
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 11 July 2017
Sec. Virology
Volume 8 - 2017 | https://doi.org/10.3389/fmicb.2017.01314
 Ángela Vázquez-Calvo1
Ángela Vázquez-Calvo1 Nereida Jiménez de Oya1
Nereida Jiménez de Oya1 Miguel A. Martín-Acebes1
Miguel A. Martín-Acebes1 Emilia Garcia-Moruno2
Emilia Garcia-Moruno2 Juan-Carlos Saiz1*
Juan-Carlos Saiz1*The Flavivirus genus contains important pathogens, such as West Nile virus (WNV), Zika virus (ZIKV), and Dengue virus (DENV), which are enveloped plus-strand RNA viruses transmitted by mosquitoes and constitute a worrisome threat to global human and animal health. Currently no licensed drugs against them are available, being, thus, still necessary the search for effective antiviral molecules. In this line, a novel antiviral approach (economical, simple to use, and environmental friendly) is the use of natural compounds. Consequently, we have tested the antiviral potential of different polyphenols present in plants and natural products, such as wine and tea, against WNV, ZIKV, and DENV. So that, we assayed the effect of a panel of structurally related polyphenols [delphinidin (D), cyanidin (Cy), catechin (C), epicatechin (EC), epigallocatechin (EGC), and epigallocatechin gallate (EGCG)] on WNV infection, and found that D and EGCG inhibited more effectively the virus production. Further analysis with both compounds indicated that they mainly affected the attachment and entry steps of the virus life cycle. Moreover, D and EGCG showed a direct effect on WNV particles exerting a virucidal effect. We showed a similar inhibition of viral production of these compounds on WNV variants that differed on acidic pH requirements for viral fusion, indicating that their antiviral activity against WNV is produced by a virucidal effect rather than by an inhibition of pH-dependent viral fusion. Both polyphenols also reduced the infectivity of ZIKV and DENV. Therefore, D and EGCG impair the infectivity in cell culture of these three medically relevant flaviviruses.
The Flavivirus genus contains important pathogens such as Zika virus (ZIKV), Dengue virus (DENV), yellow fever virus (YFV), Japanese encephalitis virus (JEV), and West Nile virus (WNV), among others. As flaviviruses, WNV, ZIKV, and DENV are enveloped plus-strand RNA viruses transmitted by mosquitoes and constitute a worrisome threat to global human and animal health (Martin-Acebes and Saiz, 2012; Saiz et al., 2016; Pang et al., 2017). WNV is maintained in an enzootic transmission cycle between mosquitoes and birds, but it can also infect other vertebrates, such as horses and humans (Martin-Acebes and Saiz, 2012). Whereas most WNV infections in humans are asymptomatic, ∼20% of cases develop a febrile illness that can progress to severe neurological disease, including meningitis, encephalitis, and flaccid paralysis, which can be fatal (Sejvar, 2014). DENV is the most common mosquito-transmitted infective agent, responsible of over 300 million unapparent cases per year in humans, of which near 100 million are clinically relevant and cause dozens of hundreds deaths yearly (Pang et al., 2017). On the other hand, since the first detection of ZIKV in the Americas in 2014, the virus has explosively spread thru the continent resulting in the infection of millions of people (Saiz et al., 2016). Until then, ZIKV infection was characterized as causing a mild disease with exceptional reports of an association with Guillain-Barré syndrome, (GBS; Blazquez and Saiz, 2016; Saiz et al., 2016). However, during the recent outbreaks in America, a 20 times increase in the number of cases of microcephaly in fetuses and newborns, and a surprising number of GBS cases have been linked to ZIKV infection (Blazquez and Saiz, 2016). In any case, and despite the efforts of the scientific community, currently no drug against any of these viruses has been licensed, and, thus, the search for effective antiviral compounds is a current milestone in the field (Martin-Acebes and Saiz, 2012; David and Abraham, 2016; Saiz et al., 2016; Saiz and Martin-Acebes, 2017).
In this regard, a novel antiviral approach (economical, simple to use, and environmental friendly) is the use of natural compounds (Martinez et al., 2015), such as the polyphenols present in plants and natural products, such as wine and tea (Hsu, 2015). Polyphenols constitute important metabolites of plants and are generally involved in defense against ultraviolet radiation or aggression by pathogens. These compounds have antioxidant properties and present protection against development of cancers, diabetes, osteoporosis, and cardiovascular and neurodegenerative diseases (Pandey and Rizvi, 2009; Li et al., 2014). Polyphenols have also demonstrated potential antibacterial, antifungal, and antiviral activities. For instances, the antiviral effect of epigallocatechin gallate (EGCG), the most abundant polyphenol in green tea, has been already reported for several viruses: hepatitis C virus, chikungunya virus, hepatitis B virus, herpes simplex virus type 1 virus, influenza A virus, vaccinia virus, adenovirus, reovirus, vesicular stomatitis virus, and, very recently, for ZIKV (Ciesek et al., 2011; Calland et al., 2012, 2015; Huang et al., 2014; Weber et al., 2015; Carneiro et al., 2016). Likewise, other polyphenols have also been described to present antiviral activity, such as honokiol, baicalein, naringin, and quercetin (Zandi et al., 2011, 2012; Johari et al., 2012; Fang et al., 2015).
In this report, we have analyzed the effect of different polyphenols on WNV infection and found that of EGCG and delphinidin (D) inhibited the multiplication of these flaviviruses in cell culture. Even more, both polyphenols also showed a direct effect on WNV, ZIKV, and DENV particles, more probably due to a virucidal activity.
Delphinidin chloride (D), cyanidin chloride (Cy), catechin (C), epicatechin (EC), epigallocatechin (EGC), and EGCG (Extrasynthèse) were resuspended in dimethyl sulfoxide (DMSO) and used at a final concentration of 10 μM.
