 Audrey Rivière
Audrey Rivière Marija Selak
Marija Selak David Lantin
David Lantin Frédéric Leroy
Frédéric Leroy- Research Group of Industrial Microbiology and Food Biotechnology, Faculty of Sciences and Bioengineering Sciences, Vrije Universiteit Brussel, Brussels, Belgium
With the increasing amount of evidence linking certain disorders of the human body to a disturbed gut microbiota, there is a growing interest for compounds that positively influence its composition and activity through diet. Besides the consumption of probiotics to stimulate favorable bacterial communities in the human gastrointestinal tract, prebiotics such as inulin-type fructans (ITF) and arabinoxylan-oligosaccharides (AXOS) can be consumed to increase the number of bifidobacteria in the colon. Several functions have been attributed to bifidobacteria, encompassing degradation of non-digestible carbohydrates, protection against pathogens, production of vitamin B, antioxidants, and conjugated linoleic acids, and stimulation of the immune system. During life, the numbers of bifidobacteria decrease from up to 90% of the total colon microbiota in vaginally delivered breast-fed infants to <5% in the colon of adults and they decrease even more in that of elderly as well as in patients with certain disorders such as antibiotic-associated diarrhea, inflammatory bowel disease, irritable bowel syndrome, obesity, allergies, and regressive autism. It has been suggested that the bifidogenic effects of ITF and AXOS are the result of strain-specific yet complementary carbohydrate degradation mechanisms within cooperating bifidobacterial consortia. Except for a bifidogenic effect, ITF and AXOS also have shown to cause a butyrogenic effect in the human colon, i.e., an enhancement of colon butyrate production. Butyrate is an essential metabolite in the human colon, as it is the preferred energy source for the colon epithelial cells, contributes to the maintenance of the gut barrier functions, and has immunomodulatory and anti-inflammatory properties. It has been shown that the butyrogenic effects of ITF and AXOS are the result of cross-feeding interactions between bifidobacteria and butyrate-producing colon bacteria, such as Faecalibacterium prausnitzii (clostridial cluster IV) and Anaerostipes, Eubacterium, and Roseburia species (clostridial cluster XIVa). These kinds of interactions possibly favor the co-existence of bifidobacterial strains with other bifidobacteria and with butyrate-producing colon bacteria in the human colon.
Introduction
Whereas, the human gut microbiota has been studied in the past mainly in the context of infectious diseases, it is known today that this enormous number of microorganisms has an indispensable role in the normal development and functioning of the human body (O'Hara and Shanahan, 2006; Sommer and Bäckhed, 2013). Within the adult gastrointestinal tract, the colon contains the most dense (>1011 bacteria per mL of luminal content) and metabolically active microbiota (Figure 1; Whitman et al., 1998; The Human Microbiome Project Consortium, 2012). The immense number of genes (>100 times the number of genes of the human genome) encoded by this microbiota, expands the host's biochemical and metabolic capabilities substantially (Bäckhed et al., 2005; The Human Microbiome Project Consortium, 2012). Examples of supporting functions of the human gut microbiota are the degradation of otherwise non-digestible food compounds; the transformation of toxic compounds; and the production of essential vitamins, important metabolic end-products, and defending bacteriocins (Sommer and Bäckhed, 2013). Microbial metabolic end-products, which account for one third of the metabolites present in the human blood, play an important role in gut homeostasis and have an impact on host metabolism and health (Wikoff et al., 2009; Hood, 2012; Louis et al., 2014; Sharon et al., 2014; Richards et al., 2016). The short-chain fatty acids (SCFAs) acetate, butyrate, and propionate (typically occurring in a 3:1:1 ratio) are quantitatively (total concentration of 50–150 mM) and metabolically the most important microbial end-products of the human colon fermentation process (Louis et al., 2014), as they display several physiological effects (Table 1).
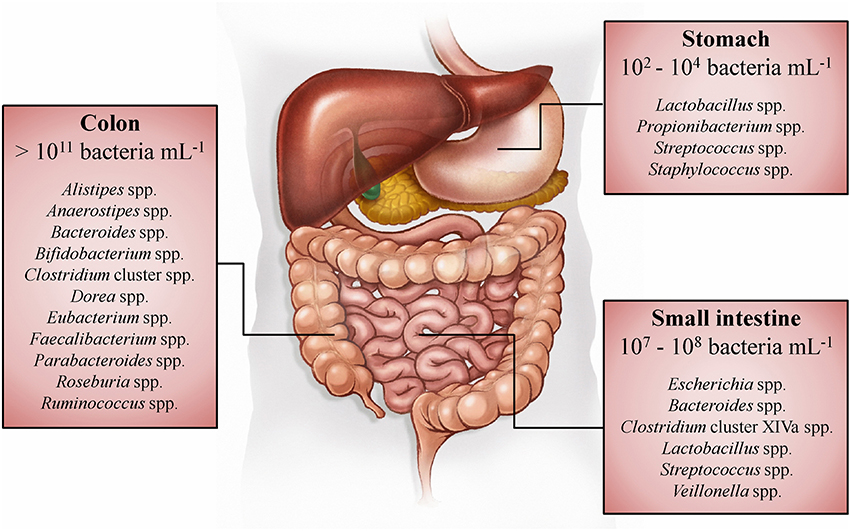
Figure 1. Spatial distribution and concentrations of bacteria along the gastrointestinal tract of humans (Tuohy and Scott, 2015). The dominant genera in the stomach, small intestine, and colon are listed, based on 16S rRNA gene sequence studies (Tap et al., 2009; Zoetendal et al., 2012; Delgado et al., 2013; Walker et al., 2014).
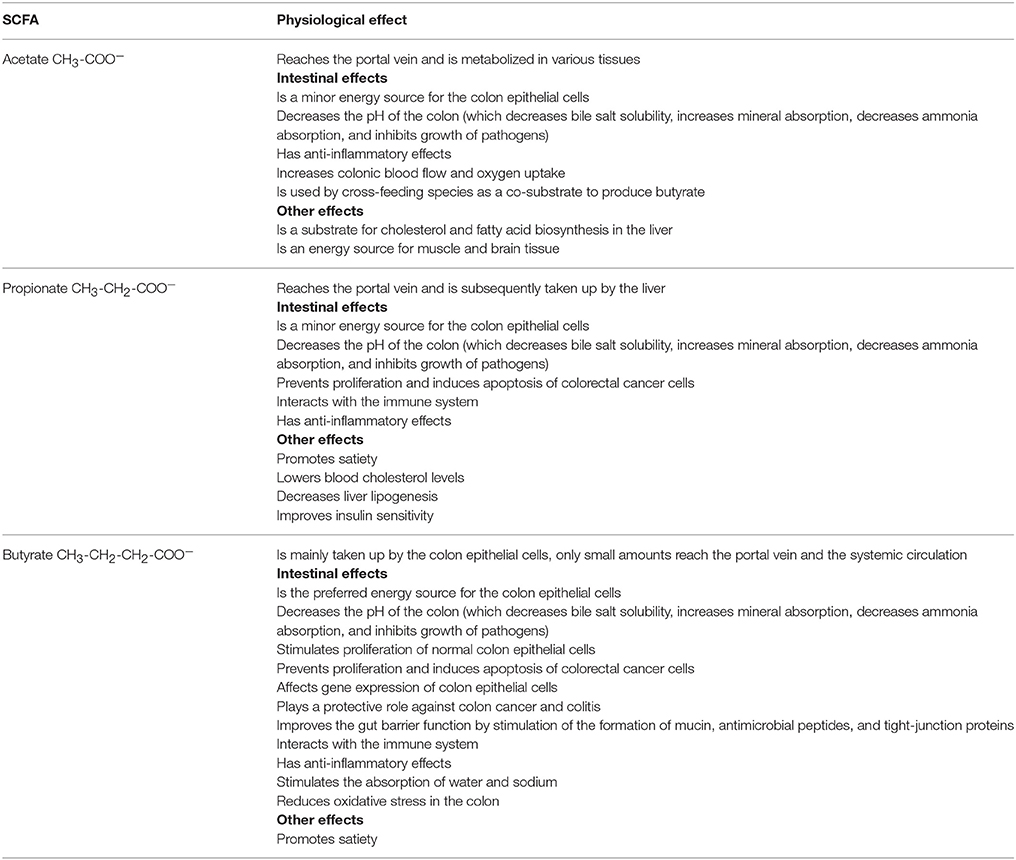
Table 1. Overview of the physiological effects of the short-chain fatty acids (SCFAs) acetate, propionate, and butyrate produced by human colon bacteria (Hamer et al., 2008; Al-Lahham et al., 2010; Havenaar, 2011; Macfarlane and Macfarlane, 2012; Chang et al., 2014; Louis et al., 2014; Tralongo et al., 2014).
Changes in the gut microbiota composition have been associated with disturbed gut barrier functions, increased gut permeability, and increased plasma lipopolysaccharide concentrations (i.e., metabolic endotoxemia), which causes low-grade inflammation that triggers the development of obesity and metabolic syndrome (Cani et al., 2008). Also other disorders, such as inflammatory bowel disease (IBD, encompassing Crohn's disease and ulcerative colitis), irritable bowel syndrome (IBS), colorectal cancer, and allergies have been linked to changes in the gut microbiota composition (de Vos and de Vos, 2012; Le Chatelier et al., 2013). During the last years, even associations have been made between the gut microbiota composition and behavioral disorders, such as depression, anxiety disorder, regressive autism, and schizophrenia (Collins et al., 2012; Braniste et al., 2014; Dinan et al., 2015). However, whereas increasing numbers of animal studies provide evidence for cause-and-effect relationships between shifts in gut microbiota composition and certain disorders (as in the case of obesity; Ridaura et al., 2013), it has not been proven yet for humans whether changes in the gut microbiota composition can cause disorders or that these changes are a consequence of the disorders themselves (de Vos and de Vos, 2012).
In recent years, a few distinct members of the human gut microbiota have received particular attention because of their dedicated metabolism and central role in gut homeostasis and because their loss adversely affects the remaining microorganisms and/or host's health. Bifidobacterium species are one such bacterial species that fulfill important functions within the human colon (Leahy et al., 2005; Rossi and Amaretti, 2011). Decreased numbers of these species in the colon have been associated with several disorders. Moreover, they have shown to interact with other colon bacteria such as butyrate-producing bacteria by cross-feeding interactions. Furthermore, decreased butyrate concentrations and decreased numbers of butyrate producers in the human colon have been associated with disorders. Therefore, this knowledge has encouraged the development of approaches to stimulate the growth and/or activity of bifidobacteria, i.e., the bifidogenic effect, and butyrate-producing colon bacteria, i.e., the butyrogenic effect, in the human colon. The most prevalent approaches to cause bifidogenic and butyrogenic effects involve the consumption of probiotics and prebiotics.
Bifidobacteria and Butyrate-Producing Colon Bacteria
Bifidobacterium Species
General Aspects
Bifidobacteria are Gram-positive, anaerobic, saccharolytic bacteria that belong to the phylum Actinobacteria; they mainly occur in the gastrointestinal tract of mammals, birds, and insects, but are present in sewage, human breast milk, fermented milk, cheeses, and water kefir too (Bottacini et al., 2014; Khodayar-Pardo et al., 2014; Laureys and De Vuyst, 2014; Laureys et al., 2016). A typical bifidobacterial genome has an average size ranging from 2.0 to 2.8 Mb and is characterized by a high guanine-plus-cytosine content, with numerous genes involved in the uptake and breakdown of carbohydrates from both diet and host origin (Ventura et al., 2014). Bifidobacteria are among the first bacteria to colonize the human gastrointestinal tract and reach their highest proportion in the colon (up to 90% of the total colon microbiota in vaginally delivered breast-fed infants) during the first 12 months of life (Tannock, 2010; Turroni et al., 2012). This abundance significantly decreases over time to <5% in adult subjects and decreases even more in the elderly (Arumugam et al., 2011; Duncan and Flint, 2013). At the time of writing, the Bifidobacterium genus comprised 51 species (Euzéby, 1997, 2016; Laureys et al., 2016), among which Bifidobacterium longum, Bifidobacterium animalis, Bifidobacterium adolescentis, Bifidobacterium bifidum, Bifidobacterium catenulatum, Bifidobacterium pseudocatenulatum, Bifidobacterium breve, Bifidobacterium pseudolongum, Bifidobacterium gallicum, Bifidobacterium angulatum, and Bifidobacterium faecale are encountered in the human colon (Turroni et al., 2009; Ventura et al., 2011; Choi et al., 2014). In general, B. bifidum and B. longum are the dominant species in infants, whereas B. adolescentis and B. longum dominate the adult gut microbiota (Turroni et al., 2012). Quantitative PCR analyses of fecal samples of 42 Belgian healthy adults have shown that the fecal microbiota of adults contains between zero and four (with an average of two) different bifidobacterial species, among which B. longum (present in 90% of the adults), B. adolescentis (present in 79% of the adults), and B. catenulatum (present in 38% of the adults) are the most frequently detected species (Ishikawa et al., 2013).
Functional Role in the Colon
From the growing body of scientific evidence associating decreased numbers of bifidobacteria with disorders, it emerges that these species have a disproportionally large impact in the human colon in relation to their relatively low numerical abundance in adults. Hence, a decrease in the relative abundances of Bifidobacterium species in the human colon has been associated with antibiotic-associated diarrhea, IBS, IBD, obesity, allergies, and regressive autism (Di Gioia et al., 2014; Grimm et al., 2014). Examples of functions carried out by bifidobacteria include the production and/or liberation of B vitamins, antioxidants, polyphenols, and conjugated linoleic acids; maturation of the immune system during early life and preservation of immune homeostasis during life; preservation of gut barrier functions and protection against pathogens by producing bacteriocins, decreasing luminal pH by the production of acids, and blocking the adhesion of pathogens to the intestinal mucosa (Leahy et al., 2005; Gorissen et al., 2010, 2012; Rossi and Amaretti, 2011; Gagnon et al., 2015). However, these functions are not characteristic for the Bifidobacterium genus or certain species, but are rather strain-specific. Another important function of the bifidobacterial genus that contributes to gut homeostasis and host health is the production of acetate and lactate during carbohydrate fermentation, organic acids that in turn can be converted into butyrate by other colon bacteria through cross-feeding interactions (Table 1; De Vuyst and Leroy, 2011; De Vuyst et al., 2014; Rivière et al., 2015).
