
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 30 August 2016
Sec. Extreme Microbiology
Volume 7 - 2016 | https://doi.org/10.3389/fmicb.2016.01301
This article is part of the Research Topic Metagenomic, genomic and single-cell approaches advancing the microbial physiology and ecology of extreme environments View all 20 articles
 Nan Mei1Anne Postec1Christophe Monnin2Bernard Pelletier3Claude E. Payri3Bénédicte Ménez4Eléonore Frouin1Bernard Ollivier1Gaël Erauso1Marianne Quéméneur1*
Nan Mei1Anne Postec1Christophe Monnin2Bernard Pelletier3Claude E. Payri3Bénédicte Ménez4Eléonore Frouin1Bernard Ollivier1Gaël Erauso1Marianne Quéméneur1*High amounts of hydrogen are emitted in the serpentinite-hosted hydrothermal field of the Prony Bay (PHF, New Caledonia), where high-pH (~11), low-temperature (< 40°C), and low-salinity fluids are discharged in both intertidal and shallow submarine environments. In this study, we investigated the diversity and distribution of potentially hydrogen-producing bacteria in Prony hyperalkaline springs by using metagenomic analyses and different PCR-amplified DNA sequencing methods. The retrieved sequences of hydA genes, encoding the catalytic subunit of [FeFe]-hydrogenases and, used as a molecular marker of hydrogen-producing bacteria, were mainly related to those of Firmicutes and clustered into two distinct groups depending on sampling locations. Intertidal samples were dominated by new hydA sequences related to uncultured Firmicutes retrieved from paddy soils, while submarine samples were dominated by diverse hydA sequences affiliated with anaerobic and/or thermophilic submarine Firmicutes pertaining to the orders Thermoanaerobacterales or Clostridiales. The novelty and diversity of these [FeFe]-hydrogenases may reflect the unique environmental conditions prevailing in the PHF (i.e., high-pH, low-salt, mesothermic fluids). In addition, novel alkaliphilic hydrogen-producing Firmicutes (Clostridiales and Bacillales) were successfully isolated from both intertidal and submarine PHF chimney samples. Both molecular and cultivation-based data demonstrated the ability of Firmicutes originating from serpentinite-hosted environments to produce hydrogen by fermentation, potentially contributing to the molecular hydrogen balance in situ.
Hydrogen (H2) can be naturally produced by both geochemical and biological processes in various environments. Geochemically, H2 can be generated during the serpentinization of ultramafic rocks by the reduction of water coupled to the oxidation of ferrous Fe contained in olivines and pyroxenes (Schrenk et al., 2013). This reaction is accompanied by the production of exceedingly alkaline waters (pH up to 12). Produced H2 constitutes a large reservoir of energy with the capacity to sustain the development of a wide range of chemolithoautotrophic microorganisms. As an illustration, in serpentinite-hosted ecosystems, H2 has been shown to be consumed by microorganisms such as anaerobic hydrogenotrophs (e.g., methanogens) and aerobic H2-oxidizing Betaproteobacteria (i.e., “Serpentinomonas” spp.; Brazelton et al., 2006; Tiago and Veríssimo, 2013; Quéméneur et al., 2014, 2015; Suzuki et al., 2014). In addition to H2 production resulting from abiotic reactions, H2 could be produced biologically by various types of microorganisms in these anoxic serpentinite-hosted environments. Among them, the Firmicutes phylum and especially the Clostridiales spp. are recognized as potential fermentative H2-producing bacteria (Xing et al., 2008; Quéméneur et al., 2011). However, the distribution and role of these microorganisms in the H2 budget of serpentinization-related systems have been scarcely addressed; so far, only two studies have investigated the potential of anaerobic microorganisms to biologically produce H2 in these hyperalkaline environments (Brazelton et al., 2012; Mei et al., 2014).
Biological H2 production is performed by phylogenetically and physiologically diverse groups of Bacteria and Archaea (i.e., anaerobes, facultative anaerobes, and photosynthetic bacteria). This process is carried out by hydrogenases that catalyze the reversible oxidation of H2 and are divided into two major phylogenetically distinct classes: [NiFe]-hydrogenases and [FeFe]-hydrogenases (Vignais and Colbeau, 2004; Lubitz et al., 2014; Peters et al., 2015). [NiFe]-hydrogenases are widely distributed among the Bacteria and Archaea domains, whereas [FeFe]-hydrogenases are primarily found in anaerobic bacteria of the orders Clostridiales, Thermotogales, and the family Desulfovibrionaceae (Vignais et al., 2001). While [NiFe]-hydrogenases are generally involved in H2 consumption, [FeFe]-hydrogenases are usually involved in H2 production in vivo (Vignais and Colbeau, 2004), with nonetheless some exceptions including electron-bifurcating [FeFe]-hydrogenases (Schut and Adams, 2009; Wang et al., 2013; Poudel et al., 2016). The hydA genes encoding the catalytic subunit of [FeFe]-hydrogenases have been used as a pertinent molecular marker to monitor compositional changes of bacterial H2-producers in fermentation bioreactors (Xing et al., 2008; Quéméneur et al., 2010, 2011). The genetic diversity of hydA genes and associated H2 production potential have also been studied by culture-independent approaches in various ecosystems including extreme environments, such as saline microbial mats of Guerrero Negro (Mexico) or geothermal springs of Yellowstone National Park (USA) discharging acidic, neutral or alkaline fluids (Boyd et al., 2009, 2010) to provide a more exhaustive picture of H2-producing community.
The hydrothermal field of the Prony Bay (PHF, New Caledonia, South Pacific) comprises several intertidal and shallow submarine hyperalkaline springs located at less than 50 m below sea level (mbsl). Similarly to the deep-sea Lost City hydrothermal field (LCHF) located at ~800 mbsl, off the Mid-Atlantic Ridge (30°N), PHF relies on an serpentinizing basement and discharges into the seawater high pH (~11) fluids enriched in hydrogen (H2: 19–24% vol in free gas) and methane (CH4: 6–13% vol in free gas; Kelley et al., 2005; Monnin et al., 2014). When alkaline fluids mix with seawater, precipitation in the form of calcium carbonates (CaCO3), and brucite [Mg(OH)2] occurs (Launay and Fontes, 1985), forming chimneys reaching up to tens of meters in height. Although PHF and LCHF display similar geochemical and mineralogical features, PHF is unique in that its hydrothermal features release low-temperature (< 40°C) and low-salinity fluids in a shallow submarine environment. Recently, molecular microbial surveys based on 16S rRNA genes analysis provided evidence of an abundant and highly diverse bacterial community inhabiting these hydrothermal chimneys with a peculiar emphasis for members of the Firmicutes (Quéméneur et al., 2014; Postec et al., 2015), as also evidenced in other serpentinization-related environments (Brazelton et al., 2013; Miller et al., 2016). Several anaerobic bacterial strains belonging to this phylum, and especially to the Clostridiales order, have been successfully isolated from the PHF submarine chimneys and described (Ben Aissa et al., 2014, 2015; Mei et al., 2014; Bes et al., 2015). To our knowledge, these bacteria are to date the unique anaerobic isolates reported from serpentinite-hosted environments. Those may impact the H2 budget.
The main goal of this study was to determine the diversity and distribution of the [FeFe]-hydrogenase encoding genes, considered as a molecular marker of H2-producing bacteria that may contribute to hydrogen production in different sites of the PHF located in intertidal or shallow submarine zones. [FeFe]-hydrogenases encoding genes related to those belonging to the order Clostridiales, phylum Firmicutes, were largely detected using both metagenomic analyses and PCR amplification and subsequent sequencing of hydA genes. Attempts to cultivate and isolate new alkaliphilic hydrogen-producing Clostridiales strains from these alkaline serpentinite-hosted environments were also successful.
The samples used for PCR-based molecular analyses as well as microbial cultures in this study were collected in October 2012 at three different sites in the Prony Bay: (i) “Rivière des Kaoris” (RK), located in the Eastern end of the Carenage Bay (22°17.969′S, 166°51.709′E), (ii) “Bain des Japonais” (BdJ), located in the Western branch of the Carenage Bay (2°17.970′S, 166°51.708′E), and (iii) “Aiguille de Prony” (also referred to as ST07), located in the North of the Bay (22°19.796′S, 166°50.058′E; Figure 1; Quéméneur et al., 2014). The samples used for metagenomic analyses were collected in November 2005 at the deepest submarine PHF site ST09 (22°21.653S, 166°52.777E; Figure 1; Postec et al., 2015). Site locations are indicated on the map displayed in Monnin et al. (2014).

Figure 1. Intertidal and submarine hydrothermal sources of Prony Hydrothermal Field (PHF) in New Caledonia. Photographs show the sampling sites: “Rivière des Kaoris” (RK), “Bain des Japonais” (BdJ), “Aiguille de Prony” (ST07), and the Prony ST09 site. Suffixes “C” and “F” in site names stand, respectively, for chimney and fluid.
The RK site is composed of large travertines (covered by seawater in high tide), close to an artificial pool (~3 m long, ~2 m wide, and ~1.5 m deep) fed by hyperalkaline fluids (pH 10.9) and it is located on the coast above sea level. The BdJ site is a carbonated platform located on the foreshore and thus uncovered at low tide. The tidal range in the Prony bay is about 1 m. In contrast, the ST07 site is a submarine edifice, 35 m high, composed of carbonated chimneys sampled by scuba divers at 16 mbsl (Figure 1). At the ST09 site, an active and partially coral encrusted submarine chimney with white top was collected at 43 mbsl (Postec et al., 2015).
