 Hamza Seghouani
Hamza Seghouani Carlos-Enrique Garcia-Rangel
Carlos-Enrique Garcia-Rangel Jérémie Füller
Jérémie Füller Jeff Gauthier
Jeff Gauthier Nicolas Derome
Nicolas Derome- 1Département de Biologie, Institut de Biologie Intégrative et des Systèmes, Université Laval, Quebec, QC, Canada
- 2Centre Hospitalier de l’Université Laval, Quebec, QC, Canada
Walleye (Sander vitreus) is the second most fished freshwater species in Canada. While much sought by anglers, walleye also supports substantial commercial fisheries. To cope with the recent decline of wild walleye populations, fish farmers produce juveniles for lake stocking. However, walleye breeding is particularly tedious, mostly due to high disease susceptibility at larval and juvenile developmental stages. The main threat is the columnaris disease, which is caused by Flavobacterium columnare, an opportunistic bacteria. As F. columnare strains exhibit increasing antibiotic resistance, there is a strong need to develop efficient and sustainable alternative strategies to control columnaris disease. Bacterial probiotics have been shown to mitigate infections either by enhancing host immune response or by inhibiting pathogen growth. Being successfully assessed in many fish/pathogen combinations, we developed a tailored probiotic strategy for walleye to prevent and treat columnaris disease. Thirty-seven endogenous bacterial strains were isolated from healthy walleye’s skin and gut, were tested in vitro against F. columnare. Significant antagonistic effect against F. columnare was measured for 2 out of 37 endogenous strains. These two probiotic strains were identified as Pseudomonas fluorescens. The antagonistic effect of these two successful probiotics was further validated in vivo during a 2-month stress trial: groups receiving probiotic treatments showed on average 53.74% survival improvement.
Introduction
Walleye (Sander vitreus) is a fecund piscivorous species usually found in moderately productive lakes. Native from North America, its geographical distribution ranges from the east of United States, to the north of St. Lawrence River in eastern Quebec, Canada (Colby et al., 1979; Wilson and Nagler, 2006). Largely known as an effective predator, walleye is one of the most economically important sport and commercial species in Canada (Bernatchez and Giroux, 1991; DFO, 2007, 2012). However, in the last decades, wild walleye populations encountered significant declines, due to overfishing (Sullivan, 2003; Hunt et al., 2011). To cope with the rarefaction of this species, fish farmers started producing juveniles for lake stocking. Noteworthy, walleye breeding is particularly tedious, because optimal rearing conditions are still very challenging, mostly in terms of nutrition (Huntingford, 2004) but also because of high disease susceptibility at early developmental stages (Suomalainen et al., 2005). The most prevalent threat is the columnaris disease, which is mainly caused by Flavobacterium columnare, that naturally inhabits both fish microbiota and in environmental microbial communities.
Flavobacterium columnare is described as one of the most important bacterial diseases of freshwater fish species (Arias et al., 2004), affecting wild and cultured fish [e.g., Arctic charr, Salvelinus alpinus (L.), Perch, Perca sp. (L.), Atlantic salmon, Salmo salar (L.); Austin and Austin, 2007]. For instance, highly virulent strain of F. columnare was able to trigger death within 24 h in coho salmon fry, Oncorhynchus kisutch (Walbaum) (Rucker et al., 1953; see in Austin and Austin, 2007). Several studies have indicated the potential for F. columnare to survive for extended periods of time in water (Kunttu et al., 2009, 2012). Under laboratory conditions, F. columnare maintains its infectious property for more than 5 months (Kunttu et al., 2012). Welker et al. (2005) confirmed that the disease can be transmitted horizontally and indirectly through the water column without essentially being in contact fish-to-fish. When surviving outside the host, F. columnare inhibits virulence gene expression in order to save energy before colonizing another fish host with compromised immune system (Kunttu et al., 2009). Indeed, the occurrence of this opportunistic disease is directly related to stress, elevated temperatures, crowding, etc. (Suomalainen et al., 2005). Columnaris disease symptoms occur internally or externally (gill or skin lesions), and appear as dark-gray or yellow lesions or ulcers (Hartman, 2009). As aquaculture intensifies, overcrowding, low water quality and intensive handling increase physiological stress and physical injury, which in turn favors opportunistic pathogens (Derome et al., 2016a,b). Under such a condition, walleye larvae and juveniles become highly susceptible to columnaris disease, which causes substantial economic loss to fish farmers.
In recent decades, prevention and control of diseases in cultured animals focused research and budgets on antibiotics and chemotherapeutic agents, which are still extensively employed. To date, there are multiple evidences that intensive use antibiotics inevitably leads not only to the emergence of drug-resistant pathogens and other microorganisms, but also to the release of active molecules in the environment, both of which represent a significant risk for public health (Miranda and Zemelman, 2001; Radu et al., 2003; World Health Organization [WHO], 2014). Therefore, there is an urgent need to develop efficient and sustainable methods to control and prevent opportunistic disease such as columnariose, to meet the increasing demand for environment friendly aquaculture. Overall, such alternative methods are expected to warrant a microbiologically healthy environment to enhance fish production and economic profits (Díaz-Rosales et al., 2009). The use of probiotics to increase disease resistance and improving the overall health of terrestrial animals, has long been established as efficient, innocuous, and sustainable (Parker, 1974; Sissons, 1989; Rolfe, 2000; Scharek et al., 2007; Boutin et al., 2013; Foureaux et al., 2014; Hai, 2015). The competition between probiotic bacteria and pathogens was reported in many fish and other aquatic species (Balcázar et al., 2000, 2004, 2007). Probiotic development usually bears on two strategies: allochthonous and autochthonous. The allochthonous strategy aims to test probiotic properties of candidates that were isolated from another host organism, whereas the autochthonous strategy targets the host microbiota to isolate promising probiotic candidates (PC), in order to ensure both efficiency against the pathogen and innocuity for the host. PC isolated from host associated microbial community (i.e., microbiota) have been shown to be efficient in fish and other vertebrates such as Solea, Solea senegalensis (Kaup) (García de La Banda et al., 2010), Brook trout, Salvelinus fontinalis (Mitchill) (Boutin et al., 2012, 2013), Zebrafish, Danio rerio (Hamilton) (Rane and Markad, 2015), and Pigs (Hou et al., 2015). Indeed, the host microbiota, which is composed with numerous microbial strains that closely interact with each other, is a dynamic system that evolves through fish development (reviewed in Llewellyn et al., 2014; Zac Stephens et al., 2015). It is now widely acknowledged that resident bacteria contribute to host disease resistance via two kind of mechanisms: (1) specifically targeting pathogens either by nutritional competition, synthesis of antimicrobial compounds, or competitive exclusion from epithelial surfaces (Bermudez-Brito et al., 2012; Kamada et al., 2013); (2) mechanisms targeting the host immune signaling pathways control (Kamada et al., 2013). Regarding resistance against columnaris and other skin diseases in fish, skin mucus is playing a major role as a physical and chemical barrier (Rottmann et al., 1992). More specifically, skin microbiota associated strains have been reported to protect their host against pathogens by competitive action for adhesion sites (Vine et al., 2004; Chabrillón et al., 2005; Boutin et al., 2013; Ige, 2013), production of organic acids and other antimicrobial compounds such as bacteriocins and siderophores (Yan et al., 2002). Therefore, autochthonous skin bacteria are relevant targets to develop efficient probiotic strains against opportunistic skin disease such as columnaris. Also, as gut microbiota is a reservoir of numerous bacterial symbionts that were proved to be efficient against opportunistic pathogens (reviewed in Gomez et al., 2013), and more specifically against Flavobacterium (Burbank et al., 2012; Ghosh et al., 2016), those bacterial strains were also considered as PC in this work.
The goal of the present study was to develop an autochthonous probiotic strategy against columnaris disease in walleye. To do so, 37 bacterial candidates were isolated from healthy adult walleye skin and gut microbiomes in order to screen in vitro their antagonistic properties vis-à-vis F. columnare. The two candidates that demonstrated highest efficiency against F. columnare were further validated in vivo to assess both their innocuity and ability to decrease mortality rates in walleye during a stress trial.
