 Natacha C. Gómez
Natacha C. Gómez Juan M. P. Ramiro2
Juan M. P. Ramiro2- 1Department of Food and Experimental Nutrition, Food Microbiology, Faculty of Pharmaceutical Sciences, Food Research Center, University of São Paulo, São Paulo, Brazil
- 2University of Jaén, Jaén, Spain
Use of probiotic biofilms can be an alternative approach for reducing the formation of pathogenic biofilms in food industries. The aims of this study were (i) to evaluate the probiotic properties of bacteriocinogenic (Lactococcus lactis VB69, L. lactis VB94, Lactobacillus sakei MBSa1, and Lactobacillus curvatus MBSa3) and non-bacteriocinogenic (L. lactis 368, Lactobacillus helveticus 354, Lactobacillus casei 40, and Weissela viridescens 113) lactic acid bacteria (LAB) isolated from Brazilian’s foods and (ii) to develop protective biofilms with these strains and test them for exclusion of Listeria monocytogenes, Escherichia coli O157:H7, and Salmonella Typhimurium. LAB were tested for survival in acid and bile salt conditions, surface properties, biosurfactant production, β-galactosidase and gelatinase activity, antibiotic resistance and presence of virulence genes. Most strains survived exposure to pH 2 and 4% bile salts. The highest percentages of auto-aggregation were obtained after 24 h of incubation. Sixty-seven percentage auto-aggregation value was observed in W. viridescens 113 and Lactobacillus curvatus MBSa3 exhibited the highest co-aggregation (69% with Listeria monocytogenes and 74.6% with E. coli O157:H7), while the lowest co-aggregation was exhibited by W. viridescens 113 (53.4% with Listeria monocytogenes and 38% with E. coli O157:H7). Tests for hemolytic activity, bacterial cell adherence with xylene, and drop collapse confirmed the biosurfactant-producing ability of most strains. Only one strain (L. lactis 368) produced β-galactosidase. All strains were negative for virulence genes cob, ccf, cylLL, cylLs, cyllM, cylB, cylA and efaAfs and gelatinase production. The antibiotic susceptibility tests indicated that the MIC for ciprofloxacin, clindamycin, gentamicin, kanamycin, and streptomycin did not exceed the epidemiological cut-off suggested by the European Food Safety Authority. Some strains were resistant to one or more antibiotics and resistance to antibiotics was species and strain dependent. In the protective biofilm assays, strains L. lactis 368 (bac-), Lactobacillus curvatus MBSa3 (bac+), and Lactobacillus sakei MBSa1 (bac+) resulted in more than six log reductions in the pathogens counts when compared to the controls. This effect could not be attributed to bacteriocin production. These results suggest that these potential probiotic strains can be used as alternatives for control of biofilm formation by pathogenic bacteria in the food industry, without conferring a risk to the consumers.
Introduction
Lactic acid bacteria (LAB) constitute part of the autochthonous microbiota of many types of foods. They are defined as a cluster of lactic-acid-producing, low G + C%, non-spore-forming, Gram-positive rods and cocci and catalase-negative bacteria which share many biochemical, physiological, and genetic properties (Abriouel et al., 2012). This group of bacteria has a particular interest for food industries due to their technological properties, being often used as starter cultures to produce fermented products (Lahtinen et al., 2011). Many reports have shown that traditional fermented foods are rich sources of LAB with probiotic characteristics (Liu et al., 2011; Favaro et al., 2014; Palomino et al., 2015).
According to FAO/WHO (2006), probiotics are live microorganisms which administered in adequate amounts confer a health benefit on the host (FAO/WHO, 2006). The principal functional properties of probiotics include tolerance to acid and bile, adherence to epithelial surfaces, and antagonistic activity toward intestinal pathogens. Probiotics may confer their health benefits by several mechanisms; by contributing to colonization resistance, reinforcing the intestinal barrier (i.e., tight junction expression, secretion of mucus, and antimicrobial peptides), modulating the immune system and instructing the intestinal microbiota composition and activity (Wan et al., 2015). This is based on either direct cell–cell contact, secreting various molecules and/or microbial cross-feeding (Jonkers, 2016). Auto-aggregation of probiotic strains seems to have influence on their adhesion to intestinal epithelial cells, while co-aggregation with pathogens may prevent colonization in the gut and their consumption reduces the viable number of pathogens while strengthening body natural defenses (Savard et al., 2011). Del Re et al. (2000) demonstrated that auto-aggregation is strongly related to adhesion. In addition, adhesion of probiotic bacteria to mucosa is one of the mechanisms by which they can overcome competition with other microorganisms. Nevertheless, production of bacteriocins and other antimicrobial substances by bacteria in biofilms and adhered to mucosal surfaces is considered relevant for the displacement of pathogens, as demonstrated in gastrointestinal tract (GIT) models (Ganzle et al., 1999). Bacteriocin-producing Lactobacillus curvatus LTH 1174 provided protection against E. coli LTH 1600 and Listeria innocua DSM20649 invasion during transit through in a dynamic model of the human stomach and small intestine (GIT model; Ganzle et al., 1999) and bacteriocin-producing Lactobacillus sakei 2a protected gnotobiotic mice against experimental challenge with Listeria monocytogenes (Bambirra et al., 2007). These data suggest that bacteriocin-producing lactobacilli prevent new strains from invading or maintaining stable populations in the colon. Therefore, bacteriocin production is often considered a probiotic trait in this context.
Studies carried out both in culture media and foods have shown that bacteriocins produced by probiotic or potentially probiotic LAB can act synergistically or have an additive effect in the antimicrobial activity when combined with other antimicrobials (Viedma et al., 2010; Gómez et al., 2012). Interestingly, LAB may simultaneously secrete organic acids, bacteriocins, and biosurfactants (Kanmani et al., 2013). The precise role of these compounds on other bacterial populations present in biofilms is not yet known, but it is well recognized that bacteriocins have stronger antimicrobial activity under acidic conditions (Gálvez et al., 2010).
The presence of biofilms is a relevant risk factor in the food industry due to the potential contamination of food products with pathogenic and spoilage microorganisms. Biofilms can be formed on surfaces becoming permanent reservoirs of bacteria. Most important, biofilms may act as reservoirs of pathogenic and spoilage bacteria, in which these microorganisms can persist against the cleaning and disinfection processes. For example, contamination of equipment with biofilms was a contributing factor to 59% of food-borne disease outbreaks investigated in France (Midelet and Carpentier, 2004). The presence of biofilms is common in food industry and represents a concern because bacteria can adhere to almost any type of surface, such as plastic, metal, glass, soil particles, wood food products (Gandhi and Chikindas, 2007).
Listeria monocytogenes is commonly found in food-processing environment, and it has been isolated from both meat and dairy processing plants (Winkelströter et al., 2013) and Mendonça et al. (2012) also demonstrated that E. coli O157:H7 has the potential to form biofilm on different surfaces commonly used in food industry. Common sites for the presence of Salmonella spp. in food-processing plants are filling or packaging equipments, floor drains, walls, cooling pipes, conveyors, collators for assembling product for packaging, racks for transporting products, hand tools or gloves, freezers, etc, which are usually made of plastics (Pompermayer and Gaylarde, 2000). In addition, a study of 122 Salmonella strains indicated that all had the ability to adhere to plastic microwell plates and that; generally, more biofilm was produced in low nutrient conditions, as can be found in specific food-processing environments, compared to high nutrient conditions (Stepanović, 2004).
The increased resistance of biofilm cells to biocides can be partially due of the exopolymeric matrix interference and this can explains why the disinfectant most effective to planktonic cells is not necessarily the most active against biofilm cells (Van Houdt and Michiels, 2010). Listeria monocytogenes cells residing in so-called refuge sites such as cracks, worn equipment and in hard to reach places such as complex machinery may be subjected to suboptimal disinfection concentrations allowing them to survive and possibly adapt to cleaning and sanitation treatments (Carpentier and Cerf, 2011).
Recent trends in the transmission and emergence of resistant pathogenic bacteria through the food chain reinforce the need to investigate several alternatives for disinfection. For this reason, there is a great interest in the development of novel strategies using natural products to control the persistence of pathogens associated with surfaces or equipment especially in food industry. Therefore, biofilms formed by LAB present in foods, agricultural products or in the GIT of mammals and used as starters in food manufacturing, may offer a promising means to counteract the establishment of pathogenic biofilms (Winkelströter et al., 2013).
A very promising approach for the control of biofilm formation is the use of probiotics to colonize hard surfaces in order to counteract the proliferation of other bacterial species, based on the competitive exclusion principle (Falagas and Makris, 2009; Hibbing et al., 2010). This concept has been designated as biocontrol when the application is antagonistic toward a certain pathogen (Gatesoupe, 1999). LAB successfully reduced Listeria monocytogenes in a ready-to-eat poultry processing plant (Zhao et al., 2013) and lactobacilli with biofilm-forming aptitudes were able to control Listeria monocytogenes on abiotic surfaces (Pérez-Ibarreche et al., 2014). In addition, several studies have shown that bacteriocin-producing LAB improved the bactericidal effect of biocides on bacterial biofilms (Lobos et al., 2009; Gómez et al., 2012).
Application of bacteriocins and/or their producer strains for inhibition of biofilm formation and/or killing of cells embedded in biofilms is a novel field of research. The objectives of this study were to evaluate the potential probiotic traits of LAB isolated from different fermented Brazilian products and their inhibition effect against Escherichia coli O157:H7, Listeria monocytogenes, and S. Typhimurium biofilm formation. Tolerance to low pH and bile salts, surface properties (aggregation and co-aggregation), biosurfactant production, gelatinase activity, antibiotic resistance and virulence genes absence were evaluated as probiotic properties of the studied LAB.
Materials and Methods
Bacterial Strains and Growth Conditions
The study was conducted with eight LAB strains isolated from foods (Table 1): bacteriocin producers Lactococcus lactis VB69 and VB94 were isolated from Brazilian charqui (Bíscola et al., 2013) and Lactobacillus sakei MBSa1 and Lactobacillus curvatus MBSa3 were isolated from salami (Barbosa et al., 2015). Non-bacteriocin producers Lactococcus lactis 368, Lactobacillus helveticus 354 isolated from goat cheese and Lactobacillus casei 40 and W. viridescens 113 isolated from ripened cheese (unpublished). The strains were identified by 16S rDNA gene sequencing, according to Cibik et al. (2000), in a CEQ2000 XL DNA Analysis System (Beckman Coulter, Brea, CA, USA). LAB strains were cultivated in De Man et al. (1960) broth (Oxoid, Basingstoke, England) at 30°C for 18 h. E. coli O157:H7 ATCC 35150, Listeria monocytogenes ATCC 7644 and S. Typhimurium ATCC 14028 were cultured in trypticase soy broth (TSB, Oxoid, Basingstoke, England) at 37°C for 20 h. All strains were maintained at -80°C in the appropriate cultivation broth containing 20% (v/v) glycerol.
