recipients and AIDS patients (Chirgwin et al., 2002; Gajurel et al., 2016).
Cytochrome bc1 complex (CYT bc1) is a membrane-bound enzyme of the respiratory electron transfer chain located in the inner mitochondrial membrane. It is a successful drug target for combatting diseases, including T. gondii, Plasmodium falciparum, and Babesia microti (Winter et al., 2008; Doggett et al., 2012; Lawres et al., 2016). CYT bc1 reduces cytochrome c and generates an electrochemical gradient by transferring protons to the intermembrane space. It also creates ubiquinone for pyrimidine biosynthesis. CYT bc1 has two active sites, the bc1 Qo site (oxidizes ubiquinol) and the bc1 Qi site (reduces ubiquinone) (Crofts, 2004).
The genetic evidence revealed that atovaquone, targets T. gondii CYT bc1 by binding to Qo domain of cytochrome b confer resistance to atovaquone (McFadden et al., 1997, 2001). M129L and I254L mutations have been identified to be related to atovaquone resistance in T. gondii (31, 32).
Of course, the investigation by Meneceur et al. (Meneceur et al., 2008) did not show any of these mutations, thus further studies will help a better understanding of resistance mechanisms.
Interestingly, Endochin-like quinolones (ELQs) have been shown to be active against atovaquone-resistant Plasmodium and Babesia (Winter et al., 2008; Lawres et al., 2016). Also, treatment with 4(1H)-pyridone compounds, GW844520, and GSK932121, showed anti-malarial activity in vivo (Capper et al., 2015). These anti-parasitic agents have a similar mechanism of action with atovaquone but by inhibiting the Qi site of CYT bc1. Furthermore, ELQ-271 and ELQ-316 showed remarkable effects against acute and latent toxoplasmosis at low doses (Doggett et al., 2012). It is likely that ELQs act at the T. gondii cytochrome b Qi site. Therefore, ELQs and 4(1H)-pyridone compounds are promising candidates for the treatment against atovaquone-resistant Toxoplasma.
1-Hydroxyquinolones are effective inhibitors of T. gondii replication. Using a drug resistant strain, Hegewald et al. (2013) described that the enzyme dihydroorotate dehydrogenase (TgDHODH) of T. gondii is a relevant target for 1-Hydroxy-2-dodecyl-4(1H) quinolone (HDQ) and compound B (1-Hydroxyquinolones derivatives). Thus, drug resistant mutants are approved tools for the identification of drug targets for future to select new anti-Toxoplasma drugs.
Antibiotics such as clindamycin, spiramycin, and azithromycin are known to be active against T. gondii. However, mutant ClnR-2 (RH) was cross-resistant to clindamycin, azithromycin, and spiramycin antibiotics (Pfefferkorn et al., 1992b; Pfefferkorn and Borotz, 1994a). Interestingly, resistance to these drugs is encoded in the rRNA genes of the 35-kb genome in T. gondii and the apicoplast protein synthesis is known as target of these antibiotics action against T. gondii (Pfefferkorn and Borotz, 1994a; McFadden et al., 2001).
ART is a natural product that is produced by Artemisia annua plant. This important compound plays an indispensable role for combating malaria (Cui et al., 2015). ART is also effective against Toxoplasma in vitro and in vivo (Schultz et al., 2014), although it is not generally used in the treatment of toxoplasmosis. Recent concerns about the development of ART resistance have led to the exploration of its mechanisms of action. Berens et al. (1998) characterized five clonal isolates that showed cross-resistance to the ART derivatives, dihydroartemisinin and artemether in laboratory studies. In a subsequent study, Nagamune et al. (2007) generated chemically derived T. gondii mutants that were resistant to growth inhibition by ART in vitro. Three ART-resistant mutants were resistant to the induction of protein secretion from micronemes, a calcium-dependent process that is triggered by artemisinin. Based on these findings, calcium homeostasis is involved in the mechanism of ART action against T. gondii and other apicomplexan parasites.