In PHF sites, H2 was the second most dominant gas (followed by CH4) after N2 (Monnin et al., 2014; Deville and Prinzhofer, 2016). The range of the H2, CH4, and N2 gas contents collected at the intertidal PHF sites (RK and BdJ) were 19–24%V, 6–13%V, and 67–69%V, respectively (Monnin et al., 2014). The fluids discharged by PHF sites had high pH-values ranging from 10.1 (ST07) to 11.1 (BdJ). The temperature of fluids measured at sampling sites ranged from 30.8°C (RK) to 37°C (BdJ). The geological context and detailed composition of waters and gases of the PHF are given by Monnin et al. (2014).
Both ST07 and ST09 submarine chimneys were transversally cut from the top part generating sections of ~10–30 cm in diameter (Figure 1). The most central parts of sections of the ST07 and ST09 carbonated chimneys and the small BdJ (BdJC) and RK (RKC) chimneys (~10 cm in height and ~3 cm in diameter) were crushed and (i) stored in sterile FalconTM tubes at −80°C prior to DNA extraction or (ii) placed overnight at 4°C in a hermetically-sealed serum bottle with a nitrogen gas headspace prior to H2-producing enrichments. Fluid end-member samples (checked by pH-values >10.5 and salinity < 2 g/L) were collected from BdJ vents (BdJF) and RK pool (RKF) using 60 mL sterile syringes, pooled in sterile plastic bottles, and stored in a portable icebox until filtration, a few hours after sampling. The fluids (2 L) were filtered through 0.2 μm pore-size IsoporeTM polycarbonate membrane filters (Millipore). The filters were then kept overnight at 4°C before cultivation or at −80°C prior to DNA extraction.
H2-producing enrichments were performed in duplicate using Hungate tubes containing the following basal medium components (per liter of distilled water): 0.1 g KH2PO4, 0.1 g K2HPO4, 0.1 g NH4Cl, 2 g NaCl, 0.1 g KCl, 0.1 g CaCl2.2H2O, 0.1 g yeast extract (Difco), 0.1 g cysteine hydrochloride and 10 mL trace mineral element solution (Balch et al., 1979). The initial pH was adjusted to 9.5 with NaOH. This pH above 9 can select alkalophilic microorganisms and enhances their diversity that is expected to be lower if pH is adjusted to 11. The basal medium was boiled and cooled down to room temperature under a continuous O2-free N2-flush. Five milliliter of this medium was then dispensed into Hungate tubes under anaerobic conditions and autoclaved (45 min, 120°C). Prior to inoculation, the following sterile solutions were injected in each tube: 0.1 mL of 2% Na2S.9H2O (reducing agent) and 0.1 mL of 8% Na2CO3 (to adjust and buffer the pH); biotrypcase (2 g/L), yeast extract (2 g/L) and glucose (2 g/L) were used as substrates. This final medium used for H2-producing enrichments was referred to as BYG medium.
The tubes were inoculated with 0.5 g of crushed chimney rock (named BdJC, RKC, or ST07 for the “Bain des Japonais,” “Rivière des Kaoris,” and “Aiguille de Prony” sample, respectively) or polycarbonate filters from 0.2 μm fluids filtration of 2 L of fluid (named respectively BdJF and RKF for the “Bain des Japonais” and “Rivière des Kaoris” samples). The suspensions were serially diluted in decimal steps using the same media (up to 10−6) and then incubated for 1 month at 37 and 55°C. Microbial growth was determined by measuring the increase in turbidity at 600 nm after insertion of Hungate tubes into the cuvette holder of a UV-visible spectrophotometer (Cary 50, Varian). One hundred microliters of the headspace was periodically collected in order to determine the H2 content in the gas phase. Gas composition (H2, O2, CH4, and CO2) was determined using a Shimadzu GC 8A gas chromatograph (GC) equipped with a thermal conductivity detector (GC/TCD; Alltech, USA). The H2 production was expressed in mM (mmol per L of culture). One to ten milliliters of the cultures were collected at the end of the experiments and then centrifuged (10,000 g, 10 min). The supernatants were stored at −20°C for further chemical analysis. The concentration of carbohydrates and soluble end-products of metabolism was determined by high-pressure liquid chromatography (HPLC) analysis and refractometric detection (Thermo Separation Products). Details of analytical operating conditions were previously described by Mei et al. (2014). All analyses were conducted in duplicate.
Positive H2-producing cultures were subcultured into the same BYG liquid medium, and then purified by repeated use of the Hungate roll-tube method (Hungate, 1969) with medium solidified with 1.6% (w/v) agar (Difco). Several colonies that had developed were picked and cultured in BYG liquid medium. The pure cultures were identified after DNA extraction followed by 16S rRNA gene amplification, cloning, and sequencing (see below).
DNA was extracted following the protocol previously described by Quéméneur et al. (2014). The matrices used were: 0.5 g of crushed carbonate chimneys or, one quarter of a 47 mm diameter polycarbonate filter or the bacterial cell pellet from 10 to 15 mL of positive H2-producing cultures and reference strains (used as control in PCR tests, see below). The concentration of DNA extracts was measured using Qubit® fluorometer (Invitrogen).
Two DNA samples from the deepest submarine PHF chimney (ST09) were used to obtain PHF metagenomes. Metagenomic libraries were prepared using 25 ng of DNA per sample. The construction kit was the Ultralow Ovation system (NuGen). The multiplex ligation adaptor mixes were L2DR_BC9 and L2DR_BC7. Fifteen PCR cycles were applied to generate sufficient materials for sequencing. Paired-end sequencing (2 × 100 nt) was performed on an Illumina HiSeq 1000 at Marine Biological Laboratory, Woods Hole, MA.
All merged paired-end reads for ST09 samples are available on the MG-RAST server (Meyer et al., 2008) under ID 4550491.3 (P27) and ID 4550492.3 (P28). There were 5,392,044 and 7,933,927 sequence reads from P27 and P28 metagenomes, respectively.
The taxonomic annotation of these merged paired-end reads was conducted in MG-RAST server. Briefly, a phylogenomic reconstruction of the ST09 samples was computed by using both the phylogenetic information contained in the SEED nr database and the similarities to the ribosomal RNA database (Meyer et al., 2008).
The PHF metagenomes (P27 and P28) were screened for hydA genes encoding the large subunit of the [FeFe]-hydrogenase. The amino acid sequences of HydA proteins (pfam02906) were retrieved from the NCBI protein database (in May 2014) and used locally as a specific database for similarity searches using PHF metagenomes as query with BLASTX tool (version 2.2.25+) with default algorithm parameters and an E-value cutoff of 10−5. The merged paired-end reads related to hydA genes with significant hits were then extracted from MG-RAST, aligned and then clustered into OTUs using an 80% identity threshold with UCLUST algorithm. The representative of each OTU was then searched with BLASTX against the NCBI nr database. The metagenomes sequences are available in NCBI public database (SRA) under accession numbers: SRX748869 (P27) and SRX748870 (P28).
Three degenerate primer sets were tested from environmental samples and reference strains to target hydA genes encoding [FeFe]-hydrogenases: (i) hydF1/hydR1 (Xing et al., 2008), (ii) FeFe-272F/ FeFe-427R (Boyd et al., 2009), and (iii) HydH1f/ HydH3r (Schmidt et al., 2010). The bacterial 16S rRNA genes from H2-producing cultures were amplified using the primer set 27F/907R (Lane, 1991). Each PCR mixture (20 μL) contained 1X GoTaq® Hot Start Green Master Mix (Promega), 200 nM of each primer, and 1–10 ng of genomic DNA. Reactions were conducted in a T100 thermal cycler (Bio-rad). The 16S rRNA genes were amplified as follows: 94°C for 2 min, followed by 30 cycles performed at 94°C for 30 s, 50°C for 30 s, and 72°C for 30 s, with a final extension at 72°C for 10 min. The hydA genes were amplified as described above, except that 40 cycles were performed and that hybridization were carried out from 50 to 62°C using a temperature gradient in order to determine the optimal annealing temperature of each primer pair. The PCR products were then checked by electrophoresis on a 1% agarose gel containing sight DNA stain. Positive and blank controls were carried out for all amplifications.
Triplicate PCR products were pooled and purified with NucleoSpin Gel and PCR Clean-up kit (Macherey-Nagel), according to the manufacturer's instructions. The purified PCR products were ligated into a pGEM®-T Easy vector and cloned into JM109 Escherichia coli competent cells (Promega), according to the manufacturer's instructions. Sequencing of the inserts was performed by Beckman Genomics (Takeley, Essex, UK) on plasmid minipreps using primers 27F and T7 for the 16S rRNA gene and the hydA genes, respectively.
Nonchimeric 16S rRNA gene sequences (checked with the online Bellerophon program) and translated hydA sequences were aligned using Muscle implemented in the MEGA6 software (Tamura et al., 2013). The program mothur was used to group sequences into operational taxonomic units (OTUs) based on 97% identity for 16S rRNA genes and 80% identity for HydA sequences (Baba et al., 2014). Mothur was also used to estimate richness (Chao, 1984) and to compute diversity indices (Shannon and Weaver, 1949; Simpson, 1949) for each hydA clone library. The Good's coverage C of each clone library was calculated according to the equation: C = 1–(n/N) where n is the number of OTU and N is the total number of clones in the library (Good, 1953). At least one HydA sequence deriving from translated hydA sequence and representing each OTU, designed as phylotype was further aligned using Muscle with related sequences retrieved from NCBI databases using BLAST tools. The MEGA6 software was also used for phylogenetic tree construction by the Maximum Likelihood method using bootstrap analysis on 1000 replicates (Felsenstein, 1985). The hydA gene and HydA sequences from environmental samples were deposited in the Genbank database under the accession numbers KT357617-KT357637. The 16S rRNA genes from bacterial cultures were deposited in the Genbank database under the accession numbers KR349722 (3b), KJ626326 (PROH2), and KR349723 (BJ2).