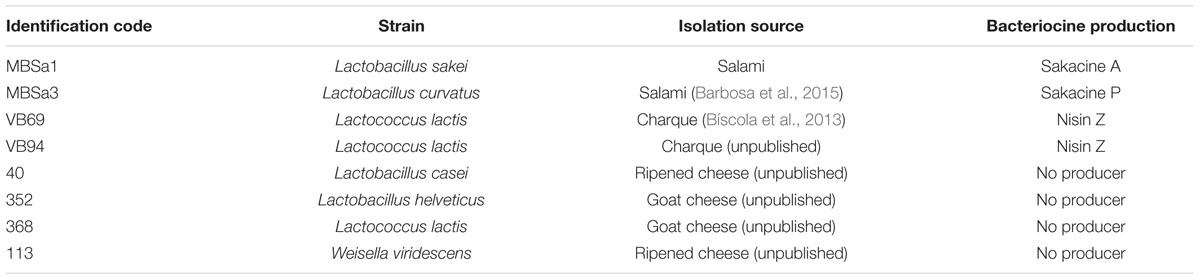
TABLE 1. Bacterial strains used in this study.
Auto-Aggregation and Co-Aggregation Assays
Aggregation abilities of LAB strains were studied as described by Collado et al. (2008), with some modifications. Bacterial cells from an overnight culture were harvested by centrifugation (5,000 × g, 20 min, 4°C), washed twice with phosphate-buffered saline PBS pH 7.1 (10 mM Na2HPO4, 1 mM KH2PO4, 140 mM NaCl, 3 mM KCl) and suspended in the same buffer. Absorbance (A600 nm) was adjusted to 0.25 ± 0.05 in order to standardize the number of bacteria (107–108 CFU/ml). The optical density (OD600 nm) of a homogenized bacterial suspension was first recorded then repeated on the same suspension left to rest for 24 h at 37°C without vortexing. The aggregation percentage was expressed as [1 - (ATime/A0) × 100] where ATime represents the absorbance of the mixture at 24 h and A0, absorbance at time 0.
For the co-aggregation assays, LAB bacterial suspensions prepared as described above were mixed with equal volumes (500 μl) of the cultures of the pathogens listed in Section “Bacterial Strains and Growth Conditions.” Mixtures were incubated at 37°C without agitation, and absorbance (OD600 nm) measured after 24 h at 37°C. The percentage of co-aggregation was calculated as [(Apathog + ALAB)/2 - (Amix)/(Apathog + ALAB)/2] × 100 (Handley et al., 1987), where Apathog and ALAB represent the absorbance in the tubes containing only the pathogen or the LAB strain, respectively, and Amix represents the absorbance of the mixture at 24 h (García-Cayuela et al., 2014).
Tolerance to Bile Salts and Acidic pH
The LAB strains were tested for bile salt tolerance (0–10%) and survival at low pH (1.5–3) according to Millette et al. (2008). The bile salt tolerance was ascertained in MRS agar containing a commercial preparation of bile salts normally used to inhibit the growth of Gram-positive bacteria in broth (Sigma–Aldrich, B-3426). The bile salt mixture was added in concentrations varying from 0 to 10% with increments of 1%. Another bile salt preparation (LP 0055; Oxoid, Basingstoke, England) was also evaluated in concentrations varying from 0 to 20% with increments of 4% to avoid differences between the different compounds. The MRS agar containing the bile salts was autoclaved for 15 min at 121°C, cooled, and plated. Aliquots of overnight MRS broth cultures (100 μl of bacteria in the stationary phase obtained after 24 h of growth) were inoculated onto the surface of the bile-salt-containing MRS agar, and incubated at 37°C for 72 h. The plates were examined visually for bacterial growth as a lawn, indicating resistance to bile salts in the tested concentration. For determination of acid tolerance, 1 ml overnight MRS broth cultures were inoculated onto 19 ml of simulated gastric fluid (3.2 g/l pepsin and 2 g/l NaCl) adjusted to different pHs (1.5, 2, 2.5, and 3) values with 5 M HCl. After incubation for 30 min at 37°C, 1 ml of the mixture was removed to determine viable counts (expressed as CFU/ml) on MRS agar taking as reference the concentration of bacteria not exposed to simulated gastric fluid. Lactobacillus rhamnosus GG (lab collection) was used as a positive control because it is a probiotic bacterium well known for its resistance to gastrointestinal conditions.
β-Galactosidase Activity
The LAB strains were grown in MRS broth at 37°C for 24 h, streaked onto MRS agar and incubated at 37°C for 48 h. One colony was transferred to a tube containing a disk of O-nitrophenyl-β-D-galactopyranoside—ONGP (Sigma–Aldrich) and 100 μl sterile saline (0.85% NaCl). A yellow color indicated the release of o-nitrophenol (chromogenic compound) and represented a positive result for the production of β-galactosidase.
Hemolytic Activity
Testing for hemolytic activity was carried out as described by Carrillo et al. (1996). Isolated strains were screened for hemolytic activity on blood agar plates containing 5% (v/v) horse blood and incubated at 30°C for 24–48 h. A clear zone around the colony indicated hemolytic activity, which was probably caused by surfactant production. The zones of clearing were scored as follows: (-) no hemolysis; (+) incomplete hemolysis, when the zone was not totally clear; (++) complete hemolysis with a diameter of lysis < 1 cm; (+++) complete hemolysis with a diameter of lysis between 1 cm and 3 cm; and (++++) complete hemolysis with a diameter of lysis > 3 cm.
Drop Collapse Test
The drop collapse test was carried out as described by Jain et al. (1991). LAB were cultivated in MRS at 37°C for 24 h, centrifuged at 12,000 × g for 5 min and 100 μl of the supernatants were added to each well of 96-well microplates (TPP, Switzerland) and then 5 μl of crude motor oil was added to the surface. A result was considered positive for biosurfactant production when the drop diameter was at least 1 mm larger than that produced by deionized water (negative control). Each test was repeated in two separate microtiter plates.
Microbial Adhesion to Hydrocarbon Test (MATH)
Bacterial cell surface hydrophobicity was assessed by measuring adhesion to hydrocarbons (MATH) as described by Kotzamanidis et al. (2010). LAB cultivated in MRS at 37°C for 24 h were washed twice in phosphate-buffered saline (PBS; 10 mM Na2HPO4, 1 mM KH2PO4, 140 mM NaCl, 3 mM KCl) and re-suspended in 3 mL of 0.1 M KNO3 to achieve approximately 108 CFU/ml (OD600 nm = 0.2). Absorbance of the suspension was measured at 600 nm (A0). One microliter of xylene was added to the cell suspension to form a two-phase system and after 10 min at room temperature, the two-phase system was mixed by vortexing for 2 min. After 20 min at room temperature (approximately 23°C), the aqueous phase was carefully removed and absorbance at 600 nm (A1) measured. The percentage of cell surface hydrophobicity (H, %) was calculated using the following formula: H (%) = (1 A1/A0) ∗ 100, where A1 represents the absorbance of the mixture after 20 min at room temperature and A0, absorbance at time 0.
Gelatinase Activity
Gelatinase production was verified by spotting 1 μl aliquots of the 24 h cultures onto the surface of five Luria Bertani agar plates (BD, Franklin Lakes, NJ, USA) supplemented with 3% (w/v) gelatin (BD). Plates were incubated at 37°C and 42°C for 48 h, 25°C for 72 h, and 10°C and 15°C for 10 days. After incubation, the plates were maintained at 4°C for 4 h and the hydrolysis of gelatin was recorded by the formation of opaque halos around the colonies (Perin et al., 2014).
Antibiotic Resistance
The resistance to antibiotics was determined by the broth microdilution protocol according to Muñoz et al. (2014) with some modifications. Antibiotics employed in this study were β-lactams (ampicillin: AMP), quinolone (ciprofloxacin: CIP), lincosamide (clindamycin: CLI), aminoglycosides (gentamicin: GEN, kanamycin: KAN and streptomycin: STR), macrolides (erythromycin: ERY), glycopeptides (vancomycin: VAN), chloramphenicol: CMP and tetracycline: TET. These antibiotics were selected based on the European Food Safety Authority recommendations for probiotics strains (European Food Safety Authority [EFSA], 2012). All antibiotics were purchased from Sigma–Aldrich, USA. To prepare the stock antibiotic solutions, each antibiotic was weighed, dissolved in sterile distilled water (except CMP which was dissolved in sterile distilled water with 0.5% of ethanol), filter-sterilized (0.2 mm) and kept at -20°C until use. The working solutions at specific concentrations were prepared daily. Overnight cultures were adjusted to OD600 nm of 0.8 (109 CFU/ml) with PBS, and used to inoculate (1% v/v) Mueller Hinton broth (Oxoid, Basingstoke, England) containing each antibiotic at tested concentrations (final volume of 100 μl per well of 96 micro-well plates). The plates were incubated at 37°C for 24 h. Resistance rates were calculated according to microbial cut-off values (mg/ml), as recommended by the European Food Safety Authority [EFSA] (2012). The microbiological breakpoints were defined according to Danielsen and Wind (2003), Flórez et al. (2005) and the European Commission (European Commission SCAN, 2007).
Virulence Genes
Total DNA extraction was performed using a Blood and Tissue mini kit Quiagen (German Town, USA). The primers used for the amplification of genes esp, agg, gelE, efaAfm and efaAfs, cylA, cylB and cylM were described by Eaton and Gasson (2001), and primers of cyl operon (cylLL and cylLS) were developed by Semedo et al. (2003). Table 2 describes the primers used in these tests. All primers were synthesized by Life Technology (Brazil). PCR amplifications were performed in a ThermoCycler AB (Applied Biosystems Veriti, NJ, USA), in 0.2-ml reaction tubes containing 25 μl of GoTaq® Green Master Mix, 2.5 μ1 (10 μM) of each primer, and 1 μl (100 ng) of DNA. Amplification reactions were as follows: initial cycle of 94°C for 1 min, 35 cycles of 94°C for 1 min, 55°C for 1 min, 72°C for 2 min, a final extension step of 72°C for 7 min and then cooling to 4°C. Amplification products were submitted to electrophoresis in 1% (w/v) agarose gel at 100 V for 30 min. A 100-bp PCR DNA ladder was used as the molecular weight marker. The gels were photographed on a Gel DocTM XR+ System (BioRad, Richmond, CA, USA), and image analysis was accomplished using Quantity One software. The positive control was Enterococcus faecalis FI 9190 (obtained from Eaton and Gasson, 2001, Institute of Food Research, Norwich Research Park, Norwich, UK). For each PCR, a negative control (sample without template) was included.

TABLE 2. Primers used to test for the presence of virulence genes.
Biofilm Assay
The quantification of biofilm production was performed as described previously by Borges et al. (2012) with some modifications. The wells of a sterile 12-well polystyrene microtiter plate (TPP, Switzerland) were filled with 2 ml of MRS broth, absorbance (A600 nm) of bacterial suspensions in MRS was adjusted to 0.25 ± 0.05 in order to standardize the number of bacteria (107–108 CFU/ml) and 200 μl of overnight was added to each well. The plates were incubated aerobically for 48 h at 30°C. To quantify the biofilm formation, the wells were gently washed three times with 2 ml of sterile distilled water. The attached bacteria were fixed with 2 ml of methanol (Romyl, Leics, UK) for 15 min, and then, microplates were emptied and dried at room temperature. Subsequently, 2 ml of a 2% (v/v) crystal violet solution was added to each well and held at ambient temperature for 5 min. Excess stain was then removed by placing the plate under gently running tap water. Stain was released from adherent cells with 2 ml of 33% (v/v) glacial acetic acid. The optical density (OD) of each well was measured at 595 nm using a plate reader (Microplate reader, Bio-Rad, Hercules; CA, USA). Each assay was performed in four replicates and conducted three individual times on different days under the same conditions, and the negative control was performed in uninoculated MRS broth. The cut-off (ODC) was defined as the mean OD value of the negative control. Based on the OD, strains were classified as non-biofilm producers (OD ≤ ODC), weak (ODC < OD ≤ 2 × ODC), moderate (2 × ODC < OD ≤ 4 × ODC) or strong biofilm producers (4 × ODC < OD; Borges et al., 2012).
