 Jennifer R. Honda
Jennifer R. Honda Ravleen Virdi
Ravleen Virdi Edward D. Chan
Edward D. Chan
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol., 30 August 2018
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.02029
This article is part of the Research TopicNTM – The New Uber-BugsView all 25 articles
Seminal microbiological work of environmental nontuberculous mycobacteria (NTM) includes the discovery that NTM inhabit water distribution systems and soil, and that the species of NTM found are geographically diverse. It is likely that patients acquire their infections from repeated exposures to their environments, based on the well-accepted paradigm that water and soil bioaerosols – enriched for NTM – can be inhaled into the lungs. Support comes from reports demonstrating NTM isolated from the lungs of patients are genetically identical to NTM found in their environment. Well documented sources of NTM include peat-rich soils, natural waters, drinking water, hot water heaters, refrigerator taps, catheters, and environmental amoeba. However, NTM have also been recovered in biofilms from ice machines, heated nebulizers, and heater-cooler units, as well as seat dust from theaters, vacuum cleaners, and cobwebs. New studies on the horizon aim to significantly expand the current knowledge of environmental NTM niches in order to improve our current understanding of the specific ecological factors driving the emergence of NTM lung disease. Specifically, the Hawaiian Island environment is currently being studied as a model to identify other point sources of exposure as it is the U.S. state with the highest number of NTM lung disease cases. Because of its geographic isolation and unique ecosystem, the Hawaiian environment is being probed for correlative factors that may promote environmental NTM colonization.
Nontuberculous mycobacteria (NTM) and their environments are intricately bound. NTM share these environments with humans and domesticated animals and repeated exposure is a well-accepted mode of acquiring these infections. While most casual NTM exposures do not results in disease, those with anatomic lung abnormalities of bronchiectasis and emphysema are particularly predisposed to develop NTM lung disease (NTM-LD). However, in individuals without obvious pre-existing risk factors, it is likely that multiple risk factors – some genetic or other acquired factors – may collude to increase their vulnerability. While there have been case reports and small case series linking genetically identical NTM in patients to their home environment, the specific factors facilitating their acquisition remains poorly characterized. These factors encompass (i) varied sources of infection, (ii) modes of aquisition, and (iii) other physical aspects of the environment such as temperature, humidity, air exchange, surface types, and turbulence created by wind and natural disasters, and (iv) human behaviors, the combination of which are likely to be relatively unique among affected individuals (Honda et al., 2015; Nishiuchi et al., 2017).
Herein, we discuss the traditional environmental niches associated with NTM organisms, but also review the lesser-recognized environmental locales that NTM colonize including non-traditional niches and non-human hosts. We also summarize previous studies that link ecological factors with risk for infection and epidemiological information. Finally, we introduce new, ongoing work to study the particular environmental drivers of NTM emergence in Hawai’i, a geographic location deemed a major hot spot for NTM-LD.
The interaction of humans with their natural and built environments along with changes humans make to their environment (e.g., installation of mechanical devices that change environmental temperature and humidity, or others), and differences in the robustness of human health may impact the emergence of infectious diseases. The origins of NTM-LD remain a mystery, but may be intrinsically linked to human interaction with their environments. As soon as early humans learned to prepare small, controlled fires, the light and heat produced gathered people together for social interactions, vastly improved food preparation, and helped communities defend against invasion – activities that significantly lengthened human survival (Chisholm et al., 2016). Because NTM are found in the environment, it is plausible that these bacteria were historically aerosolized from soil with increased fire-making, infecting human lungs already impacted by long-term exposure to campfire smoke. Nonetheless, there is no evidence to show that the number of NTM-LD cases increase after camping or large scale environmental fires. For now, the origins of lung disease caused by NTM organisms remains an area of investigation.
The ability of NTM to cause LD and its clinical relevance varies globally. Geographic distribution of NTM species provides information regarding geographic-specific drivers of exposure such as climate, environment, and host factors associated with NTM-LD that would be specific to a global region (Falkinham, 1996; Griffith et al., 2007).
Mycobacterium avium complex (MAC) is the most frequently isolated group of NTM species worldwide and the most common organism associated with NTM-LD. MAC consists of various species of slow-growing mycobacteria (SGM) including M. avium, M. intracellulare, M. chimaera, M. colombiense, M. marseillense, M. arosiense, M. timonense, M. bouchedurhonense, and M. ituriense. Subspecies of M. avium include avium, silvaticum, hominissuis, and paratuberculosis (Hoefsloot et al., 2013; Johnson and Odell, 2014). Currently, the lowest number of MAC isolates in the world are seen in South America, but this may be because there is little NTM information available from this region (Hoefsloot et al., 2013; Halstrom et al., 2015; Stout et al., 2016). High incidence of NTM-LD due to MAC, M. kansasii, M. gordonae, and M. malmoense are observed in Europe, North America and Australia.
Rapid-growing mycobacteria (RGM) including the Mycobacterium abscessus and Mycobacterium fortuitum groups also contribute to a large proportion of NTM-LD cases globally. The Mycobacterium abscessus group is comprised of three subspecies: M. abscessus subsp. abscessus, M. abscessus subsp. massiliense, and M. abscessus subsp. bolletii. In the United States (U.S.), M. abscessus complex infections are secondary only to MAC infections, comprising 3-13% of all NTM-LD cases (Lee et al., 2015). The M. abscessus group is also commonly observed in patients in East Asia; in Taiwan 17.2% of all clinical NTM isolates belong to the M. abscessus group (Lai et al., 2010). The M. fortuitum group comprises M. fortuitum, M. peregrinum, M. senegalense, M. alvei, M. houstonense, M. neworleansense, M. boenickei, M. septicum, and M. porcinum (Adekambi and Drancourt, 2004; Schinsky et al., 2004). Besides LD, M. fortuitum cause soft tissue, skeletal, catheter-related, and disseminated infections in immunocompromised patients (Brown-Elliott and Wallace, 2002).
The most commonly occurring NTM species in many parts of U.S. are MAC and M. kansasii (Hoefsloot et al., 2013). Nearly 80% of all NTM-LD in the U.S. is due to a species of MAC, followed by M, kansasii - the second most common NTM associated with LD (Falkinham, 1996; Griffith et al., 2007). The U.S. states with the highest overall risk for NTM-LD include Hawai’i, California, New York, Louisiana, Pennsylvania, Florida, Oklahoma and Wisconsin (Adjemian et al., 2012a). M. abscessus is the most commonly recovered RGM from southeastern parts of U.S. (Halstrom et al., 2015; Stout et al., 2016). For unknown reasons, NTM-LD cases are lowest in North Dakota, South Dakota, Minnesota, Michigan, New Mexico, and West Virgina (Adjemian et al., 2012a).
Higher isolation rates of MAC (44%) are seen in northern Europe as compared to southern Europe (31%) with M. avium as the most prevalent species. M. kansasii is the predominant NTM species to cause LD in London, United Kingdom. and M. lentiflavum is most frequently isolated from clinical samples in Crete, Greece (Neonakis et al., 2007; Wassilew et al., 2016). Slovakia, Poland, and the United Kingdom have the highest amount of M. kansasii isolates in Europe (Hoefsloot et al., 2013; Wassilew et al., 2016). In contrast, higher isolation rates for M. xenopi are observed in southern Europe (21%) as compared to northern Europe (6%), but are most commonly isolated in Hungary (46%) (Hoefsloot et al., 2013; Wassilew et al., 2016). RGM are more commonly seen in the United Kingdom and Greece as compared to the rest of Europe (Hoefsloot et al., 2013). While not considered to be a pathogen, M. gordonae is most commonly recovered from environmental sources in Canada and Europe (Hoefsloot et al., 2013; Halstrom et al., 2015; Stout et al., 2016).
A recent study in China observed that an increase in latitude was associated with higher isolation rates of MAC species (predominantly M. intracellulare) whereas the number of RGM (most commonly M. chelonae) increased with a decrease in latitude (Yu et al., 2016). A similar trend is observed in Taiwan, with higher cases of NTM due to MAC recovered in the north and RGM like M. abscessus in the south (Huang et al., 2017). M. scrofulaceum and M. szulgai are also intermittently found in respiratory specimens from Asia (Simons et al., 2011). Overall, elderly women are disproportionately affected by NTM-LD, but in Saudi Arabia and most of the Persian Gulf countries, elderly men are found to be more affected (perhaps due to a lifetime of extended outdoor exposure) with MAC and M. abscessus being the main causative agents (Al-Ghafli and Al-Hajoj, 2017). MAC, M. simiae, and M. marinum are most commonly observed in NTM-LD individuals in Oman (Al-Ghafli and Al-Hajoj, 2017). In Western Asia, M. fortuitum and M. flavescens are the most prevalent RGM and SGM organisms, respectively with much higher frequency of NTM in northern Iran (73.2%) (Khaledi et al., 2016). In India, M. abscessus, M. fortuitum, and M. intracellulare are most commonly isolated from clinical samples (Desikan et al., 2017). M. abscessus are more widespread in Singapore and Okinawa (Hoefsloot et al., 2013).