Vero cells (ATCC CCL-81) were cultured as described (Martin-Acebes et al., 2014; Merino-Ramos et al., 2014). All infectious virus manipulations were performed in our biosafety level 3 (BSL-3) facilities. Viral strains used were: WNV-NY99, WNV mutants with increased resistance (Martin-Acebes et al., 2013) or sensitivity (Martin-Acebes and Saiz, 2011) to NH4Cl, ZIKV PA259459, and DENV-2. Unless otherwise specified, cells were incubated with the corresponding virus for 1 h at 37°C, then viral inoculum was removed, and incubations were continued in culture medium containing 2% fetal bovine serum [time that was considered as 1 h post-infection (p.i)]. Viral titres were determined by plaque assay in semisolid agarose medium at 24 h p.i (Martin-Acebes and Saiz, 2011). The multiplicity of infection (MOI) used in each experiment was expressed as plaque forming units (PFU)/cell, and is indicated in the corresponding figure legend.
The lack of toxicity of the compounds was evaluated by determining the cellular ATP content with a CellTiter-Glo luminescent cell viability assay (Promega).
West Nile virus RNA was extracted from the supernatants of infected cultures using Speedtools RNA virus extraction kit (Biotools B&M Labs S.A.), whereas cell-associated WNV RNA was extracted using TRI Reagent (Thermo Fisher Scientific). The amount of viral RNA copies was determined by quantitative RT-PCR (Blazquez et al., 2016). Genomic equivalents to PFU per milliliter were calculated by comparison with 10-fold serial dilutions of viral RNA extracted from previously titrated sample (Blazquez and Saiz, 2010).
Fluorescent labeling of acidic organelles was performed using LysoSensor Green DND-189 probe (Life Technologies). The LysoSensor reagent exhibits a pH-dependent increase in fluorescence intensity upon acidification. Vero cells were seeded on glass bottom 35-mm dishes (Ibidi), washed with PBS, and treated (4 h) with 25 mM NH4Cl, or 10 μM of either D or EGCG in EMEM supplemented with 25 mM HEPES pH 7.4. Then, the culture medium was replaced by fresh medium containing 1 μM of LysoSensor probe, and cells were incubated during 5 min at 37°C, washed three times with the same media without LysoSensor probe, and observed under a fluorescence microscope. Images were acquired and processed using the same microscope settings. Control cells were treated in parallel with the same amount of drug solvent (H2O for NH4Cl and DMSO for polyphenols).
Analysis of variance (ANOVA) was performed with the SPSS (v.15) statistical package (SPSS Inc.). Data are presented as the mean ± standard deviation (SD). Differences with P-values of <0.05 were considered statistically significant.
The effect of a panel of structurally related polyphenols (Figure 1A) on WNV infection was analyzed in cell culture. To this end, Vero cells were infected with WNV and treated with the different polyphenols. Among the compounds tested, only D, EGC, and EGCG significantly reduced WNV yield at 10 μM concentration (Figure 1B). At this concentration no remarkable toxicity was observed, as cell viability was higher than 80% of the control (Figure 1C). Since D and EGCG exerted the highest inhibition against WNV infection, they were further analyzed. Thereby, quantitative RT-PCR showed that both polyphenols significantly reduced the amount of viral RNA in the supernatant of infected cultures (Figure 1D), which is compatible with the observed reduction on the release of infectious particles (Figure 1B), as well as the amount of cell-associated viral RNA (Figure 1E).
