
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 03 July 2018
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.01351
 Giselle C. de Oliveira Santos1†
Giselle C. de Oliveira Santos1† Cleydlenne C. Vasconcelos1†
Cleydlenne C. Vasconcelos1† Alberto J. O. Lopes2
Alberto J. O. Lopes2 Maria do S. de Sousa Cartágenes2
Maria do S. de Sousa Cartágenes2 Allan K. D. B. Filho3
Allan K. D. B. Filho3 Flávia R. F. do Nascimento4
Flávia R. F. do Nascimento4 Ricardo M. Ramos5
Ricardo M. Ramos5 Emygdia R. R. B. Pires6
Emygdia R. R. B. Pires6 Marcelo S. de Andrade2
Marcelo S. de Andrade2 Flaviane M. G. Rocha7
Flaviane M. G. Rocha7 Cristina de Andrade Monteiro7,8*
Cristina de Andrade Monteiro7,8*The Candida genus comprises opportunistic fungi that can become pathogenic when the immune system of the host fails. Candida albicans is the most important and prevalent species. Polyenes, fluoropyrimidines, echinocandins, and azoles are used as commercial antifungal agents to treat candidiasis. However, the presence of intrinsic and developed resistance against azole antifungals has been extensively documented among several Candida species. The advent of original and re-emergence of classical fungal diseases have occurred as a consequence of the development of the antifungal resistance phenomenon. In this way, the development of new satisfactory therapy for fungal diseases persists as a major challenge of present-day medicine. The design of original drugs from traditional medicines provides new promises in the modern clinic. The urgent need includes the development of alternative drugs that are more efficient and tolerant than those traditional already in use. The identification of new substances with potential antifungal effect at low concentrations or in combination is also a possibility. The present review briefly examines the infections caused by Candida species and focuses on the mechanisms of action associated with the traditional agents used to treat those infections, as well as the current understanding of the molecular basis of resistance development in these fungal species. In addition, this review describes some of the promising alternative molecules and/or substances that could be used as anticandidal agents, their mechanisms of action, and their use in combination with traditional drugs.
Candida species, opportunistic pathogens, are a major cause of morbidity and mortality worldwide and thus represents a serious threat to public health (Pfaller et al., 2014; Matthaiou et al., 2015; Pappas et al., 2016). Further, Candida species can cause vaginitis, oral candidiasis, cutaneous candidiasis, candidemia, and systemic infections (Wächtler et al., 2012). Candidemia is the most frequent hospital infection accounting for up to 15% of bloodstream infections, and Candida species are the main causative agents in 50–70% of systemic fungal infections (Cornely et al., 2012; Lionakis and Netea, 2013; Barchiesi et al., 2016).
Candida albicans is the pathogenic species most frequently isolated. However, other species such as C. glabrata, C. tropicalis, C. parapsilosis, C. krusei, C. famata, C. guilliermondii, and C. lusitaniae have been increasingly isolated, mainly in human immunodeficiency virus (HIV)-infected individuals (Brunke and Hube, 2013; Ferreira et al., 2013; Mayer et al., 2013; Patil et al., 2015; Barchiesi et al., 2016).
The pathogenesis of Candida species is poorly understood, and the rate of infections is increasing rapidly. Further, a steady increment in resistance to traditional antifungal has resulted in the need to control Candida infections through early diagnosis and prevention of candidiasis.
Among the available antifungal agents, azoles are the preferred and most frequently used drugs for treatment of Candida infections. Depending on the type of infection, the anatomical site in which it occurs and the sensitivity profile of species, other antifungals can also be used. Among these, there are polyenes, echinocandins, nucleoside analogs and allylamines (Pfaller et al., 2010a, 2013; Pappas et al., 2016). Fluconazole (FLZ), a type of azole, is often preferred in treatments of Candida infections because of its low cost and toxicity, in addition to availability in varied formulations (Pfaller et al., 2010b). However, there are many reports in the literature on the development of resistance among Candida species, especially in relation to azoles, which is essential for the determination of resistance mechanisms presented by fungi with the objective of developing new classes of antifungal for treatment of Candida infections.
The need of the hour includes the development of a more effective therapy, since the phenomenon of resistance caused the appearance of new fungal infections, in addition to facilitating the resurgence of the existing ones. In this way, the control of Candida infections is a challenge in the modern clinic. The design of new drugs from the traditional ones used in the clinic and the identification of new molecules with antifungal potential for the manufacture of new drugs, more effective and less toxic, are fundamental to face the challenge.
The present review examines infections caused by Candida species and describes our current understanding of the molecular basis of resistance development in these fungal species. In addition, this review describes some of the promising alternative molecules and/or substances that are effective pharmaceuticals for treating fungal infections and could be used as anticandidal agents, as well as their mechanisms of action.
Fungal infections are considered a serious health problem, especially in people with some impairment in the immune system and are a main cause of morbidity and mortality worldwide (Vallabhaneni et al., 2016). In the last two decades, fungal infections have shown a significant increment. This high incidence has been related to factors such as the increase in the number of patients with compromised immune system, (Ortega et al., 2010; Junqueira et al., 2012; Li et al., 2013; Terças A.L.G. et al., 2017), the increasing number of patients receiving hyperalimentation through catheters or probes and use of broad-spectrum antibiotics (Bouza and Munoz, 2008). The rising number of patients requiring organ transplantations, as well as those with leukemia and diabetes also contributes to this phenomenon (Razzaghi-Abyaneh et al., 2014).
The most frequent fungal disease affecting populations in the world is candidiasis (Lewis et al., 2012; Ferreira et al., 2013; Kwamin et al., 2013; Mayer et al., 2013; Tsai et al., 2013; Vázquez-González et al., 2013). There are several types of candidiasis as mucosal candidiasis, cutaneous candidiasis, onychomycosis and systemic candidiasis (Calderone and Fonzi, 2001; Kim and Sudbery, 2011; Wächtler et al., 2012). An important fact is that candidiasis is an infection that can affect both immunocompromised and healthy people (Li et al., 2006; Raman et al., 2013). Candidemia is another infection due Candida spp. and is the most relevant and prevalent nosocomial fungal infection associated with a high mortality rate (up to 49%) in patients with a compromised immune system (Pfaller and Diekema, 2007; Sardi et al., 2013). The association of Candida with bloodstream infections depends on patient’s condition, age, and geographic region. Candidemia is such an important infection that in 10–40% of cases it is associated with sepsis or septic shock while Candida species as main agent of sepsis or septic shock are responsible for no more than 5% of the total number of cases (Guery et al., 2009).
Many species recovered from human samples have been identified as belonging to the genus Candida that almost half has implicated in serious infections. C. albicans continues to be the most prevalent species, representing the majority of isolates of fungal infections (Delgado et al., 2009; Hise et al., 2009; Junqueira et al., 2012; Li et al., 2013; Terças A.L.G. et al., 2017). However, the prevalence of other Candida species has increase substantially. These species are C. parapsilosis, C. tropicalis, C. krusei, C. glabrata, C. guilliermondii, C. orthopsilosis, C. metapsilosis, C. famata, and C. lusitaniae (Sant’Ana Pde et al., 2002; Li et al., 2013; Kaur et al., 2016).
Candida albicans is a species that presents high degree of flexibility, being able to grow in extremely different environments regarding to the availability of nutrients, temperature variation, pH, osmolarity, and amount of available oxygen (Paramythiotou et al., 2014). This fact associated with the high resistance capacity of the species to antifungals, their virulent features, capability of forming biofilms with other species (Álvares et al., 2007; Shoham and Marr, 2012) make the genus Candida a serious risk to human health (Soll, 2008). Thus, Candida species are highly adaptable and possess numerous strategies to survive favors that might affect their overgrowth and change their susceptibility profiles.
In addition, it is difficult to identify specific Candida species, which may delay the use of precise therapeutics. For instance, microbiological tests using specific culture media do not differentiate many species of Candida. Often, it takes several days to obtain antifungal susceptibility information for Candida species (Clancy and Nguyen, 2013). Although there are newer molecular techniques available for rapid yeast detection, such as fluorescence in situ hybridization (PNA-FISH), commercially available equipment for analysis does not differentiate between C. albicans and C. parapsilosis, C. glabrata, or C. krusei. These facts are relevant because increased mortality rate is associated with delays in initiating adequate antifungal therapy (Garey et al., 2006; Bassetti et al., 2014).
Fungal cells, like human cells, are eukaryotic; both cell types are targeted by antifungal compounds, resulting in considerable side effects in patients and fewer available targets for drug action. Since the 1990s, there has been an increasing, but limited, discovery of antifungal agents (Sardi et al., 2013; Paramythiotou et al., 2014). These drugs include azoles, that inhibit ergosterol biosynthesis [FLZ, itraconazole (ITC), ketoconazole (KTC), miconazole and clotrimazole]; polyenes [amphotericin B (AMB) and nystatin]; allylamines; thiocarbamates; morpholines; 5-fluorocytosine, a deoxyribonucleic acid (DNA) analog; and echinocandins (for instance, caspofungins) (Pappas et al., 2009; Spampinato and Leonardi, 2013). Three cellular components of fungi are targeted by these drugs. In endoplasmic reticulum of the fungal cell, azoles inhibit ergosterol biosynthesis by interfering with the enzyme lanosterol 14-α-demethylase, involved in the transformation of lanosterol into ergosterol, component that is part of the plasma membrane structure of the fungus (Figure 1). Thus, accumulation of 14-α-methyl-3, 6-diol, a toxic compound, will occur. As the concentration of ergosterol is reduced, the cell membrane structure is altered, thereby inhibiting fungal growth (Sanguinetti et al., 2015).
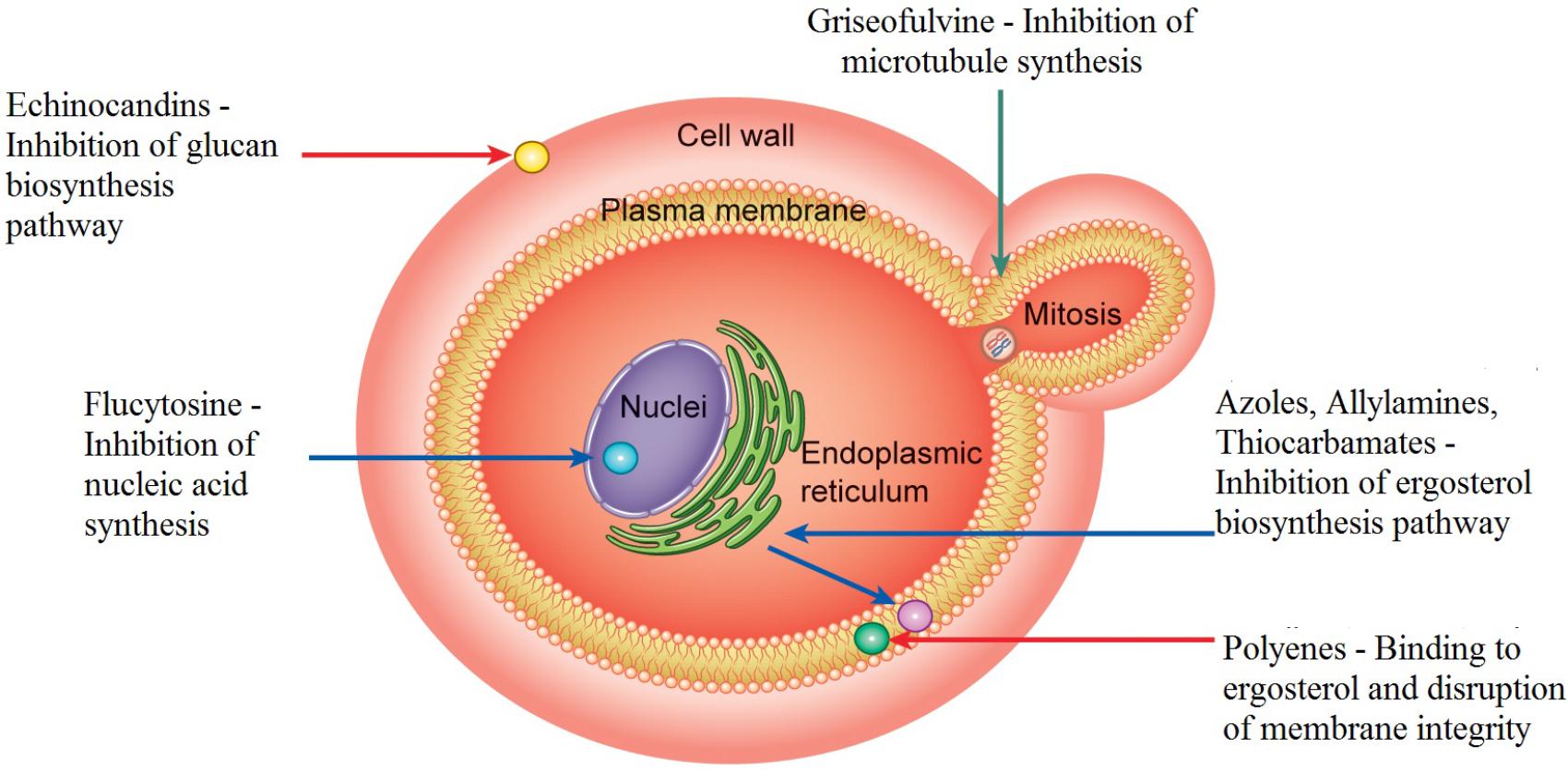
FIGURE 1. Mechanisms of action of traditional antifungal agents on cellular targets. Azoles inhibit the ergosterol synthesis in the endoplasmic reticulum of the fungal cell. They act by interfering with the enzyme lanosterol 14-α-demethylase, involved in the transformation of lanosterol into ergosterol. Polyenes act in the fungal membrane by binding to ergosterol and causing disruption of the membrane structure promoting extravasation of intracellular constituents and, consequently, cell death. Flucytosine inhibits the thymidylate-synthetase enzyme interfering with DNA. Echinocandins inhibit (1,3) β-D-glucan synthase, thereby preventing glucan synthesis, which is present in the cell membrane of fungi. Allylamines and thiocarbamates inhibit the enzyme squalene-epoxidase, which participates in the synthesis of ergosterol. Griseofulvin acts by disrupting spindle and cytoplasmic microtubule production, thereby, inhibiting fungal mitosis.
Azoles comprise a 5-member azole ring containing two (imidazole) or three nitrogen atoms (triazole) attached to a complex side chain (Georgopapadakou, 1998; Groll et al., 2003). The azole family of compounds includes the imidazoles (KTC, miconazole, econazole and clotrimazole) and triazoles (FLZ, ITC, and voriconazole, which is a synthetic triazole derivative of FLZ of second-generation), and posaconazole (hydroxylated analog of itraconazole) (Kontoyiannis et al., 2003; Maubon et al., 2014). Ergosterol is similar to cholesterol present in plasma membrane of animal cells; however, most antimycotic agents that target ergosterol binding or synthesis do not cross-react with host cells because of sufficient structural differences between these molecules (Spampinato and Leonardi, 2013).
Amphotericin B and others polyenes act in fungal membrane by binding to ergosterol and causing disruption of the membrane structure, which promotes extravasation of intracellular constituents such as potassium, magnesium, and sugars and, consequently, cell death (Perman et al., 2009; Mesa-Arango et al., 2012; Figure 1).
Flucytosine (5-FC) is a pyrimidine analog with fungistatic properties that enters the fungal cell through cytosine permease and inhibits the thymidylate-synthetase enzyme interfering with DNA. 5-FC can also be converted to 5-fluorouracil which in turn can be phosphorylated to 5-fluorodeoxyuridine monophosphate. This one being also phosphorylated can be incorporated into RNA molecules, thus interfering with the cell translation process (Cuenca-Estrella, 2010; Spampinato and Leonardi, 2013; Maubon et al., 2014; Figure 1). Further, 5-fluorodeoxyuridine is associated with considerable toxicity (Patil et al., 2015). Clinical use of 5-FC is preferred in association with AMB (Sanglard et al., 2009; Nett and Andes, 2016; Prasad et al., 2016), since the use of 5-FC alone induces stronger side-effects, such as hepatic impairment, interference with bone marrow function, and rapid occurrence of resistance especially among Candida species (Groll et al., 1998; Nett and Andes, 2016; Prasad et al., 2016).
According to the type of infection and the sensitivity/resistance profile of the isolates, as well as the site of origin of the samples all antifungal agents may be used with varying efficiency (Pfaller et al., 2010a). AMB is considered the gold standard drug for most mycoses that affect patients at risk (Mesa-Arango et al., 2012). However, AMB has high toxicity, which limits its use. Nephrotoxicity is a main effect resulting from AMB administration (Mesa-Arango et al., 2012; Nett and Andes, 2016). To minimize this problem and to increase the effectiveness of treatment, some formulations have been developed. Liposomal AMB (Ambisome®) allows for lower absorption of AMB by the reticuloendothelial system, which results in greater permanence in the bloodstream. A lipid complex of AMB (Abelcet®) comprises 50% AMB and 50% lipid compound and AMB in a colloidal dispersion (Amphocil®/Amphotech®, formed by a stable complex with cholesterol sulfate). However, the high cost of these formulations has limited their use (Kontoyiannis et al., 2003; Mesa-Arango et al., 2012; Paramythiotou et al., 2014; Nett and Andes, 2016).
Azoles represent the class of antifungals with the highest number of drugs. Azoles have fungistatic properties that affect cell growth and proliferation; a large amount of accumulated toxic sterols can eventually lead to fungal cell death (Shapiro et al., 2011; Prasad et al., 2016). Such agents are preferred in the treatment of candidemia (Spampinato and Leonardi, 2013; Maubon et al., 2014) and candidiasis. Among azoles, miconazole and KTC (imidazoles) first emerged and were the only drugs available for systemic use, with KTC being the first alternative to AMB (Groll et al., 1998; Seyedmousavi et al., 2017). Then triazoles as itraconazole and FLZ have emerged, more effective and better tolerated than KTC (Dismukes, 2000).
Fluconazole is the drug of choice for most Candida infections (Pfaller et al., 2010b; Patil et al., 2015) and is the most recommended antifungal agent, attributable to its low cost, for use in invasive candidiasis in patients who have not previously been medicated with azole antifungal agents (Shoham and Marr, 2012; Paramythiotou et al., 2014).
Azole antifungals have limitations to their use, although they are generally well-tolerated. Limitations include adverse effects such as hepatotoxicity and the emergence of resistance among fungal isolates (Carrillo-Muñoz et al., 2006). Azoles can be toxic because they act as substrates or inhibitors of several enzymes such as cytochrome P450 enzymes. Further, these limitations provide motivation for improving this class of antifungal agents (Nett and Andes, 2016).
Alterations in triazole molecule gave rise to voriconazole (structurally related to FLZ) and posaconazole (related to ITC), a second generation of antifungals. Both are available for systemic therapy and have been shown to have better specificity and antifungal potency than that of first generation triazoles (Nett and Andes, 2016).
A new class of drugs, the echinocandins, has been shown to have fungicidal effects in all Candida species (Nett and Andes, 2016). The echinocandins include caspofungin, micafungin, and anidulafungin (Grossman et al., 2014; Koehler et al., 2014; Paramythiotou et al., 2014). Echinocandins inhibit (1,3) β-D-glucan synthase, thereby preventing glucan synthesis, which is present in the cell membrane of fungi (Figure 1). β-D-Glucan synthase inhibition depletes glucan polymers in fungal cells, resulting in an abnormal cell wall that is weak and unable to resist osmotic stress (Chen and Sorrel, 2007; Kuse et al., 2007). This class of drugs has certain advantages attributable to its effects on the fungal cell wall, including a lower risk of side effects since animal cells do not have this structure. Further, echinocandins can be used in cases of azole-antifungal resistance (Spampinato and Leonardi, 2013; Grossman et al., 2014; Maubon et al., 2014; Paramythiotou et al., 2014).
Allylamines (terbinafine and naftifine) and thiocarbamates inhibit the enzyme squalene-epoxidase, which participates in the synthesis of ergosterol and is encoded by the ERG1 gene (Figure 1). Inhibition of squalene-epoxidase leads to membrane rupture and accumulation of squalene. Allylamines effects can also prevent the production of other sterol derivatives.
Another antifungal is the tricyclic spirodiketone griseofulvin that acts by interfering the cytoplasmic microtubule production, disrupting spindle formation and, thereby inhibiting fungal mitosis (Figure 1). Griseofulvin was isolated from Penicillium griseofulvum, (Francois et al., 2005).
Certain pharmacological strategies have been developed to minimize toxicity and resistance. Development and use of new antifungal formulas (liposomal AMB, AMB lipid complexes, AMB colloidal dispersions, and AMB lipid nanosphere formulations), itraconazole, and β-cyclodextrin itraconazole is one strategy. Others include combination therapies of antifungal compounds (for example, AMB + 5-FC, FLZ + 5-FC, AMB + FLZ, caspofungin + liposomal AMB, and caspofungin + FLZ) (Table 1) and nanostructuring of conventional antifungal agents (Amaral and Felipe, 2013; Spampinato and Leonardi, 2013; Stiufiuc et al., 2015; Souza and Amaral, 2017).