T. gondii CDPK1 (TgCDPK1) was found to be the target of 1NM-PP1, which is a bumped kinase inhibitor (BKIs). CDPK1 contains an atypically small glycine gatekeeper residue, which allows entry of BKIs into the ATP binding domain. Most mammalian kinases have larger gatekeeper residues, e.g., methionine. CDPK1 is involved in microneme secretion and host cell invasion and egress. When TgCDPK1 was mutated at position 128 from glycine to methionine, parasites became BKI resistant (Sugi et al., 2010). Resistance to 1NM-PP1 can also be acquired via a mutation in T. gondii mitogen-activated protein kinase like 1, which indicates that this kinase could also be a target (Sugi et al., 2013, 2015). However, CDPK1 has become an important drug target for more recently developed and largely improved BKIs in a variety of apicomplexans beside T. gondii (Van Voorhis et al., 2017).
T. gondii is sensitive to dinitroaniline compounds, which disrupt microtubules without affecting host cells. T. gondii containing alpha-tubulin point mutations are dinitroaniline resistant. Ma et al. (2008) identified T. gondii lines that have suppressed microtubule defects in G142S or F52Y mutations. In addition, secondary resistant mutations were isolated that corrects fitness defects in the T. gondii parasite. Based on the current findings, targeting parasite microtubules can be a viable strategy for developing new anti-parasitic therapies.
Anti-coccidial agents were assessed in T. gondii mutants for development of resistance in vitro. Mutants had 20- to 50-fold-reduced susceptibility to decoquinate, arprinocid-N-oxide, and CP-25,415. In addition, ionophore-resistant T. gondii mutants were explored in vitro; however, resistance to all of the mutants except ionophores occurs in coccidia in vivo. The availability of a T. gondii mutant resistant to a different drug could aid for assessing the risk of developing resistance in Eimeria species (Ricketts and Pfefferkorn, 1993).
Diclazuril, an anticoccidial compound, is a safe and effective drug that inhibits tachyzoite production of RH strain in T. gondii by >97% at therapeutic dose levels (Oz, 2014). Lindsay et al. (1995) have shown that formation of T. gondii tissue cysts was not prevented by treatment with diclazuril, in vitro. They also showed that GT-1, WTD-3 strains, and a mutant RH strain of T. gondii were resistant to 1.0 μg/ml of diclazuril.
Monensin is a polyether anti-coccidial antibiotic that has been effective against T. gondii. However, within 3 years of the drug introduction, monensin-resistant Eimeria maxima were noted. Thus, T. gondii was used for studying the monensin's mechanisms of resistance. The investigators have shown that resistance phenotype is caused by the disruption of T. gondii homologs MSH-1 (a homolog of the DNA repair enzyme, MutS). Interestingly, this enzyme localizes to the T. gondii parasite mitochondrion (Garrison and Arrizabalaga, 2009). Subsequent studies showed that the disruption of the autophagy pathway could result in drug resistance. Autophagy pathway is a potentially important model of cell death of T. gondii in response to monensin (Lavine and Arrizabalaga, 2012).
Several targets were identified against T. gondii including folate synthesis pathway, mitochondrial electron transport chain, calcium dependent ATPases, protein synthesis, mitogen-activated protein kinase 1, enzyme TgDHODH, and microtubules for PYR and SDZ, atovaquone, ART, clindamycin, spiramycin and azithromycin, 1NM-PP1 and 1-hydroxyquinolones HDQ, and compound B, respectively. Thus, drug resistant mutants are approved tools for the characterization of drug targets for future to select new anti-Toxoplasma drugs with specific activity against the parasite.
Also, mechanisms of drug resistance in T. gondii have been described. Interestingly, analogous amino acid substitutions in the Toxoplasma enzyme have been identified to confer PYR resistance in transfected parasites (Donald and Roos, 1993). Moreover, resistance to clindamycin, spiramycin and azithromycin is encoded in the rRNA genes of the 35-kb genome in T. gondii.
There are numerous reports with a focus on identifying SDZ resistance mechanisms. However, T. gondii SDZ resistance mechanism has not been proved so far. As a consequence, understanding mechanisms of drug resistance in T. gondii is essential for controlling the disease particularly among immunocompromised patients. Also, it helps identify targets that are crucial to the parasite and predicts which combinations of drugs should act synergistically (McFadden et al., 2001).
Studies in the past 10 years indicated that drug resistance to SDZ is actually increased. Most resistant strains were found in clinical cases between 2013 and 2017. However, a possible resistance was reported in three strains of T. gondii in 2008. Also, six strains resistant to SDZ were found in clinical cases between 2013 and 2017.
