
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 24 October 2017
Sec. Food Microbiology
Volume 8 - 2017 | https://doi.org/10.3389/fmicb.2017.02049
 Karen K. Dittmann1†
Karen K. Dittmann1† Luíza T. Chaul2†
Luíza T. Chaul2† Sarah H. I. Lee3
Sarah H. I. Lee3 Carlos H. Corassin3
Carlos H. Corassin3 Carlos A. Fernandes de Oliveira3
Carlos A. Fernandes de Oliveira3 Elaine C. Pereira De Martinis4
Elaine C. Pereira De Martinis4 Virgínia F. Alves2
Virgínia F. Alves2 Lone Gram1*
Lone Gram1* Virginie Oxaran1
Virginie Oxaran1Staphylococcus aureus, a major food-poisoning pathogen, is a common contaminant in dairy industries worldwide, including in Brazil. We determined the occurrence of S. aureus in five dairies in Brazil over 8 months. Of 421 samples, 31 (7.4%) were positive for S. aureus and prevalence varied from 0 to 63.3% between dairies. Sixty-six isolates from the 31 samples were typed by Multi-Locus Sequence Typing to determine if these isolates were persistent or continuously reintroduced. Seven known sequence types (STs), ST1, ST5, ST30, ST97, ST126, ST188 and ST398, and four new ST were identified, ST3531, ST3540, ST3562 and ST3534. Clonal complex (CC) 1 (including the four new ST), known as an epidemic clone, was the dominant CC. However, there were no indications of persistence of particular ST. The resistance toward 11 antibiotic compounds was assessed. Twelve profiles were generated with 75.8% of strains being sensitive to all antibiotic classes and no Methicillin-resistant S. aureus (MRSA) strains were found. The enterotoxin-encoding genes involved in food-poisoning, e.g., sea, sed, see, and seg were targeted by PCR. The two toxin-encoding genes, sed and see, were not detected. Only three strains (4.5%) harbored seg and two of these also harbored sea. Despite the isolates being Methicillin-sensitive S. aureus (MSSA), the presence of CC1 clones in the processing environment, including some harboring enterotoxin encoding genes, is of concern and hygiene must have high priority to reduce contamination.
Staphylococcus aureus is an important opportunistic pathogen that can cause infections in warm-blooded animals and is a leading cause of nosocomial infections in humans and bovine mastitis (Peton and Le Loir, 2014). S. aureus is also one of the most common causative agents of food-poisoning (Balaban and Rasooly, 2000; Le Loir et al., 2003; Hennekinne et al., 2012; Kadariya et al., 2014) leading to 241,000 illnesses per year in the United States (Scallan et al., 2011) and is one of the main agents implicated in foodborne diseases in Brazil (Gomes et al., 2013). In foods, enterotoxigenic strains of S. aureus can produce heat-stable and protease resistant staphylococcal enterotoxins (SE) causing one of the most common foodborne intoxications (Balaban and Rasooly, 2000; Dinges et al., 2000; Le Loir et al., 2003; Argudín et al., 2010) leading to rapid clinical symptoms such as abdominal cramps, nausea, emesis, and diarrhea (Le Loir et al., 2003; Murray, 2005). More than 20 distinct SE have been described to date including SEA and SED (Jones et al., 2002; Asao et al., 2003; Kérouanton et al., 2007; Pinchuk et al., 2010; Sato’o et al., 2014; Solano et al., 2014; Arfatahery et al., 2016) responsible for the majority of staphylococcal food-poisoning (SFP), and also in fewer cases SEB, SEC, SEE, SEG, SEH, and SEI (Chen et al., 2004; Ikeda et al., 2005; Ostyn et al., 2010; Pinchuk et al., 2010). In 2012, 346 outbreaks caused by SE were reported in Europe (EFSA and ECDC, 2014). However, not every S. aureus strains carry SE-encoding genes (Lindberg et al., 2000; Le Loir et al., 2003) and the risk for human beings from different S. aureus may thus vary.
Staphylococcus aureus has been isolated from several foods including several ready-to-eat products (Asao et al., 2003; van Loo et al., 2007; Schmid et al., 2009; Ostyn et al., 2010; Rizek et al., 2011; Baumgartner et al., 2014; Hao et al., 2015) and improper staff hygiene as well as poor surface sanitation are some of the main causes of cross-contamination (Hatakka et al., 2000; Asao et al., 2003; Lues and Van Tonder, 2007; André et al., 2008). S. aureus is capable of surviving on dry stainless steel and it can easily be transferred from sponges to stainless steel surfaces and subsequently to food products (Kusumaningrum et al., 2003). In the dairy industry, S. aureus can be introduced at almost every step of the production. Beside handlers and environment, the raw milk can be one route of introduction as a consequence of subclinical or clinical bovine mastitis. S. aureus can be shed into the milk (Kérouanton et al., 2007; Peton and Le Loir, 2014) and produce SE which represents one of the most common food safety concerns from raw milk products (Fagundes et al., 2010; Silveira-Filho et al., 2014). In addition, SE can be produced in the product during storage if conditions allow growth of S. aureus.
Dairy production is an important industry worldwide. In Brazil, up to 30% of the milk production comes from small producers (Lee et al., 2012) and soft cheeses such as Minas frescal type are widely consumed. This particular cheese is non-ripened semi-fat and high moisture (Saboya et al., 1998) which offers suitable growth conditions for many pathogens including S. aureus that has frequently been the cause of foodborne diseases (Veras et al., 2008). In 1996, the Brazilian Ministry of Agriculture (MAPA) implemented microbiological regulations for cheeses, subsequently extending specific maximum S. aureus contamination levels (e.g., 103 CFU/g for coagulase positive staphylococci) for Minas frescal cheese (Ministério da Agricultura Brasil, 1997). However, our knowledge regarding S. aureus occurrence in Brazilian dairy plants is still limited.
Besides food-poisoning, S. aureus is a major concern due to its ability to acquire antibiotic resistance genes and to disseminate and to withstand treatment (Khan et al., 2000a,b; Holden et al., 2004; de Lencastre et al., 2007; Pesavento et al., 2007). Methicillin-resistant Staphylococcus aureus (MRSA) strains are responsible for the majority of nosocomial infection worldwide (ECDC, 2014; EFSA and ECDC, 2014) and there is an increase in MRSA detection in the community, livestock environment and food (Normanno et al., 2007; van Loo et al., 2007; EFSA and ECDC, 2014). Thus, despite being mostly problematic in infection, it raises concern that foods could potentially also be a vehicle of antibiotic resistant S. aureus strains. Epidemiological studies have been used to track the contamination origin and the diversity of clinical isolates for many years. However, particular focus has been on MRSA isolates (Chambers and Deleo, 2009). Randomly amplified polymorphic DNA (RAPD) has been used to determine the relatedness of S. aureus from cattle, pigs, chickens, and humans (Lee, 2003) concluding that humans can acquire MRSA infections from livestock via contaminated food products. This reinforces that contamination of dairy processing environment, milk and dairy products by S. aureus is a public health concern (Fagundes et al., 2010; Oliveira et al., 2011). However, few studies have assessed the global epidemiology of S. aureus of animal origin and their dissemination into the food chain (Lee et al., 2012; Sobral et al., 2012; Lima et al., 2013). Different from RAPD and among the various molecular typing methods, multi-locus sequence typing (MLST) (Enright et al., 2000) provides global epidemiology data. Indeed, MLST can be used to track contamination sources, to indicate persistence of particular clones and to identify epidemic clones. However, studies investigating the molecular characteristic of S. aureus from dairy processing environment and dairy products in Brazil are scarce.
The objectives of this study were: (i) to estimate the incidence of S. aureus contamination in the Brazilian dairy industry of the Southeast and Midwest regions, (ii) to determine the genetic diversity of the isolates by MLST as well as to evaluate their reoccurrence and eventually persistence, and (iii) to assess some of the virulence factors involved in health risk such as antibiotic resistance spreading by determining the antibiotic profile, the hemolytic trend potential and to investigate the possible food-poisoning ability of the isolates by detecting the presence of enterotoxin encoding genes.
Staphylococcus aureus isolates collected from Brazilian dairies were investigated in this study (Supplementary Table 1) and sampled as described in Section “Sampling Procedure and Detection of Putative S. aureus.” Isolation and phenotypic tests were performed using Brain–Heart Infusion (BHI) broth (Oxoid, United Kingdom), BHI agar [BHI broth, 1.5% agar (AppliChem, Germany)], Tryptic Soy Broth [TSB (Oxoid, United Kingdom)], Tryptic Soy Agar [TSA (Oxoid, United Kingdom)] supplemented with 0.6% yeast extract (TSAYE), DNase Test Agar with Methyl Green (Difco, Detroit, MI, United States), Mueller Hinton II Agar [MHA (BD Diagnostics, Franklin Lakes, NJ, United States)], Mannitol Salt Agar (MSA) and Baird Parker agar (Oxoid, United Kingdom) supplemented with egg-yolk emulsion (BP). Blood agar was prepared by supplementing TSA with defibrinated sheep blood (Statens Serum Institut, Denmark) at 5% (vol/vol). Unless otherwise specified, isolates were grown at 37°C and liquid cultures were incubated under shaking conditions at 250 rpm. The isolates were stored in BHI containing 20% (vol/vol) glycerol (Merck, Germany) at -80°C.
