
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 14 September 2016
Sec. Extreme Microbiology
Volume 7 - 2016 | https://doi.org/10.3389/fmicb.2016.01365
This article is part of the Research Topic Recent Advances in Acidophile Microbiology: Fundamentals and Applications View all 11 articles
 Sigde Mamani1,2Danielle Moinier1
Sigde Mamani1,2Danielle Moinier1 Yann Denis3
Yann Denis3 Laurent Soulère4
Laurent Soulère4 Yves Queneau4
Yves Queneau4 Emmanuel Talla1
Emmanuel Talla1 Violaine Bonnefoy1*†
Violaine Bonnefoy1*† Nicolas Guiliani2*†
Nicolas Guiliani2*†While a functional quorum sensing system has been identified in the acidophilic chemolithoautotrophic Acidithiobacillus ferrooxidans ATCC 23270T and shown to modulate cell adhesion to solid substrates, nothing is known about the genes it regulates. To address the question of how quorum sensing controls biofilm formation in A. ferrooxidansT, the transcriptome of this organism in conditions in which quorum sensing response is stimulated by a synthetic superagonist AHL (N-acyl homoserine lactones) analog has been studied. First, the effect on biofilm formation of a synthetic AHL tetrazolic analog, tetrazole 9c, known for its agonistic QS activity, was assessed by fluorescence and electron microscopy. A fast adherence of A. ferrooxidansT cells on sulfur coupons was observed. Then, tetrazole 9c was used in DNA microarray experiments that allowed the identification of genes regulated by quorum sensing signaling, and more particularly, those involved in early biofilm formation. Interestingly, afeI gene, encoding the AHL synthase, but not the A. ferrooxidans quorum sensing transcriptional regulator AfeR encoding gene, was shown to be regulated by quorum sensing. Data indicated that quorum sensing network represents at least 4.5% (141 genes) of the ATCC 23270T genome of which 42.5% (60 genes) are related to biofilm formation. Finally, AfeR was shown to bind specifically to the regulatory region of the afeI gene at the level of the palindromic sequence predicted to be the AfeR binding site. Our results give new insights on the response of A. ferrooxidans to quorum sensing and on biofilm biogenesis.
Due to its low operating cost, biomining is a very successful geobiotechnology that actually produces approximately 15 per cent of the world’s extracted copper (Johnson, 2014). Withstanding low pH and high heavy metal concentrations, Acidithiobacillus species are acidophilic key players in biomining industry recovering valuable metals from sulfidic ores such as copper or gold (Jerez, 2009). However, these bacteria are also involved in Acid Mine/Rock Drainage (AM/RD), which represents a worldwide problem of water pollution, from natural and anthropogenic environments (Johnson, 2009, 2012). Indeed, several studies recently indicated that Acidithiobacillus species play a pivotal and structural role in acidophilic communities ranging from 6°C to 90°C (Chen et al., 2015; Liljeqvist et al., 2015; Menzel et al., 2015). Nevertheless, due to an insufficient understanding of the microbiological processes, most biohydrometallurgical plants operate far from maximum efficiency and natural AM/RD are to a large extent uncontrolled.
Acidithiobacillia has been recently defined as a new class of Proteobacteria in which the genus Acidithiobacillus is the main one characterized (Williams and Kelly, 2013). Actually, the genus Acidithiobacillus encompasses seven closely related Gram-negative, chemolithoautotrophic bioleaching species: (i) Acidithiobacillus thiooxidans, A. caldus, and A. albertensis, which oxidize only reduced inorganic sulfur compounds (RISC) and (ii) A ferrooxidans, A. ferrivorans, A. ferridurans, and A. ferriphilus that oxidize both ferrous iron and RISC (Amouric et al., 2011; Hedrich and Johnson, 2013; Williams and Kelly, 2013; Falagan and Johnson, 2015). It has been well established that all Acidithiobacillus species are able to form biofilms on the surface of ores. This bacterial attachment on the mineral has been reported to increase metal leaching due to the formation of a close and enlarged “reaction space” between the metal sulfide surface and the cell (Pogliani and Donati, 1999; Harneit et al., 2006; Rohwerder and Sand, 2007). Therefore, deciphering molecular mechanisms underlying biofilm formation in acidophilic leaching bacteria has been early pointed out as an important field of investigation.
Quorum sensing (QS) and the secondary messenger c-di-GMP signaling pathway [for recent reviews see (Hengge, 2009; Decho et al., 2011; Kalia et al., 2013; Romling et al., 2013; Hengge et al., 2015)] are the most studied mechanisms controlling biofilm development in bacteria. Both pathways have been shown to be linked in several bacterial species (Ryan et al., 2006; Waters et al., 2008; Ueda and Wood, 2009; Zhang, 2010; Kozlova et al., 2011) and to control more particularly polysaccharide production and biofilm formation (Ueda and Wood, 2009). QS is an important mechanism for the timing of collective behaviors through the regulation of population density-dependent cellular processes, such as the production of virulence factors, motility, exopolysaccharide production and biofilm formation (Parsek and Greenberg, 2005; Waters and Bassler, 2005; Ng and Bassler, 2009). In Gram-negative bacteria, the main characterized QS system involves three key molecular elements (Venturi and Subramoni, 2009): (i) N-acyl homoserine lactones (AHLs), which act as autoinducers (AIs); (ii) the AHLs synthase encoded by a luxI-like gene; (iii) a transcriptional regulator, which is encoded by a luxR-like gene and which binds AHL molecules and modulates the expression of different target genes that constitute the QS regulon. Depending on the bacterial species and also on the experimental strategies (transcriptomic or proteomic), the size of the QS regulons oscillates between 3 and 8% of the identified ORFs (Vasil, 2003; Wagner et al., 2003; Cantero et al., 2006; Qin et al., 2007; Stevens et al., 2011; Majerczyk et al., 2014).
Even if several reports related to biofilm formation regulation by acidophilic bacteria belonging to Acidithiobacillus genus have been released recently (Farah et al., 2005; Bellenberg et al., 2012, 2014; Ruiz et al., 2012; Diaz et al., 2013; Montgomery et al., 2013; Vera et al., 2013; Castro et al., 2015), the molecular cascade involved in exopolysaccharide production and biofilm formation by Acidithiobacillus species is still undeciphered. While c-di-GMP pathway has been identified in all Acidithiobacillus spp. (Ruiz et al., 2012; Diaz et al., 2013; Castro et al., 2015), the species that oxidize only RISC do not possess the genes related to canonical QS systems (Valdés et al., 2008). Indeed, a functional QS system has been reported only in the iron/RISC-oxidizing species A. ferrooxidans (Farah et al., 2005; Rivas et al., 2005; Valenzuela et al., 2007). In addition, it has been recently reported that the RISC-oxidizing species A. thiooxidans cannot adhere to pyrite if this mineral is not previously colonized by an iron-oxidizing species (Bellenberg et al., 2014) pointing out A. ferrooxidans as a key player for mineral colonization.
Acidithiobacillus ferrooxidans ATCC 23270T QS system involves two divergent genes afeI and afeR coding for the AHL synthase and the transcriptional regulator, respectively (Farah et al., 2005). AfeR has the conserved amino acid residues located in the active site of LuxR-protein family and possesses the canonical AHL and DNA binding domains based on a 3D-structural model (Soulere et al., 2008). In A. ferrooxidans ATCC 23270T, nine different AHL molecules are synthesized with medium or large acyl side chains (Valenzuela et al., 2007). In this strain, transcription of afeI is increased under the physiological conditions that promote biofilm formation, such as growth in the presence of sulfur (solid energetic substrate) or in low phosphate medium (Farah et al., 2005), suggesting a role of QS system in the attachment of A. ferrooxidans to ores (e.g., pyrite). In agreement with this hypothesis, addition of synthetic AHL that are AIs naturally synthesized by A. ferrooxidans such as C14-AHL and 3-hydroxy-C14-AHL has been shown to enhance A. ferrooxidans ATCC 23270T cell adhesion, exopolysaccharide production and biofilm development on elemental sulfur and pyrite (Ruiz et al., 2008; Gonzalez et al., 2013).
However, to date this phenotypic result is still uncoupled with genotypic data that will allow the understanding of the molecular chain reaction going from the AHL-sensing by AfeR to ore colonization. A bioinformatics analysis has recently allowed the identification of a putative QS regulon in A. ferrooxidans ATCC 23270T that encompasses 75 possible AfeR target-genes, including genes likely involved in polysaccharide biosynthesis (Banderas and Guiliani, 2013). However, biological data are required to fully identify the A. ferrooxidans genes whose expression is modulated by AHL signaling.
Here, we report the first biological study focused on deciphering the QS regulon of A. ferrooxidans ATCC 23270T. The effects of AI 3-hydroxy-C14-AHL and of tetrazolic AHL-analog 9c, on A. ferrooxidans adhesion to sulfur were first compared by fluorescence and scanning electronic microscopy. Then, DNA microarray experiments were performed to compare total RNA of A. ferrooxidans ATCC 23270T cells induced or not by tetrazole 9c. These allowed the identification of 141 genes from which at least 48 can be linked with QS pathway, exopolysaccharide production and biofilm development. If we include the genes encoding hypothetical proteins that colocalized and are coregulated with these 48 genes, this number would increase to 60 and represents 1.9% of the ATCC 23270T genome.
Acidithiobacillus ferrooxidans ATCC 23270T was used throughout this study. Escherichia coli TG1 [(supE, hsdΔ5, thi, Δ (lac-proAB), F’:traD36, proAB+, lacIq, lacZΔM15) was used for plasmid propagation. Rosetta (DE3)/pLysS strain (F- ompT hsdSB(rB- mB-) gal dcm λ [DE3 (lacI lacUV5-T7 gene 1 ind1 sam7 nin5)] pLysSRARE (CamR)] and the pET21 plasmid from Novagen were used to produce the recombinant AfeR with a hexahistidine tag fused to its C terminus (AfeR-Histag).
Acidithiobacillus ferrooxidans was grown at 30°C under oxic conditions in modified 9K medium [(0.1 g L-1 NH4)2SO4, 0.4 g L-1 MgSO4⋅7H2O; 0.04 g L-1 K2HPO4, pH 2,5] with sulfur (S0) coupons (0.5 cm2 obtained by S0 fusion) for fluorescence and electron microscopy or 200 g L-1 S0 prills for real-time PCR or microarrays analysis (Amaro et al., 1991) in the presence (5 μM) or the absence of the AHL analogs. The ferrous iron [Fe(II)] growth conditions were described in (Yarzabal et al., 2003). E. coli strains were usually grown at 37°C under oxic conditions in Luria-Bertani broth (LB) supplemented with 100 μg ml-1 ampicillin and 34 μg ml-1 chloramphenicol when necessary (Ausubel et al., 1998).
Due to its high agonistic effect reported on Vibrio fischeri QS system (Sabbah et al., 2012), the tetrazolic AHL analog (tetrazole 9c; Supplementary Figure S1) was selected to test its biological activity on biofilm formation by A. ferrooxidans. It was synthesized according to the protocol described by Sabbah et al. (2012). Briefly, this synthesis was achieved from racemic α-amino-γ-butyrolactone hydrobromide that was acylated with heptanoyl chloride. The intermediate was then cyclized with sodium azide (Biot et al., 2004) to afford tetrazole 9c (Supplementary Figure S1). A. ferrooxidans natural AI 3-hydroxy-C14-AHL was also obtained by chemical synthesis according to the protocol described previously (Chhabra et al., 2003).
Experimental procedures have been previously described (Gonzalez et al., 2013). A. ferrooxidans was grown at 30°C in modified 9K medium (Ruiz et al., 2012) at pH 2.5 with 5% (wt/vol) sulfur (S0) prills. To assess adhesion levels, sterilized S0 coupons were initially added to cell cultures. S0 coupons were daily extracted from day 1 (lag phase) to day 7 (end of the exponential phase) and adhered cells were fixed. Staining was performed with fluorochrome Syto9 for epifluorescence microscopy observations. Epifluorescence visualizations of stained coupons were performed by using fluorescence microscope (ZEISS Axiovert 200 M) equipped with a filter set 10 (FITC, emission BP 515–565) and 20 (Rhodamine, emission BP 575–640) and a digital microscope camera (Axiocam ZEISS). For scanning electronic microscopy (SEM) visualizations, S0 coupons colonized by A. ferrooxidans cells were submitted to critical point drying to avoid cell shrinking and damage. Then, dried samples were coated with a thin conductive film of gold and analyzed with a scanning electron microscope (HITACHI TM 3000, Japan) at the Pontificia Universidad Católica de Chile.