The mixtures of 16S rRNA gene amplicons were generated from a 341F/815R bacterial primer set, as previously described by Dowd et al. (2008), and were sequenced on a 454 GS-FLX Titanium sequencer (Roche Life Sciences, USA) by the Molecular Research Laboratory (Texas, USA). Raw sequence data generated by pyrosequencing were uploaded into QIIME 1.8.0 and processed as described by Caporaso et al. (2010). Briefly, sequences were qualitatively trimmed, aligned using Pynast and clustered into operational taxonomic units (OTUs) using a 97% identity threshold with UCLUST (Edgar, 2010). Taxonomic assignment was carried out with the RDP Classifier (Wang et al., 2007) with a minimum bootstrap confidence of 80%. BLAST searches against a non-redundant nucleotide database were performed for a representative sequence of each OTU. The OTU richness was assessed using non-parametric richness estimators. The Shannon and Simpson's diversity indices were also calculated. The 16S rRNA genes from environmental samples were deposited in the Genbank database under the accession numbers KT344933-KT344984.
The two PHF metagenomes obtained from the submarine ST09 site contained numerous sequences related to [FeFe]-hydrogenases (Tables S1, S2). A total of 292 and 1386 merged paired-end reads were assigned to [FeFe]-hydrogenases in samples P27 and P28, respectively. These 1678 whole putative hydA reads represented 0.011% of the total reads in average (0.005 and 0.017% for P27 and P28, respectively). The number of OTUs observed in P27 (n = 113; Table S1) was lower than that found in P28 (n = 395; Table S2) in agreement with the respective size of the two metagenomes.
Whichever the PHF metagenome considered, the HydA OTUs were mainly related to Firmicutes (64.6 and 66.4% of the total HydA reads in P27 and P28, respectively), in which Clostridiales-related sequences were dominant (Figure 2), in agreement with BLAST assignment of total bacterial merged paired-end reads which are widely associated with those of Firmicutes (28.3 and 28.8% of the total bacterial reads in P27 and P28, respectively; Figure 3). Among total merged paired-end reads related to Firmicutes sequences, Clostridia, and Bacilli accounted for the two major classes (95 and 96% of the Firmicutes reads in P27 and P28, respectively) and Clostridiales-related sequences were dominant (52% of the Firmicutes reads; Figure 3).
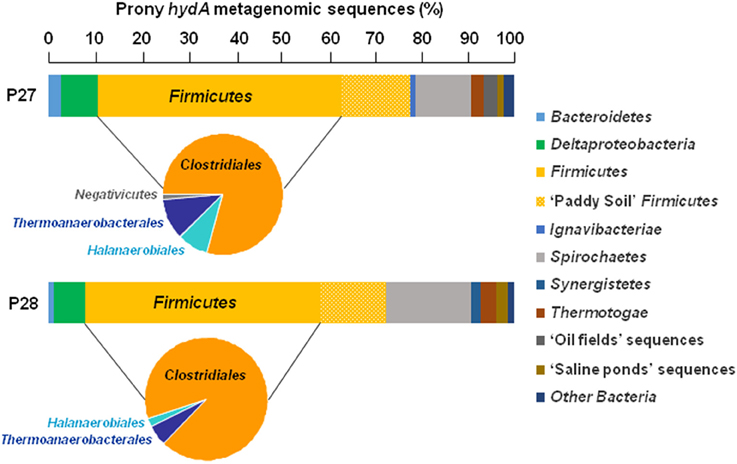
Figure 2. Distribution and taxonomic assignments of putative hydA gene sequences encoding the catalytic subunit of [Fe-Fe] hydrogenases in Prony Hydrothermal Field metagenomes. The histogram shows the relative abundance of major hydA phyla in the total hydA metagenomic sequences (n = 1678) and pie charts show distributions of “Firmicutes HydA” classes in the metagenomes P27 and P28 obtained from ST09 site.
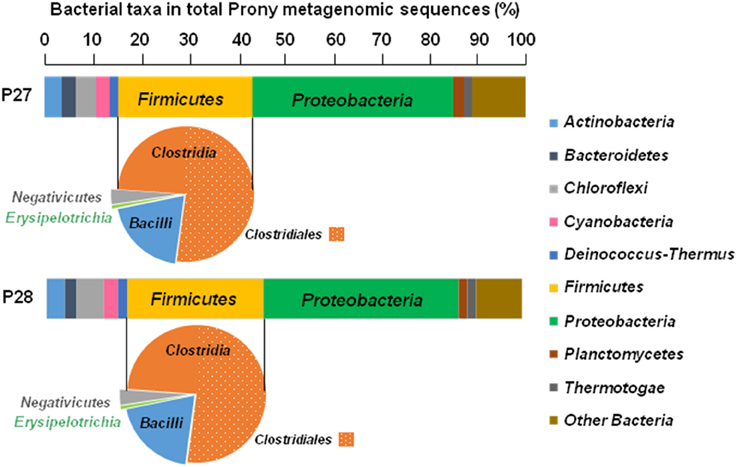
Figure 3. Distribution and taxonomic assignments of total bacterial sequences in Prony Hydrothermal Field metagenomes P27 and P28 obtained from ST09 site. The histogram shows the relative abundance of major bacterial phyla in the total bacterial metagenomic sequences and pie charts show distributions of major Firmicutes classes.
The majority of HydA OTUs had close phylogenetic relationships with Clostridium- and Desulfotomaculum-related sequences (Tables S1, S2). A relatively large proportion of HydA sequences (14.4–14.5%) from ST09 metagenomes shared high sequence similarity with those of paddy soil Firmicutes. Other HydA sequences within the Firmicutes (i.e., related to the orders Halanaerobiales, Thermoanaerobacterales, and Selenomonadales) were also observed but in less amounts (Figure 2). In addition, HydA sequences retrieved with high frequency were associated with those of Spirochaetales (11.9–18.3%), as well as Deltaproteobacteria (e.g., Desulfobulbus genus), Bacteroidetes, and Thermotogales, which are also known to be rich in fermentative H2-producers. The rest of metagenomic HydA sequences of PHF were associated with those of microbial mat inhabiting saline environments or oil fields (Boyd et al., 2009; Liu et al., 2015).
Five samples of both intertidal and submarine Prony sites (“Bain des Japonais” fluids, BdJF; “Bain des Japonais” chimney, BdJC; “Rivière des Kaoris” fluids, RKF; “Rivière des Kaoris” chimney, RKC, and ST07 chimney) were tested for amplification of the large subunit of the [FeFe]-hydrogenase encoding gene (hydA) using three degenerate primer sets (FeFe-272F/FeFe-427R, HydH1f /HydH3r, hydF1/hydR1). PCR products of ~680 bp were obtained with the primer set hydF1/hydR1 from both the PHF samples and the control genomic DNA (i.e., Desulfovibrio vulgaris and Clostridium saccharolyticum). In contrast, no hydA gene amplification occurred for the PHF samples by using primer sets FeFe-272F/FeFe-427R or HydH1f/HydH3r, despite modifications of the PCR conditions (e.g., annealing temperature) with validation on controls (i.e., PCR products with the expected band size were observed at 50°C for control). Therefore, the primer sets hydF1/hydR1 was selected for further analysis (i.e., cloning and sequencing of hydA genes of PHF DNAs).
Five hydA gene libraries were analyzed to study the HydA diversity in the five different PHF samples and to gain insight into the HydA diversity of uncultivable H2-producers. A total of 181 hydA sequences were obtained from the five environmental samples BdJC (52), BdJF (61), RKC (28), RKF (26), and ST07 (14). Both the highest richness and the highest diversity were observed in BdJ samples (Table S3).
Figure 4 shows the phylogenetic relationship of the 21 HydA OTUs corresponding to the translated sequences of the hydA gene detected in the PHF samples. They were related to the Firmicutes (81.0% of the total sequences), followed by Bacteroidetes (9.5%), and Alphaproteobacteria phyla (4.8%). The bacterial community of the submarine chimney ST07 was mainly represented (92.9% of the total sequences) by mesophilic to thermophilic fermenters or sulfate-reducers related to Firmicutes (i.e., Clostridiales or Thermoanaerobacterales) isolated from terrestrial hot spring or subterrestrial environments, such as Desulfotomaculum spp. These putative “submarine/anoxic Firmicutes” HydA sequences were also mainly detected in the metagenomes of the submarine chimney ST09. In contrast, both BdJC and RKC samples were dominated by HydA sequences associated with those of uncultured Firmicutes retrieved from a paddy field soil (78.8 and 92.9% of the total sequences, respectively), which were less represented in the metagenomes of the submarine chimney ST09. BdJF sample revealed a similar proportion of HydA sequences (73.5% of the total sequences) associated with this cluster hence referred to here as “intertidal/oxic Firmicutes HydA group.” The HydA sequences retrieved from RKF were mainly associated to the Bacteroidetes/Chlorobi group (96.2%) and also affiliated with uncultured Caldithrix–like bacteria retrieved from paddy field soil or anaerobic sludge (3.9%).
