 Marta Irla1
Marta Irla1 Tonje M. B. Heggeset2
Tonje M. B. Heggeset2 Ingemar Nærdal2
Ingemar Nærdal2 Lidia Paul1Tone Haugen2Simone B. Le2
Lidia Paul1Tone Haugen2Simone B. Le2 Trygve Brautaset2,3
Trygve Brautaset2,3 Volker F. Wendisch1*
Volker F. Wendisch1*- 1Genetics of Prokaryotes, Faculty of Biology and CeBiTec, Bielefeld University, Bielefeld, Germany
- 2SINTEF Materials and Chemistry, Department of Biotechnology and Nanomedicine, Trondheim, Norway
- 3Department of Biotechnology, Norwegian University of Science and Technology, Trondheim, Norway
Bacillus methanolicus is a thermophilic methylotroph able to overproduce amino acids from methanol, a substrate not used for human or animal nutrition. Based on our previous RNA-seq analysis a mannitol inducible promoter and a putative mannitol activator gene mtlR were identified. The mannitol inducible promoter was applied for controlled gene expression using fluorescent reporter proteins and a flow cytometry analysis, and improved by changing the -35 promoter region and by co-expression of the mtlR regulator gene. For independent complementary gene expression control, the heterologous xylose-inducible system from B. megaterium was employed and a two-plasmid gene expression system was developed. Four different replicons for expression vectors were compared with respect to their copy number and stability. As an application example, methanol-based production of cadaverine was shown to be improved from 6.5 to 10.2 g/L when a heterologous lysine decarboxylase gene cadA was expressed from a theta-replicating rather than a rolling-circle replicating vector. The current work on inducible promoter systems and compatible theta- or rolling circle-replicating vectors is an important extension of the poorly developed B. methanolicus genetic toolbox, valuable for genetic engineering and further exploration of this bacterium.
Introduction
Bacillus methanolicus is a thermophilic bacterium, able to grow on methanol as a sole carbon and energy source (Schendel et al., 1990; Arfman et al., 1992). The growth of B. methanolicus occurs in a wide temperature range between 37 and 60°C, with an optimum at 50°C. It was, however, observed that a rapid change of growth temperature from 50 to 37°C leads to the initiation of sporulation processes in the wild type strain MGA3, specifically the upregulation of stage VI sporulation protein D, the anti-sigma F factor antagonist SpoIIAA, and the stage IV sporulation protein A and the downregulation of two proteins which belong to the flagellar apparatus (Schendel et al., 1990; Müller et al., 2014). B. methanolicus MGA3 produces 60 g/L of L-glutamate in methanol-controlled high cell density fed-batch fermentations (Schendel et al., 2000; Heggeset et al., 2012). Currently, Corynebacterium glutamicum is typically used for the industrial production of L-glutamate in fermentative processes with the most common carbon sources being molasses and sugar cane, and the global annual consumption reaching 3.2 million tons (IHS Chemical, 2016). The second largest product of the amino acid market is L-lysine, a feed additive with an annual demand exceeding 2 million tons (Zahoor et al., 2012). B. methanolicus does not naturally overproduce this amino acid, however, during the last two decades several strategies have been employed to generate L-lysine producing strains. To date, the classical mutants of B. methanolicus produce up to 65 g/L of L-lysine in high cell density methanol fed-batch fermentations (Brautaset et al., 2010). Furthermore, it was shown both in wild type and lysine producing strains that heterologous expression of a lysine decarboxylase enables the synthesis of cadaverine (Nærdal et al., 2015). Cadaverine, also known as 1,5-diaminopentane, is a five-carbon linear aliphatic diamine (Schneider and Wendisch, 2011; Shimizu, 2013), that finds applications in the (bio)plastics industry since polycondensation of cadaverine with dicarboxylic acids yields polyamides or nylons of the AA,BB-type (Shimizu et al., 2003; Wendisch, 2014). The most significant advantage of B. methanolicus for the use in the amino acid industry is its ability to utilize methanol as a carbon source in combination with a high growth temperature, which leads to a reduced need for cooling. Methanol is a cheap, non-food alternative to raw materials commonly used in the biotechnological processes (Müller et al., 2015a). In the recent years, considerable progress has been made in the elucidation of the methanol utilization pathway starting from sequencing of the full genome (Heggeset et al., 2012; Irla et al., 2014), characterization of the enzymes involved in the methanol oxidation and the ribulose monophosphate (RuMP) pathway (Krog et al., 2013; Stolzenberger et al., 2013a,b; Markert et al., 2014; Ochsner et al., 2014; Wu et al., 2016), unraveling of the transcriptome by the means of microarray analysis (Heggeset et al., 2012) and RNA-seq (Irla et al., 2015), of the proteome (Müller et al., 2014), and the metabolome (Kiefer et al., 2015; Müller et al., 2015b). These findings enabled a better understanding of the metabolic processes taking place during growth on methanol, but also on the limited number of alternative C-sources for this facultative methylotroph, in particular on mannitol.
The obvious suitability of B. methanolicus for industrial application has been the main motivation behind the extensive work on the development of metabolic engineering tools. The first attempts included random mutagenesis approaches towards increased L-lysine production (Hanson et al., 1996; Brautaset et al., 2010). Furthermore, the protocol for the protoplast transformation with plasmid DNA was developed and several different origins of replication were tested for their transformation efficiency and stability (Cue et al., 1997). The protoplast-based transformation protocols are known to be laborious and difficult to perform, for this reason a more versatile electroporation procedure was developed, and for the first time the mdh promoter (mp) was used to establish plasmid based gene expression (Jakobsen et al., 2006). The only alternative vector that has been used for the heterologous gene expression thus far is pNW33N with a gfp gene cloned under control of the mdh promoter (Nilasari et al., 2012).
Despite the fact that some progress has been made in genetic manipulation of B. methanolicus, and that L-lysine and cadaverine producing strains have been created by the plasmid-based gene expression, the available toolbox is a limiting factor for the development of industrially relevant B. methanolicus strains. Here, we present the expansion of the metabolic engineering tool box by the addition of two new expression vectors and the establishment and development of xylose- and mannitol-inducible promoter systems.
Materials and Methods
Strains, Plasmids, and Primers
All strains, plasmids, and primers constructed and used in this study are listed in the Supplementary Tables. B. methanolicus MGA3 was used as the expression host, Escherichia coli strain DH5α (Stratagene) was used as the general cloning host.
Molecular Cloning
All standard recombinant DNA procedures were performed as described by Sambrook and Russell (2001). Plasmid DNA was introduced into chemically competent E. coli cells (Higa and Mandel, 1970; Hanahan, 1983). Total DNA was isolated from B. methanolicus using the MasterPureTM Gram Positive DNA Purification Kit (Epicenter) or as previously described (Eikmanns et al., 1994). The NucleoSpin® Gel and PCR Clean-up kit (Machery-Nagel) and the Qiaquick PCR Purification and Gel Extraction kits (Qiagen) were used for PCR purification and gel extraction. Plasmids were isolated using the GeneJET Plasmid Miniprep Kit (Thermo Fisher Scientific) or the Wizard® Plus SV Minipreps (Promega). Plasmid backbones were amplified with PfuTurbo DNA polymerase (Agilent), inserts with ALLinTM HiFi DNA Polymerase (highQ) or the ExpandTM High Fidelity PCR System (Roche). Dephosphorylation of plasmid DNA was performed using Antarctic Phosphatase or Calf Intestinal Alkaline Phosphatase (New England Biolabs). The DNA fragments were joined either with Rapid DNA Ligation Kit (Roche), T4 DNA ligase (New England Biolabs) or by the means the isothermal DNA assembly (Gibson et al., 2009). For colony PCR the Taq polymerase (New England Biolabs) was used. Site-directed mutagenesis was performed essentially as described by Liu and Naismith (2008) using Pfu polymerase (Agilent). All cloned DNA fragments and introduced mutations were verified by sequencing. B. methanolicus competent cells were prepared according to Jakobsen et al. (2006). SOBsuc plates [1% (w/v) agar] supplemented with suitable antibiotics were used instead of regeneration plates. SOBsuc medium is SOB medium (Difco) supplemented with 0.25 M sucrose. Electroporation was performed as previously described (Jakobsen et al., 2006).
Media and Cultivation Conditions
Escherichia coli strains were cultivated at 37°C in Lysogeny Broth (LB) or on LB–agar plates supplemented with antibiotics (ampicillin 200 μg/mL, chloramphenicol 30 μg/mL, kanamycin 50 μg/mL) when relevant. Unless otherwise stated, B. methanolicus strains were cultured at 50°C in MVcMY minimal medium with 200 mM methanol as previously described (Brautaset et al., 2004). When appropriate, media were supplemented with kanamycin 50 μg/mL (or 10 μg/mL) and/or chloramphenicol 5 μg/mL. Inducers were used at the following concentrations: mannitol [2.5, 5.0, 12.5, 25, 50, and 55 mM (1%)], arabitol (50 mM), ribitol (50 mM), xylitol (50 mM), xylose [0.01, 0.05, 0.1, 0.5, or 1% (w/v)], or CuSO4 (10, 20, 50, 100, and 200 μM). All experiments were performed in triplicates.
β-Galactosidase (LacZ) Activity Assay
For LacZ enzymatic assays, overnight cultures of B. methanolicus strains MGA3 (pTH1mp-lacZ), MGA3 (pTH1xp-lacZ), MGA3 (pTH1cup-lacZ), MGA3 (pTH1mtlAp-lacZ), or MGA3 (pHP13), were diluted to OD600 0.2 in fresh medium with appropriate antibiotics. When the cultures reached OD600 = 0.5, they were split in two equal halves. Inducer (50 μM CuSO4, 1% (w/v) xylose, or 1% (w/v) mannitol) was added to one of the two and growth was continued until OD600 1–1.5. Cells were harvested by centrifugation (5000 g, 10 min, 4°C) and the pellets were stored at -80°C. Cells were thawed, resuspended in potassium phosphate buffer (100 mM, pH 7.0) (10% of the original volume) and sonicated on ice/water for 10-15 min (Branson Sonifier 250, output control = 3 and duty cycle = 30%). Cellular debris was removed by centrifugation (10000 g, 45 min, 4°C) followed by filtration through a 0.2 μm sterile filter. Enzymatic activities were measured by monitoring the liberation of o-nitrophenol from o-nitrophenyl β-D-galactopyranoside (ONPG) at 410 nm. 100 mM potassium phosphate buffer pH 7.0 (910 μl), 68 mM ONPG (30 μl), and 30 mM MgCl2 (30 μl) were mixed and the catalysis started by the addition of cell extract (30 μl). The molar extinction coefficient used for o-nitrophenyl at 410 nm, pH 7.0 used for calculation is 3500 M-1 cm-1 and the light path 1 cm. One unit (U) is defined as the amount of enzyme able to convert 1.0 μmol of ONPG per min.
Flow Cytometry
For the fluorescent activated cell scanning analysis, overnight cultures were diluted to an initial OD600 of 0.15 and cultivated for 6 h at 50°C prior to incubation at 37°C for two hours. Samples were centrifuged at 13,000 g, 5 min, 4°C, washed twice with cold phosphate-buffered saline (PBS) and resuspended therein to a final OD600 of 0.3. The fluorescence was determined in a flow cytometer (Becton Dickinson) using the Kaluza for Gallios Acquisition Software 1.0. The fluorescence emission signal was collected with a 450/50 BP bandpass filter (FL9) for GFPuv and with a 620/30 BP, bandpass (FL3) for mCherry. The following data analysis was performed using Kaluza Analysis Software 1.3.
Plasmid Stability
To test for stability of plasmid segregation, overnight cultures were diluted in 50 mL fresh medium with and without relevant antibiotics to an initial OD600 of 0.05, grown for 12 h (six generations) and then diluted again into fresh medium to an initial OD600 of 0.05. This was repeated over the course of the whole experiment. After 6 h from inoculation, 10 mL of the cultures were aliquoted to 100 mL shaking flasks and incubated for 2 h at 37°C, 200 rpm, after which the flow cytometry analysis was carried out. This procedure was repeated every 24 h (every 12 generations) for a total of 5 days (60 generations). The stability is presented as the ratio of cells fluorescent in absence of antibiotics to the cells fluorescent in presence of antibiotics.
