
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 15 August 2018
Sec. Microbial Immunology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.01890
This article is part of the Research Topic Immune Evasion Mechanisms in the Pathogenesis of Trypanosomatid Infections View all 50 articles
 Beatriz R. S. Dias1Carina S. de Souza1Niara de Jesus Almeida1José G. B. Lima1
Beatriz R. S. Dias1Carina S. de Souza1Niara de Jesus Almeida1José G. B. Lima1 Kiyoshi F. Fukutani2Thiale B. S. dos Santos2
Kiyoshi F. Fukutani2Thiale B. S. dos Santos2 Jaqueline França-Cost2,3
Jaqueline França-Cost2,3 Claudia I. Brodskyn1Juliana P. B. de Menezes1
Claudia I. Brodskyn1Juliana P. B. de Menezes1 Maria I. Colombo4
Maria I. Colombo4 Patricia S. T. Veras1*
Patricia S. T. Veras1*CBA mouse macrophages control Leishmania major infection yet are permissive to Leishmania amazonensis. Few studies have been conducted to assess the role played by autophagy in Leishmania infection. Therefore, we assessed whether the autophagic response of infected macrophages may account for the differential behavior of these two parasite strains. After 24 h of infection, the LC3-II/Act ratio increased in both L. amazonensis- and L. major-infected macrophages compared to uninfected controls, but less than in chloroquine-treated cells. This suggests that L. amazonensis and L. major activate autophagy in infected macrophages, without altering the autophagic flux. Furthermore, L. major-infected cells exhibited higher percentages of DQ-BSA-labeled parasitophorous vacuoles (50%) than those infected by L. amazonensis (25%). However, L. major- and L. amazonensis-induced parasitophorous vacuoles accumulated LysoTracker similarly, indicating that the acidity in both compartment was equivalent. At as early as 30 min, endogenous LC3 was recruited to both L. amazonensis- and L. major-induced parasitophorous vacuoles, while after 24 h a greater percentage of LC3 positive vacuoles was observed in L. amazonensis-infected cells (42.36%) compared to those infected by L. major (18.10%). Noteworthy, principal component analysis (PCA) and an hierarchical cluster analysis completely discriminated L. major-infected macrophages from L. amazonensis-infected cells accordingly to infection intensity and autophagic features of parasite-induced vacuoles. Then, we evaluated whether the modulation of autophagy exerted an influence on parasite infection in macrophages. No significant changes were observed in both infection rate or parasite load in macrophages treated with the autophagic inhibitors wortmannin, chloroquine or VPS34-IN1, as well as with the autophagic inducers rapamycin or physiological starvation, in comparison to untreated control cells. Interestingly, both autophagic inducers enhanced intracellular L. amazonensis and L. major viability, while the pharmacological inhibition of autophagy exerted no effects on intracellular parasite viability. We also demonstrated that autophagy induction reduced NO production by L. amazonensis- and L. major-infected macrophages but not alters arginase activity. These findings provide evidence that although L. amazonensis-induced parasitophorous vacuoles recruit LC3 more markedly, L. amazonensis and L. major similarly activate the autophagic pathway in CBA macrophages. Interestingly, the exogenous induction of autophagy favors L. major intracellular viability to a greater extent than L. amazonensis related to a reduction in the levels of NO.
Leishmaniasis, a disease caused by trypanosomatids of the genus Leishmania, represents an emerging disease that continues to present a major public health problem, especially due to elevated incidence in developing countries (Santos et al., 2008; Perinoto et al., 2010). According to the World Health Organization, around 15 million people are affected worldwide in 98 countries, including 72 developing nations (World Health Organization, 2010; Alvar et al., 2012). Leishmania spp. are obligate intracellular parasites that live and multiply within modified phagolysosome compartments, denominated parasitophorous vacuoles, in macrophages, the main host cell in vertebrate hosts (Russell and Wilhelm, 1986).
Since the early 2000s, many fields of study have focused research efforts on the physiological process of autophagy. This evolutionarily conserved process is responsible for the degradation of organelles and proteins during cell differentiation and under stress conditions (Meijer and Codogno, 2004; Levine and Deretic, 2007; Duszenko et al., 2011). During the autophagic degradative process, cytosolic components become trapped into compartments called autophagosomes, which subsequently fuse with lysosomes to form autolysosomes in consecutively coordinated steps: initiation, nucleation, expansion, completion, docking and fusion, thereby resulting in intravacuolar cargo degradation. These events are dependent on the family of autophagy related genes (Atg) and proteins that play specific roles during each stage of the autophagic process (Suzuki and Ohsumi, 2007; Suzuki et al., 2010; Nazarko et al., 2011). The initiation step results in phagophore formation, which is mostly dependent on the assembling of the Beclin 1-Vps34 class III PI(3)K complex in autophagosomal membranes of differing origin (Levine and Deretic, 2007). Two conjugation systems can be subsequently recruited to support the expansion of the phagophore: the Atg12-Atg5-Atg16 complex, or the Atg8 protein conjugated to phosphoethalanolamine (PE), which is referred as LC3-II in mammals. This protein, central in the autophagic process, is generated by site-specific proteolysis and lipidation occurring near the C-terminus of the cytosolic protein Atg8 (LC3-I) (Lang et al., 1998). As LC3-II is integrated into the membranes of newly formed autophagosomes, levels of LC3-II correlate positively with numbers of cellular autophagosomes (Kabeya et al., 2000). Thus, LC3-II has been employed as a useful marker for studies investigating the dynamics of autophagic pathway activation.
In recent decades, the role played by autophagy in mammalian cells during infection arising from a variety of pathogens has been extensively investigated (Gutierrez et al., 2004; Levine and Deretic, 2007; Schnaith et al., 2007; Mestre et al., 2010). Several studies have implicated autophagic pathway activation in the triggering of an innate immune response, which results in the protection of host cells against infections arising from intracellular microorganisms, such as Mycobacterium tuberculosis and Streptococcus pyogenes (Gutierrez et al., 2004; Nakagawa et al., 2004; Gutierrez et al., 2005; Levine and Deretic, 2007). On the other hand, autophagic pathway activation can also favor the survival of some species of intracellular pathogens, such as Staphylococcus aureus and Coxiella burnetti (Gutierrez et al., 2005; Schnaith et al., 2007; Mestre et al., 2010).
To date, few studies have attempted to assess the role of autophagy in infection by parasites of the Leishmania genus. A seminal work by Schaible and colleagues demonstrated that large L. mexicana-induced parasitophorous vacuoles acquired macromolecules from the host cell cytoplasm via microautophagy (Schaible et al., 1999). These authors suggested that these macromolecules, once transferred from the host cell cytoplasm to the lumen of L. mexicana-induced parasitophorous vacuoles, seemed to be endocytosed by parasites (Russell et al., 1992). More recently, it has been demonstrated that both L. amazonensis and L. major infection induces autophagic pathway activation in susceptible BALB/c mouse macrophages (Cyrino et al., 2012; Frank et al., 2015). Investigations in the literature regarding the role played by autophagy in Leishmania infection have led to controversial data. Pinheiro et al. (2009) showed that in macrophages from susceptible BALB/c mice, but not in macrophages of L. amazonensis-resistant C57BL/6 mice, the autophagic induction resulted in increased intracellular load of L. amazonensis (Pinheiro et al., 2009). Also, these authors demonstrated that autophagy induced by starvation did not alter intracellular L. major parasitic load in susceptible BALB/c mouse macrophages (Pinheiro et al., 2009). Studies employing the genetic modification of autophagic-related genes have also presented inconclusive results. Thomas et al. (2017) demonstrated that Atg5 and Atg9 knockdown in the human monocytic cell-line, THP-1, reduced L. donovani survival, suggesting that autophagy is beneficial to Leishmania infection. This stands in contrast to BALB/c macrophages knocked-down for Atg5, which served to enhance L. major parasitic load (Frank et al., 2015). Consistent with these authors’ findings, another study recently showed that Atg5 knockdown in the L. major-resistant C57BL/6 macrophages resulted in increased parasite replication (Franco et al., 2017), which suggests that the activation of an Atg5-dependent autophagic process could result in the elimination of L. major intracellular parasites.
The CBA mouse model has demonstrated susceptibility to L. amazonensis, while controlling L. major infection in vivo (Lemos de Souza et al., 2000). Moreover, while CBA mouse macrophages were shown to be permissive to L. amazonensis in vitro, these same cells also reduced L. major infection (Gomes et al., 2003). Therefore, we addressed whether the autophagic response of infected macrophages may account for the differential behavior of these two parasite strains in CBA macrophages. Accordingly, the present study took advantage of the in vitro CBA infection model to comparatively characterize L. major and L. amazonensis-induced compartments with respect to autophagic features, in addition to investigating the effects of autophagic modulation on Leishmania infection outcome using a pharmacological approach.
The CBA mice used in the present study were provided by the animal care facility at the Gonçalo Moniz Institute - FIOCRUZ/BA, following approval by the Institutional Animal Experimentation Review Board (CEUA) under protocol number 015/2014. Animals were kept and handled in accordance with the norms recommended by the International Guiding Principles for Biomedical Research Involving Animals; all experimental protocols complied with these guidelines, as well as all resolutions established by the Brazilian National Council for the Control of Animal Experimentation (CONCEA).
All protocols, analytic methods and material used in the present study are available upon request to all interested researchers.
To obtain inflammatory peritoneal macrophages, 2.5 mL of 3% thioglycolate (Sigma, St. Louis, MO, United States) was injected into the peritoneal cavities of CBA mice, as previously described by Gomes et al. (2003). After 96 h, mice were euthanized, and peritoneal cavities were washed twice using 10 mL of 0.9% NaCl with heparin (20 U.I./mL) (Cristália, Itapira, SP, BR). Next, macrophages were centrifuged at 300 × g under 4°C for 10 min and plated in complete DMEM (Dulbecco’s modified Eagle medium) (Gibco, Grand Island, NY, United States) supplemented with 25 mM HEPES (N-2-hydroxyethyl piperazine-N’-2-ethane-sulfonic acid) (Sigma, St Louis, MO, United States) adjusted to pH 7.4, 2 mM glutamin (Gibco, Grand Island, NY, United States), 20 g/mL ciprofloxacin (Isofarma, Precabura, CE, BR) and 10% inactivated fetal bovine serum (Gibco, Grand Island, NY, United States), then incubated overnight at 37°C under 5% CO2 and 95% humidity.
Axenic cultures of L. amazonensis (MHOM/Br88/Ba-125) or L. major (MHOM/RI/-/WR-173) promastigotes were maintained for up to six successive passages in Schneider’s Insect Medium (Sigma, St Louis, MO, United States) supplemented with 50 μg/mL gentamicin (Gibco, Grand Island, NY, United States) and 10% fetal bovine serum (Gibco, Grand Island, NY, United States), following a slightly modified previously described protocol (Gomes et al., 2003). Promastigotes were grown in an incubator at 24°C and monitored daily by counting in a Neubauer chamber. Upon reaching the stationary phase, the promastigotes were subjected to separation of infective metacyclic forms using a Ficoll-Paque gradient as described by Spath and Beverley (2001).