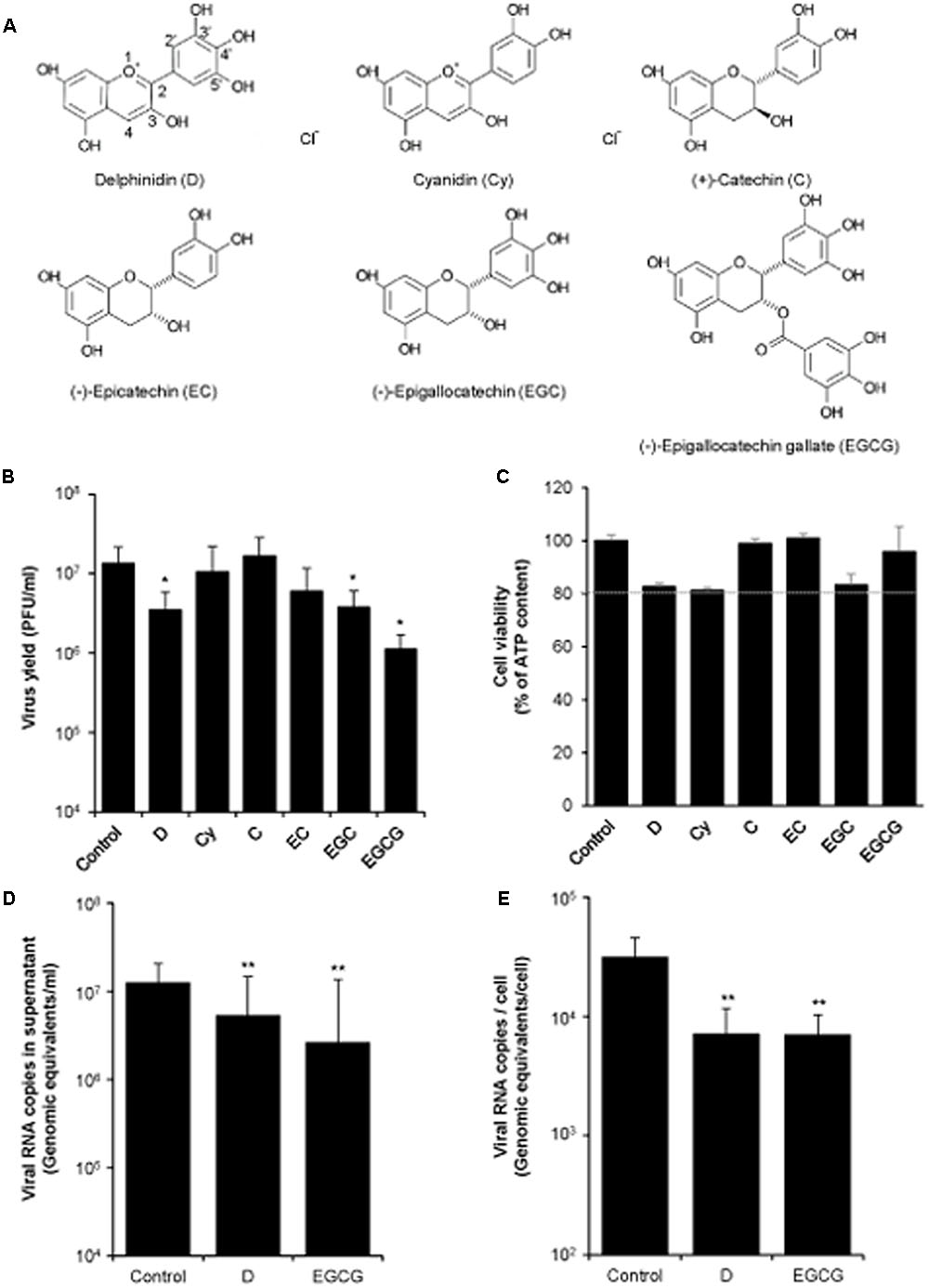
FIGURE 1. Effect of different polyphenols on WNV infection. (A) Schematic representation of the different polyphenols tested. (B) Vero cells were infected with WNV NY99 (MOI of 0.5 PFU/cell) and the different polyphenols (10 μM) were added after the first hour of infection (adsorption time). Virus yield in culture supernatant was determined by plaque assay at 24 h p.i. (C) Mock-infected Vero cells were treated (24 h) in parallel with 10 μM of each polyphenol and the cell viability was determined by measuring the cellular ATP content. Dashed line indicates 80% of cell viability of control cells. (D) Quantification by qRT-PCR of genome-containing particles in culture supernatant of Vero cells infected and treated as in (B). (E) Amount of cell-associated viral RNA in cell cultures infected and treated as in (B) determined by qRT-PCR at 24 h p.i. Data are represented as mean + SD. Statistically significant differences are indicates: ∗P < 0.05; ∗∗P < 0.005.
In order to dissect the step of the infection affected by the polyphenols, D and EGCG were added at different times of infection (Figure 2A). To analyze if they interfere with (i) viral attachment: cells were incubated with the polyphenols together with the virus for 1 h at 4°C, thus allowing virus binding to the cell monolayers, but preventing viral entry (Figure 2A, Attachment). (ii) Viral entry: viruses were adsorbed (1 h) to cell monolayers at 4°C to allow virus attachment and synchronize viral entry. Then, viral inoculum was removed and fresh medium (warmed at 37°C) containing the polyphenol was added for 2 h, time at which the medium was replaced by fresh medium without the drugs. So that, polyphenols were only present during the first 2 h of infection (Figure 2A, Entry). (iii) Late infection steps: cell monolayers were infected as in (ii), incubated with fresh medium warmed at 37°C for 2 h, and then the medium was replaced by fresh medium containing the polyphenols throughout the rest of the experiment (Figure 2A, Post-entry). Using this experimental approach it was noticed that both D and EGCG exhibited antiviral effect only at early steps of the viral infection.
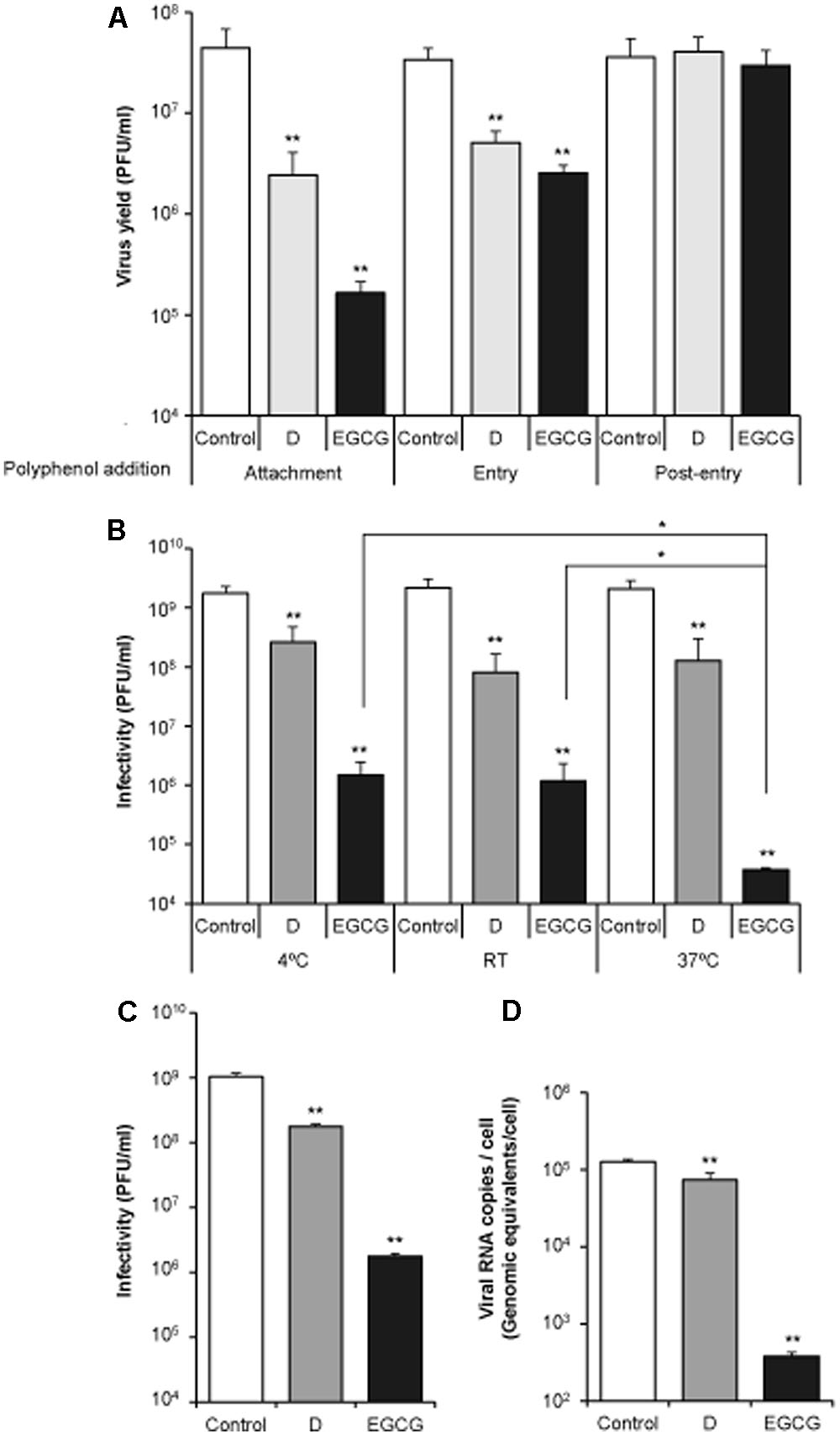
FIGURE 2. Delphinidin (D) and epigallocatechin gallate (EGCG) inhibit WNV infection by exerting a virucidal effect that impairs an early step of the infection. (A) DC and EGCG only inhibit WNV when added at an early step of infection. Vero cells were incubated with WNV (MOI of 1 PFU/cell) at 4°C to allow viral attachment and, then, shifted to 37°C to promote virus entry and infection. The polyphenols were added during this attachment step (Attachment), during the two first hours at 37°C (Entry), or from 2 h after infection until the end of the experiment (Post-entry). Virus yield in supernatant was determined by plaque assay at 24 h p.i. (B) Virucidal effect of the polyphenols on WNV infection. WNV (6 × 108 PFU) were incubated (1 h) with 10 μM of each polyphenol, or the same volume of drug vehicle (DMSO) as a control, at 4°C, RT, or 37°C. Then, the infectivity in each sample was determined by plaque assay. (C,D) Viral infectivity and amount of cell-associated viral RNA in cell cultures infected with pretreated virus as in (B) (37°C) assayed by plaque assay and qRT-PCR at 24 h p.i, respectively. Data are represented as mean + SD. Statistically significant differences are indicates: ∗P < 0.05; ∗∗P < 0.005.