Estimation of Copy Number by the Means of Droplet Digital PCR
Overnight cultures of MGA3 (pHCMC04), MGA3 (pHP13), MGA3 (pNW33Nkan), or MGA3 (pUB110Smp-lacZ) were diluted to 2% in fresh medium (supplemented with 5 μg/ml chloramphenicol or 10 μg/ml kanamycin) and cultivated until the mid-exponential growth phase (OD600 2-4). Cell pellets were harvested from 5 ml cultures by centrifugation and total DNA was extracted using the MasterPureTM Gram Positive DNA Purification Kit (Epicenter), followed by an additional purification step using the Agencourt® AMPure XP system (Beckman Coulter). DNA concentrations were determined on a Qubit® 2.0 Fluorometer using the Qubit® dsDNA BR Assay Kit (ThermoFischer Scientific). Twenty microliter ddPCR reaction mixtures containing EvaGreen Supermix (Bio-Rad), primers (0.2 μM) and gDNA template (8 or 20 pg) were prepared according to the manufacturer’s instructions and used for droplet generation (QX200 droplet generator, Bio-Rad). Forty microliter of sample was manually transferred to a 96-well plate and heat-sealed prior to amplification initiated by enzyme activation at 95°C for 5 min, followed by 40 cycles of amplification (95°C for 30 s, 60°C 1 min) and signal stabilization (4°C 5 min, 90°C 5 min), temperature ramp 2.5°C/s. Following amplification, fluorescence intensity was measured in a QX200 Droplet Reader (Bio-Rad) and the signal data were analyzed with QuantaSoft, Version 1.5.38 (Bio-Rad). Primer sequences are listed in the Supplementary Material.
High Cell Density Fed-Batch Methanol Fermentation
Fed-batch fermentation was performed at 50°C in UMN1 medium using Applikon 3 L fermenters with an initial volume of 0.75 L medium essentially as previously described (Jakobsen et al., 2009; Brautaset et al., 2010). Kanamycin (50 μg/mL) or chloramphenicol (5 μg/mL) was added to the initial batch growth medium, the pH was maintained at 6.5 by automatic addition of 12.5% (w/v) NH3 solution, and the dissolved oxygen level was maintained at 30% saturation by increasing the agitation speed and using enriched air (up to 60% O2). The methanol concentration in the fermenter was monitored by online analysis of the headspace gas with a mass spectrometer (Balzers Omnistar GSD 300 02). The headspace gas was transferred from the fermenters to the mass spectrometer in insulated heated (60°C) stainless steel tubing. The methanol concentration in the medium was maintained at a set point of 150 mM by automatic addition of methanol feed solution containing methanol, trace metals and antifoam 204 (Sigma), as previously described (Brautaset et al., 2010). All fermentations were run until the carbon dioxide content of the exhaust gas was close to zero (no cell respiration). Bacterial growth was monitored by measuring OD600. Dry cell weight was calculated using a conversion factor of one OD600 unit corresponding to 0.24 g dry cell weight per liter (Jakobsen et al., 2009). Due to significant increase in the culture volume throughout the fermentation, the biomass, cadaverine, and amino acid concentrations were corrected for the increase in volume and subsequent dilution. A volume correction factor of 1.8 was used for values presented in Table 2. The actual concentrations measured in the bioreactors were therefore accordingly lower as described previously (Jakobsen et al., 2009). Samples for determination of volumetric cadaverine and amino acid yields were collected from early exponential phase and throughout the cultivation (10–47 h).
Measurement of Cadaverine and Amino Acids
Samples were analyzed by RP-HPLC as described previously by Skjerdal et al. (1996) using pre-column derivatization with o-phtaldialdehyde and a buffer containing 0.02 M sodium acetate +2% tetrahydrofuran at pH 5.9.
Detection of α-Amylase Activity
For the detection of α-amylase activity, overnight cultures were diluted to an initial OD600 of 0.15 and cultivated for 6-8 h at 50°C. The cultures were diluted to OD600 of 1 and 15 μL of the diluted cultures were placed on the appropriate plates in the form of a drop. The plates were incubated for 12 h at 50°C to allow the cell growth and then placed at 37°C for next 24 h for in order to support activity of the heterologous α-amylase. Ten milliliter of iodine solution were placed on the plate in order to visualize the formation of the halo in the starch.
Results
Comparison of Different Replicons for Plasmid-Based Gene Expression in B. methanolicus MGA3
Genetic engineering of B. methanolicus has until now relied on only two plasmids, pNW33N and pHP13. Therefore, we decided to analyze a range of different replicons with regard to their applicability for gene overexpression in B. methanolicus MGA3. We compared four different plasmids that were able to replicate: pTH1mp (derived from pHP13), pUB110Smp, pNW33Nmp, and pBV2mp (derived from pHCMC04). As shown in Table 1, we have chosen plasmids differing in the copy number, original host organism and the replication mechanism. All rolling circle (RC) plasmids used belong to the pC194/pUB110 family, which is characterized by similarity in Rep protein and the sequences of sites involved in the replication with pNW33N and pUB110 sharing identical Rep protein sequences. The pUB110 plasmid is reported to be a high copy number plasmid in B. subtilis, pNW33N – medium, pHP13 and pBV2mp – low copy number, both of the low copy number plasmids originate from B. subtilis.

TABLE 1. Comparison of properties and gfpUV expression from methanol-inducible promoter Pmp of plasmids used in the study.
Our initial goal was to characterize the copy number, expression levels and stability of the chosen plasmids in B. methanolicus MGA3. To analyze the expression levels, gfpuv (Chalfie et al., 1994; Crameri et al., 1996) was used as a reporter controlled by the mdh promoter from B. methanolicus MGA3. Fluorescence intensity was evaluated during growth in methanol minimal medium by flow cytometry. Using the ddPCR, plasmid copy numbers were estimated for the selected plasmids and, in comparison, for the native MGA3 plasmids pBM19 and pBM69 (Table 1). The plasmid pUB110Smp-gfpuv showed the highest fluorescence levels among the plasmids tested (Table 1), followed by pNW33Nmp-gfpuv, pTH1mp-gfpuv, and pBV2mp-gfpuv, respectively, which was in accordance to the plasmid copy number results (Table 1).
Next, we compared the plasmid stability for gfpuv-expressing RC plasmids transferred to B. methanolicus MGA3. The strains were grown for 60 generations in media with and without antibiotic selection and plasmid-containing cells emitting a fluorescence signal were counted every 12 generations. As shown in Figure 1 only the pTH1mp plasmid was lost at a significant level over the course of the experiment.
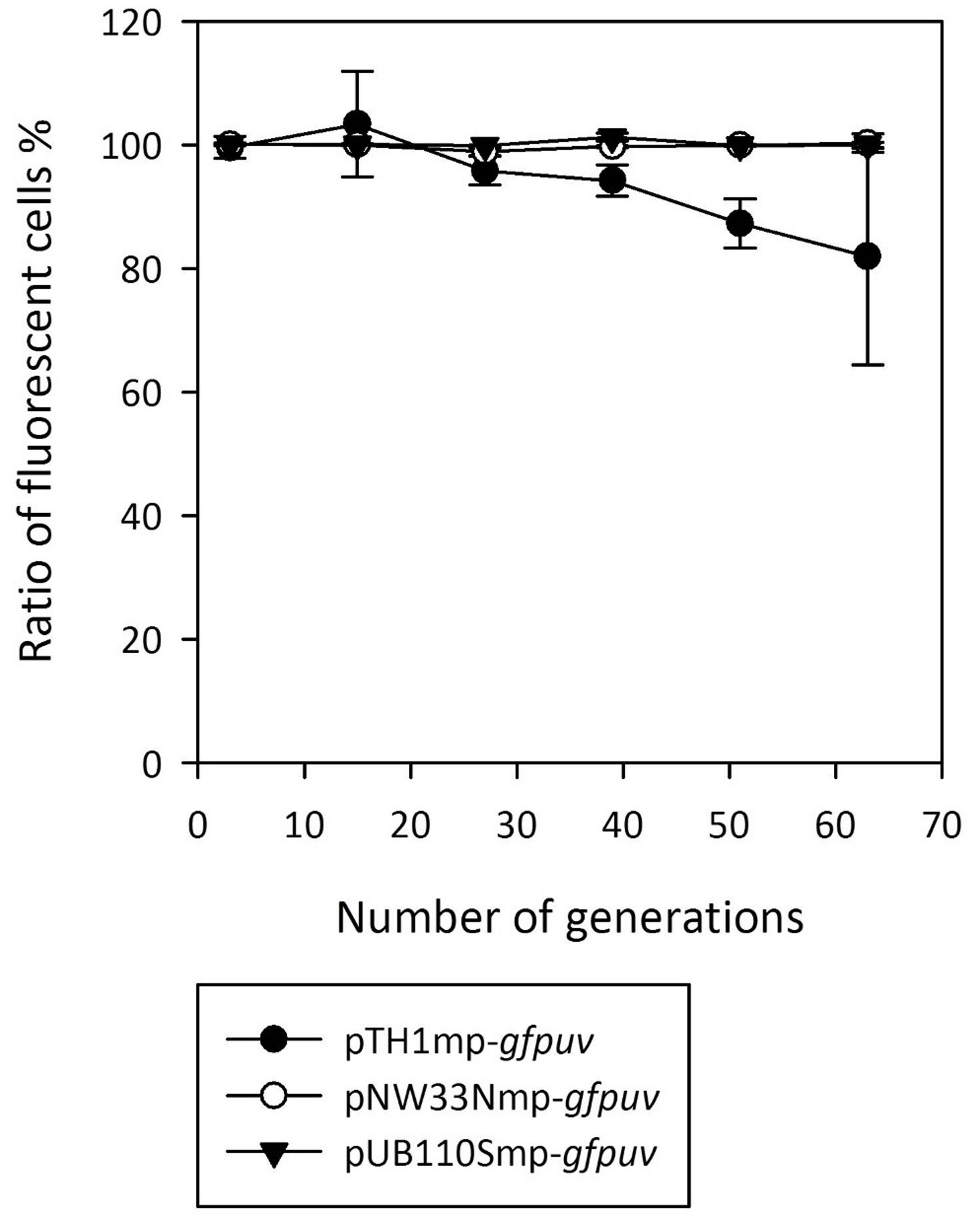
FIGURE 1. Stability of gfpuv gene expression based on the RC plasmids used in the study. The graph presents the ratio of the fluorescent cells to all the cells analyzed during growth in medium in presence and absence of antibiotics for 60 generations. Means of triplicates with standard deviation are shown.
Cadaverine Production from Methanol by Expression of a Heterologous Lysine Decarboxylase Gene from a Theta-Replicating Plasmid
The plasmids pTH1mp and pBV2mp, containing the mdh promoter were used to study cadaverine production in B. methanolicus during fed-batch methanol fermentation. We have previously reported a methanol-based cadaverine production titer of 6.5 g/L by B. methanolicus MGA3 (pTH1mp-cadA), a strain overexpressing the lysine decarboxylase cadA gene from E. coli (corrigendum to Nærdal et al., 2015). We compared cadaverine production in the strain overexpressing cadA from a theta-replicating plasmid during high cell density fed-batch fermentation. The B. methanolicus strain MGA3 (pBV2mp-cadA) was tested in duplicates under comparable fermentation conditions. Samples for cadaverine and amino acid analysis, cell dry weight and OD600 were taken throughout the cultivation. As presented in Table 2, we obtained a cadaverine production titer of 10.2 g/L based on the alternative theta-replicating pBV2mp plasmid. A substantial 55% production increase compared to the previously reported (pTH1mp-cadA)-based strain was observed. While biomass and by-product levels were similar between the two strains, the specific growth rate of MGA3 (pBV2mp-cadA) was lower than that of MGA3 (pTH1mp-cadA) (Table 2).