The only worrying trend was a very slight recent increase in SDZ resistance to Brazilian T. gondii strains obtained from livestock and humans newborns with congenital toxoplasmosis between 2016 and 2017 where T. gondii prevalence in Brazil is high (77.5%) (Pappas et al., 2009). Thus, establishing a more effective therapeutic scheme in the treatment of toxoplasmosis is critically needed.
Recent experimental studies in clinical cases have clearly shown that drug resistance in Toxoplasma is ongoing. The emergence of T. gondii strains resistant to current drugs reviewed here represents a concern not only for treatment failure but also for increased clinical severity in immunocompromised patients. Thus, understanding mechanisms of drug resistance is essential for controlling the disease and it helps identify targets that are crucial to the parasite and predicts which combinations of drugs should act synergistically. Also, establishing a more effective therapeutic scheme in the treatment of toxoplasmosis, particularly among high-risk individuals is critically needed. Additionally, monitoring the presence of resistant parasites, particularly in food products, would thus seem a prudent public health measure. Further development of a greater understanding of exact mechanisms of drug resistance in T. gondii is needed to improve the therapeutic outcomes in patients.
AD conceived the study. AD and MS designed the study protocol. MM, SS, and AT searched the databases. MM wrote the manuscript. SM and SA critically revised the manuscript. All authors read and approved the final manuscript for publication.
This work was supported by the Deputy of Research, Mazandaran University of Medical Sciences, Sari, Iran (Grant number: 9180).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank Student Research Committee, Mazandaran University of Medical Sciences, Sari, Iran.
Alday, P. H., and Doggett, J. S. (2017). Drugs in development for toxoplasmosis: advances, challenges, and current status. Drug Des. Dev. Ther. 11, 273–293. doi: 10.2147/DDDT.S60973
Alves, C., and Vitor, R. (2005). Efficacy of atovaquone and sulfadiazine in the treatment of mice infected with Toxoplasma gondii strains isolated in Brazil. Parasite 12, 171–177. doi: 10.1051/parasite/2005122171
Aspinall, T. V., Joynson, D. H., Guy, E., Hyde, J. E., and Sims, P. F. (2002). The molecular basis of sulfonamide resistance in Toxoplasma gondii and implications for the clinical management of toxoplasmosis. J. Infect. Dis. 185, 1637–1643. doi: 10.1086/340577
Berens, R. L., Krug, E. C., Nash, P. B., and Curiel, T. J. (1998). Selection and characterization of Toxoplasma gondii mutants resistant to artemisinin. J. Infect. Dis. 177, 1128–1131. doi: 10.1086/517411
Bossi, P., Caumes, E., Astagneau, P., Li, T., Paris, L., Mengual, X., et al. (1998). Epidemiologic characteristics of cerebral toxoplasmosis in 399 HIV-infected patients followed between 1983 and 1994. Rev. Med. Interne 19, 313–317.
Capper, M. J., O'Neill, P. M., Fisher, N., Strange, R. W., Moss, D., Ward, S. A., et al. (2015). Antimalarial 4 (1H)-pyridones bind to the Qi site of cytochrome bc1. Proc. Natl. Acad. Sci. U. S. A. 112, 755–760. doi: 10.1073/pnas.1416611112
Chirgwin, K., Hafner, R., Leport, C., Remington, J., Andersen, J., Bosler, E. M., et al. (2002). Randomized phase II trial of atovaquone with pyrimethamine or sulfadiazine for treatment of toxoplasmic encephalitis in patients with acquired immunodeficiency syndrome: ACTG 237/ANRS 039 study. Clin. Infect. Dis. 34, 1243–1250. doi: 10.1086/339551