Similar to the aforementioned geographic areas, LD caused by NTM are also increasingly observed in Australia (Thomson, 2010). MAC is the most commonly isolated NTM in Queensland with M. intracellulare comprising nearly 80% of the MAC isolates (Hoefsloot et al., 2013). RGM are the second most common cause of NTM-LD with M. abscessus the most commonly recovered from southern Australia (Hoefsloot et al., 2013; Wassilew et al., 2016). Unlike many other regions of the world, NTM are notifiable infections under the Queensland Public Health Act, 2005 which has facilitated the surveillance of potentially highly virulent and transmissible NTM strains (Thomson et al., 2017).
Information regarding the causative agents of NTM-LD in Africa are limited and is likely due to the overwhelming burden of tuberculosis in the regions. However, M. abscessus, M. avium, M. fortuitum, and M. nebraskense are recognized as the most frequently isolated NTM species from clinical samples in Zambia (Monde et al., 2018). M. gordonae has been recently found to be highly prevalent in water reservoirs like borehole wells, rivers dams, and tap water (Monde et al., 2018).
NTM are slow-growing compared to other types of bacteria, with the ability to form biofilms, resist high temperatures, and grow in marginal environments with low nutrient and oxygen content (Kirschner et al., 1992; Falkinham, 2009). However, cell surface hydrophobicity is the major driver sustaining NTM in the biofilms of both natural waters and man-made drinking water distribution systems, hospitals, and household plumbing. Due to their repulsion to water, NTM are found in aerosolized particles present above natural water bodies, showerheads, humidifiers, hot tubs and spas as well as in biofilms that form in these places (Falkinham, 2013). Environmental factors such as high humidity levels and high evapotranspiration (movement of water from land to the atmosphere) rates are known to be associated with an increased risk of NTM infection in susceptible individuals (Adjemian et al., 2012a). This is particularly translatable to the recovery of NTM during different seasons. For example, the species of NTM isolated from municipal water distribution systems in Brisbane, Australia differed in the samples collected in summer as compared to those collected in the winter months with higher numbers of M. gordonae, M. kansasii, M. abscessus, M. mucogenicum, and MAC isolated in the winter (Thomson R. M. et al., 2013).
Other environmental factors have a profound impact of NTM viability. For example, NTM have been isolated from water bodies with moderate salinity (1–2% NaCl) like estuaries (Chesapeake Bay) (Kirschner et al., 1992; Falkinham, 2009). But in a separate study, reduced numbers of NTM isolates were observed when water salinity exceeded 2% (Gruft et al., 1981). NTM also favor environments with acidic pH. Humic and fulvic acids and acidic brown water swamps along the southeastern coast of the U.S. support high numbers of MAC (Falkinham, 2009, 2013). Pine forest (boreal rich) and peat rich soils, brackish marshes, and drainage water are also rich in NTM (Kirschner et al., 1992; Falkinham, 2009). Minerals widely found in clay soils such as kaolin and dust have also been demonstrated to facilitate the growth of M. abscessus (Malcolm et al., 2017).
M. avium, M. fortuitum, M. chelonae, M. kansasii, M. gordonae, and M. xenopi are the NTM species most commonly found in water distribution systems, water bodies including lakes, rivers and streams as well as soil and dust (Falkinham, 2009; Ulmann et al., 2015). Falkinham et al. (2008) was the first to demonstrate that the M. avium isolated from a patient with NTM-LD had a clonal relationship with the M. avium isolated from her home showerhead biofilm. Falkinham also first reported identical NTM DNA fingerprints from patients’ sputa and matched shower water isolates, shedding light on the paradigm that inhalation of aerosols while showering is a likely mode of NTM acquisition (Falkinham, 2011). Similar reports are observed from Japan where MAC isolates recovered from bathtub inlets and showerheads showed identical pulse-field gel electrophoresis profiles when compared to their respective clinical isolates (Nishiuchi et al., 2009; Ichijo et al., 2014). Thus, showerhead biofilms remain one of the most frequently sampled environmental sources used to describe the presence of NTM globally. In an interesting turn of events, early methods such as hybridization probes and multiplex 16S rRNA gene PCR methodologies identified M. intracellulare in households and potable water (Tichenor et al., 2012; Whiley et al., 2012). However, Falkinham et al. (2008) and Wallace et al. (2013) reanalyzed environmental household water and biofilm isolates originally identified as M. intracellulare by sequencing the 280 bp 16S to 23S internal transcribed spacer region and discovered that these isolates were instead M. chimaera (Thomson, 2010; Falkinham, 2011; Koh W. J. et al., 2012; Tichenor et al., 2012). Using the same sequencing method, isolates originally called M. avium were confirmed as M. avium. Thus, M. chimaera is now widely recognized in water biofilms, while M. intracellulare is found to be absent from them. A clue to their sources may come from a study conducted in American Samoa where M. intracellulare was identified in roof-harvested rainwater (Kirs et al., 2017) and soil (Honda et al., 2016).
NTM found in households are likely piped into home plumbing systems from public utility sources where biofilms are commonly formed. However, NTM are also known inhabitants of natural freshwater ecosystems. Of two recreational lakes, RGM were the dominant NTM, but a diversity of other mycobacteria were found in high density in the water column, air-water interface, sediment, and in association with benthic algae growing on plants and fine sediment using quantitative real-time PCR and the MiSeq Illumina platform (Roguet et al., 2016). Yet, NTM remain seldomly recovered from well and groundwaters (Martin et al., 1987).
Nosocomial NTM lung infections have been reported in the literature. MAC species have been detected in hospital potable hot water distribution systems, hospital tap water used for dialysis, and in water used to prepare medical solutions, highlighting their propensity to stick to piped surfaces (Bolan et al., 1985; Safranek et al., 1987; Hector et al., 1992). After a significant increase in NTM-positive sputa was observed from patients referred to respiratory wards in Rome, an infection control investigation revealed a massive presence of NTM in the hospital water network (D’Antonio et al., 2016). In another study, 83% of U.S. dialyses centers examined showed NTM in municipal water supplies (Carson et al., 1988). More recently, global outbreaks of M. chimaera associated with heater-cooler units used during open-heart surgery have provided unique challenges for the medical community (Sax et al., 2015; Schreiber et al., 2016; Marra et al., 2017). Investigations point to model-specific designs in air flow direction, location of cooling ventilators, and the continuous cooling of unit water tanks significantly increased the risk of disseminating colonized M. chimaera into the air of operating rooms (Kuehl et al., 2018). Of the first thirty cases affected by this outbreak in the United Kingdom, 60% (18/30) died at a median of 30 months after initial surgery (Scriven et al., 2018). Poor disinfection and resistance to glutaraldehydes have been highlighted in pseudo-outbreaks of M. abscessus subsp. bolletii in bronchoscopes, endoscopes, and disinfection units (Guimaraes et al., 2016). In other areas of the hospital, patient accessible ice machines have been shown to be laden with NTM, particularly M. paraffinicum and M. fortuitum (Gebo et al., 2002; Wang et al., 2009).
M. fortuitum skin infections have been associated with pedicure-associated whirlpool footbaths in California and Georgia nail salons (Winthrop et al., 2004). Skin infections due to other NTM such as M. chelonae have been linked to contaminated water used for diluting tattoo ink and to unsterilized instrumentation (Mudedla et al., 2015). Dental unit water lines have also been shown to harbor a variety of environmental NTM species (Schulze-Robbecke et al., 1995), but remain as unproven sources of NTM lung infection. MAC-associated hypersensitivity pneumonitis (HP) has been linked to exposure to warm, bubbly water found in rarely cleaned hot tubs and spa baths (Embil et al., 1997; Sugita et al., 2000; Rickman et al., 2002). However, MAC has also been isolated from cold water sources including swimming pool water. In fact, lifeguards with long-term exposure to indoor swimming pool aerosols are susceptible to work-associated exposures and are at increased risk for MAC-associated HP (Rose et al., 1998; Koschel et al., 2006). Besides tap water, a Dutch group found NTM in swimming pools and whirlpool water (Havelaar et al., 1985). NTM-associated HP has also been linked to occupational exposures to aerosols produced through manipulation of metalworking fluids (Bernstein et al., 1995; Wilson et al., 2001).
Soil is also a widely recognized environmental niche for NTM. NTM patients’ potting soils yielded NTM that were identical by DNA fingerprinting to the NTM isolates from the same patients’ lungs (Ichiyama et al., 1988; De Groote et al., 2006). Moreover, M. avium subsp. hominissuis (MAH) was predominant in soil and dust, but not identified in German water and biofilm samples by culture (Lahiri et al., 2014). A study in Iran found that 6-15% of soil samples compared to 10–27% of water samples collected from the suburbs of Tehran had NTM isolated by culture with M. farcinogens and M. fortuitum being the most common species (Velayati et al., 2014). Furthermore, the risk for acquiring NTM is significantly higher in communities engaged in occupations that generate aerosols and are exposed to soil for a longer time (e.g., agriculture, mining, landscaping, and tunnel work) as compared to communities that have a limited exposure to soil (Hamada et al., 2016). In West Harima, Japan, NTM-LD was associated with natural resource activities, construction, mining, and soil exposure (Hamada et al., 2016). In a separate study, M. chelonae, M. fortuitum, and M. kansasii were identified in 85% of the alpine and subalpine soil, peat, humus, porous ricks, mosses, and wood examined suggesting NTM thrive in mountain ranges and elevations (Thorel et al., 2004). Rare and unique NTM species have also been described in polluted soils of Hawai’i where they functioned as polycyclic aromatic hydrocarbon pollutant-degrading organisms (Hennessee et al., 2009).