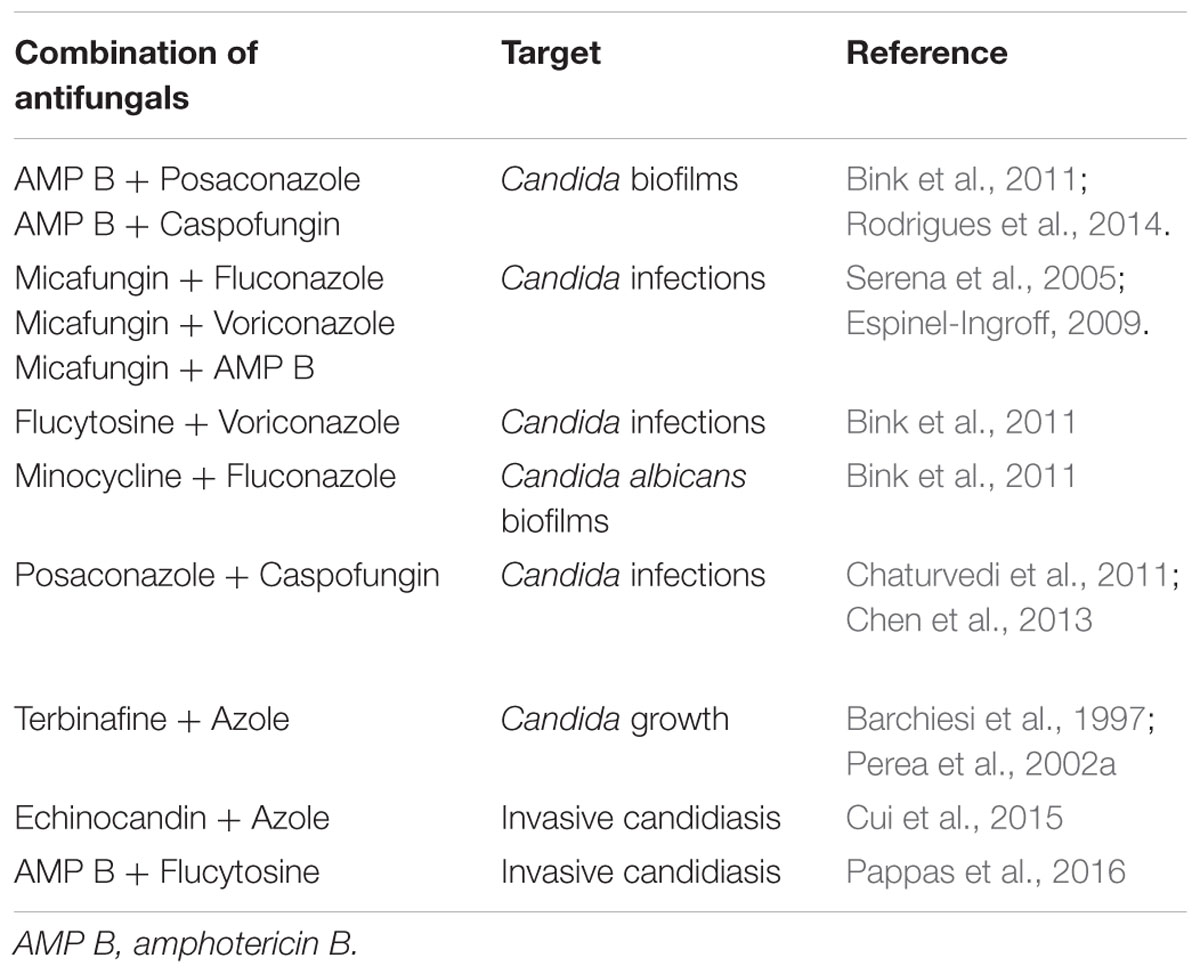
TABLE 1. Various regimes of combinatorial antifungal therapy showing better efficacy in combination than that of independent drugs.
Chaturvedi et al. (2011) evaluated the sensitivity profile of reference and clinical samples of C. albicans, C. glabrata, and C. parapsilosis in relation to antifungals like azoles and equinocandinas. They found that despites clinical isolates had relatively high azole and echinocandin MICs, some synergistic combinations were found for AMB- posaconazole against C. glabrata and AMB- anidulafungin and AMB- caspofungin against C. parapsilosis by both visual and spectrophotometric readings. Chen et al. (2013) a potential therapeutic applicability for posaconazole and caspofungin combinations in the future. Their studies reported that posaconazole exhibits in vitro and in vivo synergy with caspofungin against drug susceptible or resistant C. albicans strains (derived echinocandin-resistant mutants).
Pappas et al. (2016) drew attention to the fact that a combination of liposomal AmB, 5 mg/kg daily, and flucytosine, 25 mg/kg 4 times daily, may be considered as salvage therapy in patients who have not had a clinical response to initial AmB therapy in cases of central nervous system infections by fungus in neonates.
Examples of some others publications on combination therapy between different antifungal drugs are shown in Table 1.
However, among traditional antimycotic drugs, none has all the qualities required for an ideal agent (Wong et al., 2014). All drugs have at least one of the following restrictions: they do not have a broad spectrum of action, some are fungistatic, and others have high toxicity and low bioavailability with significant side effects in patients undergoing therapeutic regimens (Petrikkos and Skiada, 2007; Safdar et al., 2010; Lewis et al., 2012; Vollenbroich et al., 2014; Bayhan et al., 2015). Therefore, limitations of treatment and drug resistance (Canuto and Rodero, 2002; Petrikkos and Skiada, 2007; Mukherjee et al., 2011; Tscherner et al., 2011) associated with pathogenicity of the clinical isolates support the urgent need to identify substances that are more effective, with new mechanisms of action in the fight against Candida infections.
There are three types of antifungal resistance, including a primary or intrinsic form that exists prior to antifungal exposure. The second type is an acquired form that occurs after antifungal exposure and may be reversible, attributable to transient or non-reversible adaptation resulting from several genetic alterations. A clinical form refers to unfavorable outcomes in patients despite antifungal therapy and is directly linked to primary or secondary resistance (Cowen et al., 2015).
Populations are increasingly at risk of fungal infections, resulting in an increased use of antifungal agents. Consequently, higher minimum inhibitory concentrations (MIC) for antifungals against C. albicans strains have been observed and may be related to therapeutic failures. In addition, some non-albicans Candida (NAC) species have inherent resistance to azoles (Oxman et al., 2010; Lortholary et al., 2011; Fothergill et al., 2014). Low-dose prophylactic administration of azole derivatives, such as FLZ, for prolonged periods to prevent the occurrence of opportunistic infections in immunosuppressed patients also results in resistant phenotypes (Siikala et al., 2010; Rautemaa and Ramage, 2011). These facts are likely to collaborate to the increased incidence of fungal infections. Resistance to polyenes (AMB) in C. albicans is less common and is associated with the substitution of ergosterol with a precursor molecule or a general reduction of sterols in the plasma membrane (Kanafani and Perfect, 2008) (Figure 2). Enzymes such as Δ5,6-desaturase, encoded by ERG3 gene, and C-8 sterol isomerase, encoded by ERG2 gene participate in ergosterol biosynthesis and have the main alterations related to AMB resistance. These enzymes influence the amount of ergosterol required for the action of polyenes because the mutations are responsible for modifications in sterol content (Sheikh et al., 2013). For instance, Δ5,6-desaturase when mutated converts fecosterol to episterol which has low affinity for AMB. Another likely AMB-resistant mechanism is the reduction of oxidative damage via enhanced catalase activity (Kanafani and Perfect, 2008).
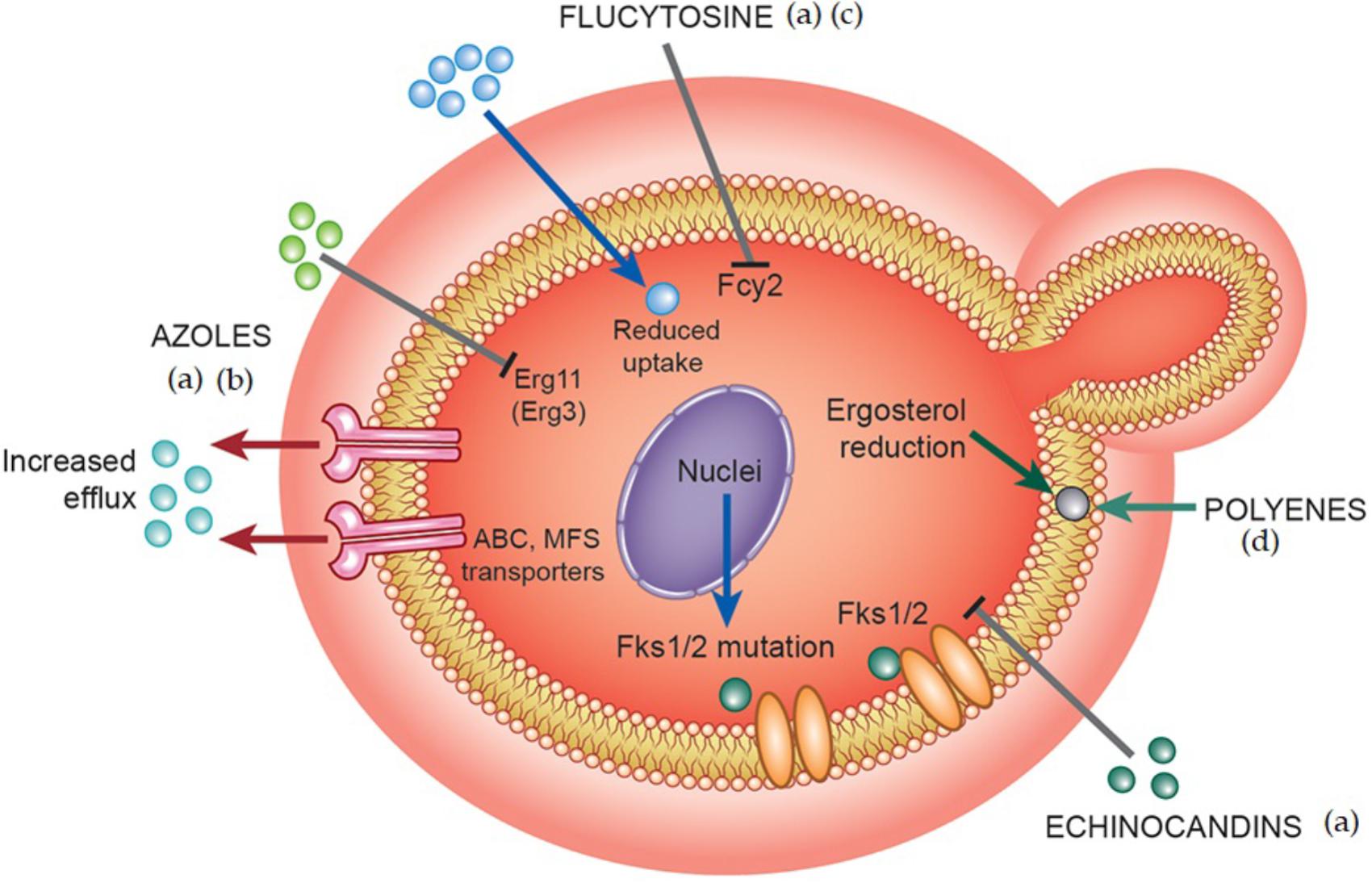
FIGURE 2. Different mechanisms of multidrug resistance adopted by fungal cells. The main mechanisms of drug resistance against azoles, polyenes, echinocandins, and flucytosine include: (a) alteration of the enzyme target (genes encoding ergosterol biosynthetic pathway enzymes – AZOLES, glucan synthases – ECHINOCANDINS, cytosine deaminase or uracil phosphoribosyltransferase – FLUCYTOSINE) leading to poor binding of toxic drugs to its enzyme target sites, (b) overexpression of drug efflux proteins leading to increased efflux – AZOLES, (c) changes in membrane property/composition affecting normal drug import – FLUCYTOSINE, (d) Reduction of sterols in plasma membrane – POLYENES. Colored balls mean antifungal molecules.
Possible mechanisms for cellular and molecular resistance to FLZ in C. albicans are described. The first is related to induction of multi-drug pumps, which decrease the concentration of drug available for the target enzyme, 14-α-demethylase, in fungal cells (Kanafani and Perfect, 2008) (Figure 2). There are two types of active transporters in C. albicans, including those encoded by the Candida drug resistance-CDR genes (Cdr1 and Cdr2) and those encoded by the multidrug resistance-MDR1 genes. Cdr1- and Cdr2-type pumps are ATP-binding cassette (ABC) transporters, and Mdr1 is a major facilitator superfamily (MFS)-type pump that transports solutes from different sides of the fungal cell plasma membrane. Overexpression of transporters encoded by CDR genes confers cross-resistance to various azole-derived compounds, while overexpression of those transporters encoded by MDR1 genes is responsible for FLZ resistance. Superexpression of these transporters prevents accumulation of the drug in the intracellular compartment (Kanafani and Perfect, 2008; Pfaller, 2012) (Figure 2).
A second mechanism of resistance involves modification of the target enzyme encoded by the ERG11 gene, also known as cytochrome P450 lanosterol 14 α-demethylase (Cyp51) (Figure 2). Mutations in this gene prevent azoles from binding to enzyme sites (Marichal et al., 1999; Flowers et al., 2015). Another mechanism of resistance to azoles is related to substitution of ergosterol by another sterol. Mutations in the ERG3 gene does not convert 14-α-methylfecosterol into 14-α-methyl-3,6-diol. This substitution causes azoles to have no fungistatic effects on the fungal cell membrane (Sanguinetti et al., 2015).
Two possible mechanisms of resistance to echinocandins have been reported. The first deals with point mutations in gene that encodes the major subunit of the glucan synthase enzyme (Fks subunit) (Figure 2). These mutations occur only in two regions of the gene (known as “hot-spot”), are dominants and can provide resistance to all echinocandin (Perlin, 2015). The consequence is that these mutations lead to the production of high MIC values. For instance, C. parapsilosis and C. guilliermondii present MIC values 4- to 100-fold greater compared to those observed for C. albicans. The FKS1 mechanism extends to other NAC species such as C. tropicalis, C. parapsilosis, C. glabrata, C. krusei, C. guilliermondii, and C. dubliniensis that show the same mutations as those of C. albicans (Katiyar et al., 2006; Perlin, 2011). In that way, FKS1-mediated resistance mechanisms can be pervasive in the fungal kingdom because it is suggested that it can be responsible for a reduced inherit sensitivity of certain Candida species and molds (Perlin, 2007).
The second mechanism of resistance involves the response to adaptation stress. When there is an inhibition of production of fungal cell wall component, microorganism is capable of increasing the production of another one. Some research showed that many Candida species respond to the inhibition of Fks synthesis producing high amounts of chitin (Chamilos et al., 2007; Shields et al., 2011). Chamilos et al. (2007), studying Candida bloodstream isolates from cancer patients, observed a process known as the paradoxical effect in some isolates of Candida, that is, isolates are capable of growing in the presence of high concentrations of echinocandins (above the MIC). This phenomenon was strikingly absent in C. glabrata isolates, but was well evidenced in C. parapsilosis, C. tropicalis, and C. krusei ones (Chamilos et al., 2007).
Resistance to 5-FC can be of two types: primary, occurring via cytosine permease (encoded by the FCY2 gene) with decreased drug uptake (Sanglard and Odds, 2002) (Figure 2); secondary, related to alterations in cytosine deaminase (encoded by FCY1) or uracil phosphoribosyltransferase (encoded by FUR1) enzymes activities. Cytosine permease is responsible by conversion of 5-FC to 5-fluorouridine or to 5-fluorouridine monophosphate (5-FUMP) (Kontoyiannis and Lewis, 2002; Espinel-Ingroff, 2008) Resistance is easily developed in fungal isolates from patients who are receiving the drug. However, most of these mechanisms have only been observed in others species of Candida, but not in C. albicans (Papon et al., 2007). Therefore, other molecular mechanisms related to resistance to 5-CF must exist, playing relevant role in fungal resistance (Schwarzmuller et al., 2014).
Costa et al. (2015) recently reported a relation between arginine metabolic enzymes and 5-FC resistance, suggesting that 5-FC resistance requires somehow high arginine production. Accordingly, L-arginine concentrations in some body fluids in healthy individuals, as vaginal fluid (Gregoire et al., 1959) or human plasma (Armengou et al., 2003) can be as high as 0.1 mM. The molecular mechanisms involved in arginine and 5-FC resistance relationship have not been clarified, but the results obtained by the authors highlight the significance of a new possibility of fighting resistance to 5-FC.
Studies aimed at elucidating the molecular mechanisms responsible for developing resistance to Candida species have predominantly focused on resistance to azoles, as they are the most commonly used drugs clinically (Cernicka and Subik, 2006; Gualco et al., 2007). Thus, the main research targets include the CDR1, CDR2, and MDR1 genes (Puri et al., 1999; Yang and Lo, 2001; Morschhäuser et al., 2007; Tsao et al., 2009). In addition, mutations in transcription factors associated with the CDR1 and CDR2 genes (Chen et al., 2004; Coste et al., 2006; Wang et al., 2006), specific mutations or superexpression of ERG11 genes, and mutations in the ERG5 or ERG3 genes (also involved in ergosterol biosynthesis), have been identified. Most of them were related to FLZ resistance (Sanglard et al., 2003; Lo et al., 2005; Martel et al., 2010).
An important and potent mechanism of multi-drug resistance (MDR) in fungi is the intracellular accumulation of antifungals by increased efflux of drugs (Prasad and Kapoor, 2005; Prasad and Rawal, 2014). In C. albicans, overexpression of genes encoding transporters proteins, mainly CDR1 and CDR2 genes (encoding Cdr1 and Cdr2, respectively, which are ABC multidrug transporter proteins) or MDR1 gene (encoding the MFS efflux pump protein Mdr1) is considered the main mechanism responsible for antimycotic resistance in Candida isolates (Franz et al., 1998; Lopez-Ribot et al., 1998; Lyons and White, 2000; Wirsching et al., 2000; Kusch et al., 2004; Niimi et al., 2004; Prasad and Kapoor, 2005; Prasad and Rawal, 2014). Increased production of the Cdr1 transporter is responsible for FLZ, KTC, and ITC resistance. In contrast, expression of the Candida drug resistance protein 2 (Cdr2p) is related to FLZ and ketoconazole resistance, but does not affect resistance to itraconazole (Tsao et al., 2009). There are related multidrug transporters with MDR function in NAC species such as those in C. glabrata (CgCdr1, CgCdr2 and Snq2) (Miyazaki et al., 1998; Sanglard et al., 1999; Torelli et al., 2008) and in C. krusei (ABC1) (Katiyar and Edlind, 2001).
Azole resistance in C. glabrata probably is also related to upregulation of homologous transporter genes CgCDR1 and CgCDR2 (Sanglard et al., 1999, 2001; Bennett et al., 2004), and genetic evidence has been provided that supports a role for multidrug transporters in azole resistance in C. glabrata (Sanglard et al., 1999).
Looi et al. (2005) investigated expression of the C. albicans and C. glabrata CDR1 and MDR1 genes associated with azole resistance in patients with vaginitis. There was overexpression of genes to varying extents in all Candida isolates tested and this result was correlated with the degree of resistance, as evidenced by antifungals MICs. The authors also observed that in one C. albicans resistant isolate there was overexpression of messenger RNA for Mdr1 after superexpression of Cdr1, which suggests a synergism between these drug efflux pumps proteins. DNA sequence analysis of the CDR1 promoter region also suggests there are several point mutations in resistant clinical isolates that are not present in susceptible isolates. Thus, this region is important for binding of transcription factors and for increasing the affinity of activators responsible by CDR1 expression in drug resistant isolates.
Sanguinetti et al. (2005) evaluated the molecular mechanisms of resistance in 29 nosocomial isolates of C. glabrata recovered during 3 years of study; of these, most were resistant to FLZ. Quantitative real-time PCR analyses provided evidence that azole resistance in these isolates probably was due the upregulation of genes CgCDR1, CgCDR2, and CgSNQ2, encoding efflux proteins in C. glabrata.
Katiyar and Edlind (2001) identified two homologous ABC transporter genes (ABC1 and ABC2) in C. krusei previously described for C. albicans. When cultures of C. krusei were exposed to imidazole and cycloheximide, ABC1 gene was upregulated. Lamping et al. (2009) showed that ABC1 is involved in the inherent resistance of C. krusei to azoles. In C. parapsilosis authors have shown that MRR1 is involved in resistance to FLZ (Souza et al., 2015; Zhang et al., 2015); however, in C. tropicalis, efflux pumps genes related to azoles resistance have not yet been identified (Barchiesi et al., 2000).
Gołąbek et al. (2015) also studied the expression of CDR1, CDR2, and MDR1 genes in 120 strains of C. albicans (60 resistant and 60 azole susceptible) obtained from clinical samples and observed that the expression of Cdr1, Cdr2, and Mdr1 was higher in azole-resistant strains than that in sensitive strains. Several transcription factors have been identified as responsible for upregulating CDR1, CDR2, and MDR1 genes, with several serving as positive MDR1 regulators (Coste et al., 2004; Wang et al., 2006) and others as negative MDR1 regulators (Chen et al., 2009).
Overexpression of the CDR1 and CDR2 genes has been suggested to influence the relationship between susceptibility and resistance to azoles and AMB. According to Ren et al. (2014), azole-resistant strains of C. albicans that overexpress CDR1 and CDR2 are hypersensitive to AMB. In contrast, knockout strains for the CDR1 and CDR2 genes are resistant to AMB, suggesting that the ergosterol content determines sensitivity to both azoles and AMB in C. albicans, and that there is an inverse susceptibility to these drugs that is directly associated with Cdr1 and Cdr2 transporters. The authors also suggest a new therapeutic approach for administering alanine phosphoricin B in situations of fungal resistance to azoles rather than increasing the administered dose of the azole agent.