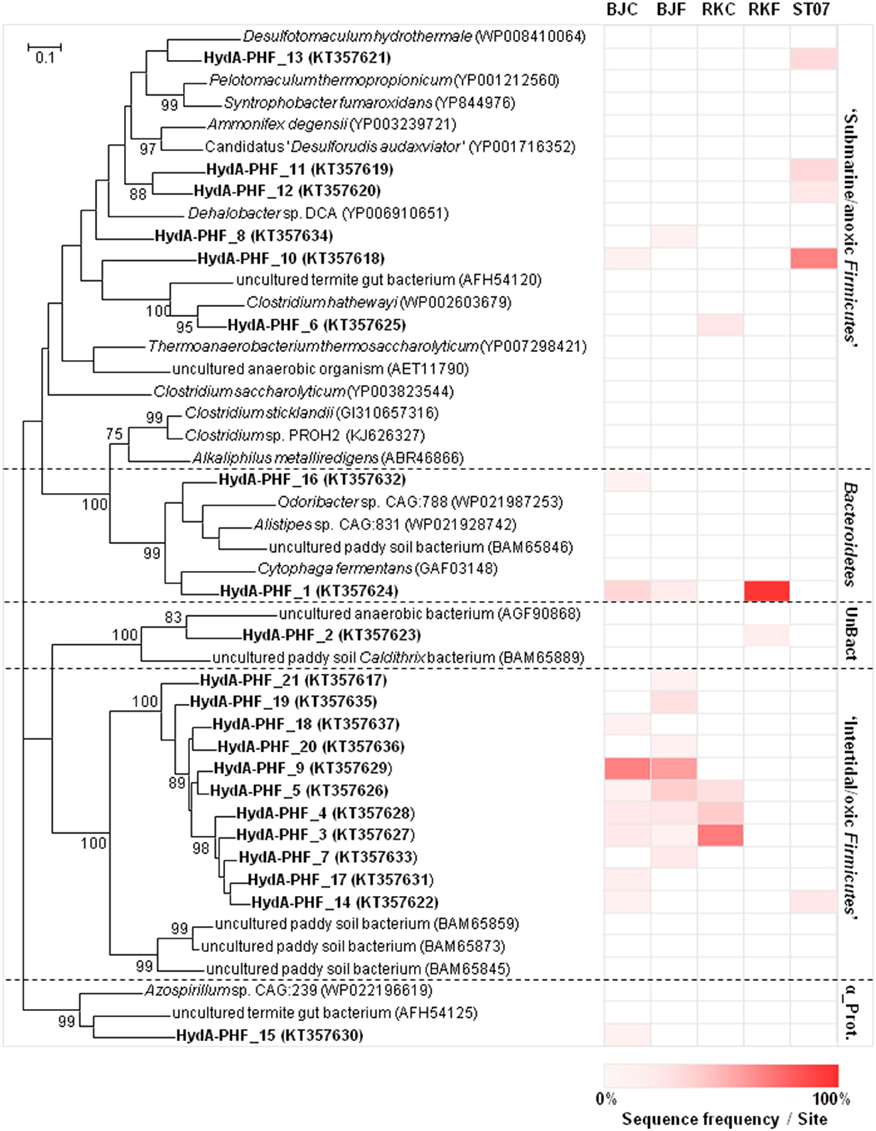
Figure 4. Maximum-likelihood phylogenetic tree of [Fe-Fe]-hydrogenase sequences obtained from intertidal and submarine PHF sites. Protein HydA sequences (188 amino acids) were obtained after translation of hydA gene sequences (obtained by cloning and Sanger sequencing) for 3 PHF hydrothermal sites: “Bain des Japonais” (BdJ), “Rivière des Kaoris” (RK), and “Aiguille de Prony” (ST07). The suffix “C” in sample names corresponds to “chimney” and the suffix “F” corresponds to “fluid”. The [FeFe]-hydrogenase sequences from PHF samples are marked in bold. Each OTU is represented by one representative sequence (at ≥80% similarity level). Percentage of OTU with respect to the total number of retrieved sequences in each PHF site is indicated by different levels of red from 0% (white) to 100% (red). Genbank accession numbers (in brackets) were obtained from the protein sequence database. Bootstrap values < 70% are not shown. The scale bar indicates 0.1% sequence divergence. UnBact and α_Prot stand for unclassified Bacteria and Alphaproteobacteria, respectively.
In order to obtain accurate and consistent estimates of the overall bacterial diversity in the studied sites, 16S rRNA pyrosequencing were conducted on the same five PHF subsamples (BdJF and RKF fluids; BdJC, RKC, and ST07 chimneys) previously used for hydA PCR amplification and subsequent sequencing. After quality/size trimming and removal of chimeric sequences, pyrosequencing of bacterial 16S rRNA PCR products yielded a total of 20,495 sequences. These sequences were assigned to 969 OTUs (RKC: 161; RKF: 143; BDJC: 323; BDJF: 125; ST07: 217; Table S3). The PHF samples were dominated by 52 bacterial OTUs (Table 1). BdJC had the highest bacterial richness while ST07 sample displayed the highest bacterial diversity (Table S3).
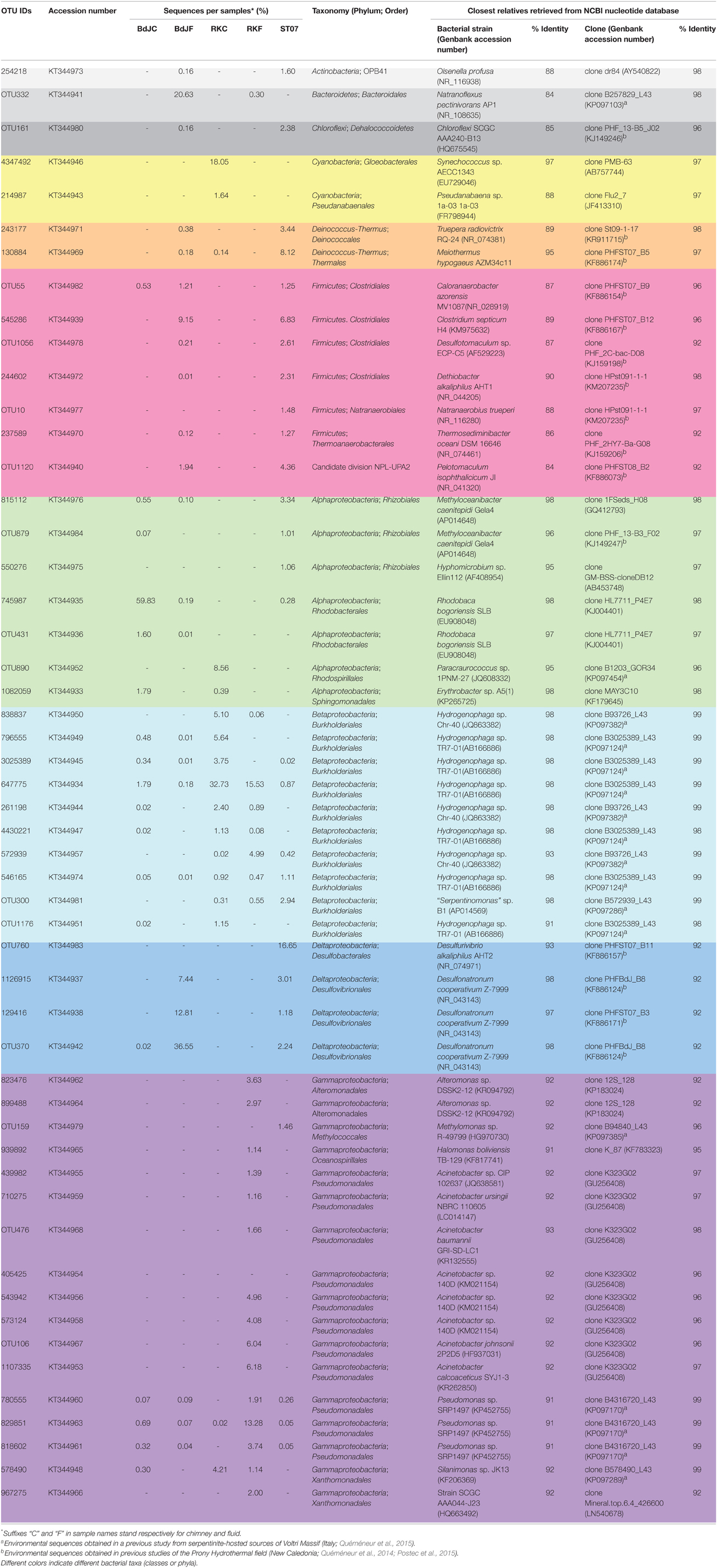
Table 1. Phylogenetic affiliation of the dominant bacterial OTUs obtained from 16S rRNA pyrosequencing analyses of Prony Hydrothermal Field (PHF) sites: “Bain des Japonais” (BdJ), “Rivière des Kaoris” (RK), and “Aiguille de Prony” (ST07).
The global structure of the bacterial communities obtained for each sample is shown in Figure 5. The total bacterial OTUs were assigned to 24 phyla and candidate phyla (including candidate phylum NPL-UPA2), and other unclassified groups (Figure 5). Bacteroidetes, Cyanobacteria, Firmicutes, Proteobacteria, and Thermus-Deinococcus accounted for the five main phyla (>10% of the total sequences each in average), representing 93.5% of the total bacterial communities.
