 Martine Denis
Martine Denis Bérengère Nagard
Bérengère Nagard Valérie Rose
Valérie Rose Annaëlle Kerouanton
Annaëlle Kerouanton- ANSES, Hygiene and Quality of Poultry and Pig Products Unit, Bretagne-Loire University, Ploufragan, France
To evaluate the impact of pig farm management on the genetic diversity and on the virulence of Campylobacter coli, we characterized isolates from 19 organic pig farms (62 isolates) and from 24 conventional pig farms (58 isolates). The 120 C. coli isolates were typed using pulsed field gel electrophoresis (PFGE) and multilocus sequence typing (MLST) and the presence of nine virulence genes was screened using real-time PCR. The capacity of adhesion and invasion of 61 isolates (32 from organic and 29 from conventional farms) were then tested on human intestinal Caco-2 cells. A total of 59 PFGE types and of 50 sequence types (STs) were identified. Twelve PFGE types and nine STs, accounting for 34 and 41.6% of the isolates, respectively, were common between the two production systems with ST854 dominating (18.3% of the isolates). Twenty-nine PFGE types and 25 STs were only found in isolates from organic farms, and 18 PFGE types and 16 STs from conventional farms. No significant differences were found in diversity despite the differences in rearing systems, except at the locus level for the glnA, gltA, and uncA genes. All isolates, regardless of their origin, carried the ceuE, iam, ciaB, and flaA genes and more than 95% of the isolates carried the cadF and cdtABC genes. No significant differences were found in pathogenicity between the two farming systems. The pathogenicity of the C. coli isolates was low compared to C. jejuni control strains tested. The plasmid gene virb11 was detected in only 13 isolates from organic farms; these isolates showed greater invasion capacity than those without this gene. Our study indicates that pig farm management does not significantly affect the diversity and the virulence of Campylobacter coli isolated from pigs. The common genotypes between conventional and organic farms may indicate that some genotypes are adapted to pigs.
Introduction
New consumer trends focus on products derived from systems that promote good animal welfare conditions and a high safety level. Organic pig production differs in many ways from conventional pig production, particularly in terms of antibiotic use, herd structure, feeding regimes, access to outdoor areas, and space allowance per pig. More specifically, the European Union regulations for organic farming (Council Directives 2007/834/EC and 2008/889/EC) require that animals have access to an outdoor area. In addition, the preventive use of chemically-synthesized allopathic veterinary medicinal products is not authorized, and may be used only for treatment of sick animals when necessary. If more than one treatment, derived products are no longer considered organic.
Recently, we investigated the carriage of antibiotic-resistant Escherichia coli in colons at the slaughterhouse and in feces on organic and conventional pig farms in four European countries (SafeOrganic project, Österberg et al., 2016). We also studied the carriage of resistant Campylobacter from the same samples in two European countries (SafeOrganic project, Kempf et al., 2017). In France, the level of antibiotic resistance in E. coli and Campylobacter coli is lower for organic pig production than for conventional production, suggesting that practices such as little or no use of antibiotics on organic pig farms can affect the level of bacterial resistance. Several studies (Saini et al., 2013; Garcia-Migura et al., 2014) indicate that extensive use of antibiotics produces a selection pressure favoring resistance among commensal bacteria from animals.
Because the management of conventional and organic pig farms has an impact on Campylobacter resistance to antibiotics (Kempf et al., 2017), we assumed that the management of these two types of pig production systems (in terms of antibiotic use and access to the outdoors) may also have an impact on the diversity of Campylobacter isolates excreted by pigs and on the virulence of these isolates. The more frequent use of antibiotics and confinement of pigs in a building in conventional farming may reduce the number of Campylobacter genotypes. In contrast, in organic farming, little or no use of antibiotics and access to an outdoor area may promote the presence of a higher number of Campylobacter genotypes. Access to an outside area increases exposure of animals to environmental sources of different microorganisms including Campylobacter jejuni (Greig et al., 2015).
Thus in this study, the C. coli previously isolated from pigs from organic and conventional farms to test their resistance to antibiotics (Kempf et al., 2017) were typed using two molecular typing methods, and tested for their virulence.
Materials and Methods
Origin of the Isolates
The Campylobacter isolates considered in this study were isolated by our laboratory, which is also the French National Reference Laboratory for Campylobacter as part of the SafeOrganic project. Sampling and isolation methods for Campylobacter are described in Kempf et al. (2017). Briefly, colon contents were sampled at one slaughterhouse from 114 pigs. These pigs came from 31 organic pig batches (56 pigs) and 31 conventional pig batches (58 pigs). These batches involved 21 organic farms and 29 conventional farms, all located within 200 km of the slaughterhouse. Out of the 50 sampled farms, 43 farms were positive for Campylobacter: 19 organic farms and 24 conventional farms.
The isolates were kept at −80°C in peptone glycerol broth. They were all identified as C. coli and tested for their antibiotic resistance (Kempf et al., 2017). Here, we randomly selected two to three isolates per positive farm for a total of 120 isolates: 62 isolates from 19 organic farms and 58 isolates from 24 conventional farms.
DNA Extraction
The 120 isolates were cultured on blood agar plates (Oxoid, Dardilly, France) for 48 h at 37°C in a micro-aerobic atmosphere (5% O2, 10% CO2, 85% N2). A few colonies from the bacterial culture were used for DNA extraction using the InstaGene® Matrix (BioRad Laboratories, Marnes-la-Coquette France) according to the manufacturer's recommendations. DNA was adjusted to 10 ng/μl and intended for use in PCRs for virulence gene detection, and multilocus sequence typing (MLST) as described below. The remaining colonies were used for genotyping by pulsed-field gel electrophoresis (PFGE).
Pulsed-Field Gel Electrophoresis (PFGE) and Analysis of Electrophoretic Profiles
DNA preparation, restriction endonuclease digestion with the KpnI enzyme and PFGE were carried out as described by the Campynet protocol (Rivoal et al., 2005). One KpnI restriction profile was obtained for each isolate.
Electrophoretic patterns were compared using BioNumerics v. 6.5 software (Applied Maths, Sint-Martens-Latem, Belgium). Similarities between profiles, based on band positions, were determined by calculating the Dice correlation coefficient, with a maximum position tolerance of 1%. A dendrogram based on the KpnI restriction profiles was constructed to represent the similarities between the isolates in the matrix. Isolates were clustered by the unweighted pair-group method using the arithmetic mean (UPGMA) (Struelens, 1996). Isolates displaying high levels of similarity were clustered together using a threshold of 80% (Denis et al., 2008) and considered as the same PFGE type.