Major facilitator superfamily (MFS) transporters were first identified in C. albicans and are the second major superfamily of transporters also related to drug efflux in this species (Saier et al., 1999; Gaur et al., 2008). MFS proteins consist of one polypeptide chain two three-dimensional regions with independent functions (“domains”) having six transmembrane alpha helical spanners (TMSs). There are two types of MFS proteins, including DHA1 (drug:HC antiporter-1) with 12 TMSs, and DHA2, which has 14 TMSs. The main multidrug protein of C. albicans is MDR1 from the DHA1 subfamily. Homologs of CaMDR1 were identified in C. glabrata (CgMDR1) and in C. dubliniensis (CdMDR1). CgMDR1 is constitutively expressed and confers specific resistance to FLZ in C. glabrata; therefore, this phenomenon could explain the intrinsic resistance of this yeast to triazoles (Moran et al., 1998; Sanglard et al., 1999). In C. dubliniensis overexpression of CdMDR1 is considered a main mechanism of FLZ resistance in isolates of this species (Moran et al., 1998).
In C. albicans, a 1587 bp gene encodes Erg11 protein. Erg11 has 595 amino acids. Up to now, approximately more than a hundred non-synonymous point mutations have been identified in clinically resistant isolates (Noel, 2012; Strzelczyk et al., 2013). Interestingly, most of these substitutions occur in 3 regions ranging from 105 to 165, 266 to 287, and 405 to 488 amino acids (“hot spots”) instead of being randomly dispersed (Marichal et al., 1999; Wang et al., 2009). However, many of the identified mutations are not resistant and are considered genetic polymorphisms occurring in living organisms. Few of these mutations have been demonstrated to support azole resistance (Noel, 2012; Strzelczyk et al., 2013). ERG11 genetic polymorphisms should be considered in the rational design of novel azole-derived drugs, attributable to certain polymorphisms identified in the gene that do not necessarily characterize changes in the amino acids and three-dimensional structure of proteins, and therefore, do not reduce the affinity between azolic components and the protein. Thus, mapping all ERG11 amino acid changes involved in azole resistance could help in the design of new azole antifungals with potent activity against resistant strains.
Recent studies corroborate these facts. For instance, Morio et al. (2010) investigated the susceptibility of FLZ, ITC and voriconazole in isolates of C. albicans and verified 23 distinct substitutions, 2 of which were suspected as being involved in azole resistance. Gołąbek et al. (2015) verified 19 changes in the ERG11 gene sequence and found that five alterations occurred in azole resistant strains only (A530C, G622A, G1309A, A1167G, and A1230G). Further, 33% of azole-resistant strains were characterized by the simultaneous presence of the A530C, G622A, and A1167G mutations. Caban et al. (2016) identified 21 specific mutations in the ERG11 gene, two of which were significantly associated with drug resistance, including a nucleotide substitution at position 798, which was related to an increase in drug resistance, and a silence mutation at position 1440, which significantly decreased the chance of a strain being resistant to drugs.
ERG11 mutations conferring azole resistance in C. tropicalis (Vandeputte et al., 2005; Jiang et al., 2013), C. krusei (Ricardo et al., 2014), C. dubliniensis (Perea et al., 2002b), and C. parapsilosis (Grossman et al., 2015) clinical isolates have also been described; however, there is no relate about these mutations in C. glabrata (Gonçalves et al., 2016).
In addition to mutations that directly affect the ERG11 gene, there are changes in transcription factors associated with this gene that may also affect its expression and, consequently, the biosynthesis pathway of ergosterol. The C. albicans Upc2p transcription factor (CAUPC2) gene is among the regulators of ERG11 gene expression.
It is known from the literature that overexpression of ERG11 gene, or maybe its upregulation, are responsible for the resistance to azole agents. Accordingly, different methods have been used to measure the level of ERG11 expression by detecting and quantifying its mRNA. A 3- to 20-fold increase in mRNA production was observed in resistant strains (Sanguinetti et al., 2015).
Two independent mechanisms have been shown to drive ERG11 overexpression. One is related to a chromosomal mutation (duplication) of ERG11 gene. This phenomenon was demonstrated first in C. glabrata isolates (Marichal et al., 1997); this occurs when an isochromosome is formed in the region having the ERG11 gene. In that way, this specific region will now have two copies of the left arm of chromosome 5, duplicating the chromosome (Selmecki et al., 2006). The second depends on a transcription factor that regulates the ergosterol biosynthesis (Upc2p, coded by the CaUPC2 gene) identified in C. albicans. Upc2p recognizes and is specifically bound to the promoters (the well-known SRE box, of sterol response element) of different ERG genes that activate gene transcription (Noel, 2012).
Antifungal drugs, including FLZ, induce the expression of CaUPC2 gene, and the Upc2p transcription factor upregulates ERG2 and ERG11 gene expression when C. albicans is grown under azole drug pressure (Allen et al., 2015). Studies have shown that strains that have undergone homozygous deletion of the CaUPC2 gene are hypersensitive to several drugs and accumulate significantly less cholesterol, suggesting a decrease in ergosterol in these strains (Silver et al., 2004).
Different strains of C. albicans and probably other Candida species express different UPC2 alleles encoding for transcription factors of different strength. In addition, three gain of function mutations (A643T, A643V, and G648D) have been characterized in sequential clinical isolates overexpressing ERG11. Upc2 proteins act constitutively and lead to loss of sensitivity to azoles (Noel, 2012).
Increased drug resistance in fungi is a problem that cannot be avoided, particularly for FLZ, which is the preferred antifungal for treating candidiasis in acquired immunodeficiency syndrome (AIDS) patients (Siikala et al., 2010; Rautemaa and Ramage, 2011). Moreover, there are fungi that have intrinsic resistant to antifungal agents commonly used in the clinic (Sanglard, 2016). In addition, biofilms, an ordinary virulence property of fungi, has as main characteristic the capacity of resistance to drugs (Chandra et al., 2005; Seneviratne et al., 2008).
Some peptides isolate from various sources of body have antimicrobial properties and are also a promise in the discovery of new antimycotics. One possibility is to use molecules with antifungal properties derived from host cells to prevent or treat fungal infections (Table 2). There are some small cationic peptides derived from large proteins that exert antifungal activities (Table 3). The main mechanism of action related to these peptides is that they intensify the passage of substances through the fungal membrane favoring permeabilization. These peptides include lysozyme, lactoferrin, defensins, histatin, and cathelicidins (Mehra et al., 2012) (Figure 3).
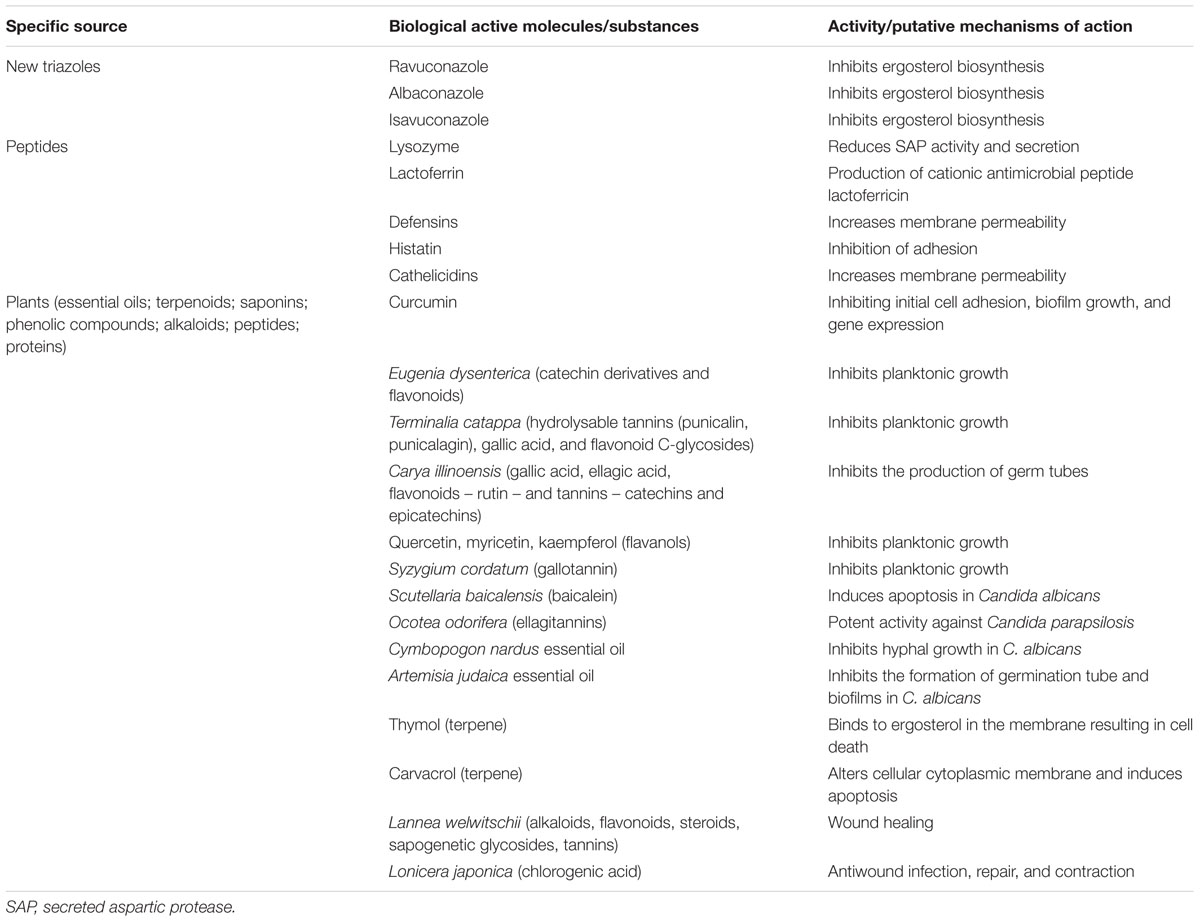
TABLE 2. Alternative products with reported antifungal activities against Candida species showing promise for antifungal drug development.
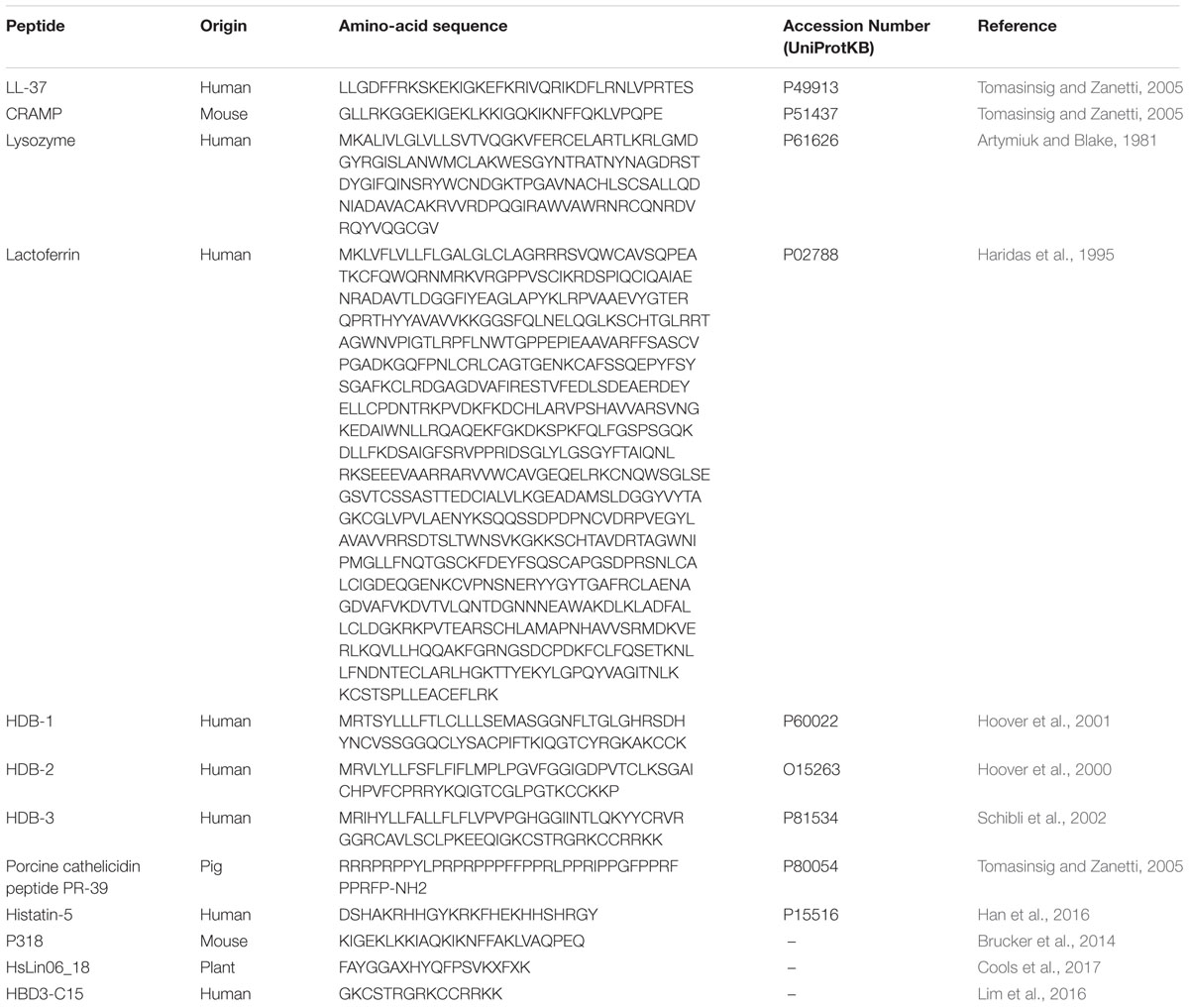
TABLE 3. Predicted amino-acid sequences (single-letter code) of antimicrobial peptides obtained from Protein Data Bank (RCSB-PDB) or from literature reference.
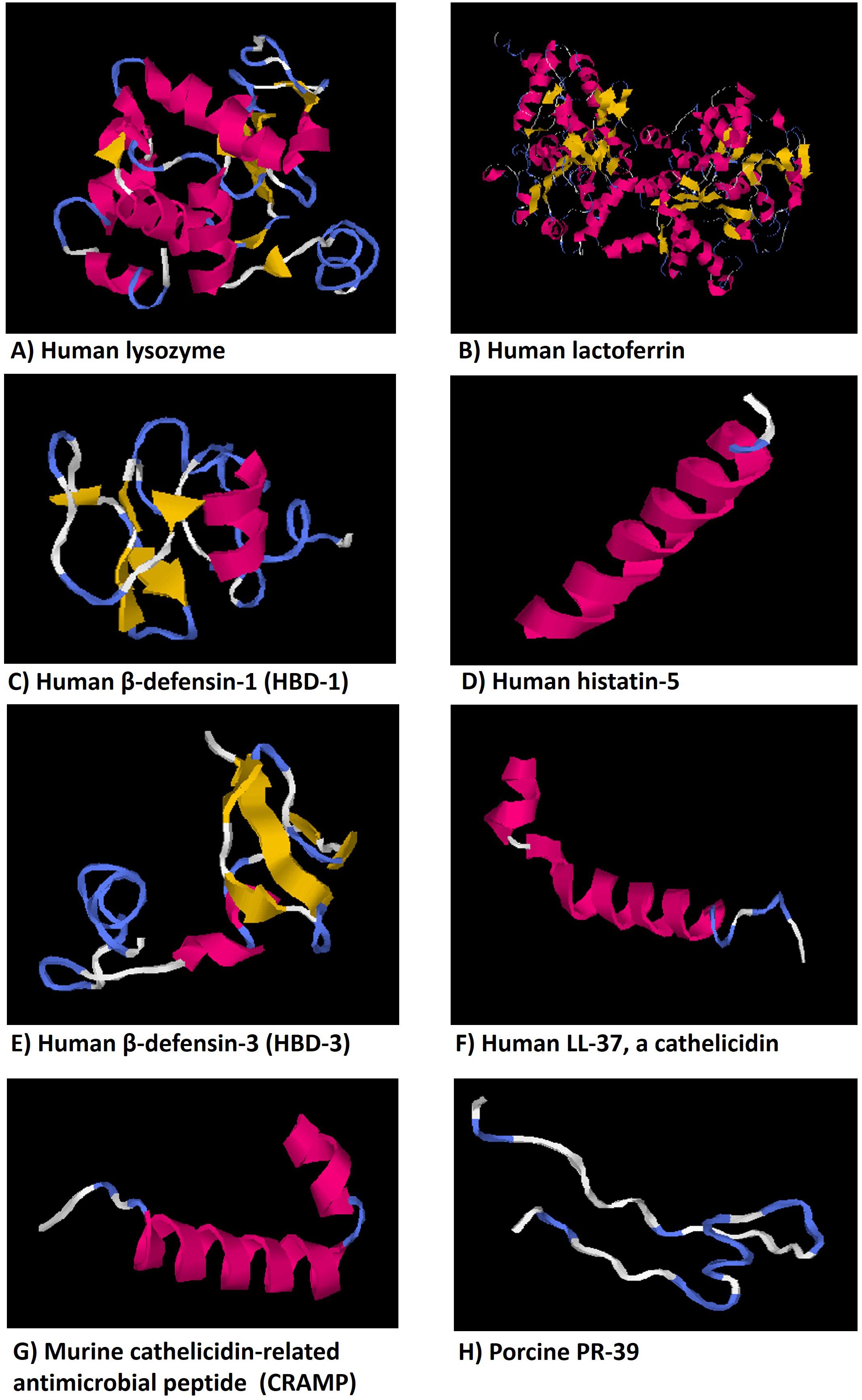
FIGURE 3. Representative models of some antifungical peptides showing alpha-helical, beta-sheet, amphiphilic structural motifs that relate forms and functions. The three-dimensional structure models of peptides were predicted using the on-line server Iterative Threading ASSEmbly Refinement (I-TASSER, Yang Zhang Lab), University of Michigan (Ann Arbor, MI, United States), from their amino acid sequences.
Accordingly, the utmost relevance in the health care field is the development of more safe and effective antimycotic agents. Therefore, this topic aimed to address new substances and/or molecules with potential antifungal activity, their clinical relevance, and mechanisms of actions. Alternative therapies with some antifungal potential include the use of synthetic agents, polymeric materials, active molecules of natural products and peptides (Table 2).
New triazoles antifungals are being developed and are under investigation, due to the limited number and the lack of effective antifungal. These include ravuconazole, albaconazole, and isavuconazole (Table 2). Preliminary studies have shown that these drugs have good pharmacokinetic profiles and low toxicity, as well as in vitro activity against Candida even in FLZ-resistant strains, with similar properties to those of FLZ and voriconazole.
Albaconazole, a broad-spectrum antifungal agent with excellent tolerability (Bartroli and Merlos, 2011), has shown great activity against Candida spp., both in vitro and in vivo studies, with better properties than those of FLZ. In fact, a single dose of albaconazole at a concentration almost 4× lower than that of FLZ recommended for acute vaginal candidiase has better efficacy (Pasqualotto and Denning, 2008). In addition, albaconazole showed low toxicity when administered to volunteer patients (Girmenia, 2009). Most isolates of C. albicans and C. glabrata are susceptible to treatment with albaconazole (Pasqualotto et al., 2010).
Isavuconazole is a new, second-generation triazole that has also a broad-spectrum antifungal activity. In vitro, isavuconazole is more active than AMB, ITC, voriconazole, 5-FC, and FLZ (MIC50 = 0.004, 0.5, 0.008, 0.03, 0.125, and 8 μg/ml, respectively) and has lower MIC50 values than those of voriconazole in the majority of Candida species (Pasqualotto et al., 2010). Isavuconazole has demonstrated activity against candidemia and invasive candidiasis, with an action mechanism similar to that of other triazoles and with activity in triazole-resistant fungi (Vermes et al., 2000).
Ravuconazole is structurally similar to isavuconazole and has demonstrated a good antimycotic action in FLZ resistant isolates. However, higher MIC values for ravuconazole have been reported for Candida isolates that display resistance to FLZ than those for susceptible isolates (Pasqualotto et al., 2010).
Lysozyme is an enzyme found in various fluids of the human body such as saliva and respiratory secretions (Table 3 and Figure 3). It is classically known for its ability to kill bacteria through its muramidase activity. Further, lysozyme is active against numerous clinical isolates of Candida species, as well as against Aspergillus fumigatus and Penicillium species (Papini et al., 1982). However, the antifungal mechanisms of action associated with lysozyme remain subject to speculation. It is likely that lysozyme acts by reducing secreted aspartic protease (SAP, involved in Candida virulence) activity and secretion in C. albicans (Wu et al., 1999); its fungicidal activity at high concentrations likely results from damage to the cell wall or plasma membrane, causing loss of osmotic equilibrium (Wu et al., 1999).
Human lactoferrin (hLF) is a peptide that binds to iron and has protease action (Table 3 and Figure 3). Lactoferrin is found in saliva and other secretions of human body. It has been found to be active against C. albicans and C. krusei (Samaranayake et al., 1997). The mechanisms of action associated with lactoferrin are likely related to production of a cationic peptide that presents a broad antimicrobial activity (Orsi, 2004). It was verified that a synthetic peptide comprising the first cationic domain of lactoferricin H (released by pepsinolysis of hLF), named hLF1-11 (Table 3), possesses a high antifungal activity, (Lupetti et al., 2007), beyond contributing to the clearance of infections, by stimulating the production of macrophages and dendritic cells (van der Does et al., 2012). The peptide hLF1-11 also inhibited C. albicans biofilm formation at early stages, interfering with biofilm cellular density and metabolic activity and to induce the down-regulation of biofilm and hyphal-associated genes (Morici et al., 2016).