NTM have been reported in kitchen sink biofilms as well as household refrigerator taps and home ice machines (Ichijo et al., 2014). MAC organisms have been found to colonize point-of-use filters used to filter tap water including carbon filers impregnated with silver (Rodgers et al., 1999; Hamilton et al., 2017). Due to the appearance of disseminated M. avium infections during the height of the HIV-AIDS epidemic, two studies tested for the presence of NTM in foods consumed by HIV-infected patients. PCR typing revealed 29 different mycobacterial isolates in 21% (25/121) of food samples tested; 41% of the 29 samples (n = 12) were M. avium (Yoder et al., 1999). One of the clinical M. avium isolates was identical to a food isolate, suggesting food as a potential source of M. avium infections. In the second study, water, food and soil samples from 290 homes of HIV-infected patients were tested for mycobacteria using DNA probes, serotyping, and multi-locus enzyme electrophoresis and compared to clinical isolates (Yajko et al., 1995). Soil, rather than water sources, were found to harbor more M. avium. While not considered a food-borne illness, M. avium subspecies DNA was also identified in raw meats (Lorencova et al., 2014). In contrast, smoked fish products did not show NTM, but 12% of samples collected from pond fish (4%), retail sold fish (61%), and frozen fish (91%) contained NTM DNA (Lorencova et al., 2013). Although unproven, the acid-resistant NTM organisms may remain viable in the stomach where food is consumed and digested. Evidence suggests patients with the nodular bronchiectatic form of NTM-LD have a high prevalence of increased esophageal acid exposure and gastroesophageal reflux disease was found to be significantly associated with RGM organisms (Winthrop et al., 2010; Koh E. et al., 2012).
Besides their well-known habitation in the numerous sources detailed above, rare NTM species (e.g., M. algericum, M. arabiense, M. heraklionense) have been cultured and identified from primary sludge samples of water treatment plants even after decontamination (Makovcova et al., 2015). M. avium, M. gordonae, and M. flavescens have been also been identified in non-traditional water sources including untreated, drinking well water in rural areas of Montana as well as in treated municipal wastewater from arid regions (Richards et al., 2015; Amha et al., 2017). Qualitative assessments were used to determine the risk of MAC exposure in Queensland, Australia, an area that utilizes rainwater catchment systems. Untreated rainwater is commonly used for showering, car washing, toilet flushing, and food preparation (Hamilton et al., 2017). But in this study, rainwater used for drinking presented the greatest risk for MAC infection; yet the species of MAC responsible was not reported. In most cases, disinfection methods to remove potential pathogens from these water sources is a decision left up to the household and can range from no disinfection methods to point-of-use filters, UV irradiation, solar disinfection, chlorine or a combination of them. More work is needed in these areas to determine the impact of using different disinfection methods to reduce exposures in NTM patients who use these water source types.
NTM have also been detected, albeit, rarely in cobwebs above hen nests, soil fertilized with chicken droppings, and moss but more commonly in dust from vacuum cleaners, and air conditioners (Nishiuchi et al., 2009; Kaevska et al., 2011). Of particular significance to smokers is the recovery and identification of M. avium from cigarettes (Eaton et al., 1995). While not directly shown to cause LD, MAC has been identified from clothing washed during laundry cycles suggesting laundry water maybe an unintentional source of household NTM (Yajko et al., 1993). Finally, M. avium was detected in samples of condensation water formed from the coagualation of steam in three different rooms inside the Russian space station, Mir (Kawamura et al., 2001).
Amoeba are free-living, freshwater associated protozoans that are ubiquitously found in water systems often cohabited by NTM. Many species of amoeba phagocytose free-living bacteria and feed on them; however, some NTM are able to resist and evade degradation. In particular, most species of MAC, including M. avium, M. intracellulare, M. chimaera, M. colombiense, M. arosiense, M. marseillense, M. timonense, and M. bouchedurhonense reside within free-living Acanthamoeba polyphaga and their exocysts as well as A. castellanii found in potable water (Cirillo et al., 1997; Taylor et al., 2009; Ben Salah and Drancourt, 2010). Delafont et al. (2014) demonstrated amoeba-mycobacteria associations in drinking water networks in a year-long sampling study. Nearly 88% of amoeba including the genera Acanthamoeba, Vermamoeba, Echinamoeba, and Protacanthamoeba recovered from drinking water were found to contain M. llatzerense and M. chelonae (Delafont et al., 2014). NTM cultured in amoeba also show increased resistance to antibiotics and enhanced virulence compared to NTM grown in chickens and mice (Cirillo et al., 1997; Falkinham et al., 2001).
NTM infections in mammals occur sporadically and are rarely transmissible between animals and seldomly considered bone fide zoonotic diseases. Except under particular scenarios, NTM infections are also generally not transmissible from human to human (Bryant et al., 2016). Nonetheless, NTM have been known to infect animals such as chickens and quails (Meissner and Anz, 1977; Morita et al., 1999). Drug-susceptible M. fortuitum and M. abscessus were identified in a cutaneous lesion on the snout and nostrils of a captive Trichechus inunguis (manatee) in the Amazon (Reisfeld et al., 2018). In captive non-domesticated hoofed animals and in immunosuppressed dogs and cats, M. avium has been reported as disseminated disease (Thorel et al., 2004). With the potential for cross species infection or transmission in the Serengeti, tissues from wildlife species and indigenous cattle were probed for mycobacteria, revealing M. intracellulare as the most frequently isolated species, followed by M. lentiflavum, M. fortuitum, and M. chelonae/abscessus. MAC organisms were also detected in animal feces and huts from pastoral Uganda (Kankya et al., 2011). Because M. avium isolates from pigs showed shared genetic characteristics to M. avium isolated from humans, pigs have been theorized as potential sources of infection (Bono et al., 1995). Among 1,249 mandibular lymph node samples collected from the wild boar, Sus scrofa, between 2007 and 2011 in Spain, M. chelonae and M. avium represented 61 and 11% of the NTM isolates (Garcia-Jimenez et al., 2015). In a separate study, a low degree of similarity between MAH isolates from Japanese patients and local pigs was found, while there was a high degree of similarity between European patient and pig isolates, suggesting geographic distinctions (Iwamoto et al., 2012).
NTM infections are among the most common chronic disease of aquatic animals (Gcebe et al., 2018). After their original discovery in 1897, NTM organisms continue to cause disease in sea bass, mullets, and amberjacks that live in temperate zones (Lescenko et al., 2003). The most common NTM pathogens of fish include M. chelonae (sea horses), M. avium, and M. fortuitum. M. marinum remains the most typical NTM found in aquatic environments and often coinhabit on shrimp, frogs, eels, oysters, and shellfish (Piersimoni and Scarparo, 2009). M. marinum is one of the only known species of NTM that grow in waters with high salt concentrations (>3% NaCl) (Kirschner et al., 1992; Falkinham, 2009). In some cases, M. marinum cause extrapulmonary infections and cutaneous “fish tank granulomas” in humans. Controlling NTM infections in the aquaculture setting is difficult, relying only on destruction of the infected stocks in the absence of effective treatments (Gcebe et al., 2018). Carassius auratus (goldfish) have been evaluated as a novel in vivo model to study the pathogenesis of M. marinum. After intraperitoneal administration of 102 and 109 CFU of M. marinum organisms, an acute and chronic infection was respectively observed with high recovery of NTM from inoculated animals (Talaat et al., 1999).
Although seldom reported, reptiles and insects can potentially carry pathogenic NTM. M. chelonae was originally isolated from the lungs of sea turtles in 1903 (Männikkö, 2011), M. intracellulare has been identified in a rusty monitor reptile with lung nodules (Friend and Russell, 1979), and M. szulgai has been isolated from a crocodile showing tuberculosis-like lung lesions (Gcebe et al., 2018). In a separate case, M. gordonae, M. avium, and M. kansasii were isolated off cockroaches from a South Taiwan hospital (Pai et al., 2003).
Mycobacterium ulcerans can infect skin and subcutaneous tissues developing into non-ulcerated nodules or lesions. Although M. ulcerans has not yet been cultured from the environment, its DNA has been detected in low levels in suspended solids/water residues and soil. High concentrations of M. ulcerans DNA are observed in the feces of Australian ringtails and brushtails possums residing in locations where human cases were reported. These findings suggest possums are naturally infected and are potential environmental reservoirs (Fyfe et al., 2010).
To understand the environmental risk factors for infection, most studies described above have either probed for the occurrence of NTM in various environmental sources or in pulmonary samples from various geographic areas. Yet, the number of studies that have overlaid and integrated ecological and geographic information with epidemiological information of NTM-LD risk are scant. However, an exciting recent study has enhanced our understanding of how ecology relates to risk for NTM infection. In Colorado, soil acidity, low manganese concentrations, and silt were significantly associated with increased disease risk for NTM (Lipner et al., 2017). This same study also identified high-risk clusters of NTM-LD and high-risk watershed locations in the same geographic region using spatial scanning methods. Another study from Queensland, Australia (2001–2010) investigated the associations between climate, soil characteristics, land use, demographic, and socio-economic variables with spatial patterns of NTM infection (Chou et al., 2014). High risk clusters of M. kansasii, M. intracellulare, and M. abscesses infections were found to be associated with areas of high agricultural, mining, and tourism activity.