The Simpson's index (D) was determined as described by Hunter (1990), and was given with a 95% confidence interval, as described by Grundmann et al. (2001). This index was used to assess the genetic diversity of the Campylobacter populations.
Multilocus Sequence Typing (MLST)
The seven housekeeping genes for MLST (aspA, glnA, gltA, glyA, pgm, tkt, and uncA) were amplified and sequenced according to previously developed experimental conditions (Dingle et al., 2001; Miller et al., 2005). PCR products were cleaned up using the ExoSAP-IT treatment (GE Healthcare), and sequence extension reactions were carried out in BigDye Ready reaction mix according to the manufacturer's instructions. Unincorporated dye terminators were removed using an ethanol precipitation method before the products were analyzed on an ABI Prism 3130 sequencer (Applied Biosystems). The sequences were assembled using the assembler implemented in BioNumerics v. 6.5 software. All allelic sequences were queried against the C. jejuni MLST database (http://pubmlst.org/campylobacter). Alleles already present in the database were assigned the numbers given there; novel alleles and sequence types (STs) were submitted to the MLST database and assigned new numbers. STs were assigned into genetically related clusters called clonal complexes (CCs), based on the sharing of four or more alleles with the central genotype that had been identified in previous studies using the BURST algorithm and UPGMA cluster analysis (Dingle et al., 2002).
Detection of Virulence Genes
Table 1 shows the nine virulence genes screened in the present study. Eight of the genes are localized on the bacterial chromosome and one on the plasmid (virB11). These genes are involved in adhesion/invasion of epithelial cells (flaA, ciaB, cadF, iam, virB11), in the acquisition of iron (ceuE), and in the production of the cytholethal distenting toxin (CDT) (cdtA, cdtB, cdtC).
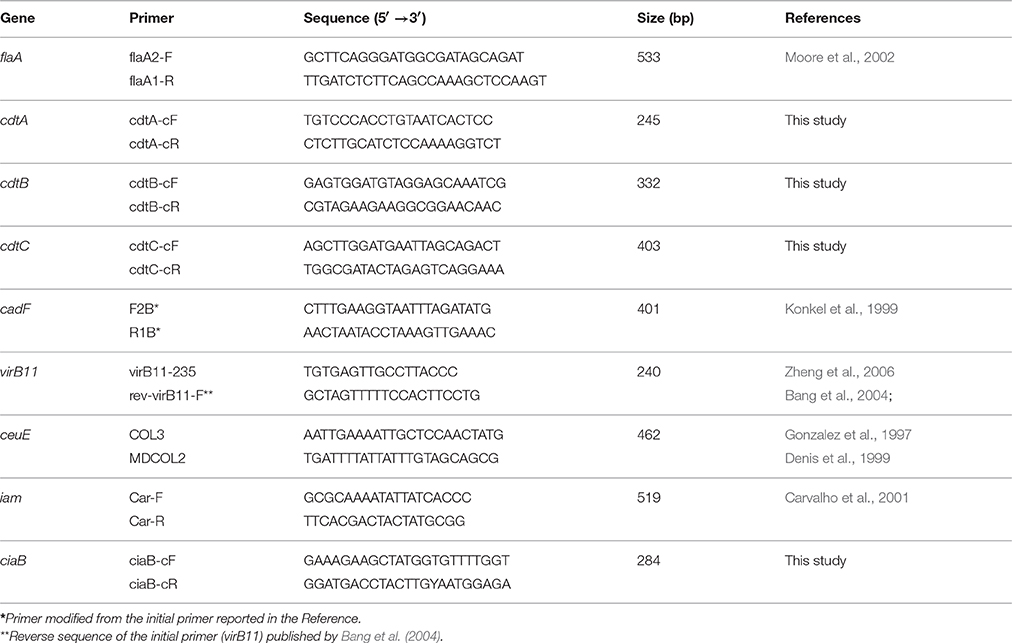
Table 1. Primer sequences for the detection of the nine virulence genes in Campylobacter coli.
The presence of these nine virulence genes in the 120 isolates was checked using real-time PCR developed for this study using primers published by Gonzalez et al. (1997), Denis et al. (1999), Konkel et al. (1999), Carvalho et al. (2001), Moore et al. (2002), Bang et al. (2004), or by Zheng et al. (2006) or using primers designed by pour laboratory for this study (Table 1).
Some published primers were slightly modified (one or two bases added or removed) to obtain primers with the same (or very similar) melting temperature (indicated by * in Table 1). We used also the reverse sequence of the initial primer (virB11) published by Bang et al. (2004).
We designed eight primers from a sequence alignment using Multalin v. 5.4.1 (INRA, France) for the detection of the three cdt genes (GenBank accession numbers: AB562905, AB274801, AB274800, AB274799, AB274798, AB274797, AB274796, AB274795, AB274794, AB274793, AB182109) and the ciaB gene (GenBank accession numbers: HG326877, CP006702, AB433217, CP004066).
The size of the PCR products was estimated by in silico PCR (FastPCR online v. 2.07, PrimerDigital) and confirmed after electrophoresis on an agarose gel.
Each PCR was carried out in a total volume of 25 μl with the mix SYBR® Green JumpStart™ Taq ReadyMix™ from Sigma-Aldrich, in which 1 μl of each primer at 10 μM was added. All nine amplifications were done using the same PCR conditions: 35 cycles, each cycle with a first step at 95°C for 1 min, a second step at 56°C for 1 min and a final step at 72°C for 1 min 40 s. The PCRs finished with an incremental step from 60 to 95°C, increasing by 0.5°C every 5 s to obtain the fusion curve.
Three isolates of human origin were used as positive controls (Table 2): C. jejuni 81–176 (with pVir plasmid) and C. jejuni NCTC11168 (without the pVir plasmid) (purchased from the Pasteur Institute Collection, Paris) and C. coli 04FM842 (purchased from the French National Reference Center) genetically close to C. coli from pigs by PFGE and with all the virulence genes except virB11.
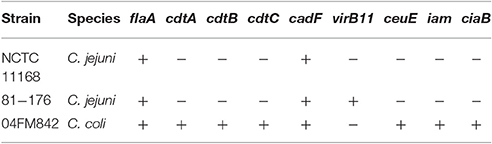
Table 2. Presence (+) of the nine genes in the three strains used as positive PCR controls.