Histatin-5 is a fragment of salivary protein histatin-3 comprising the N-terminal fragment with 24 amino acids (Table 3 and Figure 3). The peptide has strong fungicidal activity, being able to kill both yeast and filamentous forms of Candida spp. even at low concentrations (15–30 μM); histatin-5 can also exert its fungicidal activity by binding to a candidate 67 kDa protein and then interfering with non-lytic ATP efflux (Edgerton and Koshlukova, 2000). Moreover, when histatin-5 is adsorpted to microtiter plate prior C. albicans biofilm formation (90 min, 24, 48, and 72 h) it is capable of reducting significantly C. albicans colonization interfering with biofilm formation. (Moffa et al., 2015). These data corroborate to those previously obtained by Vukosavljevic et al. (2012), who demonstrated the inhibitory effect of histatin-5 when adhered to hydroxyapatite and polymethylmethacrylate (PMMA) surfaces on C. albicans colonization.
Human β-defensins (HBD) are small cationic peptides that belong to the defensins family. There are three types of human β-defensin with fungicidal activity toward C. albicans (Krishnakumari et al., 2009; Schroeder et al., 2011; Tomalka et al., 2015) that function via the same mechanism (Krishnakumari et al., 2009), including human β-defensin-1 (HBD-1), human β-defensin-2 (HBD-2), and human β-defensin-3 (HBD-3) (Table 3 and Figure 3).
Pro-inflammatory molecules such as interferon-γ, bacteria, or lipopolysaccharide can stimulate the expression of HBD-1 mRNA constitutively in endothelial tissues, by binding to toll-like receptors (TLR) (Duits et al., 2002). In addition, HBD-1 is important for control of early mucosal Candida infections and plays a critical role in HBD-2 expression (Tomalka et al., 2015). Cytokines, such as tumor necrosis factor, and also the contact with bacteria and fungi (Harder et al., 1997) and interleukin (IL)-1 (Sorensen et al., 2005) stimulate the expression of HBD-2 in epithelial tissue via TLR-2 (Hertz et al., 2003). HBD-3 expression is induced via binding of TGFα (transforming growth factor alpha) to its receptor EGFR (epidermal growth factor receptor). HBD-3 can be found in keratinocytes and airway epithelial cells (Sorensen et al., 2005). Of the three HBDs, HBD-3 is fungicidal against C. albicans, showing a minimal fungicidal concentration (MFC) of 2.5 μM; HBD-2 has a poor activity against fungi, with a MFC of 8 μM (Krishnakumari et al., 2009). Also, HBD-3 elevates Xog1 activity, an exoglucanase of C. albicans cell wall, resulting in reduced adherence of the yeast (Chang et al., 2012). Studies have shown that at least one mechanism of action associated with HBD-1, 2, and 3 increases the membrane permeability of C. albicans (Krishnakumari et al., 2009).
Other small cationic peptides include the cathelicidins [human LL-37, murine cathelicidin-related antimicrobial peptide (CRAMP), and porcine PR-39], a group of antimicrobial skin peptides produced by mast cells and by mucosal and skin epithelial cells (McCormick and Weinberg, 2000) (Table 3 and Figure 3). CRAMP is both fungicidal and fungistatic against C. albicans, with MICs as low as 15 μM. LL-37 also has fungicidal and fungistatic activity and can be cleaved into shorter peptides with a higher fungicidal activity against C. albicans. Assays have shown that LL-37 and RK-31 affect membrane permeability of Candida cells. LL-37 also inhibits adhesion of C. albicans to plastics and tissues by interacting with yeast cell wall carbohydrates (Tsai et al., 2011). When the peptide LL-37 was tested for their inhibitory effects and antibiofilm properties against C. albicans strain, using both a crystal violet and an XTT [2,3-bis- (2-methoxy-4-nitro-5-sulfophenyl) -2H -tetrazolium-5-carboxanilide] assays showed satisfactory results, mainly in relation to the prevention of biofilms. Through violet crystal analysis, LL-37 had significant efficacy both in preventing biofilm formation and in inhibiting early formed biofilms of C. albicans. However, by XTT metabolic assay, LL-37 prevented biofilm formation against C. albicans, even at sub-minimum inhibitory concentrations (sub-MIC), but did not inhibit early biofilms (Luo et al., 2017).
Although natural proteins/peptides represent a promising therapeutic agents, they are usually extracted in small amount which makes it difficult to perform biological tests (Bondaryk et al., 2017). Stimulated by these facts, some researchers are synthetizing or modifying existing natural peptides in order to obtaining new molecules with enhanced antifungal activity and reduced toxicity. For instance, a shortened peptide variant of CRAMP, named P318 (Table 3), was identified by Brucker et al. (2014) and shared 67% identity with the peptide LL-37. The peptide was discovered in the islets of Langerhans of the murine pancreas. P318 showed biofilm-specific activity as it inhibited C. albicans biofilm formation at 0.15 μM without affecting planktonic survival at that concentration.
Mollica et al. (2017) synthesized and characterized nine new cationic peptides, rich in arginine and lysine amino acids to introduce cationic charges and in phenylalanine and leucine residues to increase lipophilicity. Four from these peptides showed a potente antifungal activity against different clinical isolates of Candida spp. (MIC ranged from 62.5 to 500 μg).
Cools et al. (2017) delineated and identified a linear HsLin06_18, a 19-mer peptide (Table 3) derived from the C-terminal part of HsAFP1, an antifungal and antibiofilm plant defensin isolated from Heuchera sanguinea. Synergistic combination of HsLin06_18 with caspofungin significantly reduced in vitro biofilm formation of C. glabrata and C. albicans on catheters, as well as in vivo biofilm formation of C. albicans strain. In addition, combination dose (4.6 μM of HsLin06_18 + 0.01 μM caspofungin) was fungicidal against planktonic cells of tested strains, killing until 80% of yeast population. Lim et al. (2016) demonstrated the antifungal and antibiofilm activities of a synthetic peptide consisting of 15 amino acids at the C-terminus of human β-defensin 3 (HBD3-C15, Table 3). They observed that the biofilm of C. albicans on dentin disks was inhibited by HBD3-C15 in a dose-dependent manner.
Despite promises, much research is still needed on hemolytic activity, instability, production modes, interaction with high salt concentrations, anti-virulence activity, and poor ability to cross physiological barriers of these peptides, which could limit their use in the clinic.
The use of plants and their bioactive molecules in the treatment of candidiasis has emerged as a promising alternative to traditional drugs against resistance which has developed in the Candida genus. Antifungal substances derived from plants can selectively act on different targets with fewer side effects. In addition, the practice of phytotherapy is inexpensive; therefore, floral diversity has resulted in an increase in potential usage in populations experiencing economic difficulties. This review comments on some of the extracts of plants or their metabolites that in vitro and in vivo studies have already demonstrated a potential antifungal activity.
There are some families of plants that are more studied than others as the Combretaceae and Acanthaceae. Studies have shown that leaves, seeds, fruits, and flowers have the most enriched plant components. Leaves, as well as the seeds and fruits of plants have higher levels of phenolic compounds. The concentration of these compounds also depends on the nature of the chemical used as solvent in the extraction process as well as on the growth and storage conditions (Martins et al., 2015a). The most used solvents in extraction processes are dichloromethane, methanol, ethanol, ethyl acetate and n-butanol. (Martins et al., 2015a).
Recently, some authors have verified and evaluated the biological activity of plant products against Candida species. Lonicera japonica, a medicinal plant of folk medicine of China used to treat some diseases, was investigated by Chen et al. (2012) for the in vivo activity of an ethanol extract of its aerial parts. The extract showed a very strong antimicrobial activity against C. albicans and C. tropicalis and potent wound healing capacity; further, enhanced production of anti-inflammatory cytokines was observed. In this way, the authors suggested that both activities detected in this extracts act synergistically accelerating the process of wound healing.
Some properties of Lannea welwitschii and Justicia flava were investigated by Agyare et al. (2013). Methanolic extract of Lannea welwitschii leaves was antimicrobial against clinical strains of C. albicans and other microorganisms. The MIC for C. albicans was 2.5 mg/mL. Treatment with an extract from both plants resulted in a significant decrease in wound size and increase in wound tensile strength. A preliminary phytochemical screening of extracts revealed tannins, flavonoids, alkaloids, and glycosides as compounds. These results corroborate the use of these vegetable extracts in treatment of wounds and infections in phytotherapy.
Pereira et al. (2014) studied the activity of Pyrostegia venusta crude flower extracts, fractions, and pure compounds against isolates of Candida spp. and showed an effective broad spectrum antifungal activity. Nordin et al. (2014) reported anticandidal activity in an extract of Piper betle leaves; in fact, the extract inhibited the growth of all Candida species tested. Isa et al. (2014) also verified antimicrobial property of four different extracts of Strychnos spinosa and their fractions against American Type Culture Collection (ATCC) strains of C. albicans and C. albicans isolates (MICs of 0.16 and 0.63 mg/mL, respectively). Otari et al. (2014) described that silver nanoparticles containing Manilkara zapota seed extracts showed good activity against Candida species.
Shahzad et al. (2014) evaluated the antifungal potential of 14 polyphenols against various C. albicans clinical isolates in terms of planktonic and sessile MICs (PMICs and SMICs, respectively). Among these, 7 were able to inhibit planktonic growth. The most effective was pyrogallol (PMIC50 = 78 μg/mL) and curcumin (PMIC50 = 100 μg/mL). In addition, curcumin inhibited adhesion capability of cells and demonstrated anti-biofilm activity against C. albicans (SMIC50 = 50 μg/mL).
Martins et al. (2015b) evaluated ten different plant extracts commonly used in folk medicine for antifungal activity against Candida spp. They verified that hydro-methanolic extracts of leaves from two of these plants, Juglans regia and Eucalyptus globulus, demonstrated excellent antimycotic property against all Candida strains. Goncalves et al. (2015) described anticandidal activity in a Cynomorium coccineum methanol extract, which showed excellent action against C. guilliermondii and C. krusei, showing very low MIC values (0.025 mg/mL). Moraes et al. (2015) investigated the antimycotic property of a hydroethanolic extract of Uncaria tomentosa and some of its fractions against resistant Candida spp. and verified that the water-insoluble fraction showed significant antifungal activity.
Akroum (2017) showed antifungal activity in an acetylic extract of Vicia faba against C. albicans (MIC of 0.010 mg/mL) in vitro. Further, mortality rates were reduced in mice that were administered with the extract (20 μg/mL) for treatment of candidiasis.
Correia et al. (2016) evaluated the antifungal properties of six plants from Brazilian Cerrado commonly used in folk medicine (ethanolic and aqueous extracts) against different Candida reference strains using the disk diffusion method and determining MICs. Among these plants, the most promising were Eugenia dysenterica and Pouteria ramiflora. They showed excellent activity against C. tropicalis, C. famata, C. krusei, C. guilliermondii, and C. parapsilosis with low MICs values. A phytochemical screening of active extracts from these plants disclosed as main components flavonoids and catechins.
Terças A.G. et al. (2017) found antifungal properties in crude extract and fractions (n-butanolic and ethyl acetate ones) from Terminalia catappa leaves via the agar diffusion and microdilution tests when analyzed against Candida spp.; hydrolysable tannins (punicalin, punicalagin), gallic acid, and flavonoid C-glycosides are likely the active components. Todorovic et al. (2017) verified the antifungal activity of polyphenols (flavanol monomers such as epicatechin and catechin, and procyanidin oligomers) of alkalized/non-alkalized Theobroma cacao powders against C. albicans (ATCC 10231) and determined a MIC value of 5.0 mg/mL using the broth microdilution method.
Phytochemicals present in leaves of Carya illinoensis were first identified by Bottari et al. (2017), and the antimicrobial activity of their aqueous and ethanolic extracts was determined. Both extracts had MIC values against seven Candida reference strains between 25 mg/mL and 6.25 mg/mL. Phenolic acids (gallic acid and ellagic acid), flavonoids (rutin), and tannins (catechins and epicatechins) were likely responsible, in part, for the activity against Candida strains. Further, the extracts inhibited the production of C. albicans germ tubes.
Important biologically active molecules are found in plants (Martins et al., 2015c). Polyphenols are a kind of substance most found in plants; they are low molecular weight naturally occurring organic compounds that contain one or more phenolic groups (Daglia, 2012; Shahzad et al., 2014). Further, polyphenols perform various substantial functions in plant physiology and, therefore, can be found, in lesser or greater quantity, in all of them. Phenolic acids, flavonoids, tannins, coumarins, are some examples of phenolic compounds found in and extracted from medicinal plants (Daglia, 2012). Research has shown that polyphenols have potentially healthy effects in humans, working primarily as anticancer, antihypertensive, anti-allergen, anti-inflammatory, antioxidants, and antimicrobial agents. The antimicrobial activity of polyphenols has been extensively investigated mainly against bacteria (Daglia, 2012).
Nevertheless, the antifungal activity of some of the above-mentioned phenolic compounds remains unknown and determining the antifungal activity of such compounds remains an open area of research. Reports of studies of phenolic compounds against Candida are still scarce. There are few studies on the mechanism of action of the substance, cytotoxicity, the synergism with traditional antifungals drugs and their anti-virulence activities (such as inhibition of biofilm formation, interference of adhesion capability, interference of hyphal formation or inhibition of exoenzymes production).
Flavan-3-ols, flavonols, and tannins have received the most attention among the known polyphenols, attributable to their large spectrum of efficacy and high antimicrobial property, which have been shown in many bacteria strains and isolates. Further, virulence factors of bacteria may be influenced by polyphenols and also these substances can act in synergism with antibiotics; consequently, those polyphenols are also the most studied in relation to their anti-Candida activities. Thus, we reviewed the antifungal activities related in scientific literature of the polyphenols considered most actives and their mechanisms of action.
Flavonoids are a class of natural compounds with several known protective activities, including antifungal activity. The flavonoids include subclasses such as chalcones, flavones, isoflavones, flavonols, flavanols (flavan-3-ol), and anthocyanidins (Seleem et al., 2017). The activity of flavonols such as quercetin, myricetin, and kaempferol has been described in C. albicans. Herrera et al. (2010) showed inhibition of C. albicans growth with isolated flavonols from propolis using quercetin in an agar microdilution method, obtained a variation of MIC values from 197 to 441 μg/mL. These same authors found similar results for myricetin and kaempferol against Candida species (Herrera et al., 2010). Other studies have reported antimicrobial activity (Avila et al., 2008; Batovska et al., 2009) for other propolis polyphenols such as flavanone (pinocembrin and pinostrobin) and chalcones (2,4-dihydroxychalcone and 2,4-dihydroxy-3-methoxychalcone). The flavanols subclass (flavan-3-ol) and gallotannin, extracted from Syzygium cordatum, also showed inhibitory properties on the growth of C. albicans, with a MIC of 0.195 mg/mL in a microdilution test (Mulaudzi et al., 2012). Serpa et al. (2012) isolated baicalein, belonging to a subclass of flavones, from Scutellaria baicalensis, and induced apoptosis in C. albicans, with a MIC value of 26 μg/mL. Apigenin, a flavone isolated from propolis, had a MIC of 441 μg/mL against C. albicans, as shown using a microdilution test.
Tannins are polyphenolic compounds present in various plant parts, such as the roots, flowers, leaves, fruits, and seeds. Tannins are divided into ellagitannins (hydrolysable tannins), proanthocyanidins (condensed tannins) and gallotannins (Duval and Avérous, 2016). They have the ability to interact with and precipitate macromolecules such as proteins (dos Santos et al., 2017), as well as having antimicrobial properties. However, the mechanisms underlying the antimicrobial action of tannins in different microorganisms such as bacteria and fungi are still under investigation (Morey et al., 2016; Xu et al., 2016; dos Santos et al., 2017).
The knowledge that wood durability could be attributed to the accumulation of ellagitannins, with the ability to precipitate proteins and/or withdraw metallic cofactors acting as a microbial barrier, raised the suspicion that ellagitannins would have antifungal activity. Accordingly, ellagitannins isolated from Ocotea odorifera, a plant commonly used in Brazil in folk medicine, have a very good activity against C. parapsilosis (Yamaguchi et al., 2011). dos Santos et al. (2017) verified that encapsulated tannins from Acacia mearnsii have moderate activity against the filamentous fungi Aspergillus niger (ATCC 9642) and C. albicans (ATCC 34147).
Other phytosubstances reported to be active against yeasts and fungi include essential oils, which are derived from aromatic medicinal plants (Reichling et al., 2009; Shahid et al., 2009; Centeno et al., 2010; Vale-Silva et al., 2010; Pitman et al., 2011; Sienkiewicz et al., 2011). Essential oils are rich in monoterpenes, sesquiterpenes, and/or phenylpropanoids, considerate volatile organic compounds. Essential oils are present in various plant species. Mondello et al. (2003) proposed that tea tree oil could be used in antifungal therapy, because it showed efficacy against multidrug-resistant Candida species in vitro and against mucosal candidiasis in vivo; they further documented that terpinen-4-ol was the main substance to contribute to the anticandidal activity.
Several oils have demonstrated activity against Candida species. Essential oils from Carica papaya have inhibitory effects against Candida species, as shown via the agar diffusion method and the microdilution method, with MICs between 4 and 16 mg/mL and MFCs between 16 and 64 μg/mL (Sharma H. et al., 2016; Sharma Y. et al., 2016; He et al., 2017). Minooeianhaghighi et al. (2017) tested a combination of essential oils from Cuminum cyminum and Lavandula binaludensis against C. albicans isolates, showing growth inhibition at concentrations between 3.90 and 11.71 μg/mL. Essential oils from Cymbopogon nardus have also shown antimicrobial potential against Candida species, with MICs between 250 and 1000 μg/mL and with inhibition of hyphal growth in C. albicans at concentrations between 15.8 and 1000 μg/mL (De Toledo et al., 2016). In addition to inhibiting biofilm formation (Abu-Darwish et al., 2016), essential oils from Artemisia judaica have been shown to inhibit the formation of germination tubes in C. albicans, with 80% inhibition of filamentation at a concentration of 0.16 μL/mL. Köse et al. (2016) demonstrated the fungicidal potential of essential oils from Centaurea baseri against Candida species, with a MIC of 60 μg/mL.
Sharifzadeh et al. (2015) observed that essential oils from Trachyspermum ammi have anticandidal effects against isolates of C. albicans, some of which were resistant to FLZ. Gavanji et al. (2015) have compared the efficiency of herbal essences from Foeniculum vulgare, Satureja hortensis, C. cyminum, and Zataria multiflora against C. albicans. Essential oils from Z. multiflora showed the best anticandidal activity of those tested, with MIC and MFC values of 34 and 64 μg/mL, respectively.
Among monoterpenes there is thymol (2-isopropyl-5-methylphenol), (Sánchez et al., 2004). It is the most abundant constituent in essential oils from Thymus vulgaris (thyme) (De Lira Mota et al., 2012) and the major component of essential oils from Origanum vulgare (oregano) (Romero et al., 2012). Antifungal activity of thymol was investigated by de Castro et al. (2015) against Candida strains. They verified fungistatic and fungicidal activity, mechanisms of action and mode of interactions in combination with nystatin. Thymol had antifungal properties, with MIC of 39 μg/mL against C. albicans and C. krusei, and MIC of 78 μg/mL against C. tropicalis. Antifungal assays also showed an eightfold increase (from 39.0 to 312.5 μg/mL) in thymol MIC values against C. albicans in the presence of exogenous ergosterol, indicating that thymol binds to ergosterol in the plasma membrane, thereby increasing ion permeability and resulting in cell death. Combination of thymol and nystatin resulted in synergy.
Terpenoids exhibit a very good antimycotic activity against blastopores and filamentous forms growth of C. albicans at concentrations that are non-toxic to HeLa cells (Zore et al., 2011). Accordingly, terpenoids may be useful as a future antifungal chemotherapeutic agent, in addition to its synergistic effects with conventional drugs such as FLZ (Zore et al., 2011). Further, in experiments realized by Fan et al. (2011), rubiarbonol G, a triterpenoid from Rubia yunnanensis, showed potent antimicrobial activity against C. albicans, with a MIC of 10.5 μg/mL.
The anti-biofilm activity of terpenes, along with the efficacy of thymol, geraniol, and carvacrol in the treatment of Candida infections associated with the use of hospital devices has been related (Dalleau et al., 2008). Mechanisms underlying the effects associated with carvacrol include alterations of the cellular cytoplasmic membrane and induction of apoptosis, as shown in an in vitro macrodilution study in Candida species (Mulaudzi et al., 2012).
Phenylpropanoids are other naturally occurring compounds frequently studied for their anti-Candida properties; they are categorized as coumarins, phenylpropanoic acid, and lignans (Lu et al., 2017). Navarro-García et al. (2011) and Raut et al. (2014), found that a coumarin (scopoletin) and two phenylpropanoic acids (salicylaldehyde and anisyl alcohol) have antifungal property against C. albicans, with MICs of 25, 31, and 31 μg/mL, respectively.
Taken together these data show that plants contain molecules possessing high bioactive potential. However, the process of discovering bioactive molecules is complex and time-consuming, involving the isolation, identification, and optimization of pharmacokinetic and pharmacodynamic properties, as well as the selection of lead compounds for further drug development.