Higher rates of NTM lung infections have been associated with oceanic coastlines, accounting for 70% of annual NTM cases in the U.S. (Strollo et al., 2015). Statistically significant increases in NTM identification have been reported in the U.S. Affiliated Pacific Islands (USAPI) including American Samoa, Guam, Northern Marina Islands, Palau, Marshall Islands, and the Federated States of Micronesia (Lin et al., 2018). After analyzing respiratory cultures submitted for species identification between 2007 and 2011 at a clinical reference laboratory, the overall period prevalence of NTM isolation in this study was 106 cases/100,000 persons. The authors acknowledge that the prevalence of NTM isolation varied by island nation and may be related to urbanization. The lowest period prevalence of NTM isolation (22 cases/100,000 persons) was reported in American Samoa with 87.2% of patients living in high urban areas. The highest period prevalence (164/100,000 cases) was observed in respiratory samples from the Federated States of Micronesia which also showed the lowest percentage of urban dwellers (22.4%).
Of interest, the first nationwide population-based analysis on the prevalence of NTM-LD found that Hawai’i had the highest period prevalence (1997–2007) of any U.S. state with 396 cases/100,000 persons among persons > 65 years-old (Adjemian et al., 2012b), a rate that is four times greater than the national average. Using national Medicare claims, U.S. Census data, as well as U.S. Geological and Forest Services environmental and climatic data, high and low risk U.S. counties were identified (Adjemian et al., 2012a). High-risk regions for NTM were noted in particular geographic areas of Hawai’i. Higher daily evapotranspiration levels, higher number of surfaces covered by water, higher soil copper and sodium, and lower manganese levels were characteristics of high risk areas. More ominous, Hawai’i also shows the highest, national age-adjusted mortality rates from NTM-LD (Mirsaeidi et al., 2014).
In a recent large-scale study, researchers partnered with a network of citizen scientists to collect showerhead biofilm samples from 606 households across 49 of 50 U.S. states to simultaneously superimpose the identities of showerhead microbes over geographic areas where NTM-LD is most prevalent (Gebert et al., 2018). Multiple concluding findings were reported that reinforce known paradigms. First, Mycobacteria was the most abundant group of bacteria identified in showerhead biofilms by 16S rRNA gene sequencing, corroborating prior studies (Feazel et al., 2009; Thomson R. et al., 2013). Next, the most frequently identified NTM species by hsp65 gene sequencing were MAC, M. abscessus, and the M. fortuitum complex which are also the most problematic NTM for patients. As already observed (Garcia et al., 2013; Ovrutsky et al., 2013), mycobacteria often co-occurred with free-living amoeba. Second, similar to other studies (Feazel et al., 2009), homes using municipal treated water showed twice more mycobacteria than homes using water from wells and the former contained higher chlorine and iron concentrations. Third, by overlapping these data with previously reported epidemiological information, this study is the first to show showerhead-associated mycobacteria with the potential to cause pathogenic LD are indeed found in U.S. households in regions with the highest number of NTM-LD cases including Florida, southern California, and northeast states. It is clear mycobacterial lineages showed distinct geographic distributions. Finally, supporting prior work, this study confirms Hawai’i as the state with the most abundant mycobacterial showerhead biofilms and show these are predominated by MAC and M. abscessus organisms. Undoubtedly, Hawai’i is a U.S. hot spot for NTM-LD (Figure 1A) and the islands’ unique characteristics may contribute to greater exposures.
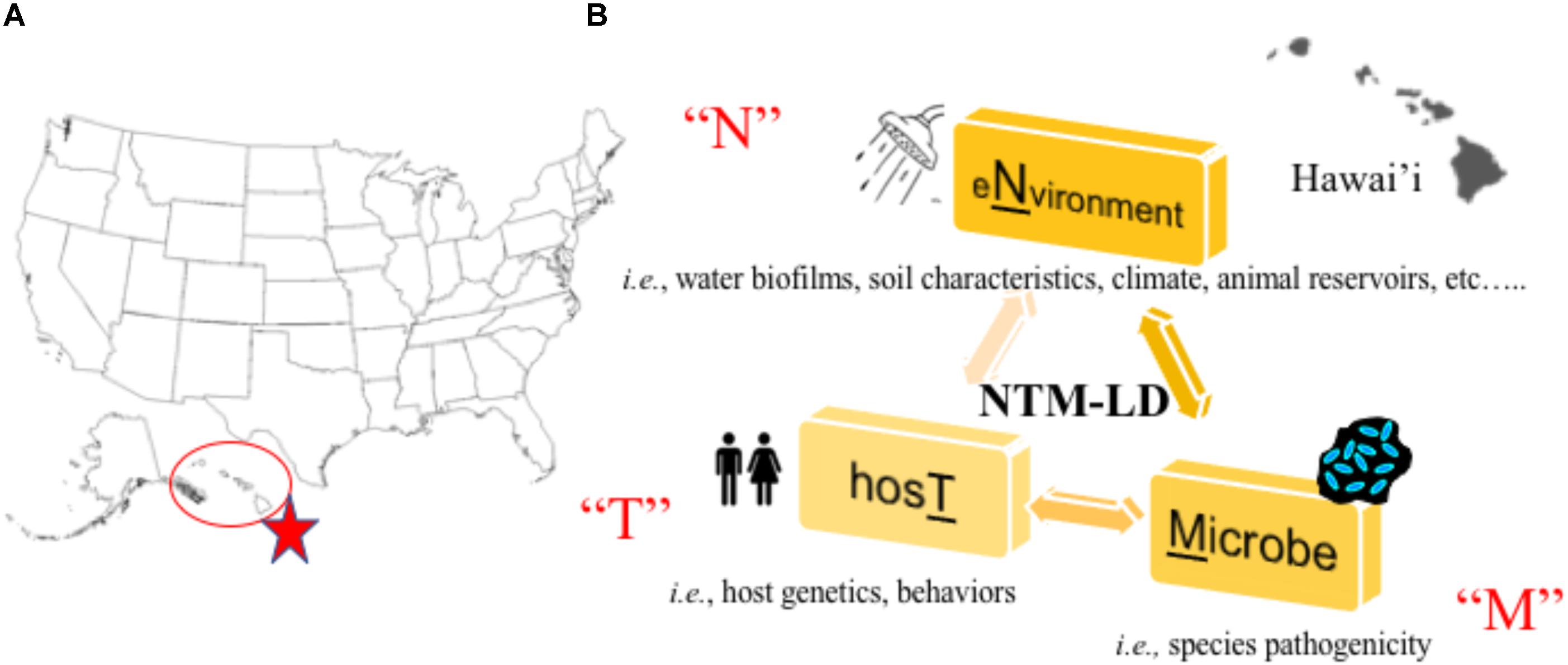
FIGURE 1. (A) Hawai’i is a hot spot for NTM lung disease in the United States (Rudis, 2018). (B) Our research program aims to study the “N-T-M” specific factors that contribute to lung disease by studying the environmental, host, and microbial factors that facilitate NTM lung disease emergence in Hawai’i.
We have previously shown increased prevalence of NTM infection in Hawai’i using patient laboratory data from a representative population of in-state residents enrolled in a closed healthcare system (Adjemian et al., 2017). Indeed, the most frequently isolated species was MAC, followed by M. fortuitum that was more common among Vietnamese and Korean patients, while M. abscessus was more associated with Japanese and Filipino patients. In this study, Native Hawaiian/Other Pacific Islander populations were less likely to have NTM infection than other racial/ethnic groups examined. A logical hypothesis is that native populations inhabited the islands for a longer period of time, evolving and developing resistance to infection.
We have also previously demonstrated not only the overwhelming presence of MAC in both clinical samples in respiratory specimens from Hawai’i, but also in water biofilm samples collected from households in Hawai’i (Honda et al., 2016). Using partial rpoB gene sequencing, M. avium was not identified in any of the samples examined, providing a contrast to previous studies from the continental U.S. M. intracellulare was found in 27% (4/15) SGM respiratory specimens examined and a soil sample. While absent from soil, M. chelonae was significantly more common in kitchen sink biofilms (9/35, 35%) compared to bathroom sink biofilms (3/34, 9%) and M. abscessus was found equally in kitchen (5/34, 15%) and bathroom sink (4/30, 13%) biofilms. M. porcinum was also frequently recovered from showerhead biofilm samples, a species normally associated with swine, but also a cause of human infections (Wallace et al., 2004; Brown-Elliott et al., 2011; Perez-Martinez et al., 2013). This observation may be particularly important in Hawai’i, which has a sizeable feral pig population (Hess et al., 2006). Importantly, we identified M. chimaera as the predominant species of MAC in these islands; in fact, M. chimaera was recovered from 56% of the 75 environmental samples tested and 67% of ten clinical isolates tested (Honda et al., 2016). By comparison, of 8,800 isolates analyzed using rpoB gene sequencing in 26 months at National Jewish Health, only 6% were M. chimaera (Dr. Max Salfinger, National Jewish Health, personal communication). Taken together, M. chimaera is emerging as a major NTM species of interest in Hawai’i, underscoring the need for further studies to define the drivers for NTM emergence there and in other Pacific Islands.