For some experiments, L. amazonensis and L. major were labeled with CFSE (21888) (Sigma, St Louis, MO, United States) as described by Chang et al. (2007). Parasites (108) were suspended in 2 mL of 0.9% NaCl solution containing 1 μM CFSE for 10 min at 37°C, protected from light. To stop the reaction, a similar volume (2 mL) of fetal bovine serum was added to each parasite suspension. After 1 min of incubation, parasite suspensions were washed twice with 0.9% NaCl solution and centrifuged at 1,800 × g for 10 min under 4°C. After counting with a Neubauer chamber, parasites were used at the experimental concentrations described below.
CBA mouse macrophages were plated in either 6-well plates (2 × 106/2 mL) or 24-well plates (2 × 105/mL) containing coverslips for 24 h at 37°C, and then infected with metacyclic promastigotes of L. amazonensis or L. major at a 5:1 (parasite:macrophage) ratio in accordance with one of the following protocols:
(i) macrophages were either infected for 30 min, or infected for 4 h followed by a 24 h-incubation period to evaluate autophagic activation and flux, as described below in Evaluation of autophagic induction and flux in macrophages infected by L. amazonensis or L. major;
(ii) macrophages were either infected for 30 min or 4 h to characterize the autophagic features within Leishmania parasitophorous vacuoles at early stages of infection. All macrophages were then washed and fixed. Alternatively, to compare L. amazonensis and L. major parasitophorous vacuole characteristics at later stages of infection, another group of macrophages was infected for 4 h and then washed with saline to remove any non-internalized promastigotes. After an additional 24 h-incubation period, these macrophages were then washed and fixed. Autophagic features were then evaluated at both early and late stages of infection as described below in Evaluation of hydrolytic activity in L. amazonensis- or L. major-induced parasitophorous vacuoles; LysoTracker labeling of L. amazonensis or L. major parasitophorous vacuoles and Evaluation of the recruitment of LC3-II to L. amazonensis or L. major parasitophorous vacuoles;
(iii) macrophages were first infected for 30 min, treated with different autophagic modulators for an additional 4 h, then washed and reincubated in modulator-free medium as described below in Modulation of the autophagic process in L. amazonensis or L. major infection. Following each infection protocol, all plates were washed to remove non-internalized parasites. Finally, all plates containing cells infected with L. amazonensis were reincubated at 35°C, while those infected with L. major were incubated at 37°C, both supplemented with 5% CO2 and 95% humidity.
For these experiments, western blotting was used to measure the ratio between the amount of membrane bound LC3-II and actin (Act) in extracts of CBA mouse macrophages. Cells were plated and infected in accordance with the protocol (i) described above in Infection of macrophages with L. amazonensis or L. major. To evaluate autophagic flux, either uninfected or cells infected with L. amazonensis or L. major were treated with chloroquine (10 μM), a drug that blocks the fusion of autophagosomes with lysosomes (Klionsky et al., 2016), for an additional 4 h. Upon the completion of infection or treatment protocols, cells were scraped, recovered and centrifuged at 720 × g for 10 min under 4°C. Cells were then resuspended in RIPA buffer [1 M Tris HCl pH 8, 1 M NaCl, Substitute Nonidet P40 (Sigma, St Louis, MO, United States), 10% Sodium Dodecyl Sulfate (Riedel-de-Haen, Seelge, GER), 5% DOC, Mili-Q Water and Protease Inhibitor (Roche)] and incubated for 30 min at 4°C. Next, cellular extracts were sonicated 3x for 10 s at 127V in a Branson Sonifier apparatus. Samples were then centrifuged at 13,000 × g under 4°C for 20 min and quantified using a Nanodrop spectrophotometer (Thermo Scientific).
Initially, 60 μg of protein from each extract was subjected to electrophoresis in a Bio-Rad Mini-Protean Tetra System. After 1 h 30 min of gel running, proteins were transferred to a nitrocellulose membrane using a Bio-RAD Trans-blot Turbo apparatus for a period of 30 min at 25 V, 1A. The membrane was then blocked with 5% skim milk in PBST (1x PBS + 0.1% Tween 20) for 1 h at room temperature. Membranes were then incubated overnight at 4°C with anti-LC3 antibody (NB600-1384, Novus Biologicals, Littleton, CO, United States) or anti–actin (Sigma) diluted in PBST + 3% milk solution. Each membrane was subsequently incubated with HRP-coupled anti-rabbit IgG or anti-mouse IgG secondary antibodies. Blots were then developed with an ECL Chemiluminescence Kit (Thermo Fisher Scientific, Rockford, IL, United States) and detected using a Luminescent Image Analyzer and Image Quant Las 4000 software. Densitometry quantification of the bands was performed using Image J software.
Hydrolytic activity in the parasitophorous vacuoles within macrophages infected by L. amazonensis or L. major was evaluated using a degradative compartment marker, DQ-BSA (DQTM Red BSA; Molecular Probes, Eugene, OR, United States), as described by Lerena and Colombo (2011). CBA mouse macrophages were plated and, after 24 h, cells were washed and incubated with 10 μg/mL of DQ-BSA for 4 h at 37°C. Next, cells were rewashed and infected with metacyclic promastigotes of L. amazonensis or L. major labeled with CFSE (λex 492 nm; λem 517 nm) at a ratio of 5:1 (parasite:macrophage) in accordance with the infection protocol (ii) described above in Infection of macrophages with L. amazonensis or L. major. Following infection, cells infected for 4 h or more were incubated with 10 μg/mL rapamycin or 1 μM VPS34-IN1 (Bago et al., 2014), a specific PI3K class III inhibitor, for 2 h to evaluate the effect of autophagy modulation on DQ-BSA-labeled Leishmania parasitophorous vacuoles. Finally, the cells were fixed in 4% paraformaldehyde for 15 min and coverslips were mounted using a ProLong Gold Antifade kit containing DAPI (Life Technologies). Images were acquired via a Leica SP8 (Leica) confocal fluorescence microscope using a 63x/1.4 objective.
LysoTracker® Red DND-99 (Molecular Probes, Eugene, OR, United States), a weakly basic amine that selectively accumulates in cellular compartments with low internal pH, was used to characterize the parasitophorous vacuoles induced by L. amazonensis or L. major with respect to the acidity present in autophagosome compartments. Cells were labeled in accordance with a protocol previously described by Aguilera et al. (2009). CBA mouse macrophages were plated and infected as described above in the protocol (ii) of Infection of macrophages with L. amazonensis or L. major. After 24 h of infection, macrophages were incubated with 100 nM of LysoTracker® for 2 h. Cells were then fixed and coverslips were mounted as described in Evaluation of hydrolytic activity in L. amazonensis- or L. major-induced parasitophorous vacuoles. Images were acquired as described above.
Macrophages were infected as described above in the protocol (ii) of Infection of macrophages with L. amazonensis or L. major. After infection procedures, labeling for the autophagic membrane marker LC3 was performed in accordance with the protocol described by Matte et al. (2016) using antibodies against LC3B (1:200) (NB600-1384, Novus Biologicals, Littleton, CO, United States), followed by AlexaFluor 488-conjugated goat anti-rabbit IgG (1:500) (ThermoScientific). Following infection, cells infected for 4 h or more were incubated with 10 μg/mL rapamycin or 1 μM VPS34-IN1 for 2 h to evaluate the effect of autophagy modulation on LC3-labeled Leishmania parasitophorous vacuoles. Cells were then fixed and mounted on coverslips as described above in Evaluation of hydrolytic activity in L. amazonensis- or L. major-induced parasitophorous vacuoles. Images were acquired as described above.
The effects of autophagy on Leishmania infection were assessed by quantifying the percentage of infected cells and the number of parasites per infected cell using optical fluorescence, or by determining the number of viable intracellular parasites by microscopy. For these assays, macrophages were first infected as described above in protocol (iii) of Infection of macrophages with L. amazonensis or L. major.
To determine the percentage of infected cells and the number of parasites per infected cell, cells infected for 30 min were reincubated with either: (i) one of the following autophagic inhibitors: 10 μM chloroquine (Sigma, St Louis, MO, United States), 100 nM wortmannin (Sigma, St Louis, MO, United States) or 1 μM VPS34-IN1; (ii) or one of the following autophagic inducers: 10 μg/mL rapamycin (Sigma, St. Louis, MO, United States), or nutrient-poor EBSS medium (Sigma, St. Louis, MO, United States). After 4 h, all cells were washed and reincubated in complete DMEM for an additional 24 h. Cells were then subsequently fixed in 4% paraformaldehyde for 15 min and coverslips were mounted using a ProLong Gold Antifade kit with DAPI (Life Technologies). The percentage of infected cells and the number of parasites per infected macrophage were determined by counting no less than 400 cells in random fields under an Olympus BX 51 fluorescence microscope using an 100x/1.4 objective in accordance with a protocol previously described by Huynh et al. (2006).
Intracellular parasite viability was assessed by determining the number of viable intracellular amastigotes. After a 30-min infection period, cells were incubated with either one of the two autophagy inhibitors (10 μM chloroquine or 1 μM VPS34-IN1) or an autophagic inducer (10 μg/mL rapamycin) or by starvation, incubating cells in nutrient-poor medium (EBSS). After 4 h, all cells were washed and incubated in complete DMEM medium for 0, 6, or 12 h, followed by a 5-day period of reincubation in Schneider’s medium at 23°C. Finally, the viable intracellular amastigotes that had transformed into motile extracellular promastigotes were collected and counted in a Neubauer chamber.
As determined by AlamarBlue® assays, the autophagy modulators employed in the present study did not present direct effects on axenic Leishmania viability at the concentrations employed in this study (data not shown).
To determine whether autophagic modulation interferes with NO production by infected macrophages and/or arginase activity, cells were primed with 50 UI/mL IFN-γ (R&D Systems, Minneapolis, MN, United States) for 24 h and subsequently infected as described above in protocol (iii) of Infection of macrophages with L. amazonensis or L. major. Next, macrophages were treated with 1 μM VPS34-IN1 or 10 μg/mL rapamycin. After 4 h, all cells were washed and reincubated in DMEM medium containing IFN-γ for an additional 24 h. Infected cells incubated in IFN-γ-free medium were used as a negative control. NO production was measured in culture supernatants by determining the accumulation of nitrite using the Griess reaction. Arginase activity was quantified by a colorimetric assay for the detection of urea as previously described (Abebe et al., 2013).
Graphing and statistical analysis were performed using the GraphPad Prism program (version Mac 5.0c). After verifying data normality, the unpaired Student’s t-test or the Mann–Whitney test were used for comparisons between two groups. For Gaussian distributions, one-way ANOVA was used, while for non-Gaussian distribution, the Kruskal–Wallis non-parametric test was employed for comparisons among three or more groups. Differences among groups were considered statistically significant when p < 0.05. Principal component analysis (PCA) and Hierarchical cluster analysis (Ward’s method) were used to test whether L. amazonensis and L. major could be clustered separately by the percentage of infected macrophages, parasite load, intracellular viability, percentage of LC3-decorated vacuoles as well as percentage of DQ-BSA and LysoTracker positives vacuoles. These data analyses were performed using JMP 11.0 (SAS) software.