Adhesion and Invasion Assay on Human Intestinal Epithelial Cells
For this assay, we selected 61 isolates (32 from the 19 organic farms and 29 from the 24 conventional farms) on the basis of their PFGE profile and ST to ensure good representativeness of the isolates from each farm.
Capacity of adhesion and invasion of the isolates was tested in vitro on Caco-2 human intestinal epithelial cells following the protocol developed in our laboratory by Guyard-Nicodème et al. (2013). Caco-2 cells (ECACC 86010202) were obtained from the European Collection of Cell Culture (ECACC, Salisbury, UK). Capacity of adhesion and invasion of the isolates was expressed as the percentage of adhesives cells (p_adh) and of invasive cells (p_inv), respectively. For each isolates, the results were the mean of at least two separate determinations.
C. jejuni and C. coli strains isolated from humans and poultry were also tested to compare the invasiveness of our isolates with other Campylobacter. The three human strains were C. jejuni 81–176 (with pVir plasmid), C. jejuni NCTC11168 (without pVir plasmid), and C. coli 04FM842 (without pVir plasmid). The poultry strains were Plouf12 (C. jejuni from poultry) and Plouf13 (C. coli from poultry), both previously tested on pig in a previous study (Leblanc Maridor et al., 2011), 17MD18, 47MD12 (two C. coli from poultry), 54MD16, and 27MD13 (two C. jejuni from poultry) isolated from a previous study (Denis et al., 2008), CRL204-08 (C. coli from poultry) purchased from the European Reference Laboratory, Uppsala, Sweden. We added also a C. coli reference strain isolated from a pig (CIP70.80T) purchased from the collection of Pasteur Institute, Paris, France.
Data Analysis
The distribution of the PFGE types or STs on organic farms was compared to that of conventional farms using the chi-square test of independence in R software (version 3.2.5). For each typing method, we considered the number of isolates from one production system that shared types common with the other system, and the number of isolates found in only one of the two production systems. The distribution was considered statistically different between the two production systems when p-values were lower than 0.05.
Using the “comparing partitions” method (http://www.comparingpartitions.info), we compared the distribution of the PFGE types of the 120 isolates with the distribution of the STs of these isolates. We also compared the distribution of the antibiotic resistance (ATB) profiles of the 120 isolates with the distribution of the PFGE types or STs of these isolates. A p-value was calculated using the jackknife pseudo-values method. We considered that the PFGE/ST, ATB/PFGE, or ATB/ST associations were weak if their distributions were significantly different (p < 0.05).
Results of adhesion to or invasion of Caco-2 cells were analyzed using the Mann-Whitney test in R software version 3.2.5. A p-value lower than 0.05 was considered significant. C. coli were also classified into three classes of pathogenicity (low, intermediate and high) using hierarchical clustering with the method “hclust ward D2” implemented in R.
Results
Genetic Diversity
We observed high genetic diversity for our 120 C. coli whatever the typing method. A total of 110 Kpn1 PFGE profiles were associated with 59 PFGE types (Figure 1) when clustered at 80% of similarity. The 62 isolates from organic farms showed 41 PFGE types and the 58 isolates from conventional farms showed 30 PFGE types. Organic farms and conventional farms shared 12 PFGE types (12/59) representing 34.16% of the isolates (41/120). Among the 59 PFGE types, 29 were found only from organic farm isolates (40 isolates) and 18 from conventional farm isolates (39 isolates). The diversity of the Campylobacter populations was slightly higher in organic isolates with an index of diversity D = 0.98 CI95%[0.97–0.99] than in conventional isolates with D = 0.96 CI95%[0.94–0.98]. The distribution of the PFGE types in the two production systems was not significantly different (χ2, p = 0.345).
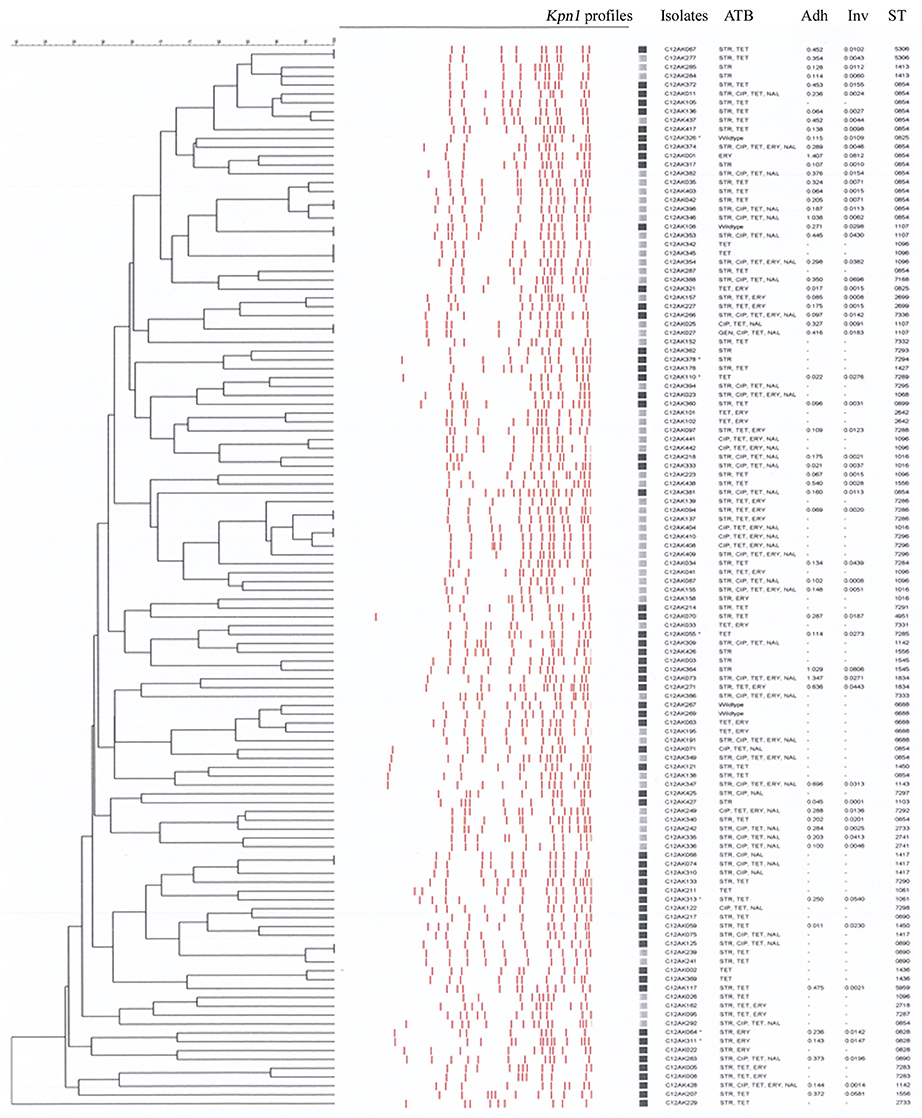
Figure 1. Dendrogram of the Kpn1 profiles of the 120 Campylobacter coli isolates from organic and conventional pig farms (optimization, 1%; tolerance, 1%; active zones, [7.0–87.0%]). Black squares, isolates from organic pig farm; Gray squares, isolates from conventional pig farms; ATB, profile of antibiotic resistance; Adh, percentage of adhesion on Caco-2 cells; Inv, percentage of invasion on Caco-2 cells; ST, sequence type.