Materials and Methods
In Vitro Experiments
Walleye Bacteria Sampling
Autochthonous bacteria were isolated from both skin and gut of healthy walleye (S. vitreus) from the Station Aquicole des Trois-Lacs (Asbestos, QC, Canada). Skin mucus samples were recovered by scraping the skin surface between the opercula and the caudal fin with a sterile razor blade. Gut mucus samples were recovered by scraping the intestine epithelial layer with a sterile Q-tip. Then, skin and gut mucus samples were diluted and homogenized 1:9 with sterile phosphate-buffered saline (1×, pH 7.4). Both pathogen and potential PC were grown on the same general growth media, i.e., Anacker and Ordal know as AO (Anacker and Ordal, 1959). The mucus dilutions were spread on fresh growth media (AO) by single-step streaking with a sterile inoculating loop. Agar plates were incubated at 20°C for 48–72 h. Individual colonies were sampled with an inoculating loop and streaked in three steps on the corresponding fresh growth media, and incubated as described above, and then stored at 4°C as solid pre-cultures. Bacterial stock cultures were prepared from pure solid culture by resuspending bacteria in excess in liquid growth medium supplemented with 15% w/v glycerol, and by storing immediately at -80°C.
Screening of Antagonistic Bacteria with Agar Diffusion Assays
The 37 autochthonous PC were screened on the basis of antagonism against F. columnare strain by diffusion of antimicrobial compounds through agar using same growth media (AO). In aseptic conditions, liquid cultures of F. columnare were prepared by resuspending bacteria from a solid culture in liquid AO medium up to an optical density at 600 nm (OD600) of 0.67. Liquid PC cultures were also prepared in a similar manner. Bacterial lawns of PC and F. columnare were prepared by streaking the whole surface of fresh agar media with a sterile cotton swab dipped in liquid bacterial strain culture. Before incubation, sections with a radius of 03 mm was excised from PC solid cultures and laid equidistantly upside down on F. columnare solid cultures. Wells were then incubated at 17°C. Inhibition surfaces around the wells were measured by scanning individually each day over a 9-day time course at a resolution of 23.6 pixels per mm, then the inhibition surfaces around the PC sections were measured using the image processing software ImageJ (NCBI, NIH)1. The radiuses were measured in the same manner using ImageJ. To obtain the final inhibition surface, the PC section area was subtracted from the inhibition area (Dheilly, 2014). Then, the PC showing inhibition radius were selected for in vivo experiment. All manipulations were executed in triplicates (Sharon et al., 2011).
Bacterial Strain Identification
The best two PC were identified to the genus level by sequencing the 16S rRNA gene. After DNA isolation using the Dneasy Blood and Tissue Kit (Qiagen), polymerase chain reaction (PCR) amplification of the 16S rRNA gene was undertaken using the universal set of bacterial primers 331F (5′-TCCTACGGGAGGCAGCAGT-3′) (Nadkarni et al., 2002) and 1389R (5′-AGGCCCGGGAACGTATTCAC-3′) (Woo et al., 2001). PCRs were conducted in volume of 50 μL using a Biometra T1 Thermocycler, using a following amplification conditions: initial denaturation at 94°C for 2 min followed by 30 cycles of 94°C for 30 s, 55°C 1 min, 72°C for 1 min 30 s and a final extension step at 72°C for 10 min. Gel electrophoresis [2% (w/v) agarose, 100 V] was used to visualize the PCR products. Fragments were sequenced using the Big Dye Terminator V3 chemistry on an ABI 3130XL sequencer (Applied Biosystems, Foster City, United States) at the Plate-forme d’Analyses Génomiques (IBIS, Université Laval, Quebec, Canada).
In Vivo Experiment
Walleye juveniles (2 cm, ∼1 g) were obtained from the Station Aquicole des Trois-Lacs (Asbestos, QC, Canada). Upon arrival, fish were acclimated in 1 m3 indoor tanks for 2 months. All fish were held under natural photoperiod conditions, constant temperature of 21°C, and fed daily with commercial fish food (Corey Aquafeeds). After acclimation, a total number of 324 fishes were distributed randomly between six independent recirculating 50 L tanks: each experimental group (PC1, PC2, control) was duplicated. Each tank was independent in terms of filtration and water recirculation using external filter (550 L/h).
It has been clearly demonstrated that in fish farms, F. columnare originates from environmental water, farm environment; then, handling practices are the principal cause triggering disease outbreaks (Pulkkinen et al., 2010; Kunttu et al., 2012). As physiological stress was identified as the most efficient disease triggering factor in intensive aquaculture (Iwama, 2011), our stress protocol aimed to mimic recurrent transfers occurring in farm conditions.
The intensity of thermal stress and mechanical stress were less extreme from previous studies (Nakano et al., 2014; Blanco Garcia et al., 2016). A combination of mechanical and thermal stresses was applied as follow: fish where captured and released into a 20-L bucket where temperature was 6°C below tank’s temperature. After a 10-min exposure to low temperature, fish where put back into their respective tanks. This stress protocol was repeated after each sampling.
Two PC isolated from walleye skin mucus and selected for their in vitro antagonistic activity were selected for this in vivo experiment. Probiotic formulations were administered twice a day (8 am and 8 pm), the mean count of isolate at each administration was 6.5 × 108 colony forming unit (CFU). Moribund fish were collected daily and euthanized by overdose of MS-222 (250 mg/L). Then, dead fish were stored at -80°C for future analysis. All experiments were conducted at the Laboratoire de Recherche en Sciences Aquatiques (LARSA – Université Laval) and carried out in accordance with the LARSA guidelines approved by the “Comité de Protection des Animaux” (CPA).
Detection of F. columnare in Fish Samples by Polymerase Chain Reaction
The detection of F. columnare was performed using the experimental procedure of Michel et al. (2002) with some modifications. Five muscle samples were taken directly from lesion and from different fish. Samples were then placed into microtubes containing 400 μL of distilled water. Using an electric homogenizer (Heidolph DIAX 100, Schwabach, Germany), the slurry was crushed and homogenized then treated in 40 μL of 40 mM Tris–ethylenediaminetetraacetic acid (EDTA) and 10 μL of 1% proteinase K. The mixture was incubated 20 min at 60°C and then 5 min at 100°C. After a 15 s centrifugation at 13,000 g, the supernatant was then stored at 4°C for PCR analysis.
As a negative control, two samples were used: healthy fish muscle and a pure culture of F. psychrophilum mixed with a healthy fish muscle tissue. As positive control, a pure culture of F. columnare mixed with a healthy fish muscle tissue. A DNeasy Blood and Tissue Kit (Qiagen) was used for DNA extraction on series of dilutions.
Polymerase Chain Reaction
Using a species-specific primer for F. columnare Col-72F (5′-GAAGGAGCTTGTTCCTTT-3′) and Col-1260R (5′-GCCTACTTGCGTAGTG-3′) as describe by Triyanto et al. (1999). A PCR reaction was performed in final volume of 50 μL, using 1 μL of Q5® High-Fidelity DNA Polymerase (M0491), 10 μL Q5 Reaction Buffer, 10 μL Q5 High GC Enhancer, 1 μL of dNTPs 10 mM, 2.5 μL of primers 10 mM, and 3 μL of template DNA samples. PCR conditions were applied as follows: samples denaturation 30 s at 98°C, then processed through 35 cycles consisting of 30 s at 98°C, 30 s at 58°C, 30 s at 72°C and 2 min at 72°C for final extension. The final products were visualized in UV light after electrophoresis in 2% agarose gel.
Furthermore, the resulting PCR products were sequenced using an Applied Biosystems ABI 3130XL DNA analyzer at the Plate-forme d’Analyses Génomiques (IBIS, Université Laval, Quebec, Canada).