The LC3-II/Act ratio was employed as an approach to evaluate activation of the autophagic pathway. At 30 min of infection, no changes in the LC3-II/Act ratio were found under either L. amazonensis or L. major infection (Figure 1). After 24 h, similar increases in the LC3-II/Act ratio were seen in both L. amazonensis- and L. major-infected macrophages (1.79- and 1.80-fold, respectively) in comparison to uninfected macrophages (Figure 1). LC3-II/Act ratio was found to be higher in positive controls compared to control untreated macrophages: 2.1-fold in macrophages treated with rapamycin and 3.2-fold in cells incubated under nutritional stress conditions (data not shown). Autophagic flux was evaluated by comparing the LC3-II band intensity in cells treated with chloroquine that blocks autophagic flux. As shown in Figure 1 in L. amazonensis- or L. major-infected cells the levels of LC3-II are increased in chloroquine treated cells, indicating that the autophagic flux was normal in the infected cells.
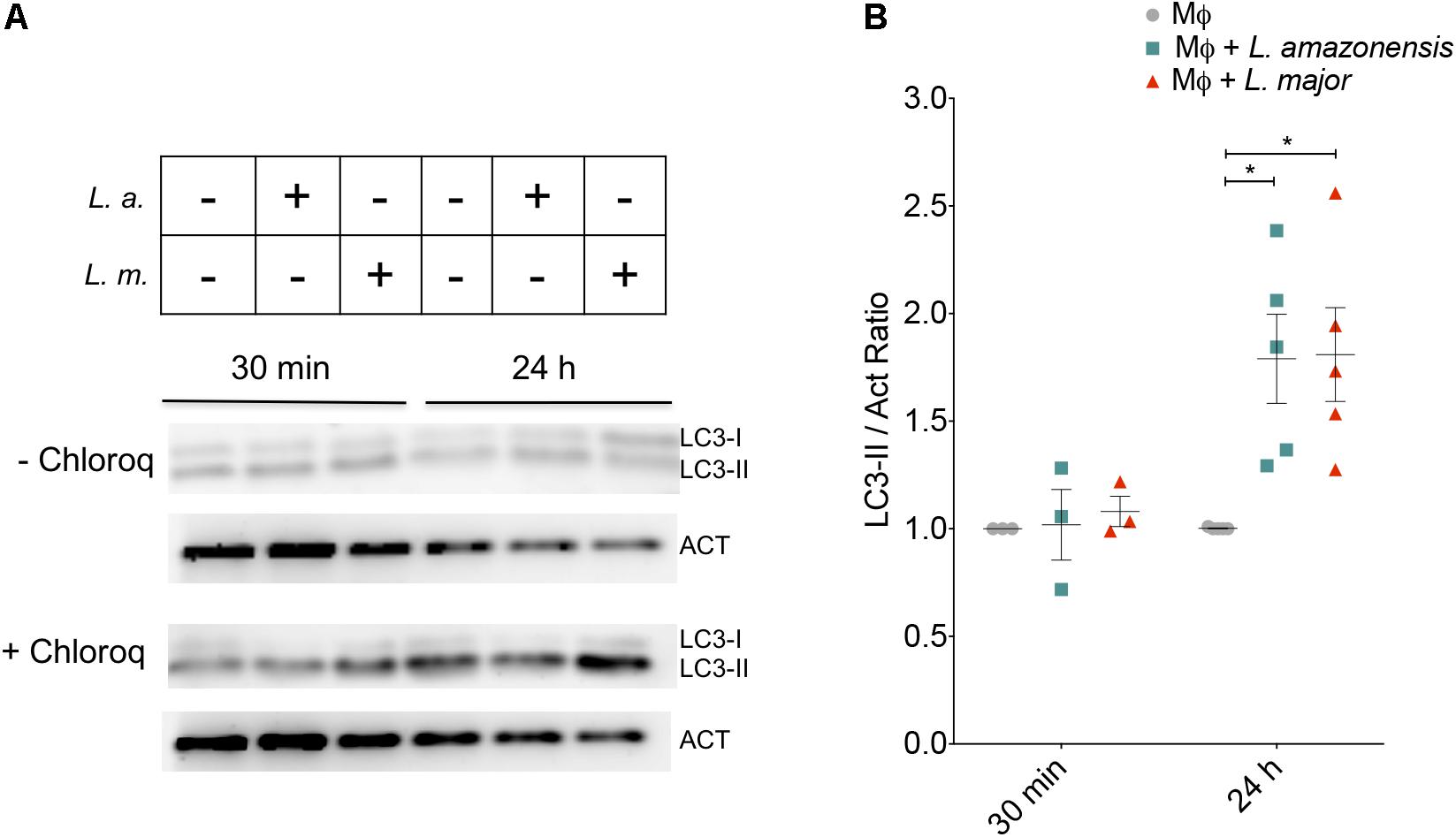
FIGURE 1. Autophagy is activated similarly in L. amazonensis- and L. major-infected cells. CBA mouse macrophages were infected with metacyclic promastigotes of L. amazonensis or L. major. (A) Western blot of LC3-II and Actin (Act) expression in CBA mouse macrophages infected with L. amazonensis or L. major for 30 min or for 4 h, followed by a 24 h-incubation period. Autophagic flux inhibition was achieved by treating the macrophages for 6 h with chloroquine (10 μM) in both uninfected and infected cells Actin was used as internal loading control. (B) Protein bands were densitometrically quantified to determined LC3II/Act ratios. Symbols represent individuals experiments, while lines are representative of means ± SE (One-way ANOVA, ∗p < 0.05). MΦ: Macrophages; MΦ + L.a.: Macrophages infected with L. amazonensis; MΦ + L.m.: Macrophages infected with L. major.
The characteristics of parasitophorous vacuoles were evaluated to determine whether dissimilarities in the capacity of L. amazonensis- and L. major-induced parasitophorous vacuoles to interact with the autophagic pathway can account to differences in parasite survival inside host cells (Alexander and Vickerman, 1975; Antoine et al., 1998; Wilson et al., 2008; Real and Mortara, 2012). First, the hydrolytic activity inside these compartments was assessed by determining the percentage of parasitophorous vacuoles labeled with DQ-BSA. After 30 min of infection, the percentage of L. amazonensis- and L. major-induced parasitophorous vacuoles labeled with DQ-BSA was similar (Figures 2A–C). By contrast, after 4 h of infection, L. major vacuoles (46%) exhibited greater positivity for DQ-BSA than those induced by L. amazonensis (33.33%) (Figure 2C). Similarly, after 24 h of infection, 50% of the L. major-induced parasitophorous vacuoles were positive for DQ-BSA versus only 25% of the L. amazonensis vacuoles (p < 0.01, Figure 2C). The positivity in DQ-BSA was enhanced by 27.58% for L. amazonensis- and 57.33% for L. major-induced parasitophorous vacuoles in macrophages in which autophagy was induced by rapamycin, as compared to untreated cells (p < 0.01, Figure 2D). In spite of this, DQ-BSA positivity was statistically similar in both the vacuoles induced by L. amazonensis and L. major (p > 0.05, Figure 2D), which indicates that L. amazonensis- and L. major-induced parasitophorous vacuoles similarly acquire degradative features in macrophages in which autophagy was induced via mTOR inhibition by rapamycin. In addition, LysoTracker labeling after 24 h of infection revealed that the percentages of parasitophorous vacuoles that accumulated this fluorescent probe are different in L. major-infected cells (42.25%) and L. amazonensis-infected macrophages (31.75), although this difference is not statistically significant (p > 0.05, Figures 2E–G).
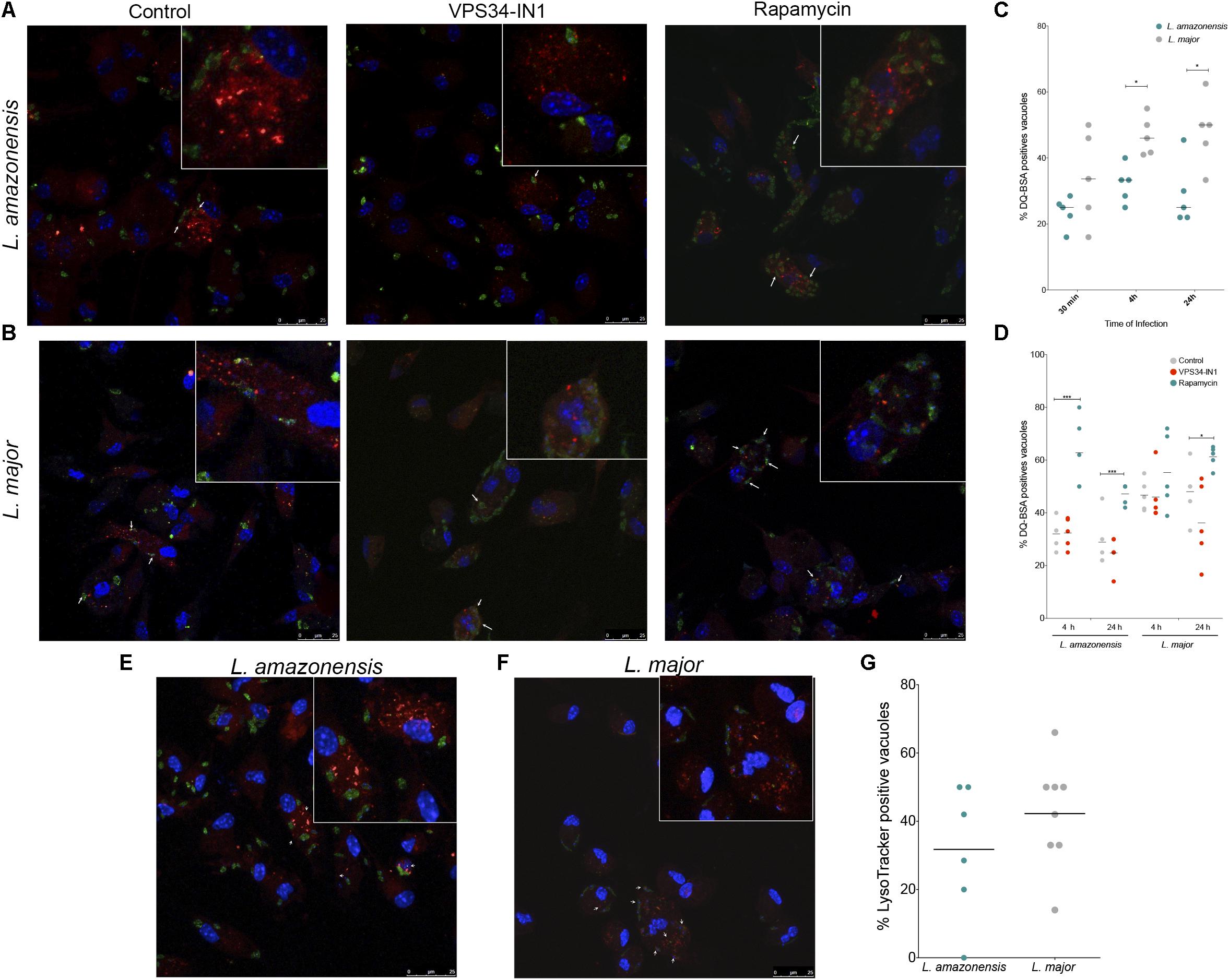
FIGURE 2. Characterization of autolysosomal features in parasitophorous vacuoles induced by L. amazonensis or L. major. (A–D) DQ-BSA labeling: CBA mouse macrophages were incubated with DQ-BSA (red), infected with metacyclic promastigotes of CFSE-labeled L. amazonensis or L. major, and then fixed. Groups of positive and negative control cells were incubated with 10 μg/mL rapamycin or 1 μM VPS34-IN1 after infection and then fixed. For confocal microscopy, cell nuclei were labeled with DAPI (blue). (A) L. amazonensis- (green) or (B) L. major- (green) infected macrophages labeled with DQ-BSA (red). (C) Percentage of L. amazonensis and L. major-induced parasitophorous vacuoles labeled with DQ-BSA (Student’s t-test, ∗p < 0.05). (D) Percentage of DQ-BSA-labeled Leishmania-induced vacuoles in macrophages treated with VPS34-IN1 or rapamycin, or in untreated control cells (One-way ANOVA, ∗∗∗p < 0.01). (E–G) LysoTracker labeling: Macrophages were infected with metacyclic promastigotes of CFSE-labeled L. amazonensis or L. major and labeled with LysoTracker (100 nM) followed by fixing. For confocal microscopy, cell nuclei were labeled with DAPI (blue). (E) L. amazonensis- (green) and (F) L. major-infected macrophages (green) labeled with LysoTracker (red). (G) Percentage of L. amazonensis- and L. major-induced parasitophorous vacuoles labeled with LysoTracker (Student’s t-test, ∗p > 0.05). White arrows indicate labeled parasitophorous vacuoles induced by Leishmania, and circles correspond to each randomly selected field analyzed.