To further explore whether D and EGCG inhibit viral infection by acting directly on the viral particle, WNV (6 × 108 PFU) was incubated with 10 μM of each compound during 1 h, and the remaining infectivity in each sample was determined by plaque assay (Figure 2B). Considering that flaviviruses can undergo temperature dependent conformational changes (Zhang et al., 2015), the assay was performed at different temperatures (4°C, RT, or 37°C). Incubation with D or EGCG reduced significantly the infectivity of WNV in all cases (Figure 2B). Even more, whereas no difference was observed among the temperatures assayed for D, in the case of EGCG incubation at 37°C resulted in a significantly higher reduction of WNV infectivity. Additionally, when viral inoculum was pretreated with the drugs for 1 h at 37°C prior to infect cell monolayers, a significant reduction on virus yield and the accumulation of intracellular viral RNA was noticed for D and EGCG in comparison to control samples (Figures 2C,D). These data further supported the virucidal effect of the compounds. Inhibitions, although relatively similar, were dose-dependent in a 1–10 μM range (data not shown). Overall, these results indicate that D and EGCG affect the early steps of WNV infection, most likely, by exerting a virucidal effect.
Polyphenols, such as catechins, can either inhibit (Imanishi et al., 2002) or enhance (Zhong et al., 2015) endosomal acidification. Therefore, and considering that the acidic pH within the endosomal system promotes viral fusion during WNV entry (Martin-Acebes and Saiz, 2012), we explored whether the antiviral effect of D and EGCG on WNV infection was related with a potential interference of these polyphenols with endosomal pH. To do so, cells were treated with the polyphenols and incubated with LysoSensor probe, which exhibits a pH-dependent increase in fluorescence intensity upon acidification (Figure 3A). As cells treated with NH4Cl, which blocks endosomal acidification, have been shown to inhibit WNV infection (Martin-Acebes et al., 2013) they were included in these experiments. Whereas control untreated cells stained with LysoSensor exhibited a fluorescence punctuated pattern that corresponded to acidic organelles, NH4Cl abolished the presence of these structures, thus confirming that this compound blocked the acidification of the endolysosomal system. Conversely, control cells were indistinguishable from D or EGCG treated-cells, suggesting that these compounds did not impair endosomal acidification (Figure 3A). Remarkably, both polyphenols also inhibited the infection of two WNV variants (Figure 3B) that differed on acidic pH requirements for vial fusion (Martin-Acebes and Saiz, 2011; Martin-Acebes et al., 2013). Even more, they also displayed virucidal effect against these mutant viruses (Figure 3C). Taken together, these results indicate that D and EGCG do not impair endosomal acidification, and suggest that their antiviral activity against WNV is produced, at least partially, by a virucidal effect rather than by an inhibition of pH-dependent viral fusion.