Crofts, A. R. (2004). The cytochrome bc1 complex: function in the context of structure. Annu. Rev. Physiol. 66, 689–733. doi: 10.1146/annurev.physiol.66.032102.150251
Cui, L., Mharakurwa, S., Ndiaye, D., Rathod, P. K., and Rosenthal, P. J. (2015). Antimalarial drug resistance: literature review and activities and findings of the ICEMR network. Am. J. Trop. Med. Hyg. 93(Suppl. 3), 57–68. doi: 10.4269/ajtmh.15-0007
Daryani, A., Montazeri, M., Pagheh, A. S., Sharif, M., Sarvi, S., Hosseinzadeh, A., et al. (2018). The potential use of melatonin to treat protozoan parasitic infections: a review. Biomed. Pharmacother. 97, 948–957. doi: 10.1016/j.biopha.2017.11.007
Doggett, J. S., Nilsen, A., Forquer, I., Wegmann, K. W., Jones-Brando, L., Yolken, R. H., et al. (2012). Endochin-like quinolones are highly efficacious against acute and latent experimental toxoplasmosis. Proc. Natl. Acad. Sci. U. S. A. 109, 15936–15941. doi: 10.1073/pnas.1208069109
Doliwa, C., Escotte-Binet, S., Aubert, D., Sauvage, V., Velard, F., Schmid, A., et al. (2013a). Sulfadiazine resistance in Toxoplasma gondii: no involvement of overexpression or polymorphisms in genes of therapeutic targets and ABC transporters. Parasite 20:19. doi: 10.1051/parasite/2013020
Doliwa, C., Escotte-Binet, S., Aubert, D., Velard, F., Schmid, A., Geers, R., et al. (2013b). Induction of sulfadiazine resistance in vitro in Toxoplasma gondii. Exp. Parasitol. 133, 131–136. doi: 10.1016/j.exppara.2012.11.019
Doliwa, C., Xia, D., Escotte-Binet, S., Newsham, E. L., Aubert, D., Randle, N., et al. (2013c). Identification of differentially expressed proteins in sulfadiazine resistant and sensitive strains of Toxoplasma gondii using difference-gel electrophoresis (DIGE). Int. J. Parasitol. Drugs Drug Resist. 3, 35–44. doi: 10.1016/j.ijpddr.2012.12.002
Donald, R., and Roos, D. S. (1993). Stable molecular transformation of Toxoplasma gondii: a selectable dihydrofolate reductase-thymidylate synthase marker based on drug-resistance mutations in malaria. Proc. Natl. Acad. Sci. U. S. A. 90, 11703–11707. doi: 10.1073/pnas.90.24.11703
Fox, B., Belperron, A., and Bzik, D. (1999). Stable transformation of Toxoplasma gondii based on a pyrimethamine resistant trifunctional dihydrofolate reductase-cytosine deaminase-thymidylate synthase gene that confers sensitivity to 5-fluorocytosine. Mol. Biochem. Parasitol. 98, 93–103. doi: 10.1016/S0166-6851(98)00154-6
Gajurel, K., Gomez, C., Dhakal, R., Vogel, H., and Montoya, J. (2016). Failure of primary atovaquone prophylaxis for prevention of toxoplasmosis in hematopoietic cell transplant recipients. Transpl. Infect. Dis.18, 446–452. doi: 10.1111/tid.12532
Garrison, E. M., and Arrizabalaga, G. (2009). Disruption of a mitochondrial MutS DNA repair enzyme homologue confers drug resistance in the parasite Toxoplasma gondii. Mol. Microbiol. 72, 425–441. doi: 10.1111/j.1365-2958.2009.06655.x
Glasner, P. D., Silveira, C., Kruszon-Moran, D., Martins, M. C., Burnier, M., Silveira, S., et al. (1992). An unusually high prevalence of ocular toxoplasmosis in southern Brazil. Am. J. Ophthalmol. 114, 136–144. doi: 10.1016/S0002-9394(14)73976-5
Hegewald, J., Gross, U., and Bohne, W. (2013). Identification of dihydroorotate dehydrogenase as a relevant drug target for 1-hydroxyquinolones in Toxoplasma gondii. Mol. Biochem. Parasitol. 190, 6–15. doi: 10.1016/j.molbiopara.2013.05.008
Howe, D. K., and Sibley, L. D. (1995). Toxoplasma gondii comprises three clonal lineages: correlation of parasite genotype with human disease. J. Infect. Dis. 172, 1561–1566. doi: 10.1093/infdis/172.6.1561