The knowledge about synergistic effects of plant extracts or their phytoconstituents with traditional agents is nowadays a type of study that is indispensable, since some screening assays, most realized in vitro, have evidenced that plant extracts are less effective than existing antifungal agents (Newman and Cragg, 2012) and that extracts with MIC of 1000 μg/mL are considered inefficient (Morales et al., 2008).
The use of drug combinations in treatment of infections by fungi is a preferred strategy clinically. Although combination of medications requires a careful evaluation of the synergistic, antagonistic, and agonist properties of the drugs involved (Lewis and Kontoyiannis, 2001), it has been used successfully in many cases of fungal infection (Serena et al., 2005; Espinel-Ingroff, 2009; Bink et al., 2011; Rodrigues et al., 2014) (Table 1). The synergistic effects of drugs are primarily attributable to cell wall damage as one antifungal component potentiates the activity of drugs exactly against some constituent of plasma membrane. Alternatively, a compromised cell wall has increased permeability and could facilitate movement of drugs across the cell membrane to their targets.
Accordingly, the synergistic effect of plant extracts or their biomolecules in combination with conventional antimicrobial agents (or with some other different extract or biosubstance) against clinical multidrug-resistant microorganisms represents a successful therapeutic approach (Mukherjee et al., 2005). Advantages of combination therapies include lower doses of antifungal agent, possible synergistic actions between antifungals, and less development of drug resistance. The objective of this strategy is to maximize the antifungal effects. Some examples are addressed herein.
Tangarife-Castaño et al. (2011) reported synergy between essential oils or plant extracts associated with antifungal drugs when used as anti-C. albicans agents. The best synergistic effects were obtained for combination between itraconazole and P. bredemeyeri extract [fractional inhibitory concentration index (FICI) range of 0.09–0.13] against C. albicans.
Chanda et al. (2013) verified a synergistic potential when methanolic extract of T. catappa leaves was combined with nystatin or AMB against C. albicans (ATCC 209), C. neoformans (National Collection of Industrial Microorganisms [NCIM] 3542), C. glabrata (NCIM 3448), C. apicola (NCIM 3367), and Trichosporon beigelii (NCIM 3404). As such, maximum synergy was observed against C. apicola.
Santos et al. (2013) related the antimycotic properties of an ethanol extract of Hyptis martiusii (EEHM) against C. albicans, C. krusei, and C. tropicalis. They verified synergistic antifungal activity for EEHM in combination with metronidazole when used against C. tropicalis.
Avijgan et al. (2014) reported a synergistic effect between an Echinophora platyloba ethanolic extract and different azoles against isolates of C. albicans from vaginal secretions of patients with recurrent vulvovaginitis. MIC and MFC values ranged from 3.1 to 6.25 mg/mL and 6.2 to 12.5 mg/mL, respectively, showing potent synergistic effects of the E. platyloba ethanol extract in combination with itraconazole and FLZ.
Combination between thymol and nystatin was found to have synergistic effects against Candida species (de Castro et al., 2015), reducing the MICs of both products by 87.4% and generating a fractional inhibitory concentration (FIC) index of 0.25.
Synergism between a water insoluble fraction (WIF) from U. tomentosa (cat’s claw) bark and the agents terbinafine or FLZ was investigated against seven resistant isolates of C. glabrata and C. krusei via the checkerboard procedure using a microdilution technique (Moraes et al., 2015). Synergism was observed between the U. tomentosa WIF and terbinafine, as well as between the U. tomentosa WIF and FLZ. The most efficacious synergistic effects leading to cell damage were unequivocally attributed to a combination of the U. tomentosa WIF and terbinafine (1.95:4.0, l g/mL), as well as the U. tomentosa WIF and FLZ (1.95:8.0, l g/mL). Moraes et al. (2015) also demonstrated, through differential scanning calorimetry and infrared analysis, that intermolecular interactions between the U. tomentosa WIF components and either terbinafine or FLZ occurring outside the cell wall are likely responsible for synergistic effects observed between substance. An action on constituents of the cell wall was suggested, independent of ABC efflux pump mechanisms.
Ngouana et al. (2015) conducted a bioguided screening with sub-fraction combinations of T. catappa, Terminalia mantaly, and Monodora tenuifolia against C. albicans, C. glabrata, C. parapsilosis, and C. neoformans isolates, as well as the C. albicans NR-29450 reference strain. They observed synergistic interactions between subfractions combinations. A combination of M. tenuifolia and T. mantaly (C36/C12) sub-fractions showed synergistic interactions and fungicidal effects against most tested strains.
Cavalcanti Filho et al. (2017) verified that the methanolic extract of Buchenavia tetraphylla is a great source of antimicrobial compounds that enhance the action of FLZ against different C. albicans isolates from vaginal secretions as well as azole-resistant isolates. The extract increased the action of FLZ in most strains through additive (20% of strains) or synergistic (60% of strains) effects.
Although many in vitro studies examining synergistic effects among potential antifungal biomolecules and traditional antifungal agents have been reported in the literature as described herein, the mechanisms underlying these synergistic effects are poorly understood. Several randomized and controlled analyzes have been performed with the objective of verifying the efficacy and risks of using traditional antifungal combinations; however, the high cost, reduced number of clinical cases and existence of confusing variables have resulted in contradictory and poor results. Therefore, it is extremely relevant the constant search for new phytocompounds to examine carefully possible synergism between them and conventional antifungal agents in order to obtain more insight. A lack of consensus in the medical clinical emphasizes the need to conduct further clinical trials using combinations of antifungals. The experiments and results addressed herein support further investigation of new plant constituents with antifungal properties and the efficacy of combination therapies involving phytocomponents and traditional antifungal agents as an important start for the development of unusual and original antifungal therapies.
Candida species are highly resistant to existing antifungal agents and can adapt to different host niches thus representing a serious risk to human health. The mechanisms underlying development of antifungal resistance are complex and involve multiple pathways and genes. Further, these mechanisms continue to change and evolve, challenging the medical clinic and exacerbating the need for discovering original therapies against Candida diseases. In this way, identification of new bioactive compounds as well as development of original formulations of antifungals and combinations involving active biomolecules and conventional agents represents the possibility for a successful therapeutic approach.
CAM, CV, GOS, FR, and AL conceived and designed the review. CAM, CV, GOS, FR, AL, MSC, AF, FN, RR, EP, and MA wrote the paper. All authors read and approved the final manuscript.
FR was funded by grant BM-00946/17 from the Fundação de Amparo à Pesquisa e ao Desenvolvimento Científico e Tecnológico do Maranhão (FAPEMA).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abu-Darwish, M. S., Cabral, C., Gonçalves, M. J., Cavaleiro, C., Cruz, M. T., Zulfiqar, A., et al. (2016). Chemical composition and biological activities of Artemisia judaica essential oil from southern desert of Jordan. J. Ethnopharmacol. 191, 161–168. doi: 10.1016/j.jep.2016.06.023
Agyare, C., Bempah, S. B., Boakye, Y. D., Ayande, P. G., Adarkwa-Yiadom, M., and Mensah, K. B. (2013). Evaluation of antimicrobial and wound healing potential of Justicia flava and Lannea welwitschii. Evid. Based Complement. Altern. Med. 2013:632927. doi: 10.1155/2013/632927
Akroum, S. (2017). Antifungal activity of acetone extracts from Punica granatum L., Quercus suber L. and Vicia faba L. J. Mycol. Med. 27, 83–89. doi: 10.1016/j.mycmed.2016.10.004
Allen, D., Wilson, D., Drew, R., and Perfect, J. (2015). Azole antifungals: 35 years of invasive fungal infection management. Expert Rev. Anti Infect. Ther. 13, 787–798. doi: 10.1586/14787210.2015.1032939
Álvares, C. A., Svidzinski, T. I. E., and Consolaro, M. E. L. (2007). Candidíase vulvovaginal: fatores predisponentes do hospedeiro e virulência das leveduras. J. Bras. Patol. Med. Lab. 43, 319–327. doi: 10.1590/S1676-24442007000500004
Amaral, A. C., and Felipe, M. S. S. (2013). Nanobiotechnology: an efficient approach to drug delivery of unstable biomolecules. Curr. Protein Pept. Sci. 14, 588–594. doi: 10.2174/1389203711209070632
Armengou, A., Hurtado, O., Leira, R., Obon, M., Pascual, C., Moro, M. A., et al. (2003). L-arginine levels in blood as a marker of nitric oxide-mediated brain damage in acute stroke: a clinical and experimental study. J. Cereb. Blood Flow Metab. 23, 978–984. doi: 10.1097/01.WCB.0000080651.64357.C6
Artymiuk, P. J., and Blake, C. C. F. (1981). Refinement of human lysozyme at 1.5 angstroms resolution. analysis of non-bonded and hydrogen-bond interactions. J. Mol. Biol. 152, 737–762. doi: 10.1016/0022-2836(81)90125-X
Avijgan, M., Mahboubi, M., Nasab, M. M., Nia, E. A., and Yousefi, H. (2014). Synergistic activity between Echinophora platyloba DC ethanolic extract and azole drugs against clinical isolates of Candida albicans from women suffering chronic recurrent vaginitis. J. Mycol. Med. 24, 112–116. doi: 10.1016/j.mycmed.2014.01.116
Avila, H. P., Smania, E. F., Monache, F. D., and Smania, A. Jr. (2008). Structure-activity relationship of antibacterial chalcones. Bioorg. Med. Chem. 16, 9790–9794. doi: 10.1016/j.bmc.2008.09.064
Barchiesi, F., Falconi Di Francesco, L., and Scalise, G. (1997). In vitro activities of terbinafine in combination with fluconazole and itraconazole against isolates of Candida albicans with reduced susceptibility to azoles. Antimicrob. Agents Chemother. 41, 1812–1814.
Barchiesi, F., Orsetti, E., Osimani, P., Catassi, C., Santelli, F., and Manso, E. (2016). Factors related to outcome of bloodstream infections due to Candida parapsilosis complex. BMC Infect. Dis. 16:387. doi: 10.1186/s12879-016-1704-y
Barchiesi, F., Schimizzi, A. M., Caselli, F., Novelli, A., Fallani, S., Giannini, D., et al. (2000). Interactions between triazoles and amphotericin B against Cryptococcus neoformans. Antimicrob. Agents Chemother. 44, 2435–2441. doi: 10.1128/AAC.44.9.2435-2441.2000
Bassetti, M., Righi, E., Ansaldi, F., Merelli, M., Trucchi, C., De Pascale, G., et al. (2014). A multicenter study of septic shock due to candidemia: outcomes and predictors of mortality. Intensive Care Med. 40, 839–845. doi: 10.1007/s00134-014-3310-z
Batovska, D., Parushev, S., Stamboliyska, B., Tsvetkova, I., Ninova, M., and Najdenski, H. (2009). Examination of growth inhibitory properties of synthetic chalcones for which antibacterial activity was predicted. Eur. J. Med. Chem. 44, 2211–2218. doi: 10.1016/j.ejmech.2008.05.010
Bayhan, G. I., Garipardic, M., Karaman, K., and Akbayram, S. (2015). Voriconazole-associated visual disturbances and hallucinations. Cutan. Ocul. Toxicol. 35, 80–82. doi: 10.3109/15569527.2015.1020544
Bennett, J. E., Izumikawa, K., and Marr, K. A. (2004). Mechanism of increased fluconazole resistance in Candida glabrata during prophylaxis. Antimicrob. Agents Chemother. 48, 1773–1777. doi: 10.1128/AAC.48.5.1773-1777.2004
Bink, A., Pellens, K., Cammue, B., and Thevissen, K. (2011). Antibiofilm strategies: how to eradicate Candida biofilms. Open Mycol. J. 5, 29–38. doi: 10.2174/1874437001105010029
Bondaryk, M., Staniszewska, M., Zielińska, P., and Urbańczyk-Lipkowska, Z. (2017). Natural antimicrobial peptides as inspiration for design of a new generation antifungal compounds. J. Fungi 3:E46. doi: 10.3390/jof3030046
Bottari, N. B., Lopes, L. Q., Pizzuti, K., Filippi, Dos Santos Alves, C., Corrêa, M. S., Bolzan, L. P., et al. (2017). Antimicrobial activity and phytochemical characterization of Carya illinoensis. Microb. Pathog. 104, 190–195. doi: 10.1016/j.micpath.2017.01.037
Bouza, E., and Munoz, P. (2008). Epidemiology of candidemia in intensive care units. Int. J. Antimicrob. Agents 32, 87–91. doi: 10.1016/S0924-8579(08)70006-2
Brucker, K., Delattin, N., Robijns, S., Steenackers, H., Verstraeten, N., Landuyt, B., et al. (2014). Derivatives of the mouse cathelicidin-related antimicrobial peptide (CRAMP) inhibit fungal and bacterial biofilm formation. Antimicrob. Agents Chemother. 58, 5395–5404. doi: 10.1128/AAC.03045-14
Brunke, S., and Hube, B. (2013). Two unlike cousins: Candida albicans and Candida glabrata infection strategies. Cell Microbiol. 15, 701–708. doi: 10.1111/cmi.12091
Caban, M., Strapagiel, D., Dziadek, J., Korycka-Machała, M., and Grzelak, A. (2016). Principles of a new protocol for prediction of azole resistance in Candida albicans infections on the basis of ERG11 polymorphisms. Curr. Microbiol. 73, 172–182. doi: 10.1007/s00284-016-1039-3
Calderone, R. A., and Fonzi, W. A. (2001). Virulence factors of Candida albicans. Trends Microbiol. 9, 327–335. doi: 10.1016/S0966-842X(01)02094-7
Canuto, M., and Rodero, F. G. (2002). Antifungal drug resistance to azoles and polyenes. Lancet Infect. Dis. 2, 550–563. doi: 10.1016/S1473-3099(02)00371-7
Carrillo-Muñoz, A. J., Giusiano, G., Ezkurra, P. A., and Quindós, G. (2006). Antifungal agents: mode of action in yeast cells. Rev. Esp. Quimioter. 19, 130–139.
Cavalcanti Filho, J. R., Silva, T. F., Nobre, W. Q., Oliveira de Souza, L. I., Silva, E., Silva Figueiredo, C. S., et al. (2017). Antimicrobial activity of Buchenavia tetraphylla against Candida albicans strains isolated from vaginal secretions. Pharm. Biol. 55, 1521–1527. doi: 10.1080/13880209.2017.1304427
Centeno, S., Calvo, M. A., Adelantado, C., and Figueroa, S. (2010). Antifungal activity of Rosmarinus officinalis and Thymus vulgaris against Aspergillus flavus and A. ochraceus. Pak. J. Biol. Sci. 13, 452–455. doi: 10.3923/pjbs.2010.452.455
Cernicka, J., and Subik, J. (2006). Resistance mechanisms in fluconazole-resistant Candida albicans isolates from vaginal candidiasis. Int. J. Antimicrob. Agents 27, 403–408. doi: 10.1016/j.ijantimicag.2005.12.005
Chamilos, G., Lewis, R. E., Albert, N., and Kontoyiannis, D. P. (2007). Paradoxical effect of echinocandins across Candida species in vitro: evidence for echinocandin-specific and Candida species-related differences. Antimicrob. Agents Chemother. 51, 2257–2259. doi: 10.1128/AAC.00095-07
Chanda, S., Rakholiya, K., Dholakia, K., and Baravalia, Y. (2013). Antimicrobial, antioxidant, and synergistic properties of two nutraceutical plants: Terminalia catappa L. and Colocasia esculenta L. Turk. J. Biol. 37, 81–91. doi: 10.3906/biy-1203-41
Chandra, J., Zhou, G., and Ghannoum, M. A. (2005). Fungal biofilms and antimycotics. Curr. Drug Targets 6, 887–894. doi: 10.2174/138945005774912762
Chang, W., Li, Y., Zhang, L., Cheng, A., and Lou, H. (2012). Retigeric acid B attenuates the virulence of Candida albicans via inhibiting adenylyl cyclase activity targeted by enhanced farnesol production. PLoS One 7:e41624. doi: 10.1371/journal.pone.0041624
Chaturvedi, V., Ramani, R., Andes, D., Diekema, D. J., Pfaller, M. A., Ghannoum, M. A., et al. (2011). Multilaboratory testing of two-drug combinations of antifungals against Candida albicans, Candida glabrata, and Candida parapsilosis. Antimicrob. Agents Chemother. 55, 1543–1548. doi: 10.1128/AAC.01510-09
Chen, C. G., Yang, Y. L., Shih, H. I., Su, C. L., and Lo, H. J. (2004). CaNdt80 is involved in drug resistance in Candida albicans by regulating CDR1. Antimicrob. Agents Chemother. 48, 4505–4512. doi: 10.1128/AAC.48.12.4505-4512.2004
Chen, C. G., Yang, Y. L., Tseng, K. Y., Shih, H. I., Liou, C. H., Lin, C. C., et al. (2009). Rep1p negatively regulating MDR1 efflux pump involved in drug resistance in Candida albicans. Fungal Genet. Biol. 46, 714–720. doi: 10.1016/j.fgb.2009.06.003
Chen, W.-C., Liou, S.-S., Tzeng, T.-F., Lee, S. L., and Liu, I. M. (2012). Wound repair and anti-inflammatory potential of Lonicera japonica in excision wound-induced rats. BMC Complement. Altern. Med. 12:226. doi: 10.1186/1472-6882-12-226
Chen, Y. L., Lehman, V. N., Averette, A. F., Perfect, J. R., and Heitman, J. (2013). Posaconazole exhibits in vitro and in vivo synergistic antifungal activity with caspofungin or FK506 against Candida albicans. PLoS One 8:e57672. doi: 10.1371/journal.pone.0057672
Clancy, C. J., and Nguyen, M. H. (2013). Finding the “missing 50%” of invasive candidiasis: how nonculture diagnostics will improve understanding of disease spectrum and transform patient care. Clin. Infect. Dis. 56, 1284–1292. doi: 10.1093/cid/cit006
Cools, T. L., Struyfs, C., Drijfhout, J. W., Kucharíková, S., Lobo Romero, C., Van Dijck, P., et al. (2017). A Linear 19-Mer plant defensin-derived peptide acts synergistically with Caspofungin against Candida albicans biofilms. Front. Microbiol. 8:2051. doi: 10.3389/fmicb.2017.02051
Cornely, O. A., Bassetti, M., Calandra, T., Garbino, J., Kullberg, B. J., Lortholary, O., et al. (2012). ESCMID guideline for the diagnosis and management of Candida diseases 2012: non-neutropenic adult patients. Clin. Microbiol. Infect. 8, 19–37. doi: 10.1111/1469-0691.12039
Correia, A. F., Dâmaris Silveira, D., Fonseca-Bazzo, Y. M., Magalhães, P. O., Fagg, C. W., da Silva, E. C., et al. (2016). Activity of crude extracts from Brazilian cerrado plants against clinically relevant Candida species. BMC Complement. Altern. Med. 16:203. doi: 10.1186/s12906-016-1164-3
Costa, C., Ponte, A., Pais, P., Santos, R., Cavalheiro, M., Yaguchi, T., et al. (2015). New mechanisms of flucytosine resistance in C. glabrata unveiled by a chemogenomics analysis in S. cerevisiae. PLoS One 10:e0135110. doi: 10.1371/journal.pone.0135110
Coste, A., Turner, V., Ischer, F., Morschha-User, J., Forche, A., Selmecki, A., et al. (2006). A mutation in Tac1p, a transcription factor regulating CDR1 and CDR2, is coupled with loss of heterozygosity at chromosome 5 to mediate antifungal resistance in Candida albicans. Genetics 172, 2139–2156. doi: 10.1534/genetics.105.054767
Coste, A. T., Karababa, M., Ischer, F., Bille, J., and Sanglard, D. (2004). TAC 1, transcriptional activator of CDR genes, is a new transcription factor involved in the regulation of Candida albicans ABC transporters CDR1 and CDR2. Eukaryot. Cell 3, 1639–1652. doi: 10.1128/EC.3.6.1639-1652.2004
Cowen, L. E., Sanglard, D., Howard, S. J., Rogers, P. D., and Perli, D. S. (2015). Mechanisms of antifungal drug resistance. Cold Spring Harb. Perspect. Med. 5:a019752. doi: 10.1101/cshperspect.a019752
Cuenca-Estrella, M. (2010). Antifúngicos en el tratamiento de las infecciones sistémicas: importancia del mecanismo de acción, espectro de actividad y resistencias. Rev. Esp. Quimioter. 23, 169–176.