Almost all isolates (91%) were from the ST828 CC; the other isolates (11) had no identified CC. Isolates were distributed among 50 STs (Figures 1, 2). The 62 isolates from organic farms were distributed into 34 STs and the 58 isolates from conventional farms into 26 STs. Nine STs (9/50), representing 41.6% of the isolates (50/120), were common to both organic and conventional farms, with a ST854 dominating (18.3% of the isolates, with 10 isolates from 8 organic farms and 12 isolates from 12 conventional farms). Among the 50 STs, 25 STs were only found in organic farm isolates (38 isolates) and 16 in conventional farm isolates (32 isolates), where ST1096 was found in 9 isolates. The diversity of the Campylobacter populations was higher in organic production systems with an index of diversity D = 0.96 CI95%[0.94–0.99] than in conventional production (D = 0.93 CI95%[0.89–0.97]). The distribution of the STs in the two production systems was not significantly different (χ2, p = 0.496).
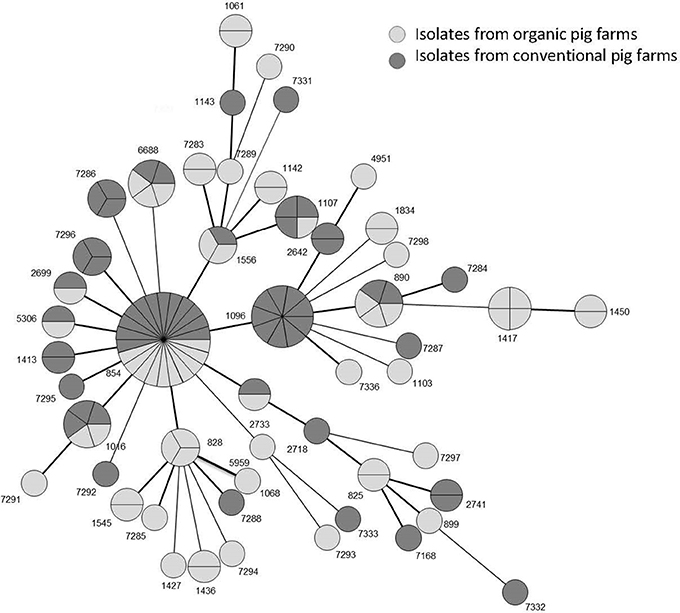
Figure 2. Distribution of the 120 isolates according their sequence type (ST) in a phylogenetic tree drawn using BioNumerics software.
The distribution of the STs was significantly different from the distribution of PFGE types (p = 0.003), indicating that STs are only weakly related to PFGE types.
Genetic diversity at individual MLST loci is shown in Table 3. The mean genetic diversity was higher in organic farms (0.435 ± 0.154) than in conventional farms (0.333 ± 0.206). Allelic diversity was higher for the pgm and tkt loci in both systems with a greater number of alleles for these two genes (Table 3). Between the two production systems, we observed a significant difference in genetic diversity for the glnA, gltA, and uncA locus, with higher genetic diversity for the organic system.
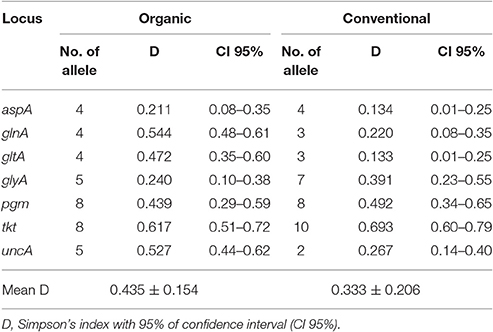
Table 3. Genetic diversity at individual loci of the multilocus sequence type for the 120 C. coli isolates from organic and conventional pig farms.
Association between Genetic Profiles and Antibiotic Resistance Profiles
The 120 Campylobacter coli were previously tested for their resistance to antibiotics. Resistant to tetracycline and erythromycin, and the number of resistances were significantly higher in isolates from conventional farms (Kempf et al., 2017).
The distribution of the ATB profiles was significantly different from the distribution of the PFGE types (p = 0.009) or the distribution of the STs (p = 0.012), indicating that ATB profiles are weakly related to PFGE types or STs.
However, the ST854 C. coli isolates predominantly showed resistance to tetracycline (90.9% of the ST854 isolates), streptomycin (90.9%), and susceptibility to erythromycin (86.3%).
Moreover, we noted that 76% of the isolates resistant to tetracycline were isolates with the allele 38 of the glnA gene (χ2, p = 0.007).
Presence of the Virulence Genes
Regardless of pig origin, all the isolates carried the ceuE, iam, ciaB, and flaA genes. Moreover, all isolates from organic pigs carried also the cadF gene and the three cdt genes. One conventional isolate did not have the cadF gene and three conventional isolates lacked the three CDT genes. The plasmid gene virB11 was detected only in eight isolates (7%), all from organic pigs. The presence of the plasmid was neither associated with a particular PFGE profile (see * in Figure 1) nor with a particular ST [ST 7285, 1450, 828 (2 isolates), 729, 1061, 825, and 724].