Genomic DNA from A. ferrooxidans was prepared with the NucleoSpin Tissue kit (Macherey Nagel). Plasmid DNA was obtained using a Wizard Plus SV DNA purification system from Promega. DNA digestions with restriction enzymes and ligation with T4 DNA ligase were performed according to New England BioLabs’ recommendations. Primers (Sigma) used in this study are described in Supplementary Table S1. For routine PCR, Go Taq polymerase (Promega) was used. For afeR cloning, PCR amplifications were carried out with Platinum Taq polymerase (Invitrogen) on genomic DNA. DNA products were analyzed on an 1% agarose gel, then concentrated and purified using Amicon® Ultra-0.5 centrifugal filter units (Millipore). Recombinant plasmids were introduced into E. coli competent cells as previously described (Chung and Miller, 1988).
Nucleotide sequence of the amplified DNA was determined by GATC Biotech (Germany).
To get reproducible results, the following experimental growth protocol was performed. The starting inoculum was obtained by growing 1 × 107 cells on 150 ml Fe (II) medium for 3 days. From this culture, 1 × 107 cells were washed three times with basal salts to remove iron traces and inoculated in 250 ml 9K modified medium containing 200 g L-1 S0 prills for 5–6 days (adaptation step). This culture was used to inoculate the same medium (400 ml) for 4 days (pre-inoculum step). This step was repeated in larger volumes in the presence of superagonist AHL analog (adding 5 μM tetrazole 9c) and in its absence (adding DMSO which is the tetrazole 9c solvent) and the cultures were grown for 2, 3, and 4 days.
The cultures were centrifuged at low speed (1,000 rpm, 5 min) to recover S0 prills. Planktonic cells were harvested from the supernatant by centrifugation and washed several times with acid water (pH 1.5) to remove S0. To get sessile cells, the collected S0 prills were washed several times with acid water to remove the remaining planktonic cells. Then, S0 prills were incubated for 5 min in acid water with 0.04% Triton X-100. They were vortexed every min and then, sonicated every 4 sec for 2 min at 4°C to recover adhered cells. S0 prills were removed by low speed centrifugation (1,000 rpm, 5 min). Sessile cells, harvested by centrifugation from the supernatant, were washed three times with acid water to remove Triton X-100.
Acidithiobacillus. ferrooxidans total RNA was extracted from planktonic and sessile cells by using a modified acid-phenol extraction method (Aiba et al., 1981) according to Quatrini et al. (2006, 2009). The modifications included a preliminary TRIZOL® reagent (Invitrogen) extraction step, a final purification step with the High Pure RNA isolation kit (Roche Applied Biosystem) and DNAse I treatments [twice with the DNAse I provided in the kit and once with the reagents from a Turbo DNA-free kit (Applied Biosystems)]. The lack of DNA contamination was checked by PCR on each RNA sample. The RNA integrity was controlled on an agarose gel.
The relative expression levels of the afeI, afeR, zwf, AFE_0233, and AFE_1339 genes were compared to that of the 16S rRNA rrs gene used as a reference standard by quantitative real-time PCR. RNAs were extracted from planktonic cells grown on S0 prills after 2, 3, and 4 days of growth and from sessile cells after 3 days of growth on sulfur prills, as described below. The real-time PCR analysis was performed on a CFX96 real-time PCR detection system with the C1000TM thermal cycler (BioRad) with the “SsoFast EvaGreen Supermix 2X” kit (Bio-Rad) following the manufacturer’s instructions and as described in (Slyemi et al., 2013). The results were analyzed with the Bio-Rad CFX Manager Software 3.0. The real-time quantitative PCR experiments were performed on RNA extracted from at least three independent cultures and duplicated for each RNA preparation with the oligonucleotides listed in Supplementary Table S1. The calculated threshold cycle (Ct) for each gene was normalized to the Ct of the rrs gene. The results are expressed in arbitrary units.
The complete genome (gene annotations and sequences) of A. ferrooxidans ATCC 23270T chromosome was downloaded from the NCBI ftp site1. The OliD program (Talla et al., 2003) was used to design oligonucleotide probe sequences matching defined criteria. An effort was placed to design oligonucleotide probes of similar lengths, with the aim to reduce cross-hybridization between related sequences. Most oligonucleotides are 55 nt long with predicted melting temperatures between 80–100°C in standard hybridization buffer (G + C contents between 30 and 70%). Oligonucleotides were selected such as to avoid self-complementary structures at 65–70°C, and cross-hybridization with the rest of the genome, and were positioned less than 1500 bp upstream of the stop codon of the CDS. The program successfully designed specific oligonucleotide probes for 3044 protein encoding genes, representing 96.7% of the total number of genes. Due to the high similarity with other sequenced regions of the ATCC 23270T genome, 103 genes (3.3%) failed to be represented by a specific oligonucleotide probe. When possible, each gene was represented by two distinct oligonucleotide probes separated by a minimum of 100 nucleotides. A total of 6294 probes from 3147 genes were thus designed. The probes were spotted twice on slides using the Agilent technology2. The array design, the experimental design, and the data for all hybridizations are available in Array Express database under accession numbers A-MTAB-592 and E-MTAB-4896.
Twelve independent hybridizations using total RNA obtained from three different cultures grown without or with 5 μM of tetrazole 9c were performed on Agilent microarrays. Total RNA was used for the synthesis of cDNA fluorescent labeled with Cy®3 and Cy®5 as previously described (Quatrini et al., 2006, 2009). Microarray hybridizations were performed at 42°C for 16 h in a microarray hybridization chamber (Agilent G2534A) following the manufacturer’s instruction. Slides were washed in washing buffer serial dilutions. Arrays were scanned for the Cy®3 and Cy®5 fluorescent signals using an Axion 4400A scanner (Molecular Devices). The data were analyzed with the image quantification software package GenePix Pro 6.0 (Axon Instruments, Inc.) as previously described (Quatrini et al., 2006, 2009). Each gene expression ratio was calculated from 12 values calculated from three biological and four technical replicates and normalized using Acuity 4.0 package (Molecular Devices). Only the four best hybridizations (in term of reproducibility) out of the six were taken into account. Genes with weak expression (median intensity <250) were discarded. A onefold deviation from the 1:1 hybridization (log2) ratio (corresponding to twofold change) was taken as indicative of differential gene expression in the conditions analyzed. The values of one Sample t-test – Benjamini–Hochberg (Adv) ≤0.05 (corresponding to 95% confidence) for at least one oligonucleotide were considered statistically significant. Only the genes filling the conditions described above were analyzed. Hierarchical cluster analysis (Pearson correlation, average linkage) was performed using Genesis software suit (Peterson et al., 2001).
Bioinformatic analyses were performed with the tools available in the MaGe annotation platform3 (Vallenet et al., 2013).
The protein concentration was determined by the modified Bradford method (Bio-Rad protein assay). The purity of the preparation was checked by 12.5% SDS-PAGE stained with Coomassie blue and by immunodetection with antibodies directed against the hexa-histidine tag using a SuperSignal West Hisprobe kit (Thermo Scientific) following the manufacturer’s instructions.
To produce wild-type AfeR fused to a hexa-histidine tag at the C-terminus, the DNA fragment corresponding to the AfeR peptide was amplified by PCR with the AFERC1 and AFERC2 oligonucleotides (Supplementary Table S1). The amplified product was digested with HindIII and XhoI and cloned into pET21 to give pET21-AfeR-Histag plasmid. Cloning was done in E. coli TG1 strain. The construction was checked by nucleotide sequencing with the petT7 and T7ter oligonucleotides (Supplementary Table S1). The recombinant plasmid was then introduced into E. coli Rosetta (DE3)/pLysS strain.
The Rosetta (DE3)/pLysS strain carrying pET21-AfeR-Histag was grown at 37°C with 100 μg ml-1 ampicillin and 34 μg ml-1 chloramphenicol to an OD600 of 0.6. Ampicillin (100 μg ml-1) and 3-hydroxy-C14-AHL (Gonzalez et al., 2013) to a final concentration of 1 μM were then added. Cells were grown 30 min at 30°C. At this stage, 0.4 mM IPTG was added and the culture was grown for a further 3 h at 30°C. The cells were harvested by centrifugation and stored at -80°C until use.
To lyse the cells, the cell pellet previously resuspended in lysis solution [50 mM Tris-HCl pH 7.4, 300 mM NaCl, 5 mM imidazole, 2% Tween-20, 1 mM phenylmethylsulfonyl fluoride (PMSF), 0.1 mg ml-1 DNase, 0.1 mg ml-1 lysosyme, and 5 μM 3-hydroxy-C14-AHL] was incubated 30 min at 4°C with gentle shaking and then sonicated. Inclusion bodies, unbroken cells, and cellular debris were removed by centrifugation at 13,000 rpm for 30 min at 4°C. The pellet was dissolved with 4 M urea in 40 mM sodium phosphate pH 7.4, 300 mM NaCl, 1 mM PMSF, 5 μM 3-hydroxy-C14-AHL, and 0.1 mg ml-1 DNase, kept on ice for 30 min with gentle stirring, and then centrifuged at 13,000 rpm for 20 min at 4°C. The supernatant, corresponding to the solubilized inclusion bodies, was filtered through a 0.45 μm membrane before loading onto a cobalt column (HisTrapTM Talon®; GE Healthcare) according to the manufacturer’s instructions. The fractions were eluted with 5, 50, 150, 250, and 500 mM imidazole, in 40 mM sodium phosphate pH 7.4, 300 mM NaCl, 1 mM PMSF, 4 M urea, and 25 μM 3-hydroxy-C14-AHL buffer. The 150 mM fractions containing the recombinant AfeR-His tag was dialysed with decreasing urea concentrations (2 M, 0.5 M, 0 M) in 50 mM HEPES pH 8, 150 mM NaCl, 10 mM DTT, and 5 μM 3-hydroxy-C14-AHL. These fractions were kept at 4°C until use.
DNA substrates for band shift assays were produced by PCR amplification with PrimeSTAR Max DNA Polymerase (Clontech) using 5′ Cy5-labeled reverse oligonucleotide (Sigma; Supplementary Table S1). The Cy5 labeled DNA (2.3 ng) was incubated in a total volume of 10 μl with increasing concentrations of the enriched recombinant AfeR-Histag preparation as indicated in the Figure. The binding reaction contained 20 mM Tris-HCl pH 8, 50 mM KCl, 1 mM DTT, 0.05 % Nonidet P40, 1 mM EDTA, 10 % glycerol, 5 μM 3-hydroxy-C14-AHL, 30 ng μl-1 herring sperm, and bovine serum albumin 100 μg ml-1. After 30 min at room temperature, the reaction mixtures were separated by electrophoresis on a 6% native polyacrylamide gel previously prerun 5 min and run for 1–2 h more in 25 mM Tris-HCl pH 8.3, 0.19 M glycine, 1 mM EDTA, 200 μM spermidine at 30 mA at 4°C. The gel was then scanned using a 635 nm laser and a LPR filter (FLA5100, Fujifilm).
To develop biological strategies for improving biomining activities and preventing environmental damages caused by AM/RD, it is well documented that mineral colonization by acidophilic bacteria such as Acidithiobacillus species is a key step to decipher (Rohwerder and Sand, 2007). If synthesis of specific exopolysaccharides rich in α-mannopyranosyl and α-glucopyranosyl sugar residues has been revealed by fluorescently labeled lectin Concanavalin A within 1 day for EPS (extracellular polymeric substances)-deficient ferrous-iron grown cells after transfer to cultures with pyrite as sole nutrient (Bellenberg et al., 2014), a clear understanding of the molecular cascade involved in exopolysaccharide production and biofilm formation by Acidithiobacillus species is actually missing. However, as a molecular relationship between QS and cell adhesion has been clearly established in A. ferrooxidans (Gonzalez et al., 2013) and it has to be pointed out that the canonical QS systems are missing in Acidithiobacillus species that can oxidize only RISC (Valdés et al., 2008), the iron-oxidizing species such as A. ferrooxidans as primary colonizers are now considered fundamental players for mineral colonization by the bioleaching community. Therefore, to address the question of how A. ferrooxidans regulates the physiological processes involved in cell adhesion, EPS production and biofilm formation, we focused on the deciphering of the QS molecular network by using a synthetic QS-activator molecule and DNA array technology.