Inhibition of Biofilm Formation
Lactic acid bacteria strains were inoculated (1% v/v) in 2 ml of MRS broth diluted to one-fifth of the concentration recommended by the manufacturer (55 g/l) and transferred (2 ml/well) to 12-well polystyrene microtiter plates (TPP, Switzerland). The plates were incubated at 30°C for 48 h for attachment of cells to the wells (biofilm formation). The broths were carefully discarded by pipetting and the biofilms visually present on the bottom and sides of the plate were washed with 2 ml PBS pH 7.1 (10 mM Na2HPO4, 1 mM KH2PO4, 140 mM NaCl, 3 mM KCl) to remove planktonic and loosely attached cells. Absorbance (A600 nm) of pathogenic bacterial suspensions in TSB was adjusted to 0.25 ± 0.05 in order to standardize the number of bacteria (107–108 CFU/ml), added to biofilms and incubated at 30°C for 24, 48, and 72h. Every 24 h, half of the broth in the wells was replaced with fresh broth. After incubation, the planktonic cultures were carefully removed and the biofilms were suspended by scrapping and vigorous shaking.. To evaluate the viable count of adherent microorganisms in the biofilm, three wells for each strain were washed three times as previously described and scraped. The obtained suspensions were transferred into sterile tubes and mixed with a vortex mixer for 30 s. Proper dilutions were prepared in saline solution 0.85% (w/v) and plated on xylose lysine deoxycholate agar (XLD) for S. Typhimurium, Modified Oxford agar (MOX) for Listeria monocytogenes and MacConkey sorbitol agar (SM) for E. coli O157:H7. The plates were incubated at 37°C for 24–48 h and bacterial counts were performed.
Listeria monocytogenes, S. Typhimurium, and E. coli O157:H7 controls were used to monitor the biofilm development of these strains without the presence of LAB biofilms.
Pathogenic planktonic cells counts were performed from the broths discarded by pipetting, following the same procedures used for biofilm cell count (data not shown).
Statistical Analysis
All experiments were carried out three times, with duplicate samples per trial, and results were expressed as average. Standard deviations were determined with Excel programme (Microsoft Corp., USA). A t-test was performed at the 95% confidence interval with PASW Statistics—SPSS 17 (IBM Co.), in order to determine the statistical significance of data.
Results
Auto-Aggregation and Co-Aggregation Assays
Aggregation values increased over time in a strain-dependent manner. W. viridescens 113 presented the highest auto-aggregation (67%), compared to the other isolates showing only moderate auto-aggregation (Figure 1). All LAB strains presented co-aggregation with pathogens (Figure 2), in a strain–pathogen combination-dependent manner. Lactobacillus curvatus MBSa3 exhibited the highest co-aggregation (69% with Listeria monocytogenes and 74.6% with E. coli O157:H7), while the lowest co-aggregation was exhibited by W. viridescens 113 (53.4% with Listeria monocytogenes and 38% with E. coli O157:H7).
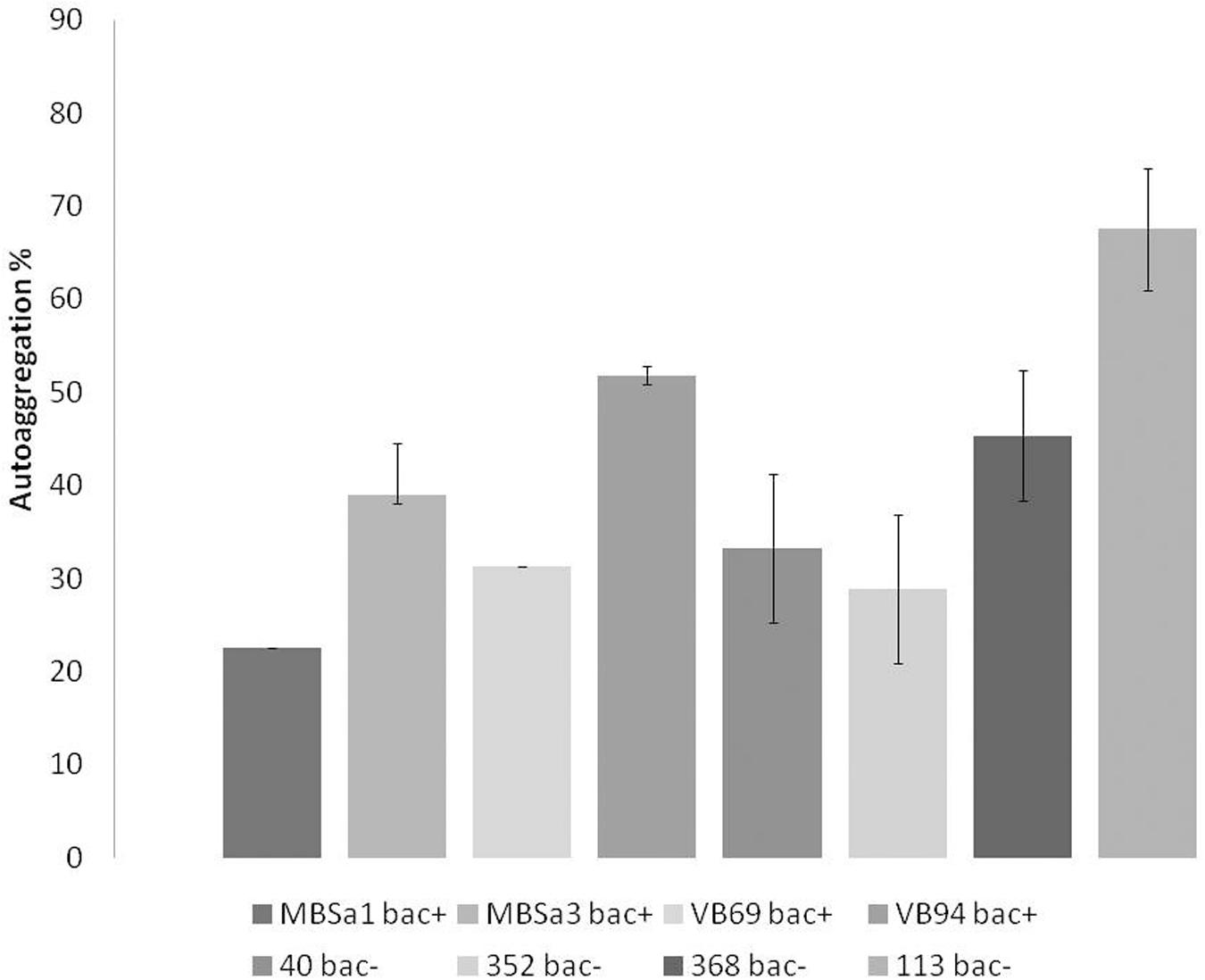
FIGURE 1. Auto-aggregation of lactic acid bacteria strains cells re-suspended in PBS (pH 7.1) evaluated after 24 h incubation at 37°C. Error bars represent standard deviations of the mean values of results from three replicated experiments (bac+ = bacteriocin producer).
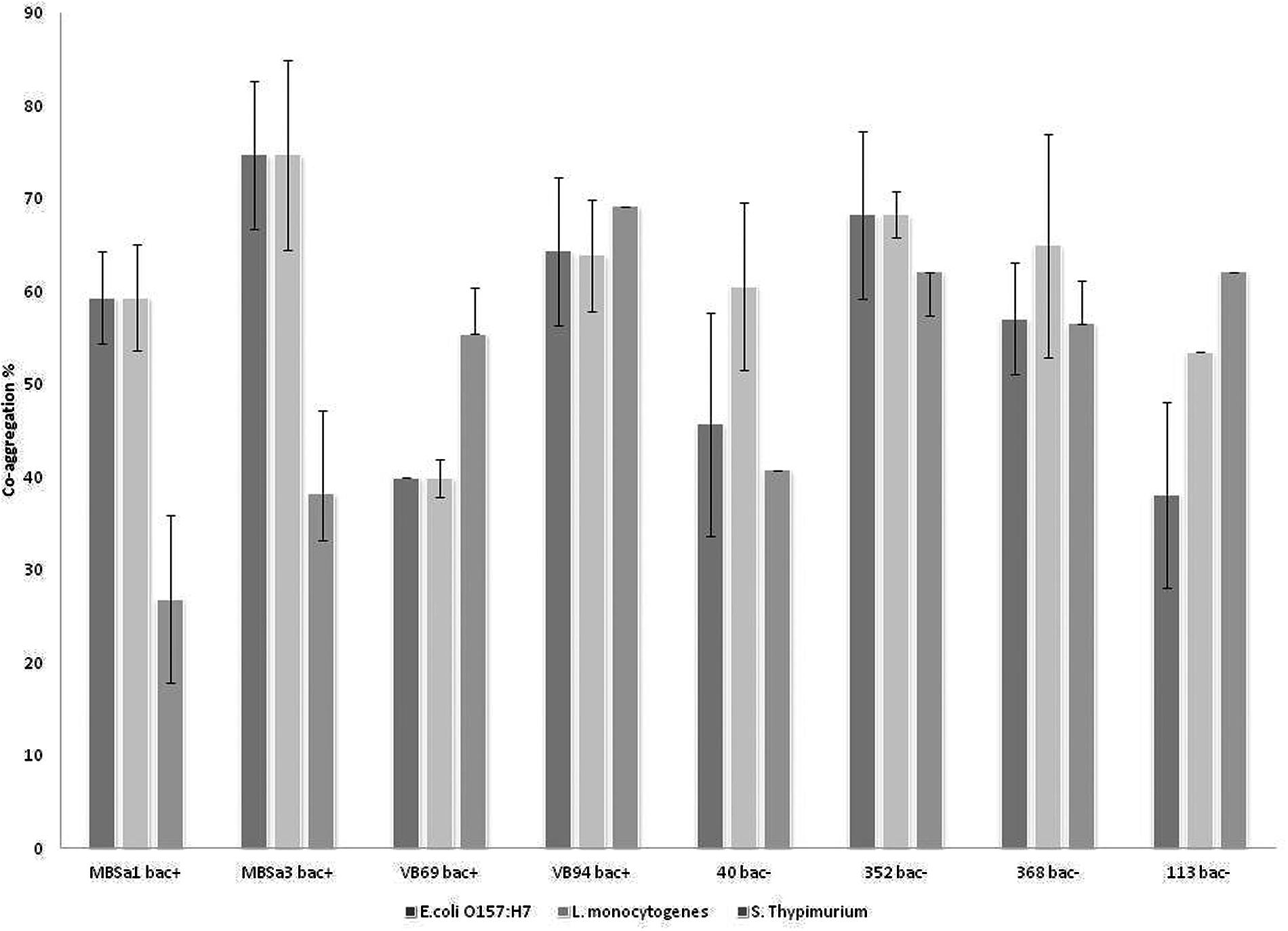
FIGURE 2. Co-aggregation values recorded for lactic acid bacteria strains with Listeria monocytogenes ATCC 7644, Salmonella Typhimurium ATCC 14028, and Escherichia coli O157:H7 ATCC 35150 after 24 h incubation at 37°C in PBS (pH 7.1). Error bars represent standard deviations of the mean values of results from three replicate experiments (bac+ = bacteriocin producer).