Capacity of the Isolates to Adhere and Invade Caco-2 Cells
The percentage of adhesion and invasion of the 61 C. coli are indicated on the dendrogram in Figure 1. Adhesion was 0.30 and 0.28% on average for organic pig and conventional pig isolates, respectively, and invasion was 0.019 and 0.015% on average for organic pig and conventional pig isolates, respectively. There were no significant differences between the isolates from organic pigs and isolates from conventional pigs for adhesion (Mann-Whitney test, p = 0.523) or invasion (Mann-Whitney test, p = 0.590) (Figure 3).
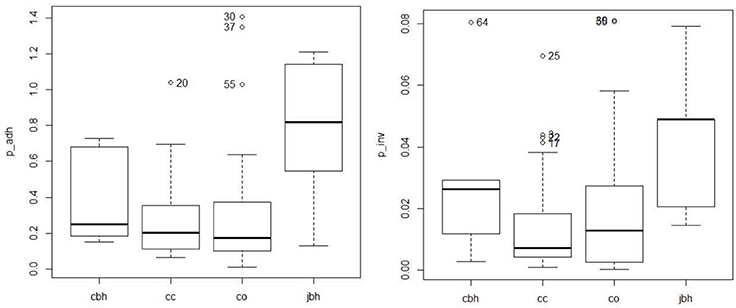
Figure 3. Distribution of the percentage of adhesion (p_adh) and the percentage of invasion (p_inv) according the origin of the isolates. cc, C. coli from conventional pig farm; co, C. coli from organic pig farm; cbh, C. coli from poultry and humans; jbk, C. jejuni from poultry and humans.
The C. coli isolates' capacity to adhere and invade Caco-2 cells was significantly lower than C. jejuni isolates isolated from poultry or humans (Mann-Whitney test, p-value for p_adh = 0.018 and p-value for p_inv = 0.014), but not significantly different from those of C. coli isolates isolated from poultry or humans (p > to 0.05) (Figure 3).
Among these isolates, eight had the plasmid gene virB11. There was no significant difference for adhesion between isolates with or without this gene (Mann-Whitney test, p = 0.058), but a significant difference was observed for invasion. Isolates with the virB11 gene had a higher capacity to invade Caco-2 cells than isolates without this gene (Mann-Whitney test, p = 0.040). We also observed that the human C. jejuni strain 81–176 carrying the virB11 gene had a higher invasion capacity (0.079%) than the human C. jejuni strain NCTC 11168 without the virB11 gene (0.015%).
The 61 C. coli were classified into three classes of pathogenicity (low, intermediate and high) from the hierarchical clustering done on the adhesion and invasion values (Figure 4). It was difficult to associate a ST with a virulence profile because there were too few isolates representing each ST, with the exception of ST854 for which 16 isolates were tested on Caco-2 cells. These ST854 isolates were distributed among the three classes of pathogenicity.
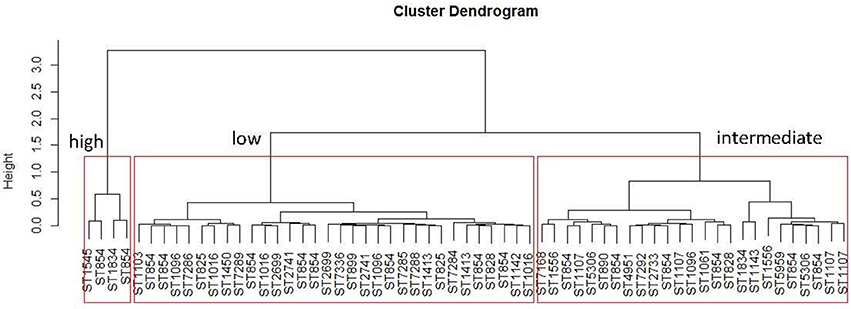
Figure 4. Distribution of the sequence types of the 61 C. coli isolates tested on Caco2-cells according their percentages of adhesion and invasion. The hierarchical clustering was done with R software using the method “hclust ward D2”. Three clusters were obtained for low, intermediate and high level pathogenicity for the C. coli tested.
Discussion
In this study, the C. coli previously isolated from organic and conventional pig farms to test their resistance to antibiotics (Kempf et al., 2017) were typed by PFGE and MLST and tested for their virulence. We wanted to determine if the production system has an impact on the genetic diversity and on the virulence of these isolates excreted by the pigs.
First, our study focused only on C. coli, the only species that was isolated from our pigs. We expected that outdoor pigs would be more exposed to C. jejuni from the environment because wild animals, particularly birds, can shed Campylobacter species other than C. coli (Greig et al., 2015). However, although Jensen et al. (2006) showed that the paddock environment of organic pigs was contaminated by non-pig strains, e.g., by wild fauna strains, they did not demonstrate that these strains could contaminate pigs, at least not C. jejuni strains. Pigs show a dominance of C. coli (Nielsen et al., 1997; Alter et al., 2005; Boes et al., 2005) and C. jejuni may co-exist with C. coli in pigs, but C. jejuni is typically present at in numbers that are 10–100-fold lower than C. coli (Madden et al., 2000; Jensen et al., 2005). In our study, we tested only two to three isolates per farm which may explain also why we never detected C. jejuni.
PFGE and MLST revealed high genetic diversity in our C. coli populations, as previously observed in French pig farms using PFGE (Denis et al., 2011). Some of the STs have been already described in other countries: ST854 and ST2718 from pig livers in Germany (von Altrock et al., 2013), ST854, ST890, ST1068, ST1096, ST1142, ST1413, ST1417, ST1436, and ST1450 from pig farms in USA (Thakur et al., 2006), and ST828, ST854, ST890, ST1016, ST1061, ST1096, ST1413, ST1556, ST2733, and ST4951 from pig feces at slaughterhouses in Switzerland (Egger et al., 2012). The most prevalent ST in our study was ST854. This ST was also reported to be predominant in pig farms in the USA (Quintana-Hayashi and Thakur, 2012) and has been detected all along the production chain, from pig farms to retail pork meat products (Abley et al., 2012). It is also the most frequently recovered ST from surface of pig livers in Germany (von Altrock et al., 2013). In our study, the ST854 isolates predominantly exhibited the TET+STR+ERY− phenotype whereas the CIP/NAL+TET+ phenotype was mainly found for this ST in the USA study (Quintana-Hayashi and Thakur, 2012).
We also noted that allelic diversity was higher at the pgm and tkt loci than the other virulence loci, with a greater number of alleles for these two loci. Quintana-Hayashi and Thakur (2012) also observed higher allelic diversity for the tkt and glyA loci in C. coli populations from swine farms.