In new and on-going work to understand the Hawai’i specific environmental, host, and NTM factors that contribute to NTM-LD emergence (Figure 1B), we are synergizing environmental and human behavioral/genetic findings with microbiological and NTM genomic data in dynamic statistical, spatial/temporal models to uncover not only disease drivers, but also potential points of intervention to prevent future infections. Isolates collected through this work will also be applied in future studies of NTM virulence. To accomplish this, a complementary team has been formed including a NTM microbiologist born and raised in Hawai’i with significant ties to the local community, a mycobacterial pulmonologist, epidemiologist, earth geochemist, climatologist, microbial ecologist, volcanic scientists, ecological modelers, local clinicians, and a team of microbial genomic and computational scientists as well as Hawai’i residents, high school/college students, their mentors, and local pig hunters.
Future studies should investigate the: (i) factors that drive the relative absence of NTM from seawater, but association with higher humidity and associated biofilms collected from different areas of the world, (ii) NTM prevalence and diversity in areas of the world with endemic tuberculosis, (iii) role of animal and protozoal reservoirs in the maintenance and spread of environmental NTM, and (iv) contributions of temperature, evapotranspiration, and air pollution including climatic determinants of disease. In the meantime, we spotlight the NTM crisis in Hawai’i as a useful model system to understand NTM transmission and disease dynamics. New information gathered from this work will then be used and applied to study NTM-LD in other areas of the world where NTM-LD is prevalent and emergent.
JH conceived the project. JH, RV, and EC wrote and edited the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
JH would like to thank the Shoot for the Cure and Padosi Foundations. The authors also acknowledge Drs. Michael Strong, Stephen Nelson, James Crooks, Krishna Pacifici and Stacey Honda as co-PI and collaborators on our work supported by the National Science Foundation, Ecology, Evolution, and Infectious Disease Program, Award # 1743587. Assistance to create the U.S. map shown in this manuscript provided by James Crooks.
Adekambi, T., and Drancourt, M. (2004). Dissection of phylogenetic relationships among 19 rapidly growing Mycobacterium species by 16S rRNA, hsp 65, sodA, recA and rpoB gene sequencing. Int. J. Syst. Evol. Microbiol. 54,2095–2105.
Adjemian, J., Frankland, T. B., Daida, Y. G., Honda, J. R., Olivier, K. N., Zelazny, A., et al. (2017). Epidemiology of nontuberculous mycobacterial lung disease and tuberculosis, Hawaii, USA. Emerg. Infect. Dis. 23, 439–447.
Adjemian, J., Olivier, K. N., Seitz, A. E., Falkinham, J. O. III, Holland, S. M., and Prevots, D. R. (2012a). Spatial clusters of nontuberculous mycobacterial lung disease in the United States. Am. J. Respir. Crit. Care Med. 186, 553–558.
Adjemian, J., Olivier, K. N., Seitz, A. E., Holland, S. M., and Prevots, D. R. (2012b). Prevalence of nontuberculous mycobacterial lung disease in U.S. Medicare beneficiaries. Am. J. Respir. Crit. Care Med. 185, 881–886.
Al-Ghafli, H., and Al-Hajoj, S. (2017). Nontuberculous mycobacteria in saudi arabia and gulf countries: a review. Can. Respir. J. 2017:5035932.
Amha, Y. M., Anwar, M. Z., Kumaraswamy, R., Henschel, A., and Ahmad, F. (2017). Mycobacteria in municipal wastewater treatment and reuse: microbial diversity for screening the occurrence of clinically and environmentally relevant species in arid regions. Environ. Sci. Technol. 51, 3048–3056.
Ben Salah, I., and Drancourt, M. (2010). Surviving within the amoebal exocyst: the Mycobacterium avium complex paradigm. BMC Microbiol. 10:99. doi: 10.1186/1471-2180-10-99
Bernstein, D. I., Lummus, Z. L., Santilli, G., Siskosky, J., and Bernstein, I. L. (1995). Machine operator’s lung. A hypersensitivity pneumonitis disorder associated with exposure to metalworking fluid aerosols. Chest 108, 636–641.
Bolan, G., Reingold, A. L., Carson, L. A., Silcox, V. A., Woodley, C. L., Hayes, P. S., et al. (1985). Infections with Mycobacterium chelonei in patients receiving dialysis and using processed hemodialyzers. J. Infect. Dis. 152, 1013–1019.
Bono, M., Jemmi, T., Bernasconi, C., Burki, D., Telenti, A., and Bodmer, T. (1995). Genotypic characterization of Mycobacterium avium strains recovered from animals and their comparison to human strains. Appl. Environ. Microbiol. 61, 371–373.
Brown-Elliott, B. A., and Wallace, R. J. Jr. (2002). Clinical and taxonomic status of pathogenic nonpigmented or late-pigmenting rapidly growing mycobacteria. Clin. Microbiol. Rev. 15, 716–746.
Brown-Elliott, B. A., Wallace, R. J. Jr., Tichindelean, C., Sarria, J. C., Mcnulty, S., Vasireddy, R., et al. (2011). Five-year outbreak of community- and hospital-acquired Mycobacterium porcinum infections related to public water supplies. J. Clin. Microbiol. 49, 4231–4238.
Bryant, J. M., Grogono, D. M., Rodriguez-Rincon, D., Everall, I., Brown, K. P., Moreno, P., et al. (2016). Emergence and spread of a human-transmissible multidrug-resistant nontuberculous Mycobacterium. Science 354, 751–757.
Carson, L. A., Bland, L. A., Cusick, L. B., Favero, M. S., Bolan, G. A., Reingold, A. L., et al. (1988). Prevalence of nontuberculous mycobacteria in water supplies of hemodialysis centers. Appl. Environ. Microbiol. 54,3122–3125.
Chisholm, R. H., Trauer, J. M., Curnoe, D., and Tanaka, M. M. (2016). Controlled fire use in early humans might have triggered the evolutionary emergence of tuberculosis. Proc. Natl. Acad. Sci. U.S.A. 113, 9051–9056.
Chou, M. P., Clements, A. C., and Thomson, R. M. (2014). A spatial epidemiological analysis of nontuberculous mycobacterial infections in Queensland, Australia. BMC Infect. Dis. 14: 279. doi: 10.1186/1471-2334-14-279
Cirillo, J. D., Falkow, S., Tompkins, L. S., and Bermudez, L. E. (1997). Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect. Immun. 65, 3759–3767.
D’Antonio, S., Rogliani, P., Paone, G., Altieri, A., Alma, M. G., Cazzola, M., et al. (2016). An unusual outbreak of nontuberculous mycobacteria in hospital respiratory wards: association with nontuberculous mycobacterial colonization of hospital water supply network. Int. J. Mycobacteriol. 5,244–247.
De Groote, M. A., Pace, N. R., Fulton, K., and Falkinham, J. O. (2006). Relationships between Mycobacterium isolated from patients with pulmonary mycobacterial infection and potting soils. Appl. Environ. Microbiol. 72, 7602–7606.
Delafont, V., Mougari, F., Cambau, E., Joyeux, M., Bouchon, D., Hechard, Y., et al. (2014). First evidence of amoebae-mycobacteria association in drinking water network. Environ. Sci. Technol. 48, 11872–11882.
Desikan, P., Tiwari, K., Panwalkar, N., Khaliq, S., Chourey, M., Varathe, R., et al. (2017). Public health relevance of Non-tuberculous mycobacteria among AFB positive sputa. Germs 7, 10–18.
Eaton, T., Falkinham, J. O. III, and Von Reyn, C. F. (1995). Recovery of Mycobacterium avium from cigarettes. J. Clin. Microbiol. 33, 2757–2758.
Embil, J., Warren, P., Yakrus, M., Stark, R., Corne, S., Forrest, D., et al. (1997). Pulmonary illness associated with exposure to Mycobacterium-avium complex in hot tub water, hypersensitivity pneumonitis or infection? Chest 111, 813–816.
Falkinham, J. O. III, Iseman, M. D., De Haas, P., and Van Soolingen, D. (2008). Mycobacterium avium in a shower linked to pulmonary disease. J. Water Health 6, 209–213.
Falkinham, J. O., Norton, C. D., and Lechevallier, M. W. (2001). Factors influencing numbers of Mycobacterium avium, Mycobacterium intracellulare, and other mycobacteria in drinking water distribution systems. Appl. Environ. Microbiol. 67, 1225–1231.
Falkinham, J. O. III. (1996). Epidemiology of infection by nontuberculous mycobacteria. Clin. Microbiol. Rev. 9, 177–215.
Falkinham, J. O. III. (2009). Surrounded by mycobacteria: nontuberculous mycobacteria in the human environment. J. Appl. Microbiol. 107,356–367.
Falkinham, J. O. III. (2011). Nontuberculous mycobacteria from household plumbing of patients with nontuberculous mycobacteria disease. Emerg. Infect. Dis. 17, 419–424.
Falkinham, J. O. III. (2013). Ecology of nontuberculous mycobacteria-where do human infections come from? Semin. Respir. Crit. Care Med. 34, 95–102.