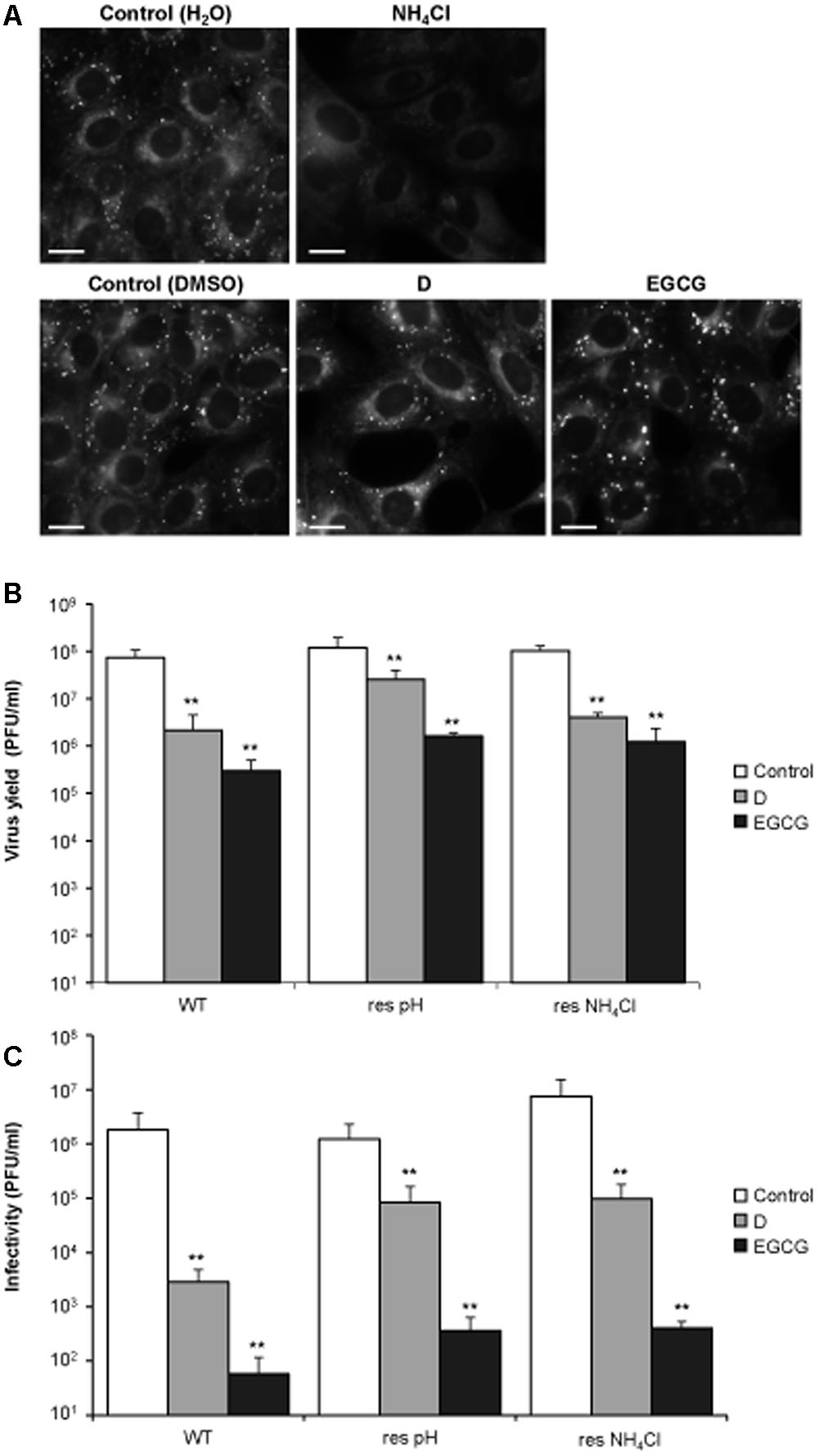
FIGURE 3. The antiviral effect of D and EGCG is not related to an impairment of endosome acidification. (A) DC and EGCG do not inhibit endosome acidification. Vero cells treated (4 h) with 25 mM NH4Cl, 10 μM DC, or 10 μM EGCG were incubated with LysoSensor probe during 5 min and analyzed by fluorescence microscopy. Bars: 10 μm. (B) Vero cells were infected (MOI of 1 PFU/cell) with WNV NY99 (termed WT), a mutant resistant to acidic-induced inactivation (termed res pH), or a mutant resistant to NH4Cl (termed res NH4Cl). DC and EGCG were added at 10 μM 1 h p.i and keep during the whole virus infection. The virus yield in the supernatants was determined by plaque assay at 24 h p.i. (C) 1–2 × 108 PFU of the different WNV strains were incubated with 10 μM during 1 h at 37°C. Then, the remaining infectivity in each sample was determined by plaque assay. Data are represented as mean + SD. Statistically significant differences are indicates: ∗∗P < 0.005.
To extend the virucidal effect of D and EGCG to other medically relevant flaviviruses, ∼106 PFU of two ZIKV strains of different genetic lineages, the African MR766 and the American PA259459 (Figures 4A,B), and of DENV-2 (Figure 4C) were incubated with 10 μM of D, or EGCG, for 1 h at 37°C, temperature at which WNV exerted the highest virucidal effect (Figure 2B). Results showed that both polyphenols reduced the infectivity of ZIKV and DENV, confirming that they exhibit a virucidal effect against medically relevant flaviviruses. In the case of ZIKV, the effect of D and, particularly of EGCG, was higher for the African strain (MR766) than for the American strain (PA259459).