Statistical Analysis
Survival times were calculated as the time of experiment started, until death. Deaths and mortality were reported daily, and stratified by tanks and treatment. The mortality proportions between treatments during 60 days were compared by chi-square tests. We used Kaplan–Meier methods, log rank test to describe survival curves and Cox’s proportional hazards multivariate regressions were used to calculate the hazard ratios for the effect of treatment on mortality. All statistical analyses were performed using the software Rstudio, version 0.98.1102.
Results
In Vitro Screening against F. columnare
Among the 37 bacterial strains issued from skin mucus and the 12 bacterial strains issued from gut epithelial layer that were initially tested with agar diffusion assays against F. columnare, PC14 and PC23 exhibited a growth circle with an inhibition zone of respectively 5 and 3 mm diameter (Figure 1 and Table 1). The plates were monitored over a 9-day period and scanned at different time (48, 120, and 216 h). The 16S rDNA gene sequence analysis showed that these two successful PC were closely related to Pseudomonas fluorescens, a Gram-negative bacteria, belonging to the Gammaproteobacteria subclass and shared 99% identity between each other. The closest hit in GenBank for CP14 and CP23 was P. fluorescens with 99.25% of average nucleotide identity (A506 complete genome accession number: NC_017911.1).
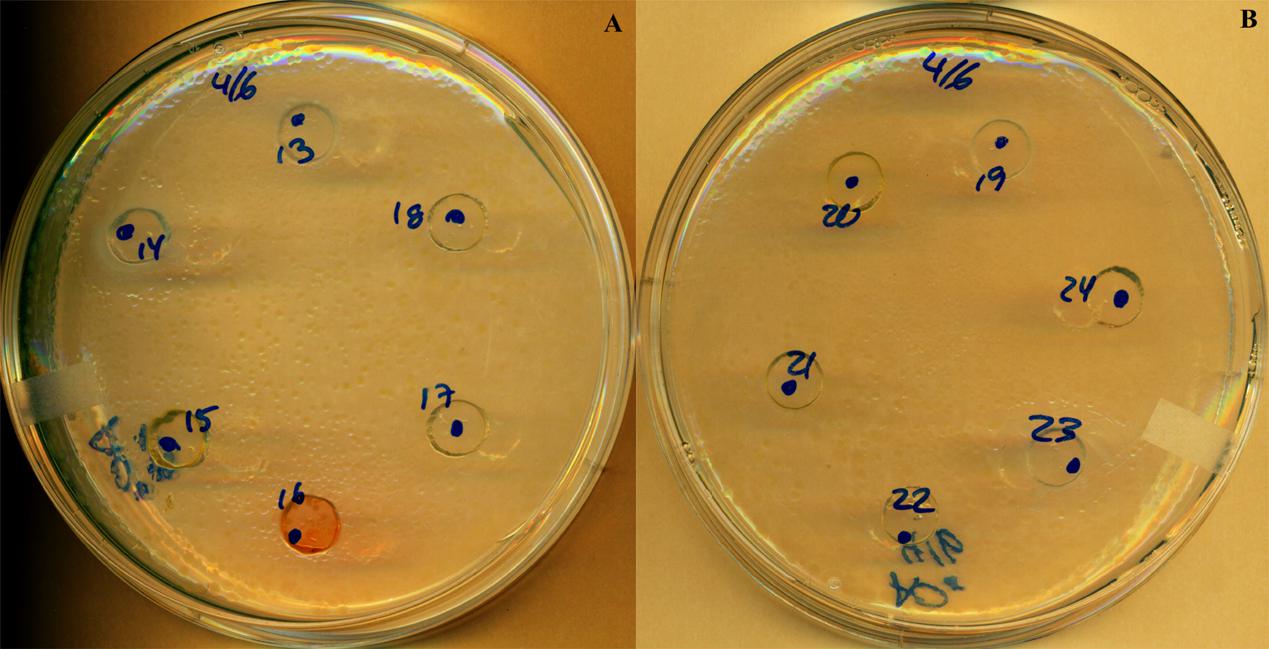
FIGURE 1. Antimicrobial activity of probiotic candidates against F. columnare demonstrated by the inhibition zones produced with the well-diffusion antagonism method at 216 h. PC14 (A), PC23 (B); all other numbers are probiotic candidates with no inhibition effects.

TABLE 1. Autochthonous bacteria from walleye exhibiting diffusible inhibitory effect on agar at 216 h against F. columnare.
Antagonistic Effects against F. columnare in the In Vivo Experiment
Fish mortalities occurred within 24 h following the stress trial. Mortality events increased further until the end of the experiment for the control group. Columnaris disease symptoms were clearly identified on 40% of moribund and dead individuals.
At the end of experiment, mortality rate was defined for each PC. PC14 and PC23 exhibited a mean mortality rate of 6.42 and 10.07%, respectively (Figures 2, 3), which was significantly lower (p < 0.01) for PC14 than what was observed in the control group 13.88%. There were no significant differences among duplicates for both treatment and control groups (p = 0.62, p = 0.81, and p = 0.72, respectively; Figure 4). Thus, the administration of PC14 reduced consistently and significantly the mortality across duplicates.
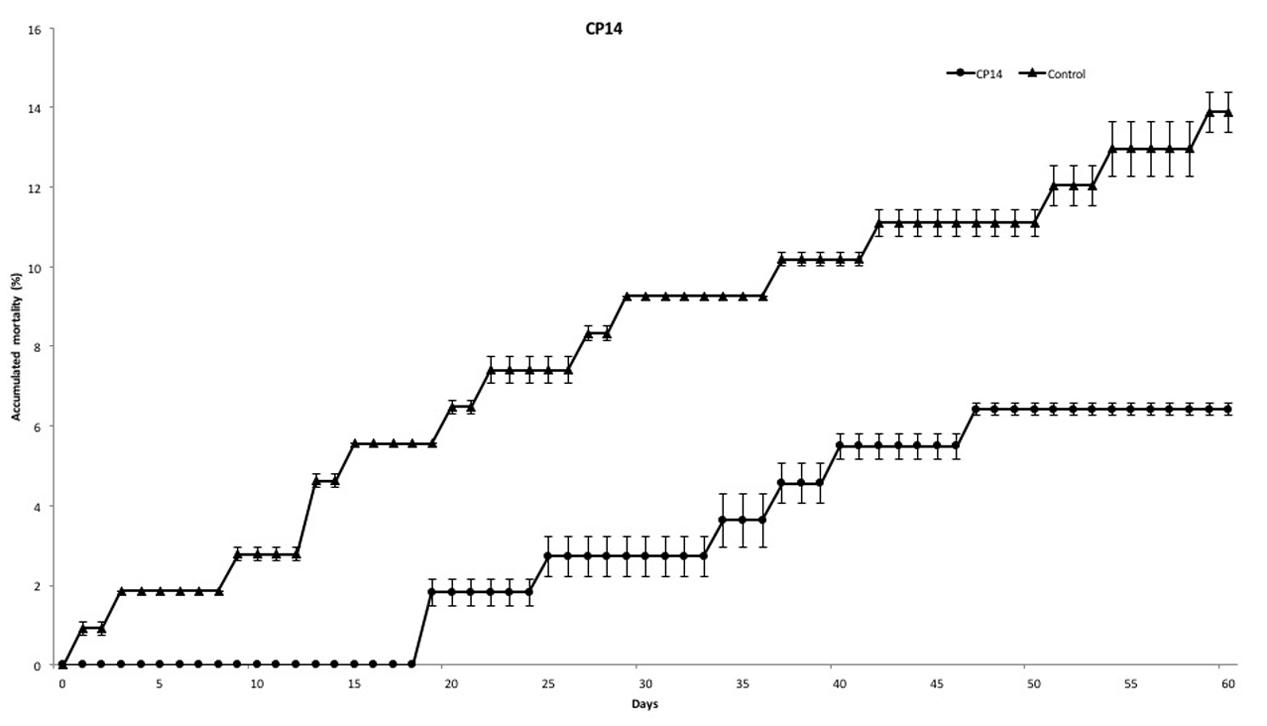
FIGURE 2. Accumulated mortality of walleye infected by F. columnare and treated with CP14. Probiotic culture was added to the tanks before and after stress.