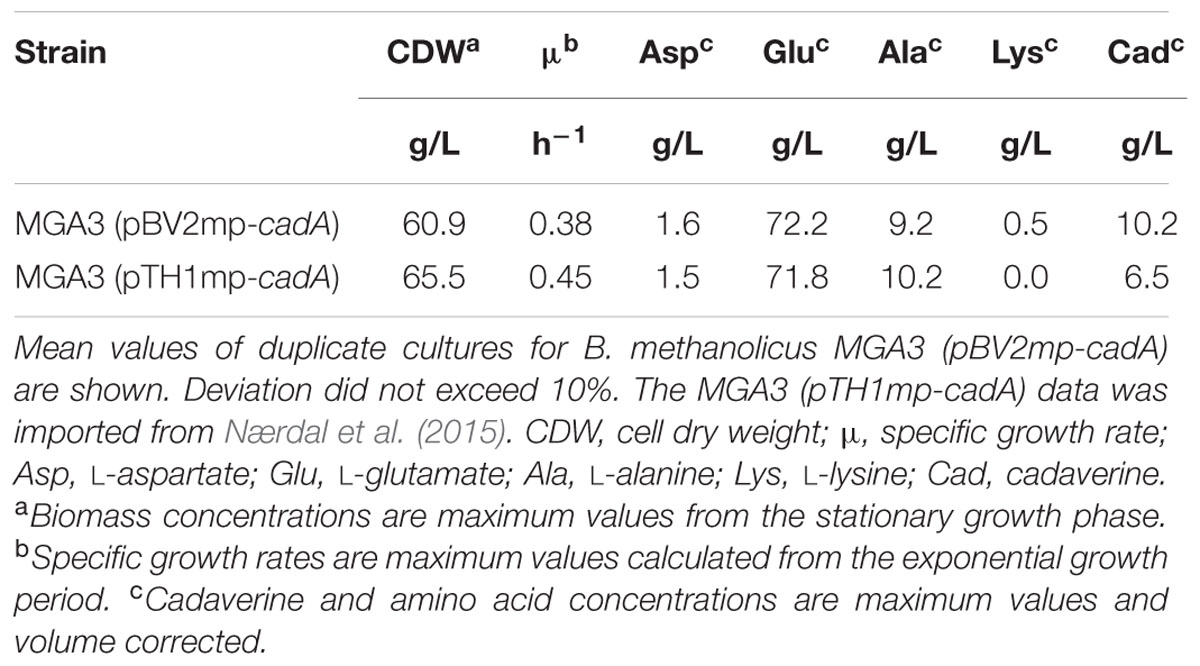
TABLE 2. Fed-batch methanol fermentation production data of strains MGA3 (pBV2mp-cadA) and MGA3 (pTH1mp-cadA).
Plasmid Compatibility
In order to establish a two plasmid-based gene expression system, we analyzed the compatibility of the chosen RC (pTH1mp and pUB110Smp)- and theta (pBV2mp)-replicating plasmids in B. methanolicus. pTH1mp and pUB110Smp share high identity (42%) of their replication protein Rep and of the origin of replication sequence (95%) and for this reason it was not clear whether they can coexist in the same cell. Similarly, we did not analyze the pUB110mp/pNW33N plasmid pair which display 100% identity of Rep protein sequences. Plasmids for expression of either gfpuv or mcherry (Shaner et al., 2004) were constructed to simultaneously analyze gene expression from two vectors. The following plasmid combinations were applied: pTH1mp-mcherry with pUB110Smp-gfpuv or pTH1mp-mcherry with pBV2mp-gfpuv. Overexpression of mcherry from pTH1mp led to red fluorescence (depicted on the y-axis in Figure 2). Similarly, overexpression of gfpuv from pUB110Smp or from pBV2mp yielded green-fluorescent cells (x-axis of Figure 2). Cells transformed with pTH1mp-mcherry and pUB110Smp-gfpuv or with pTH1mp-mcherry and pBV2mp-gfpuv showed simultaneous red and green fluorescence (Figure 2) providing evidence for two plasmid-based gene expression in B. methanolicus.
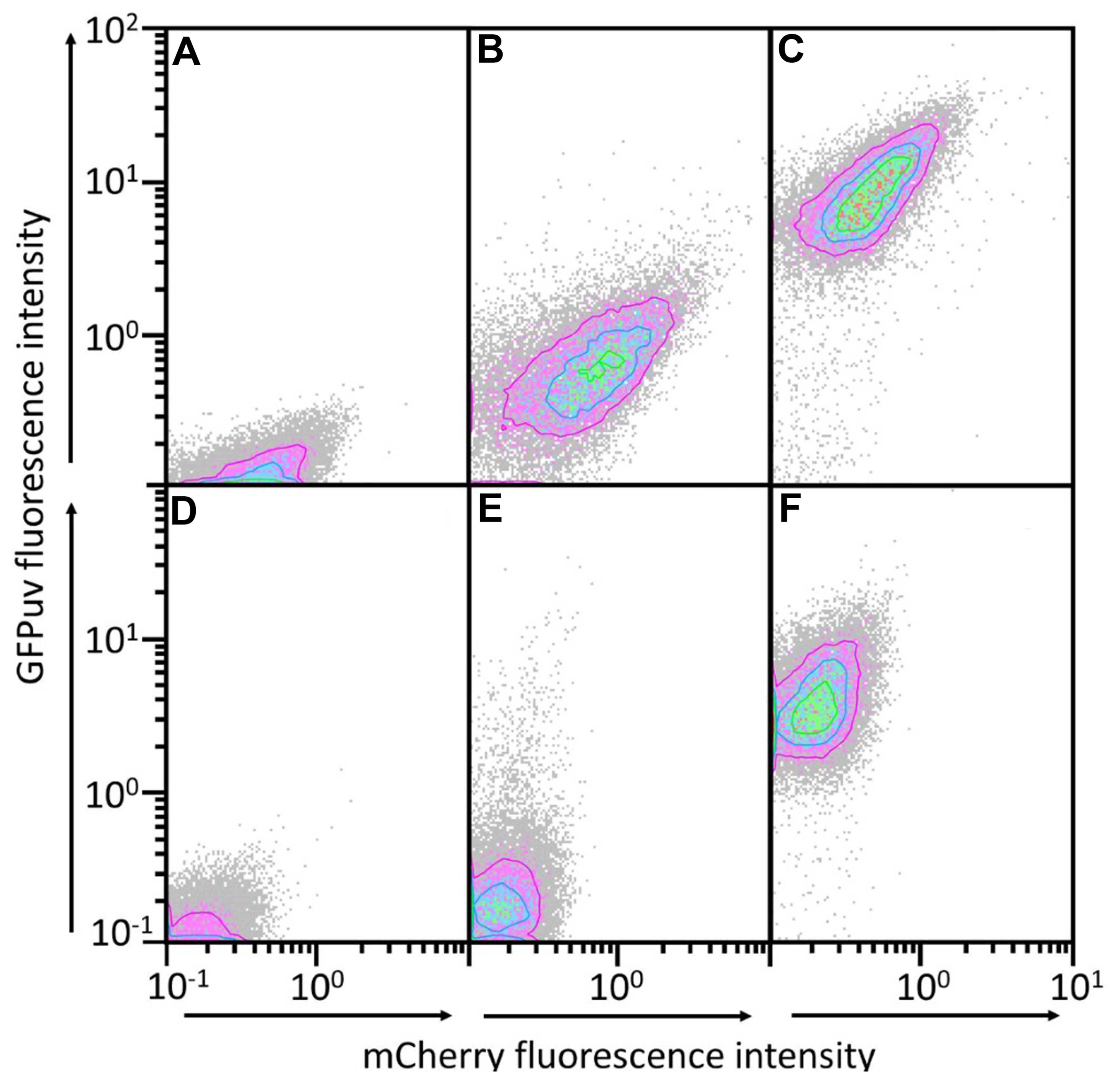
FIGURE 2. GFPuv and mCherry fluorescence contour plots of populations of B. methanolicus MGA3 transformed with two gene expression plasmids. mCherry and GFPuv fluorescence of B. methanolicus MGA3 (pTH1mp-mcherry) (A), MGA3 (pTH1mp-mCherry)(pBV2mp-gfpuv) (B), MGA3 (pTH1mp-mCherry)(pUB110Smp-gfpuv) (C), MGA3 wild type (D), MGA3 (pBV2mp-gfpuv) (E), and MGA3 (pUB110Smp-gfpuv) (F). mCherry fluorescence intensities are depicted on the x axis and GFPuv fluorescence intensities on the y axis. Contour plots of populations of 20000 cells are depicted.
Construction of Mannitol Inducible Gene Expression System
In order to choose a suitable system for inducible gene expression we screened several inducible promoter systems using the thermostable LacZ from B. coagulans as a reporter (Kovács et al., 2010). We have tested the B. megaterium xylose inducible system from plasmid pHCMC04 (Nguyen et al., 2005), a native mannitol inducible promoter from MGA3, and a copper inducible promoter from Lactobacillus sakei (Crutz-Le Coq and Zagorec, 2008). As shown in Figure 3, the xylose inducible promoter system was functional in B. methanolicus MGA3 and, when fully induced, yielded higher expression levels than the hitherto used mdh promoter. Very low expression was observed from both the mannitol-inducible promoter present in the upstream region of the mtlA gene of B. methanolicus MGA3, and the copper inducible promoter. The copper-inducible promoter showed a dose-response where the activity in cultures induced by 100 μM CuSO4 was approximately threefold higher than in cultures induced by 50 μM CuSO4 (data not shown). The inducer, however, had a toxic effect on the cells, reducing the growth rate considerably at concentrations above 50 μM CuSO4 (data not shown), making it not suitable for industrial applications.
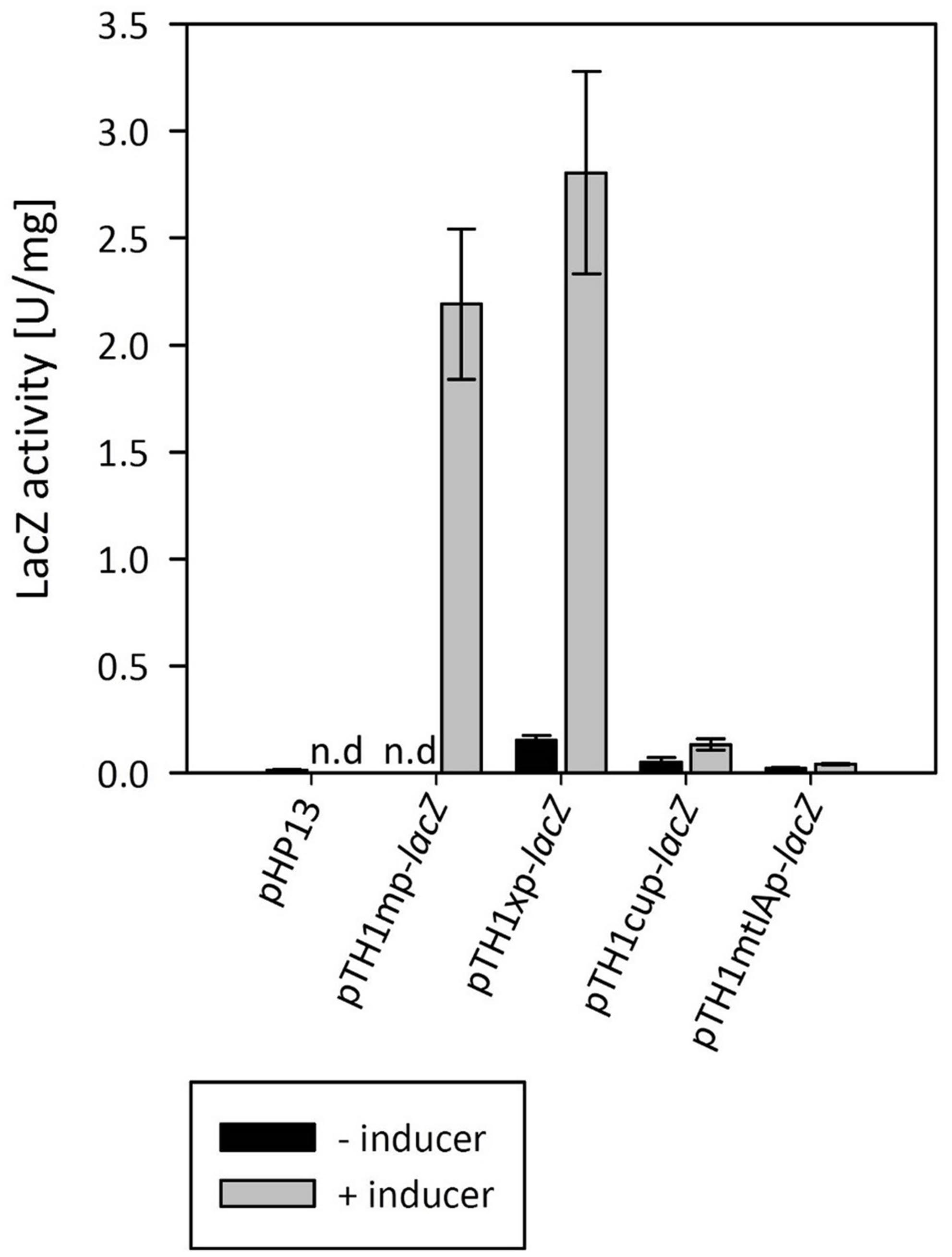
FIGURE 3. Screening of various inducible gene expression systems with lacZ used as reporter gene. The methanol, xylose, mannitol and copper inducible systems were compared. Means of triplicates with standard deviation are shown, n.d – no data.