Cui, J., Ren, B., Tong, Y., Dai, H., and Zhang, L. (2015). Synergistic combinations of antifungals and anti-virulence agents to fight against Candida albicans. Virulence 6, 362–371. doi: 10.1080/21505594.2015.1039885
Daglia, M. (2012). Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 23, 174–181. doi: 10.1016/j.copbio.2011.08.007
Dalleau, S., Cateau, E., Bergès, T., Berjeaud, J. M., and Imbert, C. (2008). In vitro activity of terpenes against Candida biofilms. Int. J. Antimicrob. Agents 31, 572–576. doi: 10.1016/j.ijantimicag.2008.01.028
de Castro, R. D., de Souza, T. M. P., Bezerra, L. M., Ferreira, G. L., Costa, E. M., and Cavalcanti, A. L. (2015). Antifungal activity and mode of action of Thymol and its synergism with nystatin against Candida species involved with infections in the oral cavity: an in vitro study. BMC Complement. Altern. Med. 15:417. doi: 10.1186/s12906-015-0947-2
De Lira Mota, K. S., de Oliveira Pereira, F., de Oliveira, W. A., Lima, I. O., and de Oliveira Lima, E. (2012). Antifungal activity of Thymus vulgaris L. essential oil and its constituent phytochemicals against Rhizopus oryzae: interaction with ergosterol. Molecules 17, 14418–14433. doi: 10.3390/molecules171214418
De Toledo, L. G., Ramos, M. A. D. S., Spósito, L., Castilho, E. M., Pavan, F. R., Lopes, É. D. O., et al. (2016). Essential oil of Cymbopogon nardus (L.) Rendle: a strategy to combat fungal infections caused by Candida species. Int. J. Mol. Sci. 17:E1252. doi: 10.3390/ijms17081252
Delgado, A. C., de Jesus Pedro, R., Aoki, F. H., Resende, M. R., Trabasso, P., Colombo, A. L., et al. (2009). Clinical and microbiological assessment of patients with a long-term diagnosis of human immunodeficiency virus infection and Candida oral colonization. Clin. Microbiol. Infect. 15, 364–371. doi: 10.1111/j.1469-0691.2009.02707.x
Dismukes, W. E. (2000). Introduction to antifungal drugs. Clin. Infect. Dis. 30, 653–657. doi: 10.1086/313748
dos Santos, C., Vargas, A., Fronza, N., and Dos Santos, J. H. Z. (2017). Structural, textural and morphological characteristics of tannins from Acacia mearnsii encapsulated using sol-gel methods: applications as antimicrobial agents. Colloids Surf. B Biointerfaces 151, 26–33. doi: 10.1016/j.colsurfb.2016.11.041
Duits, L. A., Ravensbergen, B., Rademaker, M., Hiemstra, P. S., and Nibbering, P. H. (2002). Expression of beta-defensin 1 and 2 mRNA by human monocytes, macrophages and dendritic cells. Immunology 106, 517–525. doi: 10.1046/j.1365-2567.2002.01430.x
Duval, A., and Avérous, L. (2016). Characterization and physicochemical properties of condensed tannins from Acacia catechu. J. Agric. Food Chem. 64, 1751–1760. doi: 10.1021/acs.jafc.5b05671
Edgerton, M., and Koshlukova, S. E. (2000). Salivary histatin 5 and its similarities to the other antimicrobial proteins in human saliva. Adv. Dent. Res. 14, 16–21. doi: 10.1177/08959374000140010201
Espinel-Ingroff, A. (2008). Mechanisms of resistance to antifungal agents: yeasts and filamentous fungi. Rev. Iberoam. Micol. 25, 101–106. doi: 10.1016/S1130-1406(08)70027-5
Espinel-Ingroff, A. (2009). Novel antifungal agents, targets or therapeutic strategies for the treatment of invasive fungal diseases: a review of the literature (2005–2009). Rev. Iberoam. Micol. 26, 15–22. doi: 10.1016/S1130-1406(09)70004-X
Fan, J. T., Kuang, B., Zeng, G. Z., Zhao, S. M., Ji, C. J., Zhang, Y. M., et al. (2011). Biologically active arborinane-type triterpenoids and anthraquinones from Rubia yunnanensis. J. Nat. Prod. 74, 2069–2080. doi: 10.1021/np2002918
Ferreira, A. V., Prado, C. G., Carvalho, R. R., Dias, K. S. T., and Dias, A. L. T. (2013). Candida albicans and non-C. albicans Candida species: comparison of biofilm production and metabolic activity in biofilms, and putative virulence properties of isolates from hospital environments and infections. Mycopathologia 175, 265–272. doi: 10.1007/s11046-013-9638-z
Flowers, S. A., Colón, B., Whaley, S. G., Schuler, M. A., and Rogers, P. D. (2015). Contribution of clinically derived mutations in ERG11 to azole resistance in Candida albicans. Antimicrob. Agents Chemother. 59, 450–460. doi: 10.1128/AAC.03470-14
Fothergill, A. W., Sutton, D. A., McCarthy, D. I., and Wiederhold, N. P. (2014). Impact of new antifungal breakpoints on antifungal resistance in Candida species. J. Clin. Microbiol. 52, 994–997. doi: 10.1128/JCM.03044-13
Francois, I. E., Aerts, A. M., Cammue, B. P., and Thevissen, K. (2005). Currently used antimycotics: spectrum, mode of action and resistance occurrence. Curr. Drug Targets 6, 895–907. doi: 10.2174/138945005774912744
Franz, R., Kelly, S. L., Lamb, D. C., Kelly, D. E., Ruhnke, M., and Morschhauser, J. (1998). Multiple molecular mechanisms contribute to a stepwise development of fluconazole resistance in clinical Candida albicans strains. Antimicrob. Agents Chemother. 42, 3065–3072.
Garey, K. W., Rege, M., Pai, M. P., Mingo, D. E., Suda, K. J., Turpin, R. S., et al. (2006). Time to initiation of fluconazole therapy impacts mortality in patients with candidemia: a multi-institutional study. Clin. Infect. Dis. 43, 25–31. doi: 10.1086/504810
Gaur, M., Puri, N., Manoharlal, R., Rai, V., Mukhopadhayay, G., Choudhury, D., et al. (2008). MFS transportome of the human pathogenic yeast Candida albicans. BMC Genomics 9:579. doi: 10.1186/1471-2164-9-579
Gavanji, S., Zaker, S. R., Nejad, Z. G., Bakhtari, A., Bidabadi, E. S., and Larki, B. (2015). Comparative efficacy of herbal essences with amphotricin B and ketoconazole on Candida albicans in the in vitro condition. Integr. Med. Res. 4, 112–118. doi: 10.1016/j.imr.2015.01.003
Georgopapadakou, N. H. (1998). Antifungals: mechanism of action and resistance, established and novel drugs. Curr. Opin. Microbiol. 1, 547–557. doi: 10.1016/S1369-5274(98)80087-8
Girmenia, C. (2009). New generation azole antifungals in clinical investigation. Exp. Opin. Invest. Drugs 18, 1279–1295. doi: 10.1517/13543780903176407
Gołąbek, K., Strzelczyk, J. K., Owczarek, A., Cuber, P., Ślemp-Migiel, A., and Wiczkowski, A. (2015). Selected mechanisms of molecular resistance of Candida albicans to azole drugs. Acta Biochim. Pol. 62, 247–251. doi: 10.18388/abp.2014_940
Goncalves, M. J., Piras, A., Porcedd, S., Marongiu, B., Falconieri, D., Cavaleiro, C., et al. (2015). Antifungal activity of extracts from Cynomorium coccineum growing wild in Sardinia island (Italy). Nat. Prod. Res. 29, 2247–2250. doi: 10.1080/14786419.2014.1000892
Gonçalves, S. S., Souza, A. C., Chowdhary, A., Meis, J. F., and Colombo, A. L. (2016). Epidemiology and molecular mechanisms of antifungal resistance in Candida and Aspergillus. Mycoses 59, 198–219. doi: 10.1111/myc.12469
Gregoire, A. T., Lang, W. R., and Ward, K. (1959). The qualitative identification of free amino acids in human vaginal fluid. Ann. N. Y. Acad. Sci. 83, 185–188. doi: 10.1111/j.1749-6632.1960.tb40891.x
Groll, A. H., Gea-Banacloche, J. C., Glasmacher, A., Just-Nuebling, G., Maschmeyer, G., and Walsh, T. J. (2003). Clinical pharmacology of antifungal compounds. Infect. Dis. Clin. North Am. 17, 159–191. doi: 10.1016/S0891-5520(02)00068-5
Groll, A. H., Piscitelli, S. C., and Walsh, T. J. (1998). Clinical pharmacology of systemic antifungal agents: a comprehensive review of agents in clinical use, current investigational compounds, and putative targets for antifungal drug development. Adv. Pharmacol. 44, 343–500. doi: 10.1016/S1054-3589(08)60129-5
Grossman, N. T., Chiller, T. M., and Lockhart, S. R. (2014). Epidemiology of echinocandin resistance in Candida. Curr. Fungal Infect. Rep. 8, 243–248. doi: 10.1007/s12281-014-0209-7
Grossman, N. T., Pham, C. D., Cleveland, A. A., and Lockhart, S. R. (2015). Molecular mechanisms of fluconazole resistance in Candida parapsilosis isolates from a U.S. surveillance system. Antimicrob. Agents Chemother. 59, 1030–1037. doi: 10.1128/AAC.04613-14
Gualco, L., Debbia, E. A., Bandettini, R., Pescetto, L., Cavallero, A., Ossi, M. C., et al. (2007). Antifungal resistance in Candida spp. isolated in Italy between 2002 and 2005 from children and adults. Int. J. Antimicrob. Agents 29, 179–184. doi: 10.1016/j.ijantimicag.2006.08.047
Guery, B. P., Arendrup, M. C., Auzinger, G., Azoulay, E., Sá, M. B., Johnson, E. M., et al. (2009). Management of invasive candidiasis and candidemia in adult non-neutropenic intensive care unit patients: part I. Epidemiology and diagnosis. Intensive Care Med. 35, 55–62. doi: 10.1007/s00134-008-1338-7
Han, J., Jyoti, M. A., Song, H. Y., and Jang, W. S. (2016). Antifungal activity and action mechanism of histatin 5-halocidin hybrid peptides against Candida ssp. PLoS One 11:e0150196. doi: 10.1371/journal.pone.0150196
Harder, J., Bartels, J., Christophers, E., and Schroder, J. M. (1997). A peptide antibiotic from human skin. Nature 387:861. doi: 10.1038/43088
Haridas, M., Anderson, B. F., and Baker, E. N. (1995). Structure of human diferric lactoferrin refined at 2.2 A resolution. Acta Crystallogr. D Biol. Crystallogr. 51(Pt 5), 629–646. doi: 10.1107/S0907444994013521
He, X., Ma, Y., Yi, G., Wu, J., Zhou, L., and Guo, H. (2017). Chemical composition and antifungal activity of Carica papaya Linn. seeds essential oil against Candida spp. Lett. Appl. Microbiol. 64, 350–354. doi: 10.1111/lam.12711
Herrera, C. L., Alvear, M., Barrientos, L., Montenegro, G., and Salazar, L. A. (2010). The antifungal effect of six commercial extracts of Chilean propolis on Candida spp. Cienc. Investig. Agrar. 37, 75–84. doi: 10.4067/S0718-16202010000100007
Hertz, C. J., Wu, Q., Porter, E. M., Zhang, Y. J., Weismüller, K. H., Godowski, P. J., et al. (2003). Activation of Toll-like receptor 2 on human tracheobronchial epithelial cells induces the antimicrobial peptide human beta defensin-2. J. Immunol. 171, 6820–6826. doi: 10.4049/jimmunol.171.12.6820
Hise, A. G., Tomalka, J., Ganesan, S., Patel, K., Hall, B. A., Brown, G. D., et al. (2009). An essential role for the NLRP3 inflammasome in host defense against the human fungal pathogen Candida albicans. Cell Host Microbe 5, 487–497. doi: 10.1016/j.chom.2009.05.002
Hoover, D. M., Chertov, O., and Lubkowski, J. (2001). The structure of human -defensin-1: new insights into structural properties of defensins. J. Biol. Chem. 276, 39021–39026. doi: 10.1074/jbc.M103830200
Hoover, D. M., Rajashankar, K. R., Blumenthal, R., Puri, A., Oppenheim, J. J., Chertov, O., et al. (2000). The structure of human beta-defensin-2 shows evidence of higher order oligomerization. J. Biol. Chem. 275, 32911–32918. doi: 10.1074/jbc.M006098200
Isa, A. I., Awouafack, M. D., Dzoyem, J. P., Aliyu, M., Magaji, R. A., Ayo, J. O., et al. (2014). Some Strychnos spinosa (Loganiaceae) leaf extracts and fractions have good antimicrobial activities and low cytotoxicities. BMC Complement. Altern. Med. 14:456. doi: 10.1186/1472-6882-14-456
Jiang, C., Dong, D., Yu, B., Cai, G., Wang, X., Ji, Y., et al. (2013). Mechanisms of azole resistance in 52 clinical isolates of Candida tropicalis in China. J. Antimicrob. Chemother. 68, 778–785. doi: 10.1093/jac/dks481
Junqueira, J. C., Vilela, S. F., Rossoni, R. D., Barbosa, J. O., Costa, A. C., Rasteiro, V. M., et al. (2012). Oral colonization by yeasts in HIV-positive patients in Brazil. Rev. Inst. Med. Trop. São Paulo 54, 17–24. doi: 10.1590/S0036-46652012000100004
Kanafani, Z. A., and Perfect, J. R. (2008). Antimicrobial resistance: resistance to antifungal agents: mechanisms and clinical impact. Clin. Infect. Dis. 46, 120–128. doi: 10.1086/524071
Katiyar, S., Pfaller, M., and Edlind, T. (2006). Candida albicans and Candida glabrata clinical isolates exhibiting reduced echinocandin susceptibility. Antimicrob. Agents Chemother. 50, 2892–2894. doi: 10.1128/AAC.00349-06
Katiyar, S. K., and Edlind, T. D. (2001). Identification and expression of multidrug resistance-related ABC transporter genes in Candida krusei. Med. Mycol. 39, 109–116. doi: 10.1080/mmy.39.1.109.116
Kaur, R., Dhakad, M. S., Goyal, R., Haque, A., and Mukhopadhyay, G. (2016). Identification and antifungal susceptibility testing of Candida species: a comparison of Vitek-2 system with conventional and molecular methods. J. Glob. Infect. Dis. 8, 139–146. doi: 10.4103/0974-777X.192969
Kim, J., and Sudbery, P. (2011). Candida albicans, a major human fungal pathogen. J. Microbiol. 49, 171–177. doi: 10.1007/s12275-011-1064-7
Koehler, P., Tacke, D., and Cornely, O. (2014). Our 2014 approach to candidemia. Mycoses 57, 519–524. doi: 10.1111/myc.12207
Kontoyiannis, D. P., Hachem, R., Lewis, R. E., Rivero, G. A., Torres, H. A., Thornby, J., et al. (2003). Efficacy and toxicity of caspofungin in combination with liposomal amphotericin B as primary or salvage treatment of invasive aspergillosis in patients with hematologic malignancies. Cancer 98, 292–299. doi: 10.1002/cncr.11479
Kontoyiannis, D. P., and Lewis, R. E. (2002). Antifungal drug resistance of pathogenic fungi. Lancet 359, 1135–1144. doi: 10.1016/S0140-6736(02)08162-X
Köse, Y. B., İşcan, G., Göger, F., Akalın, G., Demirci, B., and Başer, K. H. C. (2016). Chemical composition and biological activity of Centaurea baseri: new species from Turkey. Chem. Biodivers. 13, 1369–1379. doi: 10.1002/cbdv.201600070
Krishnakumari, V., Rangaraj, N., and Nagaraj, R. (2009). Antifungal activities of human beta-defensins HBD-1 to HBD-3 and their C-terminal analogs Phd1 to Phd3. Antimicrob. Agents Chemother. 53, 256–260. doi: 10.1128/AAC.00470-08
Kusch, H., Biswas, K., Schwanfelder, S., Engelmann, S., Rogers, P. D., Hecker, M., et al. (2004). A proteomic approach to understanding the development of multidrug-resistant Candida albicans strains. Mol. Genet. Genomics 271, 554–565. doi: 10.1007/s00438-004-0984-x
Kuse, E. R., Chetchotisakd, P., da Cunha, C. A., Ruhnke, M., Barrios, C., Raghunadharao, D., et al. (2007). Micafungin versus liposomal amphotericin B for candidaemia and invasive candidosis: a phase III randomised double-blind trial. Lancet 369, 1519–1527. doi: 10.1016/S0140-6736(07)60605-9
Kwamin, F., Nartey, N. O., Codjoe, F. S., and Newman, M. J. (2013). Distribution of Candida species among HIV-positive patients with oropharyngeal candidiasis in Accra, Ghana. J. Infect. Dev. Ctries. 7, 41–45. doi: 10.3855/jidc.2442
Lamping, E., Ranchod, A., Nakamura, K., Tyndall, J. D., Niimi, K., Holmes, A. R., et al. (2009). Abc1p is a multidrug efflux transporter that tips the balance in favor of innate azole resistance in Candida krusei. Antimicrob. Agents Chemother. 53, 354–369. doi: 10.1128/AAC.01095-08
Lewis, L. E., Bain, J. M., Lowes, C., Gow, N. A. R., and Erwig, L. P. (2012). Candida albicans infection inhibits macrophage cell division and proliferation. Fungal Genet. Biol. 49, 679–680. doi: 10.1016/j.fgb.2012.05.007
Lewis, R. E., and Kontoyiannis, D. P. (2001). Rationale for combination antifungal therapy. Pharmacotherapy 21, 149S–164S. doi: 10.1592/phco.21.12.149S.34505
Li, S. Y., Yang, Y. L., Chen, K. W., Cheng, H. H., Chiou, C. S., Wang, T. H., et al. (2006). Molecular epidemiology of long-term colonization of Candida albicans strains from HIV-infected patients. Epidemiol. Infect. 134, 265–269. doi: 10.1017/S0950268805004905
Li, Y. Y., Chen, W. Y., Li, X., Li, H. B., Li, H. Q., Wang, L., et al. (2013). Asymptomatic oral yeast carriage and antifungal susceptibility profile of HIV-infected patients in Kunming, Yunnan Province of China. BMC Infect. Dis. 13:46. doi: 10.1186/1471-2334-13-46
Lim, S. M., Ahn, K. B., Kim, C., Kum, J. W., Perinpanayagam, H., Gu, Y., et al. (2016). Antifungal effects of synthetic human β-defensin 3-C15 peptide. Restor. Dent. Endod. 41, 91–97. doi: 10.5395/rde.2016.41.2.91
Lionakis, M. S., and Netea, M. G. (2013). Candida and host determinants of susceptibility to invasive candidiasis. PLoS Pathog. 9:e1003079. doi: 10.1371/journal.ppat.1003079
Lo, H. J., Wang, J. S., Lin, C. Y., Chen, C. G., Hsiao, T. Y., Hsu, C. T., et al. (2005). Efg1 involved in drug resistance by regulating the expression of ERG3 in Candida albicans. Antimicrob. Agents Chemother. 49, 1213–1215. doi: 10.1128/AAC.49.3.1213-1215.2005
Looi, C. Y., D´Silva, E. C., Seow, H. F., Rosli, R., Ng, K. P., and Chong, P. P. (2005). Increased expression and hotspot mutations of the multidrug efflux transporter, CDR1 in azole-resistant Candida albicans isolates from vaginitis patients. FEMS Microbiol. Lett. 249, 283–289. doi: 10.1016/j.femsle.2005.06.036
Lopez-Ribot, J. L., McAtee, R. K., Lee, L. N., Kirkpatrick, W. R., White, T. C., Sanglard, D., et al. (1998). Distinct patterns of gene expression associated with development of fluconazole resistance in serial Candida albicans isolates from human immunodeficiency virus-infected patients with oropharyngeal candidiasis. Antimicrob. Agents Chemother. 2, 2932–2937.
Lortholary, O., Desnos-Ollivier, M., Sitbon, K., Fontanet, A., Bretagne, S., Dromer, F., et al. (2011). Recent exposure to caspofungin or fluconazole influences the epidemiology of candidemia: a prospective multicenter study involving 2,441 patients. Antimicrob. Agents Chemother. 55, 532–538. doi: 10.1128/AAC.01128-10
Lu, M., Li, T., Wan, J., Li, X., Yuan, L., and Sun, S. (2017). Antifungal effects of phytocompounds on Candida species alone and in combination with fluconazole. Int. J. Antimicrob. Agents 49, 125–136. doi: 10.1016/j.ijantimicag.2016.10.021
Luo, Y., McLean, D. T. F., Linden, G. J., McAuley, D. F., McMullan, R., and Lundy, F. T. (2017). The naturally occurring host defense peptide, LL-37, and its truncated mimetics KE-18 and KR-12 have selected biocidal and antibiofilm activities against Candida albicans, Staphylococcus aureus, and Escherichia coli in vitro. Front. Microbiol. 8:544. doi: 10.3389/fmicb.2017.00544
Lupetti, A., Brouwer, C. P. J. M., Bogaards, S. J. P., Welling, M. M., de Heer, E., Campa, M., et al. (2007). Human lactoferrin-derived peptide’s antifungal activities against disseminated Candida albicans infection. J. Infect. Dis. 196, 1416–1424. doi: 10.1086/522427
Lyons, C. N., and White, T. C. (2000). Transcriptional analyses of antifungal drug resistance in Candida albicans. Antimicrob. Agents Chemother. 44, 2296–2303. doi: 10.1128/AAC.44.9.2296-2303.2000
Marichal, P., Koymans, L., Willemsens, S., Bellens, D., Verhasselt, P., Luyten, W., et al. (1999). Contribution of mutations in the cytochrome P450 14 alpha-demethylase (Erg11p, Cyp51p) to azole resistance in Candida albicans. Microbiology 145(Pt 10), 2701–2713. doi: 10.1099/00221287-145-10-2701
Marichal, P., Vanden Bossche, H., Odds, F. C., Nobels, G., Warnock, D. W., Timmerman, V., et al. (1997). Molecular biological characterization of an azole-resistant Candida glabrata isolate. Antimicrob. Agents Chemother. 41, 2229–2237.