Tolerance to Bile Salts and Acidic pH
The results showed that tolerance for bile salts mixture from Sigma was 4% for all LAB strains. However, the tolerance to bile salts from Oxoid was 20% for W. viridescens 113 and L. lactis 94 and 8% for Lactobacillus casei 40 and L. lactis 69 (data not shown) for the rest of studied strains was 4%. The results in Figure 3 show that all tested strains, including Lactobacillus rhamnosus GG, survived to exposure to pH 2.5 for 30 min. No significant difference (p < 0.05) between the initial microbial population and the population after 30 min at pH 2.5 was observed for all strains; a reduction of viability was only observed for W. viridescens 113, approximately 2 log. However, a significant reduction of viability at pH 2.0 was observed for all tested bacteria except for L. lactis 94. In counterpart, complete survival at pH 3 and no survival at pH 1.5 were observed for all strains.
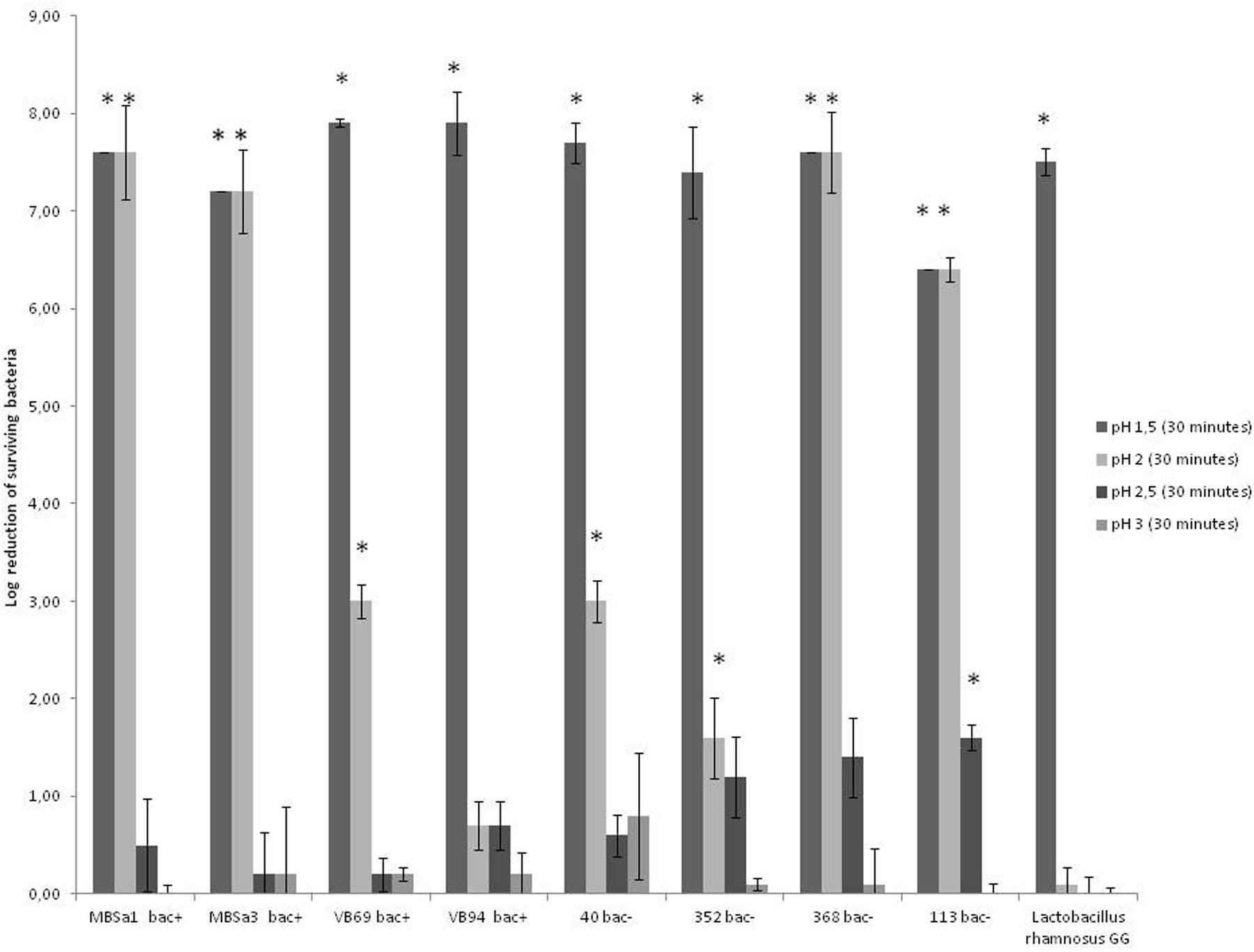
FIGURE 3. Log reduction of lactic acid bacteria strains after incubation at 37°C for 30 min in (SFG) Simulated Grastric Fluid; 3.2 g/l pepsin and 2 g/l NaCl (pH 1.5–3). Log reductions were estimated by subtracting the log of surviving to the controls (bacteria not exposed to simulated gastric fluid). Data are mean ± standard deviations. Superscript ∗ indicates a significant difference (p < 0.05) compared to the control. Detection limit is 10 CFU/ml.
Biosurfactant Production
The three screening tests indicated that all tested LAB strains were capable to produce biosurfactant (Table 3). In the hemolysis test, most strains showed zones of clearing in the blood agar with scores corresponding to (++) indicating complete hemolysis with a diameter < 1 cm. The exception was L. lactis 94 that was not hemolytic. In the MATH assay, the lowest values observed were 77.2% (Lactobacillus casei 40); 76.4% (Lactobacillus curvatus MBSa3); 81.2% (L. lactis 368) and 88.9% (Lactobacillus sakei MBSa1) for the rest of LAB studied the values was over 90% with 91.2% (Lactobacillus helveticus 352); 93.9% (W. viridescens 113); 95.1% (L. lactis 94) and 95.2% (L. lactis 69).

TABLE 3. Biosurfactant production by the LAB strains.
All strains resulted positive in the drop collapse test. Flat drops with scoring system ranging from + to ++++ corresponding to partial to complete spreading on the oil surface. The strains studied did not present complete spreading on the oil surface only a partial spreading was observed, varying between + for L. lactis 94, Lactobacillus casei 40, and Lactobacillus helveticus 352 to ++ in the rest of strains studied, L. lactis 69, W. viridescens 113 Lactobacillus sakei MBSa1 and Lactobacillus curvatus MBSa3.
Antibiotic Resistance, Presence of Virulence Genes and Gelatinase Activity
The antibiotic susceptibility tests (Table 4) indicated that the MIC for ciprofloxacin, clindamycin, gentamicin, kanamycin, and streptomycin did not exceed the epidemiological cut-off suggested by the European Food Safety Authority [EFSA] (2012) for all tested strains. All strains were sensitive to β-lactams (ampicillin: AMP), except Lactobacillus curvatus MBSa1. Some strains were resistant to one or more antibiotics: Lactobacillus casei 40 and Lactobacillus curvatus MBSa1 were resistant to erythromycin, Lactobacillus sakei MBSa3 and Lactobacillus casei 40 were resistant to chloramphenicol; Lactobacillus curvatus MBSa1, L. lactis 94 and 368 were resistant to vancomycin. Only Lactobacillus casei 40, Lactobacillus helveticus 352 and L. lactis 69 were sensitive to tetracycline. All strains were sensitive to erythromycin, except Lactobacillus casei 40 and Lactobacillus curvatus MBSa1.
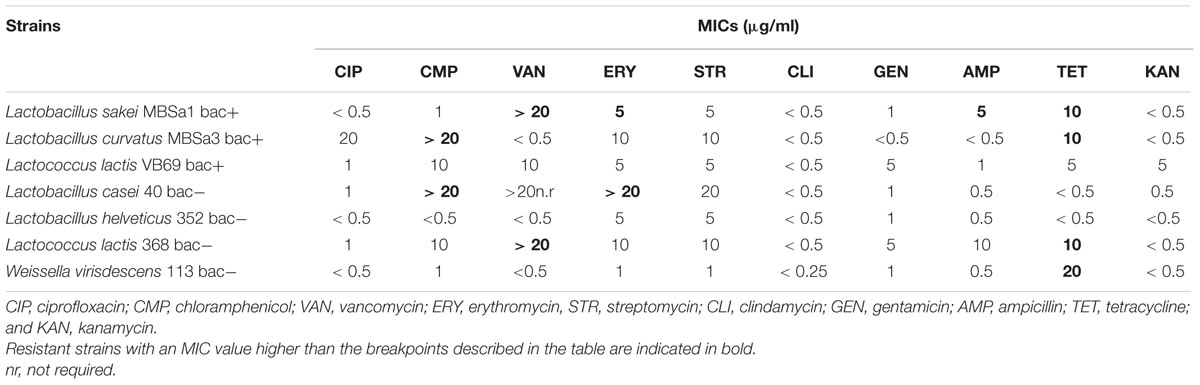
TABLE 4. Determination of minimal inhibitory concentration (MIC) against the LAB strains.
Table 5 shows the presence of the virulence genes tested in the LAB strains. All strains were negative for GelE, cob, ccf, cylLL, cylLs, cyllM, cylB, cylA and efaAfs, except W. viridescens 113 that was positive for cob and for GelE. Nevertheless, no strain presented gelatinase activity. Lactobacillus helveticus 352 was positive for Agg and efaAfm, L. lactis 368 for cpd and efaAfm too and the presence of esp was observed in L. lactis 94 and 69, Lactobacillus casei 40, and Lactobacillus curvatus MBSa3.
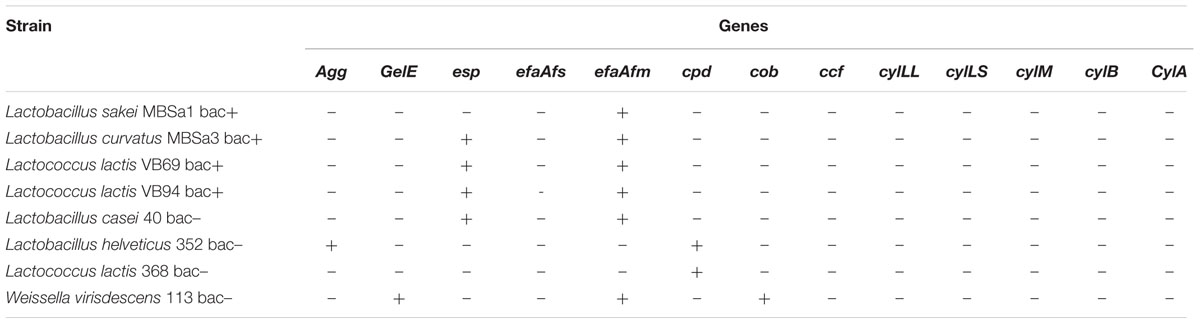
TABLE 5. Presence of virulence genes in the LAB strains.
