 Teresa Pires dos Santos
Teresa Pires dos Santos Peter Damborg
Peter Damborg Arshnee Moodley1
Arshnee Moodley1- 1Department of Veterinary Disease Biology, Faculty of Health and Medical Sciences, University of Copenhagen, Frederiksberg, Denmark
- 2Department of Biomedical Sciences, Ross University School of Veterinary Medicine, St Kitts, West Indies
Background and rationale: Methicillin-resistant Staphylococcus pseudintermedius (MRSP) is a major cause of infections in dogs, also posing a zoonotic risk to humans. This systematic review aimed to determine the global epidemiology of MRSP and provide new insights into the population structure of this important veterinary pathogen.
Methodology: Web of Science was searched systematically for articles reporting data on multilocus sequence typing (MLST) of S. pseudintermedius isolates from dogs or other animal or human patients and carriers. Data from the eligible studies were then integrated with data from the MLST database for this species. Analysis of MLST data was performed with eBURST and ClonalFrame, and the proportion of MRSP isolates resistant to selected antimicrobial drugs was determined for the most predominant clonal complexes.
Results: Fifty-eight studies published over the last 10 years were included in the review. MRSP represented 76% of the 1428 isolates characterized by the current MLST scheme. The population of S. pseudintermedius was highly diverse and included five major MRSP clonal complexes (CCs). CC71, previously described as the epidemic European clone, is now widespread worldwide. In Europe, CC258, which is more frequently susceptible to enrofloxacin and aminoglycosides, and more frequently resistant to sulphonamides/trimethoprim than CC71, is increasingly reported in various countries. CC68, previously described as the epidemic North American clone, is frequently reported in this region but also in Europe, while CC45 (associated with chloramphenicol resistance) and CC112 are prevalent in Asia. It was estimated that clonal diversification in this species is primarily driven by homologous recombination (r/m = 7.52).
Conclusion: This study provides evidence that S. pseudintermedius has an epidemic population structure, in which five successful MRSP lineages with specific traits regarding antimicrobial resistance, genetic diversity and geographical distribution have emerged upon a weakly clonal background through acquisition of SCCmec and other mobile genetic elements.
Introduction
Staphylococcus pseudintermedius is a bacterial commensal of the skin and mucosae of dogs and also the most prevalent cause of canine bacterial infections (Bannoehr and Guardabassi, 2012). Although primarily adapted to the canine host, this coagulase-positive staphylococcal species may infect other pet animals, mainly cats (Kadlec et al., 2010), and humans (Van Hoovels et al., 2006). Over the last decade, multidrug-resistant strains have emerged worldwide (Black et al., 2009; Moodley et al., 2009; Kadlec et al., 2010; Perreten et al., 2010, 2013; Ruscher et al., 2010; Gómez-Sanz et al., 2011; Laarhoven et al., 2011; Osland et al., 2012; Bardiau et al., 2013; Chanchaithong et al., 2014; Couto et al., 2014; Davis et al., 2014; Savini et al., 2014; Starlander et al., 2014; Grönthal et al., 2015; Kjellman et al., 2015; Rota et al., 2015; Ishihara et al., 2016), especially clones that have acquired the Staphylococcal Chromosomal Cassette (SCCmec) mobilizing the methicillin resistance gene mecA. Infections caused by methicillin-resistant S. pseudintermedius (MRSP) can be difficult or even impossible to treat using veterinary licensed systemic antimicrobial agents. The proportion of MRSP amongst clinical isolates varies considerably depending on the geographic region and population studied (Norström et al., 2009; Kawakami et al., 2010; De Lucia et al., 2011; Garbacz et al., 2011; Youn et al., 2011; Feng et al., 2012; Aslantaş et al., 2013; Detwiler et al., 2013; Penna et al., 2013; Chanchaithong et al., 2014; Haenni et al., 2014; Lehner et al., 2014; Windahl et al., 2015; Ishihara et al., 2016). MRSP isolation frequencies may reach up to 67% of all clinical S. pseudintermedius isolates in certain countries and veterinary hospitals (Kawakami et al., 2010). Hospitalization, frequent visits to veterinary practices, and prior antimicrobial usage are recognized risk factors for canine MRSP infection and carriage (Frank et al., 2009; Rota et al., 2011; Nienhoff et al., 2011a,b; Weese et al., 2012; Windahl et al., 2012; Eckholm et al., 2013; Lehner et al., 2014; Grönthal et al., 2015).
Multilocus sequence typing (MLST) has proven to be an invaluable DNA sequence-based technique for analysis of population structure and long-term epidemiological trends at a global level (Spratt and Maiden, 1999). A first MLST scheme based on five loci was initially proposed in 2007 to provide insight into the overall population genetic structure of the Staphylococcus intermedius Group (SIG), which includes S. intermedius and S. delphini in addition to S. pseudintermedius (Bannoehr et al., 2007). Using this scheme two major epidemic MRSP clones were detected, ST68 in North America and ST71 in Europe (Perreten et al., 2010). The first species-specific MLST scheme was launched in 2013 (Solyman et al., 2013), and its publicly available database (http://pubmlst.org/spseudintermedius/) contains records of 503 sequence types (STs) at present. Since 2007, numerous scientific papers have reported MLST data on MRSP carriage and infection in a variety of countries and continents, but to date these studies have not been reviewed systematically.
The aim of this systematic review was to provide an updated overview of the global epidemiology and evolution of MRSP, with a focus on geographical distribution of reported frequency, SCCmec content and antimicrobial resistance of the main clonal lineages, as defined by MLST. The results of the review were also used to infer the population structure of the species, including methicillin-susceptible S. pseudintermedius (MSSP), as well as to discuss the evolutionary mechanisms that could be implicated in the diversification of MRSP clones.
Methodology
Search Strategy and Study Selection Criteria
We searched the Web of Science database for articles published between 28th September 2007 and 31st May 2016 using the search term “Staphylococcus pseudintermedius.” This resulted in the retrieval of 403 articles, of which 239 reported data on MRSP. Studies were then selected if they reported MLST data on MRSP and/or MSSP isolates from screening and/or clinical samples, including data obtained by the 5-allele (Bannoehr et al., 2007; MLST-5) or the 7-allele (Solyman et al., 2013; MLST-7) scheme. Studies that exclusively used dru typing (n = 5) to characterize S. pseudintermedius strains were not included. These steps yielded 58 studies (Figure 1), including 49 studies on MRSP (with or without MSSP; Bannoehr et al., 2007; Descloux et al., 2008; Black et al., 2009, 2011; Moodley et al., 2009, 2013; Kadlec et al., 2010, 2015, 2016; Perreten et al., 2010, 2013; Ruscher et al., 2010; Stegmann et al., 2010; Boost et al., 2011; Gómez-Sanz et al., 2011, 2013a,b, 2014; Laarhoven et al., 2011; Paul et al., 2011; DiCicco et al., 2012, 2014; Feng et al., 2012; Osland et al., 2012; Walther et al., 2012; Wang et al., 2012; Bardiau et al., 2013; Haenni et al., 2013, 2014; Pilla et al., 2013; Quitoco et al., 2013; Solyman et al., 2013; Youn et al., 2013; Chanchaithong et al., 2014, 2015; Couto et al., 2014, 2016; Davis et al., 2014; Grönthal et al., 2014, 2015; Savini et al., 2014; Starlander et al., 2014; Börjesson et al., 2015; Kjellman et al., 2015; McCarthy et al., 2015; Rota et al., 2015; Damborg et al., 2016; Duim et al., 2016; Ishihara et al., 2016) and 9 studies on MSSP only (Ben Zakour et al., 2011; Tse et al., 2011; Gharsa et al., 2013, 2015; Matanovic et al., 2013; Paul et al., 2013; Gómez-Sanz et al., 2013c; Youn et al., 2014; Lozano et al., 2015).
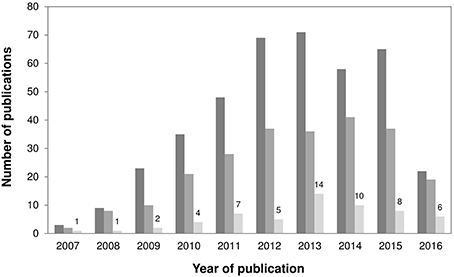
Figure 1. Bar chart showing (i) the annual number of published articles retrieved when using the search word “Staphylococcus pseudintermedius” (dark gray); (ii) the annual number of published articles retrieved involving MRSP (intermediate gray); and (iii) the annual number of published articles retrieved employing MLST (light gray), since 2007–May 2016. A total of 58 studies reporting MLST data on MRSP and MSSP were included in this review.
Data Extraction and Review Approach
A database was created to record location, date of isolation, source, host, resistance phenotype, resistance genotype, and ST of each isolate. Susceptibility data were based on the interpretation criteria used in the original articles, and isolates reported as “intermediate” were considered “susceptible.” Only one isolate per animal was included, unless multiple STs were reported in the same animal. When two isolates from the same animal belonged to the same clone but differed in antimicrobial susceptibility patterns, only the isolate displaying resistance to most drug classes was considered. The information collected in the scientific literature was integrated with that available in the S. pseudintermedius MLST database (26 June 2015), incorporated into an Excel file and summarized in Table S1. Stringent cross-checking of strain information in the database and in the papers was performed to ensure that strains were only accounted for once. Statistical comparisons of the relative prevalences of resistance were based on the chi-square test of independence or, for small numbers, on the Fisher's exact.