Feazel, L. M., Baumgartner, L. K., Peterson, K. L., Frank, D. N., Harris, J. K., and Pace, N. R. (2009). Opportunistic pathogens enriched in showerhead biofilms. Proc. Natl. Acad. Sci. U.S.A. 106, 16393–16399.
Friend, S. C., and Russell, E. G. (1979). Mycobacterium intracellulare infection in a water monitor. J. Wild. Dis. 15, 229–233.
Fyfe, J. A., Lavender, C. J., Handasyde, K. A., Legione, A. R., O’brien, C. R., Stinear, T. P., et al. (2010). A major role for mammals in the ecology of Mycobacterium ulcerans. PLoS Negl. Trop Dis. 4:e791. doi: 10.1371/journal.pntd.0000791
Garcia, A., Goni, P., Cieloszyk, J., Fernandez, M. T., Calvo-Begueria, L., Rubio, E., et al. (2013). Identification of free-living amoebae and amoeba-associated bacteria from reservoirs and water treatment plants by molecular techniques. Environ. Sci. Technol. 47, 3132–3140.
Garcia-Jimenez, W. L., Benitez-Medina, J. M., Martinez, R., Carranza, J., Cerrato, R., Garcia-Sanchez, A., et al. (2015). Non-tuberculous mycobacteria in wild boar (Sus scrofa) from Southern Spain: epidemiological, clinical and diagnostic concerns. Transbound. Emerg. Dis. 62, 72–80.
Gcebe, N., Michel, A. L., and Hlokwe, T. M. (2018). Non-tuberculous Mycobacterium species causing mycobacteriosis in farmed aquatic animals of South Africa. BMC Microbiol. 18:32. doi: 10.1186/s12866-018-1177-9
Gebert, M., Delgado-Baquerizo, M., Oliverio, A., Webster, T., Nichols, L., Honda, J., et al. (2018). Ecological analyses of mycobacteria in showerhead biofilms and their relevance to human health. bioRxiv [Preprint]. doi: 10.1101/366088
Gebo, K. A., Srinivasan, A., Perl, T. M., Ross, T., Groth, A., and Merz, W. G. (2002). Pseudo-outbreak of Mycobacterium fortuitum on a human immunodeficiency virus ward: transient respiratory tract colonization from a contaminated ice machine. Clin. Infect. Dis. 35, 32–38.
Griffith, D. E., Aksamit, T., Brown-Elliott, B. A., Catanzaro, A., Daley, C., Gordin, F., et al. (2007). An official ATS/IDSA statement: diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 175, 367–416.
Gruft, H., Falkinham, J. O. III, and Parker, B. C. (1981). Recent experience in the epidemiology of disease caused by atypical mycobacteria. Rev. Infect Dis. 3, 990–996.
Guimaraes, T., Chimara, E., Do Prado, G. V., Ferrazoli, L., Carvalho, N. G., Simeao, F. C., et al. (2016). Pseudooutbreak of rapidly growing mycobacteria due to Mycobacterium abscessus subsp bolletii in a digestive and respiratory endoscopy unit caused by the same clone as that of a countrywide outbreak. Am. J. Infect. Control. 44, e221–e226.
Halstrom, S., Price, P., and Thomson, R. (2015). Review: environmental mycobacteria as a cause of human infection. Int. J. Mycobacteriol. 4,81–91.
Hamada, S., Ito, Y., Hirai, T., Murase, K., Tsuji, T., Fujita, K., et al. (2016). Impact of industrial structure and soil exposure on the regional variations in pulmonary nontuberculous mycobacterial disease prevalence. Int. J. Mycobacteriol. 5, 170–176.
Hamilton, K. A., Ahmed, W., Toze, S., and Haas, C. N. (2017). Human health risks for Legionella and Mycobacterium avium complex (MAC) from potable and non-potable uses of roof-harvested rainwater. Water Res. 119, 288–303.
Havelaar, A. H., Berwald, L. G., Groothuis, D. G., and Baas, J. G. (1985). Mycobacteria in semi-public swimming-pools and whirlpools. Zentralbl. Bakteriol. Mikrobiol. Hyg. B 180, 505–514.
Hector, J. S., Pang, Y., Mazurek, G. H., Zhang, Y., Brown, B. A., Wallace, R. J., et al. (1992). Large restriction fragment patterns of genomic Mycobacterium fortuitum DNA as strain-specific markers and their use in epidemiologic investigation of four nosocomial outbreaks. J. Clin. Microbiol. 30, 1250–1255.
Hennessee, C. T., Seo, J. S., Alvarez, A. M., and Li, Q. X. (2009). Polycyclic aromatic hydrocarbon-degrading species isolated from Hawaiian soils: Mycobacterium crocinum sp. nov., Mycobacterium pallens sp. nov., Mycobacterium rutilum sp. nov., Mycobacterium rufum sp. nov. and Mycobacterium aromaticivorans sp. nov. Int. J. Syst. Evol. Microbiol. 59, 378–387.
Hess, S. C., Jeffrey, J. J., Ball, D. L., and Babich, L. (2006). Efficacy of feral pig removals at Hakalau forest national wildlife refuge. Hawai’i Trans. Western Sec. Wildlife Soc. 42, 53–67.
Hoefsloot, W., Van Ingen, J., Andrejak, C., Angeby, K., Bauriaud, R., Bemer, P., et al. (2013). The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples: an NTM-NET collaborative study. Eur. Respir. J. 42, 1604–1613.
Honda, J. R., Bernhard, J. N., and Chan, E. D. (2015). Natural disasters and nontuberculous mycobacteria: a recipe for increased disease? Chest 147, 304–308.
Honda, J. R., Hasan, N. A., Davidson, R. M., Williams, M. D., Epperson, L. E., Reynolds, P. R., et al. (2016). Environmental nontuberculous mycobacteria in the hawaiian islands. PLoS Negl. Trop. Dis. 10:e0005068. doi: 10.1371/journal.pntd.0005068
Huang, H. L., Cheng, M. H., Lu, P. L., Shu, C. C., Wang, J. Y., Wang, J. T., et al. (2017). Epidemiology and predictors of NTM pulmonary infection in taiwan – A retrospective, five-year multicenter study. Sci. Rep. 7:16300.
Ichijo, T., Izumi, Y., Nakamoto, S., Yamaguchi, N., and Nasu, M. (2014). Distribution and respiratory activity of mycobacteria in household water system of healthy volunteers in Japan. PLoS One 9:e110554. doi: 10.1371/journal.pone.0110554
Ichiyama, S., Shimokata, K., and Tsukamura, M. (1988). The isolation of Mycobacterium avium complex from soil, water, and dusts. Microbiol. Immunol. 32, 733–739.
Iwamoto, T., Nakajima, C., Nishiuchi, Y., Kato, T., Yoshida, S., Nakanishi, N., et al. (2012). Genetic diversity of Mycobacterium avium subsp. hominissuis strains isolated from humans, pigs, and human living environment. Infect. Genet. Evol. 12, 846–852.
Johnson, M. M., and Odell, J. A. (2014). Nontuberculous mycobacterial pulmonary infections. J. Thorac. Dis. 6, 210–220.
Kaevska, M., Slana, I., Kralik, P., Reischl, U., Orosova, J., Holcikova, A., et al. (2011). ”Mycobacterium avium subsp. hominissuis” in neck lymph nodes of children and their environment examined by culture and triplex quantitative real-time PCR. J. Clin. Microbiol. 49, 167–172.
Kankya, C., Muwonge, A., Djonne, B., Munyeme, M., Opuda-Asibo, J., Skjerve, E., et al. (2011). Isolation of Non-tuberculous mycobacteria from pastoral ecosystems of Uganda: public health significance. BMC Public Health 11:320. doi: 10.1186/1471-2458-11-320
Kawamura, Y., Li, Y., Liu, H., Huang, X., Li, Z., and Ezaki, T. (2001). Bacterial population in Russian space station ”Mir”. Microbiol. Immunol. 45, 819–828.
Khaledi, A., Bahador, A., Esmaeili, D., Tafazoli, A., Ghazvini, K., and Mansury, D. (2016). Prevalence of nontuberculous mycobacteria isolated from environmental samples in Iran: a meta-analysis. J. Res. Med. Sci. 21:58.
Kirs, M., Moravcik, P., Gyawali, P., Hamilton, K., Kisand, V., Gurr, I., et al. (2017). Rainwater harvesting in american samoa: current practices and indicative health risks. Environ. Sci. Pollut. Res. Int. 24, 12384–12392.
Kirschner, R. A. Jr., Parker, B. C., and Falkinham, J. O. III. (1992). Epidemiology of infection by nontuberculous mycobacteria. Mycobacterium avium, Mycobacterium intracellulare, and Mycobacterium scrofulaceum in acid, brown-water swamps of the southeastern United States and their association with environmental variables. Am. Rev. Respir. Dis. 145, 271–275.
Koh, E., Iizasa, T., Yamaji, H., Sekine, Y., Hiroshima, K., Yoshino, I., et al. (2012). Significance of the correlation between the expression of interleukin 6 and clinical features in patients with non-small cell lung cancer. Int. J. Surg. Pathol. 20, 233–239.