Jacobson, J. M., Davidian, M., Rainey, P. M., Hafner, R., Raasch, R. H., and Luft, B. J. (1996). Pyrimethamine pharmacokinetics in human immunodeficiency virus-positive patients seropositive for Toxoplasma gondii. Antimicrob. Agents Chemother. 40, 1360–1365. doi: 10.1128/AAC.40.6.1360
Kessl, J. J., Ha, K. H., Merritt, A. K., Meshnick, S. R., and Trumpower, B. L. (2006). Molecular basis of Toxoplasma gondii atovaquone resistance modeled in Saccharomyces cerevisiae. Mol. Biochem. Parasitol. 146, 255–258. doi: 10.1016/j.molbiopara.2005.12.002
Kovacs, J. A. (1992). Efficacy of atovaquone in treatment of toxoplasmosis in patients with AIDS. Lancet 340, 637–638. doi: 10.1016/0140-6736(92)92172-C
Kropf, C., Debache, K., Rampa, C., Barna, F., Schorer, M., Stephens, C. E., et al. (2012). The adaptive potential of a survival artist: characterization of the in vitro interactions of Toxoplasma gondii tachyzoites with di-cationic compounds in human fibroblast cell cultures. Parasitology 139, 208–220. doi: 10.1017/S0031182011001776
Lavine, M. D., and Arrizabalaga, G. (2012). Analysis of monensin sensitivity in Toxoplasma gondii reveals autophagy as a mechanism for drug induced death. PLoS ONE 7:e42107. doi: 10.1371/journal.pone.0042107
Lawres, L. A., Garg, A., Kumar, V., Bruzual, I., Forquer, I. P., Renard, I., et al. (2016). Radical cure of experimental babesiosis in immunodeficient mice using a combination of an endochin-like quinolone and atovaquone. J. Exp. Med. 27,1307–1318. doi: 10.1084/jem.20151519
Lindsay, D. S., Rippey, N. S., Toivio-Kinnucan, M. A., and Blagburn, B. L. (1995). Ultrastructural effects of diclazuril against Toxoplasma gondii and investigation of a diclazuril-resistant mutant. J. Parasitol. 81:459–466.
Luft, B. J., and Remington, J. S. (1992). Toxoplasmic encephalitis in AIDS. Clin. Infect. Dis. 15, 211–222. doi: 10.1093/clinids/15.2.211
Ma, C., Li, C., Ganesan, L., Oak, J., Tsai, S., Sept, D., et al. (2007). Mutations in α-tubulin confer dinitroaniline resistance at a cost to microtubule function. Mol. Biol. Cell 18, 4711–4720. doi: 10.1091/mbc.e07-04-0379
Ma, C., Tran, J., Li, C., Ganesan, L., Wood, D., and Morrissette, N. (2008). Secondary mutations correct fitness defects in Toxoplasma gondii with dinitroaniline resistance mutations. Genetics 180, 845–856. doi: 10.1534/genetics.108.092494
McFadden, D. C., Camps, M., and Boothroyd, J. C. (2001). Resistance as a tool in the study of old and new drug targets in Toxoplasma. Drug Resist. Updates 4, 79–84. doi: 10.1054/drup.2001.0184
McFadden, D. C., Seeber, F., and Boothroyd, J. C. (1997). Use of Toxoplasma gondii expressing -galactosidase for colorimetric assessment of drug activity in vitro. Antimicrob. Agents Chemother. 41,1849–1853. doi: 10.1128/AAC.41.9.1849
McFadden, D. C., Tomavo, S., Berry, E. A., and Boothroyd, J. C. (2000). Characterization of cytochrome b from Toxoplasma gondii and Q o domain mutations as a mechanism of atovaquone-resistance. Mol. Biochem. Parasitol. 108, 1–12. doi: 10.1016/S0166-6851(00)00184-5
McLeod, R., Boyer, K., Karrison, T., Kasza, K., Swisher, C., Roizen, N., et al. (2006). Outcome of treatment for congenital toxoplasmosis, 1981–2004: the national collaborative Chicago-based, congenital toxoplasmosis study. Clin. Infect. Dis. 42, 1383–1394. doi: 10.1086/501360
Meneceur, P., Bouldouyre, M.-A., Aubert, D., Villena, I., Menotti, J., Sauvage, V., et al. (2008). In vitro susceptibility of various genotypic strains of Toxoplasma gondii to pyrimethamine, sulfadiazine, and atovaquone. Antimicrob. Agents.Chemother. 52, 1269–1277. doi: 10.1128/AAC.01203-07