Since only very low expression was observed from the mannitol-inducible mtlA promoter, we used DNA microarray data (Heggeset et al., 2012) and RNA-seq data (Irla et al., 2015) to identify other mannitol-inducible genes. Figure 4 presents the genomic and transcriptomic organization of four genes which belong to the mannitol utilization pathway: mtlA coding for PTS system mannitol-specific EIICB component, mtlR encoding a transcriptional regulator, mtlF coding for mannitol-specific phosphotransferase enzyme IIA and mtlD encoding mannitol-1-phosphate 5-dehydrogenase. Genes mtlF and mltD are co-expressed as an operon. Transcription start sites (TSSs) were not detected either for mtlA or for mtlF-mtlD; however, a TSS was found for mtlR (Irla et al., 2015). This 5′ untranslated region (5′ UTR) of mtlR is 80 nt in length and its upstream sequence contains conserved -10 and -35 regions (bold): 5′-TTGTATTAAGGGATATAAACGTTTTATGATAAATATG-3′, furthermore the putative ribosome binding site (RBS) sequence is AGTGGAG, which differs in two positions from the B. methanolicus consensus RBS motif AGGAGG (Irla et al., 2015). We cloned the upstream sequence of the mtlR gene into the plasmid pTH1 containing the gfpuv gene and exchanged the RBS sequence to the consensus motif, which resulted in plasmid pTH1m2p-gfpuv.
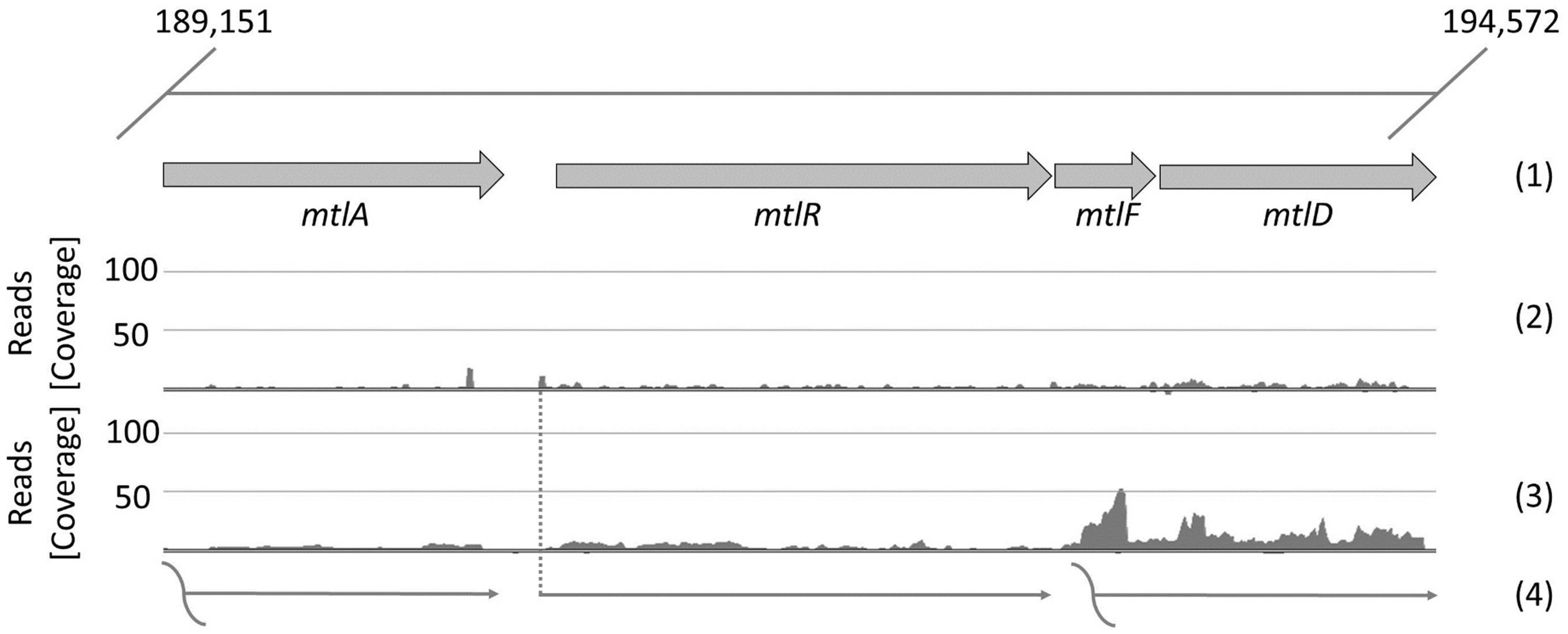
FIGURE 4. Transcriptional organization of mannitol utilization pathway genes of B. methanolicus MGA3. The first line (1) represents the genomic organization of the genes in the cluster, the next one (2) the mapped reads of 5′-ends of primary transcripts, the third (3) depicts the mapped reads of whole primary transcripts and the last (4): the putative corresponding transcripts. The level of transcription is visualized with ReadXplorer (Hilker et al., 2014) and given as absolute reads (coverage) at the corresponding genomic positions. Data are based on the RNA-seq analysis of Irla et al. (2015).
At first, several sugar alcohols were tested as potential inducers (Figure 5A). Supplementation with 50 mM mannitol induced gfpuv expression; however, neither arabitol, ribitol, nor xylitol induced reporter gene expression (Figure 5A). Subsequently, a titration experiment with different concentrations of mannitol was performed. While the addition of 5 mM mannitol did not increase reporter gene expression, high GFPuv fluorescence intensities were observed upon addition of 12.5, 25, and 50 mM mannitol (Figure 5B). It has to be noted that GFPuv fluorescence intensities in the presence of 12.5, 25, and 50 mM mannitol were comparable suggesting that full induction has been achieved (Figure 5B). The expression level of the mannitol-inducible promoter in the presence of 50 mM mannitol was similar to that obtained with the conventionally used mdh promoter. To test whether higher gene expression is possible in the mannitol inducible system, we decided to exchange the sequences of the -10 and/or the -35 region for the previously described consensus sequences (Irla et al., 2015). As shown in Table 3, the exchange of the -35 region or the -35 region together with the -10 region led to higher fluorescence levels in comparison to the native promoter. However, the double exchange caused a 3.5-folds increased background expression.
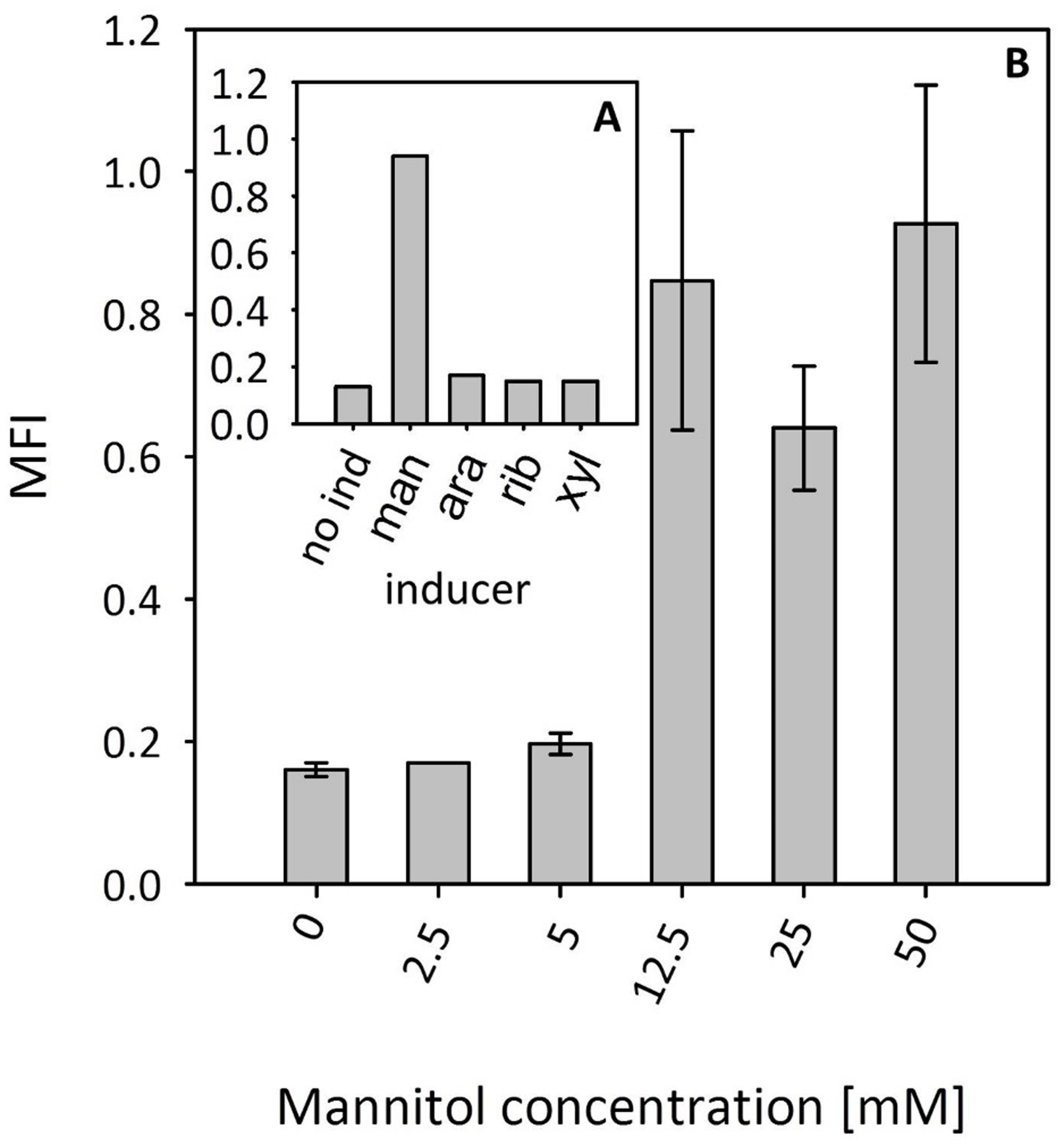
FIGURE 5. Specificity and dynamic range of induction of the B. methanolicus mtlR promoter. (A) Screening of sugar alcohols as potential inducers. Single replicates of shake flask cultures are presented. (B) Expression titration with different mannitol concentrations. Abbreviations: no ind, no inducers; man, mannitol; ara, arabitol; rib, ribitol; xyl, xylitol; MFI, median fluorescence intensity of GFPuv given in a.u. GFPuv fluorescence of exponentially growing cells was measured after 2 h incubation at 37°C and 200 rpm Mean values and standard deviations of triplicate shake flask cultures are presented.

TABLE 3. Reporter gene expression from the Bacillus methanolicus mannitol inducible mtlR promoter with changed -35 and -10 region sequences.
MtlR has been characterized as a mannitol-dependent transcriptional activator in several species (Joyet et al., 2015). The alignment of the B. methanolicus MtlR protein sequence with the sequences of characterized regulators from L. casei BL23, B. subtilis ssp. subtilis str. 168 and Geobacillus stearothermophilus ATCC 7954 (Figure 6) revealed the conserved residues important for the regulatory activity of MtlR. The high similarity to the characterized proteins suggested that MtlR of B. methanolicus most probably serves as a transcriptional activator. For this reason, we decided to test whether the plasmid-borne overexpression of mtlR increased mtlR promoter activity. As shown in Figure 7, the overexpression of this gene increased the reporter gene expression from the mannitol inducible promoter m2p by more than 2.5-fold while maintaining a low level of background expression in the absence of mannitol. Taken together, evidence is provided for a versatile mannitol inducible system for the thermophilic B. methanolicus on the basis on the previously obtained RNA-seq data.

FIGURE 6. Amino acid sequence alignment of the PRD2 (A) and EIIMtl (B) domains of various MtlR proteins. The known regulatory sites are in boldface, conserved sequence (∗), conservative mutations (:), semi-conservative mutations (.), and non-conservative mutation ( ). The alignment was performed with T-Coffee (Notredame et al., 2000). The GenBank accession numbers of the sequences are as follows: L. cas: Lactobacillus casei BL23, FM177140.1; B. sub: B. subtilis ssp. subtilis str. 168, CP010052.1; G. str: Geobacillus stearothermophilus ATCC 7954, U18943.1; B. met: B. methanolicus MGA3, CP007739.1.