LC3 labeling showed positivity even at early stages of infection, i.e., 30 min, as L. amazonensis- and L. major-induced parasitophorous vacuoles were decorated with endogenous LC3. In fact, at 30 min and 4 h after infection, both L. amazonensis- and L. major-infected macrophages exhibited similar percentages of LC3 decorated parasitophorous vacuoles (Figures 3A–C). Only after 24 h of infection differences were observed in endogenous LC3 labeling in these compartments, with 42.36% of positivity detected in L. amazonensis-induced parasitophorous vacuoles versus only 18.10% in L. major (p < 0.01, Figure 3C).
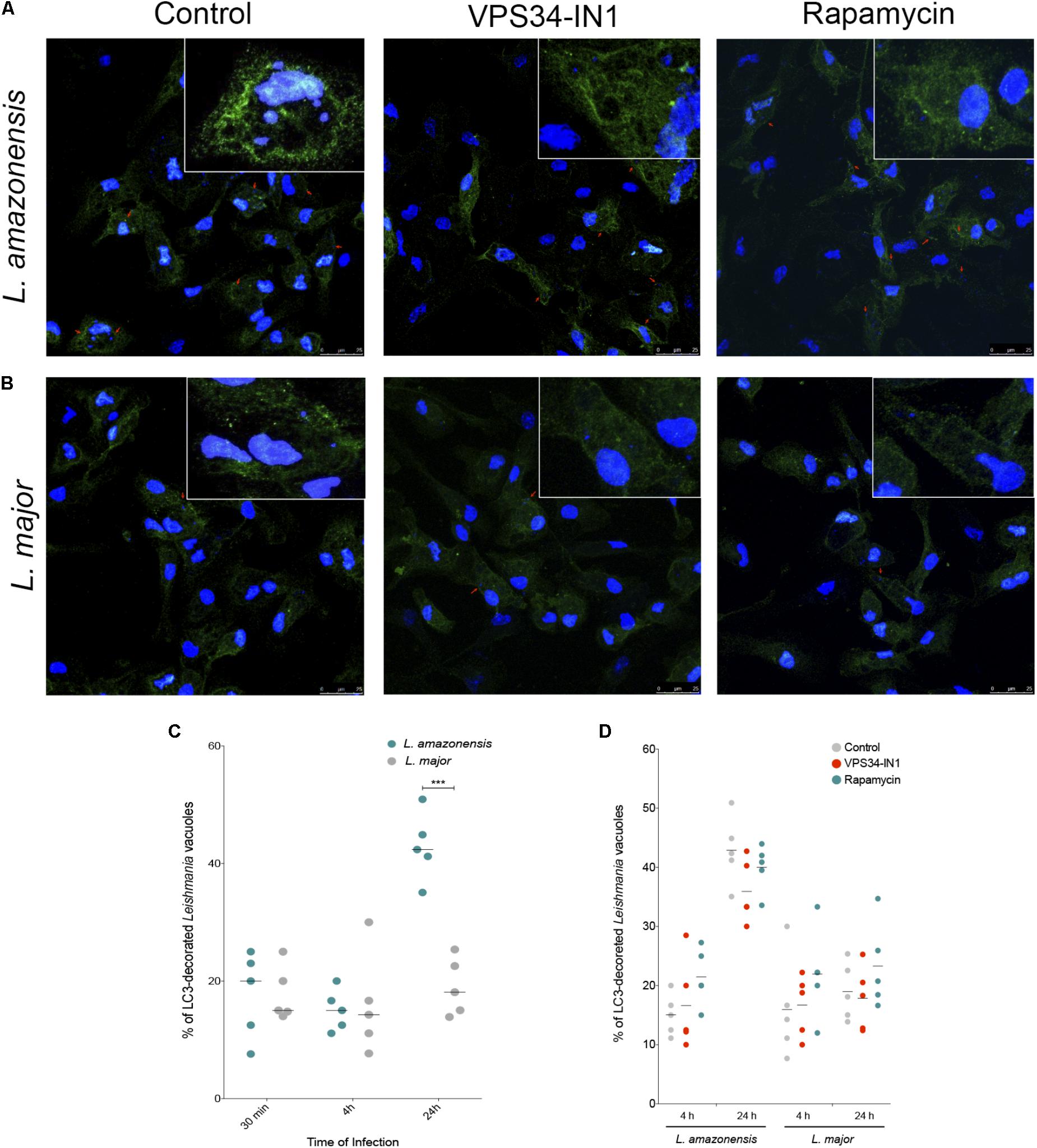
FIGURE 3. Assessment of LC3 recruitment to parasitophorous vacuoles induced by L. amazonensis and L. major. CBA mouse macrophages were infected with metacyclic promastigotes of L. major or L. amazonensis. A group of control cells were incubated with 10 μg/mL rapamycin or 1 μM VPS34-IN1 or left untreated after infection. Cells were then fixed and labeled with anti-LC3 antibody followed by the secondary anti-rabbit IgG antibody conjugated to Alexa Fluor 488 (green). For confocal microscopy, cell nuclei were labeled with DAPI (blue). Confocal microscopy images of (A) L. amazonensis- or (B) L. major-infected macrophages labeled with LC3. (C) Percentage of L. amazonensis- or L. major-induced parasitophorous vacuoles decorated with LC3-II (Student’s t-test, ∗∗∗p < 0.01). (D) Percentage of LC3-decorated Leishmania-induced vacuoles in macrophages treated with VPS34-IN1 or rapamycin, or in untreated control cells (One-way ANOVA, p > 0.05). Circles correspond to each randomly selected field analyzed.
Additional control experiments using macrophages in which autophagy was inhibited by VPS34-IN1, a specific PI3K class III inhibitor, or induced by rapamycin showed no alterations in LC3 positivity as measured by the percentage of L. amazonensis- or L. major-induced vacuoles decorated with LC3 in comparison to infected untreated macrophages (Figure 3D). This result indicates that LC3-recruiment to Leishmania-induced parasitophorous vacuoles seems to be independent of the PI3k-Akt-mTOR pathway.
The role played by autophagy in Leishmania infection was evaluated using autophagic modulators, followed by the determination of the percentage of infected macrophages and number of viable intracellular parasites. No alterations were seen in the percentage of macrophages infected with L. amazonensis or L. major (Figures 4A–D) or in the numbers of Leishmania per infected cell (Supplementary Figure S1) in comparison to untreated macrophages, regardless of autophagic inhibition by chloroquine, wortmannin or VPS34-IN1or induction by rapamycin or starvation. Also, none of the autophagic inhibitors influenced the number of viable intracellular L. amazonensis or L. major parasites (Figures 4E,F). However, we observed that induction by starvation or treatment with rapamycin enhanced the number of recovered viable parasites of both L. major and L. amazonensis species in comparison to infected untreated controls (Figures 4G,H). Pharmacologically induced autophagy by rapamycin led to significant increases of 140.25, 146.18, and 157.15% in the number of viable L. major intracellular parasites after 4, 10, and 16 h of infection, respectively, in comparison to controls (Figure 4H) for L. amazonensis, differences of 61.7 and 76.39% were seen in the number of viable parasites at 4 and 10 h after treatment, respectively (Figure 4G). Interestingly, we observed that the physiological induction of autophagy by starvation or pharmacological induction by rapamycin led to a greater enhancement in L. major intracellular parasite viability than L. amazonensis in comparison to untreated infected cells (Figures 4I,J). The physiological induction of autophagy was shown to enhance intracellular viability by 2.5 and 1.5 times in L. major versus L. amazonensis, respectively, at 10 and 16 h after treatment (p < 0.05) (Figure 4I). Additionally, the pharmacological induction of autophagy by rapamycin enhanced L. major intracellular viability by 2.17 times as compared to L. amazonensis viability at as early as 4 h after treatment (p < 0.05) (Figure 4J). Additionally, similar enhancements in intracellular parasite viability were seen in L. major compared to L. amazonensis at both 10 and 16 h after treatment (Figure 4J).