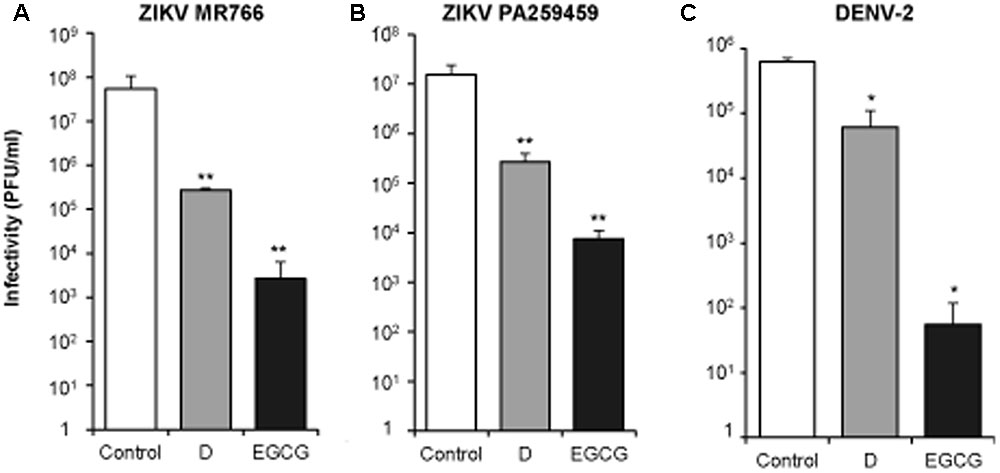
FIGURE 4. Delphinidin and EGCG exhibit virucidal effect against ZIKV and DENV. ∼106 PFU of ZIKV MR766 (A), ZIKV PA259459 (B), or DENV-2 (C) were incubated (1 h at 37°C) with 10 μM DC or EGCG, or the same volume of drug vehicle (DMSO) as a control. Then, the remaining infectivity in each sample was determined by plaque assay. Data are represented as mean + SD. Statistically significant differences are indicates: ∗P < 0.05; ∗∗P < 0.005.
Polyphenolic compounds from plant extracts possess many beneficial properties; including antiviral activities (Hsu, 2015; Parker et al., 2016; Yang et al., 2016). In this study, we have explored the in vitro antiviral capacity of different polyphenols, and shown that EGC, EGCG, and D inhibit WNV infection, and that the last two also inhibit DENV and ZIKV. Even more, in agreement with previous reports for viruses of different families (Ciesek et al., 2011; Calland et al., 2012, 2015; Huang et al., 2014; Weber et al., 2015; Carneiro et al., 2016), the antiviral activity of these compounds was only observed when the polyphenols were added during the early steps of the infection. Further analyses revealed that both, D and EGCG, act as virucidal agents by exhibiting a direct effect on WNV particles. The antiviral activity of D has been previously observed for HCV, a member of the Flaviviridae family (Calland et al., 2015), but this is the first study describing its antiviral activity against three medically relevant flaviviruses (WNV, ZIKV, and DENV).
The mode of action of EGCG and D is not yet well understood, but it has been hypothesized that, in the case of HCV, they might interact with the function of proteins at multiple binding sites (Calland et al., 2015). In this sense, molecular docking and dynamics simulation studies predicted a flavonoid binding pocket on the surface of the E protein of DENV, and showed that EGCG, baicalein, and quercetin potentially form interactions with this viral protein (Ismail and Jusoh, 2016). Therefore, and although further studies are needed it, it seems reasonable to assume a similar mechanism of action for the effect observed here. Notably, differences were observed in the inhibition exerted by EGCG and D to the two different ZIKV strains tested, one African (MR766) and one American (PA259459), especially by EGCG. This differences could be due to the different amino acid composition of the E protein, as the African strain exhibited a 4 amino acid deletion corresponding to the E protein 154 glycosylation motif found in most ZIKV strains and in many flaviviruses (Kuno and Chang, 2007; Baronti et al., 2014; Berthet et al., 2014). In fact, these two ZIKV strains have been recently reported to present different antigenicity and immunogenicity in vivo (Vazquez-Calv et al., unpublished). Moreover, EGCG showed higher virucidal activity against WNV at 37°C than at 4°C, probably because temperature-induced conformational changes (Zhang et al., 2015) allow more interactions between this compound and the WNV particle.
On the other hand, from a chemical point of view, the presence of a trihydroxyphenyl group at R2 in D and EGCG has been related to their anti-HCV activity (Calland et al., 2015). Accordingly, our results showed that an anti-WNV effect was only observed with those polyphenols having a hydroxyl group at R5’ (D, EGC, and EGCG), but not with those lacking it (Cy, C, and EC).
In addition to direct binding to the viral particle as an antiviral mechanism of action, it has been reported that these polyphenolic compounds may also interfere with the endocytic pathway by reducing virus uptake by endocytosis (Huang et al., 2014), or by modulating the endosomal pH (Imanishi et al., 2002; Zhong et al., 2015), which could impair pH-dependent viral fusion. Consistent with these observations, our results provide evidence that the tested polyphenols only have an antiviral effect when added at the early steps of WNV infection. However, no differences on endosomal acidification upon polyphenol treatment were observed. Furthermore, they also impaired the infection of WNV mutants differing in pH requirements for uncoating (Martin-Acebes and Saiz, 2011; Martin-Acebes et al., 2013). Therefore, all these data support that, in the case of WNV, their mechanism of action is not related to an inhibition of the pH necessary for viral fusion but, more likely, throughout a direct effect on the viral particle. In fact, other polyphenols have shown similar mechanisms of action, such as baicalein that exhibited anti-adsorption and virucidal activities against DENV and JEV (Johari et al., 2012), and naringin that presented anti-adsorption and anti-attachment activity against DENV (Zandi et al., 2012).
Due to their properties, polyphenolic compounds have been proposed as broad spectrum antiviral candidates (Colpitts and Schang, 2014; Hsu, 2015). Here, our results unveil the antiviral activity of two different polyphenols, D, and EGCG, against three clinically relevant members of the Flavivirus genus (WNV, ZIKV, and DENV), and point to them as potential scaffolds for future design of antiflaviviral compounds.