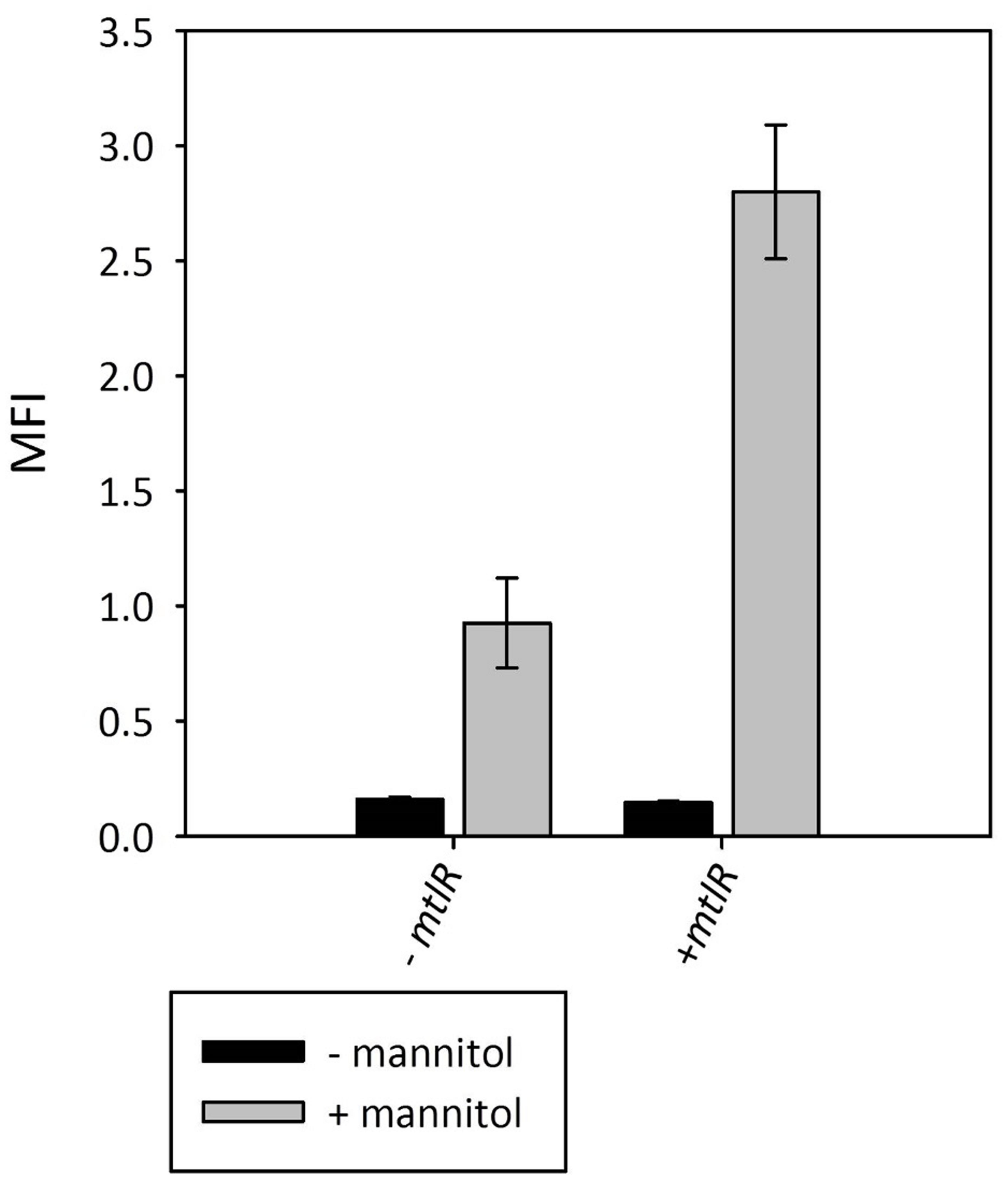
FIGURE 7. Effect of plasmid-borne overexpression of mtlR on mannitol-inducible gfpuv reporter gene expression from mtlR promoter variant m2p. MFI - median fluorescence intensity of GFPuv given in a.u. GFPuv fluorescence of exponentially growing cells was measured after 2 h incubation at 37°C and 200 rpm. Mean values and standard deviations of triplicate shake flask cultures are given.
Xylose Inducible Gene Expression in B. methanolicus
Based on our screening experiments (Figure 3), we decided to further develop the xylose inducible system for gene expression in B. methanolicus. We have subcloned the xylR regulator gene together with the promoter and the RBS sequence of the B. megaterium xylA gene into pTH1 to drive expression of gfpuv. The resulting plasmid was named pTH1xpx-gfpuv. Figure 8 shows the expression levels of gfpuv transcribed from the xylose inducible promoter in media with different xylose concentrations. Reporter gene expression increased linearly in the concentration range between 0.01% (w/v) and 0.1% (w/v) and reached a plateau at 0.5% (w/v). The fluorescence from fully induced xpx promoter is around 15-fold higher in comparison to the conventionally used mdh promoter (Table 1). Furthermore, the background gene expression with uninduced MGA3 (pTH1xpx-gfpuv) was very low (0.17 ± 0.01 a.u.) as compared to the background fluorescence (0.12 ± 0.00 a.u.) obtained for wild type B. methanolicus MGA3. Notably, mannitol did not induce expression of gfpuv from the xylose inducible promoter (data not shown).
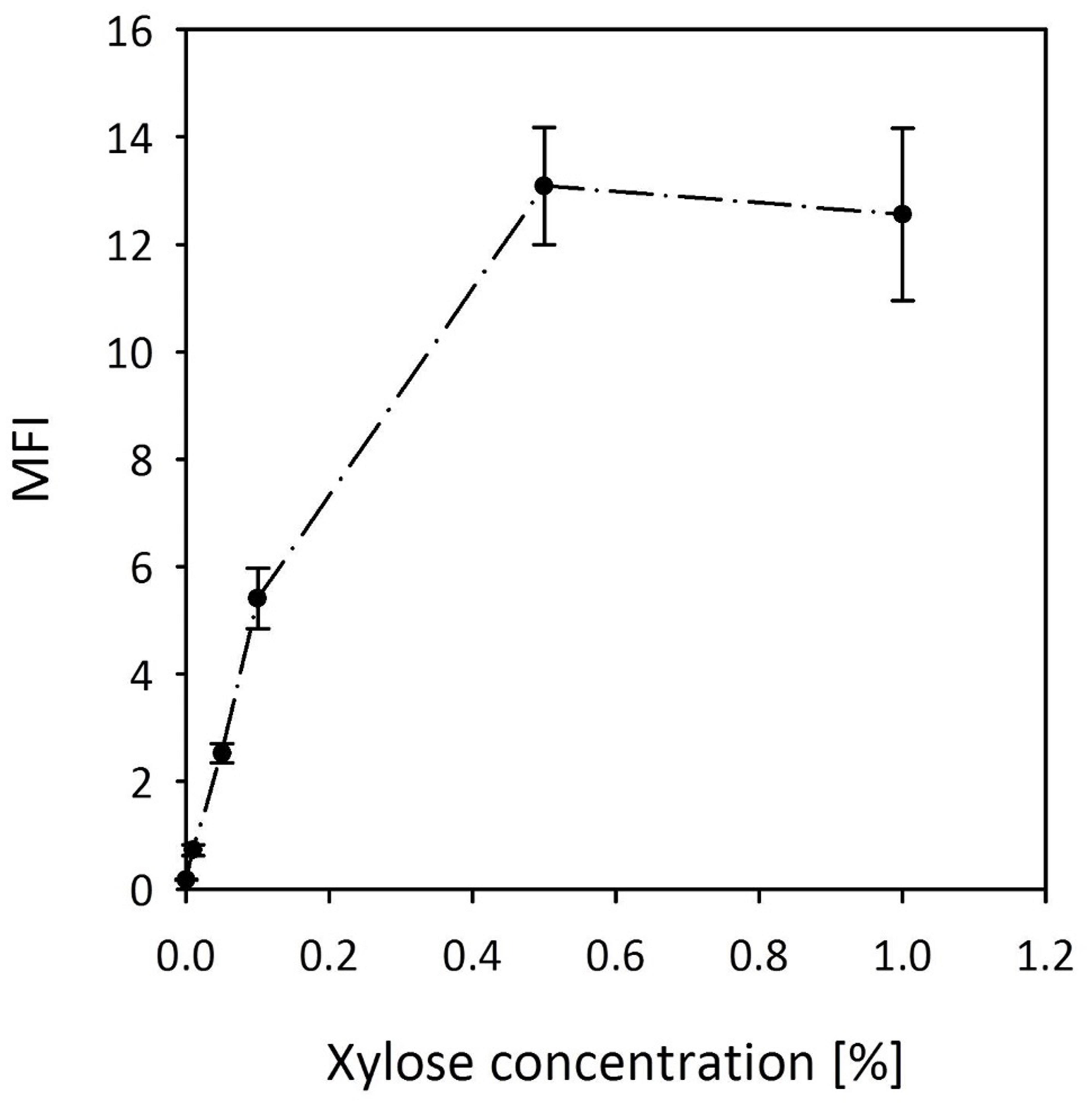
FIGURE 8. Induction of reporter gene expression from the xylose inducible promoter of pTH1xpx-gfpuv. MFI - median fluorescence intensity of GFPuv given in a.u. GFPuv fluorescence of exponentially growing cells was measured after 2 h incubation at 37°C and 200 rpm. Mean values and standard deviations of triplicate shake flask cultures are presented.
Introduction of Heterologous Starch Degradation Pathway in B. methanolicus MGA3 by Heterologous Overexpression of α-Amylase Gene from Streptomyces griseus IMRU3570
α-Amylases degrade starch to glucose and expression of heterologous α-amylase genes in glucose-positive, but starch-negative species enabled starch utilization as for example shown for C. glutamicum expressing α-amylase gene (amy) from Streptomyces griseus IMRU3570 (Seibold et al., 2006). A BLAST search of the Bacillus methanolicus genome revealed two genes putatively encoding α-amylases (BMMGA3_04340, BMMGA3_04345) and one coding for an α-glucosidase (Heggeset et al., 2012; Irla et al., 2014). For heterologous expression of amy from S. griseus plasmid pTH1xpx was used for xylose inducible expression in B. methanolicus. Starch degradation by the control strain B. methanolicus MGA3 (pTH1mp) on LB agar plates supplemented with 0.5% soluble starch and 0.05% xylose at 37°C was not observed (Figure 9A). By contrast B. methanolicus MGA3 (pTH1xpx-amy) showed a halo on starch LB plates containing xylose as an inducer and incubated at 37°C (Figure 9B) indicating that expression of amy from S. griseus plasmid allowed for starch degradation by recombinant B. methanolicus.
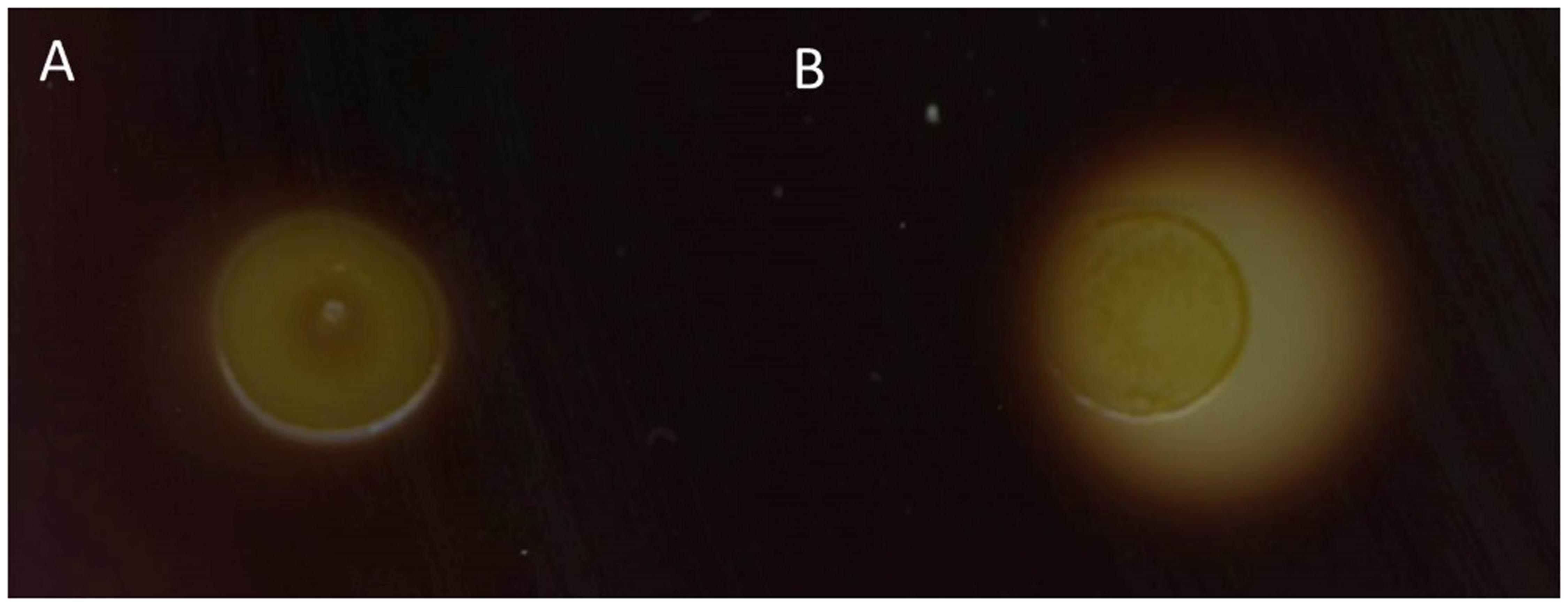
FIGURE 9. Starch degradation by B. methanolicus strains. The strains were grown on LB plates supplemented with 0.5% starch and 0.05% xylose and iodine added to detect starch degradation. The dark area in the plate indicates presence of starch and the colorless halo around cells indicates starch degradation. (A) B. methanolicus MGA3 (pTH1mp) and (B) B. methanolicus MGA3 (pTH1xpx-amy).