To further investigate the molecular mechanisms underlying this pathway, we first challenged the identification of synthetic AHL analogs capable to induce better A. ferrooxidans cell adhesion than natural AIs previously tested (Gonzalez et al., 2013). Thus, a tetrazolic derivative that displays a much higher affinity to the LuxR protein than the natural AI and acts as a superagonist of AHL signaling molecules (Sabbah et al., 2012) was tested. Its effect on biofilm formation by A. ferrooxidans was compared to the natural AI 3-hydroxy-C14-AHL (Figure 1). Growth curves revealed that both tetrazolic AHL analog and 3-hydroxy-C14-AHL have no effect on A. ferrooxidans growth compared to the control in the absence of exogenous AHL (Figure 1A). Fluorescence (Figure 1B) and SEM (Figure 1C) clearly indicated that tetrazole 9c also promoted cell adhesion. Moreover, confirming in the A. ferrooxidans model the superagonistic behavior of tetrazole 9c previously found in V. fisheri (Sabbah et al., 2012), the results obtained on day 3 strongly suggest that tetrazole 9c is biologically more efficient than the natural AI 3-hydroxy-C14-AHL in promoting biofilm formation (Figure 1C).
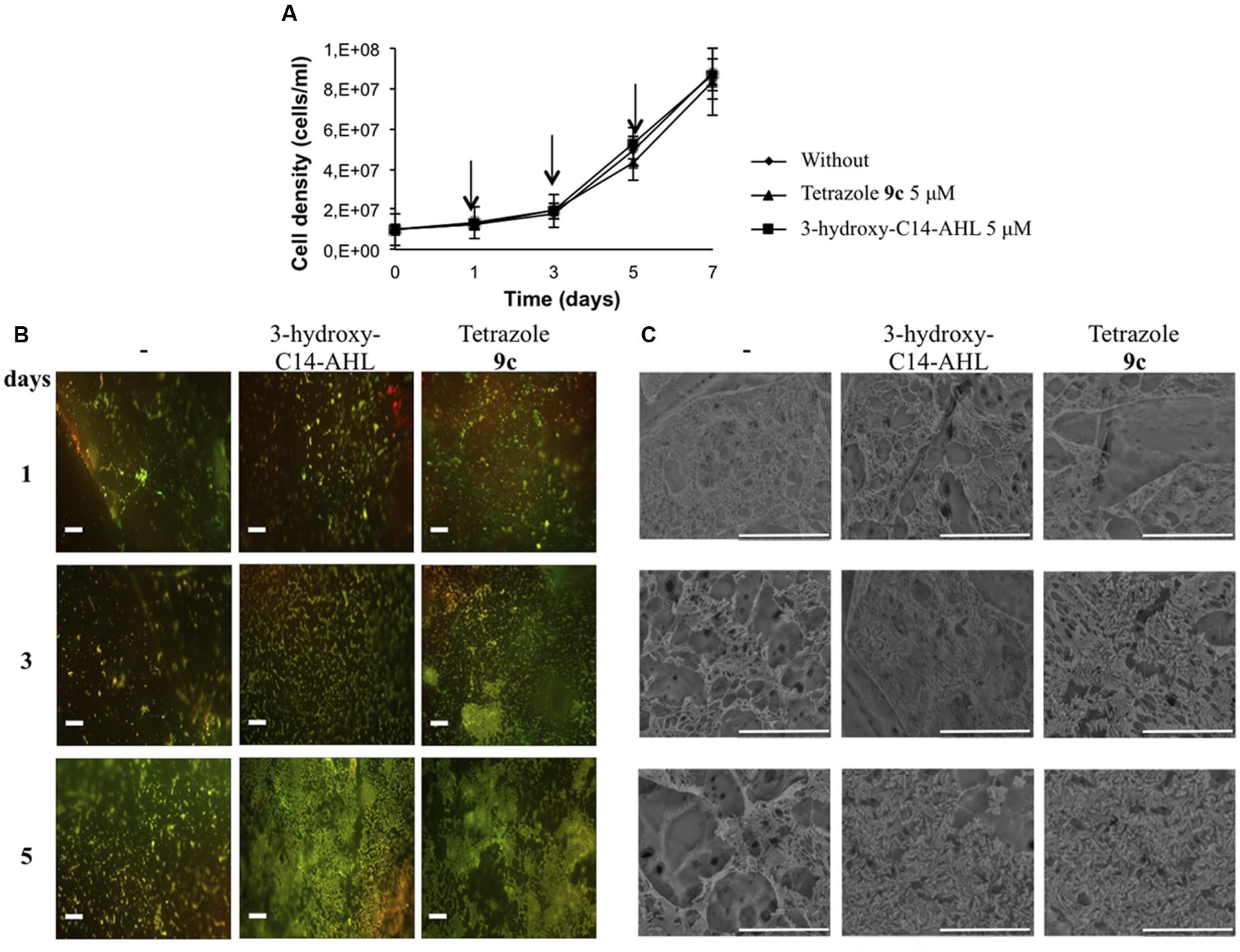
FIGURE 1. Effect of tetrazole 9c on biofilm formation. (A) Growth curves in the absence or the presence of 5 μM 3-hydroxy-C14-AHL or tetrazole 9c. Arrows indicate when aliquots were sampled for microscopy analysis. (B) Fluorescence microscopy of sulfur coupons after 1, 3, or 5 days of Acidithiobacillus ferrooxidans ATCC 23270T cells grown in the absence (-) or the presence of 5 μM 3-hydroxy-C14-AHL or tetrazole 9c. Cells were stained with fluochrome Syto9. (C) Electron microscopy of sulfur coupons treated as above. White bars represent 20 μm.
The results presented in Figure 1 suggest that QS was triggered by 5 μM tetrazole 9c between 2 and 3 days of growth versus 4–5 days in the absence of this AHL analog. To assess whether the tetrazole 9c indeed switched on QS system by inducing the transcription of the genes known to be involved in QS response (Farah et al., 2005), i.e. afeI (AFE_1999) and afeR (AFE_1997), the transcription of these genes was analyzed by quantitative real-time PCR after 2, 3, and 4 days of growth in the presence or the absence of 5μM tetrazole 9c. The results indicated that afeR expression was constitutively expressed under the conditions analyzed, while afeI was induced by tetrazole 9c from the third day of growth in planktonic cells (Table 1).
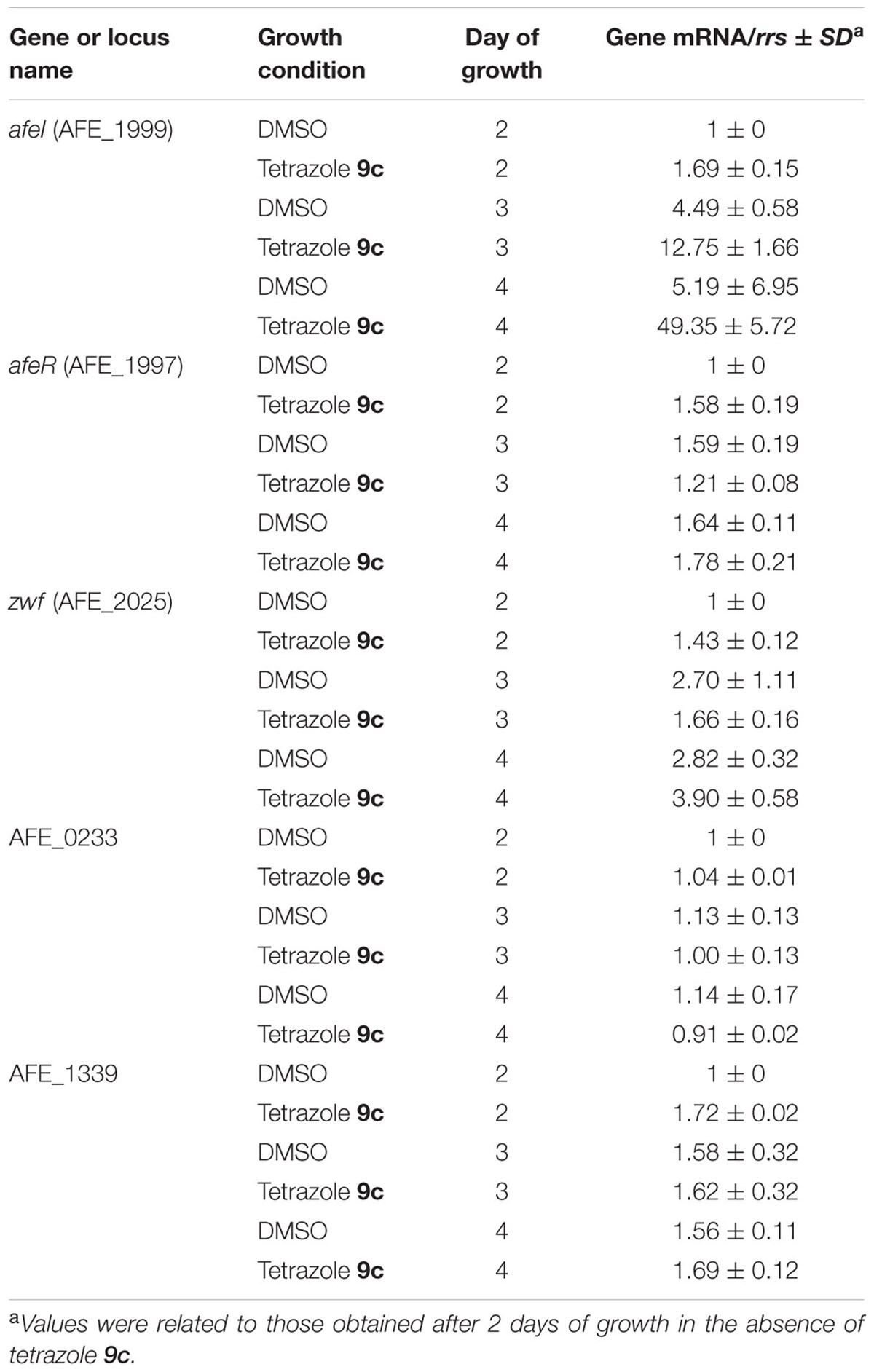
TABLE 1. Quantitative real-time PCR expression data for afeI, afeR, zwf, AFE_0233 (glycosyl transferase), and AFE_1339 (putative polysaccharide export protein) genes from Acidithiobacillus ferrooxidans ATCC 23270T planktonic cells grown with sulfur prills in the presence or the absence of 5 μM tetrazole 9c after 2, 3, and 4 days of growth.
Biofilm formation after 3 days was strongly enhanced in cells treated with 5 μM tetrazole 9c compared to cells from control experiments without agonist (Figure 1). Therefore, expression of some genes predicted to be linked to EPS biosynthesis [zwf (AFE_2025), AFE_0233, and AFE_1339] was also monitored in planktonic (Table 1) and sessile (Supplementary Table S2) cells after 3 days of growth with 5 μM tetrazole 9c. The gene zwf encodes glucose-6-phosphate 1-dehydrogenase that is involved in the intracellular levels of glucose-6P, a precursor of the EPS. AFE_0233 encodes a glycosyl transferase and is located in a gene cluster predicted to encode cell wall constituents (polysaccharides, and lipopolysaccharides). AFE_1339 encodes the putative polysaccharide export protein Wza and is located close to the gal operon proposed to be involved in the formation of EPS in iron-grown cells (Barreto et al., 2005). Besides, AfeR-AHL binding sites were predicted in the regulatory region of zwf, AFE_0233, and AFE_1339 (Banderas and Guiliani, 2013). Surprisingly, tetrazole 9c had no effect on AFE_0233, AFE_1339 and zwf transcription and only the expression of the afeI gene was induced by tetrazole 9c (Table 1; Supplementary Table S2). These data indicate that afeI, and not afeR, is regulated by QS and suggest either that zwf, AFE_0233, and AFE_1339 genes were not regulated by AfeR or that their expression was induced later during biofilm biogenesis.
Quorum sensing response and biofilm formation were obvious within 3 days of growth in the presence of the tetrazolic AHL analog 9c (Figure 1; Table 1). Consequently, total RNAs from planktonic and sessile cells of A. ferrooxidans ATCC 23270T were isolated from 3-days cultures in the presence or the absence of the superagonist AHL analog 9c. They were used to probe gene expression using microarrays displaying two specific oligonucleotides for each gene of this bacterium (3147 predicted genes). Only the genes filling the conditions described in the Materials and Methods section were analyzed. It has to be pointed out that the microarray and quantitative real-time PCR data agreed with the constitutive expression of afeR, zwf, AFE_0233, and AFE_1339 genes under the conditions tested (Table 1; Supplementary Tables S2–S4).