Koh, W. J., Jeong, B. H., Jeon, K., Lee, N. Y., Lee, K. S., Woo, S. Y., et al. (2012). Clinical significance of the differentiation between Mycobacterium avium and Mycobacterium intracellulare in M. avium complex lung disease. Chest 142, 1482–1488.
Koschel, D., Pietrzyk, C., Sennekamp, J., and Muller-Wening, D. (2006). [Swimming pool lung – Extrinsic allergic alveolitis or mycobacterial disease?]. Pneumologie 60, 285–289.
Kuehl, R., Banderet, F., Egli, A., Keller, P. M., Frei, R., Dobele, T., et al. (2018). Different types of heater-cooler units and their risk of transmission of Mycobacterium chimaera during open-heart surgery: clues from device design. Infect. Control. Hosp. Epidemiol. 39, 834–840.
Lahiri, A., Kneisel, J., Kloster, I., Kamal, E., and Lewin, A. (2014). Abundance of Mycobacterium avium ssp. hominissuis in soil and dust in Germany – Implications for the infection route. Lett. Appl. Microbiol. 59, 65–70.
Lai, C. C., Tan, C. K., Chou, C. H., Hsu, H. L., Liao, C. H., Huang, Y. T., et al. (2010). Increasing incidence of nontuberculous mycobacteria, Taiwan, 2000-2008. Emerg. Infect. Dis. 16, 294–296.
Lee, M. R., Sheng, W. H., Hung, C. C., Yu, C. J., Lee, L. N., and Hsueh, P. R. (2015). Mycobacterium abscessus complex infections in humans. Emerg. Infect. Dis. 21, 1638–1646.
Lescenko, P., Matlova, L., Dvorska, L., Bartos, M., Vavra, O., Navratil, L., et al. (2003). Mycobacterial infection in aquarium fish. Vet. Med. – Czech 48, 71–78.
Lin, C., Russell, C., Soll, B., Chow, D., Bamrah, S., Brostrom, R., et al. (2018). Increasing prevalence of nontuberculous mycobacteria in respiratory specimens from US-Affiliated Pacific island jurisdictions(1). Emerg. Infect. Dis. 24, 485–491.
Lipner, E. M., Knox, D., French, J., Rudman, J., Strong, M., and Crooks, J. L. (2017). A geospatial epidemiologic analysis of nontuberculous mycobacterial infection: an ecological study in colorado. Ann. Am. Thorac. Soc. 14, 1523–1532.
Lorencova, A., Klanicova, B., Makovcova, J., Slana, I., Vojkovska, H., Babak, V., et al. (2013). Nontuberculous mycobacteria in freshwater fish and fish products intended for human consumption. Foodborne Pathog. Dis. 10, 573–576.
Lorencova, A., Vasickova, P., Makovcova, J., and Slana, I. (2014). Presence of Mycobacterium avium subspecies and hepatitis E virus in raw meat products. J. Food Prot. 77, 335–338.
Makovcova, J., Babak, V., Slany, M., and Slana, I. (2015). Comparison of methods for the isolation of mycobacteria from water treatment plant sludge. Antonie Van Leeuwenhoek 107, 1165–1179.
Malcolm, K. C., Caceres, S. M., Honda, J. R., Davidson, R. M., Epperson, L. E., Strong, M., et al. (2017). Mycobacterium abscessus displays fitness for fomite transmission. Appl. Environ. Microbiol. 83:e00562-17.
Marra, A. R., Diekema, D. J., and Edmond, M. B. (2017). Mycobacterium chimaera infections associated with contaminated heater-cooler devices for cardiac surgery: outbreak management. Clin. Infect. Dis. 65, 669–674.
Martin, E. C., Parker, B. C., and Falkinham, J. O. III (1987). Epidemiology of infection by nontuberculous mycobacteria. VII. Absence of mycobacteria in southeastern groundwaters. Am. Rev. Respir. Dis. 136, 344–348.
Meissner, G., and Anz, W. (1977). Sources of Mycobacterium avium complex infection resulting in human diseases. Am. Rev. Respir. Dis. 116, 1057–1064.
Mirsaeidi, M., Machado, R. F., Garcia, J. G., and Schraufnagel, D. E. (2014). Nontuberculous mycobacterial disease mortality in the United States, 1999-2010: a population-based comparative study. PLoS One 9:e91879. doi: 10.1371/journal.pone.0091879
Monde, N., Munyeme, M., Muwonge, A., Muma, J. B., and Malama, S. (2018). Characterization of Non-tuberculous Mycobacterium from humans and water in an Agropastoral area in Zambia. BMC Infect. Dis. 18:20. doi: 10.1186/s12879-017-2939-y
Morita, Y., Maruyama, S., Hashizaki, F., and Katsube, Y. (1999). Pathogenicity of Mycobacterium avium complex serovar 9 isolated from painted quail (Excalfactoria chinensis). J. Vet. Med. Sci. 61, 1309–1312.
Mudedla, S., Avendano, E. E., and Raman, G. (2015). Non-tuberculous Mycobacterium skin infections after tattooing in healthy individuals: a systematic review of case reports. Dermatol. Online J. 21:13030/qt8mr3r4f0.
Neonakis, I. K., Gitti, Z., Kourbeti, I. S., Michelaki, H., Baritaki, M., Alevraki, G., et al. (2007). Mycobacterial species diversity at a general hospital on the island of Crete: first detection of Mycobacterium lentiflavum in Greece. Scand. J. Infect. Dis. 39, 875–879.
Nishiuchi, Y., Iwamoto, T., and Maruyama, F. (2017). Infection sources of a common Non-tuberculous mycobacterial pathogen, Mycobacterium avium complex. Front. Med. (Lausanne) 4:27. doi: 10.3389/fmed.2017.00027
Nishiuchi, Y., Tamura, A., Kitada, S., Taguri, T., Matsumoto, S., Tateishi, Y., et al. (2009). Mycobacterium avium complex organisms predominantly colonize in the bathtub inlets of patients’ bathrooms. Jpn. J. Infect. Dis. 62, 182–186.
Ovrutsky, A. R., Chan, E. D., Kartalija, M., Bai, X., Jackson, M., Gibbs, S., et al. (2013). Cooccurrence of free-living amoebae and nontuberculous mycobacteria in hospital water networks, and preferential growth of Mycobacterium avium in Acanthamoeba lenticulata. Appl. Environ. Microbiol. 79,3185–3192.
Pai, H. H., Chen, W. C., and Peng, C. F. (2003). Isolation of Non-tuberculous mycobacteria from hospital cockroaches (Periplaneta americana). J. Hosp. Infect. 53, 224–228.
Perez-Martinez, I., Aguilar-Ayala, D. A., Fernandez-Rendon, E., Carrillo-Sanchez, A. K., Helguera-Repetto, A. C., Rivera-Gutierrez, S., et al. (2013). Occurrence of potentially pathogenic nontuberculous mycobacteria in Mexican household potable water: a pilot study. BMC Res. Notes 6:531. doi: 10.1186/1756-0500-6-531
Piersimoni, C., and Scarparo, C. (2009). Extrapulmonary infections associated with nontuberculous mycobacteria in immunocompetent persons. Emerg. Infect. Dis. 15, 1351–1358; quiz 1544.
Reisfeld, L., Ikuta, C. Y., Ippolito, L., Silvatti, B., Ferreira Neto, J. S., Catao-Dias, J. L., et al. (2018). Cutaneous mycobacteriosis in a captive Amazonian manatee Trichechus inunguis. Dis. Aquat. Organ. 127, 231–236.
Richards, C. L., Broadaway, S. C., Eggers, M. J., Doyle, J., Pyle, B. H., Camper, A. K., et al. (2015). Detection of pathogenic and non-pathogenic bacteria in drinking water and associated biofilms on the crow reservation, Montana, USA. Microb. Ecol. 76, 52–63.
Rickman, O. B., Ryu, J. H., Fidler, M. E., and Kalra, S. (2002). Hypersensitivity pneumonitis associated with Mycobacterium avium complex and hot tub use. Mayo Clin. Proc. 77, 1233–1237.
Rodgers, M. R., Blackstone, B. J., Reyes, A. L., and Covert, T. C. (1999). Colonisation of point of use water filters by silver resistant Non-tuberculous mycobacteria. J. Clin. Pathol. 52:629.
Roguet, A., Therial, C., Saad, M., Boudahmane, L., Moulin, L., and Lucas, F. S. (2016). High mycobacterial diversity in recreational lakes. Antonie Van Leeuwenhoek 109, 619–631.
Rose, C. S., Martyny, J. W., Newman, L. S., Milton, D. K., King, T. E. Jr., et al. (1998). ”Lifeguard lung”: endemic granulomatous pneumonitis in an indoor swimming pool. Am. J. Public Health 88, 1795–1800.
Rudis, B. (2018). albersusa: Tools, Shapefiles and Data to Work with an ‘AlbersUSA’ Composite Projection. R package version 0.3.1. Avialable at: https://github.com/hrbrmstr/albersusa
Safranek, T. J., Jarvis, W. R., Carson, L. A., Cusick, L. B., Bland, L. A., Swenson, J. M., et al. (1987). Mycobacterium chelonae wound infections after plastic surgery employing contaminated gentian violet skin-marking solution. N. Engl. J. Med. 317, 197–201.