Metabolism
Bifidobacteria display a strictly fermentative metabolism, i.e., they gain energy in the form of ATP by substrate-level phosphorylation during anaerobic carbohydrate breakdown, and play an important role in the human colon with respect to the degradation of carbohydrates that resist digestion and absorption in the upper gastrointestinal tract (Pokusaeva et al., 2011; De Vuyst et al., 2014). Glycoside hydrolases (EC 3.2.1.x) constitute the most important enzyme group that colon bacteria use to degrade poly- and oligosaccharides to fermentable monosaccharides (van den Broek et al., 2008; van den Broek and Voragen, 2008). Compared with the human genome, encoding only 17 glycoside hydrolases for the digestion of food carbohydrates, bifidobacterial genomes possess high numbers of genes encoding these carbohydrases (El Kaoutari et al., 2013). As an example, the genome of B. longum NCC2705 contains 56 genes encoding glycoside hydrolases, one gene encoding a carbohydrate esterase (EC 3.1.1.x), but no genes encoding polysaccharide lyases (EC 4.2.2.x; Schell et al., 2002; Lombard et al., 2014). Bifidobacteria are particularly specialized in efficient uptake of short oligosaccharides into the cell, where they are further degraded to monosaccharides, i.e., they display a preferential oligosaccharide metabolism, providing them a competitive advantage toward other colon bacteria that degrade carbohydrates extracellularly (Van der Meulen et al., 2004, 2006b; Falony et al., 2009a; De Vuyst and Leroy, 2011; De Vuyst et al., 2014). About 5% of the total bifidobacterial gene content is dedicated to carbohydrate internalization, through either ATP-binding cassette transporters, permeases, or proton symporters (Ventura et al., 2009). For example, B. longum NCC2705 contains 15 genes that encode transport systems that could be involved in the transport of oligosaccharides (Schell et al., 2002; Parche et al., 2007). Several laboratory fermentation studies have shown that bifidobacteria can use various non-digestible carbohydrates as energy sources, encompassing plant-derived carbohydrates [such as resistant starch, pectin, inulin, arabinoxylan (AX), cellulose, and their corresponding oligosaccharides] and host-produced carbohydrates (human milk oligosaccharides and mucin), although this ability is strain-dependent too (Klijn et al., 2005; De Vuyst et al., 2014; Rivière et al., 2014; McLaughlin et al., 2015; Selak et al., 2016).
Once internalized into the cytoplasm, hexose monosaccharides (e.g., fructose and glucose) are converted into acetate and lactate by the fructose 6-phosphate phosphoketolase pathway or bifid shunt (De Vuyst et al., 2014). Bifidobacteria will initially cleave, by means of the key enzyme fructose 6-phosphate phosphoketolase, one mole of fructose 6-phosphate into one mole of erythrose 4-phosphate and one mole of acetyl-phosphate (Figure 2A). From erythrose 4-phosphate and an additional mole of fructose 6-phosphate, one mole of ribose 5-phosphate and one mole of xylulose 5-phosphate are formed by the successive action of a transaldolase and a transketolase. Two moles of xylulose 5-phosphate are subsequently converted into two moles of acetyl-phosphate and two moles of glyceraldehyde 3-phosphate by the action of a xylulose 5-phosphate phosphoketolase. These two moles of acetyl-phosphate plus the additional mole of acetyl-phosphate (produced by the fructose 6-phosphate phosphoketolase) are further converted into three moles of acetate by an acetate kinase, which is accompanied by the production of three moles of ATP. The two moles of glyceraldehyde 3-phosphate are oxidized into two moles of pyruvate by enzymes participating in the Embden-Meyerhof-Parnas pathway, which results in an additional production of two moles of ATP. In a last step, pyruvate can be reduced into lactate by means of a lactate dehydrogenase, which is accompanied by NAD+ recycling. Thus, when fermenting hexose monosaccharides, acetate and lactate are produced in a theoretical molar ratio of 1.5 and three moles of ATP are produced. Pentose monosaccharides (e.g., arabinose and xylose) can also be shuttled into the bifid shunt by their conversion into xylulose 5-phosphate (Figure 2A). However, this is not accompanied by the production of an additional mole of acetate (and thus no additional mole of ATP) as in the case of hexose fermentation, leading to a final theoretical molar ratio of acetate to lactate of 1.0 and two moles of ATP (Pokusaeva et al., 2011; De Vuyst et al., 2014). However, these theoretical ratios are rarely found during bifidobacterial carbohydrate fermentation, due to the production of formate, acetate, and ethanol from pyruvate instead of lactate (Figure 2A), which depends on the available energy source and its consumption rate (Palframan et al., 2003; Van der Meulen et al., 2004, 2006a,b; Falony et al., 2009b; De Vuyst et al., 2014). The production of formate from pyruvate by a formate acetyltransferase, at the expense of lactate, can be explained by the need for additional ATP production by means of the concomitant production of acetate when bifidobacteria are grown on complex carbohydrates to improve their fitness, despite their lower growth rate compared with simple carbohydrates. Bifidobacteria are also able to produce ethanol from acetyl-CoA with a bifunctional aldehyde-alcohol dehydrogenase, at the expense of acetate, to enable the continuation of pyruvate production by regenerating NAD+. This shift in metabolism away from lactate production has been found for the degradation of complex carbohydrates such as inulin-type fructans (ITF; oligofructose and inulin; Van der Meulen et al., 2004; Falony et al., 2009b) and arabinoxylan-oligosaccharides (AXOS; Rivière et al., 2014, 2015). Bifidobacteria can also regenerate NAD+ by the production of succinate from oxaloacetate that is in turn formed from phosphoenolpyruvate (Figure 2A; Van der Meulen et al., 2006a).
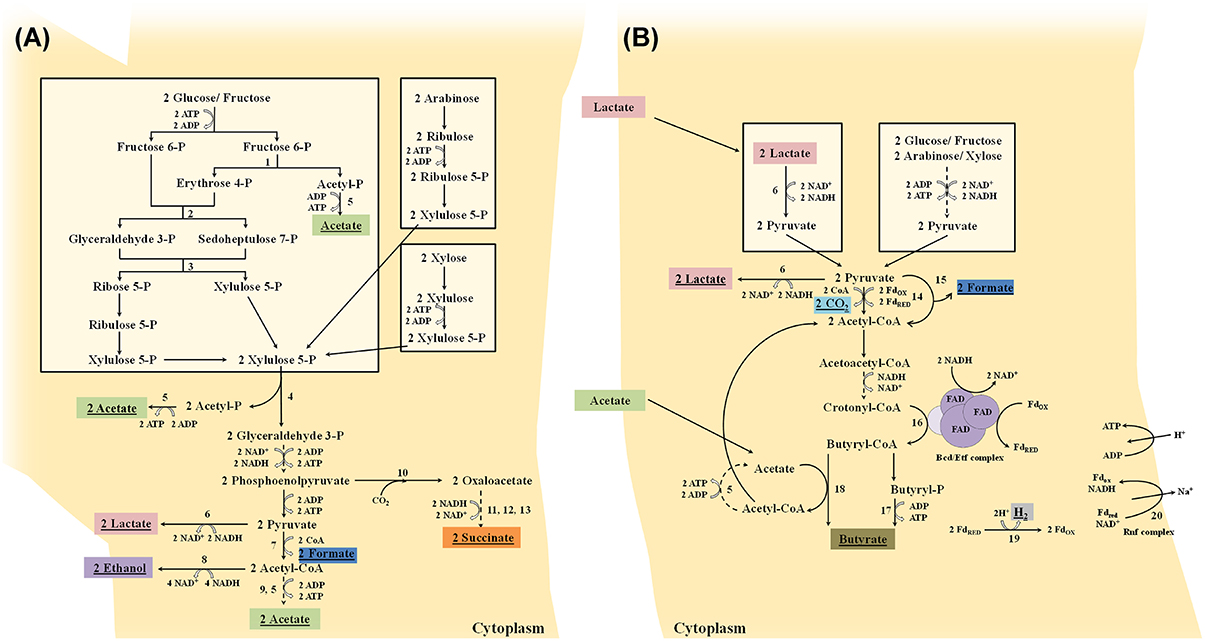
Figure 2. (A) Schematic representation of the fermentation of hexoses (glucose and fructose) and pentoses (arabinose and xylose) by bifidobacteria through the fructose 6-phosphate phosphoketolase pathway or bifid shunt. (B) Schematic representation of the fermentation of hexoses (glucose and fructose) and pentoses (arabinose and xylose) by butyrate-producing colon bacteria through the Embden-Meyerhof-Parnas pathway and pentose-phosphate pathway, respectively, and of lactate. Dashed lines represent different steps. Underlined metabolites are excreted into the extracellular medium. Fdox, oxidized ferredoxin; Fdred, reduced ferredoxin; FAD, flavin adenine dinucleotide; enzymes: 1, fructose 6-phosphate phosphoketolase; 2, transaldolase; 3, transketolase; 4, xylulose 5-phosphate phosphoketolase; 5, acetate kinase; 6, lactate dehydrogenase; 7, formate acetyltransferase; 8, bifunctional aldehyde-alcohol dehydrogenase; 9, phosphotransacetylase; 10, phosphoenolpyruvate carboxylase; 11, malate dehydrogenase; 12, fumarase; 13, succinate dehydrogenase; 14, pyruvate:ferredoxin oxidoreductase; 15, pyruvate-formate lyase; 16, butyryl-CoA dehydrogenase/electron-transferring flavoprotein (Bcd/Etf) complex; 17, butyrate kinase; 18, butyryl-CoA:acetate CoA transferase; 19, ferredoxin hydrogenase; and 20, membrane-bound ferredoxin oxidoreductase (Rnf) complex.
Butyrate-Producing Colon Bacterial Species
General Aspects
Gene-targeted approaches to investigate the butyrate-producing bacterial communities of the human gut microbiota have led to the consideration that butyrate-producing colon bacteria form a functional group rather than a monophyletic group. Most butyrate producers in the human colon belong to the Firmicutes phylum and in particular clostridial clusters IV and XIVa (Louis and Flint, 2009; Van den Abbeele et al., 2013a; Vital et al., 2014). Clostridial clusters IV and XIVa butyrate producers are Gram-positive, highly oxygen-sensitive, strictly anaerobic, saccharolytic bacteria. The two most dominant bacterial species in the human colon are Faecalibacterium prausnitzii (up to 14% of the total fecal gut microbiota, clostridial cluster IV) and Eubacterium rectale (up to 13% of the total fecal gut microbiota, clostridial cluster XIVa), and are expected to have a significant contribution to butyrate production (De Vuyst et al., 2014; Walker et al., 2014). Other important butyrate-producing bacterial species in the human colon are Roseburia spp. (clostridial cluster XIVa, such as Roseburia faecis, Roseburia inulinivorans, Roseburia intestinalis, and Roseburia hominis), Eubacterium spp. (clostridial cluster XIVa, such as Eubacterium hallii), Anaerostipes spp. (clostridial cluster XIVa, such as Anaerostipes butyraticus, Anaerostipes caccae, and Anaerostipes hadrus), and Butyricicoccus pullicaecorum (clostridial cluster IV). Some of these species (such as E. rectale, F. prausnitzii, and R. intestinalis) preferentially colonize the mucus layer, and consequently increase the butyrate bioavailability for colon epithelial cells, whereas other species such as A. caccae mainly occur in the lumen of the colon (El Aidy et al., 2013; Van den Abbeele et al., 2013a). In contrast to bifidobacteria, clostridial clusters IV and XIVa do not directly colonize the colon in high quantities after birth. In the case of F. prausnitzii, it has been shown that fecal numbers in infants younger than 6 months are undetectable, slightly increase between the age of 6 and 24 months, then suddenly increase to reach a peak during late childhood and adolescence, and finally decrease again during adulthood and especially in the elderly (Miquel et al., 2014).
Functional Role in the Colon
Clostridial clusters IV and XIVa have gained a lot of attention during the last years because of their contribution to gut homeostasis, by preserving gut barrier functions and exerting immunomodulatory and anti-inflammatory properties (Velasquez-Manoff, 2015). In addition to the beneficial properties of the butyrate produced (Table 1), F. prausnitzii produces anti-inflammatory peptides blocking nuclear factor NF-κB activation and cytokine IL-8 production in mice, which provide protection against chemically induced colitis (Qiu et al., 2013; Quévrain et al., 2016). Several studies have shown that the abundance of B. pullicaecorum, E. rectale, F. prausnitzii, and/or R. intestinalis is markedly decreased in IBD patients (Morgan et al., 2012; Eeckhaut et al., 2013; Gevers et al., 2014) and that such patients have lower concentrations of butyrate in their feces than healthy individuals (Marchesi et al., 2007; Nemoto et al., 2012). Less butyrate producers were also found in patients with colorectal cancer (Wu et al., 2013). Therefore, methods are being searched to stimulate butyrate-producing human colon bacterial species by diet (prebiotic approach) or by administering these bacteria orally (probiotic approach). In medical applications, pure butyrate by means of tablets or rectal enemas is used as a therapeutic agent for IBD treatment (Geirnaert et al., 2014).
Metabolism
Like bifidobacteria, members of clostridial clusters IV and XIVa carry out a fermentative metabolism and are often able to degrade a wide range of non-digestible carbohydrates in the human colon anaerobically, encompassing resistant starch, ITF, xylo-oligosaccharides (XOS), and AXOS (Falony et al., 2009c; Louis and Flint, 2009; Scott et al., 2014; Rivière et al., 2015; Moens et al., 2016). As an example, the genome of E. rectale ATCC 33656 encodes 52 glycoside hydrolases, encompassing β-fructofuranosidases, α-arabinofuranosidases, β-xylosidases, exo-oligoxylanases, α-amylases, α- and β-glucosidases, α- and β-galactosidases, and cellulases (Lombard et al., 2014). However, inter-genus variations have been found within clostridial clusters IV and XIVa, as not all species and even strains within one species can consume complex carbohydrates to the same extent (Falony et al., 2009c; Scott et al., 2014; Moens et al., 2016). Most of the butyrate-producing colon bacteria use a non-preferential extracellular degradation mechanism for the breakdown of oligo- and polysaccharides, with the release of monosaccharides into the extracellular medium. As illustrated during laboratory batch fermentation experiments, co-cultivation of such butyrate-producing bacteria with bifidobacteria that have a preferential carbohydrate degradation mechanism, can comprise the competitiveness of the butyrate-producing colon bacteria (Falony et al., 2006, 2009c; De Vuyst et al., 2014). For instance, the percentage of oligofructose that was consumed by F. prausnitzii DSM 17677T when co-cultivated with different bifidobacterial strains decreased with an increasing ITF degradation capacity of the latter (Moens et al., 2016).