Martel, C. M., Parker, J. E., Bader, O., Weig, M., Gross, U., Warrilow, A. G. S., et al. (2010). A clinical isolate of Candida albicans with mutations in ERG11 (encoding sterol 14α-demethylase) and ERG5 (encoding C22 desaturase) is cross resistant to azoles and amphotericin B. Antimicrob. Agents Chemother. 54, 3578–3583. doi: 10.1128/AAC.00303-10
Martins, N., Barros, B., Henriques, M., Silva, S., and Ferreira, I. C. (2015a). Activity of phenolic compounds from plant origin against Candida species. Ind. Crops Prod. 74, 648–670. doi: 10.1016/j.indcrop.2015.05.067
Martins, N., Barros, L., Henriques, M., Silva, S., and Ferreira, I. C. (2015b). In vivo anti-Candida activity of phenolic extracts and compounds: future perspectives focusing on effective clinical interventions. Biomed. Res. Int. 2015:247382. doi: 10.1155/2015/247382
Martins, N., Barros, L., Santos-Buelga, C., Henriques, M., Silva, S., and Ferreira, I. C. (2015c). Evaluation of bioactive properties and phenolic compounds in different extracts prepared from Salvia officinalis L. Food Chem. 170, 378–385. doi: 10.1016/j.foodchem.2014.08.096
Matthaiou, D. K., Christodoulopoulou, T., and Dimopoulos, G. (2015). How to treat fungal infections in ICU patients. BMC Infect. Dis. 15:205. doi: 10.1186/s12879-015-0934-8
Maubon, D., Garnaud, C., Calandra, T., Sanglard, D., and Cornet, M. (2014). Resistance of Candida spp. to antifungal drugs in the ICU: where are we now? Intensive Care Med. 40, 1241–1255. doi: 10.1007/s00134-014-3404-7
Mayer, F. L., Wilson, D., and Hube, B. (2013). Candida albicans pathogenicity mechanisms. Virulence 4, 119–128. doi: 10.4161/viru.22913
McCormick, T. S., and Weinberg, A. (2000). Epithelial cell-derived antimicrobial peptides are multifunctional agents that bridge innate and adaptive immunity. Periodontol 54, 195–206. doi: 10.1111/j.1600-0757.2010.00373
Mehra, T., Köberle, M., Braunsdorf, C., Mailänder-Sanchez, D., Borelli, C., and Schaller, M. (2012). Alternative approaches to antifungal therapies. Exp. Dermatol. 21, 778–782. doi: 10.1111/exd.12004
Mesa-Arango, A. C., Scorzoni, L., and Zaragoza, O. (2012). It only takes one to do many jobs: amphotericin B as antifungal and immunomodulatory drug. Front. Microbiol. 3:286. doi: 10.3389/fmicb.2012.00286
Minooeianhaghighi, M. H., Sepehrian, L., and Shokri, H. (2017). Antifungal effects of Lavandula binaludensis and Cuminum cyminum essential oils against Candida albicans strains isolated from patients with recurrent vulvovaginal candidiasis. J. Mycol. Med. 27, 65–71. doi: 10.1016/j.mycmed.2016.09.002
Miyazaki, H., Miyazaki, Y., Geber, A., Parkinson, T., Hitchcock, C., Falconer, D. J., et al. (1998). Fluconazole resistance associated with drug efflux and increased transcription of a drug transporter gene, PDH1, in Candida glabrata. Antimicrob. Agents Chemother. 42, 1695–1701.
Moffa, E. B., Mussi, M. C. M., Xiao, Y., Garrido, S. S., Machado, M. A. A. M., Giampaolo, E. T., et al. (2015). Histatin 5 inhibits adhesion of C. albicans to reconstructed human oral epithelium. Front. Microbiol. 6:885. doi: 10.3389/fmicb.2015.00885
Mollica, A., Macedonio, G., Stefanucci, A., Costante, R., Carradori, S., Cataldi, V., et al. (2017). Arginine- and lysine-rich peptides: synthesis, characterization and antimicrobial activity. Lett. Drug Des. Discov. 14, 1–7. doi: 10.2174/1570180814666170213161341
Mondello, F., De Bernardis, F., Girolamo, A., Salvatore, G., and Cassone, A. (2003). In vitro and in vivo activity of tea tree oil against azole-susceptible and -resistant human pathogenic yeasts. J. Antimicrob. Chemother. 51, 1223–1229. doi: 10.1093/jac/dkg202
Moraes, R. C., Lana, A. J. D., Kaiser, S., Carvalho, A. R., Oliveira, L. F. S., Fuentefria, A. M., et al. (2015). Antifungal activity of Uncaria tomentosa (Willd.) D.C. against resistant non-albicans Candida isolates. Ind. Crops Prod. 69, 7–14. doi: 10.1016/j.indcrop.2015.01.033
Morales, G., Paredes, A., Sierra, P., and Loyola, L. A. (2008). Antimicrobial activity of three Baccharis species used in the traditional medicine of Northern Chile. Molecules 13, 790–794. doi: 10.3390/molecules13040790
Moran, G. P., Sanglard, D., Donnelly, S. M., Shanley, D. B., Sullivan, D. J., and Coleman, D. C. (1998). Identification and expression of multidrug transporters responsible for fluconazole resistance in Candida dubliniensis. Antimicrob. Agents Chemother. 42, 1819–1830.
Morey, A. T., de Souza, F. C., Santos, J. P., Pereira, C. A., Cardoso, J. D., de Almeida, R. S. C., et al. (2016). Antifungal activity of condensed tannins from Stryphnodendron adstringens: effect on Candida tropicalis growth and adhesion properties. Curr. Pharm. Biotechnol. 17, 365–375. doi: 10.2174/1389201017666151223123712
Morici, P., Fais, R., Rizzato, C., Tavanti, A., and Lupetti, A. (2016). Inhibition of Candida albicans biofilm formation by the synthetic lactoferricin derived peptide hLF1-11. PLoS One 11:e0167470. doi: 10.1371/journal.pone.0167470
Morio, F., Loge, C., Besse, B., Hennequin, C., and Le Pape, P. (2010). Screening for amino acid substitutions in the Candida albicans Erg11 protein of azole-susceptible and azole-resistant clinical isolates: new substitutions and a review of the literature. Diagn. Microbiol. Infect. Dis. 66, 373–384. doi: 10.1016/j.diagmicrobio.2009.11.006
Morschhäuser, J., Barker, K. S., Liu, T. T., Blaß-Warmuth, J., Homayouni, R., and Rogers, P. (2007). The transcription factor Mrr1p controls expression of the MDR1 efflux pump and mediates multidrug resistance in Candida albicans. PLoS Pathog. 3:e164. doi: 10.1371/journal.ppat.0030164
Mukherjee, P. K., Sheehan, D., Puzniak, L., Schlamm, H., and Ghannoum, M. A. (2011). Echinocandins: are they all the same? J. Chemother. 23, 319–325. doi: 10.1179/joc.2011.23.6.319
Mukherjee, P. K., Sheehan, D. J., Hitchcock, C. A., and Ghannoum, M. A. (2005). Combination treatment of invasive fungal infections. Clin. Microbiol. Rev. 18, 163–194. doi: 10.1128/CMR.18.1.163-194.2005
Mulaudzi, R. B., Ndhlala, A. R., Kulkarni, M. G., and Van Staden, J. (2012). Pharmacological properties and protein binding capacity of phenolic extracts of some Venda medicinal plants used against cough and fever. J. Ethnopharmacol. 143, 185–193. doi: 10.1016/j.jep.2012.06.022
Navarro-García, V. M., Rojas, G., Avilés, M., Fuentes, M., and Zepeda, G. (2011). In vitro antifungal activity of coumarin extracted from Loeselia mexicana Brand. Mycoses 54, e569–e571. doi: 10.1111/j.1439-0507.2010.01993.x
Nett, J. E., and Andes, D. R. (2016). Antifungal agents: spectrum of activity, pharmacology, and clinical indications. Infect. Dis. Clin. North Am. 30, 51–83. doi: 10.1016/j.idc.2015.10.012
Newman, D. J., and Cragg, G. M. (2012). Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 75, 311–335. doi: 10.1021/np200906s
Ngouana, T. K., Mbouna, C. D. J., Kuipou, R. M. T., Tchuenmogne, M. A. T., Zeuko’o, E. M., Ngouana, V., et al. (2015). Potent and synergistic extract combinations from Terminalia catappa, Terminalia mantaly and Monodora tenuifolia against pathogenic yeasts. Medicines 2, 220–235. doi: 10.3390/medicines2030220
Niimi, M., Niimi, K., Takano, Y., Holmes, A. R., Fischer, F. J., Uehara, Y., et al. (2004). Regulated overexpression of CDR1 in Candida albicans confers multidrug resistance. J. Antimicrob. Chemother. 54, 999–1006. doi: 10.1093/jac/dkh456
Noel, T. (2012). The cellular and molecular defense mechanisms of the Candida yeasts against azole antifungals drugs. J. Mycol. Med. 22, 173–178. doi: 10.1016/j.mycmed.2012.04.004
Nordin, M. A., Wan Harun, W. H., Abdul Razak, F., and Musa, M. Y. (2014). Growth inhibitory response and ultrastructural modification of oral associated candidal reference strains (ATCC) by Piper betle L. extract. Int. J. Oral Sci. 6, 15–21. doi: 10.1038/ijos.2013.97
Orsi, N. (2004). The antimicrobial activity of lactoferrin: current status and perspectives. Biometals 17, 189–196. doi: 10.1023/B:BIOM.0000027691.86757.e2
Ortega, M., Marco, F., Soriano, A., Almela, M., Martínez, J. A., Pitart, C., et al. (2010). Candida spp. bloodstream infection: influence of antifungal treatment on outcome. J. Antimicrob. Chemother. 65, 562–568. doi: 10.1093/jac/dkp495
Otari, S. V., Patil, R. M., Ghosh, S. J., and Pawar, S. H. (2014). Green phytosynthesis of silver nanoparticles using aqueous extract of Manilkara zapota (L.) seeds and its inhibitory action against Candida species. Mater. Lett. 116, 367–369. doi: 10.1016/j.matlet.2013.11.066
Oxman, D. A., Chow, J. K., Frendl, G., Hadley, S., Hershkovitz, S., Ireland, P., et al. (2010). Candidemia associated with decreased in vitro fluconazole susceptibility: is Candida speciation predictive of the susceptibility pattern? J. Antimicrob. Chemother. 65, 1460–1465. doi: 10.1093/jac/dkq136
Papini, M., Simonetti, S., Franceschini, S., Scaringi, L., and Binazzi, M. (1982). Lysozyme distribution in healthy human skin. Arch. Dermatol. Res. 272, 167–170. doi: 10.1007/BF00510410
Papon, N., Noel, T., Florent, M., Gibot-Leclerc, S., Jean, D., Chastin, C., et al. (2007). Molecular mechanism of flucytosine resistance in Candida lusitaniae: contribution of the FCY2, FCY1, and FUR1 genes to 5-fluorouracil and fluconazole cross-resistance. Antimicrob. Agents Chemother. 51, 369–371. doi: 10.1128/AAC.00824-06
Pappas, P. G., Kauffman, C. A., Andes, D., Benjamin, D. K. Jr., Calandra, T. F., Edwards, J. E. Jr., et al. (2009). Clinical practice guidelines for the management of candidiasis: 2009 update by the infectious diseases society of America. Clin. Infect. Dis. 48, 503–535. doi: 10.1086/596757
Pappas, P. G., Kauffman, C. A., Andes, D. R., Clancy, C. J., Marr, K. A., Ostrosky-Zeichner, L., et al. (2016). Clinical practice guideline for the management of candidiasis: 2016 update by the infectious diseases society of America. Clin. Infect. Dis. 62, e1–e50. doi: 10.1093/cid/civ933
Paramythiotou, E., Frantzeskaki, F., Flevari, A., Armaganidis, A., and Dimopoulos, G. (2014). Invasive fungal infections in the ICU: how to approach, how to treat. Molecules 19, 1085–1119. doi: 10.3390/molecules19011085
Pasqualotto, A. C., Thiele, K. O., and Goldani, L. Z. (2010). Novel triazole antifungal drugs: focus on isavuconazole, ravuconazole and albaconazole. Curr. Opin. Investig. Drugs 11, 165–174.
Pasqualotto, C., and Denning, D. W. (2008). New and emerging treatments for fungal infections. J. Antimicrob. Chemother. 61, i19–i30. doi: 10.1093/jac/dkm428
Patil, S., Rao, R. S., Majumdar, B., and Anil, S. (2015). Clinical appearance of oral Candida infection and therapeutic strategies. Front. Microbiol. 6:1391. doi: 10.3389/fmicb.2015.01391
Perea, S., Gonzalez, G., Fothergill, A. W., Sutton, D. A., and Rinaldi, M. G. (2002a). In vitro activities of terbinafine in combination with fluconazole, itraconazole, voriconazole, and posaconazole against clinical isolates of Candida glabrata with decreased susceptibility to azoles. J. Clin. Microbiol. 40, 1831–1833. doi: 10.1128/JCM.40.5.1831-1833.2002
Perea, S., López-Ribot, J. L., Wickes, B. L., Kirkpatrick, W. R., Dib, O. P., Bachmann, S. P., et al. (2002b). Molecular mechanisms of fluconazole resistance in Candida dubliniensis isolates from human immunodeficiency virus-infected patients with oropharyngeal candidiasis. Antimicrob. Agents Chemother. 46, 1695–1703. doi: 10.1128/AAC.46.6.1695-1703.2002
Pereira, A. M. S., Hernandes, C., Pereira, S. I. V., Bertoni, B. W., Franca, S. C., Pereira, P. S., et al. (2014). Evaluation of anticandidal and antioxidant activities of phenolic compounds from Pyrostegia venusta (Ker Gawl.) Miers. Chem. Biol. Interact. 224, 136–141. doi: 10.1016/j.cbi.2014.10.023
Perlin, D. S. (2007). Resistance to echinocandin class antifungal drugs. Resist. Updat. 10, 121–130. doi: 10.1016/j.drup.2007.04.002
Perlin, D. S. (2011). Current perspectives on echinocandin class drugs. Future Microbiol. 6, 441–457. doi: 10.2217/fmb.11.19
Perlin, D. S. (2015). Mechanisms of echinocandin antifungal drug resistance. Ann. N. Y. Acad. Sci. 1354, 1–11. doi: 10.1111/nyas.12831
Perman, J., Canton, E., and Espinel-Ingroff, A. (2009). Antifungal drug resistance mechanisms. Expert Rev. Anti Infect. Ther. 7, 453–460. doi: 10.1586/eri.09.18
Petrikkos, G., and Skiada, A. (2007). Recent advances in antifungal chemotherapy. Int. J. Antimicrob. Agents 30, 108–117. doi: 10.1016/j.ijantimicag.2007.03.009
Pfaller, M. A. (2012). Antifungal drug resistance: mechanisms, epidemiology, and consequences for treatment. Am. J. Med. 125, S3–S13. doi: 10.1016/j.amjmed.2011.11.001
Pfaller, M. A., Andes, D. R., Diekema, D. J., Horn, D. L., Reboli, A. C., Rotstein, C., et al. (2014). Epidemiology and outcomes of invasive candidiasis due to non-albicans species of Candida in 2,496 patients: data from the prospective antifungal therapy (PATH) registry 2004–2008. PLoS One 9:e101510. doi: 10.1371/journal.pone.0101510
Pfaller, M. A., Castanheira, M., Messer, S. A., Moet, G. J., and Jones, R. N. (2010a). Variation in Candida spp. distribution and antifungal resistance rates among blood stream infection isolates by patient age: report from the SENTRY Antimicrobial Surveillance Program (2008-2009). Diagn. Microbiol. Infect. Dis. 68, 278–283. doi: 10.1016/j.diagmicrobio.2010.06.015
Pfaller, M. A., Diekema, D. J., Gibbs, D. L., Newell, V. A., Ellis, D., Tullio, V., et al. (2010b). Results from the ARTEMIS DISK global antifungal surveillance study, 1997 to 2007: a 10.5-year analysis of susceptibilities of Candida species to fluconazole and voriconazole as determined by CLSI standardized disk diffusion. J. Clin. Microbiol. 48, 1366–1377. doi: 10.1128/JCM.02117-09
Pfaller, M. A., and Diekema, D. J. (2007). Epidemiology of invasive Candidiasis: a persistent public health problem. Clin. Microbiol. Rev. 20, 133–163. doi: 10.1128/CMR.00029-06
Pfaller, M. A., Messer, S. A., Woosley, L. N., Jones, R. N., and Castanheira, M. (2013). Echinocandin and triazole antifungal susceptibility profiles of opportunistic yeast and mould clinical isolates (2010–2011): application of new CLSI clinical breakpoints and epidemiological cutoff values to characterize geographic and temporal trends of antifungal resistance. J. Clin. Microbiol. 51, 2571–2581. doi: 10.1128/JCM.00308-13
Pitman, S. K., Drew, R. H., and Perfect, J. R. (2011). Addressing current medical needs in invasive fungal infection prevention and treatment with new antifungal agents, strategies and formulations. Expert Opin. Emerg. Drugs 16, 559–586. doi: 10.1517/14728214.2011.607811
Prasad, R., and Kapoor, K. (2005). Multidrug resistance in yeast Candida. Int. Rev. Cytol. 242, 215–248. doi: 10.1016/S0074-7696(04)42005-1
Prasad, R., and Rawal, M. K. (2014). Efflux pump proteins in antifungal resistance. Front. Pharmacol. 5:202. doi: 10.3389/fphar.2014.00202
Prasad, R., Shah, A. H., and Rawal, M. K. (2016). “Antifungals: mechanism of action and drug resistance,” in Yeast Membrane Transporter Advances in Experimental Medicine and Biology, eds J. Ramos, H. Sychrová, and M. Kschischo (Cham: Springer International Publishing), 327–349.
Puri, N., Krishnamurthy, S., Habib, S., Hasnain, S. E., Goswami, S. K., and Prasad, R. (1999). CDR1, a multidrug resistance gene from Candida albicans, contains multiple regulatory domains in its promoter and the distal AP-1 element mediates its induction by miconazole. FEMS Microbiol. Lett. 180, 213–219. doi: 10.1111/j.1574-6968.1999.tb08798.x
Raman, S. B., Nguyen, M. H., Cheng, S., Badrane, H., Iczkowski, K. A., Wegener, M., et al. (2013). A competitive infection model of hematogenously disseminated candidiasis in mice redefines the role of Candida albicans IRS4 in pathogenesis. Infect. Immun. 81, 1430–1438. doi: 10.1128/IAI.00743-12
Raut, J. S., Shinde, R. B., Chauhan, N. M., and Karuppayil, S. M. (2014). Phenylpropanoids of plant origin as inhibitors of biofilm formation by Candida albicans. J. Microbiol. Biotechnol. 24, 1216–1225. doi: 10.4014/jmb.1402.02056
Rautemaa, R., and Ramage, G. (2011). Oral candidosis–clinical challenges of a biofilm disease. Crit. Rev. Microbiol. 37, 328–336. doi: 10.3109/1040841X.2011.585606
Razzaghi-Abyaneh, M., Sadeghi, G., Zeinali, E., Alirezaee, M., Ghahfarokhi, M. S., Amani, A., et al. (2014). Species distribution and antifungal susceptibility of Candida spp. isolated from superficial candidiasis in outpatients in Iran. J. Mycol. Med. 24, 43–50. doi: 10.1016/j.mycmed.2014.01.004
Reichling, J., Schnitzler, P., Suschke, U., and Saller, R. (2009). Essential oils of aromatic plants with antibacterial, antifungal, antiviral, and cytotoxic properties – an overview. Forsch. Komplementmed. 16, 79–90. doi: 10.1159/000207196
Ren, B., Huan-Qin, D., Gang, P., Yao-Jun, T., Ying, Z., Na, Y., et al. (2014). ABC transporters coupled with the elevated ergosterol contents contribute to the azole resistance and amphotericin B susceptibility. Appl. Microbiol. Biotechnol. 98, 2609–2616. doi: 10.1007/s00253-013-5425-5
Ricardo, E., Miranda, I. M., Faria-Ramos, I., Silva, R. M., Rodrigues, A. G., and Pina-Vaz, C. (2014). In vivo and in vitro acquisition of resistance to voriconazole by Candida krusei. Antimicrob. Agents Chemother. 58, 4604–4611. doi: 10.1128/AAC.02603-14
Rodrigues, M. E., Silva, S., Azeredo, J., and Henriques, M. (2014). Novel strategies to fight Candida species infection. Crit. Rev. Microbiol. 10, 1–13. doi: 10.3109/1040841X.2014.974500
Romero, A. L., Romero, R. B., Silva, E. L., Diniz, S. P. S. S., Oliveira, R. R., and Vida, J. B. (2012). Composição química e atividade do óleo essencial de Origanum vulgare sobre fungos fitopatogênicos. UNOPAR Cien. Ciênc. Biol. Saúde. 14, 231–235. doi: 10.17921/2447-8938.2012v14n4p25p
Safdar, A., Ma, J., Saliba, F., Dupont, B., Wingard, J. R., Hachem, R. Y., et al. (2010). Drug-induced nephrotoxicity caused by amphotericin B lipid complex and liposomal amphotericin B: a review and meta-analysis. Medicine 89, 236–244. doi: 10.1097/MD.0b013e3181e9441b
Saier, M. H. Jr., Beatty, J. T., Goffeau, A., Harley, K. T., Heijne, W. H. M., Huang, S. C., et al. (1999). The major facilitator superfamily. J. Mol. Microbiol. Biotechnol. 1, 257–279.