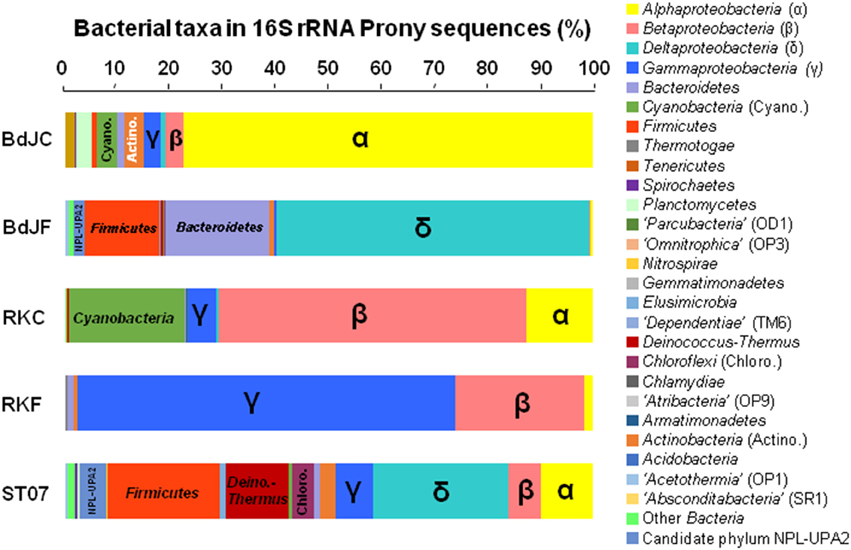
Figure 5. Distribution and taxonomic assignments of 16S rRNA bacterial sequences in chimneys and fluids of interdidal and submarine sites of PHF. Relative phylogenetic abundance was based on frequencies of the 16S rRNA gene sequences (with respect to the total number of sequences obtained by pyrosequencing) affiliated to major phylogenetic phyla or class in the bacterial communities of the “Bain des Japonais” (BdJ), “Rivière des Kaoris” (RK), and the “Aiguille de Prony” (ST07). The suffix “C” in sample names corresponds to “chimney” and the suffix “F” corresponds to “fluid.” The abbreviations of phyla and the former names of candidate phyla are indicated in parentheses.
BdJF and ST07 displayed the highest occurrence of Firmicutes and Deltaproteobacteria (Figure 5). Firmicutes were mainly represented by fermentative heterotrophs belonging to Clostridiales, Natranaerobiales (only detected in ST07), and Thermoanaerobacterales (Table 1). BdJF displayed the highest occurrence of Deltaproteobacteria sequences, which were exclusively affiliated to the alkaliphilic Desulfonatronum genus, while ST07 chimney sample comprised two alkaliphilic deltaproteobacterial groups: the first and most abundant related to the genus Desulfurivibrio, and the second related to the genus Desulfonatronum. BdJF bacterial community contained a large part (14.3%) of anaerobic, haloalkaliphilic, hydrolytic, and fermenting members of the Natronoflexus genus (Bacteroidetes phylum).
Remarkably, on the contrary to BdJF and ST07, the BdJC sample was dominated by Alphaproteobacteria, mainly represented by the anoxygenic phototrophic Rhodobaca genus. Moreover, photosynthetic Cyanobacteria were abundantly detected in RKC sample (21.9% of the bacterial community), while RKF bacteria were largely dominated by Gammaproteobacteria. Finally, H2-consuming Betaproteobacteria (i.e., Hydrogenophaga/“Serpentinomonas” members) were detected in all samples, especially in RKC and RKF, where they represented 58.4 and 24.4% of the total bacterial communities, respectively.
Two subsamples of the five PHF samples (BdJC, BdJF, RKC, RKF, and ST07) were cultivated into BYG medium at an initial pH of 9.5 in order to enrich for fermentative alkaliphilic H2-producing bacteria. In total, 70 tubes were inoculated (5 samples × 7 dilutions × 2 replicates). After 1 week of incubation at 37°C, H2 production was detected for all tested chimney samples (i.e., RKC, BdJC, and ST07; Figure 6A). The H2 production was accompanied by a decrease in both glucose concentration and pH (final pH 8.5 ± 0.5), as a result of acidic metabolite production (i.e., acetate and butyrate). No H2 production was observed in controls and enrichment cultures from fluids (BdJF and RKF) even after 1 month of incubation under the same culture conditions (37°C, pH 9.5). Moreover, no H2 production was observed after 1 month of incubation at 55°C with similar pH and substrate conditions from any tested samples (chimney samples and fluids). This result suggests that no thermo-alkaliphilic fermentative microorganisms were involved in production of H2 in PHF with these substrates.
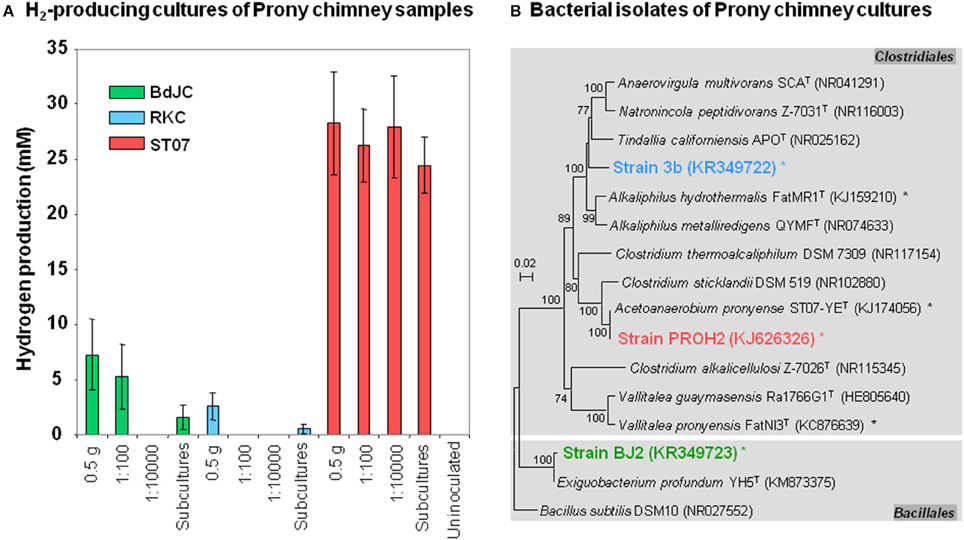
Figure 6. Hydrogen-producing cultures obtained from hydrothermal chimneys of the Prony bay: performances and identification of bacterial isolates. (A) Hydrogen-producing cultures were attempted using several dilutions of chimney subsamples from three Prony sites: “Bain des Japonais” (BdJ), “Rivière des Kaoris” (RK), and the “Aiguille de Prony” (ST07). The suffix “C” in sample names stands for “chimney.” Subcultures were also performed from the first round of positive enrichment cultures. Values are mean of biological replicates ± standard deviation (errors bars). (B) Maximum-likelihood tree based on 16S rRNA gene sequences showing the phylogenetic position of Firmicutes strains 3b, PROH2, and BJ2, isolated from hydrogen-producing cultures of Prony hydrothermal Field (PHF). Representative sequences in the tree were obtained from GenBank (accession number in the brackets). Other bacterial species isolated from PHF are indicated by asterisks. Bootstrap values >70% are indicated at nodes.
The RKC culture and the lowest dilution of BdJC (10−2) and ST07 (10−2) cultures yielding a significant H2 production were transferred into a fresh culture medium after 2 weeks of incubation. The highest H2 production (24.5 ± 2.5 mM) was reached in ST07 subcultures (Figure 6A), while extremely low H2 production (< 1.5 mM) was observed in both BdJC and RKC subcultures (after 7 days of incubation). All H2-producing subcultures (BdJC, RKC, and ST07) displayed before the isolation procedure, a weak bacterial richness with the detection of only one dominant OTU belonging to Firmicutes after 16S rRNA-based cloning/sequencing analyses (data not shown). These dominant OTUs corresponded to the bacterial strains finally isolated by the roll-tube method from each H2-producing subcultures. As shown in Figure 6B, the alkaliphilic, highly efficient H2-producing ST07 culture was dominated by the strain PROH2 (sharing 99.9 and 96.8% 16S rRNA identity with Acetoanaerobium pronyense and Clostridium sticklandii, respectively; Mei et al., 2014; Bes et al., 2015). The RKC cultures were dominated by the alkaliphilic strain 3b having Alkaliphilus hydrothermalis as its closest phylogenetic relative (92.3% 16S rRNA identity), hence representing a novel species of a new genus in the order Clostridiales, for which the name “Serpentinicella alkaliphila” has been recently proposed (Mei et al., 2016). The BdJC cultures were dominated by strain BJ2 closely related to Exiguobacterium profundum (99.5% 16S rRNA identity) a facultative anaerobe originating from a deep-sea hydrothermal vent and belonging to the order Bacillales.