Once internalized into the cytoplasm, hexoses and pentoses are degraded to pyruvate by the Embden-Meyerhof-Parnas pathway or pentose phosphate pathway, respectively. Like other fermentative bacteria, clostridial clusters IV and XIVa butyrate producers possess several alternative pathways to form different end-metabolites from pyruvate, depending on the bacterial species, carbohydrate source, hydrogen gas pressure, and necessity of redox balancing. Besides butyrate, they can form lactate, formate, hydrogen gas, and carbon dioxide (Figure 2B). Pyruvate can get reduced into lactate by means of a lactate dehydrogenase, which is accompanied by NAD+ recycling (for instance R. inulinivorans and E. rectale; Falony et al., 2009c; Rivière et al., 2015; Moens et al., 2016). The production of butyrate from pyruvate involves the conversion of pyruvate into acetyl-CoA by a pyruvate:ferredoxin oxidoreductase, with the reduction of ferredoxin and production of carbon dioxide (for instance most clostridial clusters IV and XIVa butyrate-producing colon bacteria; Falony et al., 2009c; Moens et al., 2016) and/or by a pyruvate-formate lyase with the formation of formate (for instance F. prausnitzii and E. rectale; Rivière et al., 2015; Moens et al., 2016). Two moles of acetyl-CoA are then converted via a four-step pathway into butyryl-CoA, in which the last step is carried out by a butyryl-CoA dehydrogenase/electron-transferring flavoprotein complex that catalyzes the NADH + H+-dependent reduction of crotonyl-CoA coupled to the reduction of ferredoxin. The final step from butyryl-CoA to butyrate is either catalyzed by a butyrate kinase (after phosphorylation of butyryl-CoA) or a butyryl-CoA:acetate CoA transferase (Falony et al., 2009c; Louis and Flint, 2009; Mahowald et al., 2009; De Vuyst and Leroy, 2011; Moens et al., 2016). Only a few butyrate-producing colon bacteria, encompassing Clostridium butyricum, Coprococcus eutactus, and Coprococcus comes, are known to use a butyrate kinase to produce butyrate (Louis and Flint, 2009; Vital et al., 2014). The butyryl-CoA:acetate CoA transferase step involves the consumption of external acetate (coming from for instance bifidobacterial carbohydrate breakdown through cross-feeding) as a co-substrate, thereby producing acetyl-CoA and butyrate. The acetyl-CoA produced can be converted via acetyl-phosphate into acetate, with the production of ATP, by acetate kinase, or recycled into the four-step pathway mentioned above (Falony et al., 2009c). The reduced ferredoxin can be reoxidized via a ferredoxin hydrogenase, with the concomitant production of H2, and/or via a membrane-bound ferredoxin oxidoreductase (Rnf) complex, without production of H2, but with the generation of a proton-motive force that allows additional ATP production (for instance R. inulinivorans and F. prausnitzii; Falony et al., 2009c; Moens et al., 2016). The production of butyrate thus not only leads to regeneration of NAD+ from NADH + H+ produced in the upper parts of the carbohydrate degradation pathways for ATP production, but also leads to additional ATP production (Falony et al., 2009c; Louis and Flint, 2009; Mahowald et al., 2009; De Vuyst and Leroy, 2011). Some butyrate producers, encompassing A. caccae, A. butyraticus, A. hadrus, and E. hallii, can produce butyrate from lactate instead of carbohydrates (Figure 2B; Duncan et al., 2004; Falony et al., 2006, 2009c; Belenguer et al., 2011; De Vuyst and Leroy, 2011; De Vuyst et al., 2014).
Stimulation of Bifidobacteria and Butyrate-Producing Colon Bacteria
Since decreased numbers of Bifidobacterium species and butyrate-producing bacterial species in the human colon have been reported in patients with diverse disorders and because the SCFAs produced by these species have beneficial effects (Table 1), these bacteria are potential candidates to be stimulated in the colon to prevent and restore a disturbed gut homeostasis. The most prevalent strategies to stimulate bifidobacteria and butyrate-producing colon bacteria in the human colon involve the consumption of probiotics and prebiotics (Scott et al., 2015).
Probiotics
According to the international scientific association for probiotics and prebiotics (ISAPP), probiotics are defined as “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” (Hill et al., 2014). Selected strains of Bifidobacterium species are commonly used probiotics and are added to food supplements and foods (especially dairy products). The oral consumption of bifidobacteria has been associated with beneficial effects for different digestive problems and disorders, encompassing acceleration of the gut transit time; improvement of lactose intolerance; prevention of antibiotic-associated diarrhea and necrotizing enterocolitis (in pre-term infants that usually harbor reduced numbers of bifidobacteria); and alleviation of IBS and IBD symptoms (Leahy et al., 2005; Di Gioia et al., 2014; Tojo et al., 2014; Saez-Lara et al., 2015). Also, evidence continues to emerge that bifidobacteria influence immune responses and hence may enhance resistance to infections and allergies (Di Gioia et al., 2014; Frei et al., 2015). Further, bifidobacteria display anti-inflammatory effects and negatively correlate with metabolic endotoxemia (Everard and Cani, 2013). Moreover, interest is growing to use bifidobacterial strains (such as Bifidobacterium infantis 35624) as psychobiotics, which are “live organisms that, when ingested in adequate amounts, produce a health benefit in patients suffering from psychiatric illness” (Dinan et al., 2013, 2015). The health effects that bifidobacteria exert are of course strain-related; some bifidobacterial strains are effective, whereas others are not. Moreover, the probiotic health benefits are probably not caused by the bifidobacterial strains consumed solely, but are rather the result of interactions with the resident gut microbiota (Cani and Van Hul, 2015; Scott et al., 2015). Indeed, a recent metagenomic and metatranscriptomic study of feces of 12 healthy individuals has shown that the oral administration of the probiotic strain Lactobacillus rhamnosus GG significantly changes the activity of the resident gut microbiota, without influencing the gut microbiota composition itself (Eloe-Fadrosh et al., 2015). Especially genes involved in adhesion, chemotaxis, and/or motility of Bifidobacterium spp., Eubacterium spp., and Roseburia spp. are overexpressed during probiotic consumption, suggesting that the consumption of the probiotic strain promotes interactions between the resident gut microbiota and the host. Nowadays, there is also a growing interest toward the use of other bacterial strains as probiotics, such as Akkermansia muciniphila and butyrate-producing colon bacteria, encompassing B. pullicaecorum, E. rectale, F. prausnitzii, and Roseburia spp. (Marteau, 2013; Geirnaert et al., 2014; Cani and Van Hul, 2015; Scott et al., 2015). For example, the oral administration of B. pullicaecorum 25-3T and F. prausnitzii A2-165 in rodents has shown attenuation of chemically induced colitis (Eeckhaut et al., 2013, 2014; Martín et al., 2015). However, whether these strict anaerobic colon bacteria can survive the harsh industrial production steps and deal with the regulatory hurdles (as these bacteria have no history of safe use) will partly determine their application as probiotics in the future human diet (Figueroa-González et al., 2011; Gosálbez and Ramón, 2015; Kumar et al., 2015; Scott et al., 2015).
Since the implementation of EU legislation on health claims in 2009, no health claims for probiotics in foods have been approved by the European Food Safety Authority (EFSA) neither can the term probiotic further be used as a food label in Europe (Glanville et al., 2015). The only approved health claim is the benefit on lactose digestion when consuming live Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus strains present in yogurt or fermented milk (EFSA, 2010).
In severe cases of a disturbed gut homeostasis, whereby probiotic treatments do not suffice, the gut microbiota can be restored by transplanting the complete fecal microbiota from a healthy donor into a diseased person. However, the ISAPP recommends that fecal microbiota transplantations (FMTs) should not be considered as probiotics, as they are uncharacterized mixtures of strains (Hill et al., 2014). FMTs have shown to be very effective for curing Clostridium difficile infections, although they have ambiguous outcomes for IBD and IBS (Aroniadis and Brandt, 2014; Pamer, 2014). Furthermore, a step-up FMT strategy has been proposed to treat Crohn's disease and ulcerative colitis, which consists of a FMT, followed by additional FMT steps or standard IBD medications depending on the patient's clinical response to the treatment (Cui et al., 2016). Also, it has been shown that patients with metabolic syndrome display improved insulin sensitivity after being treated with fecal microbiota of healthy individuals (Vrieze et al., 2012). These patients possess increased numbers of butyrate-producing colon bacteria and decreased numbers of Gram-negative bacteria after a FMT. Studies are being performed to see whether FMTs can also cure non-gastrointestinal disorders, such as allergies and behavioral disorders (Xu et al., 2015). However, up to now, few fecal transplants have been performed, as the selection of healthy fecal donors requires a thorough examination to avoid the transfer of pathogens and gut microbiota-associated disorders (Kapel et al., 2014). Therefore, new approaches are being searched to transplant well-defined mixtures of bacteria (de Vos, 2013; Van den Abbeele et al., 2013b). However, an additional challenge in selecting an appropriate healthy donor or bacterial synthetic community is that, despite the large amount of information about the composition and diversity of the human gut microbiota, it is difficult (if not impossible) to define a healthy gut microbiota composition, as each healthy individual harbors a unique gut microbiota (de Vos and de Vos, 2012; Faith et al., 2013; Li et al., 2016).
Prebiotics
General
Another strategy to increase bifidobacteria and butyrate-producing bacteria in the human colon is through the consumption of prebiotics, which are defined according to the ISAPP as “a selectively fermented ingredient that results in specific changes in the composition and/or activity of the gastrointestinal microbiota, thus conferring benefit(s) upon host health” (Gibson et al., 2010). To date, all well-known prebiotics are carbohydrates, although other compounds such as, for instance, polyphenols may display prebiotic properties as well (Bindels et al., 2015). Compared with probiotics, prebiotics are more stable and thus can easily be added to foods, such as yogurts, biscuits, breads, cereals, spreads, ice creams, and drinks (Gibson et al., 2010). The criteria for classifying a compound as a prebiotic have been listed as (i) resistance to gastric acidity, hydrolysis by mammalian digestive enzymes, and gastrointestinal absorption; (ii) fermentation by intestinal microbiota; and (iii) selective stimulation of the growth and/or activity of intestinal bacteria associated with health and well-being (Gibson et al., 2004). In the past, the impact of the consumption of prebiotics on the gut microbiota composition was mainly studied regarding species of Bifidobacterium and Lactobacillus (Verbeke, 2014). However, recent community-wide analyses of the gut microbiota show that prebiotics are not that selective as previously assumed, and that they stimulate other bacteria too (Bindels et al., 2015). It has indeed been shown that butyrate-producing colon bacteria, such as E. rectale, F. prausnitzii, and Roseburia spp., can consume prebiotics such as ITF (Falony et al., 2006, 2009c; Rivière et al., 2015; Moens et al., 2016). Also, the consumption of oligofructose changes the relative abundance of 102 bacterial taxa in mice, of which 16 display a more than 10-fold decrease or increase in abundance (Everard et al., 2011). Therefore, Bindels et al. (2015) proposed to define a prebiotic as “a non-digestible compound that, through its metabolization by microorganisms in the gut, modulates composition and/or activity of the gut microbiota, thus conferring a beneficial physiological effect on the host.” Alternatively, the definition of prebiotics has been challenged over time not only according to scientific considerations but also due to its importance for regulators, industry, and consumers (Hutkins et al., 2016). As for probiotics, the term prebiotic cannot be used as a health claim on food products in Europe (Salminen and van Loveren, 2012). Some claims exist for the term fiber (EFSA, 2011a,b, 2015), but not all fibers are prebiotics, whereby the latter are distinguished from the former by the selectivity of their fermentation (Slavin, 2013; Hutkins et al., 2016; Verspreet et al., 2016).
Examples of prebiotic non-digestible carbohydrates that are bifidogenic include poly- and oligosaccharides containing fructose (and a terminal glucose) as in ITF, galactose and glucose (as in galacto-oligosaccharides), glucose (as in isomalto-oligosaccharides), galactose and fructose (as in lactulose), xylose (as in XOS), and arabinose and xylose (as in AX and AXOS) (Roberfroid, 2005; Macfarlane et al., 2008; Broekaert et al., 2011; De Vuyst and Leroy, 2011; De Vuyst et al., 2014). Whereas, in the past the target genus for prebiotic stimulation was Bifidobacterium (Gibson et al., 2010), today new prebiotics are searched to stimulate other beneficial bacterial species in the human colon such as butyrate producers. Of special interest are prebiotics that cause both a bifidogenic effect and a butyrogenic effect. ITF, AX, and AXOS are such prebiotics that stimulate both bifidobacteria and the production of butyrate (Falony et al., 2006, 2009b,c; De Vuyst and Leroy, 2011; De Vuyst et al., 2014; Rivière et al., 2015).
ITF As an Example of Well-Known Prebiotics
Inulin naturally occurs in fruits and plants such as chicory roots, wheat, onion, banana, garlic, and leek, but is generally extracted from chicory roots on an industrial scale (Roberfroid, 2007). Inulin consists of a linear backbone of β-(2 → 1)-linked fructose monomers with a degree of polymerization (DP) between 2 and 65 (average DP of 10), which is often linked to a terminal glucose monomer by an α-(1 → 2)-glycosidic bond (Figures 3A,B). Oligofructose is derived from native inulin by partial enzymatic hydrolysis with an inulinase and has a DP that varies between 2 and 8 (average DP of 4). Given the relative simple structures of ITF, only few bacterial enzymes are required for their degradation in the human colon, encompassing enzymes belonging to the β-fructofuranosidase (EC 3.2.1.26) superfamily that cleave terminal fructose residues from the non-reducing ends of the fructose polymers (Figure 3B; Scott et al., 2011). Several β-fructofuranosidases have been isolated and characterized in colon bacteria, for instance in Bifidobacterium species (Warchol et al., 2002; Ehrmann et al., 2003; Omori et al., 2010; Jedrzejczak-Krzepkowska et al., 2011) and R. inulinivorans (Scott et al., 2011). Examples of beneficial effects of the consumption of ITF include increased stool frequency, increased colonic absorption of dietary minerals (calcium and magnesium), decreased proteolytic activity, and increased secretion of satiety hormones (Schaafsma and Slavin, 2015).
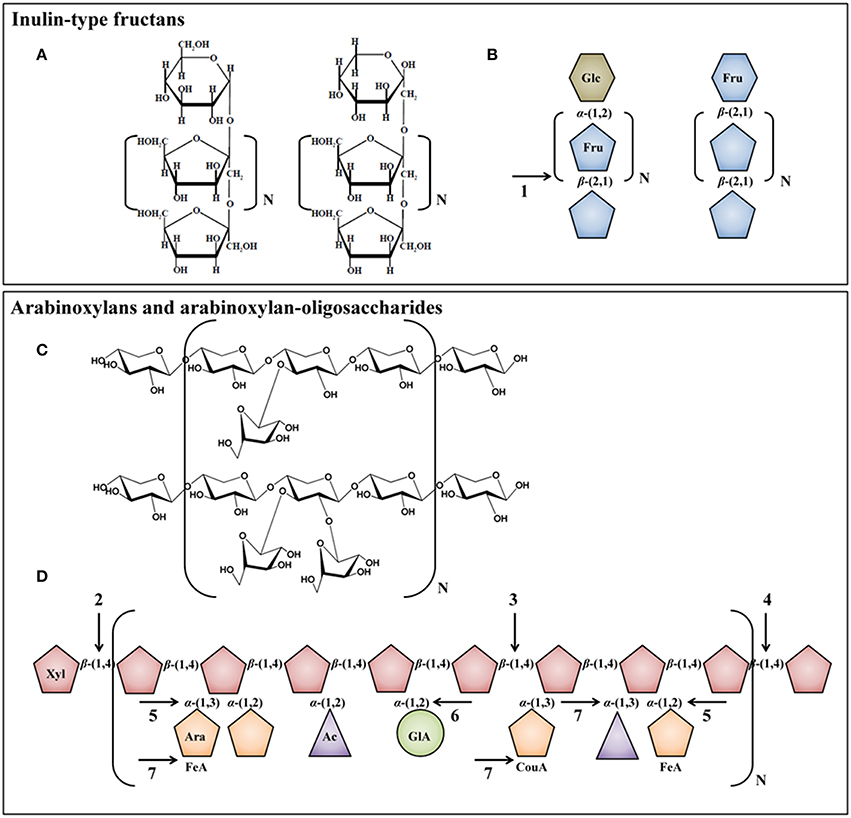
Figure 3. Chemical structures [(A) and (C)] and schematic representations [(B) and (D)] of ITF, AX, and AXOS molecules. Glc, glucose; Fru, fructose; Xyl, xylose; Ara, arabinose; FeA, ferulic acid; Ac, acetyl group; GlA, glucuronic acid; CouA, p-coumaric acid. Arrows indicate possible hydrolysis of the carbohydrates by bacterial enzymes present in the human colon: 1, β-fructofuranosidase; 2, β-xylosidase; 3, β-endoxylanase; 4, exo-oligoxylanase; 5, α-arabinofuranosidase; 6, α-glucuronidase; and 7, esterase.