Conceived and designed the experiments: AV-C, NJ, MM-A, and J-CS. Performed the experiments: AV-C and NJ. Analyzed the data: AV-C, MM-A, EG-M, and J-CS. Contributed to the writing if the manuscript: AV-C, MM-A, NJ, EM-G, and J-CS.
The work was supported in part by grants PLATESA (P2013/ABI-2906) from the Comunidad Autónoma de Madrid, and E_RTA2013-00013-C04, ZIKA-BIO-2016-01, and RTA2015-00009 from the Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA) to J-CS and AGL2014-56518-JIN from the Spanish Ministry of Economy and Competitiveness (MINECO) to MM-A. AV-C is a recipient of a “Contrato de formación postdoctoral” from MINECO.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank M. Calvo for technical assistance.
Baronti, C., Piorkowski, G., Charrel, R. N., Boubis, L., Leparc-Goffart, I., and de Lamballerie, X. (2014). Complete coding sequence of Zika virus from a French polynesia outbreak in 2013. Genome Announc. 2:e00500-14. doi: 10.1128/genomeA.00500-14
Berthet, N., Nakoune, E., Kamgang, B., Selekon, B., Descorps-Declere, S., Gessain, A., et al. (2014). Molecular characterization of three Zika flaviviruses obtained from sylvatic mosquitoes in the Central African Republic. Vector Borne Zoonotic Dis. 14, 862–865. doi: 10.1089/vbz.2014.1607
Blazquez, A. B., Martin-Acebes, M. A., and Saiz, J. C. (2016). Inhibition of west nile virus multiplication in cell culture by anti-Parkinsonian drugs. Front. Microbiol. 7:296. doi: 10.3389/fmicb.2016.00296
Blazquez, A. B., and Saiz, J. C. (2010). West Nile virus (WNV) transmission routes in the murine model: intrauterine, by breastfeeding and after cannibal ingestion. Virus Res. 151, 240–243. doi: 10.1016/j.virusres.2010.04.009
Blazquez, A. B., and Saiz, J. C. (2016). Neurological manifestations of Zika virus infection. World J. Virol 5, 135–143. doi: 10.5501/wjv.v5.i4.135
Calland, N., Albecka, A., Belouzard, S., Wychowski, C., Duverlie, G., Descamps, V., et al. (2012). (-)-Epigallocatechin-3-gallate is a new inhibitor of Hepatitis C virus entry. Hepatology 55, 720–729. doi: 10.1002/hep.24803
Calland, N., Sahuc, M. E., Belouzard, S., Pene, V., Bonnafous, P., Mesalam, A. A., et al. (2015). Polyphenols inhibit Hepatitis C virus entry by a new mechanism of action. J. Virol. 89, 10053–10063. doi: 10.1128/JVI.01473-15
Carneiro, B. M., Batista, M. N., Braga, A. C., Nogueira, M. L., and Rahal, P. (2016). The green tea molecule EGCG inhibits Zika virus entry. Virology 496, 215–218. doi: 10.1016/j.virol.2016.06.012
Ciesek, S., von Hahn, T., Colpitts, C. C., Schang, L. M., Friesland, M., Steinmann, J., et al. (2011). The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 54, 1947–1955. doi: 10.1002/hep.24610
Colpitts, C. C., and Schang, L. M. (2014). A small molecule inhibits virion attachment to heparan sulfate- or sialic acid-containing glycans. J. Virol. 88, 7806–7817. doi: 10.1128/JVI.00896-14
David, S., and Abraham, A. M. (2016). Epidemiological and clinical aspects on West Nile virus, a globally emerging pathogen. Infect. Dis. 48, 571–586. doi: 10.3109/23744235.2016.1164890
Fang, C. Y., Chen, S. J., Wu, H. N., Ping, Y. H., Lin, C. Y., Shiuan, D., et al. (2015). Honokiol, a lignan biphenol derived from the magnolia tree, inhibits dengue virus type 2 infection. Viruses 7, 4894–4910. doi: 10.3390/v7092852
Hsu, S. (2015). Compounds derived from epigallocatechin-3-gallate (EGCG) as a novel approach to the prevention of viral infections. Inflamm. Allergy Drug Targets 14, 13–18. doi: 10.2174/1871528114666151022150122
Huang, H. C., Tao, M. H., Hung, T. M., Chen, J. C., Lin, Z. J., and Huang, C. (2014). (-)-Epigallocatechin-3-gallate inhibits entry of hepatitis B virus into hepatocytes. Antiviral Res. 111, 100–111. doi: 10.1016/j.antiviral.2014.09.009
Imanishi, N., Tuji, Y., Katada, Y., Maruhashi, M., Konosu, S., Mantani, N., et al. (2002). Additional inhibitory effect of tea extract on the growth of influenza A and B viruses in MDCK cells. Microbiol. Immunol. 46, 491–494. doi: 10.1111/j.1348-0421.2002.tb02724.x
Ismail, N. A., and Jusoh, S. A. (2016). Molecular docking and molecular dynamics simulation studies to predict flavonoid binding on the surface of DENV2 E protein. Interdiscip. Sci. doi: 10.1007/s12539-016-0157-8 [Epub ahead of print].