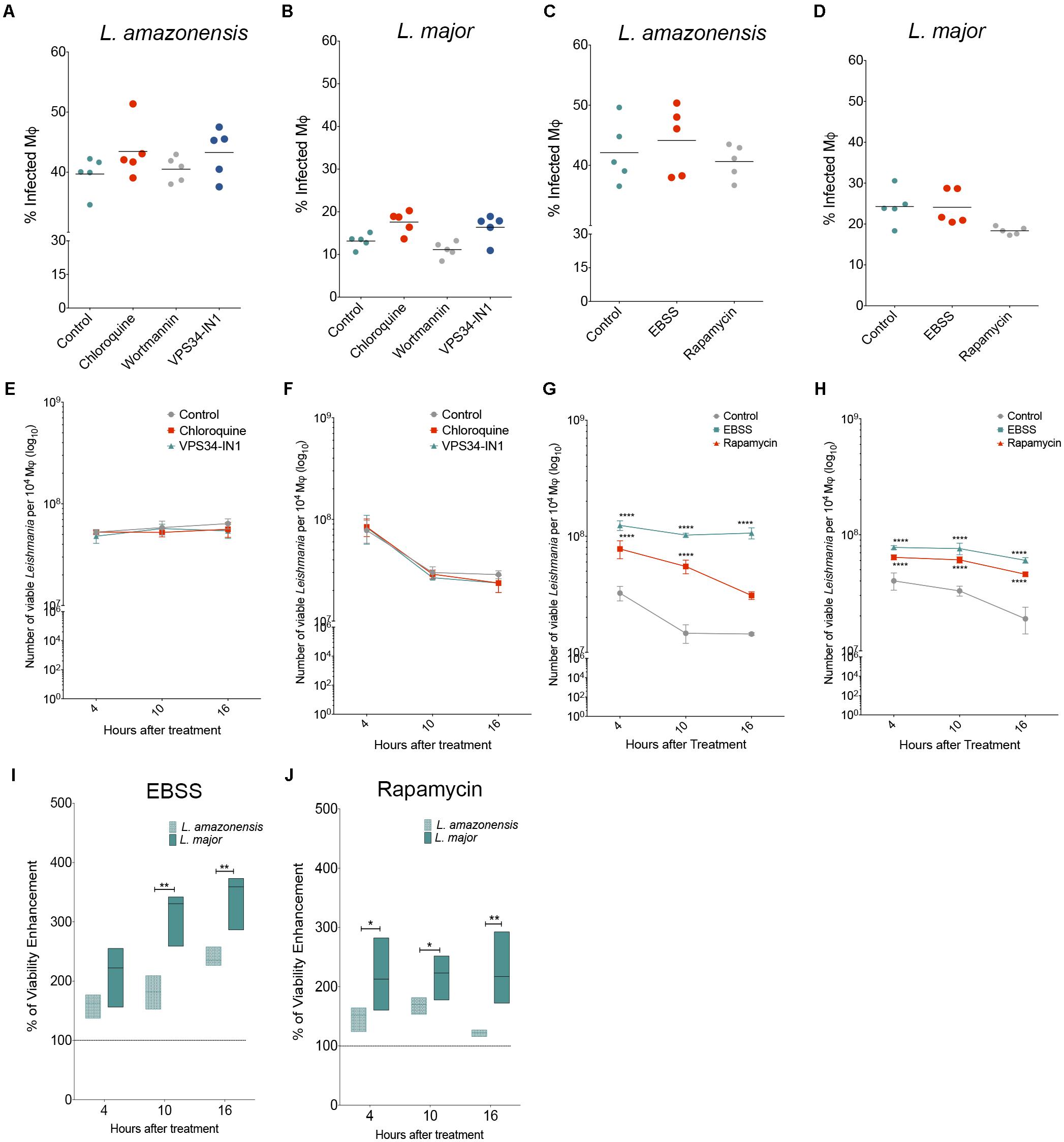
FIGURE 4. Effect of autophagy modulation on Leishmania infection. CBA mouse macrophages were infected with L. amazonensis or L. major and incubated with different autophagic modulators. (A–D) To determine the percentage of L. amazonensis- or L. major-infected cells were incubated with autophagic inhibitors (A,B): chloroquine (10 μM), wortmannin (100 nM) or VPS34-IN1 (1 μM), or with autophagic inducers (C,D): nutrient-poor EBSS medium or rapamycin (10 μg/mL). After 4 h, all cell groups were reincubated for an additional 24 h, fixed and stained with DAPI. Circles represent data from each replicate of one of two independent experiments performed in quintuplicate (Kruskal–Wallis test, Dunns post-test, p > 0.5). (E–J) To assess intracellular (E,G) L. amazonensis or (F,H) L. major viability, infected macrophages were incubated with the (E,F) autophagic inhibitors: chloroquine (10 μM) or VPS34-IN1 (1 μM) or (G,H) autophagic inducers: nutrient-poor EBSS medium or rapamycin (10 μg/mL). After incubation, cells were reincubated in Schneider’s medium to determine the number of viable parasites. (I,J) Percentage of enhancement in intracellular parasite viability after incubation with (I) EBSS or (J) rapamycin. Circles correspond to the mean of one representative out of two independent experiments performed in quintuplicate ± SD (Two-Way ANOVA test, Tukey’s multiple comparisons post-test ∗∗∗∗p < 0.0001). Lines represent median values and floating bars show quartiles (25% and 75%) of one representative experiment out of two independent experiments performed in quintuplicate (Mann–Whitney Test, ∗p < 0.05 and ∗∗p < 0.01).
To determine whether NO and arginase are involved in the enhanced L. amazonensis and L. major intracellular viability induced by autophagy, we evaluated the effect of rapamycin-induced autophagy on NO production and arginase activity in infected macrophages. Unstimulated control cells, both uninfected and infected with L. amazonensis or L. major, produced NO at undetectable levels. Uninfected macrophages stimulated with IFN-γ released 2.30 μM of NO in the culture medium (data not shown). The addition of rapamycin or VPS24-IN1 to these cell cultures altered NO production to 0.74 or 4.61 μM, respectively (data not shown). The addition of rapamycin to L. amazonensis-infected macrophages resulted in a NO production level of 4.33 μM in comparison to controls (11.40 μM) (p < 0.05, Figure 5). Additionally, VPS34-IN1 enhanced NO production by L. amazonensis-infected macrophages (35.55 μM) in comparison to control cells (11.40 μM) (p < 0.05, Figure 5). Similarly, in L. major-infected macrophages, the addition of VPS34-IN1 to cell cultures increased NO levels to 22.17 μM, while the addition of rapamycin reduced NO production to 1.46 μM in comparison to 6.35 μM of NO released by untreated L. major-infected macrophages (p < 0.05, Figure 5). Interestingly, no significant differences were observed in arginase activity between the Leishmania-infected macrophages treated with rapamycin or VPS34-IN1 and untreated control macrophages (p > 0.05, Supplementary Figure S2). This seems to suggest that the induction of autophagy favors Leishmania intracellular viability by reducing NO production.
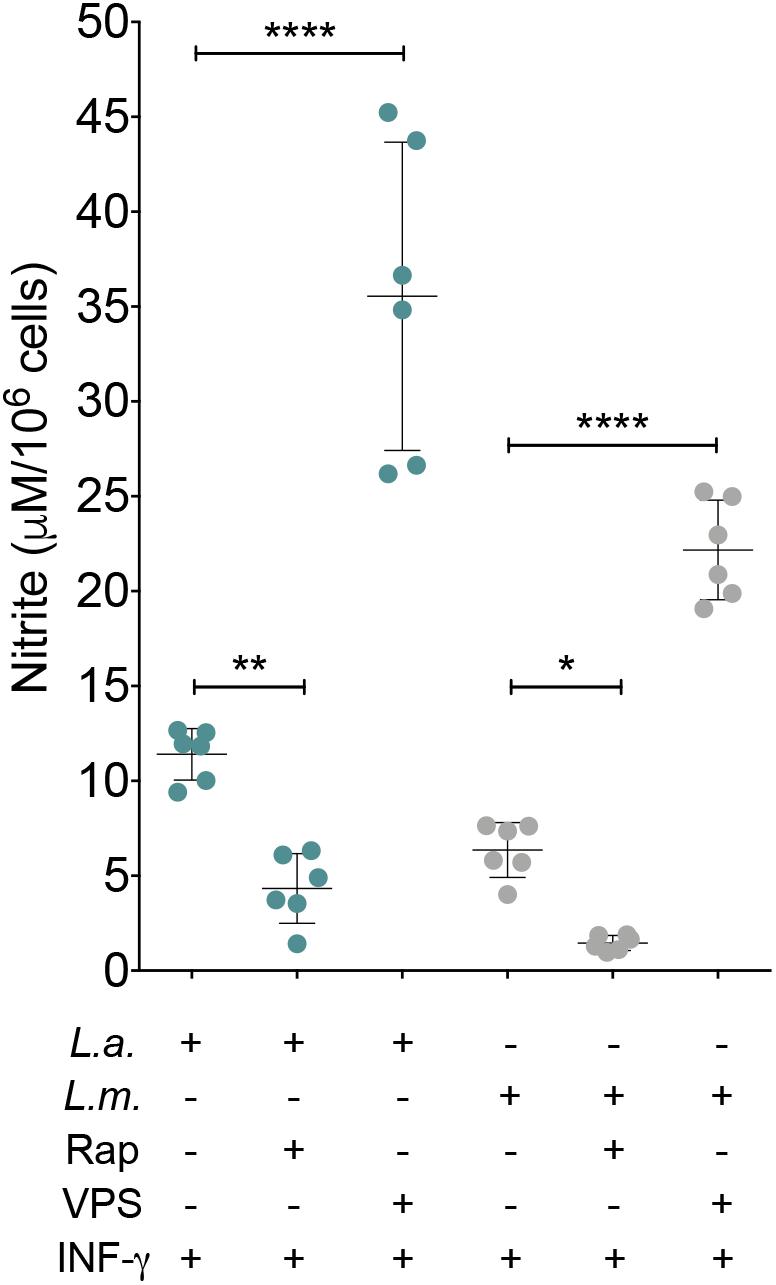
FIGURE 5. Autophagic induction reduces NO production by L. major- and L. amazonensis-infected macrophages. CBA mouse macrophages were infected with L. amazonensis or L. major and incubated with VPS34-IN1 (1 μM) or rapamycin (10 μg/mL). After 4 h, all cell groups were reincubated in modulator-free medium for an additional 24 h. Macrophage NO production was measured by detecting nitrite in culture supernatants. Circles correspond to each replica of one representative of two independent experiments performed in sextuplicate ± SD (One-way ANOVA, ∗p < 0.05, ∗∗p < 0.01 and ∗∗∗∗p < 0.0001). L.a., L. amazonensis; L.m., L. major; Rap, rapamycin; VPS, VPS34-IN1.
A PCA was used to verify whether the parameters above evaluated: characterization of parasitophorous vacuoles regarding autophagic features and the percentage of infected macrophages, parasitic load and intracellular parasite viability, could discriminate L. amazonensis and L. major infection (Figures 6A–C). The analyzed parameters allowed differentiation of two major groups separating the L. amazonensis-infected macrophages from those infected by L. major (Figures 6A,B). Additionally, heatmap analysis revealed that L. amazonensis-infected macrophages are grouped into a cluster associated to infected cells presenting higher percentage of LC3-decorated vacuoles, higher percentage of infected macrophages and higher intracellular parasite viability, while L. major-infected macrophages clustered in a group associated to higher percentage of vacuoles positive for DQ-BSA and LysoTracker (Figure 6B).