In planktonic cells, a total of 133 genes were differentially expressed, 34 induced and 99 repressed by tetrazole 9c (Supplementary Table S3). In sessile cells under the same conditions, only eight genes presented significant differences in expression, four induced and four repressed by tetrazole 9c (Supplementary Table S4). Therefore, 141 genes were QS regulated, which represent 4.5% of the total number of A. ferrooxidans gene analyzed in this study (see Materials and Methods). These genes were grouped according to their COG classification. Their percentage relative to all the A. ferrooxidans ATCC 23270T genes present in the same COG class is given in Table 2. In planktonic cells, mainly the genes involved in inorganic ion transport and metabolism (4.86%), and nucleotide transport and metabolism (3.39%) were induced in the presence of tetrazole 9c. Mainly those involved in carbohydrate transport and metabolism (11.11%), posttranslational modification, protein turnover, chaperones (8.27%), energy production and conversion (5.76%), cell motility (3.70%), and transcription (2.92%) as well as poorly characterized proteins (11%) were repressed by this AHL analog. In sessile cells, mainly induction by tetrazole 9c of secondary metabolites biosynthesis, transport and catabolism (1.61%), and signal transduction mechanisms (1.15%) was observed while repression was detected for energy production and conversion genes (1.05%). Only the genes differentially expressed in cells that were cultivated with or without the tetrazolic AHL analog and which have known or reliable predicted function are presented in Table 3 for the planktonic cells and in Table 4 for the sessile cells and are discussed below.
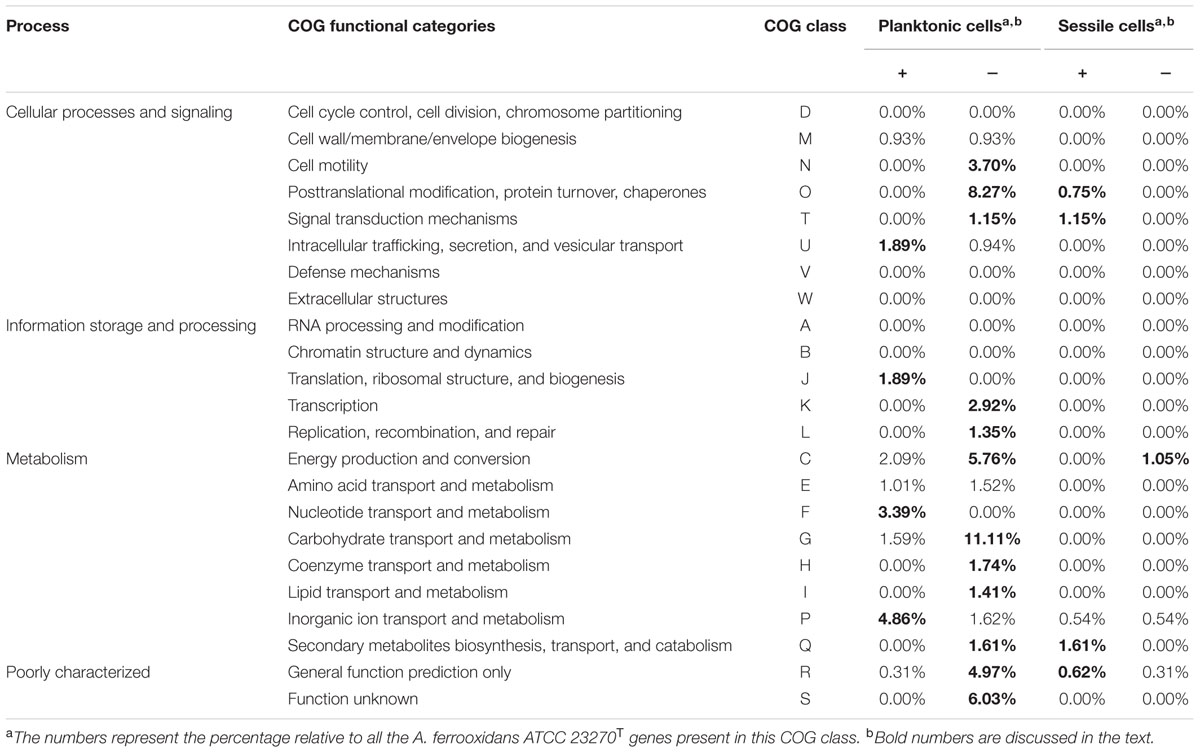
TABLE 2. COG classification of the genes differentially expressed in planktonic and sessile cells grown with (+) and without (-) tetrazole 9c.
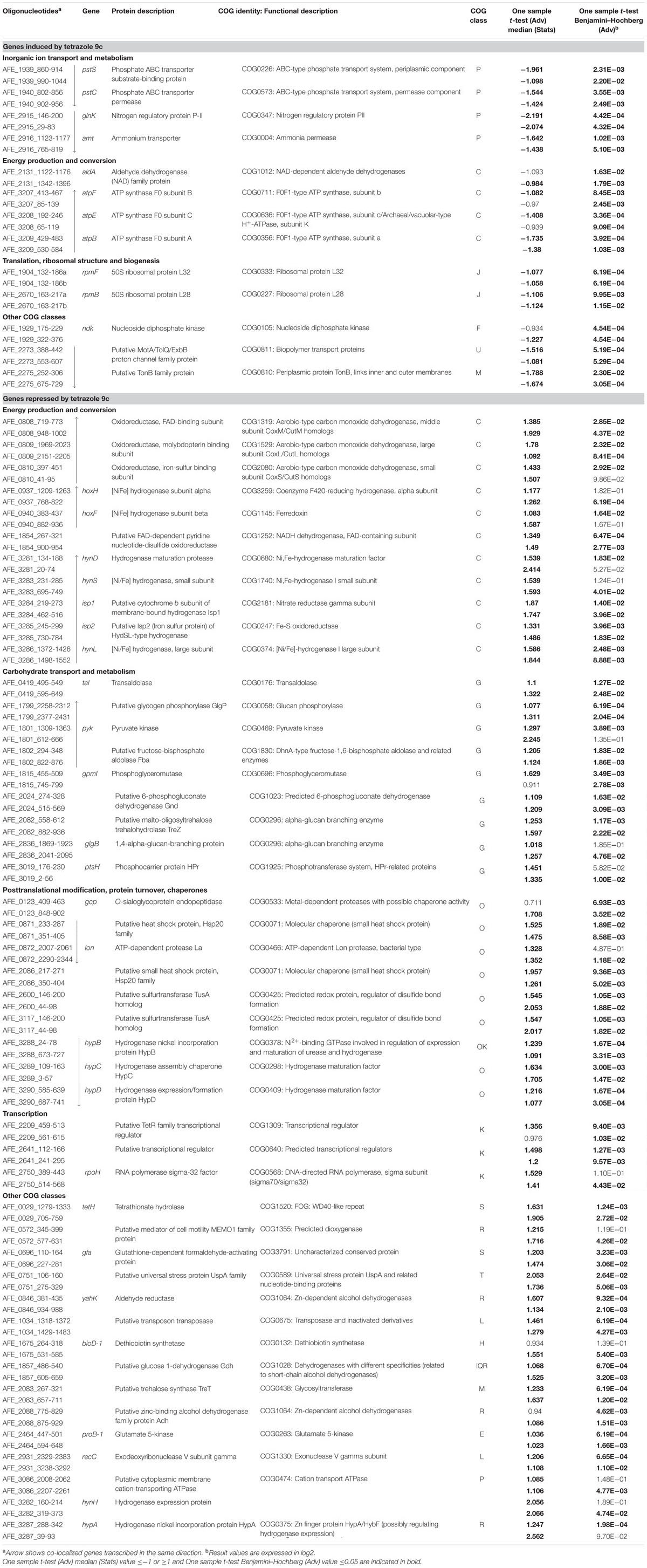
TABLE 3. Microarray expression data for genes with known or predicted function differentially expressed in planktonic cells in the presence of tetrazole 9c.

TABLE 4. Microarray expression data for genes with known or predicted function differentially expressed in sessile cells in the presence of tetrazole 9c.
In planktonic cells, tetrazole 9c modified the expression of a number of genes related to biofilm formation, few being induced and several repressed. Among the induced genes, those involved in inorganic ion transport and energy conversion were mainly found. Not surprisingly, genes involved in the transport of phosphate [pstS (AFE_1939) and pstC (AFE_1940)] and ammonium [glnK (AFE_2915) and amt (AFE_2916)] were upregulated. The phosphate specific transport (Pst) system is known to be important in biofilm formation in a number of bacteria [see (O’May et al., 2009; Heindl et al., 2014) and references therein] including Leptospirillum ferrooxidans (Moreno-Paz et al., 2010) and A. ferrooxidans (Vera et al., 2013), in which phosphate metabolism was early linked to QS regulatory pathway (Farah et al., 2005). Deep cDNA sequencing experiments also revealed that several genes related to ammonium metabolism (amt-1, amt-2, and glnK-1) were upregulated in A. ferrooxidans planktonic cells induced by hydroxyl-C14-AHL compared to not induced (unpublished data). Biofilm formation occurs also in response to the availability of nutrients supplied by the ammonium transporter (AFE_2916) which expression is regulated by GlnK (AFE_2915), as shown recently in Streptococcus mutans (Ardin et al., 2014). This might anticipate gradient of inorganic ions within and around microbial biofilm. The other gene class that was induced by tetrazole 9c in planktonic cells is involved in energy production and conversion, in particular the genes atpBEF (AFE_3207–3209) encoding the membrane-embedded proton channel F0 of the ATPase. This upregulation could allow more protons to pass through the ATP synthase complex generating a proton motive force (PMF) rather than ATP. PMF is required not only for early biofilm formation (Saville et al., 2011), but also in influx and efflux involved in QS since PMF inhibition enhances the intracellular accumulation of AHL leading to decrease in biofilm formation (Ikonomidis et al., 2008; Varga et al., 2012). Along the same lines, genes encoding a putative MolA/TolQ/ExbB proton channel family protein (AFE_2273) and TonB family protein (AFE_2275) were upregulated in the presence of the tetrazole 9c and could contribute to PMF-dependent import through the outer membrane of substrates necessary for QS and/or early EPS synthesis. Another interesting gene that was more expressed in the presence of the tetrazolic AHL analog in planktonic cells is ndk (AFE_1929) encoding a nucleoside diphosphate kinase. A ndk knockout mutant of Pseudomonas aeruginosa was shown to be deficient in polysaccharide synthesis (Kapatral et al., 2000), because it was unable to provide GTP necessary for the incorporation of mannuronate in alginate. It is therefore possible that nucleotide triphosphates are required in an early step of A. ferrooxidans EPS biosynthesis.
The genes that were repressed in the presence of the tetrazolic AHL analog in planktonic cells were mainly involved in energy production and conversion, carbohydrate transport and metabolism, posttranslational modification, protein turnover, chaperones, and transcription. Most of the energy production and conversion class genes encoded two out of the four hydrogenases described in A. ferrooxidans. One is a group one membrane-bound respiratory enzyme enabling the cell to use H2 as an energy source [hynS (AFE_3283) and hynL (AFE_3286)]. The genes encoding this hydrogenase physiological partners [isp1 (AFE_3284) and isp2 (AFE_3285)] and biogenesis machinery [hynD (AFE_3281), hynH (AFE_3282), hynL (AFE_3286), hypA (AFE_3287), hypB (AFE_3288), hypC (AFE_3289), and hypD (AFE_3290)] were also repressed under this condition. The second hydrogenase is a sulfhydrogenase, a group 3b cytoplasmic hydrogenase [hoxH (AFE_0937) and hoxF (AFE_0940)], with both hydrogenase and sulfur reductase activities, likely serving as an electron sink under highly reducing conditions by recycling redox cofactors using either protons or polysulfides as the electron acceptor. It is worth mentioning that, in different bacteria, some hydrogenases were shown to be upregulated in sessile cells, others in planktonic cells (Caffrey et al., 2008; Clark et al., 2012; Kassem et al., 2012). Our data suggest that the groups 1 and 3 hydrogenases of A. ferrooxidans are specific to the non-attached cells.