Discussion
In this study we have developed a versatile toolbox for inducible gene expression in B. methanolicus from RC- and theta-replicating plasmids. As a test case, we have applied a theta-replicating plasmid for heterologous expression of the lysine decarboxylase gene from E. coli and have shown improved cadaverine (1,5-diaminopentane) production in methanol-controlled fed-batch fermentations.
Strain development for B. methanolicus until recently relied on (over-)expression of genes or operons from a single plasmid despite the need for gene co-expression from two different plasmids and for inducible gene expression (Brautaset et al., 2010; Nærdal et al., 2015). To that end, we have extended the existing portfolio of available expression vectors (based on pHP13 and pNW33N) with the two additional replicons pUB110 and pHCMC04 (Gryczan et al., 1978; Cue et al., 1997; Nguyen et al., 2005; Nilasari et al., 2012). Plasmids pUB110, pHP13 and pNW33N replicate via a RC mechanism (Khan, 1997) and belong to the same plasmid family. This family is named pC194/pUB110 and is characterized by a similar ori sequence CTT(G)TTCTTTCTTATCTTGATA. However, they are known to have different copy numbers in B. subtilis. Typically, RC plasmids are known to replicate in thermophilic bacteria (Soutsehek-Bauer et al., 1987; Cue et al., 1997; Rhee et al., 2007). This, to the best of our knowledge, was not known for theta-replicating plasmids and we show here for the first time that the theta-replicating plasmid pHCMC04 replicates stably in the thermophilic B. methanolicus.
Cadaverine production by recombinant B. methanolicus expressing the E. coli lysine decarboxylase gene cadA was superior when using the theta-replicating plasmid pBV2mp-cadA (pHCM04 replicon) as compared to the rolling circle-replicating plasmid pTH1mp-cadA (pHP13 replicon) (Table 2). Despite the low copy number of the pHCMC04 replicon (approximately half of that for replicon pHP13) confirmed both by ddPCR and GFPuv fluorescence measurement (Table 1), cadaverine production by B. methanolicus MGA3 (pBV2mp-cadA) was about 55% higher (Table 2) than by MGA3 (pTH1mp-cadA). This observation results most probably from two factors: loss of the pHP13 replicon over cultivation time and high stability of the pHCMC04 replicon. The loss of the pHP13 replicon was somewhat surprising as it was reported to be stable in B. methanolicus (Cue et al., 1997). Plasmid pHP13 contains the ori sequence from parental plasmid pTA1060 (Haima et al., 1987), but is however lacking a 167-bp fragment outside of the ori sequence from pTA1060, which has been shown to improve stable plasmid segregation (Bron et al., 1987; Chang et al., 1987; Haima et al., 1987). By contrast, the theta-replicating plasmid pHCMC04 showed the expected high stability typically observed for this type of plasmids (Bruand et al., 1991; Titok et al., 2003; Nguyen et al., 2005). High plasmid stability is important in large-scale industrial processes requiring long seed trains or in fed-batch and continuous cultivations since subpopulations of cells which have lost plasmids due to low segregational stability usually lead to significant productivity losses (Friehs, 2004). With respect to methanol utilization in the fed-batch cultivations, it was observed that the overall carbon consumption (from methanol) of strains MGA3 (pTH1mp-cadA) and MGA3 (pBV2mp-cadA) differed by less than 5% (data not shown). Thus, the finding that cadaverine production increased by 6.2 g/L and biomass formation was reduced by 4.6 g/L (Table 2), suggested a reallocation of carbon source utilization from biomass to product formation.
Here we have characterized the plasmid pUB110 as a very feasible choice for gene expression in thermophilic B. methanolicus for two reasons: it showed the highest copy number among the tested replicons (Table 1) and showed high segregational stability (Figure 1). In B. subtilis, the pUB110 plasmid is known as a high copy number plasmid (Gryczan et al., 1978), whereas segregational stability seems to be a more complex issue. It was shown in several studies that the wild type plasmid is stable over multiple generations in different Bacillus spp. including B. subtilis (Polak and Novic, 1982; Alonso et al., 1987; Shoham and Demain, 1990), B. thuringiensis (Naglich and Andrews, 1988) and B. sphaericus (Seyler et al., 1991). Nonetheless, molecular modifications may lead to decreased stability of pUB110 for several different reasons. The segregational instability may be the function of the insert size (Bron et al., 1988; Zaghloul et al., 1994) or the high expression level of the cloned gene (Vehmaanperii and Korhola, 1986). Moreover, the lack of the so-called BA3 and BA4 regions has been described to destabilize the plasmid (Tanaka and Sueoka, 1983; Bron and Luxen, 1985; Shoham and Demain, 1990). Despite the fact that the pUB110-derived plasmid (pUB110Smp) used in this study did not contain BA3 and BA4 sequences and contained a 2.4 kbp insert, it was stable over 60 generations in B. methanolicus (Figure 1). Taken together, in thermophilic B. methanolicus pUB110Smp seems to be more feasible for molecular cloning than the hitherto used pTH1mp replicon.
Additionally, pUB110Smp as well as the theta-replicating pBV2mp were shown to be compatible with pTH1mp and could be used for independent expression of two genes in a two-plasmid approach.
Bacillus methanolicus not only grows with mannitol as the carbon source, but also shows mannitol dependent induction of at least two promoters PmtlA and PmtlR as revealed by transcriptome and proteome analyses (Heggeset et al., 2012; Müller et al., 2014; Irla et al., 2015). As compared to growth on methanol, mannitol-grown cells showed about 20-fold higher abundances of the proteins involved in mannitol utilization, i.e., EIIA and EIIBC components of the mannitol-specific PTS and mannitol-1-phosphate 5-dehydrogenase (Müller et al., 2014). The genomic organization suggests monocistronic transcription of mtlA and co-transcription of mtlRFD although RNA-seq data also suggest co-transcription of mtlFD without mtlR (see Figure 2). Reporter gene expression from PmtlR was higher than from PmtlA. Background expression from PmtlR was low and the induction when grown in the presence of mannitol was 6.5-fold for the native promoter and 13-fold for the improved version. These values are lower compared to mannitol inducible promoters in B. subtilis and Pseudomonas putida which show induction rates of about 20 for the native promoters and up to 176 for modified versions (Heravi et al., 2011; Hoffmann and Altenbuchner, 2015). The threshold concentration of mannitol required for induction of the promoter mtlR in the vector pTH1m2p was about 12.5 mM which is higher than the Monod constant, i.e., the concentration supporting growth with mannitol with a half-maximal growth rate (about 0.5 mM). This different threshold may reflect basal expression of mannitol utilization genes (e.g., mtlA, mtlFD operon). Moreover, we used the promoter of the regulatory gene mtlR rather than a promoter of a structural gene (e.g., mtlA, mtlFD operon) and dose dependency of induction of mtlA or the mtlFD operon might differ from dose dependency of induction of mtlR. Thus, it is conceivable that when mannitol is present in limited concentrations in the environment the background expression of the mannitol utilization genes is sufficient for initial mannitol utilization, whereas only higher mannitol concentrations lead to autoinduction of mtlR expression.
Induction of PmtlR was specific to mannitol, while similar sugar alcohols did not affect transcription. The fact that mannitol is one of the few carbon sources of B. methanolicus precludes its use as gratuitous inducers similar to IPTG in the E. coli lac system or the xylose system applied to B. methanolicus (see below).
The xylose inducible system originating from B. megaterium was previously successfully used in several bacterial species, including B. megaterium (Rygus and Hillen, 1991), B. subtilis (Kim et al., 1996), Staphylococcus aureus (Zhang et al., 2000), and Brevibacillus choshinensis (D’Urzo et al., 2013). Here we show that this system also works in B. methanolicus. Since xylose is not metabolized by B. methanolicus it serves as a gratuitous inducer in this bacterium. In fact, the xylose inducible system turned out to have multiple advantages, including very low background expression in the uninduced state, titratable induction, and a 75-fold induction window between the uninduced and the fully induced state. Similar high dynamic ranges of xylose induction have been reported for other Bacillus ssp. (Kim et al., 1996; Zhang et al., 2000; Bhavsar et al., 2001). However, catabolite repression of the xylose inducible promoter in multiple Bacillus ssp. is disadvantageous for biotechnological applications. Catabolite repression is due to the cis-acting catabolite responsive element (cre), which is a binding site of the catabolite repressor protein CcpA (Jacob et al., 1991; Lokman et al., 1994; Kim et al., 1996; Schmiedel and Hillen, 1996; Chaillou et al., 1998; Bhavsar et al., 2001; Miyoshi et al., 2004). To avoid this phenomenon, the cre sequence TGAAAGCGCAAACA of the xyl operon in B. megaterium, which is located within the of xylA gene (Schmiedel and Hillen, 1996), is not present in the plasmids used here. The absence of the cre sequence from the plasmids may be relevant since the genome of B. methanolicus encodes a homolog of CcpA (BMMGA3_13325). A BLAST search did not indicate that the cre sequence TGAAAGCGCAAACA is present upstream of the genes for carbon source utilization (only methanol, glucose, or mannitol are known carbon sources) of B. methanolicus. A cre sequence may be present upstream of the putative glucosamine-6-phosphate synthetase encoding gene glmS (BMMGA3_01020). In B. subtilis, glmS mRNA acts as a metabolite-responsive ribozyme (Winkler et al., 2004) and glucose-repressive glmS transcription is at least partially under CcpA-independent control (Yoshida et al., 2001). However, the regulatory mechanism of the homolog of CcpA (BMMGA3_13325) of B. methanolicus and its target genes need still to be defined. Plasmid pTH1xpx for xylose inducible gene expression was applied for heterologous expression of Streptomyces griseus-derived α-amylase gene in B. methanolicus. As shown before for mesophilic Corynebacterium glutamicum (Seibold et al., 2006), heterologous expression of the α-amylase gene from S. griseus supported starch degradation by recombinant B. methanolicus assayed at 37°C. While α-amylase from S. griseus was an obvious choice it has its limitations in thermophiles since the enzyme is known to exhibit maximal activity at 30°C with 92% of the remaining activity at 40°C, but only trace activity being observable at 50°C (Simpson and McCoy, 1953). Nonetheless, heterologous expression of amy from S. griseus serves as an example that the gene expression tools described here are suitable for pathway engineering of B. methanolicus.
Taken together, a series of plasmids for stable replication in the thermophilic B. methanolicus was developed for xylose as well as mannitol inducible gene expression. Thus, an important step for further advancing this thermophilic bacterium as a very promising candidate for industrial production of amino acids and their derivatives has been reached. Improved production of cadaverine using a theta-replicating plasmid for heterologous expression of the lysine decarboxylase gene from E. coli in methanol-controlled fed-batch fermentations was demonstrated as a first application example, starch degradation by recombinant B. methanolicus carrying xylose inducible expression plasmid pTH1xpx with the gene for α-amylase from S. griseus as a second example.