Bioinformatic Analysis
eBURST V3 (Feil et al., 2004) (http://eburst.mlst.net) was used to infer the patterns of evolutionary descent from MLST allelic data. The program settings were adjusted to define clonal complexes (CCs) as groups of sequence types (STs) sharing at least six identical alleles, with the primary founder being the ST having the most number of single locus variants (SLVs). Subgroup founders were defined according to the default settings of eBURST, i.e., as STs with at least three SLVs that diversified from a primary founder. A bootstrapping resampling procedure (1000 samples) was used to assess the statistical confidence in the primary founders.
Validation of ST memberships in various CCs was done with phylogenetic analysis based on the nucleotide sequences of the S. pseudintermedius MLST genes (ack, cpn60, fdh, pta, purA, sar, and tuf; Solyman et al., 2013). A total of 144 STs representing major eBURST group/subgroup founders (with more than three SLV links) and singletons with no double locus variant (DLV) links to eBURST-assigned CCs (Willems et al., 2011) were used in the analysis. Two independent ClonalFrame (Didelot and Falush, 2007) runs consisting of 2,000,000 MCMC iterations were performed, the first half of which was discarded as burn-in. The Gelman-Rubin statistic was calculated to assess convergence of the two runs (< 1.1) (Vos and Didelot, 2009), and the genealogy was built as a 50% majority-rule consensus tree. The Bayesian posterior probability of a given cluster (i.e., the proportion of trees supporting a given branch) was used to evaluate the confidence level in the assigned clusters; clusters with confidence levels >0.90 were considered well supported.
The rates at which recombination (rho, ρ) and mutation (theta, θ) occur were extracted from one ClonalFrame output file to assess how frequent recombination events occur relative to mutation events (ρ/θ). The ratio of probabilities that a given site is altered through recombination and mutation (r/m) was also estimated to determine how important the effect of recombination has been to clonal diversification relative to mutation. The program MultiLocus (Agapow and Burt, 2001) (version 1.2, Imperial College, available at http://www.bio.ic.ac.uk/evolve/software/multilocus) was used to test for significant linkage disequilibrium among the MLST loci (1000 randomizations were used for all P-value estimates).
Genotypic (ST) diversity (G), i.e., the probability that two randomly picked isolates belong to a different ST; genetic (allelic) diversity of a locus (h), i.e., the probability that two randomly picked STs have a different allele at a locus; and mean allelic diversity over the seven loci (H) were calculated with the software Arlequin V3.1 (Excoffier et al., 2005). Analysis of molecular variance (AMOVA) as a weighted average over loci was carried out to determine the degree of genetic variation between subpopulations (MRSP and MSSP). Descriptive statistics for MLST data including the number of variable sites/codons, nucleotide diversity and estimates of dS and dN (Tamura-Nei model) were performed using MEGA V7.0.14 (Kumar et al., 2016).
Results
Sources of S. pseudintermedius Isolates
MRSP represented 1087 (76.1%) of all 1428 S. pseudintermedius isolates that were characterized by MLST-7, and 558 (76.1%) of all 733 S. pseudintermedius isolates that were characterized according to MLST-5. Approximately 14.3% of MRSP isolates (n = 155) were characterized according to both schemes. Clinical isolates accounted for 81.8% of the isolates analyzed by MLST-7 for which an isolation source could be identified (n = 892). These were mainly isolated from skin samples associated to pyoderma, surgical site- and wound infections (64.1%). MRSP accounted for 80.5% of all 892 isolates, for 86.2% of clinical isolates and for 54.9% of carriage isolates. Most (90.5%) isolates of known source originated from dogs (n = 804, of which 83.3% were MRSP), while the remaining isolates were recovered from humans (n = 36, 52.8% of which were MRSP), cats (n = 26, 73.1% of which were MRSP) and horses (n = 3, of which one was MRSP). Among the 36 isolates from human origin, 22 were from clinical specimens (31.8% of which were MRSP) while 14 were from screening samples (85.7% of which were MRSP) (Table S1).
Nucleotide Sequence Variation at Each MLST Locus
A high genotypic diversity (G = 0.8720; SD = 0.0082) was detected among the 1428 isolates characterized by MLST-7. Among the 503 existing STs, 431 were reported only once, while 72 were reported multiple times. Sequence comparisons amongst all STs revealed an average of 18.4 variable nucleotide sites (range = 7–29) and 3.4 variable codons (range = 1–8) per locus (Table 1). The average number of alleles at the seven housekeeping genes was 25.4, ranging from 8 alleles for tuf to 44 for cpn60 and purA. The average single-locus genetic diversity (H) was high for the total population (H = 0.710; SD = 0.060). The genes cpn60, fdh, and purA showed higher nucleotide diversity (π = 0.011–0.058) than ack, pta, sar, and tuf (π = 0.005–0.008). There was a low percentage of nonsynonymous (amino-acid changing) substitutions for all seven genes (dS/dN = 3.557–35.090) (Table 1), indicating that natural selection acting on housekeeping genes is negative (dS/dN > 1, i.e., purifying selection), as it is expected for genes encoding essential cellular functions.
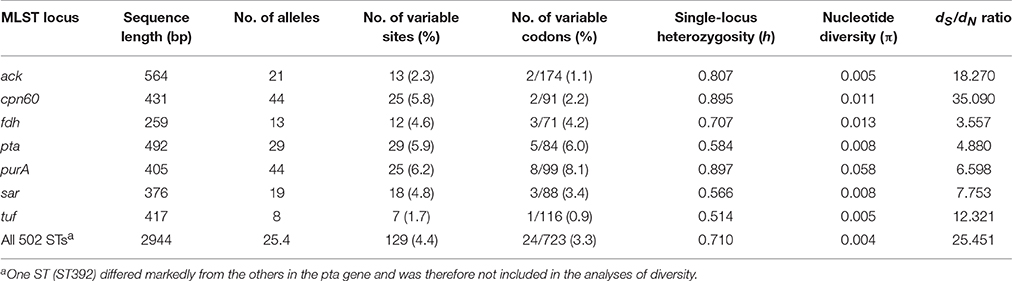
Table 1. Nucleotide variation within seven housekeeping genes analyzed by MLST.
Distribution of MRSP and MSSP Isolates
The obtained data included 335 MSSP isolates that belonged to 328 STs, and 1087 MRSP isolates that belonged to 182 STs. Genotypic diversity was higher among MSSP (G = 0.9999; SD = 0.0002) than among MRSP (G = 0.7791; SD = 0.0126). Sequence type 71 was the most predominant ST amongst MRSP (44.9%), followed by ST45 (11.0%), ST258 (6.9%), ST261 (3.5%), ST112 (1.9%), ST265 (1.7%), ST68 (1.5%), ST169, and ST181 (1.4%), which were reported ≥15 times and represented 74.2% of all 1087 MRSP isolates. Only seven STs were common to both MRSP and MSSP isolates (ST17, ST29, ST56, ST123, ST125, ST155, and ST170). AMOVA showed a small but significant genetic variation (2.31%) between MRSP and MSSP subpopulations (P < 0.05), indicating that they are slightly genetically distinct.
Identification of Main Clonal Lineages
The 503 STs of S. pseudintermedius were assigned by eBURST into two major (CC71 and CC258) and two minor CCs (CC379 and CC75), 30 groups of STs with no predicted founder (or a founder that was not well supported; bootstrapping < 50%), and 231 singletons (Figure 2). While CC258 and CC379 comprised both MRSP and MSSP isolates, CC71 only comprised MRSP and CC75 only comprised MSSP. CC71 was largely dominated by ST71 and included 15 additional STs (12 SLVs, 2 DLVs, and 1 TLV). In contrast, CC258 formed a large straggly cluster accounting for 34.4% (n = 173) of all STs and 46.7% (n = 85) of the STs associated with MRSP. The remaining MRSP isolates were widely distributed throughout the population of S. pseudintermedius in a wide variety of different lineages (Figure 2, in pink).