Moncada, P. A., and Montoya, J. G. (2012). Toxoplasmosis in the fetus and newborn: an update on prevalence, diagnosis and treatment. Expert Rev. Anti Infect. Ther. 10, 815–828. doi: 10.1586/eri.12.58
Montazeri, M., Daryani, A., Ebrahimzadeh, M., Ahmadpour, E., Sharif, M., and Sarvi, S. (2015). Effect of propranolol alone and in combination with pyrimethamine on acute murine toxoplasmosis. Jundishapur J.Microbiol. 8:e22572. doi: 10.5812/jjm.22572
Montazeri, M., Mehrzadi, S., Sharif, M., Sarvi, S., Shahdin, S., and Daryani, A. (2018). Activities of anti-Toxoplasma drugs and compounds against tissue cysts in the last three decades (1987 to 2017), a systematic review. Parasitol. Res. 117, 3045–3057. doi: 10.1007/s00436-018-6027-z
Montazeri, M., Rezaei, K., Ebrahimzadeh, M. A., Sharif, M., Sarvi, S., Ahmadpour, E., et al. (2017a). Survey on synergism effect of ketotifen in combination with pyrimethamine in treatment of acute murine toxoplasmosis. Trop. Med. Int. 45:39. doi: 10.1186/s41182-017-0079-0
Montazeri, M., Sharif, M., Sarvi, S., Mehrzadi, S., Ahmadpour, E., and Daryani, A. (2017b). A systematic review of in vitro and in vivo activities of anti-toxoplasma drugs and compounds (2006–2016). Front Microbiol. 8:25. doi: 10.3389/fmicb.2017.00025
Montoya, J. G., and Liesenfeld, O. (2004). Toxoplasmosis. Lancet 363, 1965–1976. doi: 10.1016/S0140-6736(04)16412-X
Morrissette, N. S., Mitra, A., Sept, D., and Sibley, L. D. (2004). Dinitroanilines bind α-tubulin to disrupt microtubules. Mol. Biol. Cell 15, 1960–1968. doi: 10.1091/mbc.e03-07-0530
Nagamune, K., Moreno, S. N., and Sibley, L. D. (2007). Artemisinin-resistant mutants of Toxoplasma gondii have altered calcium homeostasis. Antimicrob. Agents Chemother. 51, 3816–3823. doi: 10.1128/AAC.00582-07
Nair, S. C., Brooks, C. F., Goodman, C. D., Strurm, A., McFadden, G. I., Sundriyal, S., et al. (2011). Apicoplast isoprenoid precursor synthesis and the molecular basis of fosmidomycin resistance in Toxoplasma gondii. J. Exp. Med. 208, 1547–1559. doi: 10.1084/jem.20110039
Oliveira, C., Meurer, Y. S., Andrade, J., Costa, M. E., Andrade, M., Silva, L. A., et al. (2016). Pathogenicity and phenotypic sulfadiazine resistance of Toxoplasma gondii isolates obtained from livestock in northeastern Brazil. Mem Inst Oswaldo Cruz. 111, 391–398. doi: 10.1590/0074-02760150459
Ouellette, M. (2001). Biochemical and molecular mechanisms of drug resistance in parasites. Trop. Med. Int. Health 6, 874–882. doi: 10.1046/j.1365-3156.2001.00777.x
Oz, H. S. (2014). Novel synergistic protective efficacy of atovaquone and diclazuril on fetal-maternal toxoplasmosis. Int. J. Clin. Med. 5:921. doi: 10.4236/ijcm.2014.515124
Pappas, G., Roussos, N., and Falagas, M. E. (2009). Toxoplasmosis snapshots: global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int. J. Parasitol. 39, 1385–1394. doi: 10.1016/j.ijpara.2009.04.003
Pfefferkorn, E., and Borotz, S. E. (1994a). Comparison of mutants of Toxoplasma gondii selected for resistance to azithromycin, spiramycin, or clindamycin. Antimicrob. Agents Chemother. 38, 31–37.
Pfefferkorn, E., and Borotz, S. E. (1994b). Toxoplasma gondii: characterization of a mutant resistant to 6-thioxanthine. Exp Parasitol. 79, 374–382.
Pfefferkorn, E., Borotz, S. E., and Nothnagel, R. F. (1992a). Toxoplasma gondii: characterization of a mutant resistant to sulfonamides. Exp Parasitol. 74, 261–270.
Pfefferkorn, E., Borotz, S. E., and Nothnagel, R. F. (1993). Mutants of Toxoplasma gondii resistant to atovaquone (566C80) or decoquinate. J. Parasitol. 79, 559–564.