The number of genes belonging to the carbohydrate transport and metabolism class that were differentially expressed with/without tetrazole 9c agrees with an alteration in the carbon flow when planktonic cells switched to sessile state. It has to be pointed out that all these genes were downregulated in the presence of the tetrazole 9c. Three pathways seemed to be affected: the glycolysis [pyk (AFE_1801), AFE_1802, gpmL (AFE_1815)], the pentose phosphate pathway [tal (AFE_0419), AFE_1857, and AFE_2024] and the glycogen biosynthesis/degradation pathway [AFE_1799, AFE_2082, AFE_2083, and glgB (AFE_2836)]. In the case of glycolysis, this could mean that the pathway was directed toward β-D-fructose-1,6-bisphosphate, β-D- fructose-6-phosphate, α-D-glucose-6-phosphate, and α-D-glucose-1-phosphate production (Figure 2A). Similarly, in the pentose phosphate pathway, the repression would lead toward β-D-glucose, β-D-glucose-6-phosphate and β-D-fructose-6-phosphate direction and therefore to α-D-glucose-6-phosphate and α-D-glucose-1-phosphate accumulation (Figure 2A). Noteworthy, α-D-glucose-6-phosphate and α-D-glucose-1-phosphate are the precursors of UDP glucose, UDP-galactose, dTDP-rhamnose and GDP-mannose, which are the building blocks in EPS biosynthesis (Quatrini et al., 2007). Another interesting results was the repression of three genes predicted to be involved in trehalose synthesis [treT (AFE_2083), treZ (AFE_2082) and treY (AFE_2081)] by tetrazole 9c. In the first case, α-D-glucose-1-phosphate consumption will be prevented, in agreement with the data presented above, and, in addition, maltodextrin synthesis will be favored. In the second case, maltodextrin consumption will be avoided (Figure 2B). Notably, maltodextrin has been shown to increase E. coli adhesion (Nickerson and McDonald, 2012). Along the same lines, genes involved in maltodextrin consumption [AFE_1799 and glgB (AFE_2836)] were repressed in the presence of tetrazole 9c (Figure 2B). Therefore, in planktonic cells, it appears that tetrazole 9c directed the carbon flow toward adhesion (maltodextrin), EPS precursor biosynthesis (α-D-glucose-6-phosphate, α-D-glucose-1-phosphate) and therefore biofilm formation.
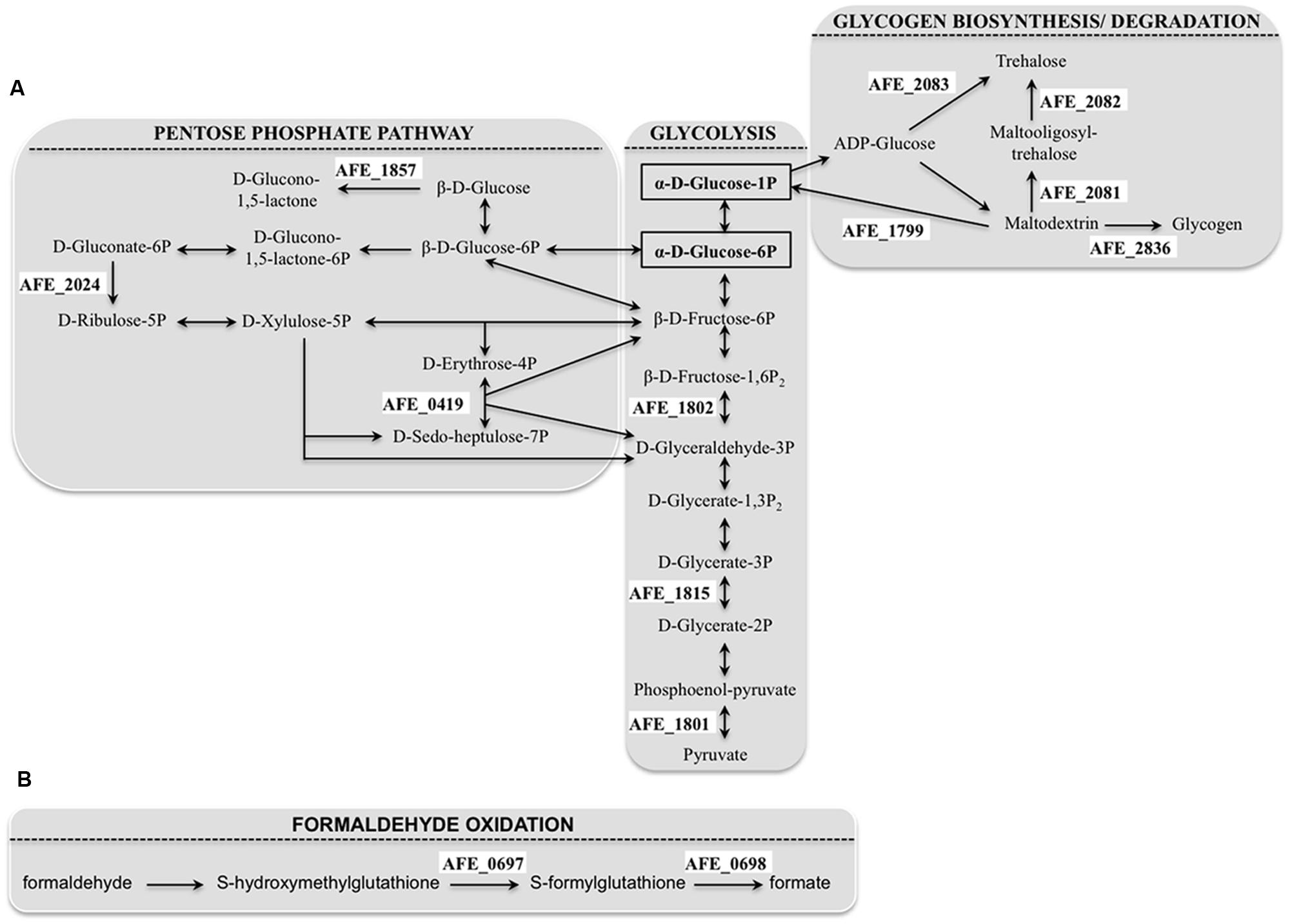
FIGURE 2. Acidithiobacillus ferrooxidans pathways showing gene repression by tetrazole 9c. (A) Genes involved in carbon flow that were downregulated in planktonic cells (for clarity, only the connections of the pathways discussed in the text have been shown). (B) Genes involved in formaldehyde oxidation that were downregulated in sessile cells. The precursors of the EPS (extracellular polymeric substances) building blocks are surrounded by a black rectangle. The genes that were repressed by tetrazole 9c are indicated in a white background.
In a number of microorganisms, including L. ferrooxidans, heat shock chaperones (Moreno-Paz et al., 2010; Singh et al., 2012; Becherelli et al., 2013; Grudniak et al., 2013) and proteases (Doern et al., 2009; Moreno-Paz et al., 2010; Singh et al., 2012; Yepes et al., 2012), in particular O-sialoglycoprotein endopeptidase (Wickstrom et al., 2013), have been shown to be required in sessile cells for biofilm development. Furthermore, the uspA gene, encoding an universal stress protein, is necessary for optimal biofilm formation in Porphyromonas gingivalis (Chen et al., 2006). In A. ferrooxidans, tetrazole 9c repressed the genes encoding the heat shock response RNA polymerase σ32 factor [rpoH (AFE2750)], Hsp20 family heat shock proteins (AFE_0871 and 2086), a putative universal stress protein (AFE_0751), as well as protease [lon (AFE_0872)] and O-sialoglycoprotein endopeptidase [gcp (AFE_0123)] in planktonic cells, indicating that these proteins are not required at the early step of biofilm biogenesis. Interestingly, bioD (AFE_1675) was repressed in the presence of the tetrazolic analog. This gene encodes dethiobiotin synthetase involved in biotin synthesis from 7-keto-8-aminopelargonate. This pathway consumes S-adenosyl-L-methionine (Streit and Entcheva, 2003). The down-regulation in the presence of tetrazole 9c of this gene could save this substrate that is required for AHL biosynthesis. Another important data is the repression of proB (AFE_2464) in the presence of tetrazole 9c. The proB gene encodes glutamate-5-kinase and its repression could lead to glutamate accumulation. Glutamate metabolism has been reported to be essential for biofilm formation. Amino acid levels in general increased in biofilm cells and are used as precursors for energy production with gluconeogenesis (Yeom et al., 2013). In harsh environments, such as acidic conditions, a high demand for amino acids as substrates for energy production may indeed exist in biofilms. Very recently, it has been proposed that amino acids, including glutamate, may also have another role as a signal for biofilm maturation and eventual disassembly (Wong et al., 2015). Finally, two genes encoding transcriptional regulators (AFE_2209 and AFE_2641) were repressed in planktonic cells in the presence of tetrazolic AHL analog. Therefore, we cannot exclude the possibility that genes differentially expressed in the presence of this superagonist AHL analog were indirectly regulated by one of these regulators rather than directly by the AfeR/AfeI QS system. It is noteworthy that members of the TetR-protein family, as is the case for AFE_2209, have been directly involved in the regulation of cellular processes and in particular the QS in different Gram-negative species (Cuthbertson and Nodwell, 2013; Longo et al., 2013).
To summarize, in planktonic cells, tetrazole 9c led to the induction of genes encoding (i) proton channel proteins to allow PMF energized transport system of AHL and substrates required for EPS synthesis, (ii) an enzyme required in an early step of polysaccharide synthesis, and (iii) transport system to anticipate phosphate and ammonium gradients within the biofilm. On the other hand, it repressed genes involved in (i) biofilm maturation (heat-shock proteins and chaperone encoding genes), (ii) biotin synthesis to prevent the consumption of S-adenosyl-L-methionine required for AHL biosynthesis, (iii) glutamate conversion to proline to use it as an energy source and/or as a signal for biofilm maturation, and (iv) carbohydrate metabolism to redirect the carbon flow toward proteins necessary for adhesion and EPS precursor biosynthesis. It seems therefore reasonable to conclude that tetrazole 9c reprograms planktonic cells toward early biofilm formation.
In sessile cells, only four genes, encoding proteins with known or predicted functions, presented significant differences in expression. Not surprisingly, the gene with the highest fold difference was afeI (AFE_1999) encoding the AHL synthase, with at least an eight-fold expression increase in the presence of tetrazole 9c indicating that indeed the QS was triggered. The three other genes fdhD (AFE_0690), adhI (AFE_0697), and fghA (AFE_0698) encoding a putative formate dehydrogenase family accessory protein FdhD, a S-(hydroxymethyl) glutathione dehydrogenase, and a S-formylglutathione hydrolase, respectively, are involved in formaldehyde oxidation to formate (Figure 2B). Their repression could lead to the accumulation of formaldehyde, shown to lead to higher biofilm density in a biofilm reactor (Ong et al., 2006). Another not exclusive possibility is that this system is to prevent formate formation that could acidify A. ferroxidans cytoplasm and lead to cell death.
Surprisingly, only three genes differentially expressed in the presence of tetrazole 9c (Supplementary Tables S3 and S4) have the AfeR binding site inferred from bioinformatic prediction (Farah et al., 2005; Banderas and Guiliani, 2013): AFE_0582 and AFE_1998 encoding hypothetical proteins as well as afeI (AFE_1999). This could be due to an indirect regulation through a regulator whose expression is controlled by QS. However, the two genes encoding a transcription regulator whose expression was downregulated in the presence of tetrazole 9c (AFE_2209 and AFE_2641) do not exhibit this predicted AfeR binding site. On the other hand, three genes [zwf (AFE_2025), AFE_0233, and AFE_1339] in which this site was predicted, are constitutively expressed in the conditions analyzed. Therefore, another possibility is that a different transcriptional regulator than AfeR binds to the proposed AfeR binding site. All in all, the QS regulon of A. ferrooxidans seems to involve a complex regulatory cascade.