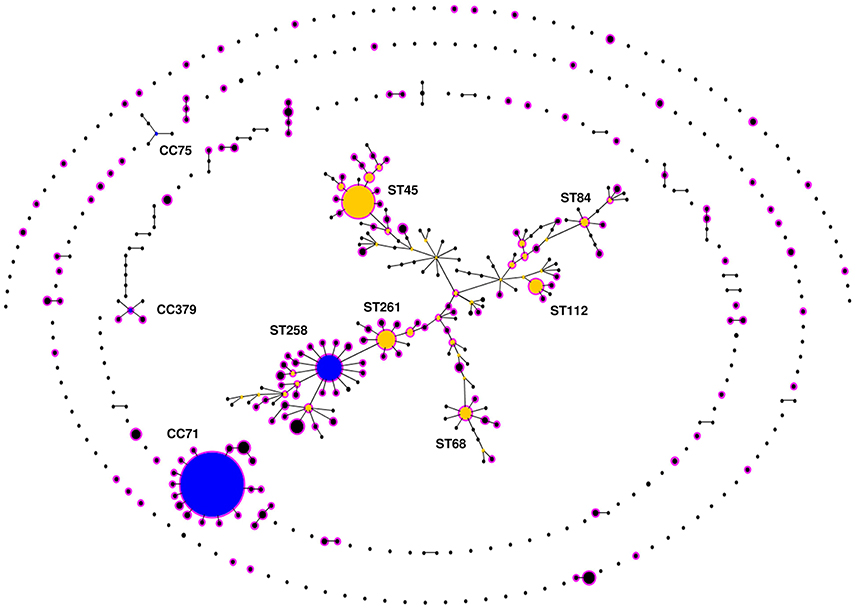
Figure 2. eBURST population snapshot of Staphylococcus pseudintermedius. Clusters of linked isolates correspond to clonal complexes (CCs). Primary founders (blue) are positioned centrally in the clusters, and subgroup founders are shown in yellow. The diameter of the circles is directly proportional to the total number of isolates reported in the MLST database (http://pubmlst.org/spseudintermedius/) and in reviewed scientific papers given in Table S1. Names of various important STs associated with methicillin resistance are indicated within a large, “straggly” CC, CC258. Methicillin resistant S. pseudintermedius (MRSP) lineages are highlighted in pink.
Phylogenetic Analysis
Phylogenetic analysis of the concatenated sequences of the seven MLST loci from 144 STs using ClonalFrame showed that MRSP were associated to a variety of different STs and often clustered with MSSP (Figure 3). However, the major group/subgroup founders (e.g., ST45, ST68, ST112, and ST258) that were linked together using eBURST (Figure 2) appeared as distinct phylogenetic lineages that do not share a recent common ancestor (Figure 3). Therefore, in our subsequent analyses, we used arbitrary CC designations (e.g., CC45, CC68, CC112, and CC258) that include the founder STs confirmed by ClonalFrame to represent phylogenetically distinct lineages, and their respective single-, double-, and triple locus variants (SLV, DLV, and TLV), as defined by eBURST.
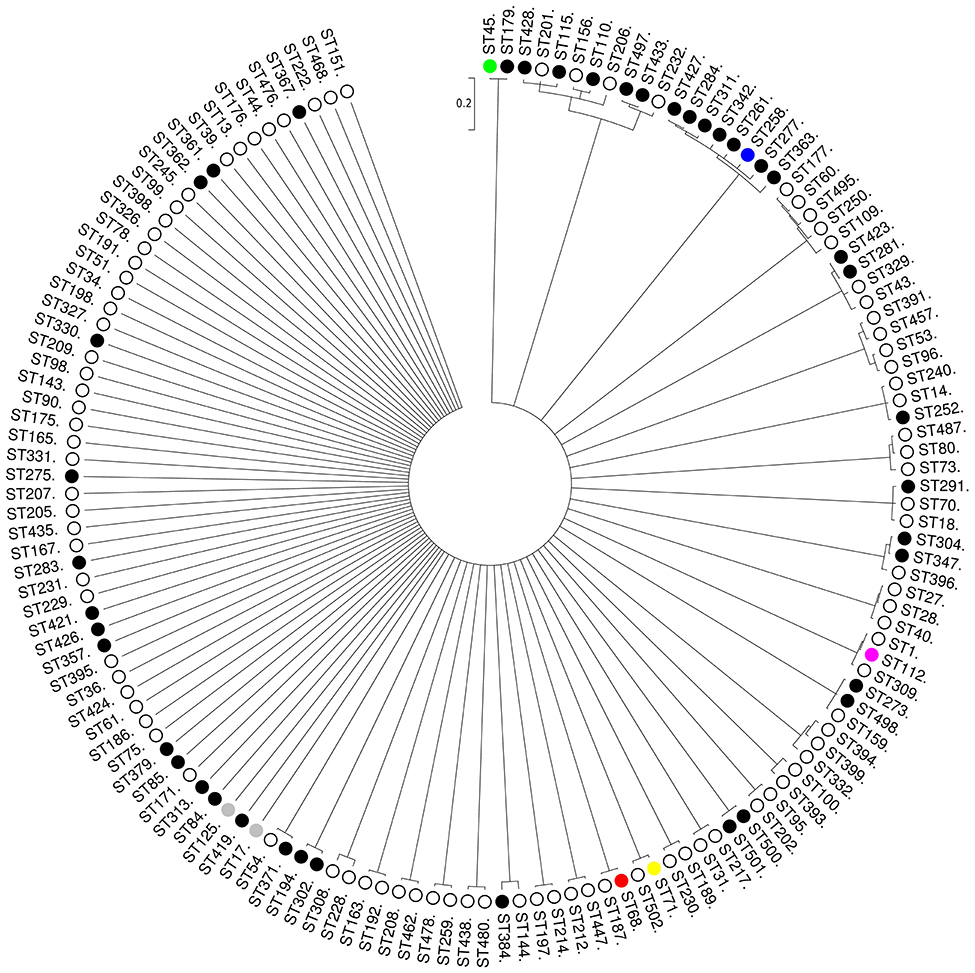
Figure 3. Unrooted consensus tree displaying the relationship between the 144 STs of the S. pseudintermedius sample (96 MSSP, 46 MRSP, and 2 MSSP/MRSP) at seven concatenated loci (2944 nucleotides in total). The 144 STs represent major eBURST group/subgroup founders (with more than three SLV links) and singletons with no double locus variant (DLV) links to eBURST-assigned CCs. The phylogenetic tree was based on all trees sampled after 2,000,000 iterations, from which half were discarded as burn-in. The dendogram was constructed from the combination of two ClonalFrame runs with a cut-off value of 0.5 as a majority rule consensus. Scale is in coalescent units. There were no well-supported nodes (i.e., with a Bayesian posterior probability > 0.90) for clusters with > 3 STs). The main MRSP clones are shown in different colors. MRSP, black; MSSP, white; both MRSP and MSSP, gray.
The clustering performed by eBURST and a phylogenetic tree built with all 503 STs (Figure S1) were in general agreement for CC71, i.e., 10 of the 16 STs from this CC clustered with high confidence (1.00) with ClonalFrame. Likewise, eight of the 15 STs from CC68 were clustered with high confidence (0.99) in the ClonalFrame phylogenetic tree, together with three eBURST singletons that were DLVs of STs belonging to CC68 (Figure S1). In contrast to CC71 and CC68 though, CC258 lineages appeared as part of a bundle of STs within a star-shaped genealogy (503 STs) or, for the analyzed 144 STs, in a node that was not well supported (0.67) (Figure 3). In the case of CC258, only eBURST could provide a hypothesis about the way this CC has emerged and diversified.
ClonalFrame analysis revealed that recombination occurred more frequently (ρ/θ = 3.68) and had a more important contribution to clonal diversification of S. pseudintermedius than mutation (r/m = 7.52). The standardized value of the index of association between the alleles, although much lower than 1, was still significantly different from 0 (rbard = −0.1218, P < 0.001), indicating that there is weak but significant linkage disequilibrium between the MLST loci, and ultimately that the S. pseudintermedius population is weakly clonal. Similar results were obtained when analyzing separately the 182 STs found amongst MRSP (rbard = 0.0312, P < 0.001), whereas there was no significant linkage disequilibrium between the MLST loci of the 328 STs detected amongst MSSP (rbard = −0.1521, P = 0.065) nor for the STs from highly supported clusters (e.g., CC71, CC68; Figure S1), a result that reinforces the recombinogenic nature of S. pseudintermedius.
Geographical Dissemination of the Main MRSP Lineages
Overall, 24 countries were represented in our dataset (Table S1). Twenty-five STs, 20 STs of which associated with MRSP, were recovered from multiple countries and continents. The majority of MRSP isolates were recovered from Europe (70.6%), followed by Asia (22.4%), North America (6.2%), Oceania (0.6%), and South America (0.2%). Among the 182 MRSP-associated STs, 16 were recovered from more than 1 continent (ST17, ST29, ST45, ST68, ST71, ST85, ST112, ST121, ST123, ST125, ST155, ST169, ST179, ST180, ST282, and ST261), and 34 were reported in more than 1 country. The most widely disseminated clone was ST71, which was isolated in 14 different countries across 3 continents (Europe, North and South America, and Asia). The other widespread MRSP lineages were distributed across two continents and included ST45 (7 countries); ST121 and ST261 (4 countries); ST68, ST169, ST265, and ST277 (3 countries); and ST17, ST29, ST85, ST112, ST123, ST125, ST155, ST179, ST180, and ST188 (2 countries). ST41, ST118, ST181, ST196, ST258, ST265, ST277, ST298, ST301, ST307, ST312, ST339, ST342, and ST414 were reported in multiple countries but one continent only. Certain MRSP clonal lineages were frequently reported in particular continents, namely CC71 and CC258 in Europe (93 and 98%, respectively); CC68 in North America (66.7%); and CC45 and CC112 in Asia (59.5 and 48.9%, respectively; Figure 4).