ITF belong to the most studied prebiotics and their bifidogenic and butyrogenic effects have been well established in various studies (De Vuyst and Leroy, 2011; De Vuyst et al., 2014). For instance, it has been shown that not all bifidobacterial strains benefit in the same way from the presence of ITF in the human colon. A comparative statistical study of 18 bifidobacterial strains, belonging to 10 different species and coming from different donors and origins, has shown the existence of four different clusters of strains differing in their mechanisms and capabilities to degrade ITF (Falony et al., 2009b). Some strains only consume fructose (cluster A), whereas others consume both fructose and oligofructose, mainly short oligosaccharides (DP up to seven) after import into the cell, i.e., they display a preferential metabolism (cluster B). Certain strains degrade both oligofructose and inulin (short chain length fractions only) extracellularly, accompanied with the release of fructose into the extracellular medium, i.e., they display a non-preferential metabolism (clusters C and D). A recent study of 190 bifidobacterial strains isolated from different donors and colon regions has shown that these ITF degradation fingerprints are not correlated with the region in the intestine, suggesting that the degradation of ITF is uniform along the human intestine (Selak et al., 2016). Yet, intra-species variability in ITF degradation capacity indicates strain-specific variations. Moreover, within one colon region bifidobacterial strains with different ITF degradation mechanisms occur, which suggests cooperation for the degradation of ITF in the colon, with opportunities for cross-feeding on strain and/or species level. Similar cross-feeding between bifidobacterial strains with complementary degradation mechanisms has also been shown for starch, xylan, and mucin glycoproteins (Egan et al., 2014; Turroni et al., 2015). Also, it has been shown that the consumption of ITF, the bifidogenic effect, and the butyrogenic effect are linked to each other, because of cross-feeding interactions between bifidobacteria and butyrate-producing colon bacteria (Figure 4; Belenguer et al., 2006; Falony et al., 2006, 2009c; Moens et al., 2016). As an end-metabolite of the bifid shunt and a co-substrate for the production of butyrate (Section AX and AXOS as an Example of Interesting Prebiotics), acetate plays a key role in cross-feeding interactions between bifidobacteria and butyrate-producing colon bacteria in the human colon. In a first type of cross-feeding, both the bifidobacterial and butyrate-producing strains consume ITF (Figure 4). The consumption of ITF by bifidobacteria provides butyrate-producing colon bacteria with exogenous acetate that is used as a co-substrate to produce butyrate by growing on ITF simultaneously [which is, for instance, the case for R. intestinalis DSM 14610 (Falony et al., 2006), R. inulinivorans DSM 16841 (Falony et al., 2009c), and F. prausnitzii DSM 17677T (Moens et al., 2016)]. However, such cross-feeding interactions can be either a pure commensal or beneficial relationship between these bacteria or can be dominated by competition, depending on the ITF degradation capacities of the bifidobacterial strains involved (Moens et al., 2016). A second type of cross-feeding takes place between bifidobacteria that consume ITF, and concomitantly produce acetate, and acetate-consuming butyrate-producing colon bacteria that are not able to degrade ITF (Figure 4). Instead of ITF, the latter bacteria consume carbohydrate breakdown products (short-chain oligosaccharides) liberated by the bifidobacterial strain (which is, for instance, the case for R. hominis DSM 16839; Belenguer et al., 2006) or lactate (for instance E. hallii DSM 17630; Belenguer et al., 2006, and A. caccae DSM 14662; Falony et al., 2006).
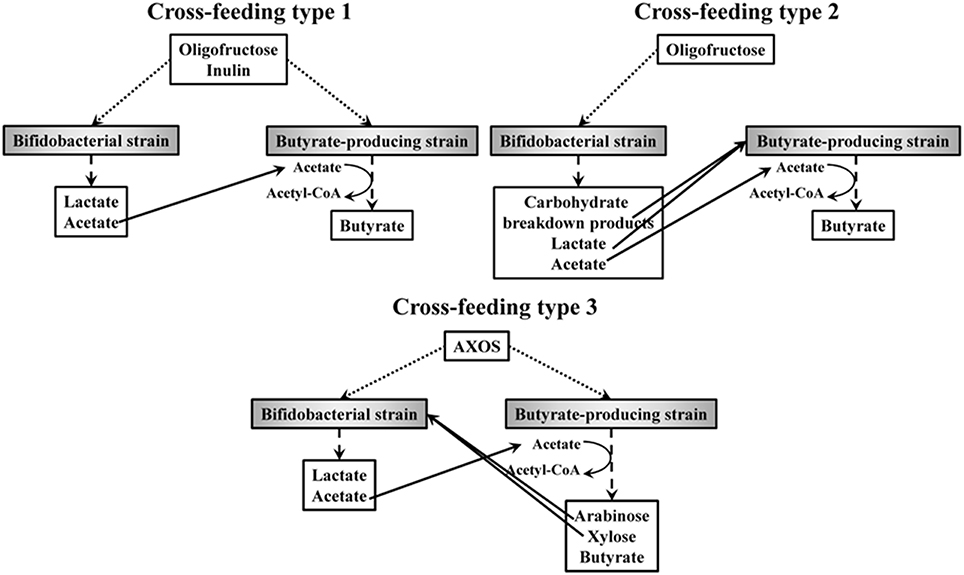
Figure 4. Different types of cross-feeding that can take place between Bifidobacterium spp. and species of butyrate-producing colon bacteria in the human colon. Arrows indicate consumption of oligofructose, inulin, and AXOS (…..), production of carbohydrate breakdown products and/or metabolic end-products (- - -), and cross-feeding interactions between the bifidobacterial and butyrate-producing strains (—).
AX and AXOS As an Example of Interesting Prebiotics
Physiological effects
Growing interest is devoted to complex non-digestible carbohydrates that ferment slowly and thereby cause bifidogenic and butyrogenic effects along the entire length of the human colon. AX and AXOS, as a broad class of heteropolysaccharides and -oligosaccharides with complex varying structures (Figures 3C,D), belong to these slow-fermenting carbohydrates and hence are able to decrease the production of bacterial toxic metabolites originating from protein and lipid metabolism in the distal colon (Section Bifidobacterium Species; Van Craeyveld et al., 2008; Grootaert et al., 2009; Sanchez et al., 2009; Neyrinck et al., 2011). This is to be explained by a stimulation of saccharolytic activities, an increase in SCFA production, and a lowering of the luminal pH in the distal part of the colon, where carbohydrates are rare and proteolytic bacteria, such as Bacteroides spp., are otherwise favored (Duncan et al., 2009). Examples of additional potential benefits of the consumption of AX and AXOS for human health include improved mineral (calcium and magnesium) absorption; increased stool frequency and improved stool consistency; reduced post-prandial glycemic response; reduced blood cholesterol levels; and increased antioxidant capacity (Grootaert et al., 2007; Broekaert et al., 2011; Damen et al., 2011; Mendis and Simsek, 2013). Moreover, the consumption of AXOS, with the increase of bifidobacterial numbers as a result, may help to restore gut barrier functions and cure metabolic endotoxemia in mice (Neyrinck et al., 2012).
Occurrence, structural properties, and degradation
AX naturally occur in the endosperm and bran (pericarp, testa, and aleuron layer) of cereal grains such as wheat, rye, rice, barley, oat, and sorghum, but in varying quantities, depending on the cereal species and the location within the cereal kernel (Izydorczyk and Biliaderis, 2006). For instance, the endosperm of wheat kernels contains ca. 2% of AX, whereas the pericarp contains ca. 38% of AX (Benamrouche et al., 2002; Maes and Delcour, 2002). AX consist of a linear backbone of 1500 to 15,000 β-(1 → 4)-linked xylose monomers, which can randomly be substituted with arabinose monomers on the C-(O)-2 or C-(O)-3 positions (monosubstituted) or on both positions (disubstituted; Figures 3C,D; Izydorczyk and Biliaderis, 1995). Distribution patterns of arabinose substituents on the xylose backbone are not regular for wheat AX; highly branched regions are interlinked by sequences of contiguous non-substituted xylose residues (Gruppen et al., 1993). The number of arabinose substituents bound to the xylose backbone is expressed as the arabinose/xylose ratio (A/X) and depends on the cereal species and the location within the kernel. For instance, in the pericarp, testa, aleuron layer, and endosperm of wheat kernels, different A/Xs are found, namely ca. 1.0, 0.1, 0.4, and 0.5, respectively (Izydorczyk and Biliaderis, 1995; Antoine et al., 2003; Barron et al., 2007). The fermentability of AX and AXOS in the human colon is strongly influenced by the complexity of the AXOS molecules and decreases with increasing DP and increasing A/X (Van Craeyveld et al., 2008; Pollet et al., 2012). Additionally, xylose residues can be esterified with glucuronic acid and acetyl groups, whereas arabinose residues can be esterified with ferulic acid and p-coumaric acid, although in low numbers (Figure 3D; Izydorczyk and Biliaderis, 1995). These esterifications are of health and physicochemical importance, since ferulic acid and p-coumaric acid are antioxidants and potent cross-linking sites for attachment to other AX chains (Bunzel et al., 2001; Ou and Sun, 2014). The presence of feruloylated and diferuloylated arabinose substituents reduces the fermentability of AX and AXOS (Hopkins et al., 2003; Snelders et al., 2014). As cereal whole grains only contain low concentrations of AX (varying between 1.8% of AX in sorghum and 12.1% of AX in rye; Izydorczyk and Biliaderis, 2006), and thus the overall intake of AX is low (especially in modern Western-type diets with high intakes of refined cereal products), AX can be extracted from cereal grains and added to food products in higher concentrations (Broekaert et al., 2009). On an industrial scale, AX are usually extracted from wheat bran that is available in large quantities in Europe (Swennen et al., 2006). AXOS, the hydrolysis products of AX, are formed not only in processed cereal-based food products such as bread and beer (Courtin et al., 2009; Broekaert et al., 2011), but can also be produced on an industrial scale by the enzymatic cleavage of AX with β-endoxylanases (Broekaert et al., 2009, 2011). This results in various substituted molecules (i.e., AXOS) and non-substituted molecules (i.e., XOS), differing in DP and A/X.
Given their complex structures, the degradation of AX and AXOS in the human colon requires the cooperative action of debranching and depolymerizing bacterial carbohydrate-active enzymes, encompassing β-endoxylanases (EC 3.2.1.8) that cleave AX into AXOS and XOS; β-xylosidases (EC 3.2.1.37) that cleave terminal xylose residues from the non-reducing ends of the xylose backbones; exo-oligoxylanases (EC 3.2.1.156) that release terminal xylose residues from the reducing ends of the xylose backbones; and α-arabinofuranosidases (EC 3.2.1.55) that remove arabinose substituents from the xylose backbones (Figure 3D). Additional enzymes are needed to cleave glucuronic acid [i.e., α-glucuronidase (EC 3.2.1.139)], ferulic acid [i.e., ferulic acid esterase (EC 3.1.1.73)], acetyl groups [i.e., acetyl xylan esterase (EC 3.1.1.72)], and p-coumaric acid [i.e., p-coumaric acid esterase (EC 3.1.1.-)] from AXOS (Figure 3D; Dodd and Cann, 2009; Lagaert et al., 2014).
To date, AX and AXOS fall under the definition of dietary fiber (European Commission, 2008; Snelders et al., 2014) but are not considered as prebiotics by the EFSA, although they meet the three criteria of prebiotics (see above; Broekaert et al., 2011). AX and AXOS are neither digested nor absorbed in the upper gastrointestinal tract and reach the human colon intact, where they are fermented by the resident colon bacteria and cause bifidogenic and butyrogenic effects (Table 2). However, as is also the case for other prebiotics, the selective stimulation criterion can be questioned. Several in vivo and in vitro studies have shown that AX and AXOS stimulate, besides bifidobacteria and butyrate-producing colon bacteria, other saccharolytic colon bacteria too, such as Bacteroides spp. and Lactobacillus spp. (Table 2). Moreover, a propionogenic effect is supposed to occur. A few studies have shown that AX and AXOS especially stimulate the production of propionate (Table 2; Hopkins et al., 2003; Van den Abbeele et al., 2011; Pollet et al., 2012). For instance, the mucin-consuming propionate-producing A. muciniphila is stimulated in the colon of humanized rats fed with long-chain AX (Table 2; Van den Abbeele et al., 2011). Whether this is a direct or indirect effect is not known yet.
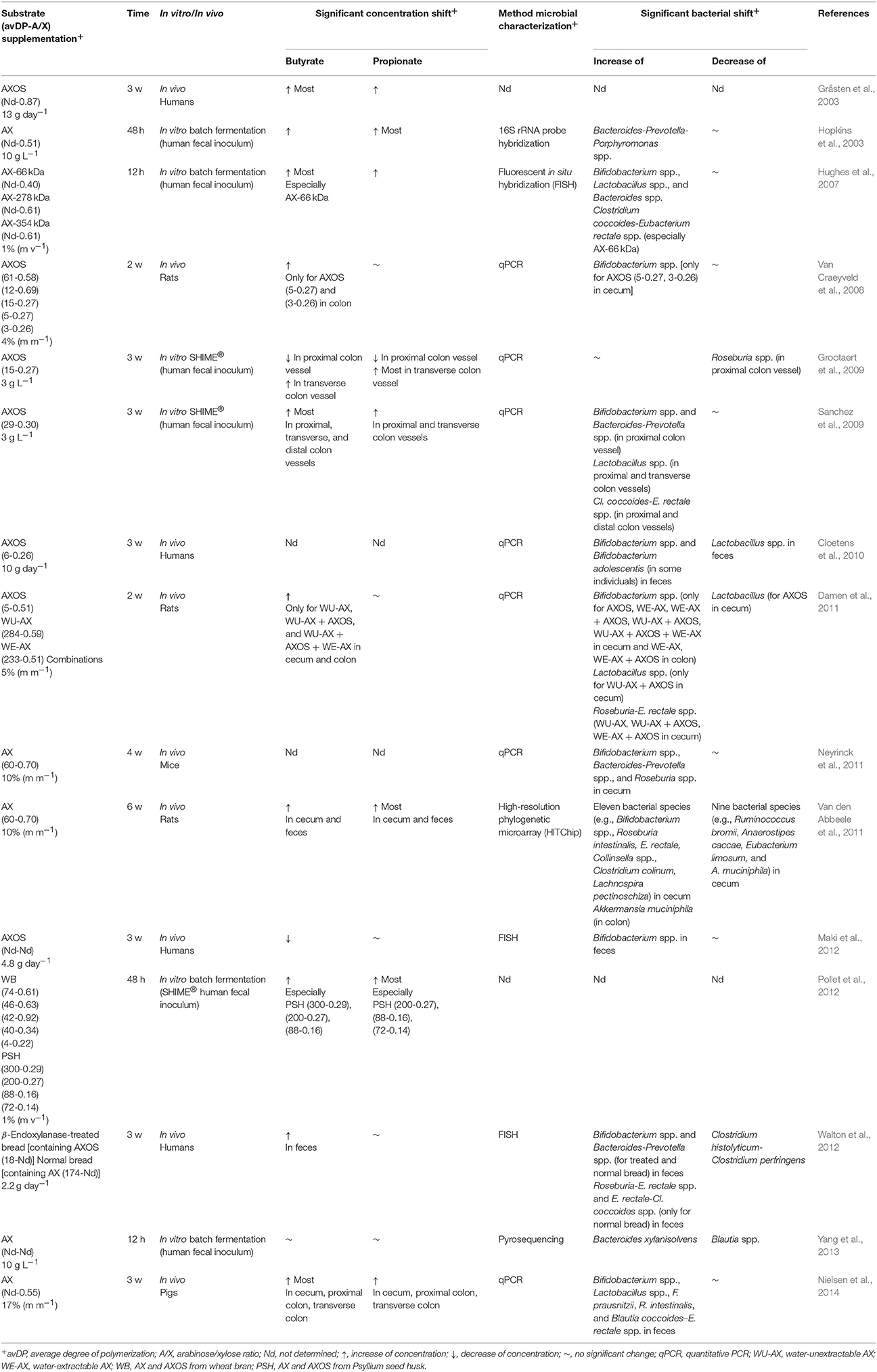
Table 2. Overview of in vitro and in vivo studies of AX and AXOS.