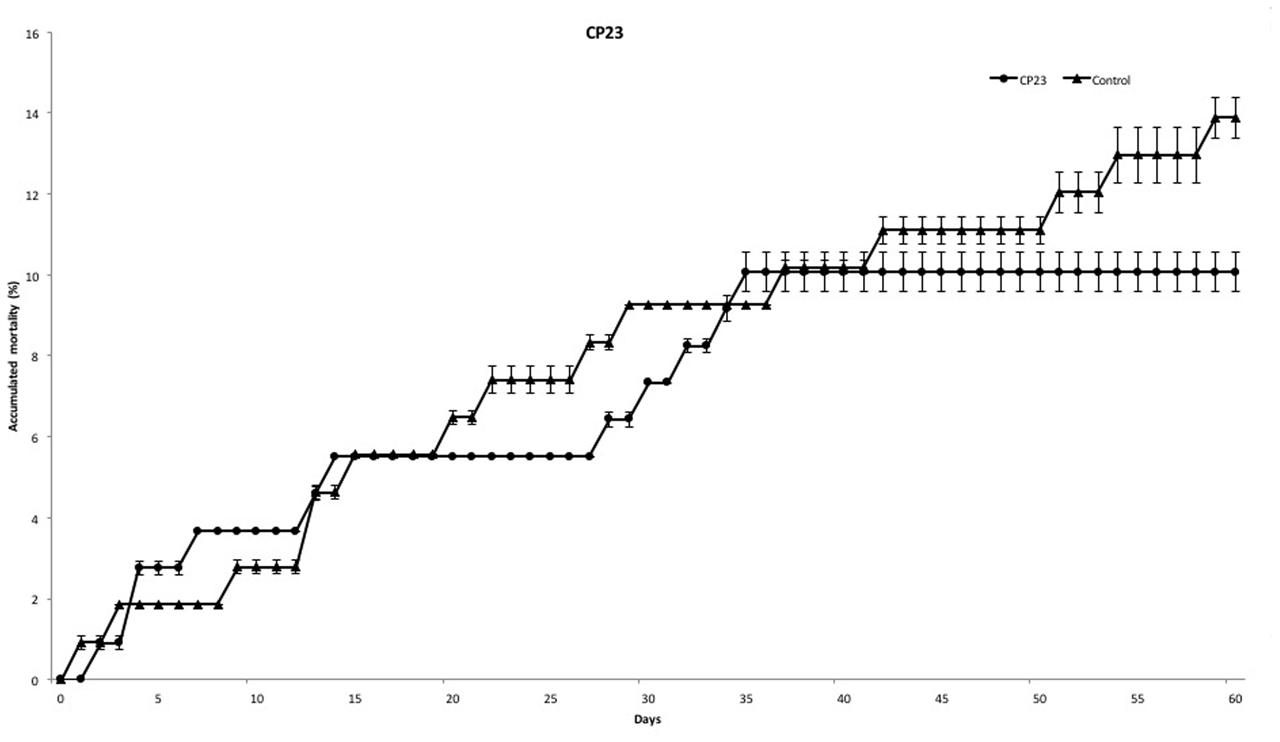
FIGURE 3. Accumulated mortality of walleye infected by F. columnare and treated with CP23. Probiotic culture was added to the tanks before and after stress.
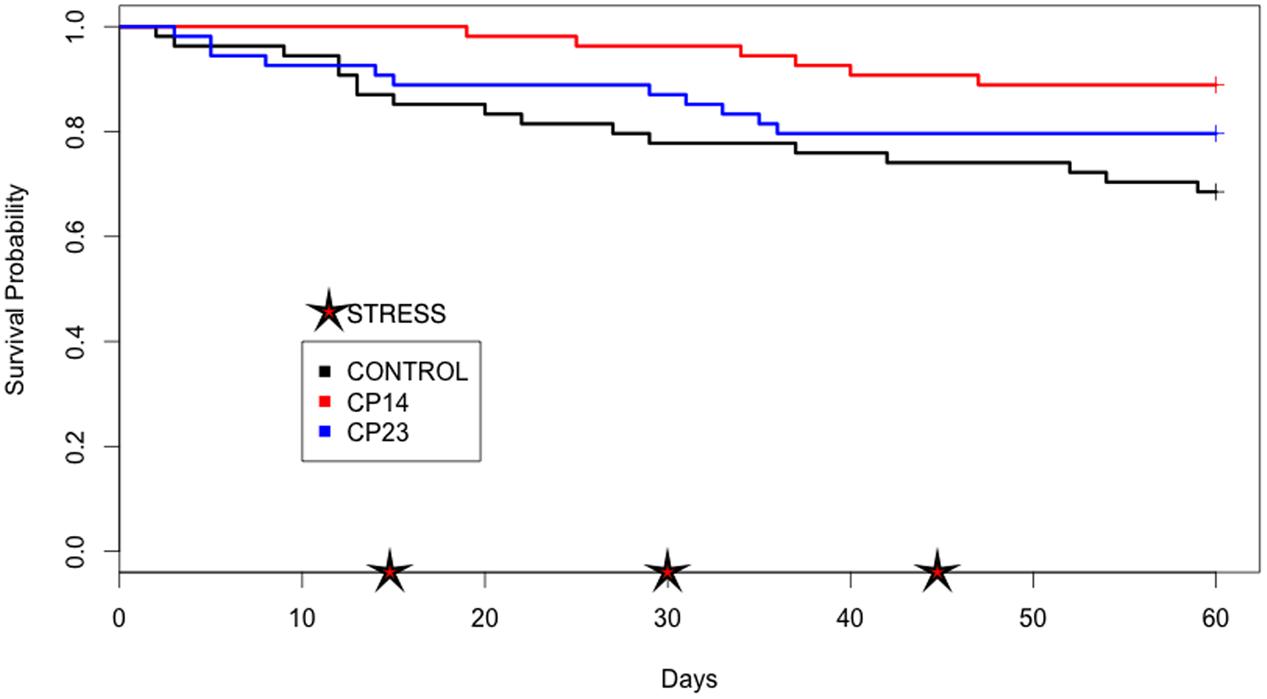
FIGURE 4. Plots of Kaplan–Meier estimate survival of control group and groups receiving probiotic (PC). Survival probability: proportion of fish that survive beyond experiment. Stress: thermal and mechanical manipulation.
PCR Analysis
The resulting PCR from the use of the specific primers Col-72F and Col-1260R was effective for all samples (Table 2). A band of 1200 bp was clearly identified for all samples with columnaris symptoms, whereas absent in all samples without columnaris symptoms. The sequencing of PCR products confirmed the presence of F. columnare in fish samples with columnaris disease symptoms (Table 2).