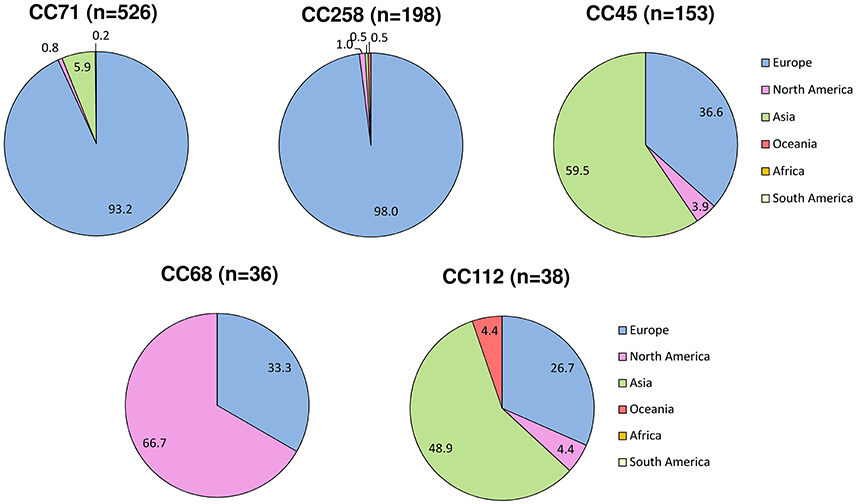
Figure 4. Geographic variation in the reported frequency of major MRSP clonal complexes based on data from the MLST database (http://pubmlst.org/spseudintermedius/) and references given in Table S1. The numbers represent the percentage (%) of MRSP isolates from each lineage that was reported in each continent.
Distribution of SCCmec among Main MRSP Clonal Lineages
The distribution of SCCmec types (when reported) varied significantly (P < 0.0001) across distinct MRSP clonal groups (Table S1). A total of five SCCmec types were reported: (i) SCCmec type II-III was significantly associated with CC71; (ii) type ψSCCmec57395 was significantly associated with CC45; (iii) SCCmecAI16-SCCczrAI16-CI was significantly associated with CC112 isolates; and (iv) SCCmec type IV was significantly associated with CC258. In contrast, SCCmec type V was present in CC68, CC45, and CC379, as well as in 21 STs unassigned to any CC, indicating both vertical and horizontal transfer of SCCmec into S. pseudintermedius of different genetic backgrounds. Based on the patterns of evolutionary descent defined by eBURST and available SCCmec typing data, we estimated that S. pseudintermedius has acquired SCCmec at least 48 times.
Antimicrobial Resistance Patterns of Main MRSP Clonal Lineages
The overall prevalence rates of resistance amongst MRSP isolates to selected non-β-lactam antimicrobials were: erythromycin, 93.5% (762/815); clindamycin, 92.7% (695/750); tetracycline, 70.7% (573/810); trimethoprim/sulfamethoxazole, 76.6% (490/640); enrofloxacin, 73.8% (454/615); gentamicin, 71.3% (574/805); chloramphenicol, 43.1% (304/706); and amikacin, 5.2% (21/404). The prevalence of antimicrobial resistance in the three most prevalent MRSP clonal groups (CC71, CC45, and CC258) are shown in Figure 5. CC258 showed significantly lower prevalence of resistance to gentamicin and enrofloxacin (13.8 and 5.6%, respectively) as compared to CC71 and CC45 (90.1–93.8 and 98.1–100.0%, respectively) (P < 0.0001), but significantly higher prevalence of resistance to trimethoprim/sulfamethoxazole (91.6%) as compared to CC71 (72.3%, P < 0.0001) and CC45 (78.8%, P = 0.012). Chloramphenicol resistance was significantly more prevalent in CC45 (95.2%) compared to CC71 (42.9%) and CC258 (12.9%) (P < 0.0001), and the prevalence of tetracycline resistance was significantly lower for CC71 isolates (51.1%) compared to CC258 and CC45 (91.1 and 96.9%, respectively) (P < 0.0001). MRSP CC71 was also significantly associated with carriage of the tetracycline resistance gene tet(K), either alone or together with tet(M) (P < 0.0001), while non-CC71 lineages were significantly associated with carriage of tet(M) alone (P < 0.0001).
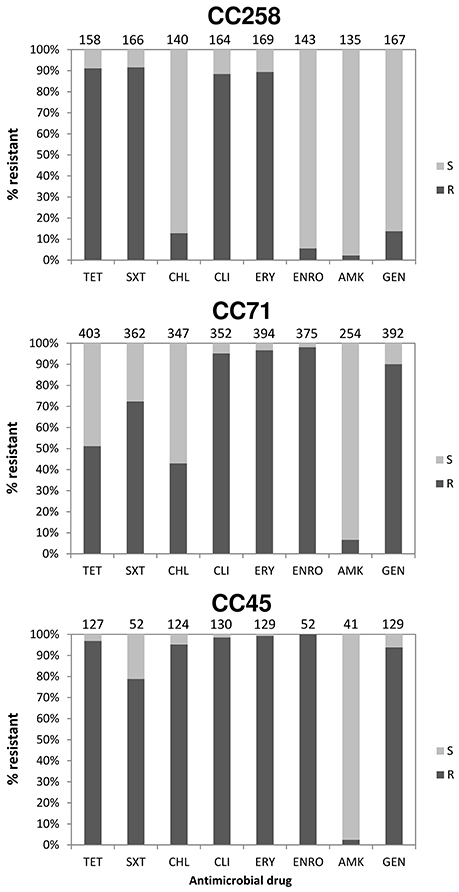
Figure 5. Proportion (%) of MRSP isolates resistant to selected antimicrobial drugs within three predominant clonal complexes (characterized by MLST-7). Antimicrobial susceptibility patterns were extracted from references given in Table S1. S, Susceptible; R, Resistance; TET, Tetracycline; SXT, Trimethoprim/Sulfamethoxazole; CHL, Chloramphenicol; CLI, Clindamycin; ERY, Erythromycin; ENRO, Enrofloxacin; AMK, Amikacin; GEN, Gentamicin.
Discussion
This is the first study providing insights into the global epidemiology of MRSP and population structure of S. pseudintermedius based on the recently established MLST-7 scheme for this species. Despite the extensive genetic diversity observed at the species level, 25 STs, 20 of which associated with MRSP, were reported in geographically-distant countries, indicating global spread of multiple high-risk MRSP clones.
Occurrence of the same clones over broad geographical areas is usually taken as evidence of clonal population structure. However, high levels of recombination (r/m = 7.52) and the index of association between the MLST loci (rbard = −0.1218, P < 0.001) revealed by our population genetics analyses show that this species is only weakly clonal. These results are consistent with an eBURST population snapshot where >25% of the STs belong to a large straggly CC (CC258), a pattern that points to potentially inaccurate links and that is often observed in populations with high recombination to mutation ratios (e.g., Burkholderia pseudomallei, Staphylococcus epidermidis, and Enterococcus faecium; Vesaratchavest et al., 2006; Miragaia et al., 2007; Turner et al., 2007; Willems et al., 2011). This clustering pattern was also recognized by Perreten et al. (2010) when using eBURST on S. pseudintermedius MLST-5 data alone. The term “epidemic clonal,” or simply “epidemic,” has been used to describe population structures of this kind, where particularly successful lineages emerge and diversify rapidly within a weakly clonal- or non-clonal population (e.g., Neisseria meningitidis; Maynard Smith et al., 2000; Vos and Didelot, 2009). This is in contrast to highly clonal species such as Staphylococcus aureus, which shows a very low recombination to mutation ratio (r/m = 0.01; Vos and Didelot, 2009) and discrete clonal lineages diversifying radially from phylogenetically well-supported founders (Feil et al., 2003). Indeed, the ClonalFrame-based phylogenetic analysis confirmed that inter-lineage recombination has likely caused incorrect links within the larger straggly group identified by eBURST. Furthermore, the genealogy shown for S. pseudintermedius is star-shaped (Figure 3), i.e., deep-level phylogenetic relationships could not be inferred, which often occurs when rates of recombination are high (Spratt and Maiden, 1999).
Both eBURST and ClonalFrame analyses showed that MRSP is found in association with numerous unrelated clones throughout the S. pseudintermedius population. These results are in agreement with recent whole genome sequencing data showing that methicillin resistance has likely evolved in genetically distinct S. pseudintermedius lineages through multiple independent SCCmec acquisitions (McCarthy et al., 2015). We estimated that S. pseudintermedius has acquired SCCmec at least 48 times, a value similar to that reported for S. epidermidis (56 times; Miragaia et al., 2007), and higher than that reported for S. aureus (at least 20 times) (Robinson and Enright, 2003). The reason for the higher number of SCCmec acquisitions in S. pseudintermedius in comparison to S. aureus might be due to an increased capacity for horizontal gene transfer, as was hypothesized by Miragaia et al. for S. epidermidis (Miragaia et al., 2007). However, the lack of complete SCCmec typing information for MRSP and the fact that at least 23 SCCmec acquisitions have occurred for one MRSA clone alone (Nübel et al., 2008) indicate that these eBURST-based estimates are likely to be underestimated.