Although the genetic diversity was higher in organic production than in conventional production for both typing methods, PFGE and MLST, there were no significant differences, except at the locus level for the glnA, gltA, and uncA genes. There were a higher number of alleles for these genes when isolates were from organic pigs. Finally, we were not able to clearly demonstrate that organic production practices with little or no use of antibiotics and outdoor access for pigs promotes a higher number of Campylobacter genotypes. There were common Campylobacter genotypes shared between both production systems. We already identified these PFGE types on French pig production farms in 2008 (Denis et al., 2011) and two STs (ST584, ST890) were also isolated by Thakur et al. (2006) from conventional and antimicrobial-free pig farms in the USA. Our results suggest that these common genotypes are adapted to the pig and that other genotypes are likely specific to the farm environment where the pigs are grown.
The production system does not select for specific virulence gene profiles, with the exception of the plasmid gene virB11. Almost all the isolates carried the ceuE, iam, ciaB, flaA, cadF genes, and the three cdt genes. High prevalence for these genes has been obtained on C. coli in many studies (Bang et al., 2003; Rozynek et al., 2005; Wieczorek and Osek, 2010, 2013; Andrzejewska et al., 2011; Acik et al., 2013; Khoshbakht et al., 2013) except for the ciaB gene. The detection of ciaB varies with species and study, ranging from 20% for C. coli (Wieczorek and Osek, 2010; Acik et al., 2013) up to 100% for C. jejuni (Datta et al., 2003; Feodoroff et al., 2010). This discrepancy may be due to the primers used; we designed new primers that facilitated the detection of the ciaB gene in all our C. coli isolates.
The plasmid gene virB11was detected in eight isolates, all from organic pigs. This gene has also been detected in 28% of the C. jejuni strains isolated from free-range broiler flocks (Hanning et al., 2010) and with a low prevalence for C. coli from various origins (Wieczorek and Osek, 2010, 2013; Acik et al., 2013). The low frequency of these isolates did not allow us to conclude that their presence in organic farms is related to the management of this type of production system.
There was no difference in the pathogenicity between organic and conventional C. coli pig isolates when tested on Caco-2 human intestinal cells. Moreover, different levels of pathogenicity were observed for the ST854 isolates, the most prevalent ST, regardless of the production system. Our C. coli isolates have low adhesion and invasion capacities, similar to C. coli from poultry and humans, compared with C. jejuni strains from poultry and humans. Guyard-Nicodème et al. (2013) reported similar results between the two species in a comparison of C. jejuni and C. coli strains isolated from poultry.
We showed that the isolates with the plasmid gene virB11 had a higher invasion capacity than isolates without this gene. Moreover, the human C. jejuni 81–176 strain carrying the plasmid also showed higher invasion capacity than the human C. jejuni NCTC 11168, which does not possess the virB11 gene. This relationship between high invasion and presence of the plasmid gene virB11 has previously been reported (see Bacon et al., 2002).
Conclusion
Our study could not conclusively demonstrate that the type of pig production system influences the C. coli population. We confirmed the high genetic diversity of C. coli in pigs in France, and showed that isolates sharing the same ST may show different levels of pathogenicity. This study helped improve the detection of virulence genes in C. coli, a species less studied than C. jejuni, and provided data on the virulence of this species, and more particularly of C. coli isolated from pigs.
Author Contributions
MD: Conception of the study, analysis and interpretation of data, and drafting of the manuscript; BN, VR, KB, MC, AK: Acquisition of the isolates and data; MD, AK: Critical revision of important intellectual content; All authors: Final approval of the version to be published and accountable for all aspects of the work, ensuring that questions related to the accuracy or integrity of any part of the work were appropriately investigated and resolved.
Funding
The sampling and isolation of isolates were conducted with the financial support of the CORE Organic II Funding Bodies, partners of the FP7 ERA-NET project, CORE Organic II (Coordination of European Transnational Research in Organic Food and Farming systems, project no. 249667). For further information, see www.coreorganic2.org. Characterization of the isolates was conducted with the financial support of FranceAgrimer (Contract number: 2013-0504).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank the manager of the slaughterhouse.
References
Abley, M. J., Wittum, T. E., Funk, J. A., and Gebreyes, W. A. (2012). Antimicrobial susceptibility, pulsed-field gel electrophoresis, and multi-locus sequence typing of Campylobacter coli in swine before, during, and after the slaughter process. Foodborne Pathog. Dis. 9, 506–512. doi: 10.1089/fpd.2011.1053
Acik, M. N., Karahan, M., Ongor, H., and Cetinkaya, B. (2013). Investigation of virulence and cytolethal distending toxin genes in Campylobacter spp. isolated from sheep in Turkey. Foodborne Pathog. Dis. 10, 589–594. doi: 10.1089/fpd.2012.1447
Alter, T., Gaull, F., Kasimir, S., Gurtler, M., Mielke, H., Linnebur, M., et al. (2005). Prevalences and transmission routes of Campylobacter spp. strains within multiple pig farms. Vet. Microbiol. 108, 251–261. doi: 10.1016/j.vetmic.2005.03.004
Andrzejewska, M., Klawe, J. J., Szczepańska, B., and Spica, D. (2011). Occurrence of virulence genes among Campylobacter jejuni and Campylobacter coli isolates from domestic animals and children. Pol. J. Vet. Sci. 14, 207–211. doi: 10.2478/v10181-011-0031-x
Bacon, D. J., Alm, R. A., Hu, L., Hickey, T. E., Ewing, C. P., Batchelor, R. A., et al. (2002). DNA sequence and mutational analyses of the pVir plasmid of Campylobacter jejuni 81-176. Infect. Immun. 70, 6242–6250. doi: 10.1128/IAI.70.11.6242-6250.2002