Bacterial populations harboring hydA genes in both intertidal and submarine PHF samples were clearly dominated by Firmicutes, as previously observed in two other serpentinite-hosted ecosystems [i.e., Lost City chimneys, discharging hot fluid (90°C) with high pH (10.8), and H2 (13 mmol/kg) and anoxic Tablelands-WHC2b fluid, pH 12.1, Eh −733 mV, 0.24 mM-H2; Brazelton et al., 2012; Figure 4]. However, different hydA patterns were observed depending on the nature and physico-chemical characteristics of the PHF samples (e.g., fluid vs. carbonate concretions/chimneys, or water depth), in agreement with previous 16S rRNA gene-based molecular studies on this hydrothermal field (Quéméneur et al., 2014; Postec et al., 2015; this study). PHF hydA populations were mainly divided in two groups. One is the “submarine/anoxic HydA group,” related to strictly anaerobic Firmicutes (e.g., Clostridium and Desulfotomaculum genera) recovered from deep environments (Haouari et al., 2008; Aüllo et al., 2013). This is the case of submarine ST07 and ST09 chimneys, as well as fluid end-members of BdJ site [BdJF, pH 10.8, Eh −352 mV, 6.4 mM-H2(g)] and RK site [RKF, pH 10.9, Eh −195, 13.4 mM-H2(g)] (data from Monnin et al., 2014) characterized by pH close to 11 and low redox potentiel. In contrast, an “intertidal/oxic HydA group,” affiliated to Firmicutes HydA sequences from paddy field soil (Japan; Baba et al., 2014), was predominant in the intertidal BdJ and RK chimneys. Irrigated paddy field soils are characterized by alternating aerobic and anaerobic conditions, when they are drained and flooded during rice cultivation periods (Lüdemann et al., 2000). Similar fluctuating exposure/concentration of oxygen may exist in both intertidal BdJ and RK chimneys, which are alternatively uncovered or covered by seawater at low or high tide, respectively (Monnin et al., 2014; Quéméneur et al., 2014). However, hydA genes are commonly detected abundantly in anoxic zones, but not in other intertidal/oxic locations (e.g., Tablelands-TLE and Great Salt Lake; Brazelton et al., 2012; Boyd et al., 2014). Therefore, this new “intertidal/oxic HydA group” distantly related to HydA sequences from cultivated microorganisms related to uncultured Firmicutes from paddy soils and may hence represent a new class of unknown [Fe-Fe]-hydrogenases of new aero-tolerant or microaerophilic microorganisms to be discovered (Figure 4).
Numerous HydA OTUs obtained from submarine PHF sites by using both metagenomic and PCR-amplified DNA sequencing analyses were closely related to sulfate-reducing Firmicutes of the Desulfotomaculum genus. This finding is consistent with a previous metagenomic investigation of [Fe-Fe]-hydrogenases in the serpentinization-driven LCHF (Brazelton et al., 2012). Desulfotomaculum spp. are well-adapted to colonize deep submarine environments where they are nonetheless better known to consume H2 for growth through sulfate reduction (Aüllo et al., 2013) rather than to produce H2 by fermentation. However, some Desulfotomaculum species possess the ability to grow in syntrophy with hydrogenotrophic methanogens (to which they transfer H2 they produce) and have even lost their ability to reduce sulfate in anoxic systems (Imachi et al., 2006), when the concentration of sulfate is quite low, as it was measured in PHF end member fluids (Monnin et al., 2014; Quéméneur et al., 2014). Some [FeFe]-hydrogenases of other sulfate-reducers (e.g., Desulfovibrio spp.) may also be bifunctional, and depending on the environmental conditions, they may produce H2 in synthrophic conditions, instead of catalyzing H2 oxidation (Meyer et al., 2013). In PHF chimney samples, no Desulfotomaculum species was previously detected by sequence analyses of dsrB genes, used as molecular marker of sulfate-reducing bacteria (Quéméneur et al., 2014; Postec et al., 2015) in contrast to the LCHF where they were found to be abundant (Brazelton et al., 2006; Gerasimchuk et al., 2010). Several attempts to cultivate sulfate-reducing bacteria, as well as anaerobic hydrogenotrophs, from these submarine serpentinite-hosted environments were unsuccessful for both locations (Postec et al., 2015), despite their abundance in both 16S rRNA and dsrB gene databases (e.g., Desulfotomaculum spp. for LCHF and Desulfonatronum spp. for PHF). However, it is well-known that the use of H2 as electron donor is generally considered as more thermodynamically favorable than use of organic acids or sugars as electron donor (Thauer et al., 1977; Amend et al., 2011). This probably means that such sulfate-reducing bacteria have a particular metabolism (and/or a synthrophic life style), and consequently may possess a novel type of hydrogenases. Altogether, these results indicate that Desulfotomaculum–related [Fe-Fe]-hydrogenases certainly play a crucial role in the biological H2 cycle in serpentinite-hosted environments but at this stage of knowledge it is difficult to stand if these [Fe-Fe]-hydrogenases were involved mostly in H2 production or consumption or both depending on environmental conditions.
An unexpectedly large HydA diversity, related to phyla that include Alphaproteobacteria, Bacteroidetes, and Firmicutes, was observed in the alkaline serpentinite-hosted PHF, where high amounts of H2, likely of mixed origin (geological and biological), is produced. This finding strongly contrasts with that reported by Boyd et al. (2010) who also used HydA sequences to study the diversity of H2-producing bacteria in basalt-hosted hydrothermal springs of the Yellowstone National Park (YNP). Indeed, they demonstrated that the HydA diversity in their samples was strongly constrained by pH (with the lowest diversity being found in springs with high pH of 9–10) and was mainly represented by uncultivated members of the Elusimicrobia phylum (known as “Termite Group 1”) at high pH. Although the geological, mineralogical and chemical setting differs in PHF, we show that neither pH solely nor in combination with in situ H2 concentration can explain the HydA diversity in the PHF system, and that alkaline habitats can also harbor a wide range of potential H2-producers, comparable to previous observations in habitats with acidic or neutral pH conditions (Xing et al., 2008; Schmidt et al., 2010). This sharp difference with what was observed in YNP hot springs may be explained by the combination of two strong environmental stresses (i.e., high pH coupled with high temperature) that seems to dramatically decrease the potential of biological H2 production. In our study, no H2 production was detected in enrichments carried out at pH 9.5 and temperature exceeding 55°C and only low proportions of the hydA sequences retrieved from PHF metagenomes could be affiliated to thermophilic H2 producers (such as Thermotogales), in agreement with results obtained from YNP springs, where hydA genes were undetected at alkaline pH and temperature above 65°C (Boyd et al., 2010).
Whatever the approach used, it appears that the order Clostridiales, phylum Firmicutes, contained the highest number of hydA genes and thus potential H2-producing candidates in PHF samples. Our molecular data are in agreement with recent extensive genomic studies, showing a predominance of hydA genes in Firmicutes genomes (Peters et al., 2015; Poudel et al., 2016). Among them, the majority of the hydA Firmicutes are related to the “G1 Hyd group,” which mainly contains representatives of H2-producing [FeFe]-hydrogenases (Poudel et al., 2016). Clostridiales were also the most frequently cultivated bacteria from PHF chimneys, allowing us to isolate numerous strains, some of which being already described as new species (e.g., Alkaliphilus hydrothermalis, Acetoanaerobium pronyense, Vallitalea pronyensis; Ben Aissa et al., 2014, 2015; Bes et al., 2015). These Clostridiales can be involved in H2 production by fermenting a wide range of organic compounds as substrates (e.g., sugars, proteins, individual amino acids, carboxylic acids), which could originate from the decay of primary microbial colonizers of such alkaline environments. Indeed, a high biomass has been previously detected in PHF chimneys with population ranging from 1 to 6 × 107 bacterial cells per gram of chimneys (Quéméneur et al., 2014). Additionally, the hydrothermal degradation at depth of serpentinite hosted ecosystems has been shown to lead to the production of organic acids circulating throughout the hydrothermal system (Pasini et al., 2013). However, further studies on fermentative H2 producers occupying these alkaline habitats are needed to ascertain their geomicrobiological role to be played in serpentinite-hosted ecosystems and to assess to what extent they contribute to the H2 budget. Most of the studies on fermentative H2 production were conducted under acidic or neutral pH conditions (optimal pH ranging from 5 to 6; Xing et al., 2008; Wang and Wan, 2009; Quéméneur et al., 2011), and only two alkaliphilic H2-producers have been isolated from alkaline environments so far (Begemann et al., 2012; Mei et al., 2014). Nonetheless, fermentative H2 production have been not only described as thermodynamically more attractive under alkaline conditions (at ambient temperature), but also reported to be enhanced and stabilized at high initial pH (Cai et al., 2004; Xiao and Liu, 2006). Besides, the alkaliphilic anaerobe, Clostridium sp. PROH2, isolated from the “Aiguille de Prony” (ST07 chimney), demonstrated efficient H2 production with H2 yields similar to that of other neutrophilic and mesothermic clostridial species studied so far. This clostridial strain was able to produce 2.71 moles of H2 per mole of glucose at high pH (9.5), low salinity and moderate temperature (37°C; Mei et al., 2014). Such pure cultures of extremophilic microorganisms constitute interesting biotechnological alternatives for producing H2 with high efficiency from vegetal biomass and organic wastes in non-sterile systems since the high-pH conditions efficiently prevent growth of most contaminants that prevail under neutrophilic conditions.
This study revealed an unexpected high diversity of [FeFe]-hydrogenase genes mostly related to Firmicutes in the hyperalkaline and serpentinite-hosted PHF. Such a high diversity may reflect either a high metabolic capability at the community level, with various fermenting bacteria occupying distinct micro-habitats in the porous structures of the carbonate chimneys and in the fluids, or individual metabolic flexibility of these indigenous microorganisms adapted to the various stresses they have to face due to harsh and fluctuating environmental conditions (e.g., Eh, pH, O2, nutrient deprivation) as recently evidenced by Pisapia et al. (in press). Their novelty can be explained by the unique feature of the serpentinite-hosted PHF, which discharges low-temperature (< 40°C) and low-salinity fluids in a shallow submarine environment. Clearly, further investigations are mandatory to assess the in situ functioning and directionality (i.e., reverse or forward) of these new and diverse [FeFe]-hydrogenases associated to members of the order Clostridiales. They also need to be complemented by isotopic investigations aiming at determining the ratio of biotic to abiotic H2 produced at the PHF, and field measurements to in situ assess the microbiological H2 production. As elevated concentrations of N2 were also reported in the fluids discharged at the PHF (Monnin et al., 2014; Deville and Prinzhofer, 2016), the involvement of serpentinite-hosted microbial communities in the deep nitrogen cycle and in the overall production of N2 will be another pending question to tackle within the next future.