Despite the lack of a recent common ancestor, MRSP lineages are, collectively, genetically distinct from MSSP, as shown by a small but significant genetic variation between the two subpopulations (2.31%, AMOVA: P < 0.05). This is also illustrated by the fact that only seven STs (ST17, ST29, ST56, ST123, ST125, ST155, and ST170) were common to both MRSP and MSSP, further strengthening that MRSP form a polyclonal S. pseudintermedius subpopulation composed of evolutionarily distinct lineages. The clustering performed by eBURST and ClonalFrame agreed to some extent for some CCs (e.g., CC71, CC68), supporting the clonal evolutionary model determined by eBURST. Our findings regarding CC258 further support previous evidence that the model of bacterial evolution implemented in eBURST is one of the most suitable for inferring patterns of evolutionary descent from MLST data over short periods of time when rates of recombination and clonal diversification are high (Feil et al., 2004).
This study also provides an updated overview of the geographic distribution of reported frequencies of five major CCs that have acquired methicillin resistance. MRSP CC71, which has been originally described as the epidemic European clone (Perreten et al., 2010), has spread worldwide, even though the vast majority of the isolates belonging to this CC have been reported in European countries (approximately 93%). Our observation that ~98% of CC258 isolates have been recently reported in Europe (2012–2016) confirms that CC258 may be partly replacing CC71, at least in certain European countries, as recently shown by a longitudinal study in the Netherlands (Duim et al., 2016). Geographical associations were however less marked for the other three main CCs. Approximately 33% of the isolates belonging to CC68, previously denominated as the epidemic North American clone (Perreten et al., 2010), originated from Europe. Approximately 67% of the CC45 and 49% of CC112 isolates originated from Asian countries, with the remaining isolates from the latter two CCs being isolated in Europe and to a lesser extent in North America and, for CC112, also in Oceania (Figure 4). Too few isolates originated from Africa and South America to infer clonal distribution in these continents. The different distribution of reported frequencies of MRSP clones across different countries and continents is likely to reflect various factors, including geographic distribution of S. pseudintermedius diversity and national patterns of antimicrobial usage.
In addition, significant differences were observed in the prevalence of antimicrobial resistance across distinct MRSP clonal groups. For instance, MRSP CC258 isolates display a very distinct resistance profile compared to CC71 and CC45, i.e., enrofloxacin and gentamicin resistance was infrequent in CC258 (6 and 14%), whereas resistance to trimethoprim/sulfamethoxazole was more frequent in CC258 (92%) than in the other two CCs (73–79%); tetracycline resistance was less frequent in CC71 isolates (51%) compared to those of CC258 and CC45 (91–97%); and chloramphenicol resistance was rare in CC258 (13%) and less frequent in CC71 (43%) compared to CC45, where it occurred in approximately 95% of the isolates (Figure 5). The differences found should reflect variations in the carriage of resistance genes among the different MRSP lineages, possibly due to truncation of the transposable elements that carry these genes, as shown recently by whole genome sequencing (McCarthy et al., 2015). Thus, our observations evidence that the therapeutic options available for MRSP infections are largely dependent on the clonal type involved. Since tet(M) is the most common tetracycline resistance determinant in the species, the strong association observed between CC71 and tet(K) confirms this to be an atypical feature of this CC, as suggested by previous studies (Perreten et al., 2010; Weese et al., 2013; Couto et al., 2014; Maaland et al., 2014).
A number of important limitations need to be considered when interpreting the data of this review. The main and unavoidable limitation is that the collection of isolates analyzed is naturally biased toward countries/continents that more often report MLST data. Therefore, some countries/continents are underrepresented in the analyzed collection (e.g., Africa, South America). Another unavoidable limitation is that we did not take into account differences in the antimicrobial susceptibility testing methods and interpretive criteria used by different studies for antimicrobial susceptibility testing. The lack of raw susceptibility data from the original studies makes it impossible to re-interpret the data according to standard interpretive criteria.
Conclusion
MLST data analysis revealed that S. pseudintermedius displays an epidemic population structure and high genetic diversity, which is most likely generated by multiple recombination events rather than mutations. The five main MRSP clonal lineages that have emerged over the last 10 years are phylogenetically distinct and differ significantly with regard to SCCmec content, antimicrobial resistance profiles and geographical distribution. Whole genome sequencing studies are needed in the future for a better understanding of the evolution of methicillin resistance within each clonal group.
Author Contributions
LG conceived the project. TP searched the database for potentially eligible articles, extracted the data, and performed the analyses. TP, LG, PD, and AM interpreted the results. LG contacted the authors of publications when information was not provided by the articles. TP, LG, PD, and AM wrote the manuscript. All the authors reviewed the final version of the manuscript prior to submission for publication.
Funding
The study was funded by the University of Copenhagen Research Centre for Control of Antibiotic Resistance (www.uc-care.ku.dk).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Ada Rota for providing data on MRSP strains.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01599
Figure S1. Phylogenetic analysis based on the nucleotide sequences of the S. pseudintermedius MLST genes for all 503 STs. The analysis was based on all trees sampled with a burn-in period obtained using ClonalFrame after 200,000 iterations, including 100,000 discarded burn-ins. Scale is in coalescent units. ST71, ST68, ST45, ST112, and ST258, representing important MRSP lineages, are indicated in yellow, red, green, pink and blue, respectively. MRSP, black; MSSP, white; both MRSP and MSSP, gray.
Table S1. Summary of MLST data from 49 studies and database on reported MRSP isolates, which accounted for 182 STs of the 503 STs defined by MLST-7.
References
Agapow, P. M., and Burt, A. (2001). Indices of multilocus linkage disequilibrium. Mol. Ecol. 1, 101–102. doi: 10.1046/j.1471-8278.2000.00014.x
Aslantaş, Ö., Türkyilmaz, S., Yilmaz, M. A., and Yilmaz, E. Ş. (2013). Prevalence of methicillin-resistant staphylococci in dogs. Kafkas Univ. Vet. Fak. Derg. 19, 37–42. doi: 10.9775/kvfd.2012.7073
Bannoehr, J., and Guardabassi, L. (2012). Staphylococcus pseudintermedius in the dog: taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 23, 253–266, e51–e52. doi: 10.1111/j.1365-3164.2012.01046.x
Bannoehr, J., Ben Zakour, N. L., Waller, A. S., Guardabassi, L., Thoday, K. L., van den Broek, A. H. M., et al. (2007). Population genetic structure of the Staphylococcus intermedius group: insights into agr diversification and the emergence of methicillin-resistant strains. J. Bacteriol. 189, 8685–8692. doi: 10.1128/JB.01150-07
Bardiau, M., Yamazaki, K., Ote, I., Misawa, N., and Mainil, J. G. (2013). Characterization of methicillin-resistant Staphylococcus pseudintermedius isolated from dogs and cats. Microbiol. Immunol. 57, 496–501. doi: 10.1111/1348-0421.2059
Ben Zakour, N. L., Bannoehr, J., van den Broek, A. H. M., Thoday, K. L., and Fitzgerald, J. R. (2011). Complete genome sequence of the canine pathogen Staphylococcus pseudintermedius. J. Bacteriol. 193, 2363–2364. doi: 10.1128/JB.00137-11