Biofilm Assay
All the strains studies were biofilm producers in MRS. The biofilm production was strain dependent (Figure 4). Based on the OD, all the strains studied were strong producer’s except W. viridescens 113. The highest values over 1, were observed for L. lactis 368 (1.65), Lactobacillus helveticus 352 (1.38) and L. lactis 94 (1.10). The values for the rest of strains were under 1, but all were strong biofilm producers except W. viridescens 113 with moderate biofilm formation.
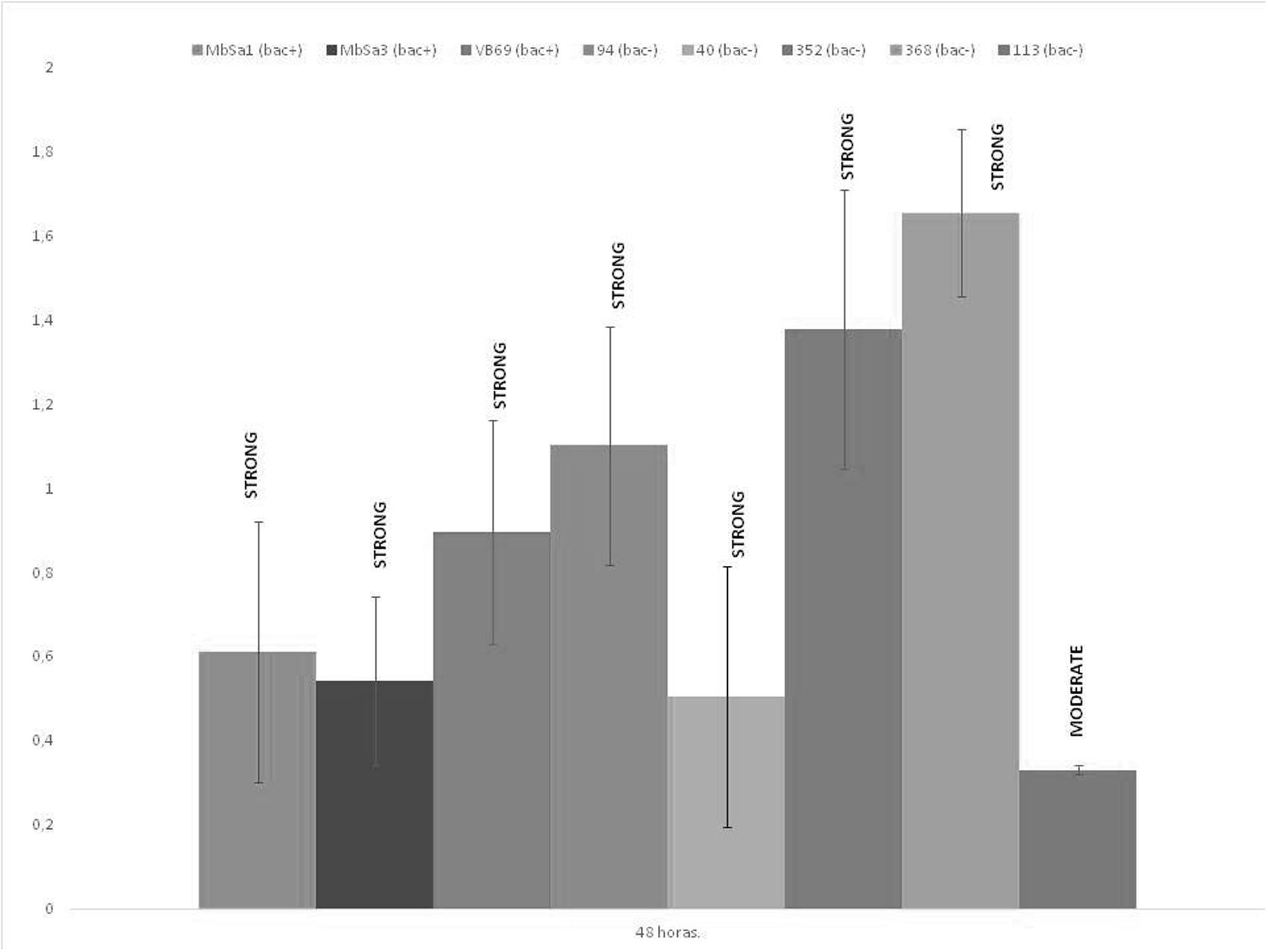
FIGURE 4. Biofilm formation of LAB using the microtiter plate assay. After incubation at 30°C for 48 h in MRS media. Data are mean ± standard deviations. The cut-off (ODC) was defined as the mean OD value of the negative control. Based on the OD, strains were classified as non-biofilm producers (OD ≤ ODC), weak (ODC < OD ≤ 2 × ODC), moderate (2 × ODC < OD 4 × ODC) or strong biofilm producers (4 × ODC < OD).
Inhibition of Biofilm Formation
The total inhibition in pathogens E. coli O157:H7, Listeria monocytogenes and S. Typhimurium biofilm formation, in 24, 48, and 72 h of exposure, was obtained for L. lactis 368 (bac-), Lactobacillus curvatus MBSa3 (bac+) and Lactobacillus sakei MBSa1 (bac+). For the other strains, the inhibition was time-dependent and varied according to the strain and target pathogen (Figure 5). The presence of sessile cells of E. coli O157:H7, Listeria monocytogenes and S. Typhimurium in the presence of LAB in 24, 48, and 72 h was significantly reduced in comparison to the pure cultures (p < 0.05). Listeria monocytogenes was not detected within L. lactis 69 (bac+) and 94 (bac+) established biofilms following 24 h and 48h interaction periods. Nevertheless, the presence of Listeria monocytogenes biofilms were observed in the cases of W. viridescens 113 (bac-), Lactobacillus casei 40 (bac-) and Lactobacillus helveticus 352 (bac-); 4 log of decrease was observed for 24 h of incubation in presence of Lactobacillus helveticus 352 (bac-) biofilm, as well as, 7 log of decrease for Lactobacillus casei 40 (bac-) during the same incubation time. After 48 h of incubation 5 log of decrease were detected in the presence of W. viridescens 113 (bac-). The presence of Listeria monocytogenes biofilms was detected during 72 h of incubation in all cases, varying between 4 log for W. viridescens 113 (bac-), Lactobacillus helveticus 352 (bac-) and Lactobacillus casei 40 (bac-) to 6 log of decrease in the cases of L. lactis 94 (bac+) and 69 (bac+). In S. Typhimurium experiment, sessile cells were not detected during 24 h of incubation in the presence of most LAB tested, only for Lactobacillus helveticus 352 (bac-) 2 log were achieved (6 log of decrease). After 48 and 72 h only in the presence of Lactobacillus casei 40 (bac-) sessile cells of S. Typhimurium were not detected. For E. coli O157:H7 only after 24 h of incubation the presence was not detected, except for Lactobacillus helveticus 352 (bac-). During 48 and 72 h approximately 3 log of E. coli was detected (5 log of decrease) in the presence of all tested LAB. In most cases, reductions between 5 and 3 log for E. coli O157:H7, 4log for S. Typhimurium and between 7 and 3 log for Listeria monocytogenes were achieved.
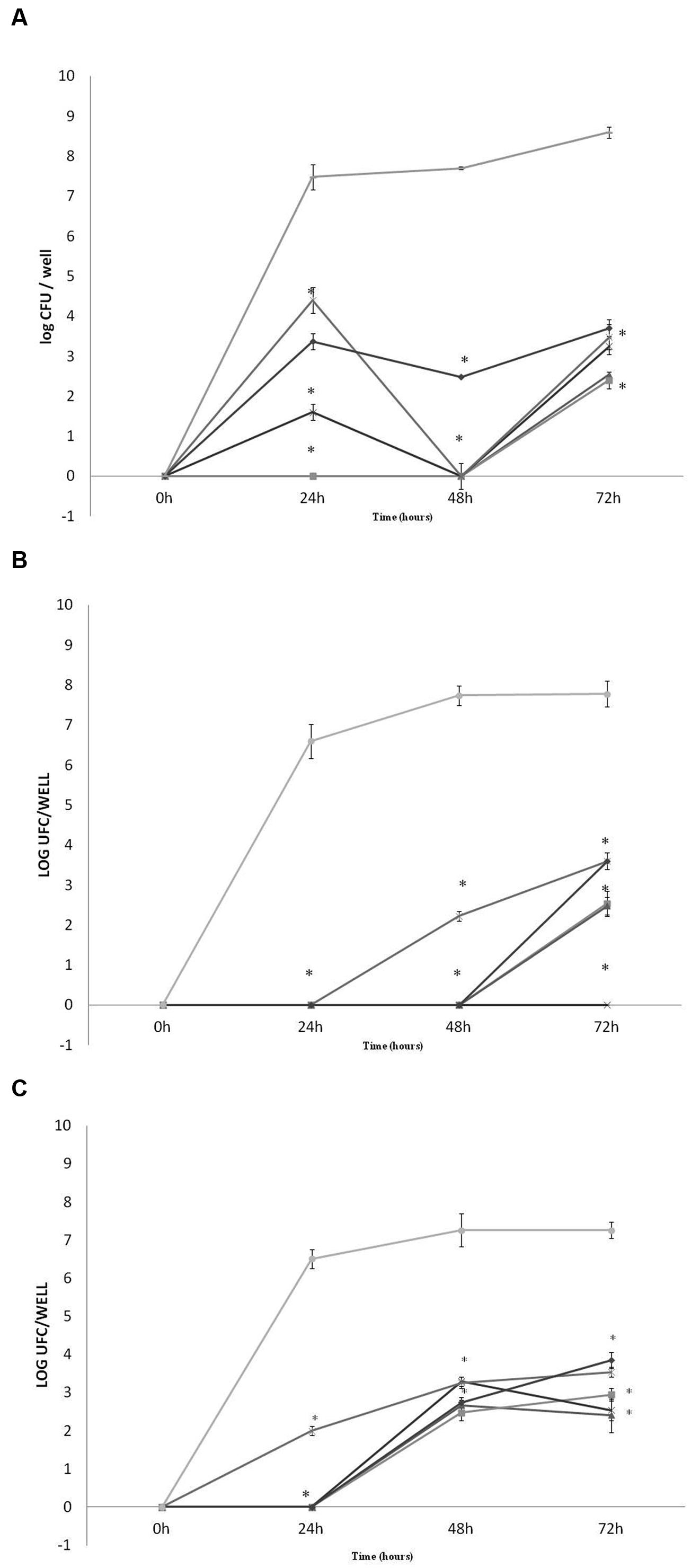
FIGURE 5. Quantification of pathogen biofilms on microtiter plates in MRS broth (A, Listeria monocytogenes ATCC 7644, B, S. Typhimurium ATCC 14028, C, E. coli O157:H7 ATCC 35150) in the presence of W. viridescens 113 bac- ( ); L. lactis 69 bac+ (
); L. lactis 69 bac+ ( ); L. lactis 94 bac+ (
); L. lactis 94 bac+ ( ); Lactobacillus casei 40 bac- (
); Lactobacillus casei 40 bac- ( ), and Lactobacillus helveticus 352 bac- (
), and Lactobacillus helveticus 352 bac- ( ), biofilms after 24, 48, and 72h at 30°C. Listeria monocytogenes, S. Typhimurium, and E. coli O157:H7 positive control (
), biofilms after 24, 48, and 72h at 30°C. Listeria monocytogenes, S. Typhimurium, and E. coli O157:H7 positive control ( ). No counts of the pathogens biofilms were detected in the presence of Lactobacillus. sakei MBSa1 bac+, Lactobacillus curvatus MBSa3 bac+ and L. lactis 368 bac- biofilms. Results are mean of triplicates and vertical bars show standard deviations. Superscript ∗ indicates a significant difference (p < 0.05) compared to the control (pathogens alone).