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This project was financially supported by the French Research Institute for Development (IRD), the EC2CO-Biohefect/Ecodyn/Dril/MicrobiEn (MicroProny) CNRS program, the deepOASES ANR project (ANR-14-CE01-0008-06), the Deep Carbon Observatory (Census for Deep Life: Comparative metagenomics of archaeal biofilms in carbonate chimneys associated with geographically isolated sites of serpentinization; PI W. J. Brazelton) and the China Scholarship Council. We acknowledge the support given by the Deep Life Community from the Alfred P. Sloan Foundation to M. L. Sogin and K. Hinrichs. We sincerely thank M. O. Schrenk and W. J. Brazelton for their helpful reading and comments on the manuscript. We are grateful to the scuba divers team of IRD Nouméa (Eric Folcher, John Butcher, and Bertrand Bourgeois) for collecting samples.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01301
Amend, J. P., McCollom, T. M., Hentscher, M., and Bach, W. (2011). Catabolic and anabolic energy for chemolithoautotrophs in deep-sea hydrothermal systems hosted in different rock types. Geochim. Cosmochim. Acta 75, 5736–5748. doi: 10.1016/j.gca.2011.07.041
Aüllo, T., Ranchou-Peyruse, A., Ollivier, B., and Magot, M. (2013). Desulfotomaculum spp. and related gram-positive sulfate-reducing bacteria in deep subsurface environments. Front. Microbiol. 4:362. doi: 10.3389/fmicb.2013.00362
Baba, R., Kimura, M., Asakawa, S., and Watanabe, T. (2014). Analysis of [FeFe]-hydrogenase genes for the elucidation of a hydrogen-producing bacterial community in paddy field soil. FEMS Microbiol. Lett. 350, 249–256. doi: 10.1111/1574-6968.12335
Balch, W. E., Fox, G. E., Magrum, L. J., Woese, C. R., and Wolfe, R. S. (1979). Methanogens: reevaluation of a unique biological group. Microbiol. Rev. 43, 260–296.
Begemann, M. B., Mormile, M. R., Sitton, O. C., Wall, J. D., and Elias, D. A. (2012). A streamlined strategy for biohydrogen production with Halanaerobium hydrogeniformans, an alkaliphilic bacterium. Front. Microbiol. 3:93. doi: 10.3389/fmicb.2012.00093
Ben Aissa, F., Postec, A., Erauso, G., Payri, C., Pelletier, B., Hamdi, M., et al. (2014). Vallitalea pronyensis sp. nov., isolated from a marine alkaline hydrothermal chimney. Int. J. Syst. Evol. Microbiol. 64, 1160–1165. doi: 10.1099/ijs.0.055756-0
Ben Aissa, F., Postec, A., Erauso, G., Payri, C., Pelletier, B., Hamdi, M., et al. (2015). Characterization of Alkaliphilus hydrothermalis strain FatMR1 sp. nov., a novel alkaliphilic anaerobic bacterium, isolated from a carbonaceous chimney of the Prony Hydrothermal Field, New Caledonia. Extremophiles 19, 183–188. doi: 10.1007/s00792-014-0697-y
Bes, M., Merrouch, M., Joseph, M., Quéméneur, M., Payri, C., Pelletier, B., et al. (2015). Acetoanaerobium pronyense sp. nov., an anaerobic mesophilic bacterium isolated from the Prony alkaline Hydrothermal Field, New Caledonia. Int. J. Syst. Evol. Microbiol. 65, 2574–2580. doi: 10.1099/ijs.0.000307
Boyd, E. S., Hamilton, T. L., Spear, J. R., Lavin, M., and Peters, J. W. (2010). [FeFe]-hydrogenase in Yellowstone National Park: evidence for dispersal limitation and phylogenetic niche conservatism. ISME J. 4, 1485–1495. doi: 10.1038/ismej.2010.76
Boyd, E. S., Hamilton, T. L., Swanson, K. D., Howells, A. E., Baxter, B. K., Meuser, J. E., et al. (2014). [FeFe]-hydrogenase abundance and diversity along a vertical redox gradient in Great Salt Lake, USA. Int. J. Mol. Sci. 15, 21947–21966. doi: 10.3390/ijms151221947
Boyd, E. S., Spear, J. R., and Peters, J. W. (2009). [FeFe]-hydrogenase genetic diversity provides insight into molecular adaptation in a saline microbial mat community. Appl. Environ. Microbiol. 75, 4620–4623. doi: 10.1128/AEM.00582-09
Brazelton, W. J., Morrill, P. L., Szponar, N., and Schrenk, M. O. (2013). Bacterial communities associated with subsurface geochemical processes in continental serpentinite springs. Appl. Environ. Microbiol. 79, 3906–3916. doi: 10.1128/AEM.00330-13
Brazelton, W. J., Nelson, B., and Schrenk, M. O. (2012). Metagenomic evidence for H2 oxidation and H2 production by serpentinite-hosted subsurface microbial communities. Front. Microbiol. 2:268. doi: 10.3389/fmicb.2011.00268
Brazelton, W. J., Schrenk, M. O., Kelley, D. S., and Baross, J. A. (2006). Methane-and sulfur-metabolizing microbial communities dominate the Lost City hydrothermal field ecosystem. Appl. Environ. Microbiol. 72, 6257–6270. doi: 10.1128/AEM.00574-06
Cai, M., Liu, J., and Wei, Y. (2004). Enhanced biohydrogen production from sewage sludge with alkaline pretreatment. Environ. Sci. Technol. 38, 3195–3202. doi: 10.1021/es0349204
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Chao, A. (1984). Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 11, 265–270.
Deville, E., and Prinzhofer, A. (2016). The origin of N2-H2-CH4-rich natural gas seepages in ophiolitic context: a major and noble gases study of fluid seepages in New Caledonia. Chem. Geol. 440, 139–147. doi: 10.1016/j.chemgeo.2016.06.011
Dowd, S. E., Callaway, T. R., Wolcott, R. D., Sun, Y., McKeehan, T., Hagevoort, R. G., et al. (2008). Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiol. 8:125. doi: 10.1186/1471-2180-8-125
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Felsenstein, J. (1985). Confidence-limits on phylogenies - an approach using the bootstrap. Evolution 39, 783–791. doi: 10.2307/2408678
Gerasimchuk, A. L., Shatalov, A. A., Novikov, A. L., Butorova, O. P., Pimenov, N. V., Lein, A. Y., et al. (2010). The search for sulfate-reducing bacteria in mat samples from the lost city hydrothermal field by molecular cloning. Microbiology 79, 96–105. doi: 10.1134/S0026261710010133
Good, I. J. (1953). The population frequencies of species and the estimation of population parameters. Biometrika 40, 237–264. doi: 10.1093/biomet/40.3-4.237
Haouari, O., Fardeau, M.-L., Cayol, J. L., Casiot, C., Elbaz-Poulichet, F., Hamdi, M., et al. (2008). Desulfotomaculum hydrothermale sp. nov., a thermophilic sulfate-reducing bacterium isolated from a terrestrial Tunisian hot spring. Int. J. Syst. Evol. Microbiol. 58, 2529–2535. doi: 10.1099/ijs.0.65339-0
Hungate, R. E. (1969). A roll tube method for cultivation of strict anaerobes. Method. Microbiol. 3B, 117–132. doi: 10.1016/S0580-9517(08)70503-8
Imachi, H., Sekiguchi, Y., Kamagata, Y., Loy, A., Qiu, Y. L., Hugenholtz, P., et al. (2006). Non-sulfate-reducing, syntrophic bacteria affiliated with Desulfotomaculum cluster I are widely distributed in methanogenic environments. Appl. Environ. Microbiol. 72, 2080–2091. doi: 10.1128/AEM.72.3.2080-2091.2006
Kelley, D. S., Karson, J. A., Früh-Green, G. L., Yoerger, D. R., Shank, T. M., Butterfield, D. A., et al. (2005). A serpentinite-hosted ecosystem: the Lost City hydrothermal field. Science 307, 1428–1434. doi: 10.1126/science.1102556
Lane, D. J. (1991). “Nucleic acid techniques in bacterial systematics,” in Nucleic Acid Techniques in Bacterial Systematics, eds E. Stackebrandt and M. Goodfellow (Hoboken, NJ: Wiley), 115–175.
Launay, J., and Fontes, J. C. (1985). Les sources thermales de Prony (Nouvelle–Caledonie) et leurs precipites chimiques. Exemple de formation de brucite primaire. Geologie de la France 1, 83–100.