Black, C. C., Eberlein, L. C., Solyman, S. M., Wilkes, R. P., Hartmann, F. A., Rohrbach, B. W., et al. (2011). The role of mecA and blaZ regulatory elements in mecA expression by regional clones of methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 151, 345–353. doi: 10.1016/j.vetmic.2011.03.026
Black, C. C., Solyman, S. M., Eberlein, L. C., Bemis, D. A., Woron, A. M., and Kania, S. A. (2009). Identification of a predominant multilocus sequence type, pulsed-field gel electrophoresis cluster, and novel staphylococcal chromosomal cassette in clinical isolates of mecA-containing, methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 139, 333–338. doi: 10.1016/j.vetmic.2009.06.029
Boost, M. V., So, S. Y. C., and Perreten, V. (2011). Low rate of methicillin-resistant coagulase-positive staphylococcal colonization of veterinary personnel in Hong Kong. Zoonoses Public Health 58, 36–40. doi: 10.1111/j.1863-2378.2009.01286.x
Börjesson, S., Gómez-Sanz, E., Ekström, K., Torres, C., and Grönlund, U. (2015). Staphylococcus pseudintermedius can be misdiagnosed as Staphylococcus aureus in humans with dog bite wounds. Eur. J. Clin. Microbiol. Infect. Dis. 34, 839–844. doi: 10.1007/s10096-014-2300-y
Chanchaithong, P., Perreten, V., Schwendener, S., Tribuddharat, C., Chongthaleong, A., Niyomtham, W., et al. (2014). Strain typing and antimicrobial susceptibility of methicillin-resistant coagulase-positive staphylococcal species in dogs and people associated with dogs in Thailand. J. Appl. Microbiol. 117, 572–586. doi: 10.1111/jam.12545
Chanchaithong, P., Prapasarakul, N., Perreten, V., and Schwendener, S. (2015). Characterization of a novel composite staphylococcal cassette chromosome mec in methicillin-resistant Staphylococcus pseudintermedius from Thailand. Antimicrob. Agents Chemother. 60, 1153–1157. doi: 10.1128/AAC.02268-15
Couto, N., Belas, A., Couto, I., Perreten, V., and Pomba, C. (2014). Genetic relatedness, antimicrobial and biocide susceptibility comparative analysis of methicillin-resistant and -susceptible Staphylococcus pseudintermedius from Portugal. Microb. Drug Resist. 20, 364–371. doi: 10.1089/mdr.2013.0043
Couto, N., Monchique, C., Belas, A., Marques, C., Gama, L. T., and Pomba, C. (2016). Trends and molecular mechanisms of antimicrobial resistance in clinical staphylococci isolated from companion animals over a 16 year period. J. Antimicrob. Chemother. 71, 1479–1487. doi: 10.1093/jac/dkw029
Damborg, P., Moodley, A., Aalbæk, B., Ventrella, G., Pires dos Santos, T., and Guardabassi, L. (2016). High genotypic diversity among methicillin-resistant Staphylococcus pseudintermedius isolated from canine infections in Denmark. BMC Vet. Res. 12:131. doi: 10.1186/s12917-016-0756-y
Davis, J. A., Jackson, C. R., Fedorka-Cray, P. J., Barrett, J. B., Brousse, J. H., Gustafson, J., et al. (2014). Carriage of methicillin-resistant staphylococci by healthy companion animals in the US. Lett. Appl. Microbiol. 59, 1–8. doi: 10.1111/lam.12254
De Lucia, M., Moodley, A., Latronico, F., Giordano, A., Caldin, M., Fondati, A., et al. (2011). Prevalence of canine methicillin resistant Staphylococcus pseudintermedius in a veterinary diagnostic laboratory in Italy. Res. Vet. Sci. 91, 346–348. doi: 10.1016/j.rvsc.2010.09.014
Descloux, S., Rossano, A., and Perreten, V. (2008). Characterization of new staphylococcal cassette chromosome mec (SCC mec) and topoisomerase genes in fluoroquinolone- and methicillin-resistant Staphylococcus pseudintermedius. J. Clin. Microbiol. 46, 1818–1823. doi: 10.1128/JCM.02255-07
Detwiler, A., Bloom, P., Petersen, A., and Rosser, E. J. Jr. (2013). Multi-drug and methicillin resistance of staphylococci from canine patients at a veterinary teaching hospital (2006–2011). Vet. Q. 33, 60–67. doi: 10.1080/01652176.2013.799792
DiCicco, M., Neethirajan, S., Singh, A., and Weese, J. S. (2012). Efficacy of clarithromycin on biofilm formation of methicillin-resistant Staphylococcus pseudintermedius. BMC Vet. Res. 8:225. doi: 10.1186/1746-6148-8-225
DiCicco, M., Weese, S., Neethirajan, S., Rousseau, J., and Singh, A. (2014). Fosfomycin susceptibility of canine methicillin-resistant Staphylococcus pseudintermedius isolates. Res. Vet. Sci. 96, 251–253. doi: 10.1016/j.rvsc.2014.02.004
Didelot, X., and Falush, D. (2007). Inference of bacterial microevolution using multilocus sequence data. Genetics 175, 1251–1266. doi: 10.1534/genetics.106.063305
Duim, B., Verstappen, K. M., Broens, E. M., Laarhoven, L. M., van Duijkeren, E., Hordijk, J., et al. (2016). Changes in the population of methicillin-resistant Staphylococcus pseudintermedius and dissemination of antimicrobial-resistant phenotypes in the Netherlands. J. Clin. Microbiol. 54, 283–288. doi: 10.1128/JCM.01288-15
Eckholm, N. G., Outerbridge, C. A., White, S. D., and Sykes, J. E. (2013). Prevalence of and risk factors for isolation of meticillin-resistant Staphylococcus spp. from dogs with pyoderma in northern California, USA. Vet. Dermatol. 24, 154–161.e34. doi: 10.1111/j.1365-3164.2012.01051.x
Excoffier, L., Laval, G., and Schneider, S. (2005). Arlequin (version 3.0): an integrated software package for population genetics data analysis. Evol. Bioinform. Online 1, 47–50.
Feil, E. J., Cooper, J. E., Grundmann, H., Robinson, D. A., Enright, M. C., Berendt, T., et al. (2003). How clonal is Staphylococcus aureus? J. Bacteriol. 185, 3307–3316. doi: 10.1128/JB.185.11.3307-3316.2003
Feil, E. J., Li, B. C., Aanensen, D. M., Hanage, W. P., and Spratt, B. G. (2004). eBURST: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J. Bacteriol. 186, 1518–1530. doi: 10.1128/JB.186.5.1518-1530.2004
Feng, Y., Tian, W., Lin, D., Luo, Q., Zhou, Y., Yang, T., et al. (2012). Prevalence and characterization of methicillin-resistant Staphylococcus pseudintermedius in pets from South China. Vet. Microbiol. 160, 517–524. doi: 10.1016/j.vetmic.2012.06.015
Frank, L. A., Kania, S. A., Kirzeder, E. M., Eberlein, L. C., and Bemis, D. A. (2009). Risk of colonization or gene transfer to owners of dogs with meticillin-resistant Staphylococcus pseudintermedius. Vet. Dermatol. 20, 496–501. doi: 10.1111/j.1365-3164.2009.00826.x
Garbacz, K., Piechowicz, L., Żarnowska, S., Haras, K., and Dąbrowska-Szponar, M. (2011). Heterogeneity of methicillin-sensitive Staphylococcus pseudintermedius strains isolated from diseased dogs. Pol. J. Vet. Sci. 14, 283–284. doi: 10.2478/v10181-011-0043-6
Gharsa, H., Slama, K. B., Gómez-Sanz, E., Gómez, P., Klibi, N., Zarazaga, M., et al. (2015). Characterisation of nasal Staphylococcus delphini and Staphylococcus pseudintermedius isolates from healthy donkeys in Tunisia. Equine Vet. J. 47, 463–466. doi: 10.1111/evj.12305
Gharsa, H., Slama, K. B., Gómez-Sanz, E., Lozano, C., Klibi, N., Jouini, A., et al. (2013). Antimicrobial resistance, virulence genes, and genetic lineages of Staphylococcus pseudintermedius in healthy dogs in Tunisia. Microb. Ecol. 66, 363–368. doi: 10.1007/s00248-013-0243-y
Gómez-Sanz, E., Simón, C., Ortega, C., Gómez, P., Lozano, C., Zarazaga, M., et al. (2014). First detection of methicillin-resistant Staphylococcus aureus ST398 and Staphylococcus pseudintermedius ST68 from hospitalized equines in Spain. Zoonoses Public Health 61, 192–201. doi: 10.1111/zph.12059
Gómez-Sanz, E., Torres, C., Benito, D., Lozano, C., and Zarazaga, M. (2013a). Animal and human Staphylococcus aureus associated clonal lineages and high rate of Staphylococcus pseudintermedius novel lineages in Spanish kennel dogs: predominance of S. aureus ST398. Vet. Microbiol. 166, 580–589. doi: 10.1016/j.vetmic.2013.07.014
Gómez-Sanz, E., Torres, C., Ceballos, S., Lozano, C., and Zarazaga, M. (2013b). Clonal dynamics of nasal Staphylococcus aureus and Staphylococcus pseudintermedius in dog-owning household members. Detection of MSSA ST398. PLoS ONE 8:e69337. doi: 10.1371/journal.pone.0069337
Gómez-Sanz, E., Torres, C., Lozano, C., Sáenz, Y., and Zarazaga, M. (2011). Detection and characterization of methicillin-resistant Staphylococcus pseudintermedius in healthy dogs in La Rioja, Spain. Comp. Immunol. Microbiol. Infect. Dis. 34, 447–453. doi: 10.1016/j.cimid.2011.08.002