Bifidogenic effects of AX and AXOS
Several in vivo studies (in rodents, pigs, and humans) and in vitro studies [during batch and simulator of human intestinal microbial ecosystem (SHIME®) fermentations with fecal slurries] have shown that AX and AXOS are bifidogenic (Table 2). An in vivo study with rats has shown that the bifidogenic effect is only caused by AXOS with low average DPs ≤ 5 and A/Xs ≤ 0.27 (Van Craeyveld et al., 2008), whereas other rodent studies have found a stimulation of bifidobacteria by AX and AXOS with high average DPs up to 284 and A/Xs up to 0.70 (Table 2; Damen et al., 2011; Neyrinck et al., 2011; Van den Abbeele et al., 2011). In the latter study, a 60-fold increase of bifidobacteria in the cecum of rats has been found, caused by the consumption of long-chain AX (average DP of 60, A/X of 0.70; Van den Abbeele et al., 2011). Apart from in vitro and animal experiments, human studies have revealed a bifidogenic effect caused by a daily intake of 10 g of AXOS per day (Cloetens et al., 2010), 5.5 g of AXOS per day (Maki et al., 2012), and 2.2 g of AX and AXOS per day (Walton et al., 2012; Table 2). However, until recently, many fundamental questions remain unanswered. For instance, how can the low numerical abundant bifidobacteria (<5%) compete with other, more abundant, saccharolytic bacteria in the human colon for AX and AXOS? Do bifidobacteria have a preference for certain AX and AXOS molecules? Are all bifidobacterial strains in the human colon stimulated by AX and AXOS? To answer these questions, a detailed knowledge of the carbohydrate-hydrolyzing capacity of bifidobacteria was missing. Indeed, in the past, studies of the degradation of AX and AXOS through mono-culture fermentations with bifidobacterial strains were restricted to monitoring of bacterial growth, pH, and SCFA production (Van Laere et al., 2000; Crittenden et al., 2002), or fermentations of purified short-chain AXOS standards were performed (Pastell et al., 2009) without revealing the complete fermentation capacity of bifidobacteria. Recently, the mechanistic variations in AXOS degradation by 36 bifidobacterial strains from different donors and origins have been investigated (Rivière et al., 2014). The results show that not all bifidobacterial strains are stimulated by AXOS to the same extent. AXOS degradation by bifidobacteria is complex and involves the consumption of arabinose substituents, whether or not followed by the consumption of the xylose backbones of AXOS, either up to xylotetraose or longer and either intracellularly or extracellularly. Several bifidobacterial strains use the arabinose substituents of AXOS solely, whereas others first consume the arabinose substituents and later import the xylose backbones (up to xylotetraose) into the cell. This extracellular arabinose substituent-oriented metabolism of bifidobacteria has been linked to the presence of genes encoding extracellular cell-associated α-arabinofuranosidases (Lagaert et al., 2010, 2014; Rivière et al., 2014). The majority of the bifidobacterial strains cannot use xylose backbones longer than xylotetraose, i.e., they display a preferential metabolism, except for one strain among the 36 tested ones, B. catenulatum LMG 11043T, that also uses longer xylose backbones, i.e., they display a non-preferential metabolism (Rivière et al., 2014). This could explain why the bifidogenic effect is strongly influenced by the complexity of the AXOS molecules and decreases with increasing DP (Table 2; Van Craeyveld et al., 2008). A multivariate data analysis of the fermentation data of these 36 bifidobacterial strains has revealed five species-independent clusters, representing five different complementary AXOS degradation mechanisms (Rivière et al., 2014). Cluster I strains, albeit not all, consume free arabinose and xylose; cluster II strains have an extracellular arabinose substituent-oriented metabolism; cluster III strains display a preferential metabolism of non-substituted xylose backbones; cluster IV strains combine the degradation mechanisms of clusters II and III; and cluster V strains display a non-preferential AXOS metabolism. The complementary degradation mechanism of bifidobacterial strains and the ability of intracellular and cell-associated degradation of xylose backbones and AXOS, could explain the selective stimulation of bifidobacteria by AXOS in the presence of other saccharolytic colon bacteria in the human colon. Whole-genome sequence annotations have revealed that some bifidobacterial strains contain genes coding for enzymes involved in the debranching of substituents and in the exo-cleavage of the xylose backbones of AX and AXOS (Schell et al., 2002; van den Broek and Voragen, 2008; van den Broek et al., 2008). Indeed, several AXOS-degrading enzymes have been isolated and characterized in bifidobacterial strains, encompassing β-xylosidases in B. adolescentis ATCC 15703 and B. animalis subsp. lactis BB-12; α-arabinofuranosidases in B. adolescentis ATCC 15703, B. adolescentis DSM 20083, B. longum B667, and B. longum NCC2705; and exo-oligoxylanases in B. adolescentis LMG 10502 (Lagaert et al., 2010, 2011, 2014). However, up to now, no β-endoxylanases have been found in the genome of bifidobacteria. The only gene (i.e., BL1543) that was first annotated as a β-endoxylanase in B. longum NCC2705 (Schell et al., 2002) has shown to be an extracellular membrane-associated α-arabinofuranosidase (Lagaert et al., 2010, 2014; Rivière et al., 2014). For the complete utilization of AX, it is likely that most of the Bifidobacterium species require cooperation with β-endoxylanase-producing bacteria, such as Bacteroides and Roseburia species (Chassard et al., 2007; Dodd et al., 2011). For instance, the genome of R. intestinalis L1-82 contains three genes possibly encoding β-endoxylanases (NCBI Resource Coordinators, 2014).
Butyrogenic effects of AX and AXOS
Besides a bifidogenic effect, AX and AXOS have shown to cause a butyrogenic effect (Table 2). In seven of the 13 in vitro and in vivo studies summarized in Table 2, bifidobacteria and butyrate-producing colon bacteria (F. prausnitzii, E. rectale, and Roseburia spp.) are stimulated simultaneously, with a significant increase of butyrate production as a result. As these butyrate-producing colon bacteria are present in high numbers in the colon, a rise in butyrate concentration does not come as a surprise (De Vuyst et al., 2014). In contrast to bifidobacteria, much less is known about the genetic AX- and AXOS-degrading potential of species of butyrate-producing colon bacteria. In silico analysis of the genome sequence of, for instance, E. rectale ATCC 33656 has shown that there are five genes possibly encoding AXOS-degrading enzymes (exo-oligoxylanase, bifunctional β-xylosidase/α-arabinofuranosidase, β-xylosidase, and two α-arabinofuranosidases; Rivière et al., 2015).
In contrast to ITF, the link between the consumption of AXOS, the bifidogenic effect, and the butyrogenic effect has been assessed only recently (Rivière et al., 2015). It has been shown that a third type of cross-feeding can take place in the presence of AXOS (Figure 4), for instance in the case of B. longum NCC2705 (an arabinose substituent degrader of AXOS) and E. rectale ATCC 33656 (a complete AXOS degrader). Both strains consume AXOS (as in cross-feeding type 1), but the bifidobacterial strain is additionally stimulated by consuming the monosaccharides released by the extracellular degradation of AXOS by the E. rectale strain, leading to cross-feeding interactions that are mutually beneficial (Figure 4). It is likely that these kinds of cross-feeding interactions between bifidobacteria and butyrate-producing colon bacteria, caused by prebiotic consumption, will take place in vivo in the human colon (Boets et al., 2013). However, the presence of other bacterial strains, with their own mechanisms of carbohydrate degradation (preferential vs. non-preferential) and own cross-feeding interactions within and between species and genera (Figure 4), complicate the attempts to fully understand the bifidogenic and butyrogenic effects of AX and AXOS in the human colon. Furthermore, the inter-individual variations in bacterial composition make it even more intricate to predict the effects of prebiotic consumption in the colon.
Conclusions
Human gut microbiota research has grown tremendously over the last years in terms of technology development and implications for human health. For instance, it has been shown that certain key bacteria within the colon, such as bifidobacteria and butyrate-producing colon bacteria, are negatively correlated with disorders such as IBD and colorectal cancer. Of the same importance is the progress that is being made into the modulation of the gut microbiota through the use of probiotics, prebiotics, and FMTs to improve human health. Whereas, in the past, the focus was on straightforward increase of bifidobacterial cell concentrations, shifts in interests are currently emphasizing that the stimulation of butyrate-producing bacteria in the human colon is of importance too. The consumption of prebiotic ITF and AXOS seems to be a promising approach to counteract decreased numbers of bifidobacteria and butyrate-producing colon bacteria. The challenge for the upcoming years will however be to first find out whether these changes in gut microbiota composition are the cause or the consequence of a disorder.
Author Contributions
AR acted as the main author. MS, DL, FL, and LD all contributed substantially to the writing and critical revision of the manuscript and approved its final version.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge their financial support of the Research Council of the Vrije Universiteit Brussel (SRP7, IRP2, and IOF342 projects) and the Hercules Foundation (grant UABR09004). MS is the recipient of a PhD fellowship of the Vrije Universiteit Brussel in the framework of a bilateral agreement with the University of Ljubljana. AR was the recipient of a PhD fellowship of the Research Foundation Flanders (FWO-Vlaanderen). DL is the recipient of a PhD fellowship in the framework of an IWT-SBO project (BRANDING IWT130028).
References
Al-Lahham, S. H., Peppelenbosch, M. P., Roelofsen, H., Vonk, R. J., and Venema, K. (2010). Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 1801, 1175–1183. doi: 10.1016/j.bbalip.2010.07.007
Antoine, C., Peyron, S., Mabille, F., Lapierre, C., Bouchet, B., Abecassis, J., et al. (2003). Individual contribution of grain outer layers and their cell wall structure to the mechanical properties of wheat bran. J. Agric. Food Chem. 51, 2026–2033. doi: 10.1021/jf0261598
Aroniadis, O. C., and Brandt, L. J. (2014). Intestinal microbiota and the efficacy of fecal microbiota transplantation in gastrointestinal disease. Gastroenterol. Hepatol. 10, 230–237.
Arumugam, M., Raes, J., Pelletier, E., Le Paslier, D., Yamada, T., Mende, D. R., et al. (2011). Enterotypes of the human gut microbiome. Nature 473, 174–180. doi: 10.1038/nature09944
Bäckhed, F., Ley, R. E., Sonnenburg, J. L., Peterson, D. A., and Gordon, J. I. (2005). Host-bacterial mutualism in the human intestine. Science 307, 1915–1920. doi: 10.1126/science.1104816
Barron, C., Surget, A., and Rouau, X. (2007). Relative amounts of tissues in mature wheat (Triticum aestivum L.) grain and their carbohydrate and phenolic acid composition. J. Cereal Sci. 45, 88–96. doi: 10.1016/j.jcs.2006.07.004
Belenguer, A., Duncan, S. H., Calder, A. G., Holtrop, G., Louis, P., Lobley, G. E., et al. (2006). Two routes of metabolic cross-feeding between Bifidobacterium adolescentis and butyrate-producing anaerobes from the human gut. Appl. Environ. Microbiol. 72, 3593–3599. doi: 10.1128/AEM.72.5.3593-3599.2006
Belenguer, A., Holtrop, G., Duncan, S. H., Anderson, S. E., Calder, A. G., Flint, H. J., et al. (2011). Rates of production and utilization of lactate by microbial communities from the human colon. FEMS Microbiol. Ecol. 77, 107–119. doi: 10.1111/j.1574-6941.2011.01086.x
Benamrouche, S., Crônier, D., Debeire, P., and Chabbert, B. A. (2002). A chemical and histological study on the effect of (1 → 4)-β-endo-xylanase treatment on wheat bran. J. Cereal Sci. 36, 253–260. doi: 10.1006/jcrs.2001.0427
Bindels, L. B., Delzenne, N. M., Cani, P. D., and Walter, J. (2015). Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 12, 303–310. doi: 10.1038/nrgastro.2015.47
Boets, E., Houben, E., Windey, K., De Preter, V., Moens, F., Gomand, S., et al. (2013). “In vivo evaluation of bacterial cross-feeding in the colon using stable isotope techniques: a pilot study,” in Digestive Disease Week, Orlando, FL. Gastroenterology 144.
Bottacini, F., Ventura, M., van Sinderen, D., and O'Connell Motherway, M. (2014). Diversity, ecology and intestinal function of bifidobacteria. Microb. Cell Fact. 13, S4. doi: 10.1186/1475-2859-13-S1-S4
Braniste, V., Al-Asmakh, M., Kowal, C., Anuar, F., Abbaspour, A., Tóth, M., et al. (2014). The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 6, 263ra158. doi: 10.1126/scitranslmed.3009759
Broekaert, W. F., Courtin, C., and Delcour, J. (2009). (Arabino)xylan Oligosaccharide Preparation. WO 2009117790 A2. PCT International Publication.