Liu, J. F., Sun, X. B., Yang, G. C., Mbadinga, S. M., Gu, J. D., and Mu, B. Z. (2015). Analysis of microbial communities in the oil reservoir subjected to CO2-flooding by using functional genes as molecular biomarkers for microbial CO2 sequestration. Front. Microbiol. 6:236. doi: 10.3389/fmicb.2015.00236
Lubitz, W., Ogata, H., Rüdiger, O., and Reijerse, E. (2014). Hydrogenases. Chem. Rev. 114, 4081–4148. doi: 10.1021/cr4005814
Lüdemann, H., Arth, I., and Liesack, W. (2000). Spatial changes in the bacterial community structure along a vertical oxygen gradient in flooded paddy soil cores. Appl. Environ. Microbiol. 66, 754–762. doi: 10.1128/AEM.66.2.754-762.2000
Mei, N., Postec, A., Erauso, G., Joseph, M., Pelletier, B., Payri, C., et al. (2016). Serpentinicella alkaliphila gen. nov., sp. nov., a novel alkaliphilic anaerobic bacterium isolated from the serpentinite-hosted Prony hydrothermal field, New Caledonia. Int. J. Syst. Evol. Microbiol. doi: 10.1099/ijsem.0.001375. [Epub ahead of print].
Mei, N., Zergane, N., Postec, A., Erauso, G., Ollier, A., Payri, C., et al. (2014). Fermentative hydrogen production by a new alkaliphilic Clostridium sp. (strain PROH2) isolated from a shallow submarine hydrothermal chimney in Prony Bay, New Caledonia. Int. J. Hydrogen Energy 39, 19465–19473. doi: 10.1016/j.ijhydene.2014.09.111
Meyer, B., Kuehl, J., Deutschbauer, A. M., Price, M. N., Arkin, A. P., and Stahl, D. A. (2013). Variation among Desulfovibrio species in electron transfer systems used for syntrophic growth. J. Bacteriol. 195, 990–1004. doi: 10.1128/JB.01959-12
Meyer, F., Paarmann, D., D'Souza, M., Olson, R., Glass, E. M., Kubal, M., et al. (2008). The metagenomics RAST server–a public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinformatics 9:386. doi: 10.1186/1471-2105-9-386
Miller, H. M., Matter, J. M., Kelemen, P., Ellison, E. T., Conrad, M. E., Fierer, N., et al. (2016). Modern water/rock reactions in Oman hyperalkaline peridotite aquifers and implications for microbial habitability. Geochim. Cosmochim. Acta 179, 217–241. doi: 10.1016/j.gca.2016.01.033
Monnin, C., Chavagnac, V., Boulart, C., Ménez, B., Gérard, M., Gérard, E., et al. (2014). Fluid chemistry of the low temperature hyperalkaline hydrothermal system of the Prony Bay (New Caledonia). Biogeosciences 11, 5687–5706. doi: 10.5194/bg-11-5687-2014
Pasini, V., Brunelli, D., Dumas, P., Sandt, C., Frederick, J., Benzerara, K., et al. (2013). Low temperature hydrothermal oil and associated biological precursors in serpentinites from Mid-Ocean Ridge. Lithos 178, 84–95. doi: 10.1016/j.lithos.2013.06.014
Peters, J. W., Schut, G. J., Boyd, E. S., Mulder, D. W., Shepard, E. M., Broderick, J. B., et al. (2015). [FeFe]-and [NiFe]-hydrogenase diversity, mechanism, and maturation. Biochim. Biophys. Acta Mol. Cell Res. 1853, 1350–1369. doi: 10.1016/j.bbamcr.2014.11.021
Pisapia, C., Gérard, E., Gérard, M., Lecourt, L., Pelletier, B., Payri, C., et al (in press). Mineralizing filamentous bacteria from the Prony bay Hydrothermal Field open new perspectives for serpentinization-based deep ecosystems. Front. Microbiol..
Postec, A., Quéméneur, M., Bes, M., Mei, N., BenaÏssa, F., Payri, C., et al. (2015). Microbial diversity in a submarine carbonate edifice from the serpentinizing hydrothermal system of the Prony Bay (New Caledonia) over a 6-year period. Front. Microbiol. 6:857. doi: 10.3389/fmicb.2015.00857
Poudel, S., Tokmina-Lukaszewska, M., Colman, D. R., Refai, M., Schut, G. J., King, P. W., et al. (2016). Unification of [FeFe]-hydrogenases into three structural and functional groups. Biochim. Biophys. Acta 1860, 1910–1921. doi: 10.1016/j.bbagen.2016.05.034
Quéméneur, M., Bes, M., Postec, A., Mei, N., Hamelin, J., Monnin, C., et al. (2014). Spatial distribution of microbial communities in the shallow submarine alkaline hydrothermal field of the Prony Bay, New Caledonia. Environ. Microbiol. Rep. 6, 665–674. doi: 10.1111/1758-2229.12184
Quéméneur, M., Hamelin, J., Latrille, E., Steyer, J.-P., and Trably, E. (2010). Development and application of a functional CE-SSCP fingerprinting method based on [Fe-Fe]-hydrogenase genes for monitoring hydrogen producing Clostridium in mixed cultures. Int. J. Hydrogen Energy 35, 13158–13167. doi: 10.1016/j.ijhydene.2010.07.076
Quéméneur, M., Hamelin, J., Latrille, E., Steyer, J.-P., and Trably, E. (2011). Functional versus phylogenetic fingerprint analyses for monitoring hydrogen-producing bacterial populations in dark fermentation cultures. Int. J. Hydrogen Energy 36, 3870–3879. doi: 10.1016/j.ijhydene.2010.12.100
Quéméneur, M., Palvadeau, A., Postec, A., Monnin, C., Chavagnac, V., Ollivier, B., et al. (2015). Endolithic microbial communities in carbonate precipitates from serpentinite-hosted hyperalkaline springs of the Voltri massif (Ligurian Alps, Northern Italy). Environ. Sci. Pollut. R. 22, 13613–13624. doi: 10.1007/s11356-015-4113-7
Schmidt, O., Drake, H. L., and Horn, M. A. (2010). Hitherto unknown [Fe-Fe]-hydrogenase gene diversity in anaerobes and anoxic enrichments from a moderately acidic fen. Appl. Environ. Microbiol. 76, 2027–2031. doi: 10.1128/AEM.02895-09
Schrenk, M. O., Brazelton, W. J., and Lang, S. Q. (2013). Serpentinization, carbon, and deep life. Rev. Mineral. Geochem. 75, 575–606. doi: 10.2138/rmg.2013.75.18
Schut, G. J., and Adams, M. W. (2009). The iron-hydrogenase of Thermotoga maritime utilizes ferredoxin and NADH synergistically: a new perspective on anaerobic hydrogen production. J. Bacteriol. 191, 4451–4457. doi: 10.1128/JB.01582-08
Shannon, C. E., and Weaver, W. (1949). The Mathematical Theory of Communication. Urbana, IL: University of Illinois Press.
Suzuki, S., Kuenen, J. G., Schipper, K., van der Velde, S., Ishii, S. I., Wu, A., et al. (2014). Physiological and genomic features of highly alkaliphilic hydrogen-utilizing Betaproteobacteria from a continental serpentinizing site. Nat. Commun. 5:3900. doi: 10.1038/ncomms4900
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Thauer, R. K., Jungermann, K., and Decker, K. (1977). Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 41, 100.
Tiago, I., and Veríssimo, A. (2013). Microbial and functional diversity of a subterrestrial high pH groundwater associated to serpentinization. Environ. Microbiol. 15, 1687–1706. doi: 10.1111/1462-2920.12034
Vignais, P. M., Billoud, B., and Meyer, J. (2001). Classification and phylogeny of hydrogenases. FEMS Microbiol. Rev. 25, 455–501. doi: 10.1111/j.1574-6976.2001.tb00587.x
Vignais, P. M., and Colbeau, A. (2004). Molecular biology of microbial hydrogenases. Curr. Issues Mol. Biol. 6, 159–188.
Wang, J., and Wan, W. (2009). Factors influencing fermentative hydrogen production: a review. Int. J. Hydrogen Energy 34, 799–811. doi: 10.1016/j.ijhydene.2008.11.015
Wang, Q., Garrity, G. M., Tiedje, J. M., and Cole, J. R. (2007). Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267. doi: 10.1128/AEM.00062-07
Wang, S., Huang, H., Kahnt, J., and Thauer, R. K. (2013). A reversible electron-bifurcating ferredoxin-and NAD-dependent [FeFe]-hydrogenase (HydABC) in Moorella thermoacetica. J. Bacteriol. 195, 1267–1275. doi: 10.1128/JB.02158-12
Xiao, B., and Liu, J. (2006). pH dependency of hydrogen fermentation from alkali-pretreated sludge. Chin. Sci. Bull. 51, 399–404. doi: 10.1007/s11434-006-0399-7
Keywords: hydrogen, microbial diversity, hydrogen producers, serpentinization, hydA genes, [FeFe]-hydrogenase, metagenomics
Citation: Mei N, Postec A, Monnin C, Pelletier B, Payri CE, Ménez B, Frouin E, Ollivier B, Erauso G and Quéméneur M (2016) Metagenomic and PCR-Based Diversity Surveys of [FeFe]-Hydrogenases Combined with Isolation of Alkaliphilic Hydrogen-Producing Bacteria from the Serpentinite-Hosted Prony Hydrothermal Field, New Caledonia. Front. Microbiol. 7:1301. doi: 10.3389/fmicb.2016.01301
Received: 18 February 2016; Accepted: 08 August 2016;
Published: 30 August 2016.
Edited by:
Andreas Teske, University of North Carolina at Chapel Hill, USAReviewed by:
John R. Spear, Colorado School of Mines, USACopyright © 2016 Mei, Postec, Monnin, Pelletier, Payri, Ménez, Frouin, Ollivier, Erauso and Quéméneur. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marianne Quéméneur, marianne.quemeneur@ird.fr
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.