Samaranayake, Y. H., Samaranayake, L. P., Wu, P. C., and So, M. (1997). The antifungal effect of lactoferrin and lysozyme on Candida krusei and Candida albicans. Apmis 105, 875–883. doi: 10.1111/j.1699-0463.1997.tb05097.x
Sánchez, M. A., Turina, A. V., García, D. A., Nolan, M. V., and Perillo, M. A. (2004). Surface activity of thymol: implications for an eventual pharmacological activity. Colloids Surf. B Biointerfaces 34, 77–86. doi: 10.1016/j.colsurfb.2003.11.007
Sanglard, D. (2016). Emerging threats in antifungal-resistant fungal pathogens. Front. Med. 3:11. doi: 10.3389/fmed.2016.00011
Sanglard, D., Coste, A., and Ferrari, S. (2009). Antifungal drug resistance mechanisms in fungal pathogens from the perspective of transcriptional gene regulation. FEMS Yeast Res. 9, 1029–1050. doi: 10.1111/j.1567-1364.2009.00578.x
Sanglard, D., Ischer, F., and Bille, J. (2001). Role of ATP-binding-cassette transporter genes in high-frequency acquisition of resistance to azole antifungals in Candida glabrata. Antimicrob. Agents Chemother. 45, 1174–1183. doi: 10.1128/AAC.45.4.1174-1183.2001
Sanglard, D., Ischer, F., Calabrese, D., Majcherczyk, P. A., and Bille, J. (1999). The ATP binding cassette transporter gene CgCDR1 from Candida glabrata is involved in the resistance of clinical isolates to azole antifungal agents. Antimicrob. Agents Chemother. 43, 2753–2765.
Sanglard, D., Ischer, F., Parkinson, T., Falconer, D., and Bille, J. (2003). Candida albicans mutations in the ergosterol biosynthetic pathway and resistance to several antifungal agents. Antimicrob. Agents Chemother. 47, 2404–2412. doi: 10.1128/AAC.47.8.2404-2412.2003
Sanglard, D., and Odds, F. C. (2002). Resistance of Candida species to antifungal agents: molecular mechanisms and clinical consequences. Lancet Infect. Dis. 2, 73–85. doi: 10.1016/S1473-3099(02)00181-0
Sanguinetti, M., Postearo, B., and Lass-Florl, C. (2015). Antifungal drug resistance among Candida species: mechanisms and clinical impact. Mycoses 58, 2–13. doi: 10.1111/myc.12330
Sanguinetti, M., Posteraro, B., Fiori, B., Ranno, S., Torelli, R., and Fadda, G. (2005). Mechanisms of azole resistance in clinical isolates of Candida glabrata collected during a hospital survey of antifungal resistance. Antimicrob. Agents Chemother. 49, 668–679. doi: 10.1128/AAC.49.2.668-679.2005
Sant’Ana Pde, L., Milan, E. P., Martinez, R., Queiroz-Telles, F., Ferreira, M. S., Alcântara, A. P., et al. (2002). Multicenter Brazilian study of oral Candida species isolated from AIDS patients. Mem. Inst. Oswaldo Cruz. 97, 253–257. doi: 10.1590/S0074-02762002000200019
Santos, K. K. A., Matias, E. F. F., Sobral-Souza, C. E., Tintino, S. R., Morais-Braga, M. F. B., Guedes, G. M. M., et al. (2013). Trypanocide, cytotoxic, and anti-Candida activities of natural products: Hyptis martiusii Benth. Eur. J. Integr. Med. 5, 427–431. doi: 10.1016/j.eujim.2013.06.001
Sardi, J. C. O., Scorzoni, L., Bernardi, T., Fusco-Almeida, A. M., and Giannini, M. M. (2013). Candida species: current epidemiology, pathogenicity, biofilm formation, natural antifungal products and new therapeutic options. J. Med. Microbiol. 62, 10–24. doi: 10.1099/jmm.0.045054-0
Schibli, D. J., Hunter, H. N., Aseyev, V., Starner, T. D., Wiencek, J. M., McCray, P. B. Jr., et al. (2002). The solution structures of the human β-defensins lead to a better understanding of the potent bactericidal activity of HBD3 against Staphylococcus aureus. J. Biol. Chem. 277, 8279–8289. doi: 10.1074/jbc.M108830200
Schroeder, B. O., Wu, Z., Nuding, S., Groscurth, S., Marcinowski, M., Beisner, J., et al. (2011). Reduction of disulphide bonds unmasks potent antimicrobial activity of human beta-defensin 1. Nature 469, 419–423. doi: 10.1038/nature09674
Schwarzmuller, T., Ma, B., Hiller, E., Istel, F., Tscherner, M., Brunke, S., et al. (2014). Systematic phenotyping of a large-scale Candida glabrata deletion collection reveals novel antifungal tolerance genes. PLoS Pathog. 10:e1004211. doi: 10.1371/journal.ppat.1004211
Seleem, D., Pardi, V., and Murata, R. M. (2017). Review of flavonoids: a diverse group of natural compounds with anti-Candida albicans activity in vitro. Arch. Oral Biol. 76, 73–83. doi: 10.1016/j.archoralbio.2016.08.030
Selmecki, A., Forche, A., and Berman, J. (2006). Aneuploidy and isochromosome formation in drug-resistant Candida albicans. Science 313, 367–370. doi: 10.1126/science.1128242
Seneviratne, C. J., Jin, L., and Samaranayake, L. P. (2008). Biofilm lifestyle of Candida: a minireview. Oral Dis. 14, 582–590. doi: 10.1111/j.1601-0825.2007.01424.x
Serena, C., Fernández-Torres, B., Pastor, F. J., Trilles, L., Lazéra, M. S., Nolard, N., et al. (2005). In vitro interactions of micafungin with other antifungal drugs against clinical isolates of four species of Cryptococcus. Antimicrob. Agents Chemother. 49, 2994–2996. doi: 10.1128/AAC.49.7.2994-2996.2005
Serpa, R., França, E. J., Furlaneto-Maia, L., Andrade, C. G., Diniz, A., and Furlaneto, M. C. (2012). In vitro antifungal activity of the flavonoid baicalein against Candida species. J. Med. Microbiol. 61, 1704–1708. doi: 10.1099/jmm.0.047852-0
Seyedmousavi, S., Rafati, H., Ilkit, M., Tolooe, A., Hedayati, M. T., and Verweij, P. (2017). “Systemic antifungal agents: current status and projected future developments,” in Methods in Molecular Biology, ed. T. Lion (New York, NY: Springer), 107–139.
Shahid, M., Shahzad, A., Sobia, F., Sahai, A., Tripathi, T., Singh, A., et al. (2009). Plant natural products as a potential source for antibacterial agents: recent trends. Anti Infect. Agents Med. Chem. 8, 211–225. doi: 10.2174/187152109788680199
Shahzad, M., Sherry, L., Rajendran, R., Edwards, C. A., Combet, E., and Ramage, C. (2014). Utilising polyphenols for the clinical management of Candida albicans biofilms. Int. J. Antimicrob. Agents 44, 269–273. doi: 10.1016/j.ijantimicag.2014.05.017
Shapiro, R. S., Robbins, N., and Cowen, L. E. (2011). Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. Rev. 75, 213–267. doi: 10.1128/MMBR.00045-10
Sharifzadeh, A., Khosravi, A. R., Shokri, H., and Sharafi, G. (2015). Antifungal effect of Trachyspermum ammi against susceptible and fluconazole-resistant strains of Candida albicans. J. Mycol. Med. 25, 143–150. doi: 10.1016/j.mycmed.2015.03.008
Sharma, H., Yunus, G. Y., Mohapatra, A. K., Kulshrestha, R., Agrawal, R., and Kalra, M. (2016). Antimicrobial efficacy of three medicinal plants Glycyrrhiza glabra, Ficus religiosa, and Plantago major on inhibiting primary plaque colonizers and periodontal pathogens: an in vitro study. Indian J. Dent. Res. 27, 200–204. doi: 10.4103/0970-9290.183135
Sharma, Y., Khan, L. A., and Manzoor, N. (2016). Anti-Candida activity of geraniol involves disruption of cell membrane integrity and function. J. Med. Mycol. 26, 244–254. doi: 10.1016/j.mycmed.2016.04.004
Sheikh, N., Jahagirdar, V., Kothadia, S., and Nagoba, B. (2013). Antifungal drug resistance in Candida species. Eur. J. Gen. Med. 10, 254–258. doi: 10.29333/ejgm/82217
Shields, R. K., Nguyen, M. H., Du, C., Press, E., Cheng, S., and Clancy, C. J. (2011). Paradoxical effect of caspofungin against Candida bloodstream isolates is mediated by multiple pathways but eliminated in human serum. Antimicrob. Agents Chemother. 55, 2641–2647. doi: 10.1128/AAC.00999-10
Shoham, S., and Marr, K. A. (2012). Invasive fungal infections in solid organ transplant recipients. Future Microbiol. 7, 639–655. doi: 10.2217/fmb.12.28
Sienkiewicz, M., Lysakowska, M., Ciecwierz, J., Denys, P., and Kowalczyk, E. (2011). Antibacterial activity of thyme and lavender essential oils. Med. Chem. 7, 674–689. doi: 10.2174/157340611797928488
Siikala, E., Rautemaa, R., Richardson, M., Saxen, H., Bowyer, P., and Sanglard, D. (2010). Persistent Candida albicans colonization and molecular mechanisms of azole resistance in autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy (APECED) patients. J. Antimicrob. Chemother. 65, 2505–2513. doi: 10.1093/jac/dkq354
Silver, P. M., Oliver, B. G., and White, T. C. (2004). Role of Candida albicans transcription factor Upc2p in drug resistance and sterol metabolism. Eukaryot. Cell 3, 1391–1397. doi: 10.1128/EC.3.6.1391-1397.2004
Soll, D. R. (2008). Candida biofilms: is adhesion sexy? Curr. Biol. 18, R717–R720. doi: 10.1016/j.cub.2008.07.014
Sorensen, O. E., Thapa, D. R., Rosenthal, A., Liu, L., Roberts, A. A., and Ganz, T. (2005). Differential regulation of beta-defensin expression in human skin by microbial stimuli. J. Immunol. 174, 4870–4879. doi: 10.4049/jimmunol.174.8.4870
Souza, A. C., Fuchs, B. B., Pinhati, H. M., Siqueira, R. A., Hagen, F., Meis, J. F., et al. (2015). Candida parapsilosis resistance to fluconazole: molecular mechanisms and in vivo impact in infected Galleria mellonella larvae. Antimicrob. Agents Chemother. 59, 6581–6587. doi: 10.1128/AAC.01177-15
Souza, A. C. O., and Amaral, A. C. (2017). Antifungal therapy for systemic mycosis and the nanobiotechnology era: improving efficacy, biodistribution and toxicity. Front. Microbiol. 8:336. doi: 10.3389/fmicb.2017.00336
Spampinato, C., and Leonardi, D. (2013). Candida infections, causes, targets, and resistance mechanisms: traditional and alternative antifungal agents. Biomed Res. Int. 2013, 204–237. doi: 10.1155/2013/204237
Stiufiuc, R., Iacovita, C., Stiufiuc, G., Florea, A., Achim, M., and Lucaciu, C. M. (2015). A new class of pegylated plasmonic liposomes: synthesis and characterization. J. Colloid Interface Sci. 437, 17–23. doi: 10.1016/j.jcis.2014.09.023
Strzelczyk, J. K., Slemp-Migiel, A., Rother, M., Gołabek, K., and Wiczkowski, A. (2013). Nucleotide substitutions in the Candida albicans ERG11 gene of azole-susceptible and azole-resistant clinical isolates. Acta Biochim. Pol. 60, 547–552.
Tangarife-Castaño, V., Correa-Royero, C., Zapata-Londoño, B., Duran, C., Stanshenko, E., and Mesa-Arango, A. C. (2011). Anti-Candida albicans activity, cytotoxicity and interaction with antifungal drugs of essential oils and extracts from aromatic and medicinal plants. Infectio 15, 160–167. doi: 10.1016/S0123-9392(11)70080-7
Terças, A. G., Monteiro, A. S., Moffa, E. B., Dos Santos, J. R. A., de Sousa, E. M., Pinto, A. R. B., et al. (2017). Phytochemical characterization of Terminalia catappa Linn. extracts and their antifungal activities against Candida spp. Front. Microbiol. 8:595. doi: 10.3389/fmicb.2017.00595
Terças, A. L. G., Marques, S. G., Moffa, E. B., Alves, M. B., de Azevedo, C. M. P. S., Siqueira, W. L., et al. (2017). Antifungal drug susceptibility of Candida species isolated from HIV-positive patients recruited at a public hospital in São Luís, Maranhão, Brazil. Front. Microbiol. 8:298. doi: 10.3389/fmicb.2017.00298
Todorovic, V., Milenkovic, M., Vidovic, B., Todorovic, Z., and Sobajic, S. (2017). Correlation between antimicrobial, antioxidant activity, and polyphenols of alkalized/ non alkalized cocoa powders. J. Food Sci. 82, 1020–1027. doi: 10.1111/1750-3841.13672
Tomalka, J., Azodi, E., Narra, H. P., Patel, K., O’Neill, S., Cardwell, C., et al. (2015). β-Defensin 1 plays a role in acute mucosal defense against Candida albicans. J. Immunol. 194, 1788–1795. doi: 10.4049/jimmunol.1203239
Tomasinsig, L., and Zanetti, M. (2005). The cathelicidins – structure, function and evolution. Curr. Protein Pept. Sci. 6, 23–34. doi: 10.2174/1389203053027520
Torelli, R., Posteraro, B., Ferrari, S., La Sorda, M., Fadda, G., Sanglard, D., et al. (2008). The ATP-binding cassette transporter-encoding gene CgSNQ2 is contributing to the CgPDR1-dependent azole resistance of Candida glabrata. Mol. Microbiol. 68, 186–201. doi: 10.1111/j.1365-2958.2008.06143.x
Tsai, P. W., Chen, Y. T., Hsu, P. C., and Lan, C. Y. (2013). Study of Candida albicans and its interactions with the host: a mini review. BioMedicine 3, 51–64. doi: 10.1016/j.biomed.2012.12.004
Tsai, P. W., Yang, C. Y., Chang, H. T., and Lan, C. Y. (2011). Human antimicrobial peptide LL-37 inhibits adhesion of Candida albicans by interacting with yeast cell-wall carbohydrates. PLoS One 6:e17755. doi: 10.1371/journal.pone.0017755
Tsao, S., Rahkhoodaee, F., and Raymond, M. (2009). Relative contributions of the Candida albicans ABC transporters Cdr1p and Cdr2p to clinical azole resistance. Antimicrob. Agents Chemother. 53, 1344–1352. doi: 10.1128/AAC.00926-08
Tscherner, M., Schwarzmüller, T., and Kuchler, K. (2011). Pathogenesis and antifungal drug resistance of the human fungal pathogen Candida glabrata. Pharmaceuticals 4, 169–186. doi: 10.3390/ph4010169
Vale-Silva, L. A., Gonçalves, M. J., Cavaleiro, C., Salgueiro, L., and Pinto, E. (2010). Antifungal activity of the essential oil of Thymus viciosoi against Candida, Cryptococcus, Aspergillus and Dermatophyte species. Planta Med. 76, 882–888. doi: 10.1055/s-0029-1240799
Vallabhaneni, S., Mody, R. K., Walker, T., and Chiller, T. (2016). The global burden of fungal diseases. Infect. Dis. Clin. North Am. 30, 1–11. doi: 10.1016/j.idc.2015.10.004
van der Does, A. M., Joosten, S. A., Vroomans, E., Bogaards, S. J. P., van Meijgaarden, K. E., Ottenhoff, T. H. M., et al. (2012). The antimicrobial peptide hLF1-11 drives monocyte-dendritic cell differentiation toward dendritic cells that promote antifungal responses and enhance Th17 polarization. J. Innate Immun. 4, 284–292. doi: 10.1159/000332941
Vandeputte, P., Larcher, G., Bergès, T., Renier, G., Chabasse, D., and Bouchara, J. P. (2005). Mechanisms of azole resistance in a clinical isolate of Candida tropicalis. Antimicrob. Agents Chemother. 49, 4608–4615. doi: 10.1128/AAC.49.11.4608-4615.2005
Vázquez-González, D., Perusquía-Ortiz, A. M., Hundeiker, M., and Bonifaz, A. (2013). Opportunistic yeast infections: candidiasis, cryptococcosis, trichosporonosis and geotrichosis. J. Dtsch. Dermatol. Ges. 11, 381–394. doi: 10.1111/ddg.12097
Vermes, A., Guchelaar, H. J., and Dankert, J. (2000). Flucytosine: a review of its pharmacology, clinical indications, pharmacokinetics, toxicity and drug interactions. J. Antimicrob. Chemother. 46, 171–179. doi: 10.1093/jac/46.2.171
Vollenbroich, R., Maeder, M. T., and Weilenmann, D. (2014). Congestive heart failure related to antifungal therapy with itraconazole. Int. J. Cardiol. 172, e170–e171. doi: 10.1016/j.ijcard.2013.12.057
Vukosavljevic, D., Custodio, W., Del Bel Cury, A. A., and Siqueira, W. L. (2012). The effect of histatin 5, adsorbed on PMMA and hydroxyapatite, on Candida albicans colonization. Yeast 29, 459–466. doi: 10.1002/yea.2925
Wächtler, B., Citiulo, F., Jablonowski, N., Förster, S., Dalle, F., Schaller, M., et al. (2012). Candida albicans-epithelial interactions: dissecting the roles of active penetration, induced endocytosis and host factors on the infection process. PLoS One 7:e36952. doi: 10.1371/journal.pone.0036952
Wang, H., Kong, F., Sorrell, T. C., Wang, B., McNicholas, P., Pantarat, N., et al. (2009). Rapid detection of ERG11 gene mutations in clinical Candida albicans isolates with reduced susceptibility to fluconazole by rolling circle amplification and DNA sequencing. BMC Microbiol. 9:167. doi: 10.1186/1471-2180-9-167
Wang, J. S., Yang, Y. L., Wu, C. G., Ouyang, K. J., Tseng, K. Y., Chen, C. G., et al. (2006). The DNA binding domain of CaNdt80p is required to activate CDR1 involved in drug resistance in Candida albicans. J. Med. Microbiol. 55, 1403–1411. doi: 10.1099/jmm.0.46650-0
Wirsching, S., Michel, S., Kohler, G., and Morschhauser, J. (2000). Activation of the multiple drug resistance gene MDR1 in fluconazole-resistant, clinical Candida albicans strains is caused by mutations in a transregulatory factor. J. Bacteriol. 182, 400–404. doi: 10.1128/JB.182.2.400-404.2000
Wong, S. S., Samaranayake, L. P., and Seneviratne, C. J. (2014). In pursuit of the ideal antifungal agent for Candida infections: high-throughput screening of small molecules. Drug Discov. Today 19, 1721–1730. doi: 10.1016/j.drudis.2014.06.009
Wu, T., Samaranayake, L. P., Leung, W. K., and Sullivan, P. A. (1999). Inhibition of growth and secreted aspartyl proteinase production in Candida albicans by lysozyme. J. Med. Microbiol. 48, 721–730. doi: 10.1099/00222615-48-8-721
Xu, Y., Burton, S., Kim, C., and Sismour, E. (2016). Phenolic compounds, antioxidant, and antibacterial properties of pomace extracts from four Virginia-grown grape varieties. J. Food Sci. Nutr. 4, 125–133. doi: 10.1002/fsn3.264
Yamaguchi, M. U., Garcia, F. P., Cortez, D. A., Ueda-Nakamura, T., Filho, B. P., and Nakamura, C. V. (2011). Antifungal effects of ellagitannin isolated from leaves of Ocotea odorifera (Lauraceae). Antonie Van Leeuwenhoek 99, 507–514. doi: 10.1007/s10482-010-9516-3
Yang, Y. L., and Lo, H. J. (2001). Mechanisms of antifungal agent resistance. J. Microbiol. Immunol. Infect. 34, 79–86.
Zhang, L., Xiao, M., Watts, M. R., Wang, H., Fan, X., Kong, F., et al. (2015). Development of fluconazole resistance in a series of Candida parapsilosis isolates from a persistent candidemia patient with prolonged antifungal therapy. BMC Infect. Dis. 15:340. doi: 10.1186/s12879-015-1086-6
Keywords: Candida infections, Candida, antifungals, resistance, alternative antifungal drugs
Citation: de Oliveira Santos GC, Vasconcelos CC, Lopes AJO, de Sousa Cartágenes MdS, Filho AKDB, do Nascimento FRF, Ramos RM, Pires ERRB, de Andrade MS, Rocha FMG and de Andrade Monteiro C (2018) Candida Infections and Therapeutic Strategies: Mechanisms of Action for Traditional and Alternative Agents. Front. Microbiol. 9:1351. doi: 10.3389/fmicb.2018.01351
Received: 10 February 2018; Accepted: 05 June 2018;
Published: 03 July 2018.
Edited by:
Octavio Luiz Franco, Universidade Católica de Brasília, BrazilReviewed by:
Adriano Mollica, Università degli Studi “G. d’Annunzio” Chieti - Pescara, ItalyCopyright © 2018 de Oliveira Santos, Vasconcelos, Lopes, de Sousa Cartágenes, Filho, do Nascimento, Ramos, Pires, de Andrade, Rocha and de Andrade Monteiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cristina de Andrade Monteiro, Y3Jpc3RpbmEubW9udGVpcm9AY2V1bWEuYnI=
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.