). No counts of the pathogens biofilms were detected in the presence of Lactobacillus. sakei MBSa1 bac+, Lactobacillus curvatus MBSa3 bac+ and L. lactis 368 bac- biofilms. Results are mean of triplicates and vertical bars show standard deviations. Superscript ∗ indicates a significant difference (p < 0.05) compared to the control (pathogens alone).
In addition, when supernatants were studied, planktonic pathogens cells were not detected, in all studied cases counts of pathogenic cells were below the detection limit (<10 CFU/ml, data not shown).
Discussion
The increased resistance to disinfection processes may be aggravated when bacterial biofilms are formed on surfaces that are recalcitrant for clean, such as cracks, holes, or tube connections. When planktonic cells are released from these colonization microenvironments, they may enter the food production chain and proliferate if proper conditions for growth occur, compromising the safety, quality, and stability of the final product. The application of the competitive biofilms formed by bacteria that produce natural antimicrobial substances and biosurfactants can provide new opportunities for the control of pathogenic bacteria and avoid food cross contamination.
Aggregation and co-aggregation among bacteria play an important role in prevention of colonization of surfaces by pathogens (García-Cayuela et al., 2014) as it is well known that co-aggregation abilities of LAB strains might interfere with the ability of the pathogenic species to infect the host and can prevent the colonization of food-borne pathogens (García-Cayuela et al., 2014). In this study, the tested LAB, especially the bacteriocin-producing Lactobacillus strains, presented high auto-aggregation and co-aggregation results, Lactobacillus curvatus MBSa3 exhibited the highest co-aggregation (69% with Listeria monocytogenes and 74.6% with E. coli O157:H7) and in this case pathogenic biofilms were not detected after three times of incubation tested, 24, 48, and 72; in other side the lowest co-aggregation was exhibited by W. viridescens 113 (53.4% with Listeria monocytogenes and 38% with E. coli O157:H7) and pathogenic cells were detected in 48 and 72 h of incubation in the presence of biofilm from strain. Nevertheless in other strains, there was apparently no relationship between the detection of pathogens and the percentage of co-aggregation with them.
Aggregation can also increase the concentration of excreted inhibitory substances (Kaewnopparat et al., 2013). Thus, these food-associated lactobacilli that co-aggregate numerous pathogens are of special interest with regard to potential applications in food-processing plants. Correlation between adhesion ability and hydrophobicity, as measured by microbial adhesion to hydrocarbons, has been reported for some lactobacilli (Wadström et al., 1987), but also conflicting results have been reported (Vinderola et al., 2004). As a result, adhesion, surface hydrophobicity, autoaggregation, and co-aggregation are phenotypic traits that potentially provide microbial colonization advantages within the intestinal tract. Aggregation abilities and cell surface hydrophobicity may not be the only components responsible for adhesion but these are some of the criteria to bear in mind of a complex mechanism that enables microorganisms to interact with the host and exert its beneficial effect (García-Cayuela et al., 2014).
The result obtained in hydrophobicity, aggregation, and co-aggregaton tests correspond with previus works like García-Cayuela et al. (2014). Di Bonaventura et al. (2008) reported a connection between hydrophobicity of cell surface and bacterial attachment, colonization, and biofilm formation. Our results show high values of hydrophobicity as well as a strong biofilm production, for most of the strains studied but there was no apparent correlation between hydrophobicity highest values and the strongest biofilm production. W. viridescens 113 shows a moderate biofilm production while displaying one of the highest hydrophobicity values. All tested LAB strains were tolerant to bile salts and acidic pH, evidencing their resistance to digestive stress and potential as probiotic agents. For a probiotic microorganism to be of benefit to human health it must survive the passage through the upper GIT and be able to function in the gut environment (Giraffa et al., 2010). Their functional requirements include tolerance to acid and bile, adherence to epithelial surfaces and antagonistic activity toward intestinal pathogens (Ramos et al., 2013; Peres et al., 2014). All LAB strains except L. lactis 368 were negative for β-galactosidase production (data not shown). This characteristic is disadvantageous for the probiotic activity of most studied LAB, as strains able to hydrolyze lactose might be useful for minimizing the effects of lactose intolerance (De Vrese et al., 2001).
Resistance of the LAB strains to antibiotics was species and strain dependent. Lactobacillus helveticus 352 and L. lactis VB69 were susceptible to all tested antibiotics, but Lactobacillus sakei MBSa1 was resistant to vancomycin, erythromycin, ampicillin, and tetracycline. Data from various studies on Lactobacillus spp. resistance to various antimicrobial agents demonstrate the existence of inter-genus and inter-species differences (Danielsen and Wind, 2003). The natural resistance to multiple classes of antibiotics is probably due to cell wall structure and membrane permeability, complemented in some cases by the efflux mechanisms (Ammor et al., 2007). However, this feature might represent a competitive advantage, especially when a probiotic product is administered with antimicrobials for treatment of an infectious disease thereby reducing the likelihood of disbiosis (microbial imbalance), rapidly rebalancing normal microbiota (Peres et al., 2014). The EFSA requires that bacteria which are to be introduced into the food chain lack acquired antimicrobial resistance determinants to prevent lateral spread of these (van Reenen and Dicks, 2011). Therefore for the cases of strains who presented antibiotic resistances, future genetic studies are needed to confirm if this resistance is due to acquired antimicrobials determinants. The presence of efaAfm in some strains seems to have no value as a risk indicator since this gene was also found in starter E. faecium strains with a long record of safe use in food (Eaton and Gasson, 2001). High frequencies of positive results were observed for, esp and efaAfm, in Lactococcus and Lactobacillus strains (Table 5). Furthermore, efaAfm and esp genes are related to the production of substances enrolled in the microbial colonization and adhesion at biotic and non-biotic surfaces (Valenzuela et al., 2009). W. viridescens 113 was positive for GelE but did not produce gelatinase, Eaton and Gasson (2001) described that gelE expression is highly influenced by the culture conditions, and the laboratory manipulation of the strains can result in the loss of the structural genes, and can explain the loss of gelatinase activity during in vitro tests. Moreover, W. virisdescens 113 and L. lactis 368 were positive for cob and cpd genes respectively, which are related to sex pheromones, although sex pheromones are not considered per se as virulence factors (Valenzuela et al., 2008). No strain was found positive for cytolisin family genes, and this confirms that the hemolysis present in blood agar was not related with these virulent genes.
Biosurfactant production is an interesting character, which can be related to the inhibition of the attachment of pathogens. The anti-adhesive and anti-biofilm-forming properties of lactobacilli have been reported in previous studies, such as Lactobacillus delbrueckii against E. coli (Abedi et al., 2013) and Lactobacillus brevis CD2 against Prevotella melaninogenica (Vuotto et al., 2013). In addition, Lactobacillus species were able to displace adhering uropathogenic Enterococcus faecalis from hydrophobic and hydrophilic substrata in a parallel-plate flow chamber (Velraeds et al., 1996). Biosurfactants from LABs have been shown to reduce adhesion of bacterial pathogens to glass, silicone rubber, surgical implants, and voice prostheses (Rodrigues et al., 2004). One xylolipid biosurfactant produced by a L. lactis strain with broad antibacterial activity against multidrug resistant E. coli and Staphylococcus aureus was described (Saravanakumari and Mani, 2010). Biosurfactants also been reported to have strong antifungal and antiviral activity (Singh and Cameotra, 2004). For the screening in biosurfactant production by haemolytic test, all the strains were positive except L. lactis 94. The strains showed, complete hemolysis with a diameter of lysis < 1 cm. In addition, drop collapse test was positive for all tested strains corresponding with partial spreading on the oil surface. None of the studies reported in the literature (Johnson and Boese-Marrazzo, 1980; Banat, 1993; Carrillo et al., 1996; Morán et al., 2002) mention the possibility of biosurfactant production without a hemolytic activity. However, in some cases hemolytic assay excluded many good biosurfactant producers (Youssef et al., 2004); hence in the present investigation the MATH assay and drop collapse test with crude oil were also done to confirm biosurfactant production.
The results of this study indicate that the tested LAB was capable to reduce Listeria monocytogenes, Salmonella and E. coli O157:H7 biofilm formation, and present probiotic characteristics and potentially no risk for the consumers. All strains were capable to hinder the development of pathogens in the first 72 h of incubation. Woo and Ahn (2013) obtained similar results of Listeria monocytogenes and Salmonella inhibition testing probiotic strains. Kim et al. (2013) showed the inactivation of E. coli O157:H7 on stainless steel upon exposure to Paenibacillus polymyxa biofilms. Zhao et al. (2013) reported the reduction of Listeria monocytogenes in a ready-to-eat poultry processing plant by LAB and Pérez-Ibarreche et al. (2014) reported that lactobacilli with biofilm-forming aptitudes were able to control Listeria monocytogenes biofilms. In this study inhibition, effect against biofilm adhesion was observed in bacteriocin producers L. lactis VB69 and VB94; Lactobacillus sakei MBSa1 and Lactobacillus curvatus MBSa3 as well as non-bacteriocin producers Lactococcus lactis—lactis 368, Lactobacillus helveticus 354, Lactobacillus casei 40 and W. viridescens 113. It seems that inhibition of pathogenic bacteria growth and adhesion is not only due to the bacteriocin production. This outcome can be attributed to a combination of factors like biosurfactant and bacteriocin production as well as mechanisms of pathogens exclusion through their trapping (killing of cells embedded in biofilms). This is in accordance with previous works like Guerrieri et al. (2009) witch suggests the need to apply the bacteriocine-producing microorganism, in biofilms. There may be an influence of EPS (exo-polysaccharide). Kim et al. (2006) found that the EPS of Lactobacillus acidophilus A4 had stronger anti-biofilm activity against the growth of entero-hemorrhagic E. coli O157: H7, S. enteritidis, S. typhimurium KKCCM 11806, Yersinia enterocolitica, Pseudomonas aeruginosa KCCM 11321, Listeria monocytogenes Scott A, and B. cereus.
Conclusion
Our results show that LAB strains from foods can be excellent candidates to form protective biofilms, in accordance with the hypothesis proposed by Falagas and Makris (2009) to use non-pathogenic microorganisms, namely probiotics, as part of daily cleaning products to lower the incidence of pathogenic microorganisms. Evidences on the efficacy of probiotics for the prevention and treatment of infections have been observed both in vitro and in vivo (Levkovich et al., 2013; Shu et al., 2013). The present study provided new information about the use of potential probiotic LAB biofilms for the control of Listeria monocytogenes, S. Typhimurium and E. coli O157:H7 biofilms formation through exclusion mechanisms. However, more experiments are needed to confirm the ability of these strains to inhibit the pathogen biofilm formation in other environments. Our initial studies are very encouraging and indicate that the LAB that we have tested are promising candidates for controlling the presence of pathogenic biofilms in food-processing facilities. The development of protective biofilms with probiotic LAB present in food could help avoiding problems of contamination into the food chain.