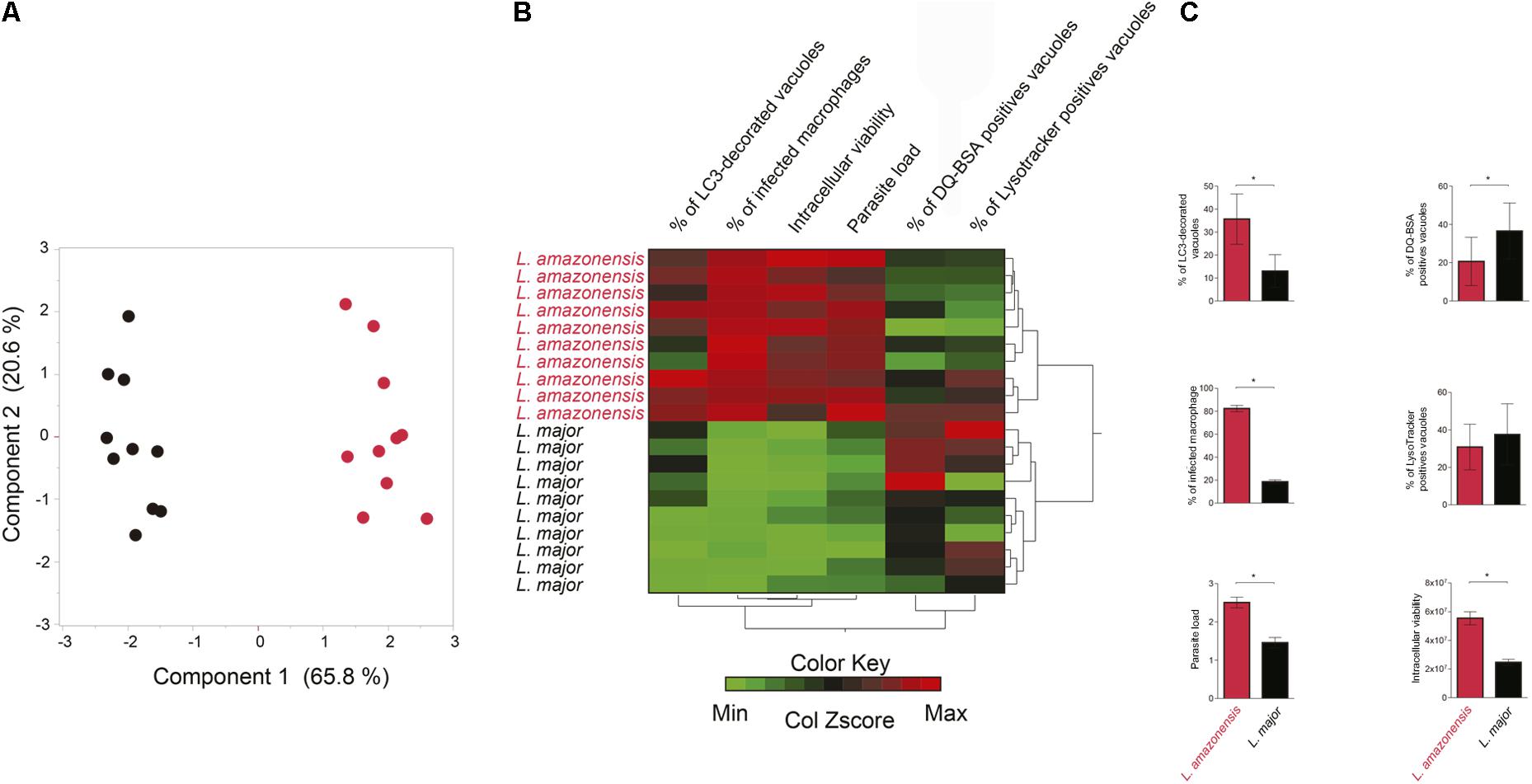
FIGURE 6. L. amazonensis- and L. major-infected macrophages are clustered in different groups regarding autophagic features and infection parameters. Cluster analysis was performed using the following variables: percentage of infected macrophages, intracellular viability, number of Leishmania per infected macrophages, percentage of LC3 decorated Leishmania vacuoles and percentage of DQ BSA and LysoTracker positives vacuoles. (A) Principal component analysis. Black dots represent L. major-infected macrophages and red dots L. amazonensis-infected macrophages. (B) Hierarchical cluster analysis (Ward’s method). Red represents maximum in the color key, green represents minimum and mean values are shown in black. (C) Statistical analysis of autophagic features and infection parameters of L. amazonensis- and L. major-infected cells evaluated in the heat map (Student’s t Test, ∗p < 0.05).
The present study demonstrated that L. amazonensis and L. major both induce autophagy in CBA macrophages, yet L. amazonensis infection was shown to produce a higher percentage of parasite-induced vacuoles decorated by endogenous LC3-II. On the other hand, the compartments induced by L. amazonensis presented lower positivity for the degradative marker DQ-BSA. With respect to autophagy modulation in L. amazonensis and L. major infection in vitro, no changes were seen in the percentage of L. amazonensis or L. major infected cells or parasite load when autophagy was induced or inhibited, either physiologically or pharmacologically. While the inhibition of autophagy did not affect Leishmania intracellular viability, exogenously induced autophagy was shown to enhance intracellular parasite viability.
The present study also found comparatively similar increases in the LC3-II/Act ratio in inflammatory peritoneal CBA mouse macrophages infected with L. amazonensis or L. major. We further demonstrated that increased levels of LC3-II were associated with the induction of autophagy per se, i.e., not via the blockage of autophagic flux, since lower levels of LC3-II were seen in infected cells compared to those treated with chloroquine, an inhibitor of autophagic flux. These findings stand in agreement with previous studies reporting increased LC3 expression in RAW cells, BALB/c and C57BL/6 bone marrow macrophages infected by L. amazonensis (Cyrino et al., 2012) or L. major (Franco et al., 2017), and BALB/c bone marrow macrophages infected with L. major (Frank et al., 2015) in comparison to uninfected cells. Consistent with our results and those reported in other in vitro studies, Mitroulis et al. (2009) observed a greater conversion of LC3-I to LC3-II in a sample of bone marrow macrophages from a male patient with visceral leishmaniasis infected with L. donovani in comparison to a bone marrow sample from a healthy patient. These findings seem to suggest that, regardless of species, Leishmania infection provokes the activation of the autophagic pathway in host cells in vitro and in vivo.
Several studies have successfully demonstrated that vacuoles induced by intracellular microorganisms can be loaded with DQ-BSA or LysoTracker (Gutierrez et al., 2004; Aguilera et al., 2009; Lerena and Colombo, 2011). While these soluble markers are not specific to autophagy, their use in labeling in conjunction with other autophagic compartment markers represents a suitable and reliable tool to monitor acidity and degradative activity in compartments, such as autolysosomes (Klionsky et al., 2007; Mitou et al., 2009; Vazquez and Colombo, 2009). Our analysis found no significant differences in the percentages of LysoTracker-labeled vacuoles induced by either L. amazonensis or L. major. Labeling of both L. amazonensis- and L. major-induced parasitophorous vacuoles with this lysosomal dye was previously demonstrated by Real and Mortara (2012). While these authors did not quantify the percentage of Leishmania-induced vacuoles labeled with LysoTracker, comparisons of the intensity of this probe revealed that the parasitophorous vacuoles induced by L. major presented lower intensity than L. amazonensis-induced compartments (Real and Mortara, 2012) suggesting that the acidity of these compartments was somehow diminished. In the present report, a higher percentage of DQ-BSA positivity was seen in L. major-induced vacuoles compared to those induced by L. amazonensis at both 4 and 24 h after infection. Previously, Beron et al. (2002) and Aguilera et al. (2009) successfully marked the large vacuoles induced by Coxiella burnetti with DQ-BSA. These vacuoles have been described as presenting morphological similarities to those induced by L. amazonensis (Antoine et al., 1990; Maurin et al., 1992; Veras et al., 1994, 1995). However, different from C. burnetii, L. amazonensis at later infection times maybe somehow preventing fusion with degradative compartments.
The recruitment of LC3 to L. major-induced parasitophorous vacuoles has been previously demonstrated (Crauwels et al., 2015; Matte et al., 2016). Crauwels et al. (2015) also reported greater percentages (92 ± 0.7%) of LC3-positive parasitophorous vacuoles induced by apoptotic L. major in comparison to those induced by viable L. major (7 ± 1%). Herein, we found similar percentages of LC3-decorated membranes of both L. amazonensis- and L. major-induced parasitophorous vacuoles as early as 30 min after infection. However, after 24 h, greater LC3-positivity was detected in L. amazonensis-induced parasitophorous vacuoles than in those induced by L. major. Interestingly, no changes in the percentage of LC3 positivity were seen in the vacuoles induced by L. amazonensis or L. major in macrophages treated with VPS34-IN1 or rapamycin when compared to untreated control macrophages. A similar result was described by Thomas et al. (2017), who found no differences in LC3-II levels between L. donovani-infected human THP-1 cells and those infected and treated with rapamycin. These authors hypothesized that the lack of an increase in LC3-II positivity observed in L. donovani-infected cells previously treated with rapamycin could be due to the fact that this parasite species inhibits classical autophagy activation via the PI3k-Akt-mTOR pathway, instead inducing this process via another pathway independent of mTOR. Accordingly, it is possible that in our CBA mouse macrophage model, this same mTOR-independent pathway was used to activate autophagy by both L. amazonensis and L. major. Alternatively, it is plausible to suggest that treatment with rapamycin resulted in a lack of enhancement in the percentage of LC3-II-decorated vacuoles in Leishmania-infected macrophages, associated with the more intense DQ-BSA labeling also found in Leishmania vacuoles, could be a result of the previously described accumulation of hydrolytic enzymes and, subsequently, increased cleavage of LC3-II in the inner membrane of these compartments (Kirisako et al., 2000; Tanida et al., 2004).
Many studies have been carried out in an attempt to determine whether parasite survival is favored by host cell autophagy under infection by different Leishmania species, or whether it functions as a defense mechanism. Of note, PCA and heat map analyses performed in the present study, completely discriminated L. major-infected macrophages, which grouped for higher positivity for markers of degradative compartments, DQ-BSA and Lysotracker, from L. amazonensis-infected cells that clustered for higher intensity of infection and LC3 positivity. Unexpectedly, autophagic inhibition exerted no effects on Leishmania spp infection, while exogenously induced autophagy was shown to favor intracellular viability of both parasites, although in a higher extent for L. major than L. amazonensis. In conjunction, these findings suggest that triggering autophagic process by PI3k-Akt-mTOR pathway seems to be beneficial to Leishmania spp intracellular survival, more intensely for those species that survive in a compartment presenting degradative characteristics. Thus, autophagy induction after infection would be detrimental to the host, since it would favor the intracellular viability of Leishmania. Consistently with our findings, Pinheiro et al. (2009) demonstrated that autophagy induced by starvation after infection led to an increase in the intracellular viability of L. amazonensis in BALB/c and A/J macrophages, as well as in the J774 cell-line. However, in contrast to our findings, these same authors found that starvation-induced autophagy did not alter the intracellular viability of L. amazonensis in infected C57BL/6 macrophages or that of L. major in infected BALB/c macrophages (Pinheiro et al., 2009). Moreover, it has also been demonstrated that pre-infection autophagic inhibition via Atg5 knockdown in bone-marrow macrophages resulted in an increase in L. major parasite burden (Frank et al., 2015). In the present study, autophagy has been modulated after infection, since, previously, we found that the induction of autophagy prior to infection inhibited the phagocytic capacity of mammalian cells in a non-specific manner (Lima et al., 2011). It is possible that the reduction in Atg5 expression induced by Frank et al. (2015) in bone marrow macrophages provoked an enhancement in L. major infection due to an increase in parasite uptake, as previously described for M. tuberculosis (Bonilla et al., 2013). Accordingly, we propose that results reported by these previous studies, which sought to assess the effects of knocking down autophagy-related genes in the context of Leishmania infection, serve to reinforce the notion that inhibiting or inducing autophagy prior to infection profoundly interferes with phagocytic capacity of cell, rather than affecting the intracellular survival of pathogens.