Five dairy processing plants in São Paulo state and Minas Gerais state (Southeast region) and Goiás state (Midwest region) in Brazil were sampled over an 8 months period from December 2013 to July 2014. In São Paulo state, two dairy plants were evaluated, one small-scale with a volume of 7,000 L cow’s milk processed per day from 60 producing farms, and the other of medium-scale that processed nearly 60,000 L per day collected from 150 farms. Two out of the three sampled dairies in Goiás state were small-scale dairies, processing about 3,000 L per day provided by around 30 farms. The third dairy was an artisanal cheese manufacturing plant, producing about 50 cheeses per day, using 250 L per day, supplied by just one cattle herd, raised on site where the cheese factory is located and using unpasteurized milk. Four categories of samples were investigated: raw material (bulked raw milk, pasteurized milk, brine, curd), food-contact surfaces (milk filter, vats, spatulas, paddle, knife, tank, picking machine strainer, hand handler, worker gloves, cheese box, shelf, gallon of milk, cloth mold, mold, plastic crates, curd tube, tables), non-food contact surfaces (pallet, wall, floor, drain, water, cleaning brush, sink, tables, gloves, and staff’s boot), and final product (cheese surfaces, cheese) as shown in Table 1 and Supplementary Table 1. The five dairies followed the same cheese processing procedure using milk and no lactic acid bacteria in the production. The milk was pasteurized except in one dairy where they used unpasteurized milk and it was chosen to represent this type of Brazilian dairy (corresponding to 20–40% of dairies in this country).
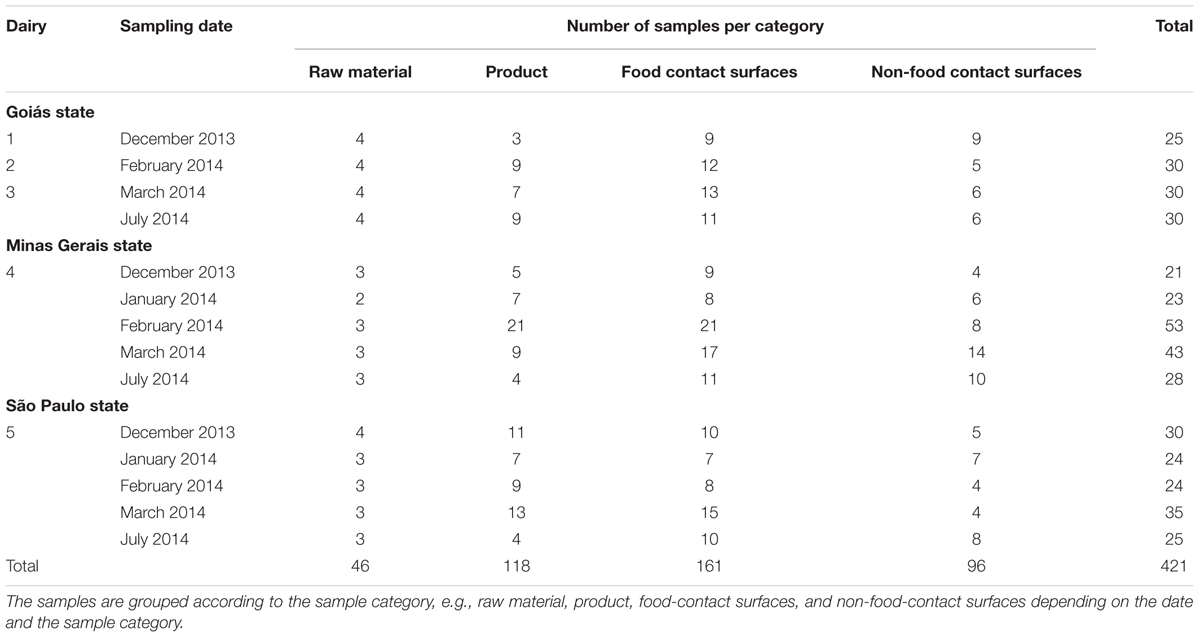
TABLE 1. Distribution of samples collected in this study in five dairies between December 2013 and July 2014 in the states of São Paulo, Minas Gerais and Goiás (Brazil).
A total of 421 samples were collected at different stages of processing following the method described by Oxaran et al. (2017). Briefly, raw material (n = 46), food-contact surfaces (n = 161), non-food-contact surfaces (n = 96), and products (n = 118) (Table 1 and Supplementary Table 1) were sampled using swab or sponge or by taking 25 g of cheese. Samples were transferred to the laboratory in cooling boxes containing ice packs. Homogenized samples in PSW were diluted 10-fold and 0.1 mL plated on BP to isolate putative S. aureus. After 24–48 h incubation at 35°C, up to three characteristic colonies of S. aureus (typical black, convex colonies, and with halo) were purified on TSAYE and incubated at 35°C for 24–48 h. Putative S. aureus isolates were stored for further analysis.
Shape and motility was determined by microscopy using an Olympus microscope (BX51). Gram-reaction was assessed by the 3% KOH method (Halebian et al., 1981). Catalase was tested using 3% hydrogen peroxide (Merck, Germany). Mannose fermentation and tolerance to high salt concentration was assessed on MSA incubated at 35°C for 48 h (Chapman, 1945). DNase activity was tested on DNase Test Agar with Methyl Green incubated at 35°C for 48 h. The production of free coagulase (Essers and Radebold, 1980) was tested using rabbit plasma (BioRad, France) and the presence of clumping factor as well as Protein A were tested using the Pastorex Staph-Plus Kit (BioRad, France) according to the manufacturer’s recommendations.
Genomic DNA was extracted using the Dynabeads® DNA DIRECT Universal kit (Invitrogen) on a 24 h culture. For each PCR reaction, 1 μL of extracted genomic DNA was used with primers listed in Table 2. PCR for the species identification and MLST were performed using the TEMPase Hot Start 2X Master Mix Blue II (Ampliqon, Denmark) and the Maxima Hot Start Taq Polymerase (Fermentas, Waltham, MA, United States) was used for toxin-encoding gene amplification, both were performed in a 25 μL reaction following the manufacturer’s instruction. Reactions were done in a Veriti Thermal Cycler (Applied Biosystems, 96 Well Model 9902). PCR products were purified using 10 U Exonuclease I (Thermo Fisher Scientific, Waltham, MA, United States) and 0.5 U Fast Alkaline Phosphatase (Thermo Fisher Scientific, Waltham, MA, United States) and incubated at 37°C for 15 min followed by an inactivation step at 85°C for 15 min (Werle et al., 1994). Macrogen Europe (The Netherlands) sequenced the gene fragments using the same primer used for the PCR amplification.

TABLE 2. List of primers used for the identification of Staphylococcus aureus based on 16 rRNA gene locus sequencing and for detection of four toxin encoding genes involved in food-poisoning, e.g., sea, sed, see, and seg.
The S. aureus identification was confirmed by 16S rRNA gene sequencing using primers 27F (Lane, 1991) and 1492R (Turner et al., 1999) (Table 2). The consensus sequence for each 16S rRNA locus was analyzed using the BLASTn algorithm on National Center for Biotechnology1.
The sequence type (ST) of each S. aureus isolate was determined by the method described by Enright et al. (2000). The internal fragment of seven housekeeping genes arcC (Carbamate kinase), aroE (shikimate dehydrogenase), glpF (glycerol kinase), gmk (guanylate kinase), pta (phosphate acetyltransferase), tpi (triosephosphate isomerase), yqiL (acetyle coenzyme A acetyltransferase), was amplified by PCR following the guideline on http://pubmlst.org/saureus/. The sequences were processed using CLC Main Workbench 7 (CLC Bio-Qiagen, Denmark) and analyzed using the S. aureus MLST database2 to assign the allele type and thus the ST. The profiles obtained including the new alleles and new profiles were submitted to the MLST database to contribute to the resource for S. aureus global epidemiology.
Hemolytic activity was tested on blood agar incubated at 35°C for 48 h and classified as α-hemolytic (partial hemolysis), β-hemolytic (complete hemolysis), α′-hemolytic (weak partial hemolysis) or γ-hemolytic (non-hemolytic bacteria).
Overnight cultures were adjusted to 0.5 McFarland (Biomerieux, France) suspensions (corresponding to 1–3 × 108 CFU/mL) and were inoculated by swabbing onto MHA in 120 mm × 120 mm Petri dishes (Almeco, Denmark). Eleven antibiotic disks (Oxoid, United Kingdom), Penicillin G (PEN, 10 U), Erythromycin (ERM, 15 μg), Tetracycline (TET, 30 μg), Clindamycin (CLIN, 2 μg), Gentamicin (GEN, 10 μg), Ciprofloxacin (CIP, 5 μg), Vancomycin (VAN, 5 μg), Trimethoprim-Sulfamethoxazole (SXT, 25 μg), Chloramphenicol (CAM, 30 μg), Cefoxitin (CEF, 30 μg), Bacitracin (BAC, 10 U) were tested. The diameter of inhibition zones was determined after 16–20 h and after 24 h of incubation at 35°C. The interpretation of the inhibition zone diameters was based on the EUCAST values (EUCAST, 2015) except for bacitracin and vancomycin inhibition zones which were compared to the Food and Drug Administration guidelines (FDA, 2006, 2009). S. aureus ATCC 25923 was used as control strain.