Gómez-Sanz, E., Torres, C., Lozano, C., and Zarazaga, M. (2013c). High diversity of Staphylococcus aureus and Staphylococcus pseudintermedius lineages and toxigenic traits in healthy pet-owning household members. Underestimating normal household contact? Comp. Immunol. Microbiol. Infect. Dis. 36, 83–94. doi: 10.1016/j.cimid.2012.10.001
Grönthal, T., Moodley, A., Nykäsenoja, S., Junnila, J., Guardabassi, L., Thomson, K., et al. (2014). Large outbreak caused by methicillin resistant Staphylococcus pseudintermedius ST71 in a Finnish veterinary teaching hospital – from outbreak control to outbreak prevention. PLoS ONE 9:e110084. doi: 10.1371/journal.pone.0110084
Grönthal, T., Ollilainen, M., Eklund, M., Piiparinen, H., Gindonis, V., Junnila, J., et al. (2015). Epidemiology of methicillin resistant Staphylococcus pseudintermedius in guide dogs in Finland. Acta Vet. Scand. 57:37. doi: 10.1186/s13028-015-0129-8
Haenni, M., Alves de Moraes, N., Châtre, P., Médaille, C., Moodley, A., and Madec, J. Y. (2014). Characterisation of clinical canine meticillin-resistant and meticillin-susceptible Staphylococcus pseudintermedius in France. J. Glob. Antimicrob. Resist. 2, 119–123. doi: 10.1016/j.jgar.2014.02.002
Haenni, M., Châtre, P., Keck, N., Franco, A., Battisti, A., and Madec, J. Y. (2013). Hospital-associated meticillin-resistant Staphylococcus pseudintermedius in a French veterinary hospital. J. Glob. Antimicrob. Resist. 1, 225–227. doi: 10.1016/j.jgar.2013.05.005
Ishihara, K., Koizumi, A., Saito, M., Muramatsu, Y., and Tamura, Y. (2016). Detection of methicillin-resistant Staphylococcus pseudintermedius ST169 and novel ST354 SCCmec II–III isolates related to the worldwide ST71 clone. Epidemiol. Infect. 144, 434–442. doi: 10.1017/S0950268815001545
Kadlec, K., Schwarz, S., Goering, R. V., and Weese, J. S. (2015). Direct repeat unit (dru) typing of methicillin-resistant Staphylococcus pseudintermedius from dogs and cats. J. Clin. Microbiol. 53, 3760–3765. doi: 10.1128/JCM.01850-15
Kadlec, K., Schwarz, S., Perreten, V., Andersson, U. G., Finn, M., Greko, C., et al. (2010). Molecular analysis of methicillin-resistant Staphylococcus pseudintermedius of feline origin from different European countries and North America. J. Antimicrob. Chemother. 65, 1826–1828. doi: 10.1093/jac/dkq203
Kadlec, K., Weiß, S., Wendlandt, S., Schwarz, S., and Tonpitak, W. (2016). Characterization of canine and feline methicillin-resistant Staphylococcus pseudintermedius (MRSP) from Thailand. Vet. Microbiol. 194, 93–97. doi: 10.1016/j.vetmic.2016.04.015
Kawakami, T., Shibata, S., Murayama, N., Nagata, M., Nishifuji, K., Iwasaki, T., et al. (2010). Antimicrobial susceptibility and methicillin resistance in Staphylococcus pseudintermedius and Staphylococcus schleiferi subsp. coagulans isolated from dogs with pyoderma in Japan. J. Vet. Medicne Sci. 72, 1615–1619. doi: 10.1292/jvms.10-0172
Kjellman, E. E., Slettemeås, J. S., Small, H., and Sunde, M. (2015). Methicillin-resistant Staphylococcus pseudintermedius (MRSP) from healthy dogs in Norway - occurrence, genotypes and comparison to clinical MRSP. Microbiologyopen 4, 857–866. doi: 10.1002/mbo3.258
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Laarhoven, L. M., de Heus, P., van Luijn, J., Duim, B., Wagenaar, J. A., and van Duijkeren, E. (2011). Longitudinal study on methicillin-resistant Staphylococcus pseudintermedius in households. PLoS ONE 6:e27788. doi: 10.1371/journal.pone.0027788
Lehner, G., Linek, M., Bond, R., Lloyd, D. H., Prenger-Berninghoff, E., Thom, N., et al. (2014). Case–control risk factor study of methicillin-resistant Staphylococcus pseudintermedius (MRSP) infection in dogs and cats in Germany. Vet. Microbiol. 168, 154–160. doi: 10.1016/j.vetmic.2013.10.023
Lozano, C., Marí, A., Aspiroz, C., Gómez-Sanz, E., Ceballos, S., Fortuño, B., et al. (2015). Nasal carriage of coagulase positive staphylococci in patients of a Primary-Healthcare-Center: genetic lineages and resistance and virulence genes. Enferm. Infecc. Microbiol. Clin. 33, 391–396. doi: 10.1016/j.eimc.2014.09.007
Maaland, M. G., Guardabassi, L., and Papich, M. G. (2014). Minocycline pharmacokinetics and pharmacodynamics in dogs: dosage recommendations for treatment of meticillin-resistant Staphylococcus pseudintermedius infections. Vet. Dermatol. 25, 182-190, e46–e47. doi: 10.1111/vde.12130
Matanovic, K., Pérez-Roth, E., Pintaric, S., and Šeol Martinec, B. (2013). Molecular characterization of high-level mupirocin resistance in Staphylococcus pseudintermedius. J. Clin. Microbiol. 51, 1005–1007. doi: 10.1128/JCM.02904-12
McCarthy, A. J., Harrison, E. M., Stanczak-Mrozek, K., Leggett, B., Waller, A., Holmes, M. A., et al. (2015). Genomic insights into the rapid emergence and evolution of MDR in Staphylococcus pseudintermedius. J. Antimicrob. Chemother. 70, 997–1007. doi: 10.1093/jac/dku496
Miragaia, M., Thomas, J. C., Couto, I., Enright, M. C., and de Lencastre, H. (2007). Inferring a population structure for Staphylococcus epidermidis from multilocus sequence typing data. J. Bacteriol. 189, 2540–2552. doi: 10.1128/JB.01484-06
Moodley, A., Riley, M. C., Kania, S. A., and Guardabassi, L. (2013). Genome sequence of Staphylococcus pseudintermedius strain E140, an ST71 European-associated methicillin-resistant isolate. Genome Announc. 1:e0020712. doi: 10.1128/genomeA.00207-12
Moodley, A., Stegger, M., Ben Zakour, N. L., Fitzgerald, J. R., and Guardabassi, L. (2009). Tandem repeat sequence analysis of staphylococcal protein A (spa) gene in methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 135, 320–326. doi: 10.1016/j.vetmic.2008.09.070
Nienhoff, U., Kadlec, K., Chaberny, I. F., Verspohl, J., Gerlach, G. F., Kreienbrock, L., et al. (2011a). Methicillin-resistant Staphylococcus pseudintermedius among dogs admitted to a small animal hospital. Vet. Microbiol. 150, 191–197. doi: 10.1016/j.vetmic.2010.12.018
Nienhoff, U., Kadlec, K., Chaberny, I. F., Verspohl, J., Gerlach, G. F., Schwarz, S., et al. (2011b). Methicillin-resistant Staphylococcus pseudintermedius among cats admitted to a veterinary teaching hospital. Vet. Microbiol. 153, 414–416. doi: 10.1016/j.vetmic.2011.05.045
Norström, M., Sunde, M., Tharaldsen, H., Mørk, T., Bergsjø, B., and Kruse, H. (2009). Antimicrobial resistance in Staphylococcus pseudintermedius in the Norwegian dog population. Microb. Drug Resist. 15, 55–59. doi: 10.1089/mdr.2009.0865
Nübel, U., Roumagnac, P., Feldkamp, M., Song, J. H., Ko, K. S., Huang, Y. C., et al. (2008). Frequent emergence and limited geographic dispersal of methicillin-resistant Staphylococcus aureus. Proc. Natl. Acad. Sci. U.S.A. 105, 14130–14135. doi: 10.1073/pnas.0804178105
Osland, A. M., Vestby, L. K., Fanuelsen, H., Slettemeås, J. S., and Sunde, M. (2012). Clonal diversity and biofilm-forming ability of methicillin-resistant Staphylococcus pseudintermedius. J. Antimicrob. Chemother. 67, 841–848. doi: 10.1093/jac/dkr576
Paul, N. C., Latronico, F., Moodley, A., Nielsen, S. S., Damborg, P., and Guardabassi, L. (2013). In vitro adherence of Staphylococcus pseudintermedius to canine corneocytes is influenced by colonization status of corneocyte donors. Vet. Res. 44:52. doi: 10.1186/1297-9716-44-52
Paul, N. C., Moodley, A., Ghibaudo, G., and Guardabassi, L. (2011). Carriage of methicillin-resistant Staphylococcus pseudintermedius in small animal veterinarians: indirect evidence of zoonotic transmission. Zoonoses Public Health 58, 533–539. doi: 10.1111/j.1863-2378.2011.01398.x
Penna, B., Mendes, W., Rabello, R., and Lilenbaum, W. (2013). Carriage of methicillin susceptible and resistant Staphylococcus schleiferi among dog with or without topic infections. Vet. Microbiol. 162, 298–299. doi: 10.1016/j.vetmic.2012.08.022
Perreten, V., Chanchaithong, P., Prapasarakul, N., Rossano, A., Blum, S. E., Elad, D., et al. (2013). Novel pseudo-staphylococcal cassette chromosome mec element (ψSCCmec57395) in methicillin-resistant Staphylococcus pseudintermedius CC45. Antimicrob. Agents Chemother. 57, 5509–5515. doi: 10.1128/AAC.00738-13