To check that the afeI induction in the presence of tetrazole 9c observed by transcriptomic data was mediated by the QS regulator AfeR, we have produced AfeR in E. coli and analyzed its binding to the regulatory region of the afeI gene. AfeR with a hexa-histidine tag fused to its C terminus (AfeR-Histag) was mainly found in the inclusion bodies, even when the 3-hydroxy-C14-AHL (Gonzalez et al., 2013) was added at the induction time. The recombinant AfeR-Histag produced in the presence of 3-hydroxy-C14-AHL was purified on an affinity cobalt column. As shown in Figure 3A, a major band of the expected mass (theoretical molecular mass: 27,876 Da including one molecule of 3-hydroxy-C14-AHL) was visualized on Coomassie blue-stained SDS-polyacrylamide gels. This same protein was recognized by anti-hexahistidine tag antibodies (Figure 3A) strongly suggesting that it was AfeR-Histag. The analysis by MALDI-TOF mass spectrometry of this protein digested with Trypsin after reduction by DTT and alkylation by iodoacetamide confirmed that it was AfeR-Histag (54% sequence coverage).
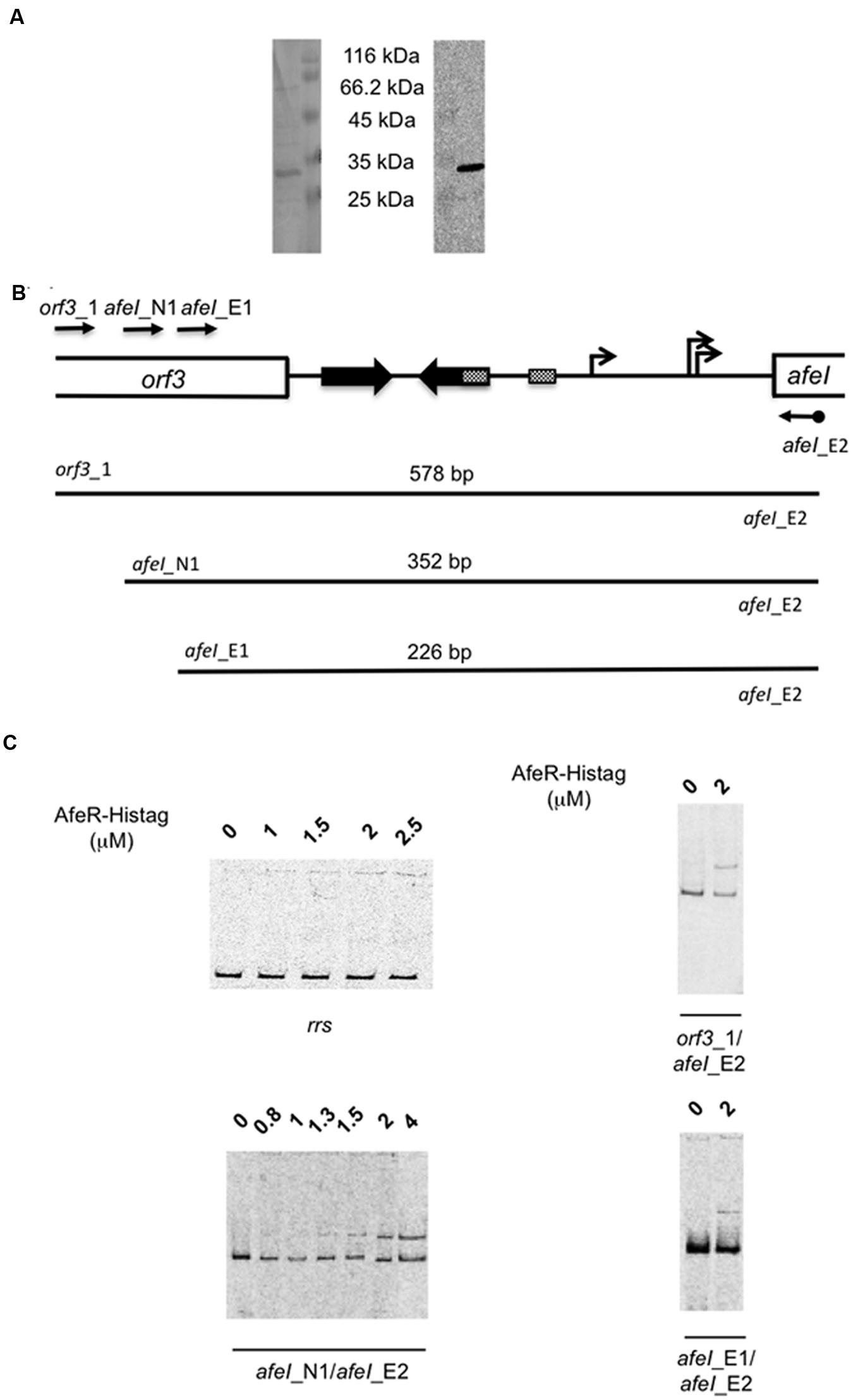
FIGURE 3. Production of the recombinant AfeR-Histag in Escherichia coli and its binding on the afeI regulatory region. (A) Coomassie brilliant blue stained SDS-PAGE (left) and Western immunoblot with antisera raised against the hexahistidine tag (right). The size of the unstained protein molecular weight marker standards (Euromedex) is indicated. (B) Schematic representation of the afeI locus with the DNA fragments analysed. (C) Gel mobility shift assays with an internal DNA fragment of the rrs gene of Thiomonas arsenitoxydans (left upper part) and the DNA fragments depicted in (B).
Binding of AfeR-Histag to the regulatory region of afeI was analyzed by EMSA in the presence of 3-hydroxy-C14-AHL. A retarded band was detected with 1.3 μM AfeR-Histag and higher concentrations (Figure 3C) with DNA fragments encompassing the palindromic sequence predicted to be the AfeR binding site (Farah et al., 2005; Banderas and Guiliani, 2013) in the afeI regulatory region (Figure 3B). This binding was specific to this region since no binding was observed on an internal fragment of the rrs gene of Thiomonas arsenitoxydans (Figure 3C). These results indicate that AfeR-Histag binds to the regulatory region of afeI in the presence of 3-hydroxy-C14-AHL, in agreement with the induction of this gene in the presence of tetrazole superagonist AHL analog 9c. Since AfeR was constitutively expressed under the conditions analyzed (i.e., with or without tetrazole 9c), these results suggest that the binding of 3-hydroxy-C14-AHL to AfeR induces a conformational change allowing its specific binding to the target DNA, as it has been proposed for several members of LuxR-like protein family (Choi and Greenberg, 1991).
The exogenous use of tetrazole superagonist AHL analog 9c allowed the first overview of the QS regulon of A. ferrooxidans, an acidophilic bacterial species involved in bioleaching processes. This study gave some insights into the molecular chain reactions involved in the first steps of mineral adherence and colonization of this bacterium. As expected, tetrazole 9c activates the positive feedback previously reported (Rivas et al., 2005) by inducing the transcription of afeI gene, likely through its binding to the transcriptional regulator AfeR, and therefore its activation, as early as the third day of growth.
The data obtained from planktonic cells revealed that tetrazole 9c triggers the QS system to drive gene expression toward sessile state by reprogramming some cellular processes. These mainly include: (i) induction of the genes encoding the F0-ATPase subunit leading to the PMF allowing AHL efflux and influx, (ii) repression of several genes involved in carbohydrate metabolism to orientate carbon flow to maltodextrin and EPS building block precursor synthesis for adhesion and biofilm formation, respectively; (iii) induction of phosphate and ammonium transporters to anticipate inorganic ion gradient within and around the biofilm structure. Whereas QS and c-di-GMP pathway have been linked in different bacterial species (Waters et al., 2008; Zhang, 2010; Kozlova et al., 2011; Suppiger et al., 2016), it is noteworthy that no change in the transcriptional profiling of the seven genes related to the c-di-GMP pathway in A. ferrooxidans (Ruiz et al., 2012; Castro et al., 2015) has been observed in the presence of tetrazole 9c. This result indicates that QS does not modulate c-di-GMP signaling in this Gram-negative species. Finally, the high transcription level of afeI gene in sessile cells observed after 3 days of growth lead not only to A. ferrooxidans biofilm stabilization but also to the synthesis of a large spectrum of AHL molecules (Farah et al., 2005; Valenzuela et al., 2017), some of which are sensed by secondary colonizers such as A. thiooxidans to form a mixed biofilm (Bellenberg et al., 2014) through a not yet identified non-canonical AHL-binding protein.
VB and NG conceived and designed the experiments. SM, DM, YD, and ET performed the experiments. VB, SM, NG, and DM analyzed the data. LS and YQ performed the chemical synthesis. NG, VB, YD, LS, YQ, and ET contributed to the reagents/materials/analysis tools. VB, NG, and ET wrote the paper. All authors read and approved the final manuscript.
SM acknowledges CONICYT to support her doctoral studies in Chile (scholarship N°21090736, 2009) and France (cotutorship “Becas Chile” N°78110005, 2011) and to allow attending international meeting. ET and VB are supported by Aix Marseille Université (AMU) and Centre National de la Recherche Scientifique (CNRS). NG is supported by Universidad de Chile (UCH). This work was partly performed in the frame of the PICS 5270 entitled “Studies of the Quorum Sensing and its function during the bioleaching process in the bacterium Acidithiobacillus ferrooxidans. An interdisciplinary challenge at the chemical/Biology/Biotechnology frontier.” This work was mainly supported by FONDECYT grants 1120295 and 1160702.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank M. Bauzan (Fermentation plant unit, IMM, Marseille, France) for growing the Escherichia coli Rosetta (DE3)/pLysS strain carrying pET21-AfeR-His tag in bioreactor and the Proteomic facility of IMM (Marseille, France) for proteomic analysis.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01365
FIGURE S1 |Chemical structure of tetrazole 9c.
Aiba, H., Adhya, S., and de Crombrugghe, B. (1981). Evidence for two functional gal promoters in intact Escherichia coli cells. J. Biol. Chem. 256, 11905–11910.
Amaro, A. M., Chamorro, D., Seeger, M., Arredondo, R., Peirano, I., and Jerez, C. A. (1991). Effect of external pH perturbations on in vivo protein synthesis by the acidophilic bacterium Thiobacillus ferrooxidans. J. Bacteriol. 173, 910–915.
Amouric, A., Brochier-Armanet, C., Johnson, D. B., Bonnefoy, V., and Hallberg, K. B. (2011). Phylogenetic and genetic variation among Fe(II)-oxidizing acidithiobacilli supports the view that these comprise multiple species with different ferrous iron oxidation pathways. Microbiology 157, 111–122. doi: 10.1099/mic.0.044537-0
Ardin, A. C., Fujita, K., Nagayama, K., Takashima, Y., Nomura, R., Nakano, K., et al. (2014). Identification and functional analysis of an ammonium transporter in Streptococcus mutans. PLoS ONE 9:e107569. doi: 10.1371/journal.pone.0107569
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., et al. (1998). Current Protocols in Molecular Biology. New York, NY: Greene publishing.