Sax, H., Bloemberg, G., Hasse, B., Sommerstein, R., Kohler, P., Achermann, Y., et al. (2015). Prolonged outbreak of Mycobacterium chimaera infection after open-chest heart surgery. Clin. Infect Dis. 61, 67–75.
Schinsky, M. F., Morey, R. E., Steigerwalt, A. G., Douglas, M. P., Wilson, R. W., Floyd, M. M., et al. (2004). Taxonomic variation in the Mycobacterium fortuitum third biovariant complex: description of Mycobacterium boenickei sp. nov. Mycobacterium houstonense sp. nov., Mycobacterium neworleansense sp. nov. and Mycobacterium brisbanense sp. nov. and recognition of Mycobacterium porcinum from human clinical isolates. Int. J. Syst. Evol. Microbiol. 54,1653–1667.
Schreiber, P. W., Kuster, S. P., Hasse, B., Bayard, C., Ruegg, C., Kohler, P., et al. (2016). Reemergence of Mycobacterium chimaera in heater-cooler units despite intensified cleaning and disinfection protocol. Emerg. Infect. Dis. 22, 1830–1833.
Schulze-Robbecke, R., Feldmann, C., Fischeder, R., Janning, B., Exner, M., and Wahl, G. (1995). Dental units: an environmental study of sources of potentially pathogenic mycobacteria. Tuber Lung. Dis. 76, 318–323.
Scriven, J. E., Scobie, A., Verlander, N. Q., Houston, A., Collyns, T., Cajic, V., et al. (2018). Mycobacterium chimaera infection following cardiac surgery in the United Kingdom: clinical features and outcome of the first 30 cases. Clin. Microbiol. Infect. [Epub ahead of print].
Simons, S., Van Ingen, J., Hsueh, P. R., Van Hung, N., Dekhuijzen, P. N., Boeree, M. J., et al. (2011). Nontuberculous mycobacteria in respiratory tract infections, eastern Asia. Emerg. Infect. Dis. 17, 343–349.
Stout, J. E., Koh, W. J., and Yew, W. W. (2016). Update on pulmonary disease due to Non-tuberculous mycobacteria. Int. J. Infect. Dis. 45, 123–134.
Strollo, S. E., Adjemian, J., Adjemian, M. K., and Prevots, D. R. (2015). The burden of pulmonary nontuberculous mycobacterial disease in the United States. Ann. Am. Thorac. Soc. 12, 1458–1464.
Sugita, Y., Ishii, N., Katsuno, M., Yamada, R., and Nakajima, H. (2000). Familial cluster of cutaneous Mycobacterium avium infection resulting from use of a circulating, constantly heated bath water system. Br. J. Dermatol. 142, 789–793.
Talaat, A. M., Trucksis, M., Kane, A. S., and Reimschuessel, R. (1999). Pathogenicity of Mycobacterium fortuitum and Mycobacterium smegmatis to goldfish, Carassius auratus. Vet. Microbiol. 66, 151–164.
Taylor, M., Ross, K., and Bentham, R. (2009). Legionella, protozoa, and biofilms: interactions within complex microbial systems. Microb. Ecol. 58,538–547.
Thomson, R., Donnan, E., and Konstantinos, A. (2017). Notification of nontuberculous mycobacteria: an australian perspective. Ann. Am. Thorac. Soc. 14, 318–323.
Thomson, R., Tolson, C., Carter, R., Coulter, C., Huygens, F., and Hargreaves, M. (2013). Isolation of nontuberculous mycobacteria (NTM) from household water and shower aerosols in patients with pulmonary disease caused by NTM. J. Clin. Microbiol. 51, 3006–3011.
Thomson, R. M. (2010). Changing epidemiology of pulmonary nontuberculous mycobacteria infections. Emerg. Infect. Dis. 16, 1576–1583.
Thomson, R. M., Carter, R., Tolson, C., Coulter, C., Huygens, F., and Hargreaves, M. (2013). Factors associated with the isolation of nontuberculous mycobacteria (NTM) from a large municipal water system in Brisbane, Australia. BMC Microbiol. 13:89. doi: 10.1186/1471-2180-13-89
Thorel, M. F., Falkinham, J. O. III, and Moreau, R. G. (2004). Environmental mycobacteria from alpine and subalpine habitats. FEMS Microbiol. Ecol. 49, 343–347.
Tichenor, W. S., Thurlow, J., Mcnulty, S., Brown-Elliott, B. A., Wallace, R. J. Jr., and Falkinham, J. O. III. (2012). Nontuberculous mycobacteria in household plumbing as possible cause of chronic rhinosinusitis. Emerg. Infect. Dis. 18, 1612–1617.
Ulmann, V., Kracalikova, A., and Dziedzinska, R. (2015). Mycobacteria in water used for personal hygiene in heavy industry and collieries: a potential risk for employees. Int. J. Environ. Res. Public Health 12, 2870–2877.
Velayati, A. A., Farnia, P., Mozafari, M., Malekshahian, D., Seif, S., Rahideh, S., et al. (2014). Molecular epidemiology of nontuberculous mycobacteria isolates from clinical and environmental sources of a metropolitan city. PLoS One 9:e114428. doi: 10.1371/journal.pone.0114428
Wallace, R. J. Jr., Brown-Elliott, B. A., Wilson, R. W., Mann, L., Hall, L., Zhang, Y., et al. (2004). Clinical and laboratory features of Mycobacterium porcinum. J. Clin. Microbiol. 42, 5689–5697.
Wallace, R. J. Jr., Iakhiaeva, E., Williams, M. D., Brown-Elliott, B. A., Vasireddy, S., et al. (2013). Absence of Mycobacterium intracellulare and the presence of Mycobacterium chimaera in household water and biofilm samples of patients in the U.S. With Mycobacterium avium complex respiratory disease. J. Clin. Microbiol. 51, 1747–1752.
Wang, S. H., Pancholi, P., Stevenson, K., Yakrus, M. A., Butler, W. R., Schlesinger, L. S., et al. (2009). Pseudo-outbreak of ”Mycobacterium paraffinicum” infection and/or colonization in a tertiary care medical center. Infect. Control Hosp. Epidemiol. 30, 848–853.
Wassilew, N., Hoffmann, H., Andrejak, C., and Lange, C. (2016). Pulmonary disease caused by Non-tuberculous mycobacteria. Respiration 91,386–402.
Whiley, H., Keegan, A., Giglio, S., and Bentham, R. (2012). Mycobacterium avium complex–the role of potable water in disease transmission. J. Appl. Microbiol. 113, 223–232.
Wilson, R. W., Steingrube, V. A., Bottger, E. C., Springer, B., Brown-Elliott, B. A., Vincent, V., et al. (2001). Mycobacterium immunogenum sp. nov., a novel species related to Mycobacterium abscessus and associated with clinical disease, pseudo-outbreaks and contaminated metalworking fluids: an international cooperative study on mycobacterial taxonomy. Int. J. Syst. Evol. Microbiol. 51, 1751–1764.
Winthrop, K. L., Albridge, K., South, D., Albrecht, P., Abrams, M., Samuel, M. C., et al. (2004). The clinical management and outcome of nail salon-acquired Mycobacterium fortuitum skin infection. Clin. Infect. Dis. 38,38–44.
Winthrop, K. L., Mcnelley, E., Kendall, B., Marshall-Olson, A., Morris, C., Cassidy, M., et al. (2010). Pulmonary nontuberculous mycobacterial disease prevalence and clinical features: an emerging public health disease. Am. J. Respir. Crit. Care Med. 182, 977–982.
Yajko, D. M., Chin, D. P., Gonzalez, P. C., Nassos, P. S., Hopewell, P. C., Reingold, A. L., et al. (1995). Mycobacterium avium complex in water, food, and soil samples collected from the environment of HIV-infected individuals. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 9, 176–182.
Yajko, D. M., Nassos, P. S., Sanders, C. A., Gonzalez, P. C., Reingold, A. L., Horsburgh, C. R., et al. (1993). Comparison of four decontamination methods for recovery of Mycobacterium avium complex from stools. J. Clin. Microbiol. 31, 302–306.
Yoder, S., Argueta, C., Holtzman, A., Aronson, T., Berlin, O. G., Tomasek, P., et al. (1999). PCR comparison of Mycobacterium avium isolates obtained from patients and foods. Appl. Environ. Microbiol. 65, 2650–2653.
Keywords: nontuberculous mycobacteria, environments, man-made, natural, Hawaii
Citation: Honda JR, Virdi R and Chan ED (2018) Global Environmental Nontuberculous Mycobacteria and Their Contemporaneous Man-Made and Natural Niches. Front. Microbiol. 9:2029. doi: 10.3389/fmicb.2018.02029
Received: 26 June 2018; Accepted: 10 August 2018;
Published: 30 August 2018.
Edited by:
Thomas Dick, Rutgers, The State University of New Jersey, Newark, United StatesReviewed by:
Joseph Oliver Falkinham, Virginia Tech, United StatesCopyright © 2018 Honda, Virdi and Chan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jennifer R. Honda, aG9uZGFqQG5qaGVhbHRoLm9yZw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.