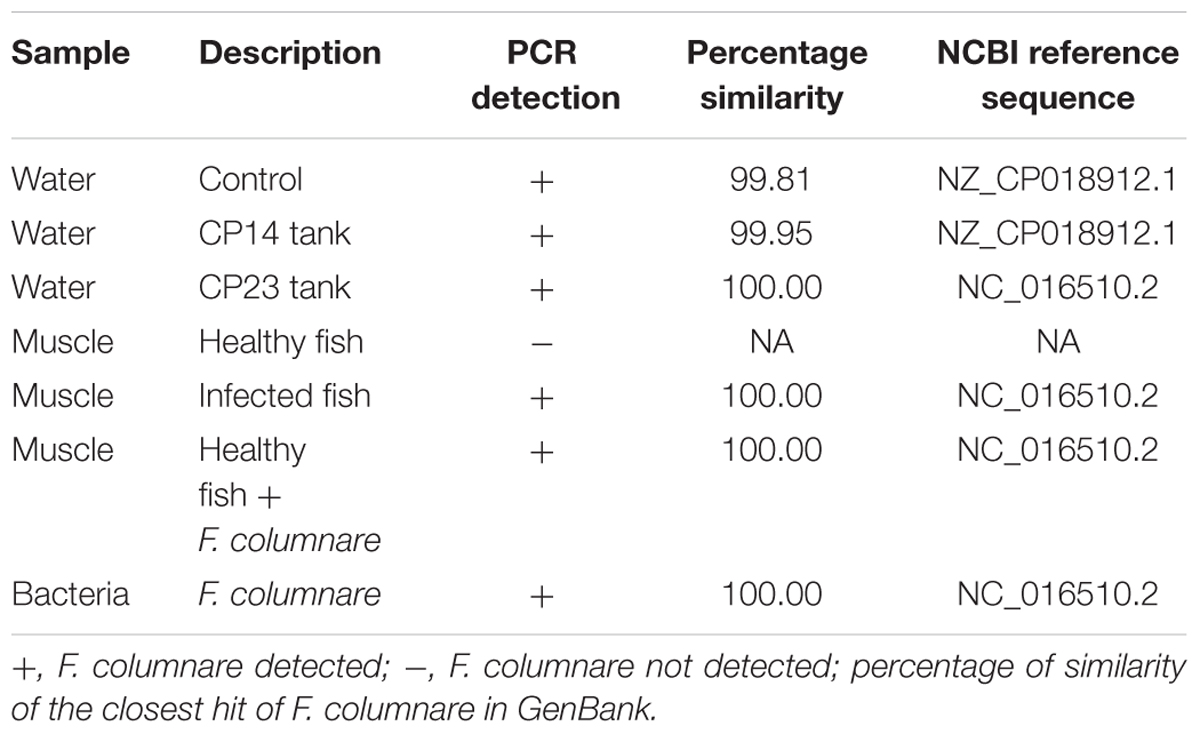
TABLE 2. PCR detection of F. columnare in samples.
Discussion
Fish recruit bacterial strains to build up their microbiota directly from the environmental water microbial community, however, microbiota assemblages are very specific to the corresponding body surface, and highly differentiated from environmental bacterial communities (Apun et al., 1999; Diler et al., 2000; Austin, 2006; Llewellyn et al., 2014, 2015; Zac Stephens et al., 2015; Sylvain et al., 2016). Many studies showed that during their long co-evolution, microbial communities and their hosts have established mutualistic interactions for many physiological aspects, providing major beneficial molecules and services to their host such as enzyme synthesis, vitamins, metabolites, antimicrobial compounds, immune system development, and maturation (Rawls et al., 2004; Salinas et al., 2006; Nayak, 2010; Tremaroli and Bäckhed, 2012).
The present study aimed to take advantage of beneficial host microbiota properties to develop an autochthonous probiotic strategy against columnaris disease in walleye. Among the 49 isolated strains screened in vitro for their potential antagonistic properties vis-à-vis F. columnare, two PC produced clear inhibition circles on F. columnare lawns (PC14 and PC23). Such result suggests their antagonistic effect against F. columnare is likely due, at least in part, to a diffusible antimicrobial compound. These two successful PC were further validated in vivo to test both their innocuity vis-à-vis S. vitreus and their ability to decrease mortality in a stress trial. According to our results, the antagonistic properties of these two PC strains measured in vitro were potentially maintained in vivo by improving significantly fish survival (+53%) under a context of stress trial which, according to both PCR analysis and sequence identification, triggered columnaris disease. However, it is not clear if the same mechanisms of action were involved in both in vitro and in vivo contexts. Overall, the successful administration of these two probiotic strains to walleye is coherent with previous studies that observed both in vitro and in vivo beneficial effects (Gram et al., 2001; Boutin et al., 2012). Gram and Ringø (2005) proposed that an effective probiotic should be identified by its capability to reduce the incidence of disease with a decrease of mortality. More recently, Boutin et al. (2012) confirmed that a positive effect of probiotic is represented by significant decrease of mortality. Still, it is premature to state whether the antibacterial properties observed in vitro are the sole mechanism that favored fish survival.
The two successful PC were closely related to P. fluorescens, belonging to the Gammaproteobacteria subclass. Interestingly, antagonistic properties against pathogenic bacteria and fungi were frequently documented in other aquatic Pseudomonas species and have been suggested to present a high interest as autochthonous PC for aquaculture (Sugita et al., 1996; Gram et al., 2001; Nayak, 2010). Furthermore, some authors concluded that the recurrent presence of Pseudomonas on fish skin represents potentially a promising probiotic strains for fish (Bly et al., 1997; Gram et al., 1999). For instance, Pseudomonas aeruginosa and P. aeruginosa YC58 improved the survival of two varieties of oysters (Pinctada mazatlanica and Crassostrea corteziensis; Aguilar-Macías et al., 2010; Campa-Cordova et al., 2011). Other Pseudomonas were successfully tested against different pathogenic organisms in vitro such as Aeromonas hydrophila (Eissa and El-Ghiet, 2011; Samal et al., 2014) and Vibrio midae (Silva-Aciares et al., 2010). Two studies showed the beneficial effect of P. fluorescens as a promising PC to control pathogens in two distantly related fish species: rainbow trout, Oncorhynchus mykiss (Walbaum) (Gram et al., 1999) and Nile tilapia, Oreochromis niloticus (L.) (Eissa et al., 2014). In our study, annotation of the two PC 16S rDNA partial sequence (<1000 nucleotides) indicated that both of them shared 99% of identity with P. fluorescens A506. This strain is registered as BlightBan® A506 and has been commercialized as a microbial pest control agent against a Erwinia amylovora, a pathogen that affect apples and pear trees (Health Canada Pest Management Regulatory Agency, 2011).
The in vivo probiotic effect of PC from our study was efficient in promoting fish survival in a context of F. columnare disease, which occurred after fish handling and thermal stress. However, regarding the current data, it is not possible to conclude whether the important mortality decrease observed in this experiment was only due to the antibacterial effect attributed to the Pseudomonas strain. Indeed, the Pseudomonas genus encompasses numerous strains, those own diverse mechanisms of action: some are producing bioactive agents such as bacteriocins, pyocin, and phenazinen (Tinh et al., 2007), other strains are triggering bacterial cell membrane lysis, or are producing inhibitors of fatty acid synthesis pathway such as acetyl-CoA, and nitrous oxide (Freiberg et al., 2004; Isnansetyo and Kamei, 2009).
The time scale of a probiotic administration experiment and mode of supplementation are an important criterion affecting the establishment of the probiotic bacteria, their persistence, and even their influence on host immune response. Studies showed that application of probiotic directly to the rearing water play a significant role to health benefits of fish, but also to the rearing environmental (Boyd and Massaut, 1999; Zhou et al., 2010).
The significant improvement of fish survival obtained after 2 months of probiotic administration suggests that autochthonous probiotic strategy is a promising avenue in aquaculture industries. As many studies showed the effectiveness of (allochthonous/autochthonous) probiotics in vivo to decrease mortality and even prevent disease in many species: shrimp, Litopenaeus vannamei (Boone) (Kongnum and Hongpattarakere, 2012), Brook trout (Boutin et al., 2012), and Nile tilapia (Villamil et al., 2012; Eissa et al., 2014). Our work confirms further how efficiently endogenous probiotic can be developed “de novo” to decrease mortality in a context of fish farming industry stressing conditions. Overall, the use of endogenous probiotics in aquaculture provides a straightforward tool to both efficiently and sustainably increase survival rates in aquaculture.
Author Contributions
HS performed in vivo experiment, data analysis, and writing manuscript. C-EG-R performed in vitro and in vivo experiment. JF worked with C-EG-R on in vitro experiment. JG brought fish and helped with in vitro experiment. ND revised the manuscript and supervised the work.
Funding
This research was funded by the CRSNG ENGAGE Grant # 463106-14 to ND.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are thankful to Station Piscicole Trois Lacs Inc., the staff of the “Regional Laboratory of Aquatic Sciences” for their help during fish rearing, and the Animal Protection Committees.