By investigating how the exogenous induction of autophagy favors Leishmania intracellular viability, we have demonstrated that rapamycin decreases NO levels but does not significantly alter arginase activity. Consistent with our finding, Pinheiro et al. (2009) previously demonstrated that starvation-induced autophagy decreased NO production by L. amazonensis-infected macrophages in association with enhanced parasite intracellular viability. This mechanism seems to be universal, since it has already been shown that autophagic induction reduces NO generation by microglia (Han et al., 2013), the RAW 264.7 macrophage cell line (Jin et al., 2009), and also by ischaemic/reperfused myocardium (Ma et al., 2018). We speculate that, in our model, rapamycin may enhance parasite viability via reduced NO production, by way of one of the two following mechanisms: the suppression of mRNA expression of inducible nitric oxide synthase (iNOS) (Han et al., 2013) or the induction of iNOS proteasomal degradation (Jin et al., 2009).
Taken together, the present findings seem to indicate that although both of the evaluated species of Leishmania activate the autophagic pathway similarly in CBA mouse macrophages, the resulting parasitophorous vacuoles present different characteristics: enhanced LC3 recruitment is observed in L. amazonensis-induced parasitophorous vacuoles in comparison to L. major-induced vacuoles, which present more degradative features (result PCA). Despite these differences, we demonstrated that autophagic induction favors intracellular parasite survival in both L. amazonensis and L. major, albeit more pronounced in L. major, which seems to be related to decreased NO production. Alternatively, it is possible that autophagosomes may provide metabolites and amino acids to L. major- and L. amazonensis-induced vacuoles, which can favor the growth and intracellular development of Leishmania. One of these metabolites may be iron, since stored within ferritin, can be mobilized and released through autophagy (Linder, 2013; Niu et al., 2016) and then transported into the leishmanial cytosol by LIT1 (Huynh et al., 2006).
BD and PV conceived and designed the experiments. BD, CS, JL, NJA, TS, and JF-C performed the experiments. BD, KF, CB, JM, MC, and PV analyzed the data. PV contributed reagents, materials, and analysis tools. BD and PV wrote the manuscript. All the authors contributed to manuscript elaboration and revision and approved the final version prior to submission.
This work was supported by grants from Fundação de Amparo à Pesquisa do Estado da Bahia (PV http://www.fapesb.ba.gov.br), Conselho Nacional de Pesquisa e Desenvolvimento Científico (PV http://www.cnpq.br) and National Institute of Science and Technology of Tropical (PV http://inct.cnpq.br/web/inct-dt). PV holds a grant from CNPq for productivity in research (307832/2015-5). The funders had no role in study design, data collection or analysis, the decision to publish, or preparation of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank Dr. Adriana Lanfredi Rangel, Dr. Claudio Pereira Figueira, and Marcio Ferreira dos Santos for technical support in confocal microscopy imaging. Finally, we are grateful to Andris K. Walter for English language revision and manuscript copyediting assistance.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01890/full#supplementary-material
FIGURE S1 | Effect of autophagic modulation on the number of Leishmania per infected macrophage. CBA mouse macrophages were infected with L. amazonensis or L. major and incubated with different autophagic modulators. To determine parasite load, L. amazonensis or L. major-infected cells were incubated with (A) autophagic inhibitors: chloroquine (10 μM), wortmannin (100 nM) or VPS34-IN1 (1 μM), or with (B) autophagic inducers: nutrient-poor EBSS medium or rapamycin (10 μg/mL). After 4 h, all cell groups were reincubated for an additional 24 h, fixed and stained with DAPI. Circles represent data from each replicate of one of two independent experiments performed in quintuplicate (Kruskal–Wallis test, Dunns post-test, p > 0.5).
FIGURE S2 | Effect of autophagic modulation on arginase activity. CBA mouse macrophages were infected with L. amazonensis or L. major and incubated with VPS34-IN1 (1 μM) or rapamycin (10 μg/mL). After 4 h, all cell groups were reincubated in modulator-free medium for an additional 24 h. Arginase activity was determined by urea production assay. Circles correspond to each replica of one experiment performed in sextuplicate ± SD (One-way ANOVA). L.a., L. amazonensis; L.m., L. major; Rap, rapamycin; VPS, VPS34-IN1.
Abebe, T., Takele, Y., Weldegebreal, T., Cloke, T., Closs, E., Corset, C., et al. (2013). Arginase activity - a marker of disease status in patients with visceral leishmaniasis in ethiopia. PLoS Negl. Trop. Dis. 7:e2134. doi: 10.1371/journal.pntd.0002134
Aguilera, M., Salinas, R., Rosales, E., Carminati, S., Colombo, M. I., and Beron, W. (2009). Actin dynamics and Rho GTPases regulate the size and formation of parasitophorous vacuoles containing Coxiella burnetii. Infect. Immun. 77, 4609–4620. doi: 10.1128/IAI.00301-09
Alexander, J., and Vickerman, K. (1975). Fusion of host cell secondary lysosomes with the parasitophorous vacuoles of Leishmania mexicana-infected macrophages. J. Protozool. 22, 502–508. doi: 10.1111/j.1550-7408.1975.tb05219.x
Alvar, J., Velez, I. D., Bern, C., Herrero, M., Desjeux, P., Cano, J., et al. (2012). Leishmaniasis worldwide and global estimates of its incidence. PLoS One 7:e35671. doi: 10.1371/journal.pone.0035671
Antoine, J. C., Prina, E., Jouanne, C., and Bongrand, P. (1990). Parasitophorous vacuoles of Leishmania amazonensis-infected macrophages maintain an acidic pH. Infect. Immun. 58, 779–787.
Antoine, J. C., Prina, E., Lang, T., and Courret, N. (1998). The biogenesis and properties of the parasitophorous vacuoles that harbour Leishmania in murine macrophages. Trends Microbiol. 6, 392–401. doi: 10.1016/S0966-842X(98)01324-9
Bago, R., Malik, N., Munson, M. J., Prescott, A. R., Davies, P., Sommer, E., et al. (2014). Characterization of VPS34-IN1, a selective inhibitor of Vps34, reveals that the phosphatidylinositol 3-phosphate-binding SGK3 protein kinase is a downstream target of class III phosphoinositide 3-kinase. Biochem. J. 463, 413–427. doi: 10.1042/BJ20140889
Beron, W., Gutierrez, M. G., Rabinovitch, M., and Colombo, M. I. (2002). Coxiella burnetii localizes in a Rab7-labeled compartment with autophagic characteristics. Infect. Immun. 70, 5816–5821. doi: 10.1128/IAI.70.10.5816-5821.2002
Bonilla, D. L., Bhattacharya, A., Sha, Y., Xu, Y., Xiang, Q., Kan, A., et al. (2013). Autophagy regulates phagocytosis by modulating the expression of scavenger receptors. Immunity 39, 537–547. doi: 10.1016/j.immuni.2013.08.026
Chang, H. K., Thalhofer, C., Duerkop, B. A., Mehling, J. S., Verma, S., Gollob, K. J., et al. (2007). Oxidant generation by single infected monocytes after short-term fluorescence labeling of a protozoan parasite. Infect. Immun. 75, 1017–1024. doi: 10.1128/IAI.00914-06
Crauwels, P., Bohn, R., Thomas, M., Gottwalt, S., Jackel, F., Kramer, S., et al. (2015). Apoptotic-like Leishmania exploit the host’s autophagy machinery to reduce T-cell-mediated parasite elimination. Autophagy 11, 285–297. doi: 10.1080/15548627.2014.998904
Cyrino, L. T., Araujo, A. P., Joazeiro, P. P., Vicente, C. P., and Giorgio, S. (2012). In vivo and in vitro Leishmania amazonensis infection induces autophagy in macrophages. Tissue Cell 44, 401–408. doi: 10.1016/j.tice.2012.08.003
Duszenko, M., Ginger, M. L., Brennand, A., Gualdron-Lopez, M., Colombo, M. I., Coombs, G. H., et al. (2011). Autophagy in protists. Autophagy 7, 127–158. doi: 10.4161/auto.7.2.13310
Franco, L. H., Fleuri, A. K. A., Pellison, N. C., Quirino, G. F. S., Horta, C. V., de Carvalho, R. V. H., et al. (2017). Autophagy downstream of endosomal Toll-like receptor signaling in macrophages is a key mechanism for resistance to Leishmania major infection. J. Biol. Chem. 292, 13087–13096. doi: 10.1074/jbc.M117.780981
Frank, B., Marcu, A., de Oliveira Almeida Petersen, A. L., Weber, H., Stigloher, C., Mottram, J. C., et al. (2015). Autophagic digestion of Leishmania major by host macrophages is associated with differential expression of BNIP3, CTSE, and the miRNAs miR-101c, miR-129, and miR-210. Parasit. Vectors 8:404. doi: 10.1186/s13071-015-0974-3
Gomes, I. N., Calabrich, A. F., Tavares Rda, S., Wietzerbin, J., de Freitas, L. A., and Veras, P. S. (2003). Differential properties of CBA/J mononuclear phagocytes recovered from an inflammatory site and probed with two different species of Leishmania. Microbes Infect. 5, 251–260. doi: 10.1016/S1286-4579(03)00025-X
Gutierrez, M. G., Master, S. S., Singh, S. B., Taylor, G. A., Colombo, M. I., and Deretic, V. (2004). Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell 119, 753–766. doi: 10.1016/j.cell.2004.11.038
Gutierrez, M. G., Vazquez, C. L., Munafo, D. B., Zoppino, F. C., Beron, W., Rabinovitch, M., et al. (2005). Autophagy induction favours the generation and maturation of the Coxiella-replicative vacuoles. Cell. Microbiol. 7, 981–993. doi: 10.1111/j.1462-5822.2005.00527.x
Han, H. E., Kim, T. K., Son, H. J., Park, W. J., and Han, P. L. (2013). Activation of autophagy pathway suppresses the expression of iNOS, IL6 and cell death of LPS-stimulated microglia cells. Biomol. Ther. 21, 21–28. doi: 10.4062/biomolther.2012.089
Huynh, C., Sacks, D. L., and Andrews, N. W. (2006). A Leishmania amazonensis ZIP family iron transporter is essential for parasite replication within macrophage phagolysosomes. J. Exp. Med. 203, 2363–2375. doi: 10.1084/jem.20060559