Bang, D. D., Borck, B., Nielsen, E. M., Scheutz, F., Pedersen, K., and Madsen, M. (2004). Detection of seven virulence and toxin genes of Campylobacter jejuni isolates from Danish turkeys by PCR and cytolethal distending toxin production of the isolates. J. Food Prot. 67, 2171–2177. doi: 10.4315/0362-028X-67.10.2171
Bang, D. D., Nielsen, E. M., Scheutz, F., Pedersen, K., Handberg, K., and Madsen, M. (2003). PCR detection of seven virulence and toxin genes of Campylobacter jejuni and Campylobacter coli isolates from Danish pigs and cattle and cytolethal distending toxin production of the isolates. J. Appl. Microbiol. 94, 1003–1014. doi: 10.1046/j.1365-2672.2003.01926.x
Boes, J., Nersting, L., Nielsen, E. M., Kranker, S., Enoe, C., Wachmann, H. C., et al. (2005). Prevalence and diversity of Campylobacter jejuni in pig herds on farms with and without cattle or poultry. J. Food Prot. 68, 722–727. doi: 10.4315/0362-028X-68.4.722
Carvalho, A. C., Ruiz-Palacios, G. M., Ramos-Cervantes, P., Cervantes, L. E., Jiang, X., and Pickering, L. K. (2001). Molecular characterization of invasive and noninvasive Campylobacter jejuni and Campylobacter coli isolates. J. Clin. Microbiol. 39, 1353–1359. doi: 10.1128/JCM.39.4.1353-1359.2001
Datta, S., Niwa, H., and Itoh, K. (2003). Prevalence of 11 pathogenic genes of Campylobacter jejuni by PCR in strains isolated from humans, poultry meat and broiler and bovine faeces. J. Med. Microbiol. 52, 345–348. doi: 10.1099/jmm.0.05056-0
Denis, M., Henrique, E., Chidaine, B., Tircot, A., Bougeard, S., and Fravalo, P. (2011). Campylobacter from sows in farrow-to-finish pig farms: risk indicators and genetic diversity. Vet. Microbiol. 154, 163–170. doi: 10.1016/j.vetmic.2011.07.001
Denis, M., Rose, V., Huneau-Salaün, A., Balaine, L., and Salvat, G. (2008). Diversity of pulsed-field gel electrophoresis profiles of Campylobacter jejuni and Campylobacter coli from broiler chickens in france. Poult. Sci. 87, 1662–1671. doi: 10.3382/ps.2008-00010
Denis, M., Soumet, C., Rivoal, K., Ermel, G., Blivet, D., Salvat, G., et al. (1999). Development of a m-PCR assay for simultaneous identification of Campylobacter jejuni and C. coli. Lett. Appl. Microbiol. 29, 406–410. doi: 10.1046/j.1472-765X.1999.00658.x
Dingle, K. E., Colles, F. M., Ure, R., Wagenaar, J. A., Duim, B., Bolton, F. J., et al. (2002). Molecular characterization of Campylobacter jejuni clones: a basis for epidemiologic investigation. Emerg. Infect. Dis. 8, 949–955. doi: 10.3201/eid0809.02-0122
Dingle, K. E., Colles, F. M., Wareing, D. R., Ure, R., Fox, A. J., Bolton, F. E., et al. (2001). Multilocus sequence typing system for Campylobacter jejuni. J. Clin. Microbiol. 39, 14–23. doi: 10.1128/JCM.39.1.14-23.2001
Egger, R., Korczak, B. M., Niederer, L., Overesch, G., and Kuhnert, P. (2012). Genotypes and antibiotic resistance of Campylobacter coli in fattening pigs. Vet. Microbiol. 155, 272–278. doi: 10.1016/j.vetmic.2011.08.012
Feodoroff, B., Ellstrom, P., Hyytiainen, H., Sarna, S., and Hanninen, M. L. and Rautelin H. (2010). Campylobacter jejuni isolates in Finnish patients differ according to the origin of infection. Gut Pathog. 2:22. doi: 10.1186/1757-4749-2-22
Garcia-Migura, L., Hendriksen, R. S., Fraile, L., and Aarestrup, F. M. (2014). Antimicrobial resistance of zoonotic and commensal bacteria in Europe: the missing link between consumption and resistance in veterinary medicine. Vet. Microbiol. 170, 1–9. doi: 10.1016/j.vetmic.2014.01.013
Gonzalez, I., Grant, K. A., Richardson, P. T., Park, S. F., and Collins, M. D. (1997). Specific identification of the enteropathogens Campylobacter jejuni and Campylobacter coli by using a PCR test based on the ceuE gene encoding a putative virulence determinant. J. Clin. Microbiol. 35, 759–763.
Greig, J., Rajić, A., Young, I., Mascarenhas, M., Waddell, L., and LeJeune, J. (2015). A scoping review of the role of wildlife in the transmission of bacterial pathogens and antimicrobial resistance to the food chain. Zoonoses Public Health 62, 269–284. doi: 10.1111/zph.12147
Grundmann, H. S., Hori, S., and Tanner, G. (2001). Determining confidence intervals when measuring genetic diversity and the discriminatory abilities of typing methods for microorganisms. J. Clin. Microbiol. 39, 4190–4192. doi: 10.1128/JCM.39.11.4190-4192.2001
Guyard-Nicodème, M., Tresse, O., Houard, E., Jugiau, E., Courtillon, C., El Manaa, K., et al. (2013). Characterization of Campylobacter spp. transferred from naturally contaminated chicken legs to cooked chicken slices via a cutting board. Int. J. Food Microbiol. 164, 7–14. doi: 10.1016/j.ijfoodmicro.2013.03.009
Hanning, I., Biswas, D., Herrera, P., Roesler, M., and Ricke, S. C. (2010). Prevalence and characterization of Campylobacter jejuni isolated from pasture flock poultry. J. Food Sci. 75, M496–M502. doi: 10.1111/j.1750-3841.2010.01747.x
Hunter, P. R. (1990). Reproducibility and indices of discriminatory power of microbial typing methods. J. Clin. Microbiol. 28, 1903–1905.
Jensen, A. N., Andersen, M. T., Dalsgaard, A., Baggesen, D. L., and Nielsen, E. M. (2005). Development of real-time PCR and hybridization methods for detection and identification of thermophilic Campylobacter spp. in pig faecal samples. J. Appl. Microbiol. 99, 292–300. doi: 10.1111/j.1365-2672.2005.02616.x
Jensen, A. N., Dalsgaard, A., Baggesen, D. L., and Nielsen, E. M. (2006). The occurrence and characterization of Campylobacter jejuni and C. coli in organic pigs and their outdoor environment. Vet. Microbiol. 16, 96–105. doi: 10.1016/j.vetmic.2006.03.006
Kempf, I., Kerouanton, A., Bougeard, S., Nagard, B., Rose, V., Mourand, G., et al. (2017). Campylobacter coli in organic and conventional pig production in France and Sweden: prevalence and antimicrobial resistance. Front. Microbiol. 8:955. doi: 10.3389/fmicb.2017.00955
Khoshbakht, R., Tabatabaei, M., Hosseinzadeh, S., Shekarforoush, S. S., and Aski, H. S. (2013). Distribution of nine virulence-associated genes in Campylobacter jejuni and C. coli isolated from broiler feces in Shiraz, Southern Iran. Foodborne Pathog. Dis. 10, 764–770. doi: 10.1089/fpd.2013.1489
Konkel, M. E., Gray, S. A., Kim, B. J., Garvis, S. G., and Yoon, J. (1999). Identification of the enteropathogens Campylobacter jejuni and Campylobacter coli based on the cadF virulence gene and its product. J. Clin. Microbiol. 37, 510–517.