Footnotes
References
Aguilar-Macías, O., Ojeda-Ramírez, J., Campa-Córdova, A., and Saucedo, P. (2010). Evaluation of natural and commercial probiotics for improving growth and survival of the pearl oyster, pinctada mazatlanica, during late hatchery and early field culturing. J. World Aquacult. Soc. 41, 447–454. doi: 10.1111/j.1749-7345.2010.00386.x
Anacker, R. L., and Ordal, E. J. (1959). Studies on the Myxobacterium chondrococcus columnaris: II, Bacteriocins. J. Bacteriol. 78, 33–40.
Apun, K., Yusof, A., and Jugang, K. (1999). Distribution of bacteria in tropical freshwater fish and ponds. Int. J. Environ. Health Res. 9, 285–292. doi: 10.1080/09603129973083
Arias, C., Welker, T., Shoemaker, C., Abernathy, J., and Klesius, P. (2004). Genetic fingerprinting of Flavobacterium columnare isolates from cultured fish. J. Appl. Microbiol. 97, 421–428. doi: 10.1111/j.1365-2672.2004.02314.x
Austin, B. (2006). The bacterial microflora of fish, revised. Sci. World J. 6, 931–945. doi: 10.1100/tsw.2006.181
Austin, B., and Austin, D. A. (2007). Bacterial Fish Pathogens Disease of Farmed and Wild Fish, 4th Edn. London: Praxis Publishing, 552.
Balcázar, J., Vendrell, D., de Blas, I., Ruiz-Zarzuela, I., Gironés, O., and Múzquiz, J. (2007). In vitro competitive adhesion and production of antagonistic compounds by lactic acid bacteria against fish pathogens. Vet. Microbiol. 122, 373–380. doi: 10.1016/j.vetmic.2007.01.023
Balcázar, J. L., De Blas, I., Ruiz-Zarzuela, I., Cunningham, D., Vendrell, D., and Múzquiz, J. L. (2000). The role of probiotics in aquaculture. Vet. Microbiol. 114, 173–186. doi: 10.1016/j.vetmic.2006.01.009
Balcázar, J. L., Vendrell, D., Blas, I. D., Ruiz-Zarzuela, I., and Múzquiz, J. L. (2004). Probiotics: a tool for the future of fish and shellfish health management. J. Aquacult. Trop. 19, 239–242.
Bermudez-Brito, M., Plaza-Diaz, J., Munoz-Quezada, S., Gomez-Llorente, C., and Gil, A. (2012). Probiotic mechanisms of action. Ann. Nutr. Metab. 61, 160–174. doi: 10.1159/000342079
Bernatchez, L., and Giroux, M. (1991). Guide des Poissons D’eau Douce du Queìbec et Leur Distribution Dans l’Est du Canada, 1st Edn. La Prairie, QC: Eìditions Broquet.
Blanco Garcia, A., Jansen, H. M., van Bakhuizen, W., van Houcke, J., and Smaal, A. (2016). “How to measure mussel vitality: evaluation of survival in air and neutral red assay as a stress response indicator for the blue mussel (Mytilus edulis) processing industry,” in Poster Session Presented at European Aquaculture Society, Edinburgh.
Bly, J., Quiniou, S., Lawson, L., and Clem, L. (1997). Inhibition of Saprolegnia pathogenic for fish by Pseudomonas fluorescens. J. Fish Dis. 20, 35–40. doi: 10.1046/j.1365-2761.1997.d01-104.x
Boutin, S., Audet, C., and Derome, N. (2013). Probiotic treatment by indigenous bacteria decreases mortality without disturbing the natural microbiota of Salvelinus fontinalis. Can. J. Microbiol. 59, 662–670. doi: 10.1139/cjm-2013-0443
Boutin, S., Bernatchez, L., Audet, C., and Derôme, N. (2012). Antagonistic effect of indigenous skin bacteria of brook charr (Salvelinus fontinalis) against Flavobacterium columnare and F. psychrophilum. Vet. Microbiol. 155, 355–361. doi: 10.1016/j.vetmic.2011.09.002
Boyd, C. E., and Massaut, L. (1999). Risks associated with the use of chemicals in pond aquaculture. Aquacult. Eng. 20, 113–132. doi: 10.1016/S0144-8609(99)00010-2
Burbank, D., LaPatra, S., Fornshell, G., and Cain, K. (2012). Isolation of bacterial probiotic candidates from the gastrointestinal tract of rainbow trout, Oncorhynchus mykiss (walbaum), and screening for inhibitory activity against Flavobacterium psychrophilum. J. Fish Dis. 35, 809–816. doi: 10.1111/j.1365-2761.2012.01432.x
Campa-Cordova, A. I., Luna-Gonzalez, A., Mazon-Suastegui, J. M., Aguirre-Guzman, G., Ascencio, F., and Gonzalez-Ocampo, H. A. (2011). Effect of probiotic bacteria on survival and growth of Cortez oyster larvae, Crassostrea corteziensis (Bivalvia: Ostreidae). Revista de Biología Tropical 59, 183–191.
Chabrillón, M., Rico, R., Balebona, M., and Morinigo, M. (2005). Adhesion to sole, Solea senegalensis Kaup, mucus of microorganisms isolated from farmed fish, and their interaction with Photobacterium damselae subsp. piscicida. J. Fish Dis. 28, 229–237. doi: 10.1111/j.1365-2761.2005.00623.x
Colby, P. J., McNicol, R. E., and Ryder, R. A. (1979). Synopsis of Biological Data on the Walleye Stizostedion v. vitreum (Mitchill 1918). FAO Fisheries Synopsis No. 119. Rome: Food and Agriculture Organization of the United Nation, 139.
Derome, N., Gauthier, J., Boutin, S., and Llewellyn, M. (2016a). “Bacterial opportunistic pathogens of fish,” in Advances in Environmental Microbiology, The Rasputin Effect: When Commensals and Symbionts Become Parasitic, ed. J. H. Christon (Heidelberg: Springer international), 81–108. doi: 10.1007/978-3-319-28170-4
Derome, N., Gauthier, J., Boutin, S., and Llewellyn, M. (2016b). “Fungal secondary invaders of fish,” in Advances in Environmental Microbiology, The Rasputin Effect: When Commensals and Symbionts Become Parasitic, Chap. 5, ed. J. H. Christon (Heidelberg: Springer international), 109–126. doi: 10.1007/978-3-319-28170-4
DFO (2007). Survey of Recreational Fishing in Canada (2005). Ottawa, ON: Fisheries and Oceans Canada Statistics Division.
DFO (2012). Survey of Recreational Fishing in Canada (2010). Ottawa, ON: Fisheries and Oceans Canada Statistics Division.
Dheilly, N. (2014). Holobiont–holobiont interactions: redefining host–parasite interactions. PLoS Pathog. 10:e1004093. doi: 10.1371/journal.ppat.1004093
Díaz-Rosales, P., Arijo, S., Chabrillón, M., Alarcón, F. J., Tapia-Paniagua, S. T., Martínez-Manzanares, E., et al. (2009). Effects of two closely related probiotics on respiratory burst activity of Senegalese sole (Solea senegalensis, Kaup) phagocytes, and protection against Photobacterium damselae subsp. piscicida. Aquaculture 293, 16–21. doi: 10.1016/j.aquaculture.2009.03.050
Diler,Ö, Altun, S.,Çalikuşu, F., and Diler, A. (2000). A study on qualitative and quantitative bacterial flora of the rainbow trout (Oncorhynchus mykiss) living in different fish farms. Turkish J. Vet. Anim. Sci. 24, 251–260.