Johari, J., Kianmehr, A., Mustafa, M. R., Abubakar, S., and Zandi, K. (2012). Antiviral activity of baicalein and quercetin against the Japanese encephalitis virus. Int. J. Mol. Sci. 13, 16785–16795. doi: 10.3390/ijms131216785
Kuno, G., and Chang, G. J. (2007). Full-length sequencing and genomic characterization of Bagaza, Kedougou, and Zika viruses. Arch. Virol. 152, 687–696. doi: 10.1007/s00705-006-0903-z
Li, A. N., Li, S., Zhang, Y. J., Xu, X. R., Chen, Y. M., and Li, H. B. (2014). Resources and biological activities of natural polyphenols. Nutrients 6, 6020–6047. doi: 10.3390/nu6126020
Martin-Acebes, M. A., Blazquez, A. B., de Oya, N. J., Escribano-Romero, E., Shi, P. Y., and Saiz, J. C. (2013). A single amino acid substitution in the core protein of West Nile virus increases resistance to acidotropic compounds. PLoS ONE 8:e69479. doi: 10.1371/journal.pone.0069479
Martin-Acebes, M. A., Merino-Ramos, T., Blazquez, A. B., Casas, J., Escribano-Romero, E., Sobrino, F., et al. (2014). The composition of West Nile virus lipid envelope unveils a role of sphingolipid metabolism in flavivirus biogenesis. J. Virol. 88, 12041–12054. doi: 10.1128/JVI.02061-14
Martin-Acebes, M. A., and Saiz, J. C. (2011). A West Nile virus mutant with increased resistance to acid-induced inactivation. J. Gen. Virol. 92, 831–840. doi: 10.1099/vir.0.027185-0
Martin-Acebes, M. A., and Saiz, J. C. (2012). West Nile virus: a re-emerging pathogen revisited. World J. Virol. 1, 51–70. doi: 10.5501/wjv.v1.i2.51
Martinez, J. P., Sasse, F., Bronstrup, M., Diez, J., and Meyerhans, A. (2015). Antiviral drug discovery: broad-spectrum drugs from nature. Nat. Prod. Rep. 32, 29–48. doi: 10.1039/c4np00085d
Merino-Ramos, T., Blazquez, A. B., Escribano-Romero, E., Canas-Arranz, R., Sobrino, F., Saiz, J. C., et al. (2014). Protection of a single dose west nile virus recombinant subviral particle vaccine against lineage 1 or 2 strains and analysis of the cross-reactivity with Usutu virus. PLoS ONE 9:e108056. doi: 10.1371/journal.pone.0108056
Pandey, K. B., and Rizvi, S. I. (2009). Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2, 270–278. doi: 10.4161/oxim.2.5.9498
Pang, T., Mak, T. K., and Gubler, D. J. (2017). Prevention and control of dengue-the light at the end of the tunnel. Lancet Infect. Dis. 17, e79–e87. doi: 10.1016/S1473-3099(16)30471-6
Parker, S., May, B., Zhang, C., Zhang, A. L., Lu, C., and Xue, C. C. (2016). A pharmacological review of bioactive constituents of Paeonia lactiflora Pallas and Paeonia veitchii Lynch. Phytother. Res. PTR 30, 1445–1473. doi: 10.1002/ptr.5653
Saiz, J. C., and Martin-Acebes, M. A. (2017). Zika virus: a race in search for antivirals. Antimicrob. Agents Chemother. 61, e411–e417. doi: 10.1128/AAC.00411-17
Saiz, J. C., Vazquez-Calvo, A., Blazquez, A. B., Merino-Ramos, T., Escribano-Romero, E., and Martin-Acebes, M. A. (2016). Zika Virus: the latest newcomer. Front. Microbiol. 7:496. doi: 10.3389/fmicb.2016.00496
Sejvar, J. J. (2014). Clinical manifestations and outcomes of West Nile virus infection. Viruses 6, 606–623. doi: 10.3390/v6020606
Weber, C., Sliva, K., von Rhein, C., Kummerer, B. M., and Schnierle, B. S. (2015). The green tea catechin, epigallocatechin gallate inhibits chikungunya virus infection. Antiviral Res. 113, 1–3. doi: 10.1016/j.antiviral.2014.11.001
Yang, C., Zhi, X., and Xu, H. (2016). Advances on semisynthesis, total synthesis, and structure-activity relationships of honokiol and magnolol derivatives. Mini Rev. Med. Chem. 16, 404–426. doi: 10.2174/1389557516666151120115558
Zandi, K., Teoh, B. T., Sam, S. S., Wong, P. F., Mustafa, M. R., and Abubakar, S. (2011). Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J. 8:560. doi: 10.1186/1743-422X-8-560
Zandi, K., Teoh, B. T., Sam, S. S., Wong, P. F., Mustafa, M. R., and Abubakar, S. (2012). Novel antiviral activity of baicalein against dengue virus. BMC Complement. Altern. Med. 12:214. doi: 10.1186/1472-6882-12-214
Zhang, X., Sun, L., and Rossmann, M. G. (2015). Temperature dependent conformational change of dengue virus. Curr. Opin. Virol. 12, 109–112. doi: 10.1016/j.coviro.2015.04.006
Keywords: Flavivirus, polyphenol, delphinidin, epigallocatechin gallate, virucidal, West Nile virus, Dengue virus, Zika virus
Citation: Vázquez-Calvo Á, Jiménez de Oya N, Martín-Acebes MA, Garcia-Moruno E and Saiz J-C (2017) Antiviral Properties of the Natural Polyphenols Delphinidin and Epigallocatechin Gallate against the Flaviviruses West Nile Virus, Zika Virus, and Dengue Virus. Front. Microbiol. 8:1314. doi: 10.3389/fmicb.2017.01314
Received: 29 March 2017; Accepted: 29 June 2017;
Published: 11 July 2017.
Edited by:
Shiu-Wan Chan, University of Manchester, United KingdomReviewed by:
Peter John Walker, The University of Queensland, AustraliaCopyright © 2017 Vázquez-Calvo, Jiménez de Oya, Martín-Acebes, Garcia-Moruno and Saiz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan-Carlos Saiz, amNzYWl6QGluaWEuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.