Broekaert, W. F., Courtin, C. M., Verbeke, K., Van de Wiele, T., Verstraete, W., and Delcour, J. A. (2011). Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 51, 178–194. doi: 10.1080/10408390903044768
Bunzel, M., Ralph, J., Marita, J. M., Hatfield, R. D., and Steinhart, H. (2001). Diferulates as structural components in soluble and insoluble cereal dietary fibre. J. Sci. Food Agric. 81, 653–660. doi: 10.1002/jsfa.861
Cani, P. D., Bibiloni, R., Knauf, C., Waget, A., Neyrinck, A. M., Delzenne, N. M., et al. (2008). Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 57, 1470–1481. doi: 10.2337/db07-1403
Cani, P. D., and Van Hul, M. (2015). Novel opportunities for next-generation probiotics targeting metabolic syndrome. Curr. Opin. Biotechnol. 32, 21–27. doi: 10.1016/j.copbio.2014.10.006
Chang, P. V., Hao, L., Offermanns, S., and Medzhitov, R. (2014). The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. U.S.A. 111, 2247–2252. doi: 10.1073/pnas.1322269111
Chassard, C., Goumy, V., Leclerc, M., Del'homme, C., and Bernalier-Donadille, A. (2007). Characterization of the xylan-degrading microbial community from human faeces. FEMS Microbiol. Ecol. 61, 121–131. doi: 10.1111/j.1574-6941.2007.00314.x
Choi, J. H., Lee, K. M., Lee, M. K., Cha, C. J., and Kim, G. B. (2014). Bifidobacterium faecale sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 64, 3134–3139. doi: 10.1099/ijs.0.063479-0
Cloetens, L., Broekaert, W. F., Delaedt, Y., Ollevier, F., Courtin, C. M., Delcour, J. A., et al. (2010). Tolerance of arabinoxylan-oligosaccharides and their prebiotic activity in healthy subjects: a randomised, placebo-controlled cross-over study. Br. J. Nutr. 103, 703–713. doi: 10.1017/S0007114509992248
Collins, S. M., Surette, M., and Bercik, P. (2012). The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 10, 735–742. doi: 10.1038/nrmicro2876
Courtin, C. M., Broekaert, W. F., Swennen, K., Aerts, G., Van Craeyveld, V., and Delcour, J. A. (2009). Occurrence of arabinoxylo-oligosaccharides and arabinogalactan peptides in beer. J. Am. Soc. Brew. Chem. 67, 112–117. doi: 10.1094/asbcj-2009-0323-01
Crittenden, R., Karppinen, S., Ojanen, S., Tenkanen, M., Fagerstrom, R., Matto, J., et al. (2002). In vitro fermentation of cereal dietary fibre carbohydrates by probiotic and intestinal bacteria. J. Sci. Food Agric. 82, 781–789. doi: 10.1002/jsfa.1095
Cui, B., Li, P., Xu, L., Peng, Z., Xiang, J., He, Z., et al. (2016). Step-up fecal microbiota transplantation (FMT) strategy. Gut Microbes. doi: 10.1080/19490976.2016.1151608. [Epub ahead of print].
Damen, B., Verspreet, J., Pollet, A., Broekaert, W. F., Delcour, J. A., and Courtin, C. M. (2011). Prebiotic effects and intestinal fermentation of cereal arabinoxylans and arabinoxylan oligosaccharides in rats depend strongly on their structural properties and joint presence. Mol. Nutr. Food. Res. 55, 1862–1874. doi: 10.1002/mnfr.201100377
Delgado, S., Cabrera-Rubio, R., Mira, A., Suárez, A., and Mayo, B. (2013). Microbiological survey of the human gastric ecosystem using culturing and pyrosequencing methods. Microb. Ecol. 63, 763–772. doi: 10.1007/s00248-013-0192-5
de Vos, W. M. (2013). Fame and future of faecal transplantations - developing next-generation therapies with synthetic microbiomes. Microb. Biotechnol. 6, 316–325. doi: 10.1111/1751-7915.12047
de Vos, W. M., and de Vos, E. A. (2012). Role of the intestinal microbiome in health and disease: from correlation to causation. Nutr. Rev. 1, S45–S56. doi: 10.1111/j.1753-4887.2012.00505.x
De Vuyst, L., and Leroy, F. (2011). Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifidobacterial competitiveness, butyrate production, and gas production. Int. J. Food Microbiol. 149, 73–80. doi: 10.1016/j.ijfoodmicro.2011.03.003
De Vuyst, L., Moens, F., Selak, M., Rivière, A., and Leroy, F. (2014). Summer meeting 2013: growth and physiology of bifidobacteria. J. Appl. Microbiol. 116, 477–491. doi: 10.1111/jam.12415
Di Gioia, D., Aloisio, I., Mazzola, G., and Biavati, B. (2014). Bifidobacteria: their impact on gut microbiota composition and their applications as probiotics in infants. Appl. Microbiol. Biotechnol. 98, 563–577. doi: 10.1007/s00253-013-5405-9
Dinan, T. G., Stanton, C., and Cryan, J. F. (2013). Psychobiotics: a novel class of psychotropic. Biol. Psychiat. 74, 720–726. doi: 10.1016/j.biopsych.2013.05.001
Dinan, T. G., Stilling, R. M., Stanton, C., and Cryan, J. F. (2015). Collective unconscious: how gut microbes shape human behavior. J. Psychiat. Res. 63, 1–9. doi: 10.1016/j.jpsychires.2015.02.021
Dodd, D., and Cann, I. K. (2009). Enzymatic deconstruction of xylan for biofuel production. Glob. Change Biol. Bioenergy 18, 2–17. doi: 10.1111/j.1757-1707.2009.01004.x
Dodd, D., Mackie, R., and Cann, I. K. (2011). Xylan degradation, a metabolic property shared by rumen and human colonic Bacteroidetes. Mol. Microbiol. 79, 292–304. doi: 10.1111/j.1365-2958.2010.07473.x
Duncan, S. H., and Flint, H. J. (2013). Probiotics and prebiotics and health in ageing populations. Maturitas 75, 44–50. doi: 10.1016/j.maturitas.2013.02.004
Duncan, S. H., Louis, P., and Flint, H. J. (2004). Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 70, 5810–5817. doi: 10.1128/AEM.70.10.5810-5817.2004
Duncan, S. H., Louis, P., Thomson, J. M., and Flint, H. J. (2009). The role of pH in determining the species composition of the human colonic microbiota. Environ. Microbiol. 11, 2112–2122. doi: 10.1111/j.1462-2920.2009.01931.x
Eeckhaut, V., Ducatelle, R., Sas, B., Vermeire, S., and Van Immerseel, F. (2014). Progress towards butyrate-producing pharmabiotics: Butyricicoccus pullicaecorum capsule and efficacy in TNBS models in comparison with therapeutics. Gut 63, 367. doi: 10.1136/gutjnl-2013-305293
Eeckhaut, V., Machiels, K., Perrier, C., Romero, C., Maes, S., Flahou, B., et al. (2013). Butyricicoccus pullicaecorum in inflammatory bowel disease. Gut 62, 1745–1752. doi: 10.1136/gutjnl-2012-303611
EFSA (2010). Scientific opinion on the substantiation of health claims related to live yoghurt cultures and improved lactose digestion (ID 1143, 2976), pursuant to Article 13 (1) of regulation (EC) No 1924/20061. EFSA J. 8, 1763. doi: 10.2903/j.efsa.2010.1763
EFSA (2011a). Scientific opinion on the substantiation of health claims related to resistant starch and reduction of post-prandial glycaemic responses (ID 681), “digestive health benefits” (ID 682) and “favours a normal colon metabolism” (ID 783) pursuant to Article 13 (1) of Regulation (EC) No 1924/2006. EFSA J. 9, 2024. doi: 10.2903/j.efsa.2011.2024
EFSA (2011b). Scientific opinion on the substantiation of health claims related to arabinoxylan produced from wheat endosperm and reduction of post-prandial glycaemic responses (ID 830) pursuant to Article 13 (1) of Regulation (EC) No 1924/2006. EFSA J. 9, 2205. doi: 10.2903/j.efsa.2011.2205
EFSA (2015). Scientific opinion on the substantiation of a health claim related to “native chicory inulin” and maintenance of normal defecation by increasing stool frequency pursuant to Article 13.5 of Regulation (EC) No 1924/2006. EFSA J. 13, 3951. doi: 10.2903/j.efsa.2015.3951
Egan, M., O'Connell Motherway, M., Kilcoyne, M., Kane, M., Joshi, L., Ventura, M., et al. (2014). Cross-feeding by Bifidobacterium breve UCC2003 during co-cultivation with Bifidobacterium bifidum PRL2010 in a mucin-based medium. BMC Microbiol. 14:282. doi: 10.1186/s12866-014-0282-7
Ehrmann, M. A., Korakli, M., and Vogel, R. F. (2003). Identification of the gene for beta-fructofuranosidase of Bifidobacterium lactis DSM10140T and characterization of the enzyme expressed in Escherichia coli. Curr. Microbiol. 46, 391–397. doi: 10.1007/s00284-002-3908-1
El Aidy, S., Van den Abbeele, P., Van de Wiele, T., Louis, P., and Kleerebezem, M. (2013). Intestinal colonization: how key microbial players become established in this dynamic process. Bioessays 35, 913–923. doi: 10.1002/bies.201300073
El Kaoutari, A., Armougom, F., Gordon, J. I., Raoult, D., and Henrissat, B. (2013). The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Microbiol. 11, 497–504. doi: 10.1038/nrmicro3050
Eloe-Fadrosh, E. A., Brady, A., Crabtree, J., Drabek, E. F., Ma, B., Mahurkar, A., et al. (2015). Functional dynamics of the gut microbiome in elderly people during probiotic consumption. MBio 6:e00231. doi: 10.1128/mBio.00231-15
European Commission (2008). Commission directive 2008/100/EC. Official Journal European Union, p. L 285/9.
Euzéby, J. P. (1997). List of bacterial names with standing in nomenclature: a folder available on the internet. Int. J. Syst. Bacteriol. 47, 590–592. doi: 10.1099/00207713-47-2-590
Euzéby, J. P. (2016). “Bifidobacterium”. List of Prokaryotic Names with Standing in Nomenclature. Available online at: http://www.bacterio.net/bifidobacterium.html (Accessed May 19, 2016).
Everard, A., and Cani, P. D. (2013). Diabetes, obesity and gut microbiota. Best Pract. Res. Clin. Gastroenterol. 27, 73–78. doi: 10.1016/j.bpg.2013.03.007
Everard, A., Lazarevic, V., Derrien, M., Girard, M., Muccioli, G. G., Neyrinck, A. M., et al. (2011). Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 60, 2775–2786. doi: 10.2337/db11-0227
Faith, J. J., Guruge, J. L., Charbonneau, M., Subramanian, S., Seedorf, H., Goodman, A. L., et al. (2013). The long-term stability of the human gut microbiota. Science 341:1237439. doi: 10.1126/science.1237439
Falony, G., Calmeyn, T., Leroy, F., and De Vuyst, L. (2009a). Coculture fermentations of Bifidobacterium species and Bacteroides thetaiotaomicron reveal a mechanistic insight into the prebiotic effect of inulin-type fructans. Appl. Environ. Microbiol. 75, 2312–2319. doi: 10.1128/AEM.02649-08
Falony, G., Lazidou, K., Verschaeren, A., Weckx, S., Maes, D., and De Vuyst, L. (2009b). In vitro kinetic analysis of fermentation of prebiotic inulin-type fructans by Bifidobacterium species reveals four different phenotypes. Appl. Environ. Microbiol. 75, 454–461. doi: 10.1128/AEM.01488-08
Falony, G., Verschaeren, A., De Bruycker, F., De Preter, V., Verbeke, K., Leroy, F., et al. (2009c). In vitro kinetics of prebiotic inulin-type fructan fermentation by butyrate-producing colon bacteria: implementation of online gas chromatography for quantitative analysis of carbon dioxide and hydrogen gas production. Appl. Environ. Microbiol. 75, 5884–5892. doi: 10.1128/AEM.00876-09
Falony, G., Vlachou, A., Verbrugghe, K., and De Vuyst, L. (2006). Cross-feeding between Bifidobacterium longum BB536 and acetate-converting, butyrate-producing colon bacteria during growth on oligofructose. Appl. Environ. Microbiol. 72, 7835–7841. doi: 10.1128/AEM.01296-06
Figueroa-González, I., Quijano, G., Ramírez, G., and Cruz-Guerrero, A. (2011). Probiotics and prebiotics - perspectives and challenges. J. Sci. Food Agric. 91, 1341–1348. doi: 10.1002/jsfa.4367
Frei, R., Akdis, M., and O'Mahony, L. (2015). Prebiotics, probiotics, synbiotics, and the immune system: experimental data and clinical evidence. Curr. Opin. Gastroenterol. 31, 153–158. doi: 10.1097/MOG.0000000000000151
Gagnon, M., Savard, P., Rivière, A., LaPointe, G., and Roy, D. (2015). Bioaccessible antioxidants in milk fermented by Bifidobacterium longum subsp. longum strains. Biomed. Res. Int. 2015:169381. doi: 10.1155/2015/169381
Geirnaert, A., Steyaert, A., Eeckhaut, V., Debruyne, B., Arends, J. B., Van Immerseel, F., et al. (2014). Butyricicoccus pullicaecorum, a butyrate producer with probiotic potential, is intrinsically tolerant to stomach and small intestine conditions. Anaerobe 30, 70–74. doi: 10.1016/j.anaerobe.2014.08.010
Gevers, D., Kugathasan, S., Denson, L. A., Vázquez-Baeza, Y., Van Treuren, W., Ren, B., et al. (2014). The treatment-naive microbiome in new-onset Crohn's disease. Cell Host Microbe 15, 382–392. doi: 10.1016/j.chom.2014.02.005
Gibson, G. R., Probert, H. M., Loo, J. V., Rastall, R. A., and Roberfroid, M. B. (2004). Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr. Res. Rev. 17, 259–275. doi: 10.1079/NRR200479
Gibson, G. R., Scott, K. P., Rastall, R. A., Tuohy, K. M., Hotchkiss, A., Dubert-Ferrandon, A., et al. (2010). Dietary prebiotics: current status and new definition. Food Sci. Technol. Bull. 7, 1–19. doi: 10.1616/1476-2137.15880
Glanville, J., King, S., Guarner, F., Hill, C., and Sanders, M. E. (2015). A review of the systematic review process and its applicability for use in evaluating evidence for health claims on probiotic foods in the European Union. Nutr. J. 14, 16. doi: 10.1186/s12937-015-0004-5
Gorissen, L., De Vuyst, L., Raes, K., De Smet, S., and Leroy, F. (2012). Conjugated linoleic and linolenic acid production kinetics by bifidobacteria differ among strains. Int. J. Food Microbiol. 155, 234–240. doi: 10.1016/j.ijfoodmicro.2012.02.012
Gorissen, L., Raes, K., Weckx, S., Dannenberger, D., Leroy, F., De Vuyst, L., et al. (2010). Production of conjugated linoleic acid and conjugated linolenic acid isomers by Bifidobacterium species. Appl. Microbiol. Biotechnol. 87, 2257–2266. doi: 10.1007/s00253-010-2713-1
Gosálbez, L., and Ramón, D. (2015). Probiotics in transition: novel strategies. Trends Biotechnol. 33, 195–196. doi: 10.1016/j.tibtech.2015.01.006
Gråsten, S., Liukkonen, K. H., Chrevatidis, A., El-Nezami, H., Poutanen, K., and Mykkänen, H. (2003). Effects of wheat pentosan and inulin on the metabolic activity of fecal microbiota and on bowel function in healthy humans. Nutr. Res. 23, 1503–1514. doi: 10.1016/S0271-5317(03)00164-7
Grimm, V., Westermann, C., and Riedel, C. U. (2014). Bifidobacteria-host interactions - an update on colonisation factors. Biomed. Res. Int. 2014:960826. doi: 10.1155/2014/960826
Grootaert, C., Delcour, J. A., Courtin, C. M., Broekaert, W. F., Verstraete, W., and Van de Wiele, T. (2007). Microbial metabolism and prebiotic potency of arabinoxylan oligosaccharides in the human intestine. Trends Food Sci. Technol. 18, 64–71. doi: 10.1016/j.tifs.2006.08.004
Grootaert, C., Van den Abbeele, P., Marzorati, M., Broekaert, W. F., Courtin, C. M., Delcour, J. A., et al. (2009). Comparison of prebiotic effects of arabinoxylan oligosaccharides and inulin in a simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 69, 231–242. doi: 10.1111/j.1574-6941.2009.00712.x
Gruppen, H., Kormelink, F. J. M., and Voragen, A. G. J. (1993). Enzymic degradation of water-unextractable cell wall material and arabinoxylans from wheat flour. J. Cereal Sci. 18, 129–143. doi: 10.1006/jcrs.1993.1041
Hamer, H. M., Jonkers, D., Venema, K., Vanhoutvin, S., Troost, F. J., and Brummer, R. J. (2008). Review article: the role of butyrate on colonic function. Aliment. Pharmacol. Ther. 27, 104–119. doi: 10.1111/j.1365-2036.2007.03562.x
Havenaar, R. (2011). Intestinal health functions of colonic microbial metabolites: a review. Benef. Microbes 2, 103–114. doi: 10.3920/BM2011.0003
Hill, C., Guarner, F., Reid, G., Gibson, G. R., Merenstein, D. J., Pot, B., et al. (2014). Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514. doi: 10.1038/nrgastro.2014.66
Hopkins, M. J., Englyst, H. N., Macfarlane, S., Furrie, E., Macfarlane, G. T., and McBain, A. J. (2003). Degradation of cross-linked and non-cross-linked arabinoxylans by the intestinal microbiota in children. Appl. Environ. Microbiol. 69, 6354–6360. doi: 10.1128/AEM.69.11.6354-6360.2003
Hughes, S. A., Shewry, P. R., Li, L., Gibson, G. R., Sanz, M. L., and Rastall, R. A. (2007). In vitro fermentation by human fecal microflora of wheat arabinoxylans. J. Agric. Food Chem. 55, 4589–4595. doi: 10.1021/jf070293g
Hutkins, R. W., Krumbeck, J. A., Bindels, L. B., Cani, P. D., Fahey, G., Goh, Y. J., et al. (2016). Prebiotics: why definitions matter. Curr. Opin. Biotechnol. 37, 1–7. doi: 10.1016/j.copbio.2015.09.001
Ishikawa, E., Matsuki, T., Kubota, H., Makino, H., Sakai, T., Oishi, K., et al. (2013). Ethnic diversity of gut microbiota: species characterization of Bacteroides fragilis group and genus Bifidobacterium in healthy Belgian adults, and comparison with data from Japanese subjects. J. Biosci. Bioeng. 116, 265–270. doi: 10.1016/j.jbiosc.2013.02.010
Izydorczyk, M. S., and Biliaderis, C. G. (1995). Cereal arabinoxylans: advances in structure and physicochemical properties. Carbohydr. Polym. 28, 33–48. doi: 10.1016/0144-8617(95)00077-1
Izydorczyk, M. S., and Biliaderis, C. G. (2006). “Arabinoxylans: technology and nutritionally functional plant polysaccharides,” in Functional Food Carbohydrates, eds C. G. Biliaderis and M. S. Izydorczyk (Boca Raton, FL: CRC Press), 249–290.