Banderas, A., and Guiliani, N. (2013). Bioinformatic prediction of gene functions regulated by quorum sensing in the bioleaching bacterium Acidithiobacillus ferrooxidans. Int. J. Mol. Sci. 14, 16901–16916. doi: 10.3390/ijms140816901
Barreto, M., Jedlicki, E., and Holmes, D. S. (2005). Identification of a gene cluster for the formation of extracellular polysaccharide precursors in the chemolithoautotroph Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 71, 2902–2909. doi: 10.1128/AEM.71.6.2902-2909.2005
Becherelli, M., Tao, J., and Ryder, N. S. (2013). Involvement of heat shock proteins in Candida albicans biofilm formation. J. Mol. Microbiol. Biotechnol. 23, 396–400. doi: 10.1159/000351619
Bellenberg, S., Diaz, M., Noel, N., Sand, W., Poetsch, A., Guiliani, N., et al. (2014). Biofilm formation, communication and interactions of leaching bacteria during colonization of pyrite and sulfur surfaces. Res. Microbiol. 165, 773–781. doi: 10.1016/j.resmic.2014.08.006
Bellenberg, S., Leon-Morales, C. F., Sand, W., and Vera, M. (2012). Visualization of capsular polysaccharide induction in Acidithiobacillus ferrooxidans. Hydrometallurgy 129–130, 82–89. doi: 10.1016/j.hydromet.2012.09.002
Biot, C., Bauer, H., Schirmer, R. H., and Davioud-Charvet, E. (2004). 5-substituted tetrazoles as bioisosteres of carboxylic acids. Bioisosterism and mechanistic studies on glutathione reductase inhibitors as antimalarials. J. Med. Chem. 47, 5972–5983. doi: 10.1021/jm0497545
Caffrey, S. M., Park, H. S., Been, J., Gordon, P., Sensen, C. W., and Voordouw, G. (2008). Gene expression by the sulfate-reducing bacterium Desulfovibrio vulgaris Hildenborough grown on an iron electrode under cathodic protection conditions. Appl. Environ. Microbiol. 74, 2404–2413. doi: 10.1128/AEM.02469-07
Cantero, L., Palacios, J. M., Ruiz-Argueso, T., and Imperial, J. (2006). Proteomic analysis of quorum sensing in Rhizobium leguminosarum biovar viciae UPM791. Proteomics 6(Suppl. 1), S97–S106. doi: 10.1002/pmic.200500312
Castro, M., Deane, S. M., Ruiz, L., Rawlings, D. E., and Guiliani, N. (2015). Diguanylate cyclase null mutant reveals that c-di-GMP pathway regulates the motility and adherence of the extremophile bacterium Acidithiobacillus caldus. PLoS ONE 10:e0116399. doi: 10.1371/journal.pone.0116399
Chen, L. X., Hu, M., Huang, L. N., Hua, Z. S., Kuang, J. L., Li, S. J., et al. (2015). Comparative metagenomic and metatranscriptomic analyses of microbial communities in acid mine drainage. ISME J. 9, 1579–1592. doi: 10.1038/ismej.2014.245
Chen, W., Honma, K., Sharma, A., and Kuramitsu, H. K. (2006). A universal stress protein of Porphyromonas gingivalis is involved in stress responses and biofilm formation. FEMS Microbiol. Lett. 264, 15–21. doi: 10.1111/j.1574-6968.2006.00426.x
Chhabra, S. R., Harty, C., Hooi, D. S., Daykin, M., Williams, P., Telford, G., et al. (2003). Synthetic analogues of the bacterial signal (quorum sensing) molecule N-(3-oxododecanoyl)-L-homoserine lactone as immune modulators. J. Med. Chem. 46, 97–104. doi: 10.1021/jm020909n
Choi, S. H., and Greenberg, E. P. (1991). The C-terminal region of the Vibrio fischeri LuxR protein contains an inducer-independent lux gene activating domain. Proc. Natl. Acad. Sci. U.S.A. 88, 11115–11119. doi: 10.1073/pnas.88.24.11115
Chung, C. T., and Miller, R. H. (1988). A rapid and convenient method for the preparation and storage of competent bacterial cells. Nucleic Acids Res. 16:3580. doi: 10.1093/nar/16.8.3580
Clark, M. E., He, Z., Redding, A. M., Joachimiak, M. P., Keasling, J. D., Zhou, J. Z., et al. (2012). Transcriptomic and proteomic analyses of Desulfovibrio vulgaris biofilms: carbon and energy flow contribute to the distinct biofilm growth state. BMC Genomics 13:138. doi: 10.1186/1471-2164-13-138
Cuthbertson, L., and Nodwell, J. R. (2013). The TetR family of regulators. Microbiol. Mol. Biol. Rev. 77, 440–475. doi: 10.1128/MMBR.00018-13
Decho, A. W., Frey, R. L., and Ferry, J. L. (2011). Chemical challenges to bacterial AHL signaling in the environment. Chem. Rev. 111, 86–99. doi: 10.1021/cr100311q
Diaz, M., Copaja, S., and Guiliani, N. (2013). Functional analysis of c-di-GMP pathway in biomining bacteria Acidithiobacillus thiooxidans. Adv. Mater. Res. 825, 133–136. doi: 10.4028/www.scientific.net/AMR.825.133
Doern, C. D., Roberts, A. L., Hong, W., Nelson, J., Lukomski, S., Swords, W. E., et al. (2009). Biofilm formation by group A Streptococcus: a role for the streptococcal regulator of virulence (Srv) and streptococcal cysteine protease (SpeB). Microbiology 155, 46–52. doi: 10.1099/mic.0.021048-0
Falagan, C., and Johnson, D. B. (2015). Acidithiobacillus ferriphilus sp. nov., a facultatively anaerobic iron- and sulfur-metabolising extreme acidophile. Int. J. Syst. Evol. Microbiol 66, 206–211. doi: 10.1099/ijsem.0.000698
Farah, C., Vera, M., Morin, D., Haras, D., Jerez, C. A., and Guiliani, N. (2005). Evidence for a functional quorum-sensing type AI-1 system in the extremophilic bacterium Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 71, 7033–7040. doi: 10.1128/AEM.71.11.7033-7040.2005
Gonzalez, A., Bellenberg, S., Mamani, S., Ruiz, L., Echeverria, A., Soulere, L., et al. (2013). AHL signaling molecules with a large acyl chain enhance biofilm formation on sulfur and metal sulfides by the bioleaching bacterium Acidithiobacillus ferrooxidans. Appl. Microbiol. Biotechnol. 97, 3729–3737. doi: 10.1007/s00253-012-4229-3
Grudniak, A. M., Pawlak, K., Bartosik, K., and Wolska, K. I. (2013). Physiological consequences of mutations in the htpG heat shock gene of Escherichia coli. Mutat. Res. 745–746, 1–5. doi: 10.1016/j.mrfmmm.2013.04.003
Harneit, K., Göksel, A., Klock, J. -H., Gehrke, T., and Sand, W. (2006). Adhesion to metal sulfide surfaces by cells of Acidithiobacillus ferrooxidans, Acidithiobacillus thiooxidans and Leptospirillum ferrooxidans. Hydrometallurgy 83, 245–254. doi: 10.1016/j.hydromet.2006.03.044
Hedrich, S., and Johnson, D. B. (2013). Acidithiobacillus ferridurans sp. nov., an acidophilic iron-, sulfur- and hydrogen-metabolizing chemolithotrophic gammaproteobacterium. Int. J. Syst. Evol. Microbiol. 63, 4018–4025. doi: 10.1099/ijs.0.049759-0
Heindl, J. E., Wang, Y., Heckel, B. C., Mohari, B., Feirer, N., and Fuqua, C. (2014). Mechanisms and regulation of surface interactions and biofilm formation in Agrobacterium. Front. Plant Sci. 5:176. doi: 10.3389/fpls.2014.00176
Hengge, R. (2009). Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol. 7, 263–273. doi: 10.1038/nrmicro2109
Hengge, R., Grundling, A., Jenal, U., Ryan, R., and Yildiz, F. (2015). Bacterial signal transduction by c-di-GMP and other nucleotide second messengers. 198:15–26 J. Bacteriol. doi: 10.1128/JB.00331-15
Ikonomidis, A., Tsakris, A., Kanellopoulou, M., Maniatis, A. N., and Pournaras, S. (2008). Effect of the proton motive force inhibitor carbonyl cyanide-m-chlorophenylhydrazone (CCCP) on Pseudomonas aeruginosa biofilm development. Lett. Appl. Microbiol. 47, 298–302. doi: 10.1111/j.1472-765X.2008.02430.x
Jerez, C. A. (2009). “Biomining microorganisms: molecular aspects and applications in biotechnology and bioremediation,” in Advances in Applied Bioremediation, eds A. Singh, R. C. Kuhad, and O. P. Ward (Berlin: Springer-Verlag), 239–256.
Johnson, D. B. (2009). “Extremophiles: acidic environments,” in Encyclopaedia of Microbiology ed. M. Schaechter (Oxford: Elsevier), 107–126.
Johnson, D. B. (2012). Geomicrobiology of extremely acidic subsurface environments. FEMS Microbiol. Ecol. 81, 2–12. doi: 10.1111/j.1574-6941.2011.01293.x
Johnson, D. B. (2014). Biomining-biotechnologies for extracting and recovering metals from ores and waste materials. Curr. Opin. Biotechnol. 30, 24–31. doi: 10.1016/j.copbio.2014.04.008
Kalia, D., Merey, G., Nakayama, S., Zheng, Y., Zhou, J., Luo, Y., et al. (2013). Nucleotide, c-di-GMP, c-di-AMP, cGMP, cAMP, (p)ppGpp signaling in bacteria and implications in pathogenesis. Chem. Soc. Rev. 42, 305–341. doi: 10.1039/c2cs35206k
Kapatral, V., Bina, X., and Chakrabarty, A. M. (2000). Succinyl coenzyme A synthetase of Pseudomonas aeruginosa with a broad specificity for nucleoside triphosphate (NTP) synthesis modulates specificity for NTP synthesis by the 12-kilodalton form of nucleoside diphosphate kinase. J. Bacteriol. 182, 1333–1339. doi: 10.1128/JB.182.5.1333-1339.2000
Kassem, I. I., Khatri, M., Esseili, M. A., Sanad, Y. M., Saif, Y. M., Olson, J. W., and Rajashekara, G. (2012). Respiratory proteins contribute differentially to Campylobacter jejuni’s survival and in vitro interaction with hosts’ intestinal cells. BMC Microbiol. 12:258. doi: 10.1186/1471-2180-12-258
Kozlova, E. V., Khajanchi, B. K., Sha, J., and Chopra, A. K. (2011). Quorum sensing and c-di-GMP-dependent alterations in gene transcripts and virulence-associated phenotypes in a clinical isolate of Aeromonas hydrophila. Microb. Pathog. 50, 213–223. doi: 10.1016/j.micpath.2011.01.007
Liljeqvist, M., Ossandon, F. J., Gonzalez, C., Rajan, S., Stell, A., Valdes, J., et al. (2015). Metagenomic analysis reveals adaptations to a cold-adapted lifestyle in a low-temperature acid mine drainage stream. FEMS Microbiol. Ecol. 91:fiv011. doi: 10.1093/femsec/fiv011
Longo, F., Rampioni, G., Bondi, R., Imperi, F., Fimia, G. M., Visca, P., et al. (2013). A new transcriptional repressor of the Pseudomonas aeruginosa quorum sensing receptor gene LasR. PLoS ONE 8:e69554. doi: 10.1371/journal.pone.0069554
Majerczyk, C., Brittnacher, M., Jacobs, M., Armour, C. D., Radey, M., Schneider, E., et al. (2014). Global analysis of the Burkholderia thailandensis quorum sensing-controlled regulon. J. Bacteriol. 196, 1412–1424. doi: 10.1128/JB.01405-13
Menzel, P., Gudbergsdottir, S. R., Rike, A. G., Lin, L., Zhang, Q., Contursi, P., et al. (2015). Comparative metagenomics of eight geographically remote terrestrial hot springs. Microb. Ecol. 70, 411–424. doi: 10.1007/s00248-015-0576-9
Montgomery, K., Charlesworth, J. C., LeBard, R., Visscher, P. T., and Burns, B. P. (2013). Quorum sensing in extreme environments. Life (Basel) 3, 131–148. doi: 10.3390/life3010131
Moreno-Paz, M., Gomez, M. J., Arcas, A., and Parro, V. (2010). Environmental transcriptome analysis reveals physiological differences between biofilm and planktonic modes of life of the iron oxidizing bacteria Leptospirillum spp. in their natural microbial community. BMC Genomics 11:404. doi: 10.1186/1471-2164-11-404
Ng, W. L., and Bassler, B. L. (2009). Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 43, 197–222. doi: 10.1146/annurev-genet-102108-134304
Nickerson, K. P., and McDonald, C. (2012). Crohn’s disease-associated adherent-invasive Escherichia coli adhesion is enhanced by exposure to the ubiquitous dietary polysaccharide maltodextrin. PLoS ONE 7:e52132. doi: 10.1371/journal.pone.0052132
O’May, G. A., Jacobsen, S. M., Longwell, M., Stoodley, P., Mobley, H. L., and Shirtliff, M. E. (2009). The high-affinity phosphate transporter Pst in Proteus mirabilis HI4320 and its importance in biofilm formation. Microbiology 155, 1523–1535. doi: 10.1099/mic.0.026500-0
Ong, S. L., Sarkar, S. K., Lee, L. Y., Hu, J. Y., Ng, H. Y., and van Loosdrecht, M. (2006). Effect of formaldehyde on biofilm activity and morphology in an ultracompact biofilm reactor for carbonaceous wastewater treatment. Water Environ. Res. 78, 372–380. doi: 10.2175/106143006X98787
Parsek, M. R., and Greenberg, E. P. (2005). Sociomicrobiology: the connections between quorum sensing and biofilms. Trends Microbiol. 13, 27–33. doi: 10.1016/j.tim.2004.11.007
Peterson, J. D., Umayam, L. A., Dickinson, T., Hickey, E. K., and White, O. (2001). The comprehensive microbial resource. Nucleic Acids Res. 29, 123–125. doi: 10.1093/nar/29.1.123
Pogliani, C., and Donati, E. (1999). The role of exopolymers in the bioleaching of a non-ferrous metal sulphide. J. Ind. Microbiol. Biotechnol. 22, 88–92. doi: 10.1038/sj.jim.2900610
Qin, N., Callahan, S. M., Dunlap, P. V., and Stevens, A. M. (2007). Analysis of LuxR regulon gene expression during quorum sensing in Vibrio fischeri. J. Bacteriol. 189, 4127–4134. doi: 10.1128/JB.01779-06
Quatrini, R., Appia-Ayme, C., Denis, Y., Jedlicki, E., Holmes, D. S., and Bonnefoy, V. (2009). Extending the models for iron and sulfur oxidation in the extreme acidophile Acidithiobacillus ferrooxidans. BMC Genomics 10:394. doi: 10.1186/1471-2164-10-394
Quatrini, R., Appia-Ayme, C., Denis, Y., Ratouchniak, J., Veloso, F., Valdes, J., et al. (2006). Insights into the iron and sulfur energetic metabolism of Acidithiobacillus ferrooxidans by microarray transcriptome profiling. Hydrometallurgy 83, 263–272. doi: 10.1007/s12223-011-0067-4
Quatrini, R., Valdes, J., Jedlicki, E., and Holmes, D. S. (2007). “The use of bioinformatics and genomic biology to advance our understanding of bioleaching microorganisms,” in Microbial Processing of Metal Sulfides, eds E. R. Donati and W. Sand (Dordrecht: Springer), 221–239.