Author Contributions
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank São Paulo Research Foundation (FAPESP) for financial support (2013/07914-8) and fellowship to the author NG (2014/06370-7).
References
Abedi, D., Feizizadeh, S., Akbari, V., and Jafarian-Dehkordi, A. (2013). In vitro anti-bacterial and anti-adherence effects of Lactobacillus delbrueckii subsp bulgaricus on Escherichia coli. Res. Pharm. Sci. 8, 260–268.
Abriouel, H., Benomar, N., Cobo, A., Caballero, N., Fuentes, M. Á. F., Pérez-Pulido, R., et al. (2012). Characterization of lactic acid bacteria from naturally-fermented Manzanilla Aloreña green table olives. Food Microbiol. 32, 308–316. doi: 10.1016/j.fm.2012.07.006
Ammor, M. S., Flórez, A. B., and Mayo, B. (2007). Antibiotic resistance in non-enterococcal lactic acid bacteria and bifidobacteria. Food Microbiol. 24, 559–570. doi: 10.1016/j.fm.2006.11.001
Bambirra, F., Lima, K. G. C., Franco, B. D. G. M., Cara, D. C., Nardi, R. M. D., Barbosa, F. H. F., et al. (2007). Protective effect of Lactobacillus sakei 2a against experimental challenge with Listeria monocytogenes in gnotobiotic mice. Lett. Appl. Microbiol. 45, 663–667. doi: 10.1111/j.1472-765X.2007.02250.x
Banat, I. M. (1993). The isolation of a thermophilic biosurfactant producing Bacillus sp. Biotechnol. Lett. 15, 591–594. doi: 10.1007/BF00138546
Barbosa, M. S., Todorov, S. D., Ivanova, I., Chobert, J. M., Haertlé, T., and Franco, B. D. G. M. (2015). Improving safety of salami by application of bacteriocins produced by an autochthonous Lactobacillus curvatus isolate. Food Microbiol. 46, 254–262. doi: 10.1016/j.fm.2014.08.004
Bíscola, V., Todorov, S. D., Capuano, V. S. C., Abriouel, H., Galvez, A., and Franco, B. D. G. M. (2013). Isolation and characterization of a nisin-like bacteriocin produced by a Lactococcus lactis strain isolated from charqui, a Brazilian fermented, salted and dried meat product. Meat Sci. 93, 607–613. doi: 10.1016/j.meatsci.2012.11.021
Borges, S., Silva, J., and Teixeira, P. (2012). Survival and biofilm formation by Group B streptococci in simulated vaginal fluid at different pHs. A Van Leeuw 101, 677–682. doi: 10.1007/s10482-011-9666-y
Carpentier, B., and Cerf, O. (2011). Review—Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 145, 1–8. doi: 10.1016/j.ijfoodmicro.2011.01.005
Carrillo, P. G., Mardaraz, C., Pitta-Alvarez, S. I., and Giulietti, A. M. (1996). Isolation and selection of biosurfactant-producing bacteria. World J. Microbiol. Biotechnol. 12, 82–84. doi: 10.1007/BF00327807
Cibik, R., Lepage, E., and Tailliez, P. (2000). Molecular Diversity of Leuconostoc mesenteroides and Leuconostoc citreum isolated from Traditional French Cheeses as Revealed by RAPD Fingerprinting, 16S rDNA Sequencing and 16S rDNA Fragment Amplification. Syst. Appl. Microbiol. 23, 267–278. doi: 10.1016/S0723-2020(00)80014-4
Collado, M. C., Meriluoto, J., and Salminen, S. (2008). Adhesion and aggregation properties of probiotic and pathogen strains. Eur. Food Res. Technol. 226, 1065–1073. doi: 10.1007/s00217-007-0632-x
Danielsen, M., and Wind, A. (2003). Susceptibility of Lactobacillus spp. to antimicrobial agents. Int. J. Food Microbiol. 82, 1–11. doi: 10.1016/S0168-1605(02)00254-4
De Man, J. C., Rogosa, M., and Sharpe, M. E. (1960). A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 23, 130–135. doi: 10.1111/j.1365-2672.1960.tb00188.x
De Vrese, M., Stegelmann, A., Richter, B., Fenselau, S., Laue, C., and Schrezenmeir, J. (2001). Probiotics – compensation for lactase insufficiency. Am. J. Clin. Nutr. 73, 421–429.
Di Bonaventura, G., Piccolomini, R., Paludi, D., D’orio, V., Vergara, A., Conter, M., et al. (2008). Influence of temperature on biofilm formation by Listeria monocytogenes on various food-contact surfaces: relationship with motility and cell surface hydrophobicity. J. Appl. Microbiol. 104, 1552–1561. doi: 10.1111/j.1365-2672.2007.03688.x
Eaton, T. J., and Gasson, M. J. (2001). Molecular screening of enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 67, 1628–1635. doi: 10.1128/AEM.67.4.1628-1635.2001
European Commission SCAN (2007). Opinion of the Scientific Committee on a request from EFSA on the introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA. EFSA J. 587, 1–16.
European Food Safety Authority [EFSA] (2012). Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. Available at: http://www.efsa.europa.eu/en/colloquiareports/colloquiaqps.htm
Falagas, M. E., and Makris, G. C. (2009). Probiotic bacteria and biosurfactants for nosocomial infection control: a hypothesis. J. Hosp. Infect. 71, 301–306. doi: 10.1016/j.jhin.2008.12.008
FAO/WHO (2006). Probiotics in Food. Health and Nutritional Properties and Guidelines for Evaluation. Rome: FAO Food and Nutrition.
Favaro, L., Basaglia, M., Casella, S., Hue, I., Dousset, X., Franco, B. D. G. M., et al. (2014). Bacteriocinogenic potential and safety evaluation of non-starter Enterococcus faecium strains isolated from home made white brine cheese. Food Microbiol. 38, 228–239. doi: 10.1016/j.fm.2013.09.008
Flórez, A. B., Delgado, S., and Mayo, B. (2005). Antimicrobial susceptibility of lactic acid bacteria isolated from a cheese environment. Can. J. Microbiol. 51, 51–58. doi: 10.1139/w04-114
Gálvez, A., Abriouel, H., Benomar, N., and Lucas, R. (2010). Microbial antagonists to food-borne pathogens and biocontrol. Curr. Opin. Biotechnol. 21, 142–148. doi: 10.1016/j.copbio.2010.01.005
Gandhi, M., and Chikindas, M. L. (2007). Listeria: a foodborne pathogen that knows how to survive. Int. J. Food Microbiol. 113, 1–15. doi: 10.1016/j.ijfoodmicro.2006.07.008
Ganzle, M. G., Hertel, C., van der Vossen, J. M., and Hammes, W. P. (1999). Effect of bacteriocin-producing lactobacilli on the survival of Escherichia coli and Listeria in a dynamic model of the stomach and the small intestine. Int. J. Food Microbiol. 48, 21–35. doi: 10.1016/S0168-1605(99)00025-2
García-Cayuela, T., Korany, A. M., Bustos, I., de Cadiñanos, L. P. G., Requena, T., Peláez, C., et al. (2014). Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 57, 44–50. doi: 10.1016/j.foodres.2014.01.010
Gatesoupe, F. J. (1999). The use of probiotics in aquaculture. Aquaculture 180, 147–165. doi: 10.1016/S0044-8486(99)00187-8
Giraffa, G., Chanishvili, N., and Widyastuti, Y. (2010). Importance of lactobacilli in food and feed biotechnology. Res. Microbiol. 161, 480–487. doi: 10.1016/j.resmic.2010.03.001
Gómez, N. C., Abriouel, H., Grande, M. J., Pulido, R. P., and Gálvez, A. (2012). Effect of enterocin AS-48 in combination with biocides on planktonic and sessile Listeria monocytogenes. Food Microbiol. 30, 51–58. doi: 10.1016/j.fm.2011.12.013
Guerrieri, E., de Niederhäusern, S., Messi, P., Sabia, C., Iseppi, R., Anacarso, I., et al. (2009). Use of lactic acid bacteria (LAB) biofilms for the control of Listeria monocytogenes in a small-scale model. Food Control 20, 861–865. doi: 10.1016/j.foodcont.2008.11.001
Handley, P. S., Harty, D. W., Wyatt, J. E., Brown, C. R., Doran, J. P., and Gibbs, A. C. (1987). A comparison of the adhesion, coaggregation and cell-surface hydrophobicity properties of fibrillar and fimbriate strains of Streptococcus salivarius. J. Gen. Microbiol. 133, 3207–3217. doi: 10.1099/00221287-133-11-3207
Hibbing, M. E., Fuqua, C., Parsek, M. R., and Peterson, S. B. (2010). Bacterial competition: surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 8, 15–25. doi: 10.1038/nrmicro2259
Jain, D. K., Collins-Thompson, D. L., Lee, H., and Trevors, J. T. (1991). A drop-collapsing test for screening surfactant-producing microorganisms. J. Microbiol. Methods 13, 271–279. doi: 10.1016/0167-7012(91)90064-W
Johnson, M. K., and Boese-Marrazzo, D. E. B. O. R. A. H. (1980). Production and properties of heat-stable extracellular hemolysin from Pseudomonas aeruginosa. Infect. Immun. 29, 1028–1033.
Jonkers, D. M. A. E. (2016). Microbial perturbations and modulation in conditions associated with malnutrition and malabsorption. Best Pract. Res. Clin. Gastroenterol. 30, 161–172. doi: 10.1016/j.bpg.2016.02.006
Kaewnopparat, S., Dangmanee, N., Kaewnopparat, N., Srichana, T., Chulasiri, M., and Settharaksa, S. (2013). In vitro probiotic properties of Lactobacillus fermentum SK5 isolated from vagina of a healthy woman. Anaerobe 22, 6–13. doi: 10.1016/j.anaerobe.2013.04.009
Kanmani, P., Satish Kumar, R., Yuvaraj, N., Paari, K. A., Pattukumar, V., and Arul, V. (2013). Probiotics and its functionally valuable products—A review. Crit. Rev. Food Sci. 53, 641–658. doi: 10.1080/10408398.2011.553752
Kim, S., Bang, J., Kim, H., Beuchat, L. R., and Ryu, J. H. (2013). Inactivation of Escherichia coli O157:H7 on stainless steel upon exposure to Paenibacillus polymyxa biofilms. Int. J. Food Microbiol. 167, 328–336. doi: 10.1016/j.ijfoodmicro.2013.10.004
Kim, Y. H., Lee, Y., Kim, S., Yeom, J., Yeom, S., Oh, K. B. S. S., et al. (2006). The role of periplasmic antioxidant enzymes (superoxide dismutase and thiol peroxidase) of the Shiga toxin producing Escherichia coli O157:H7 in the formation of biofilms. Proteomics 6, 6181–6193. doi: 10.1002/pmic.200600320
Kotzamanidis, C., Kourelis, A., Litopoulou-Tzanetaki, E., Tzanetakis, N., and Yiangou, M. (2010). Evaluation of adhesion capacity, cell surface traits and immunomodulatory activity of presumptive probiotic Lactobacillus strains. Int. J. Food Microbiol. 140, 154–163. doi: 10.1016/j.ijfoodmicro.2010.04.004
Lahtinen, S., Ouwehand, A. C., Salminen, S., and von Wright, A. (2011). Lactic Acid Bacteria: Microbiological and Functional Aspects. Boca Raton, FL: CRC Press.