The presence of the genes encoding four toxins found in SFP was determined by PCR using the primers listed in Table 2. The most two common toxin encoding genes involved in SFP, sea and sed, and also the not so common SFP toxin encoding genes, see and seg, were targeted. The S. aureus strains ATCC 29213 (sea, seg), DSM 18588 (sed), DSM 18589 (see) were used as positive controls.
Thirty-one of 421 samples were positive for S. aureus (Supplementary Table 1). Strains were isolated from between 3 and 5 colonies from each sample showing typical S. aureus colony macroscopy on Baird Parker plates. A total of 66 isolates were identified as S. aureus being Gram positive, cocci shaped cells, catalase positive, DNase positive, growing in presence of high salt concentration, fermenting mannitol, as well as producing free and bound coagulase. The 16S rRNA gene sequencing confirmed the S. aureus identification for all 66 isolates. Four out of five dairies were positive for S. aureus, leading to an overall contamination rate of 7.4% (n = 31). The average contamination rate varied among dairies from 0% (n = 0) in Dairy 1 to 63.3% (n = 19) in Dairy 2 (Table 3). The dairies 3, 4 and 5, had contamination rates of 8.3% (n = 5), 1.2% (n = 2), and 3.6% (n = 5), respectively. Based on the overall contamination rate of S. aureus positive samples (n = 31), S. aureus was isolated in all four sampling categories (Table 3): raw material (10.9%), product (7.6%), food-contact surfaces (8.1%), and non-food-contact surfaces (4.2%).
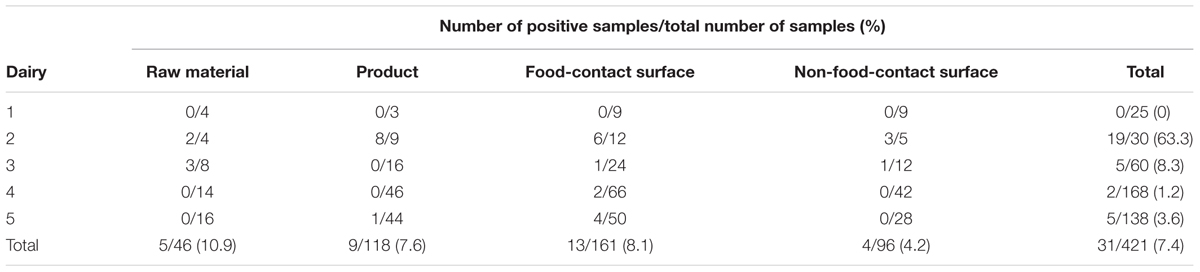
TABLE 3. Overall occurrence of S. aureus per dairy in the five dairies investigated grouped per sampling point category, e.g., raw material, product, food-contact surfaces, and non-food-contact surfaces.
Using MLST, the 66 S. aureus isolates were separated into 11 ST which were grouped into six clonal complexes (CCs) (Table 4). Of the 66 isolates, only 19 could be assigned to a known ST: ST1 (n = 7), ST188 (n = 3), ST97 (n = 3), ST398 (n = 2), ST5 (n = 1), ST30 (n = 2), and ST126 (n = 1). Besides the seven known ST, four new ST were found according to the MLST database2. The four new STs were assigned to 47 of the isolates. Due to a single point mutation, the new ST3531 (n = 41), ST3534 (n = 1), and ST3540 (n = 2) harbored a new allele type of the yqiL locus, 508, gmk locus, 291, or glpF locus, 515, respectively. All new allele types and thus ST were recorded on the MLST database. All of the seven allele types of the new ST, ST3562 (n = 3), were already assigned but the concatenation was novel and the isolate profile was submitted to the MLST database.
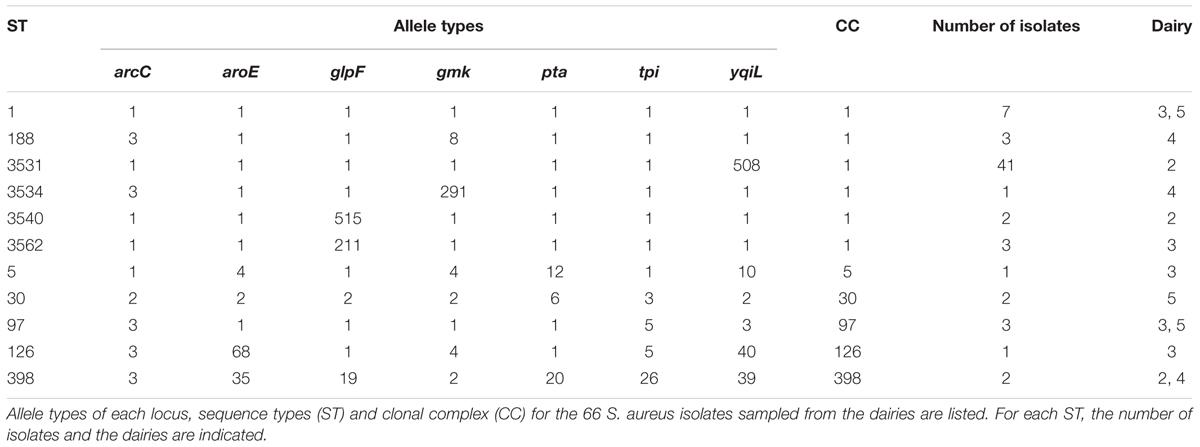
TABLE 4. MLST scheme of the S. aureus isolates from dairies in the Southeast and Midwest regions of Brazil.
ST3531 (n = 41) and ST3540 (n = 2) were only found in Dairy 2 with ST3531 being the predominant type (n = 41 out of 44 positive isolates). In this dairy, ST398 (n = 1) was also found. The ST3562 (n = 3) as well as ST5 (n = 1) and ST126 (n = 1) were specific to Dairy 3 and ST1 (n = 4) and ST97 (n = 2) were also identified. ST188 (n = 3), ST3534 (n = 1), and ST398 (n = 1) were isolated in Dairy 4. Dairy 5 was contaminated with ST1 (n = 3), ST30 (n = 2), and ST97 (n = 1).
The assigned ST were compared to the sampling points in each dairy and there was no correlation between particular ST at one specific sampling point. However, ST3531 was found at many different sampling points (wall, curd, cloth mold, cheese surface, sink, handler’s hand, floor, brine, processing tank, cheese, and paddle) in Dairy 2 (Supplementary Table 2). ST1 was found at two different sampling points both in Dairy 3 (curd and unpasteurized milk) and in Dairy 5 (bucket and cold chamber shelf). The results also showed that one sampling point, the unpasteurized milk (Dairy 3) was contaminated with S. aureus of three different ST (ST1, ST126, ST97). The ST were compared to the sampling time points in each dairy. Only ST1 and ST30 in Dairy 3 and 5, respectively, were occurring at two different time points indicating a re-introduction of these ST.
Out of the 66 isolates, 29 lysed the red blood cells completely and were β-hemolytic. Thirteen strains were α-hemolytic. Twenty-three isolates produced a weak α′-hemolytic zone and one isolate (Sa8) did not display hemolytic activity, being γ-hemolytic (Supplementary Table 2).
The 66 isolates were all susceptible to four antibiotics: trimethoprim-sulfamethoxazole, chloramphenicol, cefoxitin, and bacitracin (Table 5). Resistance toward one or more antibiotics was observed for 24.2% of the isolates (n = 16). Seven isolates were only resistant to one antibiotic being either ciprofloxacin (1.5%, n = 1) or penicillin G (9.1%, n = 6). Combination of resistance toward two to four antibiotics was observed for nine isolates (13.6%) (Table 5). None of the 66 isolates were methicillin resistant. No isolates were resistant to vancomycin but 21 isolates showed an intermediate tolerance to this antibiotic. One of the vancomycin intermediate tolerant isolates (Sa44) was also intermediate tolerant to tetracycline (Supplementary Table 2).
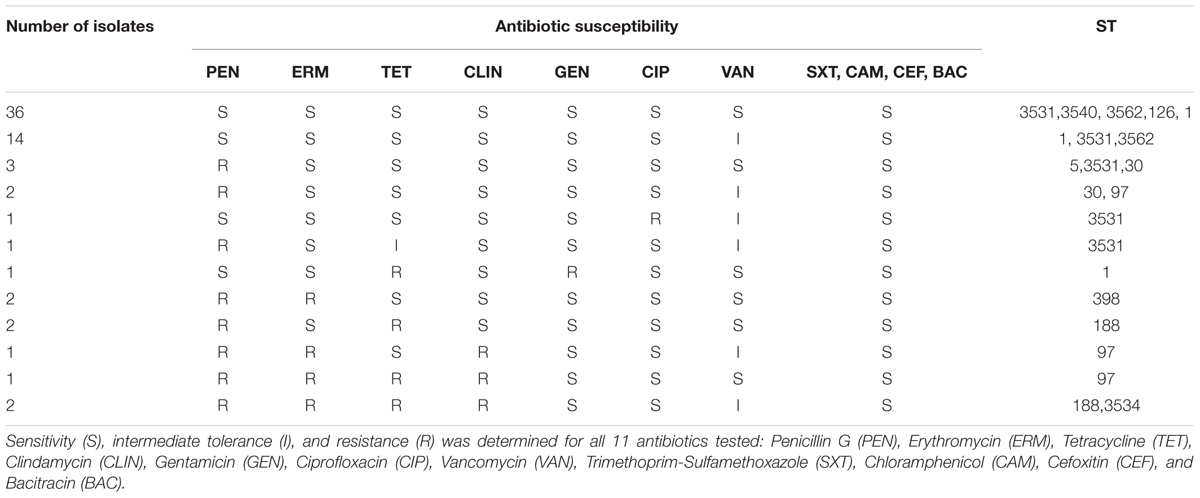
TABLE 5. Number of isolates per antibiotic susceptibility profile related to the sequence type (ST).