Pfefferkorn, E., Eckel, M. E., and McAdams, E. (1988). Toxoplasma gondii: in vivo and in vitro studies of a mutant resistant to arprinocid-N-oxide. Exp. Parasitol. 65, 282–289. doi: 10.1016/0014-4894(88)90133-6
Pfefferkorn, E., and Kasper, L. H. (1983). Toxoplasma gondii: genetic crosses reveal phenotypic suppression of hydroxyurea resistance by fluorodeoxyuridine resistance. Exp. Parasitol. 55, 207–218. doi: 10.1016/0014-4894(83)90015-2
Pfefferkorn, E., Nothnagel, R. F., and Borotz, S. E. (1992b). Parasiticidal effect of clindamycin on Toxoplasma gondii grown in cultured cells and selection of a drug-resistant mutant. Antimicrob. Agents Chemother. 36, 1091–1096.
Pfefferkorn, E., and Pfefferkorn, L. C. (1977). Toxoplasma gondii: characterization of a mutant resistant to 5-fluorodeoxyuridine. Exp. Parasitol. 42, 44–55. doi: 10.1016/0014-4894(77)90060-1
Pfefferkorn, E., and Pfefferkorn, L. C. (1978). The biochemical basis for resistance to adenine arabinoside in a mutant of Toxoplasma gondii. J. Parasitol. 64, 486–492. doi: 10.2307/3279789
Pfefferkorn, L. C., and Pfefferkorn, E. (1980). Toxoplasma gondii: genetic recombination between drug resistant mutants. Exp. Parasitol. 50, 305–316. doi: 10.1016/0014-4894(80)90034-X
Porter, S. B., and Sande, M. A. (1992). Toxoplasmosis of the central nervous system in the acquired immunodeficiency syndrome. N. Engl. J. Med. 327, 1643–1648. doi: 10.1056/NEJM199212033272306
Rajapakse, S., Chrishan Shivanthan, M., Samaranayake, N., Rodrigo, C., and Deepika Fernando, S. (2013). Antibiotics for human toxoplasmosis: a systematic review of randomized trials. Pathog. Glob. Health 107, 162–169. doi: 10.1179/2047773213Y.0000000094
Reynolds, M. G., Oh, J., and Roos, D. S. (2001). In Vitro generation of novel pyrimethamine resistance mutations in the Toxoplasma gondii dihydrofolate reductase. Antimicrob. Agents Chemother. 45, 1271–1277. doi: 10.1128/AAC.45.4.1271-1277.2001
Reynolds, M. G., and Roos, D. S. (1998). A biochemical and genetic model for parasite resistance to antifolates Toxoplasma gondii provides insights into pyrimethamine and cycloguanil resistance in Plasmodium falciparum. J. Biol. Chem. 273, 3461–3469. doi: 10.1074/jbc.273.6.3461
Ricketts, A. P., and Pfefferkorn, E. (1993). Toxoplasma gondii: susceptibility and development of resistance to anticoccidial drugs in vitro. Antimicrob. Agents Chemother. 37, 2358–2363. doi: 10.1128/AAC.37.11.2358
Sander, J., and Midtvedt, T. (1971). Development of sulphonamide resistance in Toxoplasma gondii. APMIS 79, 531–533.