Leblanc Maridor, M., Denis, M., Rosero, A., Federighi, M., Seegers, H., and Belloc, C. (2011). Evaluation of genomic instability of Campylobacter spp. strains during in vitro assay and in vivo passage in pigs. Vet. Microbiol. 154, 171–179.
Madden, R. H., Moran, L., and Scates, P. (2000). Optimising recovery of Campylobacter spp. from the lower porcine gastrointestinal tract. J. Microbiol. Meth. 42, 115–119. doi: 10.1016/S0167-7012(00)00183-4
Miller, W. G., On, S. L., Wang, G., Fontanoz, S., Lastovica, A. J., and Mandrell, R. E. (2005). Extended multilocus sequence typing system for Campylobacter coli, C. lari, C. upsaliensis, and C. helveticus. J. Clin. Microbiol. 43, 2315–2329. doi: 10.1128/JCM.43.5.2315-2329.2005
Moore, J. E., Lanser, J., Heuzenroeder, M., Ratcliff, R. M., Millar, B. C., and Madden, R. H. (2002). Molecular diversity of Campylobacter coli and C. jejuni isolated from pigs at slaughter by flaA-RFLP analysis and ribotyping. J. Vet. Med. Ser. B 49, 388–393. doi: 10.1046/j.1439-0450.2002.00595.x
Nielsen, E. M., Engberg, J., and Madsen, M. (1997). Distribution of serotypes of Campylobacter jejuni and C. coli from Danish patients, poultry, cattle and swine. FEMS Immunol. Med. Microbiol. 19, 47–56. doi: 10.1111/j.1574-695X.1997.tb01071.x
Österberg, J., Wingstrand, A., Nygaard Jensen, A., Kerouanton, A., Cibin, V., Barco, L., et al. (2016). Antibiotic resistance in Escherichia coli from pigs in organic and conventional farming in four European countries. PLoS ONE 11:e0157049. doi: 10.1371/journal.pone.0157049
Quintana-Hayashi, M. P., and Thakur, S. (2012). Phylogenetic analysis reveals common antimicrobial resistant Campylobacter coli population in antimicrobial-free (ABF) and commercial swine systems. PLoS ONE 7:e44662. doi: 10.1371/journal.pone.0044662
Rivoal, K., Ragimbeau, C., Salvat, G., Colin, P., and Ermel, G. (2005). Genomic diversity of Campylobacter coli and Campylobacter jejuni isolates recovered from free-range broiler farms and comparison with isolates of various origins. Appl. Environ. Microbiol. 71, 6216–6227. doi: 10.1128/AEM.71.10.6216-6227.2005
Rozynek, E., Dzierzanowska-Fangrat, K., Jozwiak, P., Popowski, J., Korsak, D., and Dzierzanowska, D. (2005). Prevalence of potential virulence markers in polish Campylobacter jejuni and Campylobacter coli isolates obtained from hospitalized children and from chicken carcasses. J. Med. Microbiol. 54, 615–619. doi: 10.1099/jmm.0.45988-0
Saini, V., McClure, J. T., Scholl, D. T., De Vries, T. J., and Barkema, H. W. (2013). Herd-level relationship between antimicrobial use and presence or absence of antimicrobial resistance in gram-negative bovine mastitis pathogens on Canadian dairy farms. J. Dairy Sci. 96, 4965–4976. doi: 10.3168/jds.2012-5713
Struelens, M. J. (1996). Consensus guidelines for appropriate use and evaluation of microbial epidemiologic typing systems. Clin. Microbiol. Infect. 2, 2–11. doi: 10.1111/j.1469-0691.1996.tb00193.x
Thakur, S., Morrow, W. E., Funk, J. A., Bahnson, P. B., and Gebreyes, W. A. (2006). Molecular epidemiologic investigation of Campylobacter coli in swine production systems, using multilocus sequence typing. Appl. Environ. Microbiol. 72, 5666–5669. doi: 10.1128/AEM.00658-06
von Altrock, A., Hamedyb, A., Merlec, R., and Waldmanna, K. H. (2013). Campylobacter spp. – prevalence on pig livers and antimicrobial Susceptibility. Prev. Vet. Med. 109, 152–157. doi: 10.1016/j.prevetmed.2012.09.010
Wieczorek, K., and Osek, J. (2010). Simultaneous occurrence of selected food-borne bacterial pathogens on bovine hides, carcasses and beef meat. Pol. J. Vet. Sci. 13, 645–651. doi: 10.2478/v10181-010-0001-8
Wieczorek, K., and Osek, J. (2013). Characteristics and antimicrobial resistance of Campylobacter isolated from pig and cattle carcasses in Poland. Pol. J. Vet. Sci. 16, 501–508. doi: 10.2478/pjvs-2013-0070
Keywords: Campylobacter, pig, organic, PFGE, MLST, virulence
Citation: Denis M, Nagard B, Rose V, Bourgoin K, Cutimbo M and Kerouanton A (2017) No Clear Differences between Organic or Conventional Pig Farms in the Genetic Diversity or Virulence of Campylobacter coli Isolates. Front. Microbiol. 8:1016. doi: 10.3389/fmicb.2017.01016
Received: 20 January 2017; Accepted: 22 May 2017;
Published: 26 June 2017.
Edited by:
Avelino Alvarez-Ordóñez, Universidad de León, SpainReviewed by:
Héctor Argüello, University of Córdoba, ColombiaJordi Rovira, University of Burgos, Spain
Copyright © 2017 Denis, Nagard, Rose, Bourgoin, Cutimbo and Kerouanton. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martine Denis, bWFydGluZS5kZW5pc0BhbnNlcy5mcg==