Levkovich, T., Poutahidis, T., Smillie, C., Varian, B. J., Ibrahim, Y. M., Lakritz, J. R., et al. (2013). Probiotic bacteria induce a ‘glow of health’. PLoS ONE 8:e53867. doi: 10.1371/journal.pone.0053867
Liu, S., Ye H., and Zhi-jiang Z. (2011). Lactic acid bacteria in traditional fermented Chinese foods. Food Res. Int. 3, 643–651. doi: 10.1016/j.foodres.2010.12.034
Lobos, O., Padilla, A., and Padilla, A. (2009). In vitro antimicrobial effect of bacteriocin PsVP-10 in combination with chlorhexidine and triclosan against Streptococcus mutans and Streptococcus sobrinus strains. Arch. Oral Biol. 54, 230–234. doi: 10.1016/j.archoralbio.2008.11.007
Mendonça, R. C. S., Morelli, A. M. F., Pereira, J. A. M., de Carvalho, M. M., and de Souza, N. L. (2012). Prediction of Escherichia coli O157: H7 adhesion and potential to form biofilm under experimental conditions. Food Control 23, 389–396. doi: 10.1016/j.foodcont.2011.08.004
Midelet, G., and Carpentier, B. (2004). Impact of cleaning and disinfection agents on biofilm structure and on microbial transfer to a solid model food. J. Appl. Microbiol. 97, 262–270. doi: 10.1111/j.1365-2672.2004.02296.x
Millette, M., Cornut, G., Dupont, C., Shareck, F., Archambault, D., and Lacroix, M. (2008). Capacity of human nisin- and pediocin-producing lactic acid bacteria to reduce intestinal colonization by vancomycin-resistant enterococci. Appl. Environ. Microbiol. 74, 1997–2003. doi: 10.1128/AEM.02150-07
Morán, A. C., Martinez, M. A., and Siñeriz, F. (2002). Quantification of surfactin in culture supernatants by hemolytic activity. Biotechnol. Lett. 24, 177–180. doi: 10.1023/A:1014140820207
Muñoz, M. D. C., Benomar, N., Lerma, L. L., Gálvez, A., and Abriouel, H. (2014). Antibiotic resistance of Lactobacillus pentosus and Leuconostoc pseudomesenteroides isolated from naturally-fermented Aloreña table olives throughout fermentation process. Int. J. Food Microbiol. 172, 110–118. doi: 10.1016/j.ijfoodmicro.2013.11.025
Palomino, J. M., Árbol, J. T., Benomar, N., Abriouel, H., Cañamero, M. M., Gálvez, A., et al. (2015). Application of Lactobacillus plantarum Lb9 as starter culture in caper berry fermentation. LWT-Food Sci. Technol. 60, 788–794. doi: 10.1016/j.lwt.2014.09.061
Peres, C. M., Alves, M., Hernandez-Mendoza, A., Moreira, L., Silva, S., Bronze, M. R., et al. (2014). Novel isolates of lactobacilli from fermented Portuguese olive as potential probiotics. LWT-Food Sci. Technol. 59, 234–246. doi: 10.1016/j.lwt.2014.03.003
Pérez-Ibarreche, M., Castellano, P., and Vignolo, G. (2014). Evaluation of anti-Listeria meat borne Lactobacillus for biofilm formation on selected abiotic surfaces. Meat Sci. 96, 295–303. doi: 10.1016/j.meatsci.2013.07.010
Perin, L. M., Miranda, R. O., Todorov, S. D., Franco, B. D. G. M., and Nero, L. A. (2014). Virulence, antibiotic resistance and biogenic amines of bacteriocinogenic lactococci and enterococci isolated from goat milk. Int. J. Food Microbiol. 185, 121–126. doi: 10.1016/j.ijfoodmicro.2014.06.001
Pompermayer, D. M., and Gaylarde, C. C. (2000). The influence of temperature on the adhesion of mixed cultures of Staphylococcus aureus and Escherichia coli to polypropylene. Food Microbiol. 17, 361–365. doi: 10.1006/fmic.1999.0291
Ramos, C. L., Thorsen, L., Schwan, R. F., and Jespersen, L. (2013). Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiol. 36, 22–29. doi: 10.1016/j.fm.2013.03.010
Re, B., Sgorbati, B., Miglioli, M., and Palenzona, D. (2000). Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 31, 438–442. doi: 10.1046/j.1365-2672.2000.00845.x
Rodrigues, L., van der Mei, H., Teixeira, J., and Oliveira, R. (2004). Biosurfactant from Lactococcus lactis 53 inhibits microbial adhesion on silicone rubber. Appl. Microbiol. Biotechnol. 66, 306–311. doi: 10.1007/s00253-004-1674-7
Saravanakumari, P., and Mani, K. (2010). Structural characterization of a novel xylolipid biosurfactant from Lactococcus lactis and analysis of antibacterial activity against multi-drug resistant pathogens. Bioresource Technol. 101, 8851–8854. doi: 10.1016/j.biortech.2010.06.104
Savard, P., Lamarche, B., Paradis, M. E., Thiboutot, H., Laurin,É., and Roy, D. (2011). Impact of Bifidobacterium animalis subsp. lactis BB-12 and, Lactobacillus acidophilus LA-5-containing yoghurt, on fecal bacterial counts of healthy adults. Int. J. Food Microbiol. 149, 50–57. doi: 10.1016/j.ijfoodmicro.2010.12.026
Semedo, T., Santos, M. A., Lopes, M. F., Marques, J. J. F., Crespo, M. T., and Tenreiro, R. (2003). Virulence factors in food, clinical and reference enterococci: a common trait in the genus? Syst. Appl. Microbiol. 26, 13–22. doi: 10.1078/072320203322337263
Shu, M., Wang, Y., Yu, J., Kuo, S., Coda, A., Jiang, Y., et al. (2013). Fermentation of Propionibacterium acnes, a commensal bacterium in the human skin microbiome, as skin probiotics against methicillin-resistant Staphylococcus aureus. PLoS ONE 8:e55380. doi: 10.1371/journal.pone.0055380
Singh, P., and Cameotra, S. (2004). Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol. 22, 142–146. doi: 10.1016/j.tibtech.2004.01.010
Stepanović, S., Cirković, I., and Ranin, L. (2004). Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 38, 428–432. doi: 10.1111/j.1472-765X.2004.01513.x
Valenzuela, A. S., ben Omar, N., Abriouel, H., López, R. L., Veljovic, K., Cañamero, M. M., et al. (2009). Virulence factors, antibiotic resistance, and bacteriocins in enterococci from artisan foods of animal origin. Food Control 20, 381–385. doi: 10.1016/j.foodcont.2008.06.004
Valenzuela, A. S., Omar, N. B., Abriouel, H., López, R. L., Ortega, E., Cañamero, M. M., et al. (2008). Risk factors in enterococci isolated from foods in Morocco: determination of antimicrobial resistance and incidence of virulence traits. Food Chem. Toxicol. 46, 2648–2652. doi: 10.1016/j.fct.2008.04.021
Van Houdt, R., and Michiels, C. W. (2010). Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 109, 1117–1131. doi: 10.1111/j.1365-2672.2010.04756.x
van Reenen, C. A., and Dicks, L. M. (2011). Horizontal gene transfer amongst probiotic lactic acid bacteria and other intestinal microbiota: what are the possibilities? A review. Arch. Microbiol. 193, 157–168. doi: 10.1007/s00203-010-0668-3
Velraeds, M., van der Mei, H., Reid, G., and Busscher, H. (1996). Physico-chemical and biochemical characterization of biosurfactants released by Lactobacillus strains. Colloid Surface B 8, 51–61. doi: 10.1016/S0927-7765(96)01297-0
Viedma, P. M., Ercolini, D., Ferrocino, I., Abriouel, H., Omar, N. B., López, R. L., et al. (2010). Effect of polythene film activated with enterocin EJ97 in combination with EDTA against Bacillus coagulans. LWT-Food Sci. Technol. 43, 514–518. doi: 10.1016/j.lwt.2009.09.020
Vinderola, C. G., Medici, M., and Perdigon, G. (2004). Relationship between interaction sites in the gut, hydrophobicity, mucosal immunomodulating capacities and cell wall protein profiles in indigenous and exogenous bacteria. J. Appl. Microbiol. 96, 230–243. doi: 10.1046/j.1365-2672.2004.02158.x
Vuotto, C., Barbanti, F., Mastrantonio, P., and Donelli, G. (2013). Lactobacillus brevis CD2 inhibits Prevotella melaninogenica biofilm. Oral Dis. 20, 668–674. doi: 10.1111/odi.12186
Wadström, T., Andersson, K., Sydow, M., Axelsson, L., Lindgren, S., and Gullmar, B. (1987). Surface properties of lactobacilli isolated from the small intestine of pigs. J. Appl. Microbiol. 62, 513–520. doi: 10.1111/j.1365-2672.1987.tb02683.x
Wan, L. Y. M., Chen, Z. J., Shah, N. P., and El-Nezami, H. (2015). Modulation of intestinal epithelial defense responses by probiotic bacteria. Crit. Rev. Food Sci. Nutr. doi: 10.1080/10408398.2014.905450 [Epub ahead of print].
Winkelströter, L. K., Reis, F. B., Silva, E. P., Alves, V., and De Martins, E. C. P. (2013). Unraveling microbial biofilms of importance for food microbiology. Microb. Ecol. 68, 35–46. doi: 10.1007/s00248-013-0347-4
Woo, J., and Ahn, J. (2013). Probiotic-mediated competition, exclusion and displacement in biofilm formation by food-borne pathogens. Lett. Appl. Microbiol. 4, 307–313. doi: 10.1111/lam.12051
Youssef, N. H., Duncan, K. E., Nagle, D. P., Savage, K. N., Knapp, R. M., and McInerney, M. J. (2004). Comparison of methods to detect biosurfactant production by diverse microorganisms. J. Microbiol. Methods 56, 339–347. doi: 10.1016/j.mimet.2003.11.001
Keywords: biofilm, probiotic, lactic acid bacteria, exclusion, pathogens, biocontrol
Citation: Gómez NC, Ramiro JMP, Quecan BXV and de Melo Franco BDG (2016) Use of Potential Probiotic Lactic Acid Bacteria (LAB) Biofilms for the Control of Listeria monocytogenes, Salmonella Typhimurium, and Escherichia coli O157:H7 Biofilms Formation. Front. Microbiol. 7:863. doi: 10.3389/fmicb.2016.00863
Received: 22 February 2016; Accepted: 23 May 2016;
Published: 10 June 2016.
Edited by:
Romain Briandet, Institut National de la Recherche Agronomique, FranceReviewed by:
Efstathios D. Giaouris, University of the Aegean, GreeceDjamel Drider, University of Lille 1, France
Olivier Habimana, University College Dublin, Ireland
Copyright © 2016 Gómez, Ramiro, Quecan and de Melo Franco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Natacha C. Gómez, bmNnb21lekB1c3AuYnI=