Schultz, T. L., Hencken, C. P., Woodard, L. E., Posner, G. H., Yolken, R. H., Jones-Brando, L., et al. (2014). A thiazole derivative of artemisinin moderately reduces Toxoplasma gondii cyst burden in infected mice. J. Parasitol. 100, 516–521. doi: 10.1645/13-451.1
Sepúlveda-Arias, J. C., Gómez-Marin, J. E., Bobić, B., Naranjo-Galvis, C. A., and Djurković-Djaković, O. (2014). Toxoplasmosis as a travel risk. Travel Med. Infect. Dis. 12, 592–601. doi: 10.1016/j.tmaid.2014.05.007
Shwab, E. K., Zhu, X. Q., Majumdar, D., Pena, H. F., Gennari, S. M., Dubey, J. P., et al. (2014). Geographical patterns of Toxoplasma gondii genetic diversity revealed by multilocus PCR- RFLP genotyping. Parasitology 141, 453–461. doi: 10.1017/S0031182013001844
Silva, L. A., Reis-Cunha, J. L., Bartholomeu, D. C., and Vítor, R. W. A. (2017). Genetic polymorphisms and phenotypic profiles of sulfadiazine-resistant and sensitive Toxoplasma gondii isolates obtained from newborns with congenital toxoplasmosis in minas gerais, Brazil. PloS ONE 12:e0170689. doi: 10.1371/journal.pone.0170689
Sims, P. F. G. (2009). “Drug resistance in Toxoplasma gondii,” in Antimicrobial Drug Resistance. Infectious Disease, ed D. L. Mayers (New York, NY: Humana Press), 1121–1126. doi: 10.1007/978-1-60327-595-8_31
Sugi, T., Kato, K., Kobayashi, K., Watanabe, S., Kurokawa, H., Gong, H., et al. (2010). Use of the kinase inhibitor analog 1NM-PP1 reveals a role for Toxoplasma gondii CDPK1 in the invasion step. Eukaryotic Cell 9, 667–670. doi: 10.1128/EC.00351-09
Sugi, T., Kawazu, S.-,i., Horimoto, T., and Kato, K. (2015). A single mutation in the gatekeeper residue in TgMAPKL-1 restores the inhibitory effect of a bumped kinase inhibitor on the cell cycle. Int. J. Parasitol. Drugs Drug Resist. 5, 1–8. doi: 10.1016/j.ijpddr.2014.12.001
Sugi, T., Kobayashi, K., Takemae, H., Gong, H., Ishiwa, A., Murakoshi, F., et al. (2013). Identification of mutations in TgMAPK1 of Toxoplasma gondii conferring resistance to 1NM-PP1. Int. J. Parasitol. Drugs Drug Resist. 3, 93–101. doi: 10.1016/j.ijpddr.2013.04.001
Tenter, A. M., Heckeroth, A. R., and Weiss, L. M. (2000). Toxoplasma gondii: from animals to humans. Int. J. Parasitol. 30, 1217–1258. doi: 10.1016/S0020-7519(00)00124-7
Tomavo, S., and Boothroyd, J. C. (1995). Interconnection between organellar functions, development and drug resistance in the protozoan parasite, Toxoplasma gondii. Int. J. Parasitol. 25, 1293–1299. doi: 10.1016/0020-7519(95)00066-B
Vaillant, V., Valk, H. D., Baron, E., Ancelle, T., Colin, P., Delmas, M.-C., et al. (2005). Foodborne infections in France. Foodborne Pathog. Dis. 2, 221–232. doi: 10.1089/fpd.2005.2.221
Van Voorhis, W. C., Doggett, J. S., Parsons, M., Hulverson, M. A., Choi, R., Arnold, S. L., et al. (2017). Extended-spectrum antiprotozoal bumped kinase inhibitors: a review. Exp. Parasitol. 180, 71–83. doi: 10.1016/j.exppara.2017.01.001
Villena, I., Aubert, D., Leroux, B., Dupouy, D., Talmud, M., Chemla, C., et al. (1998). Pyrimethamine-sulfadoxine treatment of congenital toxoplasmosis: follow-up of 78 cases between 1980 and 1997. Scand. J. Infect. Dis. 30, 295–300. doi: 10.1080/00365549850160963
Wang, Z.-D., Liu, H.-H., Ma, Z.-X., Ma, H.-Y., Li, Z.-Y., Yang, Z.-B., et al. (2017). Toxoplasma gondii infection in immunocompromised patients: a systematic review and meta-analysis. Front. Microbiol. 8:389. doi: 10.3389/fmicb.2017.00389
Keywords: Toxoplasma gondii, toxoplasmosis, drugs, drug resistance, mechanisms of resistance
Citation: Montazeri M, Mehrzadi S, Sharif M, Sarvi S, Tanzifi A, Aghayan SA and Daryani A (2018) Drug Resistance in Toxoplasma gondii. Front. Microbiol. 9:2587. doi: 10.3389/fmicb.2018.02587
Received: 30 July 2017; Accepted: 10 October 2018;
Published: 29 October 2018.
Edited by:
Octavio Luiz Franco, Universidade Católica de Brasília, BrazilReviewed by:
Guan Zhu, Texas A&M University, United StatesCopyright © 2018 Montazeri, Mehrzadi, Sharif, Sarvi, Tanzifi, Aghayan and Daryani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ahmad Daryani, ZGFyeWFuaWlAeWFob28uY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.