Eissa, N., El-Gheit, El, A., and Shaheen, A. A. (2014). Protective effect of Pseudomonas fluorescens as a probiotic in controlling fish pathogens. Am. J. Biosci. 2, 175–181. doi: 10.11648/j.ajbio.20140205.12
Eissa, N., and El-Ghiet, E. A. (2011). Efficacy of Pseudomonas fluorescens as biological control agents against Aeromonas hydrophila infection in Oreochromis niloticus. World J. Fish Mar. Sci. 3, 564–569. doi: 10.11648/j.ajbio.20140205.12
Foureaux, R., Messora, M., de Oliveira, L., Napimoga, M., Pereira, A., Ferreira, M., et al. (2014). Effects of probiotic therapy on metabolic and inflammatory parameters of rats with ligature-induced periodontitis associated with restraint stress. J. Periodontol. 85, 975–983. doi: 10.1902/jop.2013.130356
Freiberg, C., Brunner, N., Schiffer, G., Lampe, T., Pohlmann, J., Brands, M., et al. (2004). Identification and characterization of the first class of potent bacterial Acetyl-CoA carboxylase inhibitors with antibacterial activity. J. Biol. Chem. 279, 26066–26073. doi: 10.1074/jbc.m402989200
García de La Banda, I., Lobo, C., León-Rubio, J., Tapia-Paniagua, S., Balebona, M., Moriñigo, M., et al. (2010). Influence of two closely related probiotics on juvenile Senegalese sole (Solea senegalensis, Kaup 1858) performance and protection against Photobacterium damselae subsp. piscicida. Aquaculture 306, 281–288. doi: 10.1016/j.aquaculture.2010.05.008
Ghosh, B., Cain, K. D., Nowak, B. F., and Bridle, A. R. (2016). Microencapsulation of a putative probiotic Enterobacter species, C6-6, to protect rainbow trout, Oncorhynchus mykiss (Walbaum), against bacterial coldwater disease. J. Fish Dis. 39, 1–11. doi: 10.1111/jfd.12311
Gomez, D., Sunyer, J., and Salinas, I. (2013). The mucosal immune system of fish: the evolution of tolerating commensals while fighting pathogens. Fish Shellf. Immunol. 35, 1729–1739. doi: 10.1016/j.fsi.2013.09.032
Gram, L., Løvold, T., Nielsen, J., Melchiorsen, J., and Spanggaard, B. (2001). In vitro antagonism of the probiont Pseudomonas fluorescens strain AH2 against Aeromonas salmonicida does not confer protection of salmon against furunculosis. Aquaculture 199, 1–11. doi: 10.1016/s0044-8486(01)00565-8
Gram, L., Melchiorsen, J., Spanggaard, B., Huber, I., and Nielsen, T. F. (1999). Inhibition of Vibrio anguillarum by Pseudomonas fluorescens AH2, a possible probiotic treatment of fish. Appl. Environ. Microbiol. 65, 969–973.
Gram, L., and Ringø, E. (2005). “Prospects of fish probiotics,” in Microbial Ecology of the Growing Animal, eds W. H. Holzapfel and P. J. Naughton (Amsterdam: Elsevier), 379–417. doi: 10.1016/S1877-1823(09)70050-5
Hai, N. V. (2015). The use of probiotics in aquaculture. J. Appl. Microbiol. 4, 917–935. doi: 10.1111/jam.12886
Hartman, G. F. (2009). A biological synopsis of walleye (Sander vitreus). Can. Manuscript Rep. Fish. Aquatic Sci. 2888:48.
Health Canada Pest Management Regulatory Agency (2011). ARCHIVED - Pseudomonas fluorescens Strain A506 - Proposed Registration Decision PRD2011-18 - Health Canada Consultation Document. HC-SC.GC.CA. Available at: http://www.hc-sc.gc.ca/-spc/pest/part/consultations/_prd2011-18/prd2011-18-eng.php [accessed March 14, 2016].
Hou, C., Zeng, X., Yang, F., Liu, H., and Qiao, S. (2015). Study and use of the probiotic Lactobacillus reuteri in pigs: a review. J. Anim. Sci. Biotechnol. 6:14. doi: 10.1186/s40104-015-0014-3
Hunt, L., Arlinghaus, R., Lester, N., and Kushneriuk, R. (2011). The effects of regional angling effort, angler behavior, and harvesting efficiency on landscape patterns of overfishing. Ecol. Appl. 21, 2555–2575. doi: 10.1890/10-1237.1
Huntingford, F. (2004). Implications of domestication and rearing conditions for the behaviour of cultivated fishes. J. Fish Biol. 65, 122–142. doi: 10.1111/j.0022-1112.2004.00562.x
Ige, B. (2013). Probiotics use in intensive fish farming. Afr. J. Microbiol. Res. 7, 2701–2711. doi: 10.5897/ajmr12x.021
Isnansetyo, A., and Kamei, Y. (2009). Bioactive substances produced by marine isolates of Pseudomonas. J. Ind. Microbiol. Biotechnol. 36, 1239–1248. doi: 10.1007/s10295-009-0611-2
Iwama, G. (2011). Fish Stress and Health in Aquaculture, 1st Edn. Cambridge: Cambridge University Press.
Kamada, N., Chen, G., Inohara, N., and Núñez, G. (2013). Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 14, 685–690. doi: 10.1038/ni.2608
Kongnum, K., and Hongpattarakere, T. (2012). Effect of Lactobacillus plantarum isolated from digestive tract of wild shrimp on growth and survival of white shrimp (Litopenaeus vannamei) challenged with Vibrio harveyi. Fish Shellf. Immunol. 32, 170–177. doi: 10.1016/j.fsi.2011.11.008
Kunttu, H. M., Sundberg, L. R., Pulkkinen, K., and Valtonen, E. T. (2012). Environment may be the source of Flavobacterium columnare outbreaks at fish farms. Environ. Microbiol. Rep. 4, 398–402. doi: 10.1111/j.1758-2229.2012.00342.x
Kunttu, H. M., Valtonen, E. T., Jokinen, E. I., and Suomalainen, L. R. (2009). Saprophytism of a fish pathogen as a transmission strategy. Epidemics 1, 96–100. doi: 10.1016/j.epidem.2009.04.003
Llewellyn, M., Boutin, S., Hoseinifar, S., and Derome, N. (2014). Teleost microbiomes: the state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 5:207. doi: 10.3389/fmicb.2014.00207
Llewellyn, M., McGinnity, P., Dionne, M., Letourneau, J., Thonier, F., Carvalho, G., et al. (2015). The biogeography of the atlantic salmon (Salmo salar) gut microbiome. ISME J. 10, 1280–1284. doi: 10.1038/ismej.2015.189
Michel, C., Messiaen, S., and Bernardet, J. (2002). Muscle infections in imported neon tetra, Paracheirodon innesi Myers: limited occurrence of microsporidia and predominance of severe forms of columnaris disease caused by an Asian genomovar of Flavobacterium columnare. J. Fish Dis. 25, 253–263. doi: 10.1046/j.1365-2761.2002.00364.x
Miranda, C. D., and Zemelman, R. (2001). Antibiotic resistant bacteria in fish from the concepcion bay, Chile. Mar. Pollut. Bull. 42, 1096–1102. doi: 10.1016/S0025-326X(01)00093-5
Nadkarni, M., Hunter, N., Jacques, N., and Martin, F. (2002). Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology 148, 257–266. doi: 10.1099/00221287-148-1-257
Nakano, T., Kameda, M., Shoji, Y., Hayashi, S., Yamaguchi, T., and Sato, M. (2014). Effect of severe environmental thermal stress on redox state in salmon. Redox Biol. 2, 772–776. doi: 10.1016/j.redox.2014.05.007
Nayak, S. (2010). Probiotics and immunity: a fish perspective. Fish Shellf. Immunol. 29, 2–14. doi: 10.1016/j.fsi.2010.02.017
Pulkkinen, K., Suomalainen, L. R., Read, A. F., Ebert, D., Rintamäki, P., and Valtonen, E. T. (2010). Intensive fish farming and the evolution of pathogen virulence: the case of columnaris disease in Finland. Proc. R. Soc. Lond. B Biol. Sci. 277, 593–600. doi: 10.1098/rspb.2009.1659
Radu, S., Ahmad, N., Ling, F. H., and Reezal, A. (2003). Prevalence and resistance to antibiotics for Aeromonas species from retail fish in Malaysia. Int. J. Food Microbiol. 81, 261–266. doi: 10.1016/S0168-1605(02)00228-3
Rane, M., and Markad, A. (2015). Effects of probiotic on the growth and survival of zebra fish (Danio rerio). Int. J. Sci. Res. 4, 1839–1841. doi: 10.1071/RD12187
Rawls, J., Samuel, B., and Gordon, J. (2004). From the cover: gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proc. Natl. Acad. Sci. U.S.A. 101, 4596–4601. doi: 10.1073/pnas.0400706101
Rolfe, R. D. (2000). The role of probiotic cultures in the control of gastrointestinal health. J. Nutr. 130, 396S–402S.