Author Contributions
MI, TH, IN, LP, TH, SL carried out the experimental procedure and the data analysis of the present study. MI prepared a draft of the manuscript. MI, TH, IN, SL, TB, and VW finalized the manuscript. TB and VW coordinated the study. All authors read and approved the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the EU7 FWP project PROMYSE and the ERASysAPP project MetApp. MI acknowledges support from the CLIB Graduate Cluster Industrial Biotechnology at Bielefeld University, Germany which is financed by a grant from the Federal Ministry of Innovation, Science and Research (MIWF) of the federal state North Rhine-Westphalia, Germany. We acknowledge support for the Article Processing Charge by the Deutsche Forschungsgemeinschaft and the Open Access Publication Fund of Bielefeld University. We thank Dr. Oskar Zelder and Dr. Rober Thummer for providing the pUB110 plasmid and for scientific discussion, Dr. Oscar P. Kuipers for providing the pNZlacZ-plasmid and Dr. Anne-Marie Crutz-Le Coq for providing the pRV613 plasmid. Per O. Hansen, Elisabeth Elgsæter, Nils Kirschnick, Bin Liu, and Julia Koch are thanked for technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01481
References
Alonso, J. C., Viret, J., and Tailor, R. H. (1987). Plasmid maintenance in Bacillus subtilis recombination-deficient mutants. Mol. Gen. Genet. 208, 349–352. doi: 10.1007/BF00330464
Arfman, N., Dijkhuizen, L., Kirchhof, G., Ludwig, W., Schleifer, K. H., Bulygina, E. S., et al. (1992). Bacillus methanolicus sp. nov., a new species of thermotolerant, methanol-utilizing, endospore-forming bacteria. Int. J. Syst. Bacteriol. 42, 439–445. doi: 10.1099/00207713-42-3-439
Bhavsar, A. P., Zhao, X., and Brown, E. D. (2001). Development and characterization of a xylose-dependent system for expression of cloned genes in Bacillus subtilis: conditional complementation of a teichoic acid mutant development and characterization of a xylose-dependent system for expression of. Appl. Environ. Microbiol. 67, 1–3. doi: 10.1128/AEM.67.1.403
Brautaset, T., Jakobsen,Ø. M., Degnes, K. F., Netzer, R., Nærdal, I., Krog, A., et al. (2010). Bacillus methanolicus pyruvate carboxylase and homoserine dehydrogenase I and II and their roles for L-lysine production from methanol at 50°C. Appl. Microbiol. Biotechnol. 87, 951–964. doi: 10.1007/s00253-010-2559-6
Brautaset, T., Jakobsen,Ø. M. M., Flickinger, M. C., Valla, S., and Ellingsen, T. E. E. (2004). Plasmid-dependent methylotrophy in thermotolerant Bacillus methanolicus. J. Bacteriol. 186, 1229–1238. doi: 10.1128/JB.186.5.1229
Bron, S., Bosma, P., van Belkum, M., and Luxen, E. (1987). Stability function in the Bacillus subtilis plasmid pTA1060 since natural plasmids in Bacillus subtilis. Plasmid 18, 8–15. doi: 10.1016/0147-619X(87)90073-4
Bron, S., and Luxen, E. (1985). Segregational instability of pUB110-derived recombinant plasmids in Bacillus subtilis. Plasmid 14, 235–244. doi: 10.1016/0147-619X(85)90007-1
Bron, S., Luxen, E., and Swart, P. (1988). Instability of recombinant pUB110 plasmids in Bacillus subtilis: plasmid-encoded stability function and effects of DNA inserts. Plasmid 19, 231–241. doi: 10.1016/0147-619X(88)90041-8
Bruand, C., Ehrlich, S. D., and Janniere, L. (1991). Unidirectional theta replication of the structurally stable Enterococcus faecalis plasmid pAM beta 1. EMBO J. 10, 2171–2177.
Chaillou, P., Bor, Y., Batt, C. A., and Pouwels, P. H. (1998). Molecular cloning and functional expression in Lactobacillus plantarum 80 of xylT, encoding the D-xylose-H+ symporter of Lactobacillus brevis. Appl Environ. Microbiol. 64, 4720–4728.
Chalfie, M., Tu, Y., Ward, W. W., Euskirchen, G., and Prasher, D. (1994). Green fluorescent protein as a marker for gene expression. Science 263, 1258–1262. doi: 10.1126/science.8303295
Chang, S., Chang, S.-Y., and Gray, O. (1987). Structural and genetic analyses of a par locus that regulates plasmid partition in Bacillus subtilis. J. Bacteriol. 169, 3952–3962.
Crameri, A., Whitehorn, E. A., Tate, E., and Stemmer, W. P. C. (1996). Improved green fluorescent protein by molecular evolution using DNA shuffling. Nat. Biotechnol. 14, 315–319. doi: 10.1038/nbt0396-315
Crutz-Le Coq, A. M., and Zagorec, M. (2008). Vectors for lactobacilli and other Gram-positive bacteria based on the minimal replicon of pRV500 from Lactobacillus sakei. Plasmid 60, 212–220. doi: 10.1016/j.plasmid.2008.08.002
Cue, D., Lam, H., Dillingham, R. L., Hanson, R. S., and Flickinger, M. C. (1997). Genetic manipulation of Bacillus methanolicus, a gram-positive, thermotolerant methylotroph. Appl. Environ. Microbiol. 63, 1406–1420.
D’Urzo, N., Martinelli, M., Nenci, C., Brettoni, C., Telford, J. L., and Maione, D. (2013). High-level intracellular expression of heterologous proteins in Brevibacillus choshinensis SP3 under the control of a xylose inducible promoter. Microb. Cell Fact. 12:12. doi: 10.1186/1475-2859-12-12
Eikmanns, B. J., Thum-schmitz, N., Eggeling, L., Ludtke, K., and Sahm, H. (1994). Nucleotide sequence, expression and transcriptional analysis of the Corynebacterium glutamicum gltA gene encoding citrate synthase. Microbiology 140, 1817–1828. doi: 10.1099/13500872-140-8-1817
Friehs, K. (2004). Plasmid copy number and plasmid stability. Adv. Biochem. Eng. Biotechnol. 86, 47–82. doi: 10.1007/b12440
Gibson, D. G., Young, L., Chuang, R., Venter, J. C., Hutchison, C. A. III, and Smith, H. O. (2009). Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 6, 12–16. doi: 10.1038/NMETH.1318
Gryczan, T. J., Contente, S., and Dubnau, D. (1978). Characterization of Staphylococcus aureus plasmids introduced by transformation into Bacillus subtilis. J. Bacteriol. 134, 318–329.
Haima, P., Bron, S., and Venema, G. (1987). The effect of restriction on shotgun cloning and plasmid stability in Bacillus subtilis Marburg. Mol. Gen. Genet. 209, 335–342. doi: 10.1007/BF00329663
Hanahan, D. (1983). Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 166, 557–580. doi: 10.1016/S0022-2836(83)80284-8
Hanson, R. S., Dillingham, R. D., Olson, P., Lee, G. H., Cue, D., Schendel, F. J., et al. (1996). “Production of L-lysine and some other amino acids by mutants of B. methanolicus,” in Microbial Growth on C1 Compounds, Vol. 3, eds M. E. Lidstrom and F. Robert Tabita (Amsterdam: Springer), 227–236. doi: 10.1007/978-94-009-0213-8_31
Heggeset, T. M., Krog, A., Balzer, S., Wentzel, A., Ellingsen, T. E., and Brautaseta, T. (2012). Genome sequence of thermotolerant Bacillus methanolicus: features and regulation related to methylotrophy and production of L-lysine and L-glutamate from methanol. Appl. Environ. Microbiol. 78, 5170–5181. doi: 10.1128/AEM.00703-12
Heravi, K. M., Wenzel, M., and Altenbuchner, J. (2011). Regulation of mtl operon promoter of Bacillus subtilis: requirements of its use in expression vectors. Microb. Cell Fact. 10, 1–19. doi: 10.1186/1475-2859-10-83
Higa, A., and Mandel, M. (1970). Calcium-dependent bacteriophage DNA infection. J. Mol. Biol. 53, 159–162. doi: 10.1016/0022-2836(70)90051-3
Hilker, R., Stadermann, K. B., Doppmeier, D., Kalinowski, J., Stoye, J., Straube, J., et al. (2014). ReadXplorer–visualization and analysis of mapped sequences. Bioinformatics 30, 2247–2254. doi: 10.1093/bioinformatics/btu205
Hoffmann, J., and Altenbuchner, J. (2015). Functional characterization of the mannitol promoter of Pseudomonas fluorescens DSM 50106 and its application for a mannitol-inducible expression system for Pseudomonas putida KT2440. PLoS ONE 10:e0133248. doi: 10.1371/journal.pone.0133248
IHS Chemical (2016). IHS Chemical. Available at: http://www.businesswire.com/news/home/20151006005707/en/Global-Demand-Flavor-Enhancer-MSG-Grows-Incomes [accessed April 28, 2016].
Irla, M., Neshat, A., Brautaset, T., Rückert, C., Kalinowski, J., and Wendisch, V. F. (2015). Transcriptome analysis of thermophilic methylotrophic Bacillus methanolicus MGA3 using RNA-sequencing provides detailed insights into its previously uncharted transcriptional landscape. BMC Genomics 16:73. doi: 10.1186/s12864-015-1239-4
Irla, M., Neshat, A., Winkler, A., Albersmeier, A., Heggeset, T. M., Brautaset, T., et al. (2014). Complete genome sequence of Bacillus methanolicus MGA3, a thermotolerant amino acid producing methylotroph. J. Biotechnol. 188C, 110–111. doi: 10.1016/j.jbiotec.2014.08.013
Jacob, S., Allmansberger, R., Giirtner, D., and Hillen, W. (1991). Catabolite repression of the operon for xylose utilization from Bacillus subtilis W23 is mediated at the level of transcription and depends on a cis site in the xylA reading frame. Mol. Gen. Genet. 229, 189–196. doi: 10.1007/BF00272155
Jakobsen,Ø. M., Benichou, A., Flickinger, M. C., Valla, S., Ellingsen, T. E., and Brautaset, T. (2006). Upregulated transcription of plasmid and chromosomal ribulose monophosphate pathway genes is critical for methanol assimilation rate and methanol tolerance in the methylotrophic bacterium Bacillus methanolicus. J. Bacteriol. 188, 3063–3072. doi: 10.1128/JB.188.8.3063
Jakobsen,Ø. M., Brautaset, T., Degnes, K. F., Heggeset, T. M., Balzer, S., Flickinger, M. C., et al. (2009). Overexpression of wild-type aspartokinase increases L-lysine production in the thermotolerant methylotrophic bacterium Bacillus methanolicus. Appl. Environ. Microbiol. 75, 652–661. doi: 10.1128/AEM.01176-08
Joyet, P., Derkaoui, M., Bouraoui, H., and Deutscher, J. (2015). PTS-mediated regulation of the transcription activator MtlR from different species: surprising differences despite strong sequence conservation. J. Mol. Microbiol. Biotechnol. 25, 94–105. doi: 10.1159/000369619
Khan, S. A. (1997). Rolling-circle replication of bacterial plasmids. Microbiol. Mol. Biol. Rev. 61, 442–455.