Jin, H. K., Ahn, S. H., Yoon, J. W., Park, J. W., Lee, E. K., Yoo, J. S., et al. (2009). Rapamycin down-regulates inducible nitric oxide synthase by inducing proteasomal degradation. Biol. Pharm. Bull. 32, 988–992. doi: 10.1248/bpb.32.988
Kabeya, Y., Mizushima, N., Ueno, T., Yamamoto, A., Kirisako, T., Noda, T., et al. (2000). LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 19, 5720–5728. doi: 10.1093/emboj/19.21.5720
Kirisako, T., Ichimura, Y., Okada, H., Kabeya, Y., Mizushima, N., Yoshimori, T., et al. (2000). The reversible modification regulates the membrane-binding state of Apg8/Aut7 essential for autophagy and the cytoplasm to vacuole targeting pathway. J. Cell Biol. 151, 263–276. doi: 10.1083/jcb.151.2.263
Klionsky, D. J., Abdelmohsen, K., Abe, A., Abedin, M. J., Abeliovich, H., Acevedo Arozena, A., et al. (2016). Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 12, 1–222. doi: 10.1080/15548627.2015.1100356
Klionsky, D. J., Cuervo, A. M., and Seglen, P. O. (2007). Methods for monitoring autophagy from yeast to human. Autophagy 3, 181–206. doi: 10.4161/auto.3678
Lang, T., Schaeffeler, E., Bernreuther, D., Bredschneider, M., Wolf, D. H., and Thumm, M. (1998). Aut2p and Aut7p, two novel microtubule-associated proteins are essential for delivery of autophagic vesicles to the vacuole. EMBO J. 17, 3597–3607. doi: 10.1093/emboj/17.13.3597
Lemos de Souza, V., Ascencao Souza, J., Correia Silva, T. M., Sampaio Tavares Veras, P., and Rodrigues de-Freitas, L. A. (2000). Different Leishmania species determine distinct profiles of immune and histopathological responses in CBA mice. Microbes Infect. 2, 1807–1815. doi: 10.1016/S1286-4579(00)01340-X
Lerena, M. C., and Colombo, M. I. (2011). Mycobacterium marinum induces a marked LC3 recruitment to its containing phagosome that depends on a functional ESX-1 secretion system. Cell. Microbiol. 13, 814–835. doi: 10.1111/j.1462-5822.2011.01581.x
Levine, B., and Deretic, V. (2007). Unveiling the roles of autophagy in innate and adaptive immunity. Nat. Rev. Immunol. 7, 767–777. doi: 10.1038/nri2161
Lima, J. G., de Freitas Vinhas, C., Gomes, I. N., Azevedo, C. M., dos Santos, R. R., Vannier-Santos, M. A., et al. (2011). Phagocytosis is inhibited by autophagic induction in murine macrophages. Biochem. Biophys. Res. Commun. 405, 604–609. doi: 10.1016/j.bbrc.2011.01.076
Linder, M. C. (2013). Mobilization of stored iron in mammals: a review. Nutrients 5, 4022–4050. doi: 10.3390/nu5104022
Ma, L. L., Ma, X., Kong, F. J., Guo, J. J., Shi, H. T., Zhu, J. B., et al. (2018). Mammalian target of rapamycin inhibition attenuates myocardial ischaemia-reperfusion injury in hypertrophic heart. J. Cell Mol. Med. 22, 1708–1719. doi: 10.1111/jcmm.13451
Matte, C., Casgrain, P. A., Seguin, O., Moradin, N., Hong, W. J., and Descoteaux, A. (2016). Leishmania major promastigotes evade LC3-associated phagocytosis through the action of GP63. PLoS Pathog. 12:e1005690. doi: 10.1371/journal.ppat.1005690
Maurin, M., Benoliel, A. M., Bongrand, P., and Raoult, D. (1992). Phagolysosomes of Coxiella burnetii-infected cell lines maintain an acidic pH during persistent infection. Infect. Immun. 60, 5013–5016.
Meijer, A. J., and Codogno, P. (2004). Regulation and role of autophagy in mammalian cells. Int. J. Biochem. Cell Biol. 36, 2445–2462. doi: 10.1016/j.biocel.2004.02.002
Mestre, M. B., Fader, C. M., Sola, C., and Colombo, M. I. (2010). Alpha-hemolysin is required for the activation of the autophagic pathway in Staphylococcus aureus-infected cells. Autophagy 6, 110–125. doi: 10.4161/auto.6.1.10698
Mitou, G., Budak, H., and Gozuacik, D. (2009). Techniques to study autophagy in plants. Int. J. Plant Genomics 2009:451357. doi: 10.1155/2009/451357
Mitroulis, I., Kourtzelis, I., Papadopoulos, V. P., Mimidis, K., Speletas, M., and Ritis, K. (2009). In vivo induction of the autophagic machinery in human bone marrow cells during Leishmania donovani complex infection. Parasitol. Int. 58, 475–477. doi: 10.1016/j.parint.2009.07.002
Nakagawa, I., Amano, A., Mizushima, N., Yamamoto, A., Yamaguchi, H., Kamimoto, T., et al. (2004). Autophagy defends cells against invading group A Streptococcus. Science 306, 1037–1040. doi: 10.1126/science.1103966
Nazarko, V. Y., Nazarko, T. Y., Farre, J. C., Stasyk, O. V., Warnecke, D., Ulaszewski, S., et al. (2011). Atg35, a micropexophagy-specific protein that regulates micropexophagic apparatus formation in Pichia pastoris. Autophagy 7, 375–385. doi: 10.4161/auto.7.4.14369
Niu, Q., Li, S., Chen, D., Chen, Q., and Chen, J. (2016). Iron acquisition in Leishmania and its crucial role in infection. Parasitology 143, 1347–1357. doi: 10.1017/S0031182016000858
Perinoto, A. C., Maki, R. M., Colhone, M. C., Santos, F. R., Migliaccio, V., Daghastanli, K. R., et al. (2010). Biosensors for efficient diagnosis of leishmaniasis: innovations in bioanalytics for a neglected disease. Anal. Chem. 82, 9763–9768. doi: 10.1021/ac101920t
Pinheiro, R. O., Nunes, M. P., Pinheiro, C. S., D’Avila, H., Bozza, P. T., Takiya, C. M., et al. (2009). Induction of autophagy correlates with increased parasite load of Leishmania amazonensis in BALB/c but not C57BL/6 macrophages. Microbes Infect. 11, 181–190. doi: 10.1016/j.micinf.2008.11.006
Real, F., and Mortara, R. A. (2012). The diverse and dynamic nature of Leishmania parasitophorous vacuoles studied by multidimensional imaging. PLoS Negl. Trop. Dis. 6:e1518. doi: 10.1371/journal.pntd.0001518
Russell, D. G., and Wilhelm, H. (1986). The involvement of the major surface glycoprotein (gp63) of Leishmania promastigotes in attachment to macrophages. J. Immunol. 136, 2613–2620.
Russell, D. G., Xu, S., and Chakraborty, P. (1992). Intracellular trafficking and the parasitophorous vacuole of Leishmania mexicana-infected macrophages. J. Cell Sci. 103(Pt 4), 1193–1210.
Santos, D. O., Coutinho, C. E., Madeira, M. F., Bottino, C. G., Vieira, R. T., Nascimento, S. B., et al. (2008). Leishmaniasis treatment–a challenge that remains: a review. Parasitol. Res. 103, 1–10. doi: 10.1007/s00436-008-0943-2
Schaible, U. E., Schlesinger, P. H., Steinberg, T. H., Mangel, W. F., Kobayashi, T., and Russell, D. G. (1999). Parasitophorous vacuoles of Leishmania mexicana acquire macromolecules from the host cell cytosol via two independent routes. J. Cell Sci. 112(Pt 5), 681–693.
Schnaith, A., Kashkar, H., Leggio, S. A., Addicks, K., Kronke, M., and Krut, O. (2007). Staphylococcus aureus subvert autophagy for induction of caspase-independent host cell death. J. Biol. Chem. 282, 2695–2706. doi: 10.1074/jbc.M609784200
Spath, G. F., and Beverley, S. M. (2001). A lipophosphoglycan-independent method for isolation of infective Leishmania metacyclic promastigotes by density gradient centrifugation. Exp. Parasitol. 99, 97–103. doi: 10.1006/expr.2001.4656
Suzuki, K., Kondo, C., Morimoto, M., and Ohsumi, Y. (2010). Selective transport of alpha-mannosidase by autophagic pathways: identification of a novel receptor, Atg34p. J. Biol. Chem. 285, 30019–30025. doi: 10.1074/jbc.M110.143511
Suzuki, K., and Ohsumi, Y. (2007). Molecular machinery of autophagosome formation in yeast, Saccharomyces cerevisiae. FEBS Lett. 581, 2156–2161. doi: 10.1016/j.febslet.2007.01.096
Tanida, I., Ueno, T., and Kominami, E. (2004). LC3 conjugation system in mammalian autophagy. Int. J. Biochem. Cell Biol. 36, 2503–2518. doi: 10.1016/j.biocel.2004.05.009
Thomas, S. A., Nandan, D., Kass, J., and Reiner, N. E. (2017). Countervailing, time-dependent effects on host autophagy promotes intracellular survival of Leishmania. J. Biol. Chem. 293, 2617–2630. doi: 10.1074/jbc.M117.808675
Vazquez, C. L., and Colombo, M. I. (2009). Assays to assess autophagy induction and fusion of autophagic vacuoles with a degradative compartment, using monodansylcadaverine (MDC) and DQ-BSA. Methods Enzymol. 452, 85–95. doi: 10.1016/S0076-6879(08)03606-9
Veras, P. S., de Chastellier, C., Moreau, M. F., Villiers, V., Thibon, M., Mattei, D., et al. (1994). Fusion between large phagocytic vesicles: targeting of yeast and other particulates to phagolysosomes that shelter the bacterium Coxiella burnetii or the protozoan Leishmania amazonensis in Chinese hamster ovary cells. J. Cell Sci. 107(Pt 11), 3065–3076.
Veras, P. S., Moulia, C., Dauguet, C., Tunis, C. T., Thibon, M., and Rabinovitch, M. (1995). Entry and survival of Leishmania amazonensis amastigotes within phagolysosome-like vacuoles that shelter Coxiella burnetii in Chinese hamster ovary cells. Infect. Immun. 63, 3502–3506.
Keywords: Leishmania, macrophages, autophagy, LC3, parasitophorous vacuole
Citation: Dias BRS, de Souza CS, Almeida NJ, Lima JGB, Fukutani KF, dos Santos TBS, França-Cost J, Brodskyn CI, de Menezes JPB, Colombo MI and Veras PST (2018) Autophagic Induction Greatly Enhances Leishmania major Intracellular Survival Compared to Leishmania amazonensis in CBA/j-Infected Macrophages. Front. Microbiol. 9:1890. doi: 10.3389/fmicb.2018.01890
Received: 25 March 2018; Accepted: 27 July 2018;
Published: 15 August 2018.
Edited by:
Celio Geraldo Freire-de-Lima, Universidade Federal do Rio de Janeiro, BrazilReviewed by:
Mario Alberto Flores-Valdez, Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco (CIATEJ - CONACYT), MexicoCopyright © 2018 Dias, de Souza, Almeida, Lima, Fukutani, dos Santos, França-Cost, Brodskyn, de Menezes, Colombo and Veras. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patricia S. T. Veras, cGF0cmljaWEudmVyYXNAZmlvY3J1ei5icg==; cHN0dmVyYXNAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.