Jedrzejczak-Krzepkowska, M., Tkaczuk, K. L., and Bielecki, S. (2011). Biosynthesis, purification and characterization of β-fructofuranosidase from Bifidobacterium longum KN29.1. Proc. Biochem. 46, 1963–1972. doi: 10.1016/j.procbio.2011.07.005
Kapel, N., Thomas, M., Corcos, O., Mayeur, C., Barbot-Trystram, L., Bouhnik, Y., et al. (2014). Practical implementation of faecal transplantation. Clin. Microbiol. Infec. 20, 1098–1105. doi: 10.1111/1469-0691.12796
Khodayar-Pardo, P., Mira-Pascual, L., Collado, M. C., and Martínez-Costa, C. (2014). Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J. Perinatol. 34, 599–605. doi: 10.1038/jp.2014.47
Klijn, A., Mercenier, A., and Arigoni, F. (2005). Lessons from the genomes of bifidobacteria. FEMS Microbiol. Rev. 29, 491–509. doi: 10.1016/j.fmrre.2005.04.010
Kumar, H., Salminen, S., Verhagen, H., Rowland, I., Heimbach, J., Bañares, S., et al. (2015). Novel probiotics and prebiotics: road to the market. Curr. Opin. Biotechnol. 32, 99–103. doi: 10.1016/j.copbio.2014.11.021
Lagaert, S., Pollet, A., Courtin, C. M., and Volckaert, G. (2014). β-Xylosidases and α-L-arabinofuranosidases: accessory enzymes for arabinoxylan degradation. Biotechnol. Adv. 32, 316–332. doi: 10.1016/j.biotechadv.2013.11.005
Lagaert, S., Pollet, A., Delcour, J. A., Lavigne, R., Courtin, C. M., and Volckaert, G. (2010). Substrate specificity of three recombinant α-L-arabinofuranosidases from Bifidobacterium adolescentis and their divergent action on arabinoxylan and arabinoxylan oligosaccharides. Biochem. Biophys. Res. Commun. 26, 644–650. doi: 10.1016/j.bbrc.2010.10.075
Lagaert, S., Pollet, A., Delcour, J. A., Lavigne, R., Courtin, C. M., and Volckaert, G. (2011). Characterization of two β-xylosidases from Bifidobacterium adolescentis and their contribution to the hydrolysis of prebiotic xylooligosaccharides. Appl. Microbiol. Biotechnol. 92, 1179–1185. doi: 10.1007/s00253-011-3396-y
Laureys, D., Cnockaert, M., De Vuyst, L., and Vandamme, P. (2016). Bifidobacterium aquikefiri sp. nov., isolated from water kefir. Int. J. Syst. Evolut. Microbiol. 66, 1281–1286. doi: 10.1099/ijsem.0.000877
Laureys, D., and De Vuyst, L. (2014). Microbial species diversity, community dynamics, and metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol. 80, 2564–2572. doi: 10.1128/AEM.03978-13
Leahy, S. C., Higgins, D. G., Fitzgerald, G. F., and van Sinderen, D. (2005). Getting better with bifidobacteria. J. Appl. Microbiol. 98, 1303–1315. doi: 10.1111/j.1365-2672.2005.02600.x
Le Chatelier, E., Nielsen, T., Qin, J., Prifti, E., Hildebrand, F., Falony, G., et al. (2013). Richness of human gut microbiome correlates with metabolic markers. Nature 500, 541–546. doi: 10.1038/nature12506
Li, S., Zhu, A., Benes, V., Costea, P. I., Hercog, R., Hildebrand, F., et al. (2016). Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science 352, 586–589. doi: 10.1126/science.aad8852
Lombard, V., Ramulu, H. G., Drula, E., Coutinho, P. M., and Henrissat, B. (2014). The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 42, D490–D495. doi: 10.1093/nar/gkt1178
Louis, P., and Flint, H. J. (2009). Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 294, 1–8. doi: 10.1111/j.1574-6968.2009.01514.x
Louis, P., Hold, G. L., and Flint, H. J. (2014). The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 12, 661–672. doi: 10.1038/nrmicro3344
Macfarlane, G. T., and Macfarlane, S. (2012). Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 95, 50–60. doi: 10.5740/jaoacint.SGE_Macfarlane
Macfarlane, G. T., Steed, H., and Macfarlane, S. (2008). Bacterial metabolism and health-related effects of galacto-oligosaccharides and other prebiotics. J. Appl. Microbiol. 104, 305–344. doi: 10.1111/j.1365-2672.2007.03520.x
Maes, C., and Delcour, J. A. (2002). Structural characterisation of water-extractable and water-unextractable arabinoxylans in wheat bran. J. Cereal Sci. 35, 315–326. doi: 10.1006/jcrs.2001.0439
Mahowald, M. A., Rey, F. E., Seedorf, H., Turnbaugh, P. J., Fulton, R. S., Wollam, A., et al. (2009). Characterizing a model human gut microbiota composed of members of its two dominant bacterial phyla. Proc. Natl. Acad. Sci. U.S.A. 106, 5859–5864. doi: 10.1073/pnas.0901529106
Maki, K. C., Gibson, G. R., Dickmann, R. S., Kendall, C. W., Chen, C. Y., Costabile, A., et al. (2012). Digestive and physiologic effects of a wheat bran extract, arabino-xylan-oligosaccharide, in breakfast cereal. Nutrition 28, 1115–1121. doi: 10.1016/j.nut.2012.02.010
Marchesi, J. R., Holmes, E., Khan, F., Kochhar, S., Scanlan, P., Shanahan, F., et al. (2007). Rapid and noninvasive metabonomic characterization of inflammatory bowel disease. J. Proteome Res. 6, 546–551. doi: 10.1021/pr060470d
Marteau, P. (2013). Butyrate-producing bacteria as pharmabiotics for inflammatory bowel disease. Gut 62, 1673. doi: 10.1136/gutjnl-2012-304240
Martín, R., Miquel, S., Chain, F., Natividad, J. M., Jury, J., Lu, J., et al. (2015). Faecalibacterium prausnitzii prevents physiological damages in a chronic low-grade inflammation murine model. BMC Microbiol. 15:67. doi: 10.1186/s12866-015-0400-1
McLaughlin, H. P., Motherway, M. O., Lakshminarayanan, B., Stanton, C., Paul Ross, R., Brulc, J., et al. (2015). Carbohydrate catabolic diversity of bifidobacteria and lactobacilli of human origin. Int. J. Food Microbiol. 203, 109–121. doi: 10.1016/j.ijfoodmicro.2015.03.008
Mendis, M., and Simsek, S. (2013). Arabinoxylans and human health. Food Hydrocoll. 42, 239–243. doi: 10.1016/j.foodhyd.2013.07.022
Miquel, S., Martín, R., Bridonneau, C., Robert, V., Sokol, H., Bermúdez-Humarán, L. G., et al. (2014). Ecology and metabolism of the beneficial intestinal commensal bacterium Faecalibacterium prausnitzii. Gut Microbes 5, 146–151. doi: 10.4161/gmic.27651
Moens, F., Weckx, S., and De Vuyst, L. (2016). Bifidobacterial inulin-type fructan degradation capacity determines cross-feeding interactions between bifidobacteria and Faecalibacterium prausnitzii. Int. J. Food Microbiol. 231, 76–85 doi: 10.1016/j.ijfoodmicro.2016.05.015
Morgan, X. C., Tickle, T. L., Sokol, H., Gevers, D., Devaney, K. L., Ward, D. V., et al. (2012). Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 13:R79. doi: 10.1186/gb-2012-13-9-r79
NCBI Resource Coordinators (2014). Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 42, D7–D17. doi: 10.1093/nar/gkt1146
Nemoto, H., Kataoka, K., Ishikawa, H., Ikata, K., Arimochi, H., Iwasaki, T., et al. (2012). Reduced diversity and imbalance of fecal microbiota in patients with ulcerative colitis. Dig. Dis. Sci. 57, 2955–2964. doi: 10.1007/s10620-012-2236-y
Neyrinck, A. M., Possemiers, S., Druart, C., van de Wiele, T., De Backer, F., Cani, P. D., et al. (2011). Prebiotic effects of wheat arabinoxylan related to the increase in bifidobacteria, Roseburia and Bacteroides/Prevotella in diet-induced obese mice. PLoS ONE 6:e20944. doi: 10.1371/journal.pone.0020944
Neyrinck, A. M., Van Hée, V. F., Piront, N., De Backer, F., Toussaint, O., Cani, P. D., et al. (2012). Wheat-derived arabinoxylan oligosaccharides with prebiotic effect increase satietogenic gut peptides and reduce metabolic endotoxemia in diet-induced obese mice. Nutr. Diabetes 2, e28. doi: 10.1038/nutd.2011.24
Nielsen, T. S., Lærke, H. N., Theil, P. K., Sørensen, J. F., Saarinen, M., Forssten, S., et al. (2014). Diets high in resistant starch and arabinoxylan modulate digestion processes and SCFA pool size in the large intestine and faecal microbial composition in pigs. Br. J. Nutr. 112, 1837–1849. doi: 10.1017/S000711451400302X
O'Hara, A. M., and Shanahan, F. (2006). The gut flora as a forgotten organ. EMBO Rep. 7, 688–693. doi: 10.1038/sj.embor.7400731
Omori, T., Ueno, K., Muramatsu, K., Kikuchi, M., Onodera, S., and Shiomi, N. (2010). Characterization of recombinant β-fructofuranosidase from Bifidobacterium adolescentis G1. Chem. Centr. J. 4:9. doi: 10.1186/1752-153X-4-9
Ou, J., and Sun, Z. (2014). Feruloylated oligosaccharides: structure, metabolism and function. J. Funct. Foods 7, 90–100. doi: 10.1016/j.jff.2013.09.028
Palframan, R. J., Gibson, G. R., and Rastall, R. A. (2003). Carbohydrate preferences of Bifidobacterium species isolated from the human gut. Curr. Issues Intest. Microbiol. 4, 71–75.
Pamer, E. G. (2014). Fecal microbiota transplantation: effectiveness, complexities, and lingering concerns. Mucosal Immunol. 7, 210–214. doi: 10.1038/mi.2013.117
Parche, S., Amon, J., Jankovic, I., Rezzonico, E., Beleut, M., Barutcu, H., et al. (2007). Sugar transport systems of Bifidobacterium longum NCC2705. J. Mol. Microbiol. Biotechnol. 12, 9–19. doi: 10.1159/000096455
Pastell, H., Westermann, P., Meyer, A. S., Tuomainen, P., and Tenkanen, M. (2009). In vitro fermentation of arabinoxylan-derived carbohydrates by bifidobacteria and mixed faecal microbiota. J. Agric. Food Chem. 57, 8598–8606. doi: 10.1021/jf901397b
Pokusaeva, K., Fitzgerald, G. F., and van Sinderen, D. (2011). Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 6, 285–306. doi: 10.1007/s12263-010-0206-6
Pollet, A., Van Craeyveld, V., Van de Wiele, T., Verstraete, W., Delcour, J. A., and Courtin, C. M. (2012). In vitro fermentation of arabinoxylan oligosaccharides and low molecular mass arabinoxylans with different structural properties from wheat (Triticum aestivum L.) bran and psyllium (Plantago ovata Forsk) seed husk. J. Agric. Food Chem. 60, 946–954. doi: 10.1021/jf203820j
The Human Microbiome Project Consortium (2012). Structure, function and diversity of the healthy human microbiome. Nature 486, 207–214. doi: 10.1038/nature11234
Qiu, X., Zhang, M., Yang, X., Hong, N., and Yu, C. (2013). Faecalibacterium prausnitzii upregulates regulatory T cells and anti-inflammatory cytokines in treating TNBS-induced colitis. J. Crohns Colitis 7, e558–e568. doi: 10.1016/j.crohns.2013.04.002
Quévrain, E., Maubert, M. A., Michon, C., Chain, F., Marquant, R., Tailhades, J., et al. (2016). Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn's disease. Gut 65, 415–425. doi: 10.1136/gutjnl-2014-307649
Richards, L. B., Li, M., van Esch, B. C. A. M., Garssen, J., and Folkerts, G. (2016). The effects of short-chain fatty acids on the cardiovascular system. Pharma Nutr. 4, 68–111. doi: 10.1016/j.phanu.2016.02.001
Ridaura, V. K., Faith, J. J., Rey, F. E., Cheng, J., Duncan, A. E., Kau, A. L., et al. (2013). Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 341:1241214. doi: 10.1126/science.1241214
Rivière, A., Gagnon, M., Weckx, S., Roy, D., and De Vuyst, L. (2015). Mutual cross-feeding interactions between Bifidobacterium longum NCC2705 and Eubacterium rectale ATCC 33656 explain the bifidogenic and butyrogenic effects of arabinoxylan-oligosaccharides. Appl. Environ. Microbiol. 81, 7767–7781. doi: 10.1128/AEM.02089-15
Rivière, A., Moens, F., Selak, M., Maes, D., Weckx, S., and De Vuyst, L. (2014). The ability of bifidobacteria to degrade arabinoxylan oligosaccharide constituents and derived oligosaccharides is strain dependent. Appl. Environ. Microbiol. 80, 204–217. doi: 10.1128/AEM.02853-13
Roberfroid, M. B. (2005). Introducing inulin-type fructans. Br. J. Nutr. 93, S13–S25. doi: 10.1079/bjn20041350
Roberfroid, M. B. (2007). Inulin-type fructans: functional food ingredients. J. Nutr. 137, 2493–2502. doi: 10.1201/9780203504932
Rossi, M., and Amaretti, A. (2011). “Probiotic properties of bifidobacteria” in Bifidobacteria, Genomics and Molecular Aspects, eds B. Mayo and D. van Sinderen (Norwich: Caister Academic Press), 97–123.