Rivas, M., Seeger, M., Holmes, D. S., and Jedlicki, E. (2005). A Lux-like quorum sensing system in the extreme acidophile Acidithiobacillus ferrooxidans. Biol. Res. 38, 283–297. doi: 10.4067/S0716-97602005000200018
Rohwerder, T., and Sand, W. (2007). “Mechanisms and biochemical fundamentals of bacterial metal sulfide oxidation,” in Microbial Processing of Metal Sulfides, eds E. R. Donati and W. Sand (Dordrecht: Springer), 35–58.
Romling, U., Galperin, M. Y., and Gomelsky, M. (2013). Cyclic di-GMP: the first 25 years of a universal bacterial second messenger. Microbiol. Mol. Biol. Rev. 77, 1–52. doi: 10.1128/MMBR.00043-12
Ruiz, L. M., Castro, M., Barriga, A., Jerez, C. A., and Guiliani, N. (2012). The extremophile Acidithiobacillus ferrooxidans possesses a c-di-GMP signalling pathway that could play a significant role during bioleaching of minerals. Lett. Appl. Microbiol. 54, 133–139. doi: 10.1111/j.1472-765X.2011.03180.x
Ruiz, L. M., Valenzuela, S., Castro, M., Gonzalez, A., Frezza, M., Soulere, L., et al. (2008). AHL communication is a widespread phenomenon in biomining bacteria and seems to be involved in mineral-adhesion efficiency. Hydrometallurgy 94, 133–137. doi: 10.1016/j.hydromet.2008.05.028
Ryan, R. P., Fouhy, Y., Lucey, J. F., Crossman, L. C., Spiro, S., He, Y. W., et al. (2006). Cell-cell signaling in Xanthomonas campestris involves an HD-GYP domain protein that functions in cyclic di-GMP turnover. Proc. Natl. Acad. Sci. U.S.A. 103, 6712–6717. doi: 10.1073/pnas.0600345103
Sabbah, M., Fontaine, F., Grand, L., Boukraa, M., Efrit, M. L., Doutheau, A., et al. (2012). Synthesis and biological evaluation of new N-acyl-homoserine-lactone analogues, based on triazole and tetrazole scaffolds, acting as LuxR-dependent quorum sensing modulators. Bioorg. Med. Chem. 20, 4727–4736. doi: 10.1016/j.bmc.2012.06.007
Saville, R. M., Rakshe, S., Haagensen, J. A., Shukla, S., and Spormann, A. M. (2011). Energy-dependent stability of Shewanella oneidensis MR-1 biofilms. J. Bacteriol. 193, 3257–3264. doi: 10.1128/JB.00251-11
Singh, V. K., Syring, M., Singh, A., Singhal, K., Dalecki, A., and Johansson, T. (2012). An insight into the significance of the DnaK heat shock system in Staphylococcus aureus. Int. J. Med. Microbiol. 302, 242–252. doi: 10.1016/j.ijmm.2012.05.001
Slyemi, D., Moinier, D., Talla, E., and Bonnefoy, V. (2013). Organization and regulation of the arsenite oxidase operon of the moderately acidophilic and facultative chemoautotrophic Thiomonas arsenitoxydans. Extremophiles 17, 911–920. doi: 10.1007/s00792-013-0573-1
Soulere, L., Guiliani, N., Queneau, Y., Jerez, C. A., and Doutheau, A. (2008). Molecular insights into quorum sensing in Acidithiobacillus ferrooxidans bacteria via molecular modelling of the transcriptional regulator AfeR and of the binding mode of long-chain acyl homoserine lactones. J. Mol. Model 14, 599–606. doi: 10.1007/s00894-008-0315-y
Stevens, A. M., Queneau, Y., Soulere, L., von Bodman, S., and Doutheau, A. (2011). Mechanisms and synthetic modulators of AHL-dependent gene regulation. Chem. Rev. 111, 4–27. doi: 10.1021/cr100064s
Streit, W. R., and Entcheva, P. (2003). Biotin in microbes, the genes involved in its biosynthesis, its biochemical role and perspectives for biotechnological production. Appl. Microbiol. Biotechnol. 61, 21–31. doi: 10.1007/s00253-002-1186-2
Suppiger, A., Eshwar, A. K., Stephan, R., Kaever, V., Eberl, L., and Lehner, A. (2016). The DSF type quorum sensing signalling system RpfF/R regulates diverse phenotypes in the opportunistic pathogen Cronobacter. Sci. Rep. 6:18753. doi: 10.1038/srep18753
Talla, E., Tekaia, F., Brino, L., and Dujon, B. (2003). A novel design of whole-genome microarray probes for Saccharomyces cerevisiae which minimizes cross-hybridization. BMC Genomics 4:38. doi: 10.1186/1471-2164-4-38
Ueda, A., and Wood, T. K. (2009). Connecting quorum sensing, c-di-GMP, pel polysaccharide, and biofilm formation in Pseudomonas aeruginosa through tyrosine phosphatase TpbA (PA3885). PLoS Pathog. 5:e1000483. doi: 10.1371/journal.ppat.1000483
Valdés, J., Pedroso, I., Quatrini, R., and Holmes, D. S. (2008). Comparative genome analysis of Acidithiobacillus ferrooxidans, A. thiooxidans and A. caldus: insights into their metabolism and ecophysiology. Hydrometallurgy 94, 180–184. doi: 10.1016/j.hydromet.2008.05.039
Valenzuela, S., Banderas, A., Jerez, C. A., and Guiliani, N. (2007). “Cell-cell communication in bacteria,” in Microbial Processing of Metal Sulfides, eds E. R. Donati and W. Sand (Dordrecht: Springer), 253–264.
Valenzuela, S., Banderas, A., Jerez, C. A., and Guiliani, N. (2017). “Cell-cell communication in bacteria: a promising new approach to improve bioleaching efficiency?,” in Microbial Processing of Metal Sulfides, eds E. R. Donati and W. Sand (Dordrecht: Springer), 253–264.
Vallenet, D., Belda, E., Calteau, A., Cruveiller, S., Engelen, S., Lajus, A., et al. (2013). MicroScope–an integrated microbial resource for the curation and comparative analysis of genomic and metabolic data. Nucleic Acids Res. 41, D636–D647. doi: 10.1093/nar/gks1194
Varga, Z. G., Armada, A., Cerca, P., Amaral, L., Mior Ahmad Subki, M. A., Savka, M. A., et al. (2012). Inhibition of quorum sensing and efflux pump system by trifluoromethyl ketone proton pump inhibitors. In Vivo 26, 277–285.
Vasil, M. L. (2003). DNA microarrays in analysis of quorum sensing: strengths and limitations. J. Bacteriol. 185, 2061–2065. doi: 10.1128/JB.185.7.2061-2065.2003
Venturi, V., and Subramoni, S. (2009). Future research trends in the major chemical language of bacteria. HFSP J. 3, 105–116. doi: 10.2976/1.3065673
Vera, M., Krok, B., Bellenberg, S., Sand, W., and Poetsch, A. (2013). Shotgun proteomics study of early biofilm formation process of Acidithiobacillus ferrooxidans ATCC 23270 on pyrite. Proteomics 13, 1133–1144. doi: 10.1002/pmic.201200386
Wagner, V. E., Bushnell, D., Passador, L., Brooks, A. I., and Iglewski, B. H. (2003). Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J. Bacteriol. 185, 2080–2095. doi: 10.1128/JB.185.7.2080-2095.2003
Waters, C. M., and Bassler, B. L. (2005). Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 21, 319–346. doi: 10.1146/annurev.cellbio.21.012704.131001
Waters, C. M., Lu, W., Rabinowitz, J. D., and Bassler, B. L. (2008). Quorum sensing controls biofilm formation in Vibrio cholerae through modulation of cyclic di-GMP levels and repression of vpsT. J. Bacteriol. 190, 2527–2536. doi: 10.1128/JB.01756-07
Wickstrom, C., Chavez de Paz, L., Davies, J. R., and Svensater, G. (2013). Surface-associated MUC5B mucins promote protease activity in Lactobacillus fermentum biofilms. BMC Oral Health 13:43. doi: 10.1186/1472-6831-13-43
Williams, K. P., and Kelly, D. P. (2013). Proposal for a new class within the phylum Proteobacteria, Acidithiobacillia classis nov., with the type order Acidithiobacillales, and emended description of the class Gammaproteobacteria. Int. J. Syst. Evol. Microbiol. 63, 2901–2906. doi: 10.1099/ijs.0.049270-0
Wong, H. S., Maker, G. L., Trengove, R. D., and O’Handley, R. M. (2015). Gas chromatography-mass spectrometry-based metabolite profiling of Salmonella enterica serovar typhimurium differentiates between biofilm and planktonic phenotypes. Appl. Environ. Microbiol. 81, 2660–2666. doi: 10.1128/AEM.03658-14
Yarzabal, A., Duquesne, K., and Bonnefoy, V. (2003). Rusticyanin gene expression of Acidithiobacillus ferrooxidans ATCC 33020 in sulfur- and in ferrous iron-media. Hydrometallurgy 71, 107–114. doi: 10.1016/S0304-386X(03)00146-4
Yeom, J., Shin, J. H., Yang, J. Y., Kim, J., and Hwang, G. S. (2013). (1)H NMR-based metabolite profiling of planktonic and biofilm cells in Acinetobacter baumannii 1656-2. PLoS ONE 8:e57730. doi: 10.1371/journal.pone.0057730
Yepes, A., Schneider, J., Mielich, B., Koch, G., Garcia-Betancur, J. C., Ramamurthi, K. S., et al. (2012). The biofilm formation defect of a Bacillus subtilis flotillin-defective mutant involves the protease FtsH. Mol. Microbiol. 86, 457–471. doi: 10.1111/j.1365-2958.2012.08205.x
Keywords: quorum sensing regulon, acyl homoserine lactone, superagonist, extracellular polymeric substances, biofilm, transcriptomic, Acidithiobacillus ferrooxidans, acidophile
Citation: Mamani S, Moinier D, Denis Y, Soulère L, Queneau Y, Talla E, Bonnefoy V and Guiliani N (2016) Insights into the Quorum Sensing Regulon of the Acidophilic Acidithiobacillus ferrooxidans Revealed by Transcriptomic in the Presence of an Acyl Homoserine Lactone Superagonist Analog. Front. Microbiol. 7:1365. doi: 10.3389/fmicb.2016.01365
Received: 30 June 2016; Accepted: 17 August 2016;
Published: 14 September 2016.
Edited by:
Axel Schippers, Federal Institute for Geosciences and Natural Resources, GermanyReviewed by:
Jeannette Marrero-Coto, Leibniz University of Hanover, GermanyCopyright © 2016 Mamani, Moinier, Denis, Soulère, Queneau, Talla, Bonnefoy and Guiliani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Violaine Bonnefoy, Ym9ubmVmb3lAaW1tLmNucnMuZnI= Nicolas Guiliani, bmd1aWxpYW5AdWNoaWxlLmNs
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.