Kiefer, P., Schmitt, U., Müller, J. E. N., Hartl, J., Meyer, F., Ryffel, F., et al. (2015). DynaMet: a fully automated pipeline for dynamic LC-MS data. Anal. Chem. 87, 9679–9686. doi: 10.1021/acs.analchem.5b01660
Kim, L., Mogk, A., and Schumann, W. (1996). A xylose-inducible Bacillus subtilis integration vector and its application. Gene 181, 71–76. doi: 10.1016/S0378-1119(96)00466-0
Kovács, A. T., van Hartskamp, M., Kuipers, O. P., and van Kranenburg, R. (2010). Genetic tool development for a new host for biotechnology, the thermotolerant bacterium Bacillus coagulans. Appl. Environ. Microbiol. 76, 4085–4088. doi: 10.1128/AEM.03060-09
Krog, A., Heggeset, T. M., Müller, J. E. N., Kupper, C. E., Schneider, O., Vorholt, J. A., et al. (2013). Methylotrophic Bacillus methanolicus encodes two chromosomal and one plasmid born NAD+ dependent methanol dehydrogenase paralogs with different catalytic and biochemical properties. PLoS ONE 8:e59188. doi: 10.1371/journal.pone.0059188
Liu, H., and Naismith, J. H. (2008). An efficient one-step site-directed deletion, insertion, single and multiple-site plasmid mutagenesis protocol. BMC Biotechnol. 8:91. doi: 10.1186/1472-6750-8-91
Lokman, B. C., Leer, R. J., van Sorge, R., and Pouwels, P. H. (1994). Promoter analysis and transcriptional regulation of Lactobacillus pentosus genes involved in xylose catabolism. Mol. Gen. Genet. 245, 117–125. doi: 10.1007/BF00279757
Markert, B., Stolzenberger, J., Brautaset, T., and Wendisch, V. F. (2014). Characterization of two transketolases encoded on the chromosome and the plasmid pBM19 of the facultative ribulose monophosphate cycle methylotroph Bacillus methanolicus. BMC Microbiol. 14:7. doi: 10.1186/1471-2180-14-7
Miyoshi, A., Jamet, E., Commissaire, J., Renault, P., Langella, P., and Azevedo, V. (2004). A xylose-inducible expression system for Lactococcus lactis. FEMS Microbiol. Lett. 239, 205–212. doi: 10.1016/j.femsle.2004.08.018
Müller, J. E. N., Heggeset, T. M., Wendisch, V. F., Vorholt, J. A., and Brautaset, T. (2015a). Methylotrophy in the thermophilic Bacillus methanolicus, basic insights and application for commodity production from methanol. Appl. Microbiol. Biotechnol. 99, 535–551. doi: 10.1007/s00253-014-6224-3
Müller, J. E. N., Meyer, F., Litsanov, B., Kiefer, P., and Vorholt, J. A. (2015b). Core pathways operating during methylotrophy of Bacillus methanolicus MGA3 and induction of a bacillithiol-dependent detoxification pathway upon formaldehyde stress. Mol. Microbiol. 98, 1089–1100. doi: 10.1111/mmi.13200
Müller, J. E. N., Litsanov, B., Bortfeld-Miller, M., Trachsel, C., Grossmann, J., Brautaset, T., et al. (2014). Proteomic analysis of the thermophilic methylotroph Bacillus methanolicus MGA3. Proteomics 14, 725–737. doi: 10.1002/pmic.201300515
Naglich, J. G., and Andrews, R. E. J. (1988). Tn916-dependent conjugal transfer of pC194 and pUB110 from Bacillus subtilis into Bacillus thuringiensis subsp. israelensis. Plasmid 20, 113–126. doi: 10.1016/0147-619X(88)90014-5
Nærdal, I., Pfeifenschneider, J., Brautaset, T., and Wendisch, V. F. (2015). Methanol-based cadaverine production by genetically engineered Bacillus methanolicus strains. Microb. Biotechnol. 8, 342–350. doi: 10.1111/1751-7915.12257. Microb. Biotechnol. 2019, 12, 182–183.
Nguyen, H. D., Nguyen, Q. A., Ferreira, R. C., Ferreira, L. C. S., Tran, L. T., and Schumann, W. (2005). Construction of plasmid-based expression vectors for Bacillus subtilis exhibiting full structural stability. Plasmid 54, 241–248. doi: 10.1016/j.plasmid.2005.05.001
Nilasari, D., Rech, S., and Komives, C. (2012). Expression of recombinant green fluorescent protein in Bacillus methanolicus. Biotechnol. Prog. 28, 662–668. doi: 10.1002/btpr.1522
Notredame, C., Higgins, D. G., and Heringa, J. (2000). T-Coffee: a novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 302, 205–217. doi: 10.1006/jmbi.2000.4042
Ochsner, A. M., Müller, J. E. N., Mora, C. A., and Vorholt, J. A. (2014). In vitro activation of NAD-dependent alcohol dehydrogenases by Nudix hydrolases is more widespread than assumed. FEBS Lett. 588, 2993–2999. doi: 10.1016/j.febslet.2014.06.008
Polak, J., and Novic, R. P. (1982). Closely related plasmids from Staphylococcus aureus and soil bacilli. Plasmid 7, 152–162. doi: 10.1016/0147-619X(82)90074-9
Rhee, M. S., Kim, J., Qian, Y., Ingram, L., and Shanmugam, K. (2007). Development of plasmid vector and electroporation condition for gene transfer in sporogenic lactic acid bacterium, Bacillus coagulans. Plasmid 58, 13–22. doi: 10.1016/j.plasmid.2006.11.006
Rygus, T., and Hillen, W. (1991). Inducible high-level expression of heterologous genes in Bacillus megaterium using the regulatory elements of the xylose-utilization operon. Appl. Microbiol. Biotechnol. 35, 594–599. doi: 10.1007/BF00169622
Sambrook, J., and Russell, D. (2001). Molecular Cloning: A Laboratory Manual, 3 Edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Schendel, F. J., Bremmon, C. E., Flickinger, M. C., and Guettler, M. (1990). L-lysine production at 50 degrees C by mutants of a newly isolated and characterized methylotrophic Bacillus sp. Appl. Environ. Microbiol. 56, 963–970.
Schendel, F. J., Dillingham, R., Hanson, R. S., Sano, K., and Matsui, K. (2000). Production of glutamate using wild type bacillus methanolicus. US 6083728.
Schmiedel, D., and Hillen, W. (1996). Contributions of XylR CcpA and cre to diauxic growth of Bacillus megaterium and to xylose isomerase expression in the presence of glucose and xylose. Mol. Gen. Genet. 250, 259–266. doi: 10.1007/s004380050075
Schneider, J., and Wendisch, V. F. (2011). Biotechnological production of polyamines by bacteria: recent achievements and future perspectives. Appl. Microbiol. Biotechnol. 91, 17–30. doi: 10.1007/s00253-011-3252-0
Seibold, G., Auchter, M., Berens, S., Kalinowski, J., and Eikmanns, B. J. (2006). Utilization of soluble starch by a recombinant Corynebacterium glutamicum strain: growth and lysine production. J. Biotechnol. 124, 381–391. doi: 10.1016/j.jbiotec.2005.12.027
Seyler, R. W. J., Yousten, A. A., and Burke, W. F. Jr. (1991). Plasmid stability in Bacillus sphaericus 2362 during recycling in mosquito larvae. J. Invertebr. Pathol. 58, 362–366. doi: 10.1016/0022-2011(91)90181-O
Shaner, N. C., Campbell, R. E., Steinbach, P. A., Giepmans, B. N. G., Palmer, A. E., and Tsien, R. Y. (2004). Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 22, 1567–1572. doi: 10.1038/nbt1037
Shimizu, H., Tanaka, H., Nakato, A., Nagahisa, K., Kimura, E., and Shioya, S. (2003). Effects of the changes in enzyme activities on metabolic flux redistribution around the 2-oxoglutarate branch in glutamate production by Corynebacterium glutamicum. Bioprocess Biosyst. Eng. 25, 291–298. doi: 10.1007/s00449-002-0307-8
Shimizu, K. (2013). Metabolic regulation of a bacterial cell system with emphasis on Escherichia coli metabolism. ISRN Biochem. 2013:645983. doi: 10.1155/2013/645983
Shoham, Y., and Demain, A. L. (1990). Effect of medium composition on the maintenance of a recombinant plasmid in Bacillus subtilis. Enzym. Microb. Technol. 12, 330–336. doi: 10.1016/0141-0229(90)90160-R
Simpson, F. J., and McCoy, E. (1953). The amylases of five streptomycetes. Appl. Microbiol. 1, 228–236.
Skjerdal, O. T., Sletta, H., Flenstad, S. G., Josefsen, K. D., Levine, D. W., and Ellingsen, T. E. (1996). Changes in intracellular composition in response to hyperosmotic stress of NaCl, sucrose or glutamic acid in Brevibacterium lactofermentum and Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 44, 635–642. doi: 10.1007/BF00172497
Soutsehek-Bauer, E., Scholz, W., Grill, E., and Staudenbauer, W. L. (1987). Thermostability and superhelicity of plasmid DNA in Bacillus stearothermophilus. Mol. Gen. Genet. 209, 575–579. doi: 10.1007/BF00331165
Stolzenberger, J., Lindner, S. N., Persicke, M., Brautaset, T., and Wendisch, F. (2013a). Characterization of fructose 1,6-bisphosphatase and sedoheptulose 1,7-bisphosphatase from the facultative ribulose monophosphate cycle methylotroph Bacillus methanolicus. J. Bacteriol. 195, 5112–5122. doi: 10.1128/JB.00672-13
Stolzenberger, J., Lindner, S. N., and Wendisch, V. F. (2013b). The methylotrophic Bacillus methanolicus MGA3 possesses two distinct fructose 1,6-bisphosphate aldolases. Microbiology 159, 1770–1781. doi: 10.1099/mic.0.067314-0
Tanaka, T., and Sueoka, N. (1983). Site-specific in vitro binding of plasmid pUB110 to Bacillus subtilis membrane fraction. J. Bacteriol. 154, 1184–1194.
te Riele, H., Michel, B., and Ehrlich, S. D. (1986). Single-stranded plasmid DNA in Bacillus subtilis and Staphylococcus aureus. Proc. Natl. Acad. Sci. U.S.A. 83, 2541–2545. doi: 10.1073/pnas.83.8.2541
Titok, M. A., Chapuis, J., Selezneva, Y. V., Lagodich, A. V., Prokulevich, V. A., Ehrlich, S. D., et al. (2003). Bacillus subtilis soil isolates: plasmid replicon analysis and construction of a new theta-replicating vector. Plasmid 49, 53–62. doi: 10.1016/S0147-619X(02)00109-9
Vehmaanperii, J. O., and Korhola, M. P. (1986). Stability of the recombinant plasmid carrying the Bacillus amyloliquefaciens a-amylase gene in B. subtilis. Appl. Microbiol. Biotechnol. 23, 456–461. doi: 10.1007/BF02346060
Wendisch, V. F. (2014). Microbial production of amino acids and derived chemicals: synthetic biology approaches to strain development. Curr. Opin. Biotechnol. 30, 51–58. doi: 10.1016/j.copbio.2014.05.004
Winkler, W. C., Nahvi, A., Roth, A., Collins, J. A., and Breaker, R. R. (2004). Control of gene expression by a natural metabolite-responsive ribozyme. Nature 428, 281–286. doi: 10.1038/nature02362
Wu, T.-Y., Chen, C.-T., Liu, J. T.-J., Bogorad, I. W., Damoiseaux, R., and Liao, J. C. (2016). Characterization and evolution of an activator-independent methanol dehydrogenase from Cupriavidus necator N-1. Appl. Microbiol. Biotechnol. doi: 10.1007/s00253-016-7320-3 [Epub ahead of print].
Yoshida, K., Kobayashi, K., Miwa, Y., Kang, C., Matsunaga, M., Yamaguchi, H., et al. (2001). Combined transcriptome and proteome analysis as a powerful approach to study genes under glucose repression in Bacillus subtilis. Nucleic Acids Res. 29, 683–692. doi: 10.1093/nar/29.3.683
Zaghloul, T. I., Abdelaziz, A., and Mostafa, M. H. (1994). High level of expression and stability of the cloned alkaline protease (aprA) gene in Bacillus subtilis. Enzyme Microb. Technol. 16, 534–537. doi: 10.1016/0141-0229(94)90026-4
Zahoor, A., Lindner, S. N., and Wendisch, V. F. (2012). Metabolic engineering of Corynebacterium glutamicum aimed at alternative carbon sources and new products. Comput. Struct. Biotechnol. J. 3:e201210004. doi: 10.5936/csbj.201210004
Keywords: Bacillus methanolicus, thermophile, methylotroph, genetic tool box, theta-replicating plasmids, gene expression
Citation: Irla M, Heggeset TM, Nærdal I, Paul L, Haugen T, Le SB, Brautaset T and Wendisch VF (2016) Genome-Based Genetic Tool Development for Bacillus methanolicus: Theta- and Rolling Circle-Replicating Plasmids for Inducible Gene Expression and Application to Methanol-Based Cadaverine Production. Front. Microbiol. 7:1481. doi: 10.3389/fmicb.2016.01481
Received: 13 May 2016; Accepted: 06 September 2016;
Published: 22 September 2016.
Edited by:
Kian Mau Goh, Universiti Teknologi Malaysia, MalaysiaReviewed by:
María Sofía Urbieta, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), ArgentinaKheng Oon Low, Malaysia Genome Institute, Malaysia
Copyright © 2016 Irla, Heggeset, Nærdal, Paul, Haugen, Le, Brautaset and Wendisch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Volker F. Wendisch, dm9sa2VyLndlbmRpc2NoQHVuaS1iaWVsZWZlbGQuZGU=