Saez-Lara, M. J., Gomez-Llorente, C., Plaza-Diaz, J., and Gil, A. (2015). The role of probiotic lactic acid bacteria and bifidobacteria in the prevention and treatment of inflammatory bowel disease and other related diseases: a systematic review of randomized human clinical trials. Biomed. Res. Int. 2015:505878. doi: 10.1155/2015/505878
Salminen, S., and van Loveren, H. (2012). Probiotics and prebiotics: health claim substantiation. Microb. Ecol. Health Dis. 23:18568. doi: 10.3402/mehd.v23i0.18568
Sanchez, J. I., Marzorati, M., Grootaert, C., Baran, M., Van Craeyveld, V., Courtin, C. M., et al. (2009). Arabinoxylan-oligosaccharides (AXOS) affect the protein/carbohydrate fermentation balance and microbial population dynamics of the simulator of human intestinal microbial ecosystem. Microb. Biotechnol. 2, 101–113. doi: 10.1111/j.1751-7915.2008.00064.x
Schaafsma, G., and Slavin, J. L. (2015). Significance of inulin fructans in the human diet. Compr. Rev. Food Sci. Food Saf. 14, 37–47. doi: 10.1111/1541-4337.12119
Schell, M. A., Karmirantzou, M., Snel, B., Vilanova, D., Berger, B., Pessi, G., et al. (2002). The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. U.S.A. 99, 14422–14427. doi: 10.1073/pnas.212527599
Scott, K. P., Antoine, J. M., Midtvedt, T., and van Hemert, S. (2015). Manipulating the gut microbiota to maintain health and treat disease. Microb. Ecol. Health Dis. 26, 25877. doi: 10.3402/mehd.v26.25877
Scott, K. P., Martin, J. C., Chassard, C., Clerget, M., Potrykus, J., Campbell, G., et al. (2011). Substrate-driven gene expression in Roseburia inulinivorans: importance of inducible enzymes in the utilization of inulin and starch. Proc. Natl. Acad. Sci. U.S.A. 1, 4672–4679. doi: 10.1073/pnas.1000091107
Scott, K. P., Martin, J. C., Duncan, S. H., and Flint, H. J. (2014). Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. FEMS Microbiol. Ecol. 87, 30–40. doi: 10.1111/1574-6941.12186
Selak, M., Rivière, A., Moens, F., Van den Abbeele, P., Geirnaert, A., Rogelj, I., et al. (2016). Inulin-type fructan fermentation by bifidobacteria depends on the strain rather than the species and region in the human intestine. Appl. Microbiol. Biotechnol. 100, 4097–4107. doi: 10.1007/s00253-016-7351-9
Sharon, G., Garg, N., Debelius, J., Knight, R., Dorrestein, P. C., and Mazmanian, S. K. (2014). Specialized metabolites from the microbiome in health and disease. Cell Metab. 20, 719–730. doi: 10.1016/j.cmet.2014.10.016
Slavin, J. (2013). Fiber and prebiotics: mechanisms and health benefits. Nutrients 5, 1417–1435. doi: 10.3390/nu5041417
Snelders, J., Olaerts, H., Dornez, E., Van de Wiele, T., Aura, A. M., Vanhaecke, L., et al. (2014). Structural features and feruloylation modulate the fermentability and evolution of antioxidant properties of arabinoxylanoligosaccharides during in vitro fermentation by human gut derived microbiota. J. Funct. Foods 10, 1–12. doi: 10.1016/j.jff.2014.05.011
Sommer, F., and Bäckhed, F. (2013). The gut microbiota - masters of host development and physiology. Nat. Rev. Microbiol. 11, 227–238. doi: 10.1038/nrmicro2974
Swennen, K., Courtin, C. M., Lindemans, G. C. J. E., and Delcour, J. A. (2006). Large-scale production and characterisation of wheat bran arabinoxylooligosaccharides. J. Sci. Food Agric. 86, 1722–1731. doi: 10.1002/jsfa.2470
Tannock, G. W. (2010). “Analysis of bifidobacterial populations in bowel ecology studies” in Bifidobacteria, Genomics and Molecular Aspects, eds B. Mayo and D. van Sinderen (Norwich: Caister Academic Press), 1–15.
Tap, J., Mondot, S., Levenez, F., Pelletier, E., Caron, C., Furet, J. P., et al. (2009). Towards the human intestinal microbiota phylogenetic core. Environ. Microbiol. 11, 2574–2584. doi: 10.1111/j.1462-2920.2009.01982.x
Tojo, R., Suárez, A., Clemente, M. G., de los Reyes-Gavilán, C. G., Margolles, A., Gueimonde, M., et al. (2014). Intestinal microbiota in health and disease: role of bifidobacteria in gut homeostasis. World J. Gastroenterol. 20, 15163–15176. doi: 10.3748/wjg.v20.i41.15163
Tralongo, P., Tomasello, G., Sinagra, E., Damiani, P., Leone, A., Palumbo, V. D., et al. (2014). The role of butyric acid as a protective agent against inflammatory bowel diseases. Euromediterranean Biomed. J. 9, 24–35. doi: 10.3269/1970-5492.2014.9.4
Tuohy, K. M., and Scott, K. P. (2015). “The microbiota of the human gastrointestinal tract: a molecular view” in Diet-Microbe Interactions in the Gut, eds K. M. Tuohy and D. Del Rio (London: Elsevier), 1–15.
Turroni, F., Özcan, E., Milani, C., Mancabelli, L., Viappiani, A., van Sinderen, D., et al. (2015). Glycan cross-feeding activities between bifidobacteria under in vitro conditions. Front. Microbiol. 6:1030. doi: 10.3389/fmicb.2015.01030
Turroni, F., Foroni, E., Pizzetti, P., Giubellini, V., Ribbera, A., Merusi, P., et al. (2009). Exploring the diversity of the bifidobacterial population in the human intestinal tract. Appl. Environ. Microbiol. 75, 1534–1545. doi: 10.1128/AEM.02216-08
Turroni, F., Peano, C., Pass, D. A., Foroni, E., Severgnini, M., Claesson, M. J., et al. (2012). Diversity of bifidobacteria within the infant gut microbiota. PLoS ONE 7:e36957. doi: 10.1371/journal.pone.0036957
Van Craeyveld, V., Swennen, K., Dornez, E., Van de Wiele, T., Marzorati, M., Verstraete, W., et al. (2008). Structurally different wheat-derived arabinoxylooligosaccharides have different prebiotic and fermentation properties in rats. J. Nutr. 138, 2348–2355. doi: 10.3945/jn.108.094367
Van den Abbeele, P., Belzer, C., Goossens, M., Kleerebezem, M., De Vos, W. M., Thas, O., et al. (2013a). Butyrate-producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. 7, 949–961. doi: 10.1038/ismej.2012.158
Van den Abbeele, P., Gerard, P., Rabot, S., Bruneau, A., El Aidy, S., Derrien, M., et al. (2011). Arabinoxylans and inulin differentially modulate the mucosal and luminal gut microbiota and mucin-degradation in humanized rats. Environ. Microbiol. 13, 2667–2680. doi: 10.1111/j.1462-2920.2011.02533.x
Van den Abbeele, P., Verstraete, W., El Aidy, S., Geirnaert, A., and Van de Wiele, T. (2013b). Prebiotics, faecal transplants and microbial network units to stimulate biodiversity of the human gut microbiome. Microb. Biotechnol. 6, 335–340. doi: 10.1111/1751-7915.12049
van den Broek, L. A. M., Hinz, S. W. A., Beldman, G., Vincken, J. P., and Voragen, A. G. J. (2008). Bifidobacterium carbohydrases - their role in breakdown and synthesis of (potential) prebiotics. Nutr. Food. Res. 52, 146–163. doi: 10.1002/mnfr.200700121
van den Broek, L. A. M., and Voragen, A. G. J. (2008). Bifidobacterium glycoside hydrolases and (potential) prebiotics. Innov. Food Sci. Emerg. Technol. 9, 401–407. doi: 10.1016/j.ifset.2007.12.006
Van der Meulen, R., Adriany, T., Verbrugghe, K., and De Vuyst, L. (2006a). Kinetic analysis of bifidobacterial metabolism reveals a minor role for succinic acid in the regeneration of NAD+ through its growth-associated production. Appl. Environ. Microbiol. 72, 5204–5210. doi: 10.1128/AEM.00146-06
Van der Meulen, R., Avonts, L., and De Vuyst, L. (2004). Short fractions of oligofructose are preferentially metabolized by Bifidobacterium animalis DN-173 010. Appl. Environ. Microbiol. 70, 1923–1930. doi: 10.1128/AEM.70.4.1923-1930.2004
Van der Meulen, R., Makras, L., Verbrugghe, K., Adriany, T., and De Vuyst, L. (2006b). In vitro kinetic analysis of oligofructose consumption by Bacteroides and Bifidobacterium spp. indicates different degradation mechanisms. Appl. Environ. Microbiol. 72, 1006–1012. doi: 10.1128/AEM.72.2.1006-1012.2006
Van Laere, K. M. J., Hartemink, R., Bosveld, M., Schols, H. A., and Voragen, A. G. J. (2000). Fermentation of plant cell wall derived polysaccharides and their corresponding oligosaccharides by intestinal bacteria. J. Agric. Food Chem. 48, 1644–1652. doi: 10.1021/jf990519i
Velasquez-Manoff, M. (2015). Gut microbiome: the peacekeepers. Nature 518, S3–S11. doi: 10.1038/518S3a
Ventura, M., O'Flaherty, S., Claesson, M. J., Turroni, F., Klaenhammer, T. R., van Sinderen, D., et al. (2009). Genome-scale analyses of health-promoting bacteria: probiogenomics. Nat. Rev. Microbiol. 7, 61–71. doi: 10.1038/nrmicro2047
Ventura, M., Turroni, F., Bottacini, F., Giubellini, V., and van Sinderen, D. (2011). “Bifidobacterial ecology and comparative genomics: perspectives and prospects,” in Bifidobacteria, Genomics and Molecular Aspects, eds B. Mayo and D. van Sinderen (Norwich: Caister Academic Press), 31–44.
Ventura, M., Turroni, F., Lugli, G. A., and van Sinderen, D. (2014). Bifidobacteria and humans: our special friends, from ecological to genomics perspectives. J. Sci. Food Agric. 94, 163–168. doi: 10.1002/jsfa.6356
Verbeke, K. (2014). “Prebiotics and synbiotics: how do they affect health?” in Clinical Insights: Probiotics, Prebiotics and Gut Health, eds M. H. Floch and A. Kim (London: Future Medicine Ltd.), 47–61.
Verspreet, J., Damen, D., Broekaert, W. F., Verbeke, K., Delcour, J. A., and Courtin, C. M. (2016). A critical look at prebiotics within the dietary fiber concept. Annu. Rev. Food Sci. Technol. 7, 167–190. doi: 10.1146/annurev-food-081315-032749
Vital, M., Howe, A. C., and Tiedje, J. M. (2014). Revealing the bacterial butyrate synthesis pathways by analyzing (meta)genomic data. MBio 5, e00889–e00814. doi: 10.1128/mBio.00889-14
Vrieze, A., Van Nood, E., Holleman, F., Salojärvi, J., Kootte, R. S., Bartelsman, J. F., et al. (2012). Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 143, 913–916. doi: 10.1053/j.gastro.2012.06.031
Walker, A. W., Duncan, S. H., Louis, P., and Flint, H. J. (2014). Phylogeny, culturing, and metagenomics of the human gut microbiota. Trends Microbiol. 22, 267–274. doi: 10.1016/j.tim.2014.03.001
Walton, G. E., Lu, C., Trogh, I., Arnaut, F., and Gibson, G. R. (2012). A randomised, double-blind, placebo controlled cross-over study to determine the gastrointestinal effects of consumption of arabinoxylan-oligosaccharides enriched bread in healthy volunteers. Nutr. J. 11:36. doi: 10.1186/1475-2891-11-36
Warchol, M., Perrin, S., Grill, J. P., and Schneider, F. (2002). Characterization of a purified beta-fructofuranosidase from Bifidobacterium infantis ATCC 15697. Lett. Appl. Microbiol. 35, 462–467. doi: 10.1046/j.1472-765X.2002.01224.x
Whitman, W. B., Coleman, D. C., and Wiebe, W. J. (1998). Prokaryotes: the unseen majority. Proc. Natl. Acad. Sci. U.S.A. 95, 6578–6583. doi: 10.1073/pnas.95.12.6578
Wikoff, W. R., Anfora, A. T., Liu, J., Schultz, P. G., Lesley, S. A., Peters, E. C., et al. (2009). Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. U.S.A. 106, 3698–3703. doi: 10.1073/pnas.0812874106
Wu, N., Yang, X., Zhang, R., Li, J., Xiao, X., Hu, Y., et al. (2013). Dysbiosis signature of fecal microbiota in colorectal cancer patients. Microb. Ecol. 66, 462–470. doi: 10.1007/s00248-013-0245-9
Xu, M. Q., Cao, H. L., Wang, W. Q., Wang, S., Cao, X. C., Yan, F., et al. (2015). Fecal microbiota transplantation broadening its application beyond intestinal disorders. World J. Gastroenterol. 21, 102–111. doi: 10.3748/wjg.v21.i1.102
Yang, J., Martínez, I., Walter, J., Keshavarzian, A., and Rose, D. J. (2013). In vitro characterization of the impact of selected dietary fibers on fecal microbiota composition and short chain fatty acid production. Anaerobe 23, 74–81. doi: 10.1016/j.anaerobe.2013.06.012
Keywords: bifidobacteria, butyrate-producing colon bacteria, cross-feeding, prebiotics, probiotics, arabinoxylan-oligosaccharides, inulin-type fructans
Citation: Rivière A, Selak M, Lantin D, Leroy F and De Vuyst L (2016) Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 7:979. doi: 10.3389/fmicb.2016.00979
Received: 29 April 2016; Accepted: 07 June 2016;
Published: 28 June 2016.
Edited by:
Marco Ventura, University of Parma, ItalyReviewed by:
Sinead Christine Leahy, AgResearch Limited, New ZealandChristian U. Riedel, University of Ulm, Germany
Copyright © 2016 Rivière, Selak, Lantin, Leroy and De Vuyst. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luc De Vuyst, bGR2dXlzdEB2dWIuYWMuYmU=