Perreten, V., Kadlec, K., Schwarz, S., Grönlund Andersson, U., Finn, M., Greko, C., et al. (2010). Clonal spread of methicillin-resistant Staphylococcus pseudintermedius in Europe and North America: an international multicentre study. J. Antimicrob. Chemother. 65, 1145–1154. doi: 10.1093/jac/dkq078
Pilla, R., Bonura, C., Malvisi, M., Snel, G. G. M., and Piccinini, R. (2013). Methicillin-resistant Staphylococcus pseudintermedius as causative agent of dairy cow mastitis. Vet. Rec. 173:19. doi: 10.1136/vr.101485
Quitoco, I. M. Z., Ramundo, M. S., Silva-Carvalho, M. C., Souza, R. R., Beltrame, C. O., de Oliveira, T. F., et al. (2013). First report in South America of companion animal colonization by the USA1100 clone of community-acquired meticillin-resistant Staphylococcus aureus (ST30) and by the European clone of methicillin-resistant Staphylococcus pseudintermedius (ST71). BMC Res. Notes 6:336. doi: 10.1186/1756-0500-6-336
Robinson, D. A., and Enright, M. C. (2003). Evolutionary models of the emergence of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 47, 3926–3934. doi: 10.1128/AAC.47.12.3926-3934.2003
Rota, A., Corrò, M., Drigo, I., Bortolami, A., and Börjesson, S. (2015). Isolation of coagulase-positive staphylococci from bitches' colostrum and milk and genetic typing of methicillin-resistant Staphylococcus pseudintermedius strains. BMC Vet. Res. 11:160. doi: 10.1186/s12917-015-0490-x
Rota, A., Milani, C., Drigo, I., Drigo, M., and Corrò, M. (2011). Isolation of methicillin-resistant Staphylococcus pseudintermedius from breeding dogs. Theriogenology 75, 115–121. doi: 10.1016/j.theriogenology.2010.07.016
Ruscher, C., Lübke-Becker, A., Semmler, T., Wleklinski, C. G., Paasch, A., Soba, A., et al. (2010). Widespread rapid emergence of a distinct methicillin- and multidrug-resistant Staphylococcus pseudintermedius (MRSP) genetic lineage in Europe. Vet. Microbiol. 144, 340–346. doi: 10.1016/j.vetmic.2010.01.008
Savini, V., Carretto, E., Polilli, E., Marrollo, R., Santarone, S., Fazii, P., et al. (2014). Small colony variant of methicillin-resistant Staphylococcus pseudintermedius ST71 presenting as a sticky phenotype. J. Clin. Microbiol. 52, 1225–1227. doi: 10.1128/JCM.02861-13
Smith, J. M., Feil, E. J., and Smith, N. H. (2000). Population structure and evolutionary dynamics of pathogenic bacteria. Bioessays 22, 1115–1122. doi: 10.1002/1521-1878(200012)22:12<1115::AID-BIES9>3.0.CO;2-R
Solyman, S. M., Black, C. C., Duim, B., Perreten, V., van Duijkeren, E., Wagenaar, J. A., et al. (2013). Multilocus sequence typing for characterization of Staphylococcus pseudintermedius. J. Clin. Microbiol. 51, 306–310. doi: 10.1128/JCM.02421-12
Spratt, B. G., and Maiden, M. C. J. (1999). Bacterial population genetics, evolution and epidemiology. Philosofical Trans. R. Soc. B Biol. Sci. 354, 701–710. doi: 10.1098/rstb.1999.0423
Starlander, G., Börjesson, S., Grönlund-Andersson, U., Tellgren-Roth, C., and Melhus, Å. (2014). Cluster of infections caused by methicillin-resistant Staphylococcus pseudintermedius in humans in a tertiary hospital. J. Clin. Microbiol. 52, 3118–3120. doi: 10.1128/JCM.00703-14
Stegmann, R., Burnens, A., Maranta, C. A., and Perreten, V. (2010). Human infection associated with methicillin-resistant Staphylococcus pseudintermedius ST71. J. Antimicrob. Chemother. 65, 2047–2048. doi: 10.1093/jac/dkq241
Tse, H., Tsoi, H. W., Leung, S. P., Urquhart, I. J., Lau, S. K. P., Woo, P. C. Y., et al. (2011). Complete genome sequence of the veterinary pathogen Staphylococcus pseudintermedius strain HKU10-03, isolated in a case of canine pyoderma. J. Bacteriol. 193, 1783–1784. doi: 10.1128/JB.00023-11
Turner, K. M. E., Hanage, W. P., Fraser, C., Connor, T. R., and Spratt, B. G. (2007). Assessing the reliability of eBURST using simulated populations with known ancestry. BMC Microbiol. 7:30. doi: 10.1186/1471-2180-7-30
Van Hoovels, L., Vankeerberghen, A., Boel, A., Van Vaerenbergh, K., and De Beenhouwer, H. (2006). First case of Staphylococcus pseudintermedius infection in a Human. J. Clin. Microbiol. 44, 4609–4612. doi: 10.1128/JCM.01308-06
Vesaratchavest, M., Tumapa, S., Day, N. P. J., Wuthiekanun, V., Chierakul, W., Holden, M. T. G., et al. (2006). Nonrandom distribution of Burkholderia pseudomallei clones in relation to geographical location and virulence. J. Clin. Microbiol. 44, 2553–2557. doi: 10.1128/JCM.00629-06
Vos, M., and Didelot, X. (2009). A comparison of homologous recombination rates in bacteria and archaea. ISME J. 3, 199–208. doi: 10.1038/ismej.2008.93
Walther, B., Hermes, J., Cuny, C., Wieler, L. H., Vincze, S., Abou, Y., et al. (2012). Sharing more than friendship — nasal colonization with coagulase-positive staphylococci (CPS) and co-habitation aspects of dogs and their owners. PLoS ONE 7:e35197. doi: 10.1371/journal.pone.0035197
Wang, Y., Yang, J., Logue, C. M., Liu, K., Cao, X., Zhang, W., et al. (2012). Methicillin-resistant Staphylococcus pseudintermedius isolated from canine pyoderma in North China. J. Appl. Microbiol. 112, 623–630. doi: 10.1111/j.1365-2672.2012.05233.x
Weese, J. S., Faires, M. C., Frank, L. A., Reynolds, L. M., and Basttiti, A. (2012). Factors associated with methicillin-resistant versus methicillin-susceptible Staphylococcus pseudintermedius infection in dogs. J. Am. Vet. Med. Assoc. 240, 1450–1455. doi: 10.2460/javma.240.12.1450
Weese, J. S., Sweetman, K., Edson, H., and Rousseau, J. (2013). Evaluation of minocycline susceptibility of methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 162, 968–971. doi: 10.1016/j.vetmic.2012.10.002
Willems, R. J. L., Hanage, W. P., Bessen, D. E., and Feil, E. J. (2011). Population biology of Gram-positive pathogens: high-risk clones for dissemination of antibiotic resistance. FEMS Microbiol. Rev. 35, 872–900. doi: 10.1111/j.1574-6976.2011.00284.x
Windahl, U., Bengtsson, B., Nyman, A. K., and Holst, B. S. (2015). The distribution of pathogens and their antimicrobial susceptibility patterns among canine surgical wound infections in Sweden in relation to different risk factors. Acta Vet. Scand. 57:11. doi: 10.1186/s13028-015-0102-6
Windahl, U., Reimegård, E., Holst, B. S., Egenvall, A., Fernström, L., Fredriksson, M., et al. (2012). Carriage of methicillin-resistant Staphylococcus pseudintermedius in dogs–a longitudinal study. BMC Vet. Res. 8:34. doi: 10.1186/1746-6148-8-34
Youn, J. H., Moodley, A., Park, Y. H., and Sugimoto, C. (2013). Genome sequence of methicillin-resistant Staphylococcus pseudintermedius sequence type 233 (ST233) strain K7, of human origin. Genome Announc. 1:e00310-13. doi: 10.1128/genomeA.00310-13
Youn, J. H., Park, Y. H., Hang'ombe, B., and Sugimoto, C. (2014). Prevalence and characterization of Staphylococcus aureus and Staphylococcus pseudintermedius isolated from companion animals and environment in the veterinary teaching hospital in Zambia, Africa. Comp. Immunol. Microbiol. Infect. Dis. 37, 123–130. doi: 10.1016/j.cimid.2014.01.003
Youn, J. H., Yoon, J. W., Koo, H. C., Lim, S. K., and Park, Y. H. (2011). Prevalence and antimicrogram of Staphylococcus intermedius group isolates from veterinary staff, companion animals, and the environment in veterinary hospitals in Korea. J. Vet. Diagnost. Investig. 23, 268–274. doi: 10.1177/104063871102300211
Keywords: Staphylococcus pseudintermedius, MLST, antimicrobial resistance, companion animals, epidemiology, systematic review, MRSP
Citation: Pires dos Santos T, Damborg P, Moodley A and Guardabassi L (2016) Systematic Review on Global Epidemiology of Methicillin-Resistant Staphylococcus pseudintermedius: Inference of Population Structure from Multilocus Sequence Typing Data. Front. Microbiol. 7:1599. doi: 10.3389/fmicb.2016.01599
Received: 01 August 2016; Accepted: 26 September 2016;
Published: 18 October 2016.
Edited by:
Daniela Ceccarelli, Wageningen Bioveterinary Research, NetherlandsReviewed by:
Dinesh Sriramulu, Shres Consultancy (Life Sciences), IndiaGiovanni Gherardi, Università Campus Bio-Medico, Italy
Copyright © 2016 Pires dos Santos, Damborg, Moodley and Guardabassi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Teresa Pires dos Santos, dHBzQHN1bmQua3UuZGs=
Luca Guardabassi, bGd1YXJkYWJhc3NpQHJvc3N2ZXQuZWR1Lmtu