
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
FOCUSED REVIEW article
Front. Microbiol. , 25 August 2017
Volume 8 - 2017 | https://doi.org/10.3389/fmicb.2017.01638
 Aurélien Bailly1,2*
Aurélien Bailly1,2* Laure Weisskopf2,3
Laure Weisskopf2,3Microbial lifeforms associated with land plants represent a rich source for crop growth- and health-promoting microorganisms and biocontrol agents. Volatile organic compounds (VOCs) produced by the plant microbiota have been demonstrated to elicit plant defenses and inhibit the growth and development of numerous plant pathogens. Therefore, these molecules are prospective alternatives to synthetic pesticides and the determination of their bioactivities against plant threats could contribute to the development of control strategies for sustainable agriculture. In our previous study we investigated the inhibitory impact of volatiles emitted by Pseudomonas species isolated from a potato field against the late blight-causing agent Phytophthora infestans. Besides the well-documented emission of hydrogen cyanide, other Pseudomonas VOCs impeded P. infestans mycelial growth and sporangia germination. Current advances in the field support the emerging concept that the microbial volatilome contains unexploited, eco-friendly chemical resources that could help select for efficient biocontrol strategies and lead to a greener chemical disease management in the field.
Since the Neolithic Revolution about 12,000 years ago, the onset of plant domestication and the progressive systematization of agricultural practices have gradually led to monophyletic cropping systems, prone to pathogen outbreaks. Although the modern eras' mechanization, irrigation and chemical field management tremendously increased crop yields, today's agriculture faces the critical dilemma to meet global food demand and preserve environmental resources. In the context of climate change, productivity pressure and societal uncertainty over genetic manipulation, plant diseases and their management increasingly threaten food security and ecosystems. The promotion and intensification of sustainable farming practices relies on new biotechnological developments. Our growing understanding of the benefits brought by plant-associated microbes to crop health and growth has led to the realization that the plant-microbiome constitutes an untapped source of potential biocontrol agents, new valuable molecules and farming strategies (Mueller and Sachs, 2015).
KEY CONCEPT 1. Microbiome and Microbiota.
Often misused as synonyms, these two terms describe distinct definitions of microbial communities. The microbiota denotes the microorganisms that reside in an environmental niche. The microbiome refers to the collective genomes of these microorganisms.
Among the large diversity of microbial secondary metabolites, low molecular-weight volatile organic compounds (VOCs) have received growing attention in the past decade. Since the early reports describing the health- and growth-promoting effects of bacterial VOCs on model plants (Ryu et al., 2003, 2004), an increasing number of studies has evidenced the great potential of these gaseous molecules in crop enhancement and protection (reviewed in Bailly and Weisskopf, 2012; Kanchiswamy et al., 2015b). Microbial VOCs (mVOCs) are typically released in a multifarious and dynamic bouquet, essentially originating from the catabolic background, and comprise a majority of low-complexity, rather lipophilic compounds (Schulz and Dickschat, 2007; Blom et al., 2011a; Penuelas et al., 2014; Schenkel et al., 2015). Thus, mVOCs are seen as bona fide semiochemicals able to evaporate to the extracellular space, reach target organisms and partition into biological membranes or intracellular compartments. Indeed, microbial emissions have been shown to trigger significant volatile-mediated responses in bacteria (Garbeva et al., 2014; Audrain et al., 2015; Schulz-Bohm et al., 2015; Tyc et al., 2015), fungi (Effmert et al., 2012; Schmidt et al., 2015; Werner et al., 2016), plants (Bailly and Weisskopf, 2012; Pieterse et al., 2014; Kanchiswamy et al., 2015b), and invertebrates (D'alessandro et al., 2013; Davis et al., 2013). Although the molecular mechanisms underlying mVOCs perception by plants remain unclear, numerous studies have demonstrated that this system results in a potent priming of the plant basal immune system, termed induced systemic resistance (ISR), conferring broad-spectrum resistance against pathogens. In contrast to pattern-triggered immunity (PTI) and subsequent mounting of SAR, ISR elicitation does not negatively impact growth and productivity; in fact, many ISR-triggering microorganisms were selected for their plant growth-promoting and stress-relieving properties (reviewed in Van Hulten et al., 2006; Choudhary et al., 2007; Yang et al., 2009; Heil, 2010; Huot et al., 2014; Pieterse et al., 2014). Given the origin and chemical properties of mVOCs, these interkingdom cues represent a prospective pool of new functions that need further investigation and development to be delivered to the field (Fernando et al., 2005; Kanchiswamy et al., 2015a,b; Chung et al., 2016).
KEY CONCEPT 2. Volatile organic compounds.
VOCs are low-molecular weight, carbon-containing compounds (excluding very simple chemical species, such as carbon monoxide or carbon dioxide) that display high vapor pressure and low boiling point. Biogenic VOCs have been described as bona fide semiochemicals in most phyla.
KEY CONCEPT 3. Induced Systemic Resistance.
Induced resistance is a general term describing an induced state of resistance in plants triggered by the local perception of biotic or abiotic cues. Induced systemic resistance (ISR) describes the elicitation of latent plant defenses that systemically protects naive plant parts against future attackers, also termed defense priming. ISR activation depends on jasmonic acid and ethylene hormonal responses and is distinct from the systemic acquired resistance (SAR) engaged by the cellular recognition of microbe-associated molecular patterns (MAMPs), characterized by increased levels of the phytohormone salicylic acid.
Our recent work has focused on late blight, the major worldwide potato disease caused by the oomycete Phytophthora infestans. Although this particular pathosystem is obviously distinct from other fungal or bacterial plant diseases, our line of reasoning within this focused review could be extended to a wide range of plant pathogens. Under favorable conditions, P. infestans easily spreads from plant to plant through densely planted monocultures and rapidly ravages entire fields (Fry, 2008), and disease forecasting has become a key tool for growers. While conventional field practices control late blight via repeated, preventive applications of broad-spectrum fungicides, organic farming greatly relies on copper-based products toxic to the environment (Dorn et al., 2007; Cooke et al., 2011; Nechwatal and Zellner, 2015). The search for alternative organic solutions using either horticultural extracts, biosurfactants, or applications of plant beneficial bacteria or compounds eliciting plant defenses has not yet yielded reliable market products (Dupuis et al., 2007; Diallo et al., 2011). However, the increase in stringent policies regarding copper release into the environment exerts pressure for the continuation of investigations. In vitro work has demonstrated that mVOCs specifically contribute to the inhibition of growth and development of several phytopathogenic fungal or fungal-like genera, including members of Aspergillus (Vespermann et al., 2007; Hua et al., 2014; Chaves-Lopez et al., 2015; Gong et al., 2015), Botrytis (Huang et al., 2011; Li et al., 2012; Rouissi et al., 2013; Zhang et al., 2013; Parafati et al., 2015), Fusarium (Vespermann et al., 2007; Minerdi et al., 2009; Yuan et al., 2012; Tenorio-Salgado et al., 2013; Wang et al., 2013; Cordero et al., 2014), Penicillium (Rouissi et al., 2013), Sclerotinia (Fiddaman and Rossall, 1993, 1994; Fernando et al., 2005; Vespermann et al., 2007; Giorgio et al., 2015), Rhizoctonia (Fiddaman and Rossall, 1993, 1994; Kai et al., 2007; Vespermann et al., 2007; Liu et al., 2008), Alternaria (Andersen et al., 1994; Chaurasia et al., 2005; Trivedi et al., 2008; Zhao et al., 2011; Groenhagen et al., 2013), Pythium (Chaurasia et al., 2005; Sanchez-Fernandez et al., 2016), and Phytophthora (Zhao et al., 2011; Ann, 2012; Sharma et al., 2015).
Our recent study has therefore been centered on the hypothesis that the volatilomes of bacteria naturally associated with potato plants contain active compounds against P. infestans and that, once isolated, these antagonists would make ideal candidate biopriming control agents (Hu et al., 2014; Spence et al., 2014; Mahmood et al., 2016).
Throughout their whole lifecycle, land plants are continuously covered by environmental microorganisms colonizing their surfaces, invading intra- and intercellular spaces or building intimate symbiosis. Microbes have evolved life strategies displaying commensal, beneficial, or pathogenic behaviors toward plants to access the metabolic resources they offer. Plants are constantly challenged with biotic cues that need to be processed to balance growth, development and defense programs and achieve optimal fitness (Huot et al., 2014). They have developed a multilayered monitoring strategy that relies on the capacity of each individual cell to perceive molecular effectors and translate them into a systemic signal triggering an alert status in distant organs and on-site defense responses. In addition, recent insights into the host-specific composition of microbial communities have suggested that plants, to some extent, select for their microbiota (Bulgarelli et al., 2013; Schlaeppi and Bulgarelli, 2015). The intense competition between microbes for nutrients and favorable niches both at the rhizosphere and phyllosphere levels might in effect provide a functional addition to the plant immune system, and host-specific microbiomes can essentially be seen as an extension of the plant genome. As laboratory model organisms capable of increasing plant health may prove difficult to transfer to field conditions, the isolation of highly-adapted strains from the plant in situ microbiota has a much greater chance of success in antagonist selection processes.
KEY CONCEPT 4. Rhizosphere and phyllosphere.
In microbiology, the term rhizosphere refers to the thin volume of soil directly influenced by plant root exudates and root-associated microorganisms, while the phyllosphere describes the above-ground plant surfaces hosting microbial species. Both represent dynamic habitats with drastically different resources and environmental conditions for microorganisms.
We therefore isolated 137 morphologically distinct bacterial strains on different growth media from the rhizosphere and phyllosphere of field-grown potato plants previously infected with P. infestans (Hunziker et al., 2015). Subsequent phylogenetic identification of 92 of these strains to the genus or species level using 16S and rpoD gene amplicons revealed that Actinobacteria and Proteobacteria were the most abundant among the isolated organisms. Although our sampling and isolation methods were not exhaustive and higher resolution of OTUs has been described elsewhere (Inceoglu et al., 2011; Barnett et al., 2015), the retrieved strains are bona fide potato-associated bacteria. From the 32 bioactive strains pre-selected from a series of dual-culture assays against B. cinerea, R. solani, and P. infestans growth, Pseudomonas species had the highest inhibitory potential.
In order to evaluate the volatile-mediated activity of these strains, we then co-cultured bacterial colonies and five discrete target potato pathogens in physically-separated compartments using the I-plate Petri dish system (Hunziker et al., 2015). This work revealed that (1) P. infestans was the most VOC-susceptible target organism, (2) Pseudomonas species displayed the highest volatile-mediated activity, and (3) that hydrogen cyanide production could account for a large part of the observed inhibition. The large difference between the susceptibility of P. infestans and true fungi to mVOCs could partly be explained by the differing nature of its cell wall. Additionally, in our hands, the increase in VOCs-mediated inhibition of fungal and fungal-like species' radial growth seemed to correlate with slower growth speed (Groenhagen et al., 2013; De Vrieze et al., 2015; Hunziker et al., 2015).
The contribution of volatile HCN to the biocontrol properties of Pseudomonas strains against fungal pathogens has been known for 20 years, since the demonstration of the suppression of Thielaviopsis-induced tobacco black root rot by the cyanogenic P. protegens CHA0 but not by its isogenic mutant P. protegens CHA77 (Voisard et al., 1989; Haas and Defago, 2005; Rudrappa et al., 2008; Lanteigne et al., 2012). In the same extent, other inorganics of bacterial origin, such as ammonia, or hydrogen sulfide are suspected to account for a significant part of the target organism growth inhibition (Bernier et al., 2011; Shatalin et al., 2011; Weise et al., 2013). However, throughout our experimental work, no correlation was found between P. infestans mycelial growth and bacterial NH3 production (Hunziker et al., 2015). Moreover, the oomycete was still significantly inhibited when exposed to the volatile blend of the cyanide-deficient mutant CHA77, thus indicating that beside HCN and NH3, Pseudomonas strains release other potent volatiles against P. infestans.
This indicated that the identification and quantification of the volatile chemical species composing the natural emissions of cyanogenic and non-cyanogenic bacteria is a prerequisite to the evaluation of their contribution to the inhibitory impact on the target pathogen (Kai et al., 2007, 2009; Campos et al., 2010; Effmert et al., 2012). Such approaches, essentially based on molecule-trapping techniques and gas chromatography-mass spectrometry (GC-MS) platforms, became the standard in the field (Schulz and Dickschat, 2007), thus generating vast amounts of data in which non-abundant and/or non-readily available chemical species are generally overlooked. Indeed, while a large body of literature has reported the inhibitory activity of a broad range of bacterial volatilomes against several discrete fungal or fungal-like pathogens, the identity of single active VOCs remains elusive. In many studies, the application of identified compounds as physiologically relevant amounts of synthetic molecules rarely reached the inhibitory effects observed with natural VOC bouquets, suggesting that volatile blends act in a multifactorial manner (Yuan et al., 2012; Groenhagen et al., 2013; Chaves-Lopez et al., 2015). Recent studies have tentatively reconstituted artificial mixtures of several prominent volatile species and reported their greater effects when compared to single compound applications, suggesting that volatiles interact synergistically (Cortes-Barco et al., 2010a; Fialho et al., 2010, 2011a,b; Mitchell et al., 2010; Naznin et al., 2013; Riyaz-Ul-Hassan et al., 2013).
Extending our initial investigation of 8 Pseudomonas volatilomes (Hunziker et al., 2015), we collected and identified the compounds emitted by CHA0, CHA77, and 16 of our selected Pseudomonas strains grown on lysogeny broth plates for 24 h, under conditions mimicking our I-plate assays. We hypothesized that each strain's specific volatile-mediated inhibition potential could be explained by either a different population or different amounts of single VOCs in the volatile blends. The obtained chemoprofiles comprised volatile motifs previously identified in Pseudomonas biogenic emissions, with 1-undecene and dimethyl disulfide (DMDS) being the most prominent species (Lemfack et al., 2014; Hunziker et al., 2015), and appeared relatively conserved, thus supporting the concept that volatile signatures could help discriminate microbial genera or species (Thorn et al., 2011; Shestivska et al., 2015; Dryahina et al., 2016; Neerincx et al., 2016). However, detailed comparisons of the collected GC-MS data failed to identify the chemical features responsible for the strains VOCs-mediated inhibitory effects (Figure 1). The genetic proximity of our Pseudomonas isolates did not necessarily translate into identical chemoprofiles (Shestivska et al., 2012; De Vrieze et al., 2015), and it appeared that the impact of the rhizospheric or phyllospheric origin was negligible in our sampling. A previous study investigating the effects of volatiles emitted by closely-related Burkholderia ambifaria strains with discrete isolation origins on various target organisms also reported very similar, yet different VOC chemoprofiles, leading to very subtle changes in the targets' responses (Groenhagen et al., 2013). It is highly plausible that the rich LB medium on which we grew our strains during headspace collection did not select for and reflect the particular metabolic potential of our test-strains, but we expect the collected spectral data to mirror our inhibition assays. Moreover, univariate pair fold change analysis between CHA0 and CHA77 chemoprofiles displayed over 90 significantly different mass features (t-test, p < 0.005), including enrichments in dimethyl trisulfide (DMTS), s-methyl methanethiosulfonate (MMTS) and aminoacetophenone production in the non-cyanogenic mutant (Figure 2). Interestingly, HCN itself is not detected in standard GC-MS methods and so does not impact the differences observed between mutant and wild-type strains' chemoprofiles, thus a change in the synthesis of one particular volatile can lead to a drastic alteration of the overall volatile profiles emitted by otherwise isogenic strains. When focusing on non-cyanogenic isolates, relatively poor PCA clusterings of the total ion GC-MS chromatograms tended to separate the most active strains' chemoprofiles from low activity ones (Figure 3), although no single compound or chemical pattern seemed to unequivocally explain the blend's effect. The abundance or the detection frequency of compounds that do not substantially contribute to the total effect of the whole volatile blend may impede the description of bioactive patterns. Thus, in order to strengthen chemoprofiling data and identify key chemical species, the precise determination of the inhibition potential of individual substances is essential.
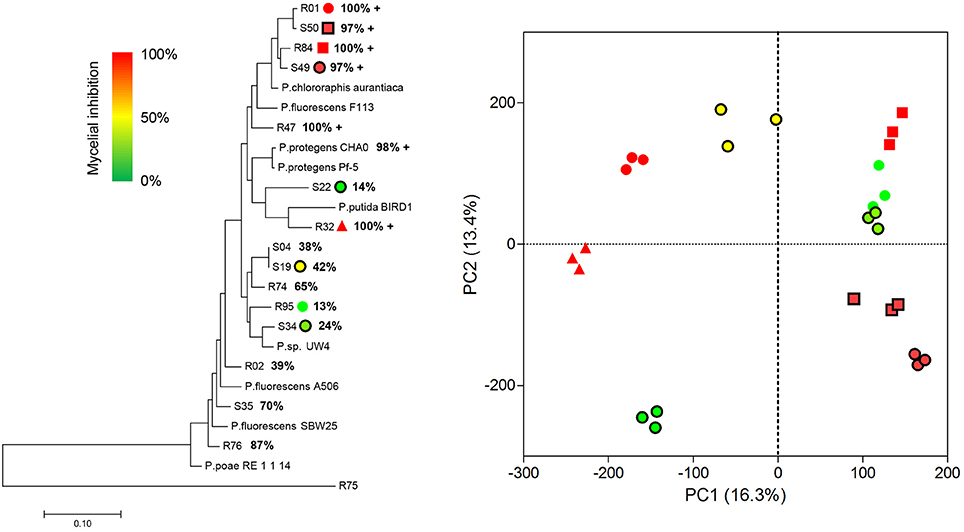
Figure 1. Volatile organic compounds production in Pseudomonas sp. does not mirror phylogenetic relationships. Left, maximum-likelihood-based phylogenetic tree of 24 Pseudomonas sp. calculated from multi-locus sequence alignments of 16s rRNA, gyrB, rpoD, and rpoB genes concatenations using the MEGA software. Bars indicate mean average base substitutions between sequences. Note that the Flavobacterium sp. R75 stands as an outlier. Bold percentage values represent the volatile-mediated inhibition of P. infestans mycelial growth for each tested strain. R and S refer to strains from rhizospheric and phyllospheric origin, respectively; +, cyanogenic stain. Right, 2-D scores plot between the highest PC scores from a principal components analysis of the total ion chromatograms of 9 selected Pseudomonas isolates. Individual biological replicates of each strain are identified according to the legend in the tree (left); each data point is color-coded according to the VOCs-mediated P. infestans mycelial growth inhibition exerted by the corresponding strain. Spectral data were processed using the MetaboAnalyst 3.0 software following this procedure: detection of Gaussian-fitted peaks (4 s fwhm, binning, integrated area of original peak), followed by alignment and grouping according to their masses and retention time after retention time correction. Data were subsequently filtered based on interquantile range, then normalized by the sample median and finally generalized log-transformed. Data scaling and outlier removal were left in automatic mode. Explained variances are shown in brackets.
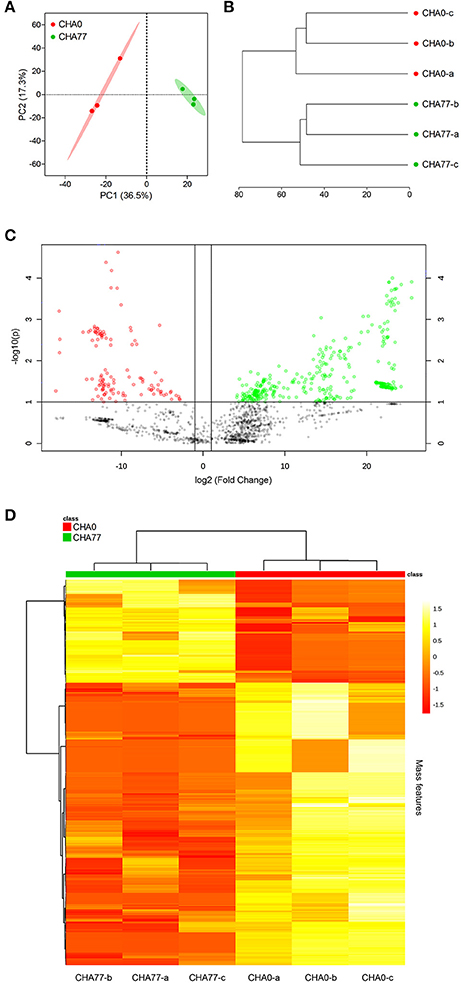
Figure 2. Pseudomonas protegens CHA0 and its isogenic, cyanide-deficient mutant CHA77 express distinct volatilomes. (A) PCA 2-D scores plot from total ion chromatograms processed as in Figure 1. Shaded areas represent the 95% confidence interval; explained variances are shown in brackets. (B) Hierarchical clustering dendrogram of the biological replicate volatilomes (Euclidean distance similarity and Ward's linkage clustering). (C) Volcano plot of mass features differentially detected in CHA77 compared to CHA0 volatilomes selected by fold change (threshold = 2) and t-test values (threshold = 0.1). Note the higher number of mass features with positive fold change (green) compared to negative fold change (red) in the HCN-deficient mutant. (D) Heat map and hierarchical clustering of the top 300 t-test selected mass features. Spectral data were processed as described for Figure 1.
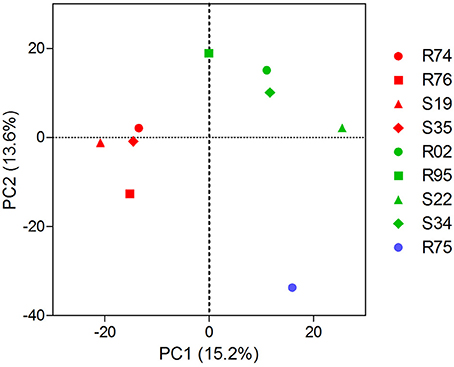
Figure 3. Non-cyanogenic Pseudomonas sp. chemoprofiles may contain compounds explaining the different inhibition potential against P. infestans. 2-D scores plot between the highest PC scores from a principal components analysis of the total ions chromatograms of 9 non-cyanogenic strains processed as in Figure 1. Data points represent the centroids of 3 biological replicates. Note that, besides a net separation between high-activity (red) and low-activity strains (green), the variance explained by each component is low. The chemoprofiles of the non-Pseudomonas strain R75 (blue) separate from Pseudomonas chemoprofiles. Explained variances are shown in brackets. Spectral data were processed as described for Figure 1.
The large amount of data contained in mass spectra, coupled to the difficulties in identification of the chemical structures they refer to, make systematic testing of the bioactivities of individual compounds a daunting task. Moreover, the limited number of comparative studies involving different microbial genera (Kai et al., 2009; Blom et al., 2011a,b; Berrada et al., 2012) does not allow the assessment of candidate active volatiles that may be present or absent in the respective volatilomes, leaving a striking knowledge gap. With the aim of assigning a weight function to our chemoprofiles, we attempted to characterize the precise contribution of 40 commercially available pure substances identified from the natural emissions of our isolates by assessing their biological activity against several stages of P. infestans life cycle. Although non-exhaustive, this series of assays revealed that a majority of Pseudomonas volatiles possess low to mild inhibitory power against Phytophthora and probably act synergistically on the target organism (De Vrieze et al., 2015). Although limited to a small panel of simple compounds, the quantitative relative IC50 values derived from dose-dependent P. infestans mycelial growth and sporangial germination inhibition assay allow for basic structure-activity relationship exploration (Figure 4). First, P. infestans sporangia appeared more sensitive to mVOCs exposure than mycelia, especially to aliphatic compounds, such as long chain aldehydes (undecanal and tridecanal), alkenes (1-undecene and 1-dodecene), and short-chained ketones (2-octanone, 2-heptanone, 4-heptanone, 3-hexanone, and 4-hydroxy-4-methyl-2-pentanone), while 2-dodecanol or undecane were found to be inactive. Interestingly, similar activities have been previously reported against Alternaria alternata germ tube growth for this chemical family (Andersen et al., 1994), implying that these lipoxygenase products may cause broad-spectrum interference to fungal and fungal-like germ tube development. This is further supported by the fact that the exposure to a subset of ketones, such as 3-hexanone triggered severe malformations in P. infestans germ tubes (De Vrieze et al., 2015). The closely-related compounds furfuryl alcohol and acetyl furan, as well as three of the six phenyl-ketones tested also performed well against P. infestans germination. Acetophenone derivatives are well-described antifungals thought to target the fungal cell wall (Soberon et al., 2015; De Aguiar et al., 2016). A high level of inhibition also resulted from exposure to diphenylamine and 2,5-dimethylpyrazine treatments; however we cautiously consider these compounds as artifacts originating from the medium. Secondly, both mycelia and sporangia showed high sensitivity to sulfur-containing compounds, such as bis(methylthiomethyl)sulfide, s-methylbunathioate, MMTS and DMTS, isovaleric acid and nitropentane. Nitroalkanes are renowned toxic substances for animals, but to the best of our knowledge, no particular study has investigated their effect on fungal growth. However, some very potent non-volatile antimicrobials display an active nitro-group, for instance nitrofurazone, metronidazole and chloramphenicol. DMDS and to a lesser extent DMTS and MMTS have been repeatedly shown to exert broad-range antifungal activities, probably via their capacity to reduce protein sulfhydryl groups and readily oxidize into highly reactive sulfur-acids, and are considered as prominent antimicrobials in the Brassicacea and Allioideae (Fernando et al., 2005; Kocic-Tanackov et al., 2012; Groenhagen et al., 2013; Zhou et al., 2014). DMDS-containing products are already marketed as soil fumigants for the suppression of soil-borne plant diseases. However, DMDS poorly performed in our assays, with the exception of zoospore activity. The inhibition of P. infestans radial growth and sporangia germination by isovaleric acid is especially interesting as this compound was shown to trigger the germination of Agaricus bisporus spores (Rast and Stauble, 1970) but not of ectomycorrhizal fungi (Fries, 1978), and to inhibit Fusarium growth (Monnet et al., 1988). Most other free fatty acids are considered as broad-range fungal inhibitors interfering with membrane composition (Pohl et al., 2011).
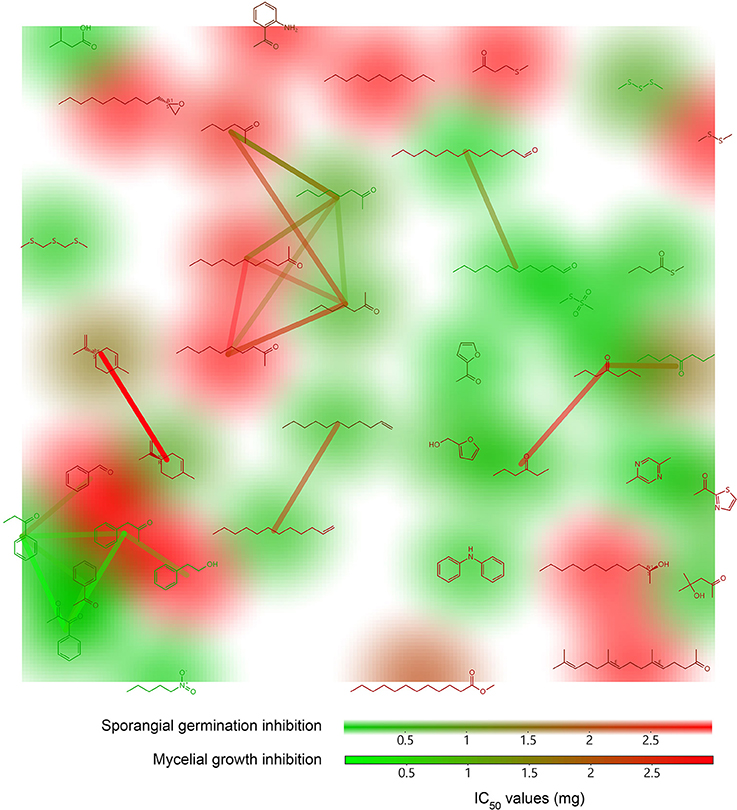
Figure 4. Structure-activity landscape plots of 40 Pseudomonas volatile organic compounds tested against P. infestans Neighbor analysis, Murcko scaffold analysis and FragFp structure similarity analysis were performed using the Actelion Pharmaceuticals DataWarrior 4.4.3 software. Structures are colored according to their P. infestans mycelial growth inhibitory activity while structure backgrounds display sporangia germination inhibition. Connection lines between different structures indicate the number of neighbor molecules (red, low number; green, high number).
Finally, few compounds were solely active against P. infestans mycelia without also impacting sporangia development and function, and these belong to the chemical groups discussed above. Furthermore, these molecules, namely 2-phenylethanol, 2-phenylacetone, 2-octanone and 4-octanone, showed mild-to-low inhibitory power. Remarkably, several reports focusing on the antifungal activity of ketones identified from Bacillus species (Fernando et al., 2005; Arrebola et al., 2010; Yuan et al., 2012; Zhang et al., 2013) concluded that long-chain ketones, such as 2-nonanone and 2-decanone demonstrated strong inhibition activity against fungal species. However, in our work, long-chain ketones treatments did not provide satisfactory inhibition of P. infestans, although 2-undecanone exposure led to a strong densification of the mycelial mat (De Vrieze et al., 2015). Similarly, a study by Chaves-Lopez et al. (2015) focusing on single volatiles from Bacillus documented that only short-chain ketones like 2-butanone were efficient against Fusarium oxysporum and Moniliophthora perniciosa growth. These discrepancies may simply be explained by the different sensitivities of the studied target organisms to mVOCs, but they may also find their source in the low pharmacological resolution of the employed methodology. In Yuan et al. (2012), F. oxysporum was exposed to 200 μl of a subset of Bacillus VOCs while in Chaves-Lopez et al. (2015), 25 and 50 μl of another subset of Bacillus VOCs were applied. Regardless of the boiling point of the particular compounds, these amounts represent tens to hundreds of milligrams introduced into the headspace, far beyond the actual production capacity of the bacterial cultures (De Vrieze et al., 2015; Shestivska et al., 2015). As VOCs readily diffuse to the environment, attention should be paid to substances with the highest potency and low dose efficacy. We therefore advocate the systematic assignment of pharmacological values based on standardized bioassays against the investigated target organisms, to the chemical species identified from microbial volatilomes. Our current work strives for the successful implementation of these valuable data layers that will allow deeper assessment of the ecological impact of biogenic microbial emissions and greatly help in pinpointing potent molecules or cornering active chemical backbones produced by various bacterial genera. Alternatively, these compounds could provide leads to drug discovery strategies, as exemplified by volatile benzothiazole (Herrera Cano et al., 2015; Zhao et al., 2016) and 2,4-diacetylphloroglucinol (Lanteigne et al., 2012), or help to select for the most appropriate antagonists from a panel of bioactive mVOCs.
As an emanation of the metabolome of a given organism in a given condition, the collected volatile blends represent only snapshots of a more complex phenomenon. Different substrate use, various growth conditions and genetic mutations are just some of the factors that directly influence the chemical composition of volatilomes (Fiddaman and Rossall, 1994; Blom et al., 2011a; De Vrieze et al., 2015). Furthermore, the natural conditions and environmental cues leading to the production of particular volatile species or signatures have not yet been resolved, and their biological relevance in biocontrol contexts remain to be assessed outside in vitro systems (issues reviewed in Schmidt et al., 2015; Chung et al., 2016). Therefore, the definition of the volatilome is not fixed to the capacity to enzymatically produce a particular compound as engraved in the genomes, but is instead relative to the dynamics of headspace compound release. In analogy to transcriptional patterns, the effect of mVOC emissions on a given target organism may depend on the production of a combination of key chemical species. Yet, in order to better characterize the impact and functions of mVOCs in interspecies relationships, a transition from low-scale individual studies to global data mining platforms is required. The experience gained in other -omics fields, especially the emergence of transcriptomic data, has led to the organization of public data repositories and the creation of resourceful toolsets that have tremendously stimulated research over the last 15 years, such as the NCBI Gene Expression Omnibus (Edgar et al., 2002). However, to a large extent, the sum of complex chemical information related to volatile production by microorganisms gathered in laboratories scattered around the world remains underexploited. As no centralized platforms exist that would allow comparative, statistically-driven exploration of published datasets, the era of metadata analysis of volatilomes has been delayed. The standardized procedures instigated in breath research (King et al., 2011; De Lacy Costello et al., 2014; Broza et al., 2015) should inspire investigators interested in the volatilomes of plant-associated microbiota. Recent attempts to pull together and unify data issued from the literature has resulted in the mVOCs (http://bioinformatics.charite.de/mvoc/; Lemfack et al., 2014) or the KNApSAcK Metabolite Ecology (http://kanaya.naist.jp/KNApSAcK/; Abdullah et al., 2015) databases, yet such initiatives require further development to become valuable instruments. Ideally, standardized NMR/MS peak lists or LC/GC-MS spectra (converted into exchange formats, such as NetCDF or mzXML) obtained from biological replicates would populate a growing database of discrete organisms, strains and experimental conditions that could serve as a basis for exploratory statistical analyses using existing metabolomics tool suites, such as MetaboAnalyst (Xia et al., 2015) or XCMS (Smith et al., 2006). Such advances would help to fill critical knowledge gaps, i.e., the determination of a core volatilome in a given species, the co-occurrence of underrepresented low-abundance mVOCs and the actual composition of emissions released by microorganisms growing in the rhizosphere or phyllosphere. Taken together, this information will provide key concepts to convert the explorative academic knowledge into concrete crop disease control solutions.
KEY CONCEPT 5. Volatilome.
The volatilome, also referred to as volatolome, defines the sum of volatile or semi-volatile organic compounds emitted by a biological system under specific experimental conditions. As the transcriptome describes the dynamic expression of genes through mRNA level detection, the quantitative identification of chemical species in the volatilome reflects the dynamic metabolic activity of the studied organism.
The concept of exploiting microbial populations hosted by plants to benefit crop health against one or more plant pathogens and productivity is ancient, but has received increasing attention in the past decades, especially in view of the potential biological and ecological functions conferred by rhizospheric and phyllospheric bacterial species (Zahir et al., 2003; Choudhary et al., 2011; Kim et al., 2011; Kumar et al., 2016). Biocontrol strains can bestow disease suppression via competition or parasitism against the targeted pathogens, antibiotic production, cell wall degradation or plant ISR elicitation. The most effective antagonists should display a range of microbicidal properties, as illustrated by Pseudomonas species potent in the production of a variety of phenazines, DAPG, pyrollnitrin, HCN (Lanteigne et al., 2012; Loper et al., 2012), and in our opinion, novel efficient mVOCs. These potentials are encoded in the genomes of the microbes and therefore, ever-decreasing DNA sequencing costs allow the prospective mining of genomes for desired functions (Loper et al., 2012).
There has been remarkable progress in defining biocontrol agents and their spread to the environment (Bale et al., 2008) which raises hopes for operational, intensive and yet sustainable agriculture in the next decades. However, regardless of the efforts made toward intensification of bioprospecting, the current bottleneck in delivering tangible applications to farmers results from difficulties in producing formulations suitable for modern agriculture (Lucy et al., 2004; Choudhary et al., 2011; Pérez-Montaño et al., 2014; Velivelli et al., 2014). The potency of biological agents and of (soluble) microbial derivatives has already been well-documented, and had grown into agronomical products (e.g., Mycostop® and Rhizoplus®, utilizing Bacillus species; Biocon® and Ecofit®, with Trichoderma sp. as active ingredient, or Cerall® and Proradix® containing Pseudomonas sp.), but the development of VOC-derived technologies is still embryonic. However, the successful volatile-based mating disruption of pest insects semiochemicals (Reddy and Guerrero, 2010; Lance et al., 2016) stands for an encouraging proof-of-concept. The rather sharp transition from the laboratory to the field has often been smoothed by a switch from model plants like Arabidopsis thaliana to economically important plants and greenhouse experiments. The volatile compound 2,3-butanediol, well-studied in Arabidopsis (Ryu et al., 2003, 2004; Farag et al., 2006; Han et al., 2006; Cho et al., 2008; Cortes-Barco et al., 2010a,b), was reported to reduce Colletotrichum-mediated anthracnose symptoms in Nicotiana benthamiana (Cortes-Barco et al., 2010b) and to protect Agrostis stolonifera against the fungal pathogens Microdochium nivale, Rhizoctonia solani, or Sclerotinia homoeocarpa (Cortes-Barco et al., 2010a). Still, in a recent field trial, attempts to reproduce in vitro results obtained with 3-pentanol and 2-butanone showed limited protection against a pathogenic Pseudomonas syringae (Song and Ryu, 2013), thus underlining the difficulties in delivering mVOC-based technology. In the case of our potato-Phytophthora pathosystem, we verified that the isolated Pseudomonas strains did not compromise plant health or growth in greenhouse pot cultures. The inoculated potato cultivars did not display any phytotoxicity symptoms or growth defects; but neither strain treatments resulted in growth enhancement (Guyer et al., 2015 and unpublished results). In addition, the ability of our candidate bioncontrol strains to colonize roots and survive on the potato phylloplane was assessed after sprout inoculation or leaf spraying, respectively. The large majority of the isolates demonstrated good rhizocompetence and successfully colonized plant shoots, both in the greenhouse and the field conditions (Guyer et al., 2015). As microbial competition for nutrients and ecological niches on the plant surfaces certainly contributes to the antagonistic activity of competent bacterial strains (Innerebner et al., 2011; Ghirardi et al., 2012; Vorholt, 2012), isolates naturally associated with potato plants have the highest chance to be artificially reintroduced to a crop for control purposes. The promising protective effects measured in dual culture assays and leaf disc infection experiments however, have not yet been transposed to successful field trials (Guyer et al., 2015).
The direct contribution of microbial VOCs in disease suppression in the open field remains elusive, and a study by Sharifi and Ryu (2016) argues that the VOCs-mediated elicitation of ISR is the primary factor in warding off pathogens, while direct inhibition via volatiles has only a minor impact. However, recent investigations by Tahir and colleagues demonstrated that VOCs emitted by well-studied suppressive Bacillus species act at multiple levels against the tobacco wilt agent Ralstonia solanacearum. Indeed, while in vitro work showed that exposure to the Bacillus volatile compounds decreased Ralstonia growth and viability and led to substantial defects in cell integrity and mobility, they as well triggered major changes in the expression of Ralstonia genes fundamental to disease progression (Tahir et al., 2017). Furthermore, tobacco plants exposed to Bacillus emissions and pure identified VOCs increased their transcription levels in key defense-related genes, such as NPR1 and EDS1, thus engaging systemic resistance and resulting in suppression (Tahir et al., 2017). It is therefore conceivable that bacterial volatiles contribute both directly and indirectly to the observed biocontrol properties of Bacillus, and that bacterial VOCs bouquets generally act as multifactorial, sequential or simultaneous signals on both pathogens and hosts.
The argument that volatiles dissipate in the environment and never reach efficient inhibiting doses may be valid at a macroscopic scale; nevertheless, competition between microbes on plant surfaces occurs in matrixes like the root mucilage or closed compartments, such as the sub-stomatal chamber, where well-adapted bacterial species may prosper and accumulate higher levels of VOCs. As these environments represent favored entry points for pathogens, we believe that the volatilome forms part of the bacterial arsenal and provides a supplementary line of plant defense. Future disease management integrating the use of biological agents for their water-soluble and volatile features in decision-making processes will lead to alternative solutions to effectively reduce pesticide and fertilizer use in an economically and environmentally sound manner.
AB and LW wrote the manuscript.
Agroscope Research Program “Microbial Biodiversity.”
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors a grateful to Dr. Kirsty Agnoli for the careful proofreading of this manuscript. Financial support from the Swiss National Science Foundation (grant 31003A_149271 to LW) is gratefully acknowledged.

Abdullah, A. A., Altaf-Ul-Amin, M., Ono, N., Sato, T., Sugiura, T., Morita, A. H., et al. (2015). Development and mining of a volatile organic compound database. Biomed. Res. Int. 2015:139254. doi: 10.1155/2015/139254
Andersen, R. A., Hamiltonkemp, T. R., Hildebrand, D. F., Mccracken, C. T., Collins, R. W., and Fleming, P. D. (1994). Structure-antifungal activity relationships among volatile C-6 and C-9 aliphatic-aldehydes, ketones, and alcohols. J. Agric. Food Chem. 42, 1563–1568. doi: 10.1021/jf00043a033
Ann, Y. C. (2012). Rhizobacteria of pepper (Piper nigrum) and their antifungal activities. Afr. J. Microbiol. Res. 6, 4185–4193. doi: 10.5897/AJMR12.583
Arrebola, E., Sivakumar, D., and Korsten, L. (2010). Effect of volatile compounds produced by Bacillus strains on post-harvest decay in citrus. Biol. Control 53, 122–128. doi: 10.1016/j.biocontrol.2009.11.010
Audrain, B., Farag, M. A., Ryu, C.-M., and Ghigo, J.-M. (2015). Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 39, 222–233. doi: 10.1093/femsre/fuu013
Bailly, A., and Weisskopf, L. (2012). The modulating effect of bacterial volatiles on plant growth: current knowledge and future challenges. Plant Signal. Behav. 7, 79–85. doi: 10.4161/psb.7.1.18418
Bale, J. S., Van Lenteren, J. C., and Bigler, F. (2008). Biological control and sustainable food production. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 363, 761–776. doi: 10.1098/rstb.2007.2182
Barnett, B. A., Holm, D. G., Koym, J. W., Wilson, R. G., and Manter, D. K. (2015). Site and clone effects on the potato root-associated core microbiome and its relationship to tuber yield and nutrients. Am. J. Potato Res. 92, 1–9. doi: 10.1007/s12230-014-9405-9
Bernier, S. P., Letoffe, S., Delpierre, M., and Ghigo, J. M. (2011). Biogenic ammonia modifies antibiotic resistance at a distance in physically separated bacteria. Mol. Microbiol. 81, 705–716. doi: 10.1111/j.1365-2958.2011.07724.x
Berrada, I., Benkhemmar, O., Swings, J., Bendaou, N., and Amar, M. (2012). Selection of halophilic bacteria for biological control of tomato gray mould caused by Botrytis cinerea. Phytopathol. Mediterr. 51, 625–630. doi: 10.14601/Phytopathol_Mediterr-10627
Blom, D., Fabbri, C., Connor, E. C., Schiestl, F. P., Klauser, D. R., Boller, T., et al. (2011a). Production of plant growth modulating volatiles is widespread among rhizosphere bacteria and strongly depends on culture conditions. Environ. Microbiol. 13, 3047–3058. doi: 10.1111/j.1462-2920.2011.02582.x
Blom, D., Fabbri, C., Eberl, L., and Weisskopf, L. (2011b). Volatile-mediated killing of Arabidopsis thaliana by bacteria is mainly due to hydrogen cyanide. Appl. Environ. Microbiol. 77, 1000–1008. doi: 10.1128/AEM.01968-10
Broza, Y. Y., Mochalski, P., Ruzsanyi, V., Amann, A., and Haick, H. (2015). Hybrid volatolomics and disease detection. Angew. Chem. Int. Ed. Engl. 54, 11036–11048. doi: 10.1002/anie.201500153
Bulgarelli, D., Schlaeppi, K., Spaepen, S., Ver Loren Van Themaat, E., and Schulze-Lefert, P. (2013). Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 64, 807–838. doi: 10.1146/annurev-arplant-050312-120106
Campos, V. P., De Pinho, R. S. C., and Freire, E. S. (2010). Volatiles produced by interacting microorganisms potentially useful for the control of plant pathogens. Ciencia E Agrotecnol. 34, 525–535. doi: 10.1590/S1413-70542010000300001
Chaurasia, B., Pandey, A., Palni, L. M. S., Trivedi, P., Kumar, B., and Colvin, N. (2005). Diffusible and volatile compounds produced by an antagonistic Bacillus subtilis strain cause structural deformations in pathogenic fungi in vitro. Microbiol. Res. 160, 75–81. doi: 10.1016/j.micres.2004.09.013
Chaves-Lopez, C., Serio, A., Gianotti, A., Sacchetti, G., Ndagijimana, M., Ciccarone, C., et al. (2015). Diversity of food-borne Bacillus volatile compounds and influence on fungal growth. J. Appl. Microbiol. 119, 487–499. doi: 10.1111/jam.12847
Cho, S. M., Kang, B. R., Han, S. H., Anderson, A. J., Park, J. Y., Lee, Y. H., et al. (2008). 2R, 3R-butanediol, a bacterial volatile produced by Pseudomonas chlororaphis O6, is involved in induction of systemic tolerance to drought in Arabidopsis thaliana. Mol. Plant Microbe Interact. 21, 1067–1075. doi: 10.1094/MPMI-21-8-1067
Choudhary, D. K., Prakash, A., and Johri, B. N. (2007). Induced systemic resistance (ISR) in plants: mechanism of action. Indian J. Microbiol. 47, 289–297. doi: 10.1007/s12088-007-0054-2
Choudhary, D. K., Sharma, K. P., and Gaur, R. K. (2011). Biotechnological perspectives of microbes in agro-ecosystems. Biotechnol. Lett. 33, 1905–1910. doi: 10.1007/s10529-011-0662-0
Chung, J. H., Song, G. C., and Ryu, C. M. (2016). Sweet scents from good bacteria: case studies on bacterial volatile compounds for plant growth and immunity. Plant Mol. Biol. 90, 677–687. doi: 10.1007/s11103-015-0344-8
Cooke, L. R., Schepers, H. T. A. M., Hermansen, A., Bain, R. A., Bradshaw, N. J., Nielsen, B. J., et al. (2011). Epidemiology and Integrated Control of Potato Late Blight in Europe. Potato Res. 54, 183–222. doi: 10.1007/s11540-011-9187-0
Cordero, P., Principe, A., Jofre, E., Mori, G., and Fischer, S. (2014). Inhibition of the phytopathogenic fungus Fusarium proliferatum by volatile compounds produced by Pseudomonas. Arch. Microbiol. 196, 803–809. doi: 10.1007/s00203-014-1019-6
Cortes-Barco, A. M., Hsiang, T., and Goodwin, P. H. (2010a). Induced systemic resistance against three foliar diseases of Agrostis stolonifera by (2R,3R)-butanediol or an isoparaffin mixture. Ann. Appl. Biol. 157, 179–189. doi: 10.1111/j.1744-7348.2010.00417.x
Cortes-Barco, A. M., Goodwin, P. H., and Hsiang, T. (2010b). Comparison of induced resistance activated by benzothiadiazole, (2R,3R)-butanediol and an isoparaffin mixture against anthracnose of Nicotiana benthamiana. Plant Pathol 59, 643–653. doi: 10.1111/j.1365-3059.2010.02283.x
D'alessandro, M., Erb, M., Ton, J., Brandenburg, A., Karlen, D., Zopfi, J., et al. (2013). Volatiles produced by soil-borne endophytic bacteria increase plant pathogen resistance and affect tritrophic interactions. Plant Cell Environ. 37, 813–826. doi: 10.1111/pce.12220
Davis, T. S., Crippen, T. L., Hofstetter, R. W., and Tomberlin, J. K. (2013). Microbial volatile emissions as insect semiochemicals. J. Chem. Ecol. 39, 840–859. doi: 10.1007/s10886-013-0306-z
De Aguiar, F. L. L., De Morais, S. M., Dos Santos, H. S., Albuquerque, M. R. J. R., Bandeira, P. N., De Brito, E. H. S., et al. (2016). Antifungal activity and synergistic effect of acetophenones isolated from species Croton against dermatophytes and yeasts. J. Med. Plants Res. 10, 216–222. doi: 10.5897/JMPR2016.6048
De Lacy Costello, B., Amann, A., Al-Kateb, H., Flynn, C., Filipiak, W., Khalid, T., et al. (2014). A review of the volatiles from the healthy human body. J. Breath Res. 8:014001. doi: 10.1088/1752-7155/8/1/014001
De Vrieze, M., Pandey, P., Bucheli, T. D., Varadarajan, A. R., Ahrens, C. H., Weisskopf, L., et al. (2015). Volatile organic compounds from native potato-associated Pseudomonas as potential anti-oomycete agents. Front. Microbiol. 6:1295. doi: 10.3389/fmicb.2015.01295
Diallo, S., Crépin, A., Barbey, C., Orange, N., Burini, J.-F., and Latour, X. (2011). Mechanisms and recent advances in biological control mediated through the potato rhizosphere. FEMS Microbiol. Ecol. 75, 351–364. doi: 10.1111/j.1574-6941.2010.01023.x
Dorn, B., Musa, T., Krebs, H., Fried, P. M., and Forrer, H. R. (2007). Control of late blight in organic potato production: evaluation of copper-free preparations under field, growth chamber and laboratory conditions. Eur. J. Plant Pathol. 119, 217–240. doi: 10.1007/s10658-007-9166-0
Dryahina, K., Sovova, K., Nemec, A., and Spanel, P. (2016). Differentiation of pulmonary bacterial pathogens in cystic fibrosis by volatile metabolites emitted by their in vitro cultures: Pseudomonas aeruginosa, Staphylococcus aureus, Stenotrophomonas maltophilia and the Burkholderia cepacia complex. J. Breath Res. 10:037102. doi: 10.1088/1752-7155/10/3/037102
Dupuis, B., Rolot, J. L., Stilmant, D., Labbe, V., and Laguesse, L. (2007). Evaluation of innovative products to reduce copper applications to control potato late blight in organic production systems. Commun. Agric. Appl. Biol. Sci. 72, 353–359.
Edgar, R., Domrachev, M., and Lash, A. E. (2002). Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 30, 207–210. doi: 10.1093/nar/30.1.207
Effmert, U., Kalderas, J., Warnke, R., and Piechulla, B. (2012). Volatile mediated interactions between bacteria and fungi in the soil. J. Chem. Ecol. 38, 665–703. doi: 10.1007/s10886-012-0135-5
Farag, M. A., Ryu, C. M., Sumner, L. W., and Pare, P. W. (2006). GC-MS SPME profiling of rhizobacterial volatiles reveals prospective inducers of growth promotion and induced systemic resistance in plants. Phytochemistry 67, 2262–2268. doi: 10.1016/j.phytochem.2006.07.021
Fernando, W. G. D., Ramarathnam, R., Krishnamoorthy, A. S., and Savchuk, S. C. (2005). Identification and use of potential bacterial organic antifungal volatiles in biocontrol. Soil Biol. Biochem. 37, 955–964. doi: 10.1016/j.soilbio.2004.10.021
Fialho, M. B., De Moraes, M. H. D., Tremocoldi, A. R., and Pascholati, S. F. (2011a). Potential of antimicrobial volatile organic compounds to control Sclerotinia sclerotiorum in bean seeds. Pesqui. Agropecu. Bras. 46, 137–142. doi: 10.1590/S0100-204X2011000200004
Fialho, M. B., Ferreira, L. F. R., Monteiro, R. T. R., and Pascholati, S. F. (2011b). Antimicrobial volatile organic compounds affect morphogenesis-related enzymes in Guignardia citricarpa, causal agent of citrus black spot. Biocontrol Sci. Technol. 21, 797–807. doi: 10.1080/09583157.2011.580837
Fialho, M. B., Toffano, L., Pedroso, M. P., Augusto, F., and Pascholati, S. F. (2010). Volatile organic compounds produced by Saccharomyces cerevisiae inhibit the in vitro development of Guignardia citricarpa, the causal agent of citrus black spot. World J. Microbiol. Biotechnol. 26, 925–932. doi: 10.1007/s11274-009-0255-4
Fiddaman, P. J., and Rossall, S. (1993). The production of antifungal volatiles by Bacillus-subtilis. J. Appl. Bacteriol. 74, 119–126. doi: 10.1111/j.1365-2672.1993.tb03004.x
Fiddaman, P. J., and Rossall, S. (1994). Effect of substrate on the production of antifungal volatiles from Bacillus subtilis. J. Appl. Bacteriol. 76, 395–405. doi: 10.1111/j.1365-2672.1994.tb01646.x
Fries, N. (1978). Basidiospore germination in some mycorrhiza-Forming hymenomycetes. Trans. Br. Mycol. Soc. 70, 319–324. doi: 10.1016/S0007-1536(78)80128-4
Fry, W. (2008). Phytophthora infestans: the plant (and R gene) destroyer. Mol. Plant Pathol. 9, 385–402. doi: 10.1111/j.1364-3703.2007.00465.x
Garbeva, P., Hordijk, C., Gerards, S., and De Boer, W. (2014). Volatile-mediated interactions between phylogenetically different soil bacteria. Front. Microbiol. 5:289. doi: 10.3389/fmicb.2014.00289
Ghirardi, S., Dessaint, F., Mazurier, S., Corberand, T., Raaijmakers, J. M., Meyer, J. M., et al. (2012). Identification of traits shared by rhizosphere-competent strains of fluorescent Pseudomonads. Microb. Ecol. 64, 725–737. doi: 10.1007/s00248-012-0065-3
Giorgio, A., De Stradis, A., Lo Cantore, P., and Iacobellis, N. S. (2015). Biocide effects of volatile organic compounds produced by potential biocontrol rhizobacteria on Sclerotinia sclerotiorum. Front. Microbiol. 6:1056. doi: 10.3389/fmicb.2015.01056
Gong, A. D., Li, H. P., Shen, L., Zhang, J. B., Wu, A. B., He, W. J., et al. (2015). The Shewanella algae strain YM8 produces volatiles with strong inhibition activity against Aspergillus pathogens and aflatoxins. Front. Microbiol. 6:1091. doi: 10.3389/fmicb.2015.01091
Groenhagen, U., Baumgartner, R., Bailly, A., Gardiner, A., Eberl, L., Schulz, S., et al. (2013). Production of bioactive volatiles by different Burkholderia ambifaria strains. J. Chem. Ecol. 39, 892–906. doi: 10.1007/s10886-013-0315-y
Guyer, A., De Vrieze, M., Bonisch, D., Gloor, R., Musa, T., Bodenhausen, N., et al. (2015). The anti-phytophthora effect of selected potato-associated Pseudomonas strains: from the laboratory to the field. Front. Microbiol. 6:1309 doi: 10.3389/fmicb.2015.01309
Haas, D., and Defago, G. (2005). Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 3, 307–319. doi: 10.1038/nrmicro1129
Han, S. H., Lee, S. J., Moon, J. H., Park, K. H., Yang, K. Y., Cho, B. H., et al. (2006). GacS-dependent production of 2R, 3R-butanediol by Pseudomonas chlororaphis O6 is a major determinant for eliciting systemic resistance against Erwinia carotovora but not against Pseudomonas syringae pv. tabaci in tobacco. Mol. Plant Microbe Interact. 19, 924–930. doi: 10.1094/MPMI-19-0924
Heil, M. (2010). “Within-plant signalling by volatiles triggers systemic defences,” in Plant Communication from an Ecological Perspective. Signaling and Communication in Plants, eds F. Baluška and V. Ninkovic (Berlin: Springer), 99–112. doi: 10.1007/978-3-642-12162-3_7
Herrera Cano, N., Ballari, M. S., Lopez, A. G., and Santiago, A. N. (2015). New synthesis and biological evaluation of benzothiazole derivates as antifungal agents. J. Agric. Food Chem. 63, 3681–3686. doi: 10.1021/acs.jafc.5b00150
Hu, W., Gao, Q., Hamada, M. S., Dawood, D. H., Zheng, J., Chen, Y., et al. (2014). Potential of Pseudomonas chlororaphis subsp. aurantiaca Strain Pcho10 as a biocontrol agent against fusarium graminearum. Phytopathology 104, 1289–1297. doi: 10.1094/PHYTO-02-14-0049-R
Hua, S. S., Beck, J. J., Sarreal, S. B., and Gee, W. (2014). The major volatile compound 2-phenylethanol from the biocontrol yeast, Pichia anomala, inhibits growth and expression of aflatoxin biosynthetic genes of Aspergillus flavus. Mycotoxin Res. 30, 71–78. doi: 10.1007/s12550-014-0189-z
Huang, R., Li, G. Q., Zhang, J., Yang, L., Che, H. J., Jiang, D. H., et al. (2011). Control of post-harvest botrytis fruit rot of strawberry by volatile organic compounds of Candida intermedia. Phytopathology 101, 859–869. doi: 10.1094/PHYTO-09-10-0255
Hunziker, L., Bonisch, D., Groenhagen, U., Bailly, A., Schulz, S., and Weisskopf, L. (2015). Pseudomonas Strains naturally associated with potato plants produce volatiles with high potential for inhibition of Phytophthora infestans. Appl. Environ. Microbiol. 81, 821–830. doi: 10.1128/AEM.02999-14
Huot, B., Yao, J., Montgomery, B. L., and He, S. Y. (2014). Growth-defense tradeoffs in plants: a balancing act to optimize fitness. Mol. Plant 7, 1267–1287. doi: 10.1093/mp/ssu049
Inceoglu, O., Abu Al-Soud, W., Salles, J. F., Semenov, A. V., and Van Elsas, J. D. (2011). Comparative analysis of bacterial communities in a potato field as determined by pyrosequencing. PLoS ONE 6:e23321. doi: 10.1371/journal.pone.0023321
Innerebner, G., Knief, C., and Vorholt, J. A. (2011). Protection of Arabidopsis thaliana against leaf-pathogenic Pseudomonas syringae by Sphingomonas strains in a controlled model system. Appl. Environ. Microbiol. 77, 3202–3210. doi: 10.1128/AEM.00133-11
Kai, M., Effmert, U., Berg, G., and Piechulla, B. (2007). Volatiles of bacterial antagonists inhibit mycelial growth of the plant pathogen Rhizoctonia solani. Arch. Microbiol. 187, 351–360. doi: 10.1007/s00203-006-0199-0
Kai, M., Haustein, M., Molina, F., Petri, A., Scholz, B., and Piechulla, B. (2009). Bacterial volatiles and their action potential. Appl. Microbiol. Biotechnol. 81, 1001–1012. doi: 10.1007/s00253-008-1760-3
Kanchiswamy, C. N., Malnoy, M., and Maffei, M. E. (2015a). Bioprospecting bacterial and fungal volatiles for sustainable agriculture. Trends Plant Sci. 20, 206–211. doi: 10.1016/j.tplants.2015.01.004
Kanchiswamy, C. N., Malnoy, M., and Maffei, M. E. (2015b). Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 6:151. doi: 10.3389/fpls.2015.00151
Kim, Y. C., Leveau, J., Mcspadden Gardener, B. B., Pierson, E. A., Pierson, L. S., and Ryu, C.-M. (2011). The multifactorial basis for plant health promotion by plant-associated bacteria. Appl. Environ. Microbiol. 77, 1548–1555. doi: 10.1128/AEM.01867-10
King, J., Unterkofler, K., Teschl, G., Teschl, S., Koc, H., Hinterhuber, H., et al. (2011). A mathematical model for breath gas analysis of volatile organic compounds with special emphasis on acetone. J. Math. Biol. 63, 959–999. doi: 10.1007/s00285-010-0398-9
Kocic-Tanackov, S., Dimic, G., Levic, J., Tanackov, I., Tepic, A., Vujicic, B., et al. (2012). Effects of onion (Allium cepa L.) and garlic (Allium sativum L.) essential oils on the Aspergillus versicolor growth and sterigmatocystin production. J. Food Sci. 77, M278–M284. doi: 10.1111/j.1750-3841.2012.02662.x
Kumar, M., Tomar, R. S., Lade, H., and Paul, D. (2016). Methylotrophic bacteria in sustainable agriculture. World J. Microbiol. Biotechnol. 32, 1–9. doi: 10.1007/s11274-016-2074-8
Lance, D. R., Leonard, D. S., Mastro, V. C., and Walters, M. L. (2016). Mating disruption as a suppression tactic in programs targeting regulated lepidopteran pests in US. J. Chem. Ecol. 42, 590–605. doi: 10.1007/s10886-016-0732-9
Lanteigne, C., Gadkar, V. J., Wallon, T., Novinscak, A., and Filion, M. (2012). Production of DAPG and HCN by Pseudomonas sp. LBUM300 contributes to the biological control of bacterial canker of tomato. Phytopathology 102, 967–973. doi: 10.1094/PHYTO-11-11-0312
Lemfack, M. C., Nickel, J., Dunkel, M., Preissner, R., and Piechulla, B. (2014). mVOC: a database of microbial volatiles. Nucleic Acids Res. 42, D744–D748. doi: 10.1093/nar/gkt1250
Li, Q. L., Ning, P., Zheng, L., Huang, J. B., Li, G. Q., and Hsiang, T. (2012). Effects of volatile substances of Streptomyces globisporus JK-1 on control of Botrytis cinerea on tomato fruit. Biol. Control 61, 113–120. doi: 10.1016/j.biocontrol.2011.10.014
Liu, W.-W., Mu, W., Zhu, B.-Y., Du, Y.-C., and Liu, F. (2008). Antagonistic activities of volatiles from four strains of Bacillus spp. and Paenibacillus spp. against soil-borne plant pathogens. Agric. Sci. China 7, 1104–1114. doi: 10.1016/S1671-2927(08)60153-4
Loper, J. E., Hassan, K. A., Mavrodi, D. V., Davis, E. W., Lim, C. K., Shaffer, B. T., et al. (2012). Comparative genomics of plant-associated Pseudomonas spp.: insights into diversity and inheritance of traits involved in multitrophic interactions. PLoS Genet. 8:e1002784. doi: 10.1371/journal.pgen.1002784
Lucy, M., Reed, E., and Glick, B. R. (2004). Applications of free living plant growth-promoting rhizobacteria. Antonie Van Leeuwenhoek 86, 1–25. doi: 10.1023/B:ANTO.0000024903.10757.6e
Mahmood, A., Turgay, O. C., Farooq, M., and Hayat, R. (2016). Seed biopriming with plant growth promoting rhizobacteria: a review. FEMS Microbiol. Ecol. 92:fiw112. doi: 10.1093/femsec/fiw112
Minerdi, D., Bossi, S., Gullino, M. L., and Garibaldi, A. (2009). Volatile organic compounds: a potential direct long-distance mechanism for antagonistic action of Fusarium oxysporum strain MSA 35. Environ. Microbiol. 11, 844–854. doi: 10.1111/j.1462-2920.2008.01805.x
Mitchell, A. M., Strobel, G. A., Moore, E., Robison, R., and Sears, J. (2010). Volatile antimicrobials from Muscodor crispans, a novel endophytic fungus. Microbiology 156, 270–277. doi: 10.1099/mic.0.032540-0
Monnet, D., Vidal, D., and Creach, O. (1988). Influence of metabolic and physical factors on production of diacetoxyscirpenol by Fusarium-Sambucinum Fuckel. Appl. Environ. Microbiol. 54, 2167–2169.
Mueller, U. G., and Sachs, J. L. (2015). Engineering microbiomes to improve plant and animal health. Trends Microbiol. 23, 606–617. doi: 10.1016/j.tim.2015.07.009
Naznin, H. A., Kimura, M., Miyazawa, M., and Hyakumachi, M. (2013). Analysis of volatile organic compounds emitted by plant growth-promoting Fungus Phoma sp GS8-3 for growth promotion effects on tobacco. Microbes Environ. 28, 42–49. doi: 10.1264/jsme2.ME12085
Nechwatal, J., and Zellner, M. (2015). Potential suitability of various leaf treatment products as copper substitutes for the control of late blight (Phytophthora infestans) in organic potato farming. Potato Res. 58, 261–276. doi: 10.1007/s11540-015-9302-8
Neerincx, A. H., Geurts, B. P., Habets, M. F., Booij, J. A., Van Loon, J., Jansen, J. J., et al. (2016). Identification of Pseudomonas aeruginosa and Aspergillus fumigatus mono- and co-cultures based on volatile biomarker combinations. J. Breath Res. 10:016002. doi: 10.1088/1752-7155/10/1/016002
Parafati, L., Vitale, A., Restuccia, C., and Cirvilleri, G. (2015). Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 47, 85–92. doi: 10.1016/j.fm.2014.11.013
Penuelas, J., Asensio, D., Tholl, D., Wenke, K., Rosenkranz, M., Piechulla, B., et al. (2014). Biogenic volatile emissions from the soil. Plant Cell Environ. 37, 1866–1891. doi: 10.1111/pce.12340
Pérez-Montaño, F., Alías-Villegas, C., Bellogín, R. A., Del Cerro, P., Espuny, M. R., Jiménez-Guerrero, I., et al. (2014). Plant growth promotion in cereal and leguminous agricultural important plants: from microorganism capacities to crop production. Microbiol. Res. 169, 325–336. doi: 10.1016/j.micres.2013.09.011
Pieterse, C. M., Zamioudis, C., Berendsen, R. L., Weller, D. M., Van Wees, S. C., and Bakker, P. A. (2014). Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 52, 347–375. doi: 10.1146/annurev-phyto-082712-102340
Pohl, C. H., Kock, J. L. F., and Thibane, V. S. (2011). “Antifungal free fatty acids: a review,” in Science against Microbial Pathogens: Communicating Current Research and Technological Advances, ed A. Méndez-Vilas (Badajoz: Formatex), 61–71.
Rast, D., and Stauble, E. J. (1970). On mode of action of isovaleric acid in stimulating germination of Agaricus-bisporus spores. New Phytol. 69, 557–566. doi: 10.1111/j.1469-8137.1970.tb07608.x
Reddy, G. V., and Guerrero, A. (2010). New pheromones and insect control strategies. Vitam. Horm. 83, 493–519. doi: 10.1016/S0083-6729(10)83020-1
Riyaz-Ul-Hassan, S., Strobel, G., Geary, B., and Sears, J. (2013). An endophytic Nodulisporium sp. from central america producing volatile organic compounds with both biological and fuel potential. J Microbiol Biotechnol 23, 29–35. doi: 10.4014/jmb.1208.04062
Rouissi, W., Ugolini, L., Martini, C., Lazzeri, L., and Mari, M. (2013). Control of post-harvest fungal pathogens by antifungal compounds from Penicillium expansum. J. Food Prot. 76, 1879–1886. doi: 10.4315/0362-028X.JFP-13-072
Rudrappa, T., Splaine, R. E., Biedrzycki, M. L., and Bais, H. P. (2008). Cyanogenic Pseudomonads influence multitrophic interactions in the rhizosphere. PLoS ONE 3:e2073. doi: 10.1371/journal.pone.0002073
Ryu, C. M., Farag, M. A., Hu, C. H., Reddy, M. S., Kloepper, J. W., and Pare, P. W. (2004). Bacterial volatiles induce systemic resistance in Arabidopsis. Plant physiol. 134, 1017–1026. doi: 10.1104/pp.103.026583
Ryu, C. M., Farag, M. A., Hu, C. H., Reddy, M. S., Wei, H. X., Pare, P. W., et al. (2003). Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 100, 4927–4932. doi: 10.1073/pnas.0730845100
Sanchez-Fernandez, R. E., Diaz, D., Duarte, G., Lappe-Oliveras, P., Sanchez, S., and Macias-Rubalcava, M. L. (2016). Antifungal volatile organic compounds from the endophyte Nodulisporium sp. strain GS4d2II1a: a qualitative change in the intraspecific and interspecific interactions with Pythium aphanidermatum. Microb. Ecol. 71, 347–364. doi: 10.1007/s00248-015-0679-3
Schenkel, D., Lemfack, M. C., Piechulla, B., and Splivallo, R. (2015). A meta-analysis approach for assessing the diversity and specificity of belowground root and microbial volatiles. Front. Plant Sci. 6:707. doi: 10.3389/fpls.2015.00707
Schlaeppi, K., and Bulgarelli, D. (2015). The plant microbiome at work. Mol. Plant Microbe Interact. 28, 212–217. doi: 10.1094/MPMI-10-14-0334-FI
Schmidt, R., Etalo, D. W., De Jager, V., Gerards, S., Zweers, H., De Boer, W., et al. (2015). Microbial small talk: volatiles in fungal-bacterial interactions. Front. Microbiol. 6:1495. doi: 10.3389/fmicb.2015.01495
Schulz, S., and Dickschat, J. S. (2007). Bacterial volatiles: the smell of small organisms. Nat. Prod. Rep. 24, 814–842. doi: 10.1039/b507392h
Schulz-Bohm, K., Zweers, H., De Boer, W., and Garbeva, P. (2015). A fragrant neighborhood: volatile mediated bacterial interactions in soil. Front. Microbiol. 6:1212. doi: 10.3389/fmicb.2015.01212
Sharifi, R., and Ryu, C. M. (2016). Making healthier or killing enemies? Bacterial volatile-elicited plant immunity plays major role upon protection of Arabidopsis than the direct pathogen inhibition. Commun. Integr. Biol. 9:e1197445. doi: 10.1080/19420889.2016.1197445
Sharma, R., Chauhan, A., and Shirkot, C. K. (2015). Characterization of plant growth promoting Bacillus strains and their potential as crop protectants against Phytophthora capsici in tomato. Biol. Agric. Horticult. 31, 230–244. doi: 10.1080/01448765.2015.1009860
Shatalin, K., Shatalina, E., Mironov, A., and Nudler, E. (2011). H2S: a universal defense against antibiotics in bacteria. Science 334, 986–990. doi: 10.1126/science.1209855
Shestivska, V., Dryahina, K., Nunvar, J., Sovova, K., Elhottova, D., Nemec, A., et al. (2015). Quantitative analysis of volatile metabolites released in vitro by bacteria of the genus Stenotrophomonas for identification of breath biomarkers of respiratory infection in cystic fibrosis. J. Breath Res. 9:027104. doi: 10.1088/1752-7155/9/2/027104
Shestivska, V., Spanel, P., Dryahina, K., Sovova, K., Smith, D., Musilek, M., et al. (2012). Variability in the concentrations of volatile metabolites emitted by genotypically different strains of Pseudomonas aeruginosa. J. Appl. Microbiol. 113, 701–713. doi: 10.1111/j.1365-2672.2012.05370.x
Smith, C. A., Want, E. J., O'maille, G., Abagyan, R., and Siuzdak, G. (2006). XCMS: processing mass spectrometry data for metabolite profiling using Nonlinear peak alignment, matching, and identification. Anal. Chem. 78, 779–787. doi: 10.1021/ac051437y
Soberon, J. R., Lizarraga, E. F., Sgariglia, M. A., Juarez, M. B. C., Sampietro, D. A., Ben Altabef, A., et al. (2015). Antifungal activity of 4-hydroxy-3-(3-methyl-2-butenyl)acetophenone against Candida albicans: evidence for the antifungal mode of action. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 108, 1047–1057. doi: 10.1007/s10482-015-0559-3
Song, G. C., and Ryu, C. M. (2013). Two volatile organic compounds trigger plant self-defense against a bacterial pathogen and a sucking insect in cucumber under open field conditions. Int. J. Mol. Sci. 14, 9803–9819. doi: 10.3390/ijms14059803
Spence, C., Alff, E., Johnson, C., Ramos, C., Donofrio, N., Sundaresan, V., et al. (2014). Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 14:130. doi: 10.1186/1471-2229-14-130
Tahir, H. A. S., Gu, Q., Wu, H., Niu, Y., Huo, R., and Gao, X. (2017). Bacillus volatiles adversely affect the physiology and ultra-structure of Ralstonia solanacearum and induce systemic resistance in tobacco against bacterial wilt. Sci. Rep. 7:40481. doi: 10.1038/srep40481
Tenorio-Salgado, S., Tinoco, R., Vazquez-Duhalt, R., Caballero-Mellado, J., and Perez-Rueda, E. (2013). Identification of volatile compounds produced by the bacterium Burkholderia tropica that inhibit the growth of fungal pathogens. Bioengineered 4, 236–243. doi: 10.4161/bioe.23808
Thorn, R. M., Reynolds, D. M., and Greenman, J. (2011). Multivariate analysis of bacterial volatile compound profiles for discrimination between selected species and strains in vitro. J. Microbiol. Methods 84, 258–264. doi: 10.1016/j.mimet.2010.12.001
Trivedi, P., Pandey, A., and Palni, L. M. S. (2008). In vitro evaluation of antagonistic properties of Pseudomonas corrugato. Microbiol. Res. 163, 329–336. doi: 10.1016/j.micres.2006.06.007
Tyc, O., Zweers, H., De Boer, W., and Garbeva, P. (2015). Volatiles in inter-specific bacterial interactions. Front. Microbiol. 6:1412. doi: 10.3389/fmicb.2015.01412
Van Hulten, M., Pelser, M., Van Loon, L. C., Pieterse, C. M., and Ton, J. (2006). Costs and benefits of priming for defense in Arabidopsis. Proc. Natl. Acad. Sci. USA. 103, 5602–5607. doi: 10.1073/pnas.0510213103
Velivelli, S. L., De Vos, P., Kromann, P., Declerck, S., and Prestwich, B. D. (2014). Biological control agents: from field to market, problems, and challenges. Trends Biotechnol. 32, 493–496. doi: 10.1016/j.tibtech.2014.07.002
Vespermann, A., Kai, M., and Piechulla, B. (2007). Rhizobacterial volatiles affect the growth of fungi and Arabidopsis thaliana. Appl. Environ. Microbiol. 73, 5639–5641. doi: 10.1128/AEM.01078-07
Voisard, C., Keel, C., Haas, D., and Defago, G. (1989). Cyanide Production by Pseudomonas-Fluorescens helps suppress black root-rot of tobacco under gnotobiotic conditions. Embo J. 8, 351–358.
Vorholt, J. A. (2012). Microbial life in the phyllosphere. Nat. Rev. Microbiol. 10, 828–840. doi: 10.1038/nrmicro2910
Wang, Z. F., Wang, C. L., Li, F. J., Li, Z. J., Chen, M. H., Wang, Y. R., et al. (2013). Fumigant activity of volatiles from Streptomyces alboflavus TD-1 against Fusarium moniliforme Sheldon. J. Microbiol. 51, 477–483. doi: 10.1007/s12275-013-2586-y
Weise, T., Kai, M., and Piechulla, B. (2013). Bacterial ammonia causes significant plant growth inhibition. PLoS ONE 8:e63538. doi: 10.1371/journal.pone.0063538
Werner, S., Polle, A., and Brinkmann, N. (2016). Belowground communication: impacts of volatile organic compounds (VOCs) from soil fungi on other soil-inhabiting organisms. Appl. Microbiol. Biotechnol. 100, 8651–8665. doi: 10.1007/s00253-016-7792-1
Xia, J., Sinelnikov, I. V., Han, B., and Wishart, D. S. (2015). MetaboAnalyst 3.0–making metabolomics more meaningful. Nucleic Acids Res. 43, W251–W257. doi: 10.1093/nar/gkv380
Yang, J., Kloepper, J. W., and Ryu, C. M. (2009). Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 14, 1–4. doi: 10.1016/j.tplants.2008.10.004
Yuan, J., Raza, W., Shen, Q. R., and Huang, Q. W. (2012). Antifungal Activity of Bacillus amyloliquefaciens NJN-6 volatile compounds against Fusarium oxysporum f. sp cubense. Appl. Environ. Microbiol. 78, 5942–5944. doi: 10.1128/AEM.01357-12
Zahir, Z. A., Arshad, M., and Frankenberger, W. T. Jr. (2003). Plant growth promoting rhizobacteria: applications and perspectives in agriculture. Adv. Agron. 81, 97–168. doi: 10.1016/S0065-2113(03)81003-9
Zhang, X. Y., Li, B. Q., Wang, Y., Guo, Q. G., Lu, X. Y., Li, S. Z., et al. (2013). Lipopeptides, a novel protein, and volatile compounds contribute to the antifungal activity of the biocontrol agent Bacillus atrophaeus CAB-1. Appl. Microbiol. Biotechnol. 97, 9525–9534. doi: 10.1007/s00253-013-5198-x
Zhao, L. J., Yang, X. N., Li, X. Y., Mu, W., and Liu, F. (2011). Antifungal, insecticidal and herbicidal properties of volatile components from Paenibacillus polymyxa Strain BMP-11. Agric. Sci. China 10, 728–736. doi: 10.1016/S1671-2927(11)60056-4
Zhao, S., Zhao, L., Zhang, X., Liu, C., Hao, C., Xie, H., et al. (2016). Design, synthesis, and structure-activity relationship studies of benzothiazole derivatives as antifungal agents. Eur. J. Med. Chem. 123, 514–522. doi: 10.1016/j.ejmech.2016.07.067
Keywords: volatile organic compounds, Pseudomonas, phytophthora, potato, biocontrol, microbiome
Citation: Bailly A and Weisskopf L (2017) Mining the Volatilomes of Plant-Associated Microbiota for New Biocontrol Solutions. Front. Microbiol. 8:1638. doi: 10.3389/fmicb.2017.01638
Received: 06 October 2016; Accepted: 14 August 2017;
Published: 25 August 2017.
Edited by:
Helene Sanfacon, Agriculture and Agri-Food Canada, CanadaReviewed by:
Marco Kai, University of Rostock, GermanyCopyright © 2017 Bailly and Weisskopf. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: YXVyZWxpZW4uYmFpbGx5QGhvdG1haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.