Rucker, R. R., Earp, B. J., and Ordal, E. J. (1953). Infectious diseases of Pacific salmon. Trans. Am. Fish. Soc. 83, 297–312. doi: 10.1577/1548-8659(1953)83[297:IDOPS]2.0.CO;2
Rottmann, R. W., Francis-Floyd, R., and Durborow, R. (1992). The Role of Stress in Fish Disease. Stoneville, MS: Southern Regional Aquaculture Center, 474.
Salinas, I., Díaz-Rosales, P., Cuesta, A., Meseguer, J., Chabrillón, M., Moriñigo, M., et al. (2006). Effect of heat-inactivated fish and non-fish derived probiotics on the innate immune parameters of a teleost fish (Sparus aurata L.). Vet. Immunol. Immunopathol. 111, 279–286. doi: 10.1016/j.vetimm.2006.01.020
Samal, S. K., Das, B. K., and Pal, B. B. (2014). Isolation, biochemical characterization, antibiotic susceptibility study of Aeromonas hydrophila isolated from freshwater fish. Int. J. Curr. Microbiol. Appl. Sci. 3, 259–267.
Scharek, L., Guth, J., Filter, M., and Schmidt, M. (2007). Impact of the probiotic bacteria Enterococcus faecium NCIMB 10415 (SF68) and Bacillus cereus var. toyoi NCIMB 40112 on the development of serum IgG and faecal IgA of sows and their piglets. Arch. Anim. Nutr. 61, 223–234. doi: 10.1080/17450390701431540
Sharon, G., Segal, D., Zilber-Rosenberg, I., and Rosenberg, E. (2011). Symbiotic bacteria are responsible for diet-induced mating preference in Drosophila melanogaster, providing support for the hologenome concept of evolution. Gut Microbes 2, 190–192. doi: 10.4161/gmic.2.3.16103
Silva-Aciares, F., Carvajal, P., Mejías, C., and Riquelme, C. (2010). Use of macroalgae supplemented with probiotics in the Haliotis rufescens (Swainson, 1822) culture in Northern Chile. Aquacult. Res. 42, 953–961. doi: 10.1111/j.1365-2109.2010.02678.x
Sissons, J. (1989). Potential of probiotic organisms to prevent diarrhoea and promote digestion in farm animals – a review. J. Sci. of Food Agric. 49, 1–13. doi: 10.1002/jsfa.2740490102
Sugita, H., Shibuya, K., Shimooka, H., and Deguchi, Y. (1996). Antibacterial abilities of intestinal bacteria in freshwater cultured fish. Aquaculture 145, 195–203. doi: 10.1016/s0044-8486(96)01319-1
Sullivan, M. (2003). Active management of walleye fisheries in alberta: dilemmas of managing recovering fisheries. N. Am. J. Fish. Manag. 23, 1343–1358. doi: 10.1577/m01-232am
Suomalainen, L., Tiirola, M., and Valtonen, E. (2005). Influence of rearing conditions on Flavobacterium columnare infection of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 28, 271–277. doi: 10.1111/j.1365-2761.2005.00631.x
Sylvain, F., Cheaib, B., Llewellyn, M., Gabriel Correia, T., Barros Fagundes, D., Luis Val, A., et al. (2016). pH drop impacts differentially skin and gut microbiota of the Amazonian fish tambaqui (Colossoma macropomum). Sci. Rep. 6:32032. doi: 10.1038/srep32032
Tinh, N., Dierckens, K., Sorgeloos, P., and Bossier, P. (2007). A review of the functionality of probiotics in the larviculture food chain. Mar. Biotechnol. 10, 1–12. doi: 10.1007/s10126-007-9054-9
Tremaroli, V., and Bäckhed, F. (2012). Functional interactions between the gut microbiota and host metabolism. Nature 489, 242–249. doi: 10.1038/nature11552
Triyanto, Kumamaru, A., and Wakabayashi, H. (1999). The use of PCR Targeted 16S rDNA for Identification of genomovars of Flavobacterium columnare. Fish Pathol. 34, 217–218. doi: 10.3147/jsfp.34.217
Villamil, L., Reyes, C., and Martínez-Silva, M. (2012). In vivo and in vitro assessment of Lactobacillus acidophilus as probiotic for tilapia (Oreochromis niloticus, Perciformes:Cichlidae) culture improvement. Aquacult. Res. 45, 1116–1125. doi: 10.1111/are.12051
Vine, N., Leukes, W., Kaiser, H., Daya, S., Baxter, J., and Hecht, T. (2004). Competition for attachment of aquaculture candidate probiotic and pathogenic bacteria on fish intestinal mucus. J. Fish Dis. 27, 319–326. doi: 10.1111/j.1365-2761.2004.00542.x
Welker, T. L., Shoemaker, C. A., Arias, C. R., and Klesius, P. H. (2005). Transmission and detection of Flavobacterium columnare in channel catfish Ictalurus punctatus. Dis. Aquat. Organ. 63, 129–138. doi: 10.3354/dao063129
Wilson, S., and Nagler, J. (2006). Age, but not salinity, affects the upper lethal temperature limits for juvenile walleye (Sander vitreus). Aquaculture 257, 187–193. doi: 10.1016/j.aquaculture.2005.10.045
Woo, P., Cheung, E., Leung, K., and Yuen, K. (2001). Identification by 16S ribosomal RNA gene sequencing of an Enterobacteriaceae species with ambiguous biochemical profile from a renal transplant recipient. Diagn. Microbiol. Infect. Dis. 39, 85–93. doi: 10.1016/s0732-8893(01)00206-1
World Health Organization [WHO] (2014). Antimicrobial Resistance. Fact Sheet N°194. Available at: http://www.who.int/mediacentre/factsheets/fs194/en/ [accessed July 15, 2016].
Yan, L., Boyd, K., and Grant Burgess, J. (2002). Surface attachment induced production of antimicrobial compounds by marine epiphytic bacteria using modified roller bottle cultivation. Mar. Biotechnol. 4, 356–366. doi: 10.1007/s10126-002-0041-x
Zac Stephens, W., Burns, A., Stagaman, K., Wong, S., Rawls, J., Guillemin, K., et al. (2015). The composition of the zebrafish intestinal microbial community varies across development. ISME J. 10, 644–654. doi: 10.1038/ismej.2015.140
Keywords: probiotics, Sander vitreus, Flavobacterium columnare, walleye diseases, autochthonous bacteria
Citation: Seghouani H, Garcia-Rangel C-E, Füller J, Gauthier J and Derome N (2017) Walleye Autochthonous Bacteria as Promising Probiotic Candidates against Flavobacterium columnare. Front. Microbiol. 8:1349. doi: 10.3389/fmicb.2017.01349
Received: 24 April 2017; Accepted: 03 July 2017;
Published: 18 July 2017.
Edited by:
Raquel Peixoto, Federal University of Rio de Janeiro, BrazilReviewed by:
Harold J. Schreier, University of Maryland, Baltimore County, United StatesJuan Jose Alava, University of British Columbia, Canada
Copyright © 2017 Seghouani, Garcia-Rangel, Füller, Gauthier and Derome. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nicolas Derome, bmljb2xhcy5kZXJvbWVAYmlvLnVsYXZhbC5jYQ==