Using PCR, none of the isolates was positive for detection of gene encoding toxins D and E, e.g., sed and see respectively. Three isolates out of 66 were positive for seg, including two isolates harboring both genes seg and sea (Supplementary Table 2).
Minas frescal cheese is a widely consumed soft cheese in Brazil and it has been associated with S. aureus food-poisoning (de Almeida Filho and Nader Filho, 2000; Araújo et al., 2002). Most of the dairies investigated in the present study were positive for S. aureus. This is in concordance with the general trend that this pathogen is a common contaminant of dairy industries worldwide, including Brazil (de Almeida Filho and Nader Filho, 2000; Araújo et al., 2002; Carvalho et al., 2007; André et al., 2008; Fagundes et al., 2010) where contamination rates can reach up to 77.0% (Araújo et al., 2002; André et al., 2008; Moraes et al., 2009). Here, we identified one dairy with a high level of contamination (63.3%, Dairy 2). The four other dairies had a lower contamination rate (from 0 to 8.3%) which is similar to the rate described by Lee et al. (2012) but lower than generally reported in the literature (Araújo et al., 2002). S. aureus was identified in all four sampling categories (raw material, product, food-contact surfaces, and non-food-contact surfaces) but the contamination rates varied. The highest overall contamination rate was observed for the raw material category (10.9%), however, only two out of the five dairies were positive for S. aureus presence in this category. This included S. aureus isolation from the milk in the only dairy producing cheese from unpasteurized milk (Dairy 3), however this type of dairy did not show the highest contamination rate of any of the other sampling categories. Contamination of raw milk by S. aureus has previously been reported and linked to milk from animals with mastitis (Jørgensen et al., 2005; da Costa Krewer et al., 2015). The frequency of S. aureus contamination of dairy cows is quite high in Brazil (da Costa Krewer et al., 2015) and this could also be one of the contamination sources for Dairy 3. However, this is likely not the only pathogen introduction route for all of the investigated dairies. The overall contamination rates of the food-contact surfaces and products were slightly lower than that of the raw material. Nevertheless, at least one food-contact surface sample was positive for S. aureus in all of the dairies where the pathogen was detected. S. aureus was also isolated from the handler’s hands, gloves, and utensils in three of the dairies (Dairy 2, 4, and 5). Several studies have reported that contamination by S. aureus is mainly due to transfer between handlers and the equipment thus incriminating the lack of good manufacturing practices (GMPs) (Lee et al., 2012; Kümmel et al., 2016). Altogether, it hints towards a human spread of the pathogen in the investigated dairies and a need for improvement in the hygienic practices. However, further monitoring is needed to confirm this speculation.
We typed the S. aureus isolates choosing MLST among the different epidemiological methods that have previously been applied to study the relatedness of S. aureus isolates in food production plants (Tondo et al., 2000; Lee, 2003; Kérouanton et al., 2007; Rabello et al., 2007; Agersø et al., 2012; Lee et al., 2012; Yan et al., 2012; Shepheard et al., 2013; Rešková et al., 2014; McMillan et al., 2016). Indeed, MLST provide data that can be compared on a global scale and allow typing of important S. aureus clones such as ST398 involved in human and animal infections that are non-typable by the standard PFGE method (using smaI) due to DNA methylation (Bens et al., 2006). The 66 isolates identified in this study belonged to six different CC and to seven known ST (ST1, ST5, ST30, ST97, ST126, ST188, and ST398) and four new ST (ST3531, ST3534, ST3540, ST3562). According to the MLST database2 and previous studies on milk and dairy products, all of the known ST have previously been isolated in Brazil (Vivoni et al., 2006; Rabello et al., 2007; Silva et al., 2013, 2014). In three instances, more than one ST was identified from one single sample, e.g., ST3531 and ST3540 being identified from the same cheese surface sample (Supplementary Table 2), which shows that more than one S. aureus clone were present and thereby supports the requirement of analyzing more than one colony per sample (Mongkolrattanothai et al., 2011; Votintseva et al., 2014). We did not find any indications of persistence as it has been seen in other studies (Koreňová et al., 2015) but our results indicate that S. aureus was continuously reintroduced into the dairies.
The majority of the isolates (44 out of 66) including the four novel ST belonged to CC1. Although several other studies have found large diversity regarding S. aureus contamination (Oliveira et al., 2016), Silva et al. (2013) also found that CC1 (ST1) was the main contaminant of milk from Brazilian cows infected with mastitis due to MSSA and also identified a new ST belonging to CC1. So far, the clinically related data represents the largest amount of available data on S. aureus epidemiology in Brazil and using other molecular typing methods, it has been shown that the genetic diversity of this pathogen in Brazilian hospitals is limited (Teixeira et al., 1995) and thus is in concordance with our results observing a low genetic diversity as well as the predominance of the CC1. All together it reinforces that clinical clones are now spreading into other niches and potentially into the food industry (Pesavento et al., 2007; Choudhury et al., 2012). In addition, isolates from different sampling points in one dairy (Dairy 2) belonged to the same ST3531, being per se epidemiologically related and thus suggesting a potential dissemination of one clone in the dairy. Investigating S. aureus from milk and the milking environment in São Paulo by PFGE (Lee et al., 2012), reached the same conclusion. To a lesser extent, the other ST belonging to different CC observed in this study have been sporadically reported in bovine dairy or products. Of concern, ST398 has previously been described as a human public health threat in Europe due to its ability to disseminate in humans and pigs (Armand-Lefevre et al., 2005; Voss et al., 2005; Price et al., 2012) as well as its potential antibiotic resistance pattern. In Brazil, MRSA ST398 has been reported in bovine mastitis (Silva et al., 2014). The two ST398 isolates found in this study were MSSA, and in China, MSSA ST398 has also been found in food products (Yan et al., 2014; Li et al., 2015b). Although the main pathogenicity determinants of MSSA ST398 are still unknown, the presence of these isolates along the food processing chain is of concern for human health. Indeed, MSSA ST398 has recently been involved in human infections (Valentin-Domelier et al., 2011; Mediavilla et al., 2012; David et al., 2013). In previous reports from Brazilian dairies, ST1, ST97, and ST126 were commonly identified (Rabello et al., 2007; Silva et al., 2013; Oliveira et al., 2016) and ST97 has also been found in dairies worldwide (Smith et al., 2005). ST126 that belongs to CC126 is usually associated with ruminants but not humans (Smith et al., 2005; Aires-de-Sousa et al., 2007; Rabello et al., 2007), hence, suggesting a bovine origin for those isolates. The ST5/CC5 was originally described in poultry infections but it can also be found among human isolates (Lowder et al., 2009). Moreover, ST5 was also present in dairy environments in the US (Matyi et al., 2013) and in the Brazilian dairy industry (Silva et al., 2013). Highlighted by those results, ST found in the dairy plants investigated here belonged to ST or CC previously involved into food-poisoning (ST1, ST5, ST30, and ST188) worldwide (Cha et al., 2006; Yan et al., 2012; Li et al., 2015a).
Le Loir et al. (2003) have reported that 25% of S. aureus strains isolated from food are enterotoxigenic and staphylococcal enterotoxins are responsible for food-poisoning outbreaks worldwide including from dairy products (Do Carmo et al., 2004; Loncarevic et al., 2005). Here, a small number of strains (4.5%) harbored SE-encoding genes involved in SFP. The number of strains harboring SE-encoding genes varies, especially depending on the geographical area (Jørgensen et al., 2005; Neder et al., 2011; Riva et al., 2015) and has ranged from 0 to 93.6% (Do Carmo et al., 2004; Fagundes et al., 2010; Rall et al., 2014; Silveira-Filho et al., 2014). SEA is the most prevalent toxin involved in SFP worldwide and sea as well as seb were the predominant SE-encoding genes found to cause SFP in Brazil (Veras et al., 2008). Here, only two isolates harbored sea, which is less frequent than previously reported. The second SE-encoding gene found in three isolates was seg. The seg/sei genes were recently reported to be the most prevalent among mastitis milking cows and cheese manufacturing in Brazil (Arcuri et al., 2010; Silveira-Filho et al., 2014). Linking ST to toxin gene presence, only ST5 and ST30 strains harbored SE-encoding genes in this study. In fact, assessing the MLST profile to the toxigenic potential of S. aureus food sources in China (Chang et al., 2016) has illustrated the specific ability of ST5 strains to harbor and to produce SE while the other ST, including ST1, ST188, ST97 and ST398, did not, as we also noticed.
Staphylococcus aureus is known for its ability to gain antimicrobial resistance and attention has been given to multi-drug as well as methicillin resistant strains and in particular to highly virulent MRSA strains. Several studies have shown that about 20% of the isolates were MRSA (Türkyilmaz et al., 2010; Riva et al., 2015), with in particular in Brazil methicillin resistance occurrence ranges from 5.5 to 37.2% in milk, dairy processing environment and dairy products (André et al., 2008; Silveira-Filho et al., 2014; Oliveira et al., 2016). However in this study, none of the isolates were classified as MRSA which corroborates with one previous study (Silveira-Filho et al., 2014). Additionally, no isolates were resistant toward vancomycin concurring with previous reports in the Brazilian dairy industry (André et al., 2008; Silveira-Filho et al., 2014; Oliveira et al., 2016). Hence, there are yet no indications of any multi-drug resistant MRSA emerging in the Brazilian dairy industry. The highest resistance rates were observed for penicillin G (20.9%), erythromycin (9.0%), and tetracycline (9.0%) similar to findings by Oliveira et al. (2016). Resistance to penicillin is frequently found in dairy herds worldwide, the occurrence rates ranging from 10 to 70% of the isolates being resistant with in particular 47.6% in Uruguay and Argentina (Calvinho et al., 2002; Gianneechini et al., 2002) as well as 69.9% in Brazil (André et al., 2008). However, resistance to the widespread administration of these specific antibiotics to control and treat infections in dairy farms including mammary infections reached lower value here. Thus, in this specific region the use of β-lactams as antibiotic treatment may have only triggered the increase of resistance occurrence to β-lactams such as penicillin. Moreover, the co-resistance pattern of tetracycline and macrolide (erythromycin) has frequently been detected in ST1 (MRSA) in Italy (Battisti et al., 2010; Franco et al., 2011; Carfora et al., 2015) which corroborates with our data. Clindamycin, gentamicin, and ciprofloxacin are commonly used in Brazil to treat bovine mastitis and it has been suggested that these antibiotics trigger a selective pressure in dairy farms. Indeed, all of the isolates from dairy farms in Northeastern Brazil collected by Silveira-Filho et al. (2014) were resistant toward gentamicin and bacitracin. However, the resistance observed toward these antibiotics was very limited here as also observed by others (André et al., 2008). Nevertheless, the resistance to more than one antibiotic of some isolates, observed here, could pave the way of the resistant determinant dissemination.
Some Brazilian dairy processing plants and cheese products have frequent occurrence of S. aureus. The genetic diversity is low and could indicate a few sources of contamination and, if identified could form the basis for an intervention strategy.
Study concept and design: VO and LG. Planning and sampling at dairies: LC, SL, CC, EPDM, VA, and CFdO. Analysis and interpretation of data: KD, SL, LC, and VO. Drafting of the manuscript: VO. Critical revision of the manuscript: LG, KD, EPDM, VA, CC, VO, and CFdO.
The study was funded by the Danish Council for Strategic Research (project number 0603-00552B), the State of São Paulo Research Foundation/Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP #2012/50507-1), The State of Goiás Research Foundation (FAPEG #2012/10267001047) and CAPES for the scholarship.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This publication made use of the S. aureus MLST website (http://pubmlst.org/saureus/) sited at the University of Oxford (Jolley and Maiden, 2010). The development of this site has been funded by the Wellcome Trust. Jette Melchiorsen and Vanessa Maciel de Souza are warmly acknowledged for their excellent technical help.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02049/full#supplementary-material
Agersø, Y., Hasman, H., Cavaco, L. M., Pedersen, K., and Aarestrup, F. M. (2012). Study of methicillin resistant Staphylococcus aureus (MRSA) in Danish pigs at slaughter and in imported retail meat reveals a novel MRSA type in slaughter pigs. Vet. Microbiol. 157, 246–250. doi: 10.1016/j.vetmic.2011.12.023
Aires-de-Sousa, M., Parente, C. E. S. R., Vieira-da-Motta, O., Bonna, I. C. F., Silva, D. A., and de Lencastre, H. (2007). Characterization of Staphylococcus aureus isolates from buffalo, bovine, ovine, and caprine milk samples collected in Rio de Janeiro State, Brazil. Appl. Environ. Microbiol. 73, 3845–3849. doi: 10.1128/AEM.00019-07
André, M. C. D. P. B., Campos, M. R. H., Borges, L. J., Kipnis, A., Pimenta, F. C., and Serafini, Á. B. (2008). Comparison of Staphylococcus aureus isolates from food handlers, raw bovine milk and Minas Frescal cheese by antibiogram and pulsed-field gel electrophoresis following SmaI digestion. Food Control 19, 200–207. doi: 10.1016/j.foodcont.2007.03.010
Araújo, V. S., Pagliares, V. A., Queiroz, M. L., and Freitas-Almeida, A. C. (2002). Occurrence of Staphylococcus and enteropathogens in soft cheese commercialized in the city of Rio de Janeiro, Brazil. J. Appl. Microbiol. 92, 1172–1177. doi: 10.1046/j.1365-2672.2002.01656.x
Arcuri, E. F., Angelo, F. F., Guimarães, M. F. M., Talon, R., Borges, M., de, F., et al. (2010). Toxigenic status of Staphylococcus aureus isolated from bovine raw milk and Minas frescal cheese in Brazil. J. Food Prot. 73, 2225–2231. doi: 10.4315/0362-028X-73.12.2225
Arfatahery, N., Davoodabadi, A., Abedimohtasab, T., LeLoir, Y., Baron, F., Gautier, M., et al. (2016). Characterization of toxin genes and antimicrobial susceptibility of Staphylococcus aureus isolates in fishery products in Iran. Sci. Rep. 6:34216. doi: 10.1038/srep34216
Argudín, M. Á., Mendoza, M. C., and Rodicio, M. R. (2010). Food poisoning and Staphylococcus aureus enterotoxins. Toxins 2, 1751–1773. doi: 10.3390/toxins2071751
Armand-Lefevre, L., Ruimy, R., and Andremont, A. (2005). Clonal comparison of Staphylococcus aureus isolates from healthy pig farmers, human controls, and pigs. Emerg. Infect. Dis. 11, 711–714. doi: 10.3201/eid1105.040866
Asao, T., Kumeda, Y., Kawai, T., Shibata, T., Oda, H., Haruki, K., et al. (2003). An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiol. Infect. 130, 33–40. doi: 10.1017/S0950268802007951
Balaban, N., and Rasooly, A. (2000). Staphylococcal enterotoxins. Int. J. Food Microbiol. 61, 1–10. doi: 10.1016/S0168-1605(00)00377-9
Battisti, A., Franco, A., Merialdi, G., Hasman, H., Iurescia, M., Lorenzetti, R., et al. (2010). Heterogeneity among methicillin-resistant Staphylococcus aureus from Italian pig finishing holdings. Vet. Microbiol. 142, 361–366. doi: 10.1016/j.vetmic.2009.10.008
Baumgartner, A., Niederhauser, I., and Johler, S. (2014). Virulence and resistance gene profiles of Staphylococcus aureus strains isolated from ready-to-eat foods. J. Food Prot. 77, 1232–1236. doi: 10.4315/0362-028X.JFP-14-027
Bens, C. C. P. M., Voss, A., and Klaassen, C. H. W. (2006). Presence of a novel DNA methylation enzyme in methicillin-resistant Staphylococcus aureus isolates associated with Pig farming leads to uninterpretable results in standard pulsed-field gel electrophoresis analysis. J. Clin. Microbiol. 44, 1875–1876. doi: 10.1128/JCM.44.5.1875-1876.2006
Calvinho, L. F., Toselli, F. G., Weimann, W. R., Canavesio, V. R., Neder, V. E., and Iguzquiza, I. A. (2002). Antimicrobial sensitivity of coagulase-positive staphylococcal strains isolated from bovine mastitis in the central dairy catchment area of Argentina. Rev. Argent. Microbiol. 34, 171–175.
Carfora, V., Caprioli, A., Marri, N., Sagrafoli, D., Boselli, C., Giacinti, G., et al. (2015). Enterotoxin genes, enterotoxin production, and methicillin resistance in Staphylococcus aureus isolated from milk and dairy products in Central Italy. Int. Dairy J. 42, 12–15. doi: 10.1016/j.idairyj.2014.10.009
Carvalho, J. D. G., Viotto, W. H., Kuaye, A. Y., Walkíria, V. H., and Kuaye, A. Y. (2007). The quality of Minas Frescal cheese produced by different technological processes. Food Control 18, 262–267. doi: 10.1016/j.foodcont.2005.10.005
Cha, J. O., Lee, J. K., Jung, Y. H., Yoo, J. I., Park, Y. K., Kim, B. S., et al. (2006). Molecular analysis of Staphylococcus aureus isolates associated with staphylococcal food poisoning in South Korea. J. Appl. Microbiol. 101, 864–871. doi: 10.1111/j.1365-2672.2006.02957.x
Chambers, H. F., and Deleo, F. R. (2009). Waves of resistance: Staphylococcus aureus in the antibiotic Era. Nat. Rev. Microbiol. 7, 629–641. doi: 10.1038/nrmicro2200
Chang, Y., Gao, H., Zhu, Z., Ye, S., Yang, Y., Shen, X., et al. (2016). High prevalence and properties of enterotoxin-producing Staphylococcus aureus ST5 strains of food sources in China. Foodborne Pathog. Dis. 13, 386–390. doi: 10.1089/fpd.2015.2085
Chapman, G. H. (1945). The significance of sodium chloride in studies of staphylococci. J. Bacteriol. 50, 201–203.
Chen, T.-R., Chiou, C.-S., and Tsen, H.-Y. (2004). Use of novel PCR primers specific to the genes of staphylococcal enterotoxin G, H, I for the survey of Staphylococcus aureus strains isolated from food-poisoning cases and food samples in Taiwan. Int. J. Food Microbiol. 92, 189–197. doi: 10.1016/j.ijfoodmicro.2003.10.002
Choudhury, R., Panda, S., and Singh, D. V. (2012). Emergence and dissemination of antibiotic resistance: a global problem. Indian J. Med. Microbiol. 30, 384–390. doi: 10.4103/0255-0857.103756
da Costa Krewer, C., Santos Amanso, E., Veneroni Gouveia, G., de Lima Souza, R., da Costa, M. M., and Aparecido Mota, R. (2015). Resistance to antimicrobials and biofilm formation in Staphylococcus spp. isolated from bovine mastitis in the Northeast of Brazil. Trop. Anim. Health Prod. 47, 511–518. doi: 10.1007/s11250-014-0752-9
David, M. Z., Siegel, J., Lowy, F. D., Zychowski, D., Taylor, A., Lee, C. J., et al. (2013). Asymptomatic carriage of sequence type 398, spa type t571 methicillin-susceptible Staphylococcus aureus in an urban jail: a newly emerging, transmissible pathogenic strain. J. Clin. Microbiol. 51, 2443–2447. doi: 10.1128/JCM.01057-13
de Almeida Filho, E. S., and Nader Filho, A. (2000). Occurrence of Staphylococcus aureus in cheese made in Brazil. Rev. Saude Publica 34, 578–580. doi: 10.1590/S0034-89102000000600003
de Lencastre, H., Oliveira, D., and Tomasz, A. (2007). Antibiotic resistant Staphylococcus aureus: a paradigm of adaptive power. Curr. Opin. Microbiol. 10, 428–435. doi: 10.1016/j.mib.2007.08.003
Dinges, M. M., Orwin, P. M., and Schlievert, P. M. (2000). Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 13, 16–34. doi: 10.1128/CMR.13.1.16-34.2000
Do Carmo, L. S., Cummings, C., Linardi, V. R., Dias, R. S., De Souza, J. M., De Sena, M. J., et al. (2004). A case study of a massive staphylococcal food poisoning incident. Foodborne Pathog. Dis. 1, 241–246. doi: 10.1089/fpd.2004.1.241
ECDC (2014). Antimicrobial Resistance Surveillance in Europe 2013. Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net). Stockholm: European Centre for Disease Prevention and Control. doi: 10.2900/39777
EFSA and ECDC (2014). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2012. Parma: EFSA. doi: 10.2903/j.efsa.2014.3547
Enright, M. C., Day, N. P. J., Davies, C. E., Peacock, S. J., and Spratt, B. G. (2000). Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 38, 1008–1015.
Essers, L., and Radebold, K. (1980). Rapid and reliable identification of Staphylococcus aureus by a latex agglutination test. J. Clin. Microbiol. 12, 641–643.
EUCAST (2015). The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 5.0, 2015. Available at: http://www.eucast.org
Fagundes, H., Barchesi, L., Nader Filho, A., Ferreira, L. M., Oliveira, C. A. F., Filho, A. N., et al. (2010). Occurrence of Staphylococcus aureus in raw milk produced in dairy farms in Sao Paulo State, Brazil. Braz. J. Microbiol. 41, 376–380. doi: 10.1590/S1517-83822010000200018
FDA (2006). U.S. Food & Drug Administration. Susceptibility Test Interpretive Criteria for Bacitracin. Silver Spring, MD: FDA.
FDA (2009). U.S. Food & Drug Administration. Susceptibility Test Interpretive Criteria for Vancomycin. Silver Spring, MD: FDA.
Franco, A., Hasman, H., Iurescia, M., Lorenzetti, R., Stegger, M., Pantosti, A., et al. (2011). Molecular characterization of spa type t127, sequence type 1 methicillin-resistant Staphylococcus aureus from pigs. J. Antimicrob. Chemother. 66, 1231–1235. doi: 10.1093/jac/dkr115
Gianneechini, R. E., Concha, C., and Franklin, A. (2002). Antimicrobial susceptibility of udder pathogens isolated from dairy herds in the west littoral region of Uruguay. Acta Vet. Scand. 43, 31–41. doi: 10.1186/1751-0147-43-31
Gomes, B. C., Franco, B. D. G., de, M., and De Martinis, E. C. P. (2013). Microbiological food safety issues in Brazil: bacterial pathogens. Foodborne Pathog. Dis. 10, 197–205. doi: 10.1089/fpd.2012.1222
Halebian, S., Harris, B., Finegold, S. M., and Rolfei, R. D. (1981). Rapid method that aids in distinguishing Gram-positive from gram-negative anaerobic rapid method that aids in distinguishing gram-positive from gram-negative anaerobic bacteria. J. Clin. Microbiol. 13, 444–448.
Hao, D., Xing, X., Li, G., Wang, X., Zhang, M., Zhang, W., et al. (2015). Prevalence, toxin gene profiles, and antimicrobial resistance of Staphylococcus aureus isolated from quick-frozen dumplings. J. Food Prot. 78, 218–223. doi: 10.4315/0362-028X.JFP-14-100
Hatakka, M., Björkroth, K. J., Asplund, K., Mäki-Petäys, N., and Korkeala, H. J. (2000). Genotypes and enterotoxicity of Staphylococcus aureus isolated from the hands and nasal cavities of flight-catering employees. J. Food Prot. 63, 1487–1491. doi: 10.4315/0362-028X-63.11.1487
Hennekinne, J.-A., De Buyser, M.-L., and Dragacci, S. (2012). Staphylococcus aureus and its food poisoning toxins: characterization and outbreak investigation. FEMS Microbiol. Rev. 36, 815–836. doi: 10.1111/j.1574-6976.2011.00311.x
Holden, M. T. G., Feil, E. J., Lindsay, J. A., Peacock, S. J., Day, N. P. J., Enright, M. C., et al. (2004). Complete genomes of two clinical Staphylococcus aureus strains: evidence for the rapid evolution of virulence and drug resistance. Proc. Natl. Acad. Sci. U.S.A. 101, 9786–9791. doi: 10.1073/pnas.0402521101
Ikeda, T., Tamate, N., Yamaguchi, K., and Makino, S. (2005). Quantitative analysis of Staphylococcus aureus in skimmed milk powder by real-time PCR. J. Vet. Med. Sci. 67, 1037–1041. doi: 10.1292/jvms.67.1037
Jarraud, S., Mougel, C., Thioulouse, J., Lina, G., Meugnier, H., Forey, F., et al. (2002). Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect. Immun. 70, 631–641. doi: 10.1128/IAI.70.2.631-641.2002
Jolley, K. A., and Maiden, M. C. (2010). BIGSdb: scalable analysis of bacterial genome variation at the population level. BMC Bioinformatics 11:595. doi: 10.1186/1471-2105-11-595
Jones, T. F., Kellum, M. E., Porter, S. S., Bell, M., and Schaffner, W. (2002). An outbreak of community-acquired foodborne illness caused by methicillin-resistant Staphylococcus aureus. Emerg. Infect. Dis. 8, 82–84. doi: 10.3201/eid0801.010174
Jørgensen, H. J., Mørk, T., Høgåsen, H. R., and Rørvik, L. M. (2005). Enterotoxigenic Staphylococcus aureus in bulk milk in Norway. J. Appl. Microbiol. 99, 158–166. doi: 10.1111/j.1365-2672.2005.02569.x
Kadariya, J., Smith, T. C., and Thapaliya, D. (2014). Staphylococcus aureus and staphylococcal food-borne disease: an ongoing challenge in public health. Biomed Res. Int. 2014:827965. doi: 10.1155/2014/827965
Kérouanton, A., Hennekinne, J. A., Letertre, C., Petit, L., Chesneau, O., Brisabois, A., et al. (2007). Characterization of Staphylococcus aureus strains associated with food poisoning outbreaks in France. Int. J. Food Microbiol. 115, 369–375. doi: 10.1016/j.ijfoodmicro.2006.10.050
Khan, S. A., Nawaz, M. S., Khan, A. A., and Cerniglia, C. E. (2000a). Transfer of erythromycin resistance from poultry to human clinical strains of Staphylococcus aureus. J. Clin. Microbiol. 38, 1832–1838.
Khan, S. A., Nawaz, M. S., Khan, A. A., Steele, R. S., and Cerniglia, C. E. (2000b). Characterization of erythromycin-resistant methylase genes from multiple antibiotic resistant Staphylococcus spp isolated from milk samples of lactating cows. Am. J. Vet. Res. 61, 1128–1132.
Koreňová, J., Rešková, Z., Véghová, A., and Kuchta, T. (2015). Tracing Staphylococcus aureus in small and medium-sized food-processing factories on the basis of molecular sub-species typing. Int. J. Environ. Health Res. 25, 384–392. doi: 10.1080/09603123.2014.958135
Kümmel, J., Stessl, B., Gonano, M., Walcher, G., Bereuter, O., Fricker, M., et al. (2016). Staphylococcus aureus entrance into the dairy chain: tracking S. aureus from dairy cow to cheese. Front. Microbiol. 7:1603. doi: 10.3389/fmicb.2016.01603
Kusumaningrum, H. D., Riboldi, G., Hazeleger, W. C., and Beumer, R. R. (2003). Survival of foodborne pathogens on stainless steel surfaces and cross-contamination to foods. Int. J. Food Microbiol. 85, 227–236. doi: 10.1016/S0168-1605(02)00540-8
Lane, D. J. J. (1991). “16S/23S rRNA sequencing,” in Nucleic Acid Techniques in Bacterial Systematics, eds E. Stackebrandt and M. Goodfellow (New York, NY: John Wiley & Sons), 115–175. doi: 10.1007/s00227-012-2133-0
Le Loir, Y., Baron, F., and Gautier, M. (2003). Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2, 63–76.
Lee, J. H. (2003). Methicillin (oxacillin)-resistant Staphylococcus aureus strains isolated from major food animals and their potential transmission to humans. Appl. Environ. Microbiol. 69, 6489–6494. doi: 10.1128/AEM.69.11.6489-6494.2003
Lee, S. H. I., Camargo, C. H., Gonçalves, J. L., Cruz, A. G., Sartori, B. T., Machado, M. B., et al. (2012). Characterization of Staphylococcus aureus isolates in milk and the milking environment from small-scale dairy farms of São Paulo, Brazil, using pulsed-field gel electrophoresis. J. Dairy Sci. 95, 7377–7383. doi: 10.3168/jds.2012-5733
Li, G., Wu, C., Wang, X., and Meng, J. (2015b). Prevalence and characterization of methicillin susceptible Staphylococcus aureus ST398 isolates from retail foods. Int. J. Food Microbiol. 196, 94–97. doi: 10.1016/j.ijfoodmicro.2014.12.002
Li, G., Wu, S., Luo, W., Su, Y., Luan, Y., and Wang, X. (2015a). Staphylococcus aureus ST6-t701 isolates from food-poisoning outbreaks (2006-2013) in Xi’an, China. Foodborne Pathog. Dis. 12, 203–206. doi: 10.1089/fpd.2014.1850
Lima, G. C., Loiko, M. R., Casarin, L. S., and Tondo, E. C. (2013). Assessing the epidemiological data of Staphylococcus aureus food poisoning occurred in the State of Rio Grande do Sul, Southern Brazil. Braz. J. Microbiol. 44, 759–763. doi: 10.1590/S1517-83822013005000063
Lindberg, E., Nowrouzian, F., Adlerberth, I., and Wold, A. E. (2000). Long-time persistence of superantigen-producing Staphylococcus aureus strains in the intestinal microflora of healthy infants. Pediatr. Res. 48, 741–747. doi: 10.1203/00006450-200012000-00007
Loncarevic, S., Jørgensen, H. J., Løvseth, A., Mathisen, T., and Rørvik, L. M. (2005). Diversity of Staphylococcus aureus enterotoxin types within single samples of raw milk and raw milk products. J. Appl. Microbiol. 98, 344–350. doi: 10.1111/j.1365-2672.2004.02467.x
Lowder, B. V., Guinane, C. M., Ben Zakour, N. L., Weinert, L. A., Conway-Morris, A., Cartwright, R. A., et al. (2009). Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc. Natl. Acad. Sci. U.S.A. 106, 19545–19550. doi: 10.1073/pnas.0909285106
Lues, J. F. R., and Van Tonder, I. (2007). The occurrence of indicator bacteria on hands and aprons of food handlers in the delicatessen sections of a retail group. Food Control 18, 326–332. doi: 10.1016/j.foodcont.2005.10.010
Matyi, S. A., Dupre, J. M., Johnson, W. L., Hoyt, P. R., White, D. G., Brody, T., et al. (2013). Isolation and characterization of Staphylococcus aureus strains from a Paso del Norte dairy. J. Dairy Sci. 96, 3535–3542. doi: 10.3168/jds.2013-6590
McMillan, K., Moore, S. C., McAuley, C. M., Fegan, N., and Fox, E. M. (2016). Characterization of Staphylococcus aureus isolates from raw milk sources in Victoria, Australia. BMC Microbiol. 16:169. doi: 10.1186/s12866-016-0789-1
Mediavilla, J. R., Chen, L., Uhlemann, A.-C., Hanson, B. M., Rosenthal, M., Stanak, K., et al. (2012). Methicillin-susceptible Staphylococcus aureus ST398, New York and New Jersey, USA. Emerg. Infect. Dis. 18, 700–702. doi: 10.3201/eid1804.111419
Ministério da Agricultura Brasil (1997). Pecuária e Abastecimento, Portaria No. 352, de 04/09/1997. Aprova o Regulamento Técnico para Fixacão de Identidade e Qualidade de Queijos Minas Frescal Diário Of. da União. Brasília: Ministério da Agricultura Brasil.
Mongkolrattanothai, K., Gray, B. M., Mankin, P., Stanfill, A. B., Pearl, R. H., Wallace, L. J., et al. (2011). Simultaneous carriage of multiple genotypes of Staphylococcus aureus in children. J. Med. Microbiol. 60, 317–322. doi: 10.1099/jmm.0.025841-0
Moraes, P. M., Viçosa, G. N., Yamazi, A. K., Ortolani, M. B. T., and Nero, L. A. (2009). Foodborne pathogens and microbiological characteristics of raw milk soft cheese produced and on retail sale in Brazil. Foodborne Pathog. Dis. 6, 245–249. doi: 10.1089/fpd.2008.0156
Murray, R. J. (2005). Recognition and management of Staphylococcus aureus toxin-mediated disease. Intern. Med. J. 35(Suppl. 2), S106–S119. doi: 10.1111/j.1444-0903.2005.00984.x
Neder, V. E., Canavesio, V. R., and Calvinho, L. F. (2011). Presence of enterotoxigenic Staphylococcus aureus in bulk tank milk from Argentine dairy farms. Rev. Argent. Microbiol. 43, 104–106. doi: 10.1590/S0325-75412011000200006
Normanno, G., La Salandra, G., Dambrosio, A., Quaglia, N. C., Corrente, M., Parisi, A., et al. (2007). Occurrence, characterization and antimicrobial resistance of enterotoxigenic Staphylococcus aureus isolated from meat and dairy products. Int. J. Food Microbiol. 115, 290–296. doi: 10.1016/j.ijfoodmicro.2006.10.049
Oliveira, C. J. B., Tiao, N., de Sousa, F. G. C., de Moura, J. F. P., Santos Filho, L., and Gebreyes, W. A. (2016). Methicillin-resistant Staphylococcus aureus from Brazilian dairy farms and identification of novel sequence types. Zoonoses Public Health 63, 97–105. doi: 10.1111/zph.12209
Oliveira, L. P., de, Soares e Barros, L. S., Silva, V. C., and Cirqueira, M. G. (2011). Study of Staphylococcus aureus in raw and pasteurized milk consumed in the Reconcavo area of the State of Bahia, Brazil. J. Food Process Technol. 2:128. doi: 10.4172/2157-7110.1000128
Ostyn, A., De Buyser, M. L., Guillier, F., Groult, J., Felix, B., Salah, S., et al. (2010). First evidence of a food poisoning outbreak due to staphylococcal enterotoxin type E, France, 2009. Euro Surveill. 15:19528.
Oxaran, V., Lee, S. H. I., Chaul, L. T., Corassin, C. H., Barancelli, G. V., Alves, V. F., et al. (2017). Listeria monocytogenes incidence changes and diversity in some Brazilian dairy industries and retail products. Food Microbiol. 68, 16–23. doi: 10.1016/j.fm.2017.06.012
Pesavento, G., Ducci, B., Comodo, N., and Lo Nostro, A. (2007). Antimicrobial resistance profile of Staphylococcus aureus isolated from raw meat: a research for methicillin resistant Staphylococcus aureus (MRSA). Food Control 18, 196–200. doi: 10.1016/j.foodcont.2005.09.013
Peton, V., and Le Loir, Y. (2014). Staphylococcus aureus in veterinary medicine. Infect. Genet. Evol. 21, 602–615. doi: 10.1016/j.meegid.2013.08.011
Pinchuk, I. V., Beswick, E. J., and Reyes, V. E. (2010). Staphylococcal enterotoxins. Toxins 2, 2177–2197. doi: 10.3390/toxins2082177
Price, L. B., Stegger, M., Hasman, H., Aziz, M., Larsen, J., Andersen, P. S., et al. (2012). Staphylococcus aureus CC398: host adaptation and emergence of methicillin resistance in livestock. mBio 3:e00520-12. doi: 10.1128/mBio.00305-11
Rabello, R. F., Moreira, B. M., Lopes, R. M. M., Teixeira, L. M., Riley, L. W., and Castro, A. C. D. (2007). Multilocus sequence typing of Staphylococcus aureus isolates recovered from cows with mastitis in Brazilian dairy herds. J. Med. Microbiol. 56, 1505–1511. doi: 10.1099/jmm.0.47357-0
Rall, V. L. M., Miranda, E. S., Castilho, I. G., Camargo, C. H., Langoni, H., Guimarães, F. F., et al. (2014). Diversity of Staphylococcus species and prevalence of enterotoxin genes isolated from milk of healthy cows and cows with subclinical mastitis. J. Dairy Sci. 97, 829–837. doi: 10.3168/jds.2013-7226
Rešková, Z., Koreňová, J., and Kuchta, T. (2014). Effective application of multiple locus variable number of tandem repeats analysis to tracing Staphylococcus aureus in food-processing environment. Lett. Appl. Microbiol. 58, 376–383. doi: 10.1111/lam.12200
Riva, A., Borghi, E., Cirasola, D., Colmegna, S., Borgo, F., Amato, E., et al. (2015). Methicillin-resistant Staphylococcus aureus in raw milk: prevalence, SCCmec typing, enterotoxin characterization, and antimicrobial resistance patterns. J. Food Prot. 78, 1142–1146. doi: 10.4315/0362-028X.JFP-14-531
Rizek, C. F., Matté, M. H., Dropa, M., Mamizuka, E. M., de Almeida, L. M., Lincopan, N., et al. (2011). Identification of Staphylococcus aureus carrying the mecA gene in ready-to-eat food products sold in Brazil. Foodborne Pathog. Dis. 8, 561–563. doi: 10.1089/fpd.2010.0706
Saboya, L. V., Oliveira, A. J., Furtado, M. M., and Spadoti, L. M. (1998). Efeitos físico-químicos da adição de leite reconstituído na fabricação de queijo minas frescal. Ciên. Tecnol. Aliment. 18, 368–378. doi: 10.1590/S0101-20611998000400002
Sato’o, Y., Omoe, K., Naito, I., Ono, H. K., Nakane, A., Sugai, M., et al. (2014). Molecular epidemiology and identification of a Staphylococcus aureus clone causing food poisoning outbreaks in Japan. J. Clin. Microbiol. 52, 2637–2640. doi: 10.1128/JCM.00661-14
Scallan, E., Hoekstra, R. M., Angulo, F. J., Tauxe, R. V., Widdowson, M.-A., Roy, S. L., et al. (2011). Foodborne illness acquired in the United States—major pathogens. Emerg. Infect. Dis. 17, 7–15. doi: 10.3201/eid1701.091101
Schmid, D., Fretz, R., Winter, P., Mann, M., Höger, G., Stöger, A., et al. (2009). Outbreak of staphylococcal food intoxication after consumption of pasteurized milk products, June 2007, Austria. Wien. Klin. Wochenschr. 121, 125–131. doi: 10.1007/s00508-008-1132-0
Sharma, N. K., Rees, C. E. D., and Dodd, C. E. R. (2000). Development of a single-reaction multiplex PCR toxin typing assay for Staphylococcus aureus strains. Appl. Environ. Microbiol. 66, 1347–1353. doi: 10.1128/AEM.66.4.1347-1353.2000
Shepheard, M. A., Fleming, V. M., Connor, T. R., Corander, J., Feil, E. J., Fraser, C., et al. (2013). Historical zoonoses and other changes in host tropism of Staphylococcus aureus, identified by phylogenetic analysis of a population dataset. PLOS ONE 8:e62369. doi: 10.1371/journal.pone.0062369
Silva, N. C. C., Guimarães, F. F., Manzi, M. P., Budri, P. E., Gómez-Sanz, E., Benito, D., et al. (2013). Molecular characterization and clonal diversity of methicillin-susceptible Staphylococcus aureus in milk of cows with mastitis in Brazil. J. Dairy Sci. 96, 6856–6862. doi: 10.3168/jds.2013-6719
Silva, N. C. C., Guimarães, F. F., Manzi, M. P., Júnior, A. F., Gómez-Sanz, E., Gómez, P., et al. (2014). Methicillin-resistant Staphylococcus aureus of lineage ST398 as cause of mastitis in cows. Lett. Appl. Microbiol. 59, 665–669. doi: 10.1111/lam.12329
Silveira-Filho, V. M., Luz, I. S., Campos, A. P. F., Silva, W. M., Barros, M. P. S., Medeiros, E. S., et al. (2014). Antibiotic resistance and molecular analysis of Staphylococcus aureus isolated from cow’s milk and dairy products in northeast Brazil. J. Food Prot. 77, 583–591. doi: 10.4315/0362-028X.JFP-13-343
Smith, E. M., Green, L. E., Medley, G. F., Bird, H. E., Fox, L. K., Schukken, Y. H., et al. (2005). Multilocus sequence typing of intercontinental bovine Staphylococcus aureus isolates. J. Clin. Microbiol. 43, 4737–4743. doi: 10.1128/JCM.43.9.4737-4743.2005
Sobral, D., Schwarz, S., Bergonier, D., Brisabois, A., Feßler, A. T., Gilbert, F. B., et al. (2012). High throughput multiple locus variable number of tandem repeat analysis (MLVA) of Staphylococcus aureus from human, animal and food sources. PLOS ONE 7:e33967. doi: 10.1371/journal.pone.0033967
Solano, C., Echeverz, M., and Lasa, I. (2014). Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 18, 96–104. doi: 10.1016/j.mib.2014.02.008
Teixeira, L. A., Resende, C. A., Ormonde, L. R., Rosenbaum, R., Figueiredo, A. M., de Lencastre, H., et al. (1995). Geographic spread of epidemic multiresistant Staphylococcus aureus clone in Brazil. J. Clin. Microbiol. 33, 2400–2404.
Tondo, E. C., Guimarães, M. C., Henriques, J. A., and Ayub, M. A. (2000). Assessing and analysing contamination of a dairy products processing plant by Staphylococcus aureus using antibiotic resistance and PFGE. Can. J. Microbiol. 46, 1108–1114. doi: 10.1139/cjm-46-12-1108
Türkyilmaz, S., Tekbiyik, S., Oryasin, E., and Bozdogan, B. (2010). Molecular epidemiology and antimicrobial resistance mechanisms of methicillin-resistant Staphylococcus aureus isolated from bovine milk. Zoonoses Public Health 57, 197–203. doi: 10.1111/j.1863-2378.2009.01257.x
Turner, S., Pryer, K. M., Miao, V. P., and Palmer, J. D. (1999). Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 46, 327–338. doi: 10.1111/j.1550-7408.1999.tb04612.x
Valentin-Domelier, A.-S., Girard, M., Bertrand, X., Violette, J., François, P., Donnio, P.-Y., et al. (2011). Methicillin-susceptible ST398 Staphylococcus aureus responsible for bloodstream infections: an emerging human-adapted subclone? PLOS ONE 6:e28369. doi: 10.1371/journal.pone.0028369
van Loo, I. H. M., Diederen, B. M. W., Savelkoul, P. H. M., Woudenberg, J. H. C., Roosendaal, R., van Belkum, A., et al. (2007). Methicillin-resistant Staphylococcus aureus in meat products, the Netherlands. Emerg. Infect. Dis. 13, 1753–1755. doi: 10.3201/eid1311.070358
Veras, J. F., do Carmo, L. S., Tong, L. C., Shupp, J. W., Cummings, C., Dos Santos, D. A., et al. (2008). A study of the enterotoxigenicity of coagulase-negative and coagulase-positive staphylococcal isolates from food poisoning outbreaks in Minas Gerais, Brazil. Int. J. Infect. Dis. 12, 410–415. doi: 10.1016/j.ijid.2007.09.018
Vivoni, A. M., Diep, B. A., de Gouveia Magalhães, A. C., Santos, K. R. N., Riley, L. W., Sensabaugh, G. F., et al. (2006). Clonal composition of Staphylococcus aureus isolates at a Brazilian university hospital: identification of international circulating lineages. J. Clin. Microbiol. 44, 1686–1691. doi: 10.1128/JCM.44.5.1686-1691.2006
Voss, A., Loeffen, F., Bakker, J., Klaassen, C., and Wulf, M. (2005). Methicillin-resistant Staphylococcus aureus in pig farming. Emerg. Infect. Dis. 11, 1965–1966. doi: 10.3201/eid1112.050428
Votintseva, A. A., Miller, R. R., Fung, R., Knox, K., Godwin, H., Peto, T. E. A., et al. (2014). Multiple-strain colonization in nasal carriers of Staphylococcus aureus. J. Clin. Microbiol. 52, 1192–1200. doi: 10.1128/JCM.03254-13
Werle, E., Schneider, C., Renner, M., Völker, M., and Fiehn, W. (1994). Convenient single-step, one tube purification of PCR products for direct sequencing. Nucleic Acids Res. 22, 4354–4355. doi: 10.1093/nar/22.20.4354
Yan, X., Wang, B., Tao, X., Hu, Q., Cui, Z., Zhang, J., et al. (2012). Characterization of Staphylococcus aureus strains associated with food poisoning in Shenzhen, China. Appl. Environ. Microbiol. 78, 6637–6642. doi: 10.1128/AEM.01165-12
Keywords: Staphylococcus aureus, dairy industry, food safety, MLST, antibiotic resistance, staphylococcal enterotoxin, Brazil
Citation: Dittmann KK, Chaul LT, Lee SHI, Corassin CH, Fernandes de Oliveira CA, Pereira De Martinis EC, Alves VF, Gram L and Oxaran V (2017) Staphylococcus aureus in Some Brazilian Dairy Industries: Changes of Contamination and Diversity. Front. Microbiol. 8:2049. doi: 10.3389/fmicb.2017.02049
Received: 13 December 2016; Accepted: 06 October 2017;
Published: 24 October 2017.
Edited by:
Danilo Ercolini, University of Naples Federico II, ItalyReviewed by:
Giuseppe Spano, University of Foggia, ItalyCopyright © 2017 Dittmann, Chaul, Lee, Corassin, Fernandes de Oliveira, Pereira De Martinis, Alves, Gram and Oxaran. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lone Gram, Z3JhbUBiaW